Abstract
Influenza A virus (IAV) matrix protein 2 (M2), an ion channel, is crucial for virus infection, and therefore, an important anti-influenza drug target. Adamantanes, also known as M2 channel blockers, are one of the two classes of Food and Drug Administration-approved anti-influenza drugs, although their use was discontinued due to prevalent drug resistance. Fast emergence of resistance to current anti-influenza drugs have raised an urgent need for developing new anti-influenza drugs against resistant forms of circulating viruses. Here we propose a simple theoretical criterion for fast virtual screening of molecular libraries for candidate anti-influenza ion channel inhibitors both for wild type and adamantane-resistant influenza A viruses. After in silico screening of drug space using the EIIP/AQVN filter and further filtering of drugs by ligand based virtual screening and molecular docking we propose the best candidate drugs as potential dual inhibitors of wild type and adamantane-resistant influenza A viruses. Finally, guanethidine, the best ranked drug selected from ligand-based virtual screening, was experimentally tested. The experimental results show measurable anti-influenza activity of guanethidine in cell culture.
Introduction
Influenza is a serious global public health concern. Regardless of the availability of antiviral drugs and vaccines, according to the World Health Organization's estimates, influenza is the cause of 3 to 5 million cases of severe illness and about 290,000–650,000 deaths in seasonal outbreaks worldwide (WHO Influenza, 2018). Annual “flu” vaccination has a primary role in preventing influenza A and B virus infections and increasing population immunity even though the efficacy of the seasonal flu vaccines may vary from year to year (Bridges et al., 2013). Since the current flu vaccination approach is imperfect, a substantial portion of the population is susceptible to infection even after vaccination every year. Therefore, alternative strategies should be considered to improve our therapeutic abilities for those patients that develop clinical flu. This would be especially important in the pandemic setting with rapid virus transmission due to the currently limited ability for fast, new vaccine production (Bridges et al., 2013). Current treatment and prophylaxis against seasonal influenza is limited to the only licensed class of antivirals, namely neuraminidase inhibitors (NAIs). Oseltamivir and zanamivir are currently licensed worldwide while peramivir and laninamivir are approved in some countries (Ison, 2017). The frequency of NAI resistance in currently circulating strains is low, <1% (Hurt et al., 2016), but resistance to oseltamivir, the most widely used NAI, was extensive amongst former seasonal H1N1 viruses in 2008. (Hurt et al., 2009), and was detected in localized clusters of oseltamivir-resistant H1N1pdm09 (Hurt et al., 2011). Nevertheless, the therapeutic window for treatment with these drugs is very short and patients benefit the most when treated 24–48 h after the onset of “flu” symptoms (Ison, 2017).
The first of the two classes of FDA-approved anti-influenza drugs are adamantanes, amantadine and rimantadine, that inhibit viral replication by blocking the wild-type (WT) M2 proton channel.
IAV matrix protein 2 (M2), an ion channel protein, is one of the most conserved viral proteins and essential for efficient virus replication, and is thus an important anti-influenza drug target (Takeda et al., 2002). Matrix protein 2 (M2) is a 97-residue-long viral protein that encompasses a 19-residue-long hydrophobic transmembrane domain (TM) that forms a homotetrameric proton-selective channel involved in proton conductance and drug binding (Lamb et al., 1985; Sakaguchi et al., 1997). Adamantanes have been used successfully against influenza A virus infection for more than 30 years because of their wide accessibility and low price (Dolin et al., 1982). However, as a consequence of the lack of activity against influenza B (Mould et al., 2003), adverse effects, and the rapid emergence of resistance during treatment or even in the absence of selective drug pressure, the Centers for Disease Control and Prevention (CDC) have strongly recommended against the use of this class of drugs (CDC, 2006). The molecular basis for resistance to adamantanes is connected with several amino acid substitutions and the M2-S31N variant is found in more than 95% of the currently circulating influenza A viruses (Dong et al., 2015). The expansion of M2 viruses with S31N mutation in the early 2000s is not a consequence of drug selection pressure but is connected to advantageous substitutions elsewhere in the virus, in a process denoted as genetic “hitch-hiking” (Simonsen et al., 2007). On the other hand, the latest report of M2 S31 and D31 viruses in Australia suggests that the role of the M2 N31 residue in viral fitness is no longer as important as it used to be (Hurt et al., 2017). Considering all these facts, new effective anti-influenza M2 inhibitors that target both WT and S31N mutant are greatly needed. Several high-resolution M2 structures that provide important insights into the favorable structural features can be employed for designing new M2 inhibitors (Hong and DeGrado, 2012).
A recent, and very popular drug discovery approach—drug repurposing (DR), wherein old drugs are given new indication by exploring new molecular pathways and targets for intervention (Strittmatter, 2014)- offers potential economic advantage and shorter regulatory process for the clinical approval. The continuous increase of drug-resistant pathogens is a great challenge for treatment of infectious diseases and DR serves as an alternative approach for rapid identification of effective therapeutics (Zheng et al., 2018). Drug repurposing (DR) applied to viral infectious diseases integrates both screening of bioactive small-molecule collections and computational methods to find a molecule, a pathway, or a biological activity that could be used against the virus of interest (Mercorelli et al., 2018). Two clinical trials against influenza viruses with repositioned drugs are currently underway: (1) The first trial (phase 2b/3clinical trial) combines clarithromycin and naproxen along with oseltamivir in a triple-drug combination and; (2) the second trial is focused on testing efficacy of an antiparasitic drug, nitazoxanide, against influenza viruses (Phase III) (Mercorelli et al., 2018).
In this study we propose a simple theoretical criterion for fast virtual screening of molecular libraries for candidate anti-influenza M2 ion channel inhibitors both for wild type and adamantane-resistant influenza A viruses. After in silico screening of drug space using the EIIP/AQVN filter, and further filtering of drugs by ligand based virtual screening and molecular docking, we proposed the five best candidate drugs as potential dual inhibitors of wild type and adamantane-resistant influenza A viruses.
Materials and Methods
For screening of drugs for repurposing to select candidates for influenza M2 inhibitors, 2,627 approved small molecule drugs from DrugBank (http://www.drugbank.ca) were screened. To define the predictive criterion for the selection of Influenza M2 candidates, the learning set (Supplementary Tables 1, 2) was composed of all active compounds from ChEMBL Target Report Card (https://www.ebi.ac.uk/chembl/target/inspect/CHEMBL613740) (EMBL-EBI. ChEMBL). (EMBL-EBI. ChEMBL. Available online: https://www.ebi.ac.uk/chembl/ (accessed on June 30, 2018) against influenza A virus M2 (Target ID CHEMBL613740) both for wild type (WT) and S31N, with corresponding IC50 values. The total number of reported compounds for WT and S39N of M2 channel were 50 and 49, respectively. After removal of duplicates and inactive compounds, the final number of compounds was 15 for WT and 12 for the S31N mutant (Supplementary Tables 1, 2). The control data sets were compounds from PubChem compounds database (http://www.ncbi.nlm.nih.gov/pccompound).
Virtual Screening
The virtual screening (VS) protocol included the application of subsequent filters to select candidate dual inhibitors of M2 ion channel. The first EIIP/AQVN filter approach was employed for in silico screening of the ChEMBL Target Report Card (https://www.ebi.ac.uk/chembl/target/inspect/CHEMBL613740) and DrugBank (http://www.drugbank.ca) (Wishart et al., 2006) and then proceeded by ligand-based screening.
EIIP/AQVN
The EIIP for organic molecules can be determined by the following simple equation derived from the “general model pseudopotential (Veljkovic et al., 2011).
where Z* is the average quasi valence number (AQVN) determined by
Where Zi is the valence number of the ith atomic component, ni is the number of atoms of the ith component, m is the number of atomic components in the molecule, and N is the total number of atoms. EIIP values calculated according to Equations (1, 2) are expressed in Rydberg units (Ry).
Ligand-Based Virtual Screening
To screen selected compounds from Drugbank, both learning set compounds and candidates from the previous step were converted to 3D sdf format from smiles. GRIND descriptors of molecules were calculated, based on molecular interaction field (MIF) probes (Duran et al., 2009). Computation method for descriptor generation was GRID with step 0.5. Applied probes (mapped regions of molecule surface) were DRY (hydrophobic interactions) O (hydrogen bond acceptor) N1 (hydrogen bond donor) and TIP (molecular shape descriptor). Discretization Method was AMANDA (Duran et al., 2008), with scale factor 0.55. Cut off was set to: DRY −0.5 O −2.6 N1 −4.2 TIP −0.75. Encoding Method was MACC2 and weights were the following: DRY: −0.5, O: −2.6, N1: −4.2, TIP: −0.75. Number of PCA components was set to five. Explained variance of such obtained model was 58.84%. Then, learning set compounds were imported and served for screening the candidate compound database. All calculations were carried in Pentacle software version 1.06 for Linux (Pastor et al., 2000).
Molecular Docking
Receptor Preparation
Crystal structures of the wild type M2 channel and the S31N mutant channel were downloaded from RCSB PDB database (https://www.rcsb.org/) with PDBIDs 2KQT (Cady et al., 2010) and 2LY0 (Wang et al., 2013) respectively. All ligands, ions and water molecules were removed from structures. All hydrogen atoms were added on protein structures and then truncated to only polar hydrogen atoms during the preparation process. The receptor was prepared in ADT Tools 1.5.6 (Sanner, 1999; Morris et al., 2009).
Ligand Preparation
Ligands were converted from 3Dsdf to mol2 format and imported to Avogadro software in order to protonate them at physiological pH. Molecules were prepared for MOPAC 2016 (Stewart, 2016) and geometrically optimized on PM7 (Stewart, 2013) level of theory. They were further prepared for molecular docking in ADT Tools.
Molecular Docking
A grid box with dimensions 24 × 24 × 24 A was placed in the center of the binding site of the protein receptor. Exhaustiveness was set to 50. Molecular docking was carried in Autodock Vina (Trott and Olson, 2010).
In vitro Efficacy Testing of Guanethidine Against Influenza a (h1n1) Virus
Influenza A/CA/07/2009 (H1N1) virus was premixed with 1, 10, and 100 μM of guanethidine and incubated at 37C for 1 hr. Positive control wells were prepared by mixing influenza A/CA/07/2009 (H1N1) virus with 10 μM of merimepodib. MDCK cells were then infected in triplicates with influenza A/CA/07/2009 (H1N1) virus / drug mixture. After ~1 h of incubation at 37°C and 5% CO2, cells were washed with serum free media and 1 × of each compound dose was added to the cells. Virus control wells as well as untreated control wells were included in triplicates. Cells were incubated at 37C and 5% CO2 and samples were collected at 0, 1, 2, and 3 days post-infection. Samples were stored at −80°C until the day of analysis. The influenza virus titer in MDCK cells via TCID50 was performed for each sample collected at days 0, 1, and 2 post-infection.
Results
The virtual screening (VS) protocol in this study was based on the application of sequential filters to select candidate dual inhibitors of the M2 ion channel. Previously it was shown for molecular targets in diverse pathological states that small molecules with similar AQVN and EIIP values interact with the common therapeutic targets (Veljkovic et al., 2011, 2013). This resulted in determining criteria for virtual screening of molecular libraries for compounds with similar therapeutic properties (Veljkovic et al., 2013). The learning set consists of M2 WT (Supplementary Table 1), and M2 S31N mutant (Supplementary Table 2) ion channel inhibitors from the ChEMBL Target Report Card (https://www.ebi.ac.uk/chembl/target/inspect/CHEMBL613740) (EMBL-EBI. ChEMBL). The AQVN/EIIP descriptor values were calculated for the learning set (Figure 1) and range for selection was based on their distribution. AQVN descriptor values were in range 2.21–2.32 for WT and 2.21–2.44 for S31N mutant. More than 80% of the compounds of WT inhibitors and 83% M2 S31N mutant ion channel inhibitors from the learning set were inside the common active domain for both while having AQVN and EIIP values within the intervals of (2.21–2.32) and (0.071–0.089). Inside this common active domain is also amantadine with AQVN/EIIP 2.214/0.0717. The reported domain was selected as a criterion for the selection of compounds representing candidate dual M2 WT and S31 mutant ion channel inhibitors (Figure 1). By applying the EIIP/AQVN-based virtual screening criterion, 39 drugs were chosen (Table 1) out of 2,627 approved drugs from the DrugBank (http://www.drugbank.ca) (Wishart et al., 2006).
Figure 1
Table 1
| Drugbank accession number | Name | Chemical formula | AQVN | EIIP |
|---|---|---|---|---|
| DB00915 | Amantadine | C10H17N | 2.214286 | 0.071739 |
| DB06689 | Ethanolamine Oleate | C18H34O2.C2H7NO | 2.215385 | 0.071958 |
| DB00153 | Ergocalciferol | C28H44O | 2.219178 | 0.072708 |
| DB00898 | Ethanol | C2H6O | 2.222222 | 0.073303 |
| DB01105 | Sibutramine | C17H26ClN | 2.222222 | 0.073303 |
| DB01158 | Bretylium | C11H17BrN | 2.233333 | 0.075425 |
| DB00804 | Dicyclomine | C19H35NO2 | 2.245614 | 0.077673 |
| DB00146 | Calcidiol | C27H44O2 | 2.246575 | 0.077845 |
| DB01436 | Alfacalcidol | C27H44O2 | 2.246575 | 0.077845 |
| DB00154 | Dihomo-Î3-linolenic acid | C20H34O2 | 2.25 | 0.078451 |
| DB00592 | Piperazine | C4H10N2 | 2.25 | 0.078451 |
| DB01191 | Dexfenfluramine | C12H16F3N | 2.25 | 0.078451 |
| DB01431 | Allylestrenol | C21H32O | 2.259259 | 0.080048 |
| DB00375 | Colestipol | C8H23N5.C3H5ClO | 2.26087 | 0.080319 |
| DB00330 | Ethambutol | C10H24N2O2 | 2.263158 | 0.080701 |
| DB00162 | Vitamin A | C20H30O | 2.27451 | 0.082539 |
| DB01365 | Mephentermine | C11H17N | 2.275862 | 0.082751 |
| DB01170 | Guanethidine | C10H22N4 | 2.277778 | 0.08305 |
| DB00132 | Alpha-Linolenic Acid | C18H30O2 | 2.28 | 0.083392 |
| DB06809 | Plerixafor | C28H54N8 | 2.288889 | 0.084723 |
| DB08868 | Fingolimod | C19H33NO2 | 2.290909 | 0.085017 |
| DB00858 | Drostanolone | C20H32O2 | 2.296296 | 0.085784 |
| DB00136 | Calcitriol | C27H44O3 | 2.297297 | 0.085924 |
| DB00910 | Paricalcitol | C27H44O3 | 2.297297 | 0.085924 |
| DB00376 | Trihexyphenidyl | C20H31NO | 2.301887 | 0.086554 |
| DB01022 | Phylloquinone | C31H46O2 | 2.303797 | 0.086811 |
| DB00191 | Phentermine | C10H15N | 2.307692 | 0.087326 |
| DB00313 | Valproic Acid | C8H16O2 | 2.307692 | 0.087326 |
| DB01577 | Methamphetamine | C10H15N | 2.307692 | 0.087326 |
| DB06204 | Tapentadol | C14H23NO | 2.307692 | 0.087326 |
| DB06709 | Methacholine | C8H18NO2 | 2.310345 | 0.087669 |
| DB08887 | Icosapent ethyl | C22H34O2 | 2.310345 | 0.087669 |
| DB01187 | Iophendylate | C19H29IO2 | 2.313725 | 0.088098 |
| DB01337 | Pancuronium | C35H60N2O4 | 2.316832 | 0.088483 |
| DB00947 | Fulvestrant | C32H47F5O3S | 2.318182 | 0.088648 |
| DB01083 | Orlistat | C29H53NO5 | 2.318182 | 0.088648 |
| DB00387 | Procyclidine | C19H29NO | 2.32 | 0.088868 |
| DB00942 | Cycrimine | C19H29NO | 2.32 | 0.088868 |
| DB08804 | Nandrolone decanoate | C28H44O3 | 2.32 | 0.088868 |
Approved drugs screened for candidate anti-M2 inhibitors.
All 39 selected drugs were imported in Pentacle software, protonated at pH 7.4, and aligned toward principal moment of inertia. In ligand based virtual screening, we used centroid distance method as criteria for similarity between learning set and candidate compounds. Top 5 candidates from DrugBank selection are presented in Table 2.
Table 2
| Drugbank ID | Name | Similarity distance from centroid | Structure | AQVN | EIIP |
|---|---|---|---|---|---|
| DB01170 | Guanethidine | 1.3446 | 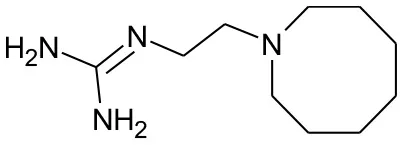 | 2.277778 | 0.08305 |
| DB00191 | Phentermine | 1.4234 | 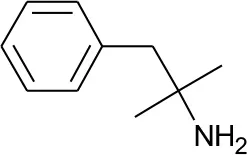 | 2.307692 | 0.087326 |
| DB01577 | Methamphetamine | 1.4334 | 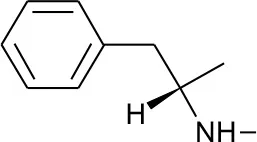 | 2.307692 | 0.087326 |
| DB01191 | Dexfenfluramine | 1.5377 | 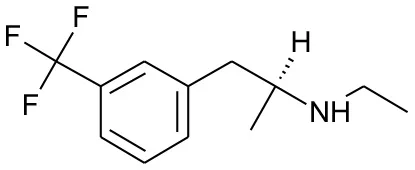 | 2.25 | 0.078451 |
| DB00942 | Cycrimine | 1.6057 | 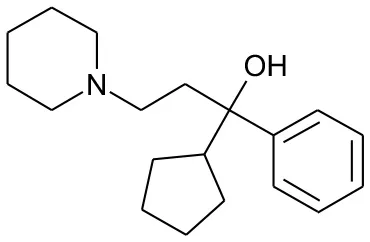 | 2.32 | 0.088868 |
Five best candidates from virtual screening, with Drugbank ID, Similarity distance, structure, and EIIP descriptor values.
We further carried out molecular docking of five candidates to both the wild type M2 channel and S31N mutant channel. The docking energies obtained are presented in Table 3. The candidate with the lowest binding energy and equal affinity to both WT channel and S31N mutant channel was cycrimine, with docking energy −8.3 kcal/mol. Docked conformations of cycrimine are presented on Figures 1, 2. In both WT and S31N mutant of M2 channel, cycrimine conserves corresponding intermolecular receptor-ligand interactions, Ala 30 and Ser 31 in case of WT and Asn 31 in the case of the S31N mutant. As presented on Figures 2, 3, the orientation of cycrimine, compared to crystal coordinates of amandatine and M2WJ332 show cycrimine's ability to form hydrogen bond interactions with Ser 31 i.e., Asn 31, while keeping hydrophobic interactions with Ala 30. This could be a possible explanation as to why cycrimine shows relatively high and equal affinity to both WT and S31N mutant M2 channels (−8.3 kcal/mol, or 800 nM). Other compounds show similar binding patterns as cycrimine; however, in most other cases the affinity ratio is in favor of the WT M2 channel protein.
Table 3
| Drugbank ID | Name | Docking energy on WT M2 channel (kcal/mol) | Docking energy on S31N mutant M2 channel (kcal/mol) | Affinity ratio WT:S31N* |
|---|---|---|---|---|
| DB00942 | Cycrimine | −8.3 | −8.3 | 1 |
| DB01191 | Dexfenfluramine | −6.3 | −6.0 | 0.6 |
| DB01170 | Guanethidine | −5.9 | −5.7 | 0.71 |
| DB00191 | Phentermine | −5.3 | −4.8 | 0.43 |
| DB01577 | Methamphetamine | −4.8 | −4.5 | 0.6 |
Docking energies of five best candidates from virtual screening, with Drugbank ID, Docking energies, and affinity ratio.
.
Figure 2
Figure 3
In order to validate our in silico screening approach, we examined the antiviral activity of guanethidine, the top candidate from the DrugBank selection (Table 2), in vitro. As a positive control, influenza virus was premixed with 10 uM of merimepodib, an IMPDH inhibitor with known antiviral activity against a variety of viruses including influenza (Markland et al., 2000; Tong et al., 2018). Addition of guanethidine to cells infected with 2009 H1N1 pandemic influenza virus resulted in significantly lower viral titers in a dose-dependent manner. Both 100 and 10 μM guanethidine treatment resulted in significant reductions in viral titers at day 1 post-infection, with 100 μM of guanethidine producing a 1–2 log reduction in viral titers.
Discussion
Current prevention and treatment options for influenza A and B infections are insufficient due to increased clinical use of licensed antivirals leading to the emergence of resistant viral strains (Hayden and de Jong, 2011). In a quest for new preventive and therapeutic options to minimize drug resistance and threats of outbreaks of pandemic viruses, the main obstacle is the fact that drug development is an expensive, time-consuming, and risky enterprise. Therefore, drug repurposing represents a promising therapeutic strategy for many viral diseases including anti-influenza A and B treatment. Various predictive computational approaches have been developed to identify drug repositioning opportunities against influenza viruses (Sencanski et al., 2015). Previously, the EIIP/AQVN criterion has been proven to be an efficient filter in virtual screening of molecular libraries for candidate inhibitors of HIV and Ebola virus infection (Tintori et al., 2007; Veljkovic et al., 2015a,b). Using this approach, ibuprofen was selected as an inhibitor of the Ebola virus infection, and this prediction was later experimentally confirmed (Zhao et al., 2016; Paessler et al., 2018.)
To select drug candidates for M2 inhibitors, the virtual screening protocol in our study was based on the application of successive filters. The previous study of EIIP/AQVN distribution of compounds from the PubChem database (http://www.ncbi.nlm.nih.gov/pccompound) showed that 92.5% compounds from PubChem are homogenously distributed inside EIIP and AQVN intervals (0.00–0.11 Ry) and (2.4–3.3), respectively (Veljkovic et al., 2011). The domain that encompasses the majority of known chemical compounds was designated as a “basic EIIP/AQVN chemical space” (BCS). Results of the application of VS based on the EIIP/AQVN filter in this study showed that the active group of candidate M2 inhibitors is very specific, belonging to the sparse cluster of compounds that are out of BCS. This finding indicates that testing only a minor fraction of the compounds from the active EIIP/AQVN domain has a greater chance to inhibit M2 function than compounds with any other EIIP/AQVN values. The proposed AQVN/EIIP interval for the selection of dual M2 inhibitor candidates encompasses only 3% of all chemical molecules. It is therefore not surprising that the previous results from the high-throughput screening had a 10- to 100-fold lower hit rate compared to screens for other targets (Balgi et al., 2013). These results confirm that the M2 is a challenging target for selective inhibition and drug development. In the course of further analysis in our study, the next following two filters were applied in selection of candidates with dual inhibition against M2 WT and M2 S31N mutant protein. First, by applying ligand based virtual screening, the candidates were selected using lowest distance from centroid in the PCA model. This model was based on variables constructed from MIF descriptors of compounds from the learning set. Therefore, their pharmacophore similarity was criteria for the selection. Structure-based approach, as the next step, allows the docking of selected compounds from the first step into both crystal structures of M2 WT and S31N mutant proteins. In this step, the output docking energies (binding free energies) were used as criteria to rank the candidates. In order to address candidate compounds in more detail, the affinity ratio was also calculated. Two of the best candidates are presented in Figures 2, 3, and in Tables 2, 3. The biological significance of docking energy is to select the best candidate that targets both the WT and S31N mutant form of M2. i.e., a dual target candidate. We carried out docking energy comparisons for all candidates, and calculated their ratio with prior conversion to Ki values (docking energy has logarithmic dependency of Ki). Ratio of equilibrium constants between two equilibrium systems (in our case, WT and S31N mutant M2 receptors) that share same ligand in the same environment gives information about that ligand's preference to a certain receptor type. The closer the value of Ki ratio is to 1, the more the ligand is prone to target both receptor types equally and therefore, the ligand is a better dual target candidate. It should also be emphasized that it was assumed that the selected dual inhibitors targeting both M2 proteins might have advantages over mono inhibitors. This is reflected in a higher genetic barrier that enables dual inhibitors to preserve activity if the mutant reverts back to WT sequence (Ma et al., 2016; Wang, 2016).
We selected guanethidine as the best ranked compound from ligand-based virtual screening for further experimental validation. In the experiments with influenza A/CA/07/2009 (H1N1) it was shown that guanethidine inhibits influenza virus production (Figure 4) in a dose-dependent manner.
Figure 4
Another of the best ranked inhibitors from our computational study, methamphetamine, has actually documented good inhibitory activity against influenza A (Chen et al., 2012). It was previously demonstrated that methamphetamine inhibits influenza A virus replication in vitro primarily via acting at the viral replication stage in which M2 plays a major role. Another drug among the best ranked candidates for repurposing against M2 is cycrimine, a drug used to reduce levels of acetylcholine to balance levels of dopamine in the treatment of Parkinson's disease (Kafer and Poch, 1957; Fahn, 2015). Interestingly, the anti-influenza drug amantadine, previously repurposed for treatment of Parkinson's disease, also causes anticholinergic-like side effects (Horstink et al., 2006). In addition, as amantadine and cycrimine are in the same EIIP/AQVN domain, it can be expected from previous studies that they share same therapeutic targets. Other drugs selected as potential M2 inhibitors are ergocalciferol, calcidiol, alfacalcidol. This result is very interesting as vitamin D metabolites were previously connected to potential anti-influenza activity (Gruber-Bzura, 2018). Another FDA approved drug with documented anti-influenza properties, Alpha-Linolenic Acid, was also selected as a potential M2 inhibitor in our study, indicating the usefulness of the proposed screen (Bai et al., 2012).
In conclusion, the results presented here suggest that guanethidine represents a promising molecular template for further development of drugs against influenza virus. Other selected drugs from our computational study present valuable starting points for further experimental investigations in a quest for safe, new treatments for human and animal influenza infections.
Statements
Data availability statement
All datasets generated for this study are included in the manuscript and/or the Supplementary Files.
Author contributions
SP, SG, DR, and VV conceived and designed the study. VP developed the analysis tools. DR, SG, MS, VV, NV, JP, EM, and NB analyzed the data. EM and NB performed the experiments. DR, MS, VP, NV, JP, VV, EM, NB, SP, and SG drafted the work. SG, DR, VV, and SP wrote the paper. DR, MS, VP, NV, JP, VV, EM, NB, SP, and SG agreed on the final approval of the manuscript to be published and agreed to be accountable for all aspects of the work.
Acknowledgments
This work was supported by the Ministry of Education, Science and Technological Development of the Republic of Serbia (Grant no. 173001).
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fcimb.2019.00067/full#supplementary-material
Supplementary Table 1The learning set (M2 WT).
Supplementary Table 2The learning set (M2 S31N).
References
1
BaiS. W.ChenC. Y.JiJ.XieQ. M.MaY.SunB. L.et al. (2012). Inhibition effect of alpha-lipoic acid on the propagation of influenza a virus in MDCK cells. Pak. Vet. J. 32, 101–106.
2
BalgiA. D.WangJ.ChengD. Y.MaC.PfeiferT. A.ShimizuY.et al. (2013). Inhibitors of the influenza A virus M2 proton channel discovered using a high-throughput yeast growth restoration assay. PLoS ONE8:e55271. 10.1371/journal.pone.0055271
3
BridgesC.PeasahS.MeltzerM. (2013). The control of influenza and cost effectiveness of interventions, in Influenza Textbook, eds WebsterR. G.MontoA. S.BracialeT. J.LambR. A. (Hoboken, NY: Wiley-Blackwell), 419–433.
4
CadyS. D.Schmidt-RohrK.WangJ.SotoC. S.DegradoW. F.HongM. (2010). Structure of the amantadine binding site of influenza M2 proton channels in lipid bilayers. Nature463, 689–692. 10.1038/nature08722
5
CDC (2006). High levels of adamantane resistance among influenza A (H3N2) viruses and interim guidelines for use of antiviral agents-United States, 2005-06 influenza season. MMWR Morb. Mortal. Wkly. 55, 44–46.
6
ChenY. H.WuK. L.ChenC. H. (2012). Methamphetamine reduces human influenza A virus replication. PLoS ONE7:e48335. 10.1371/journal.pone.0048335
7
DolinR.ReichmanR. C.MadoreH. P.MaynardR.LintonP. N.Webber-JonesJ. (1982). A controlled trial of amantadine and rimantadine in the prophylaxis of influenza A infection. N. Engl. J. Med. 307, 580–584. 10.1056/NEJM198209023071002
8
DongG.PengC.LuoJ.WangC.HanL.WuB.et al. (2015). Adamantane-resistant influenza A viruses in the World (1902–2013): frequency and distribution of M2 gene mutations. PLoS ONE10:e0119115. 10.1371/journal.pone.0119115
9
DuranA.ComesañaG.PastorM. (2008). Development and validation of AMANDA, a new algorithm for selecting highly relevant regions in molecular interaction fields. J. Chem. Inf. Model. 48, 1813–1823. 10.1021/ci800037t
10
DuranA.ZamoraI.PastorM. (2009). Suitability of GRIND-based principal properties for the description of molecular similarity and ligand-based virtual screening. J. Chem. Inf. Model. 49, 2129–2138. 10.1021/ci900228x
11
FahnS.. The medical treatment of Parkinson disease from James Parkinson to George Cotzias. (2015). Mov. Disord. 30, 4–18. 10.1002/mds.26102
12
Gruber-BzuraB. M. (2018). Vitamin D and influenza-prevention or therapy?Int. J. Mol. Sci. 16:19. 10.3390/ijms19082419
13
HaydenF. G.de JongM. D. (2011). Emerging influenza antiviral resistance threats. J. Infect. Dis. 203, 6–10. 10.1093/infdis/jiq012
14
HongM.DeGradoW. F. (2012). Structural basis for proton conduction and inhibition by the influenza M2 protein. Protein Sci.21, 1620–1633. 10.1002/pro.2158
15
HorstinkM.TolosaE.BonuccelliU.DeuschlG.FriedmanA.KanovskyP.et al. (2006). European Federation of Neurological Societies; Movement Disorder Society-European Section. Review of the therapeutic management of Parkinson's disease. Report of a joint task force of the European Federation of Neurological Societies and the Movement Disorder Society-European Section. Part I: early (uncomplicated) Parkinson's disease. Eur. J. Neurol. 13, 1170–1185. 10.1111/j.1468-1331.2006.01547.x
16
HurtA.KomadinaN.DengY. M.KayeM.SullivanS.SubbaraoK.et al. (2017). Detection of adamantane-sensitive influenza A(H3N2) viruses in Australia, 2017: a cause for hope?Euro Surveill. 22:17-00731. 10.2807/1560-7917.ES.2017.22.47.17-00731
17
HurtA. C.BesselaarT. G.DanielsR. S.ErmetalB.FryA.GubarevaL.et al. (2016). Global update on the susceptibility of human influenza viruses to neuraminidase inhibitors, 2014–2015. Antiviral Res.132, 178–185. 10.1016/j.antiviral.2016.06.001
18
HurtA. C.ErnestJ.DengY. M.IannelloP.BesselaarT. G.BirchC.et al. (2009). Emergence and spread of oseltamivir-resistant A (H1N1) influenza viruses in Oceania, South East Asia and South Africa. Antiviral Res.83, 90–93. 10.1016/j.antiviral.2009.03.003
19
HurtA. C.HardieK.WilsonN. J.DengY. M.OsbournM.GehrigN.et al. (2011). Community transmission of oseltamivir-resistant A (H1N1) pdm09 influenza. N. Engl. J. Med.365, 2541–2542. 10.1056/NEJMc1111078
20
IsonM. G. (2017). Antiviral treatments. Clin. Chest Med. 38, 139–153. 10.1016/j.ccm.2016.11.008
21
KaferJ. P.PochG. F. (1957). Cycrimine, a new drug in the treatment of Parkinson's disease and Parkinsonism. Prensa Med. Argent44, 1071–1075.
22
LambR. A.ZebedeeS. L.RichardsonC. D. (1985). Influenza virus M2 protein is an integral membrane protein expressed on the infected-cell surface. Cell40, 627–633. 10.1016/0092-8674(85)90211-9
23
MaC.ZhangJ.WangJ. (2016). Pharmacological characterization of the spectrum of antiviral activity and genetic barrier to drug resistance of M2-S31N channel blockers. Mol. Pharmacol. 90, 188–198. 10.1124/mol.116.105346
24
MarklandW.McQuaidT. J.JainJ.KwongA. D. (2000). Broad-spectrum antiviral activity of the IMP dehydrogenase inhibitor VX-497: a comparison with ribavirin and demonstration of antiviral additivity with alpha interferon. Antimicrob. Agents Chemother. 44, 859–866. 10.1128/AAC.44.4.859-866.2000
25
MercorelliB.PalùG.LoregianA. (2018). Drug repurposing for viral infectious diseases: how far are we?Trends Microbiol. 26, 865–876. 10.1016/j.tim.2018.04.004
26
MorrisG. M.HueyR.LindstromW.SannerM. F.BelewR. K.GoodsellD. S.et al. (2009). Autodock4 and AutoDockTools4: automated docking with selective receptor flexiblity. J. Comput. Chem. 16, 2785–2791. 10.1002/jcc.21256
27
MouldJ. A.PatersonR. G.TakedaM.OhigashiY.VenkataramanP.LambR. A.et al. (2003). Influenza B virus BM2 protein has ion channel activity that conducts protons across membranes. Dev. Cell5, 175–184. 10.1016/S1534-5807(03)00190-4
28
PaesslerS.HuangC.SencanskiM.VeljkovicN.PerovicV.GlisicS.et al. (2018). Ibuprofen as a template molecule for drug design against Ebola virus. Front. Biosci.23, 947–953. 10.2741/4627
29
PastorM.McLayI.PickettS.ClementiS. (2000). Grid-Independent descriptors (GRIND): a novel class of alignment-independent three-dimensional molecular descriptors. Med. Chem. 43, 3233–3243. 10.1021/jm000941m
30
SakaguchiT.TuQ.PintoL. H.LambR. A. (1997). The active oligomeric state of the minimalistic influenza virus M2 ion channel is a tetramer. Proc. Natl. Acad. Sci. U.S.A.94, 5000–5005. 10.1073/pnas.94.10.5000
31
SannerM. F. (1999). Python: a programming language for software integration and development. J. Mol. Graphics Mod. 1, 57–61.
32
SencanskiM.RadosevicD.PerovicV.GemovicB.StanojevicM.VeljkovicN.et al. (2015). Natural products as promising therapeutics for treatment of influenza disease. Curr. Pharm. Des. 21, 5573–5588. 10.2174/1381612821666151002113426
33
SimonsenL.ViboudC.GrenfellB. T.DushoffJ.JenningsL.SmitM.et al. (2007). The genesis and spread of reassortment human influenza A/H3N2 viruses conferring adamantane resistance. Mol. Biol. Evol. 24, 1811–1820. 10.1093/molbev/msm103
34
StewartJ. J. P. (2013). Optimization of parameters for semiempirical methods VI: more modifications to the NDDO approximations and re-optimization of parameters. J. Mol. Mod.19, 1–32. 10.1007/s00894-012-1667-x
35
StewartJ. J. P. (2016). MOPAC2016. Stewart Computational Chemistry. Colorado Springs, CO. Availaible online at: http://OpenMOPAC.net.
36
StrittmatterS. M. (2014). Overcoming drug development bottlenecks with repurposing: old drugs learn new tricks. Nat. Med. 20, 590–591. 10.1038/nm.3595
37
TakedaM.PekoszA.ShuckK.PintoL. H.LambR. A. (2002). Influenza a virus M2 ion channel activity is essential for efficient replication in tissue culture. J. Virol. 76, 1391–1399. 10.1128/JVI.76.3.1391-1399.2002
38
TintoriC.ManettiF.VeljkovicN.PerovicV.VercammenJ.HayesS.et al. (2007). Novel virtual screening protocol based on the combined use of molecular modeling and electron-ion interaction potential techniques to design HIV-1 integrase inhibitors. J. Chem. Inf. Model. 47,1536–1544. 10.1021/ci700078n
39
TongX.SmithJ.BukreyevaN.KomaT.ManningJ. T.KalkeriR.et al. (2018). Merimepodib, an IMPDH inhibitor, suppresses replication of Zika virus and other emerging viral pathogens. Antiviral Res. 149, 34–40. 10.1016/j.antiviral.2017.11.004
40
TrottO.OlsonA. J. (2010). AutoDock Vina: improving the speed and accuracy of docking with a new scoring function, efficient optimization and multithreading. J. Comput. Chem. 31, 455–461. 10.1002/jcc.21334
41
VeljkovicN.GlisicS.PerovicV.VeljkovicV. (2011). The role of long-range intermolecular interactions in discovery of new drugs. Exp. Opin. Drug Disc. 6, 1263–1270. 10.1517/17460441.2012.638280
42
VeljkovicN.GlisicS.PrljicJ.PerovicV.VeljkovicV. (2013). Simple and general criterion for “in silico” screening of candidate HIV drugs. Curr. Pharm. Biotechnol.14, 561–569. 10.2174/138920101405131111105301
43
VeljkovicV.GoeijenbierM.GlisicS.VeljkovicN.PerovicV.SencanskiM.et al. (2015a). In silico analysis suggests repurposing of ibuprofen for prevention and treatment of EBOLA virus disease. F1000Res. 4:104. 10.12688/f1000research.6436.1
44
VeljkovicV.LoiseauP. M.FigadereB.GlisicS.VeljkovicN.PerovicV.et al. (2015b). Virtual screen for repurposing approved and experimental drugs for candidate inhibitors of EBOLA virus infection. F1000Res. 4:34. 10.12688/f1000research.6110.1
45
WangJ. (2016). M2 as a target to combat influenza drug resistance: what does the evidence say?Future Virol. 11, 1–4. 10.2217/fvl.15.95
46
WangJ.WuY.MaC.FiorinG.WangJ.PintoL. H.et al. (2013). Structure and inhibition of the drug-resistant S31N mutant of the M2 ion channel of influenza A virus. Proc. Natl. Acad. Sci. U.S.A. 110, 1315–1320. 10.1073/pnas.1216526110
47
WHO Influenza (2018). Factsheet; March. Available online at: http://www.who.int/en/news-room/fact-sheets/detail/influenza-(seasonal) (Accessed August 25, 2018).
48
WishartD. S.KnoxC.GuoA. CShrivastavaS.HassanaliM.StothardP.et al. (2006). DrugBank: a comprehensive resource for in silico drug discovery and exploration. Nucleic Acids Res.34(Database issue):D668–D672. 10.1093/nar/gkj067
49
ZhaoY.RenJ.HarlosK.JonesD. M.ZeltinaA.BowdenT. A.et al. (2016). Toremifene interacts with and destabilizes the Ebola virus glycoprotein. Nature535, 169–172. 10.1038/nature18615
50
ZhengW.SunW.SimeonovA. (2018). Drug repurposing screens and synergistic drug-combinations for infectious diseases. Br. J. Pharmacol. 175,181–191. 10.1111/bph.13895
Summary
Keywords
influenza A, IAV matrix protein 2, drug repurposing, virtual screening, drug resistance
Citation
Radosevic D, Sencanski M, Perovic V, Veljkovic N, Prljic J, Veljkovic V, Mantlo E, Bukreyeva N, Paessler S and Glisic S (2019) Virtual Screen for Repurposing of Drugs for Candidate Influenza a M2 Ion-Channel Inhibitors. Front. Cell. Infect. Microbiol. 9:67. doi: 10.3389/fcimb.2019.00067
Received
24 October 2018
Accepted
04 March 2019
Published
26 March 2019
Volume
9 - 2019
Edited by
Rodolfo García-Contreras, National Autonomous University of Mexico, Mexico
Reviewed by
Alan G. Goodman, Washington State University, United States; Richard Yuqi Zhao, University of Maryland, School of Medicine, United States
Updates
Copyright
© 2019 Radosevic, Sencanski, Perovic, Veljkovic, Prljic, Veljkovic, Mantlo, Bukreyeva, Paessler and Glisic.
This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Slobodan Paessler slpaessl@utmb.edu
This article was submitted to Clinical Microbiology, a section of the journal Frontiers in Cellular and Infection Microbiology
Disclaimer
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.