 A. Amir Hassan
A. Amir Hassan Carla P. Coutinho
Carla P. Coutinho Isabel Sá-Correia
Isabel Sá-Correia- 1iBB - Institute for Bioengineering, Instituto Superior Técnico, Universidade de Lisboa, Lisbon, Portugal
- 2Department of Bioengineering, Instituto Superior Técnico, Universidade de Lisboa, Lisbon, Portugal
Burkholderia cepacia complex (Bcc) bacteria can adapt to the lung environment of cystic fibrosis (CF) patients resulting in the emergence of a very difficult to eradicate heterogeneous population leading to chronic infections associated with rapid lung function loss and increased mortality. Among the important phenotypic modifications is the variation of the lipopolysaccharide (LPS) structure at level of the O-antigen (OAg) presence, influencing adherence, colonization and the ability to evade the host defense mechanisms. The present study was performed to understand whether the loss of OAg expression during CF infection can be considered a general phenomenon in different Bcc species favoring its chronicity. In fact, it is still not clear why different Bcc species/strains differ in their ability to persist in the CF lung and pathogenic potential. The systematic two-decade-retrospective-longitudinal-screening conducted covered 357 isolates retrieved from 19 chronically infected patients receiving care at a central hospital in Lisbon. The study involved 21 Bcc strains of six/seven Bcc species/lineages, frequently or rarely isolated from CF patients worldwide. Different strains/clonal variants obtained during infection gave rise to characteristic OAg-banding patterns. The two most prevalent and feared species, B. cenocepacia and B. multivorans, showed a tendency to lose the OAg along chronic infection. B. cenocepacia recA lineage IIIA strains known to lead to particularly destructive infections exhibit the most frequent OAg loss, compared with lineage IIIB. The switch frequency increased with the duration of infection and the level of lung function deterioration. For the first time, it is shown that the rarely found B. cepacia and B. contaminans, whose representation in the cohort of patients examined is abnormally high, keep the OAg even during 10- or 15-year infections. Data from co-infections with different Bcc species reinforced these conclusions. Concerning the two other rarely found species examined, B. stabilis exhibited a stable OAg expression phenotype over the infection period while for the single clone of the more distantly related B. dolosa species, the OAg-chain was absent from the beginning of the 5.5-year infection until the patient dead. This work reinforces the relevance attributed to the OAg-expression switch suggesting marked differences in the various Bcc species.
Introduction
Chronic respiratory infections involving opportunistic pathogens remain the leading cause of premature death of cystic fibrosis (CF) patients (Cullen and McClean, 2015), in particular the long-term infections caused by Pseudomonas aeruginosa and bacteria of the Burkholderia cepacia complex (Bcc). The Bcc is a group of more than 20 closely-related species (Depoorter et al., 2016) particularly feared by CF patients due to their very difficult eradication, inter-patient transmission, and related increased morbidity and decreased life expectancy (Mahenthiralingam et al., 2005; Drevinek and Mahenthiralingam, 2010; Kenna et al., 2017). The majority of CF patients who acquire Bcc develops a chronic infection while a minority may clear the infection spontaneously (Mahenthiralingam et al., 2005). Pulmonary colonization with Bcc can cause an accelerated decline in lung function and the “cepacia syndrome” that is characterized by an uncontrolled deterioration with septicaemia and necrotizing pneumonia that usually results in early death (Mahenthiralingam et al., 2005). Inside the CF lung, Bcc bacteria face stressful and changing environmental conditions as a consequence of host immune defenses, inflammatory responses, antimicrobials and reactive oxygen species (ROS), fluctuating levels of nutrients, oxygen deprivation, high osmolarity, low pH, biofilm growth, and the presence of other co-infecting microbes (Hogardt and Heesemann, 2010; Döring et al., 2011; Cullen and McClean, 2015). Consequently, during long-term infection, the initial Bcc strain-population accumulates genetic alterations leading to genotypic and phenotypic diversification forming a heterogeneous bacterial community very difficult to eradicate therapeutically (Lieberman et al., 2011, 2014; Madeira et al., 2011, 2013; Silva et al., 2011, 2016; Moreira et al., 2014; Nunvar et al., 2017). Such within-patient emergence of multiple clonal variants was first described for the more prevalent CF pathogen Pseudomonas aeruginosa (Markussen et al., 2014; Winstanley et al., 2016) and proposed to provide a pool of mutations affecting virulence and antimicrobial resistance (Lorè et al., 2012; Marvig et al., 2013; Faure et al., 2018). Several studies have shown that, during chronic infection, alterations in the lipopolysaccharide (LPS) molecule, a complex glycolipid covering the cell surface and shielding Gram-negative bacteria from adverse host environments, may occur (Pier, 2007; Maldonado et al., 2016; Faure et al., 2018). The LPS is one of the most important components of pathogenic Gram-negative bacteria cellular envelope being considered a major virulence factor and contributing to adhesion, immune defenses evasion, host colonization and adaptation to the infection niche (Vinion-Dubiel and Goldberg, 2003; Pier, 2007; Maldonado et al., 2016). The LPS structure is composed by three constituents: a highly acylated lipid A, also known as endotoxin, covalently linked to the central core oligosaccharide, and the O-antigen (OAg), composed by repeating polysaccharide units of variable length (Vinion-Dubiel and Goldberg, 2003). Most of the heterogeneity in LPS molecules is found in the OAg polysaccharide and lipid-A components (Pier, 2007; Maldonado et al., 2016; Faure et al., 2018). The OAg component of the LPS is essential in several pathogens for motility (swarming and twitching) (Toguchi et al., 2000; Berry et al., 2009; Bowden et al., 2013), protection against oxidative stress (Berry et al., 2009) and evasion from host immune defenses (Murray et al., 2003; Saldias et al., 2009; Kotrange et al., 2011; Kintz et al., 2017). LPS OAg is believed to be an immunodominant molecule that can modulate host-pathogen interaction (Ranf, 2016) and proposed to be under selective pressure in Gram-negative bacteria (King et al., 2009; Maldonado et al., 2016; Kintz et al., 2017). Longitudinal studies of P. aeruginosa isolates from CF patients with chronic lung infections revealed the conversion from smooth (in the early isolates) to rough LPS with short or no OAg side chain at the late-stage of infections rendering the bacteria non-typable and less immunogenic (King et al., 2009; Maldonado et al., 2016; Demirdjian et al., 2017). In Bcc bacteria, a few recent reports also described the variation of the presence and/or modification of the OAg during persistent infection of the CF lung and on how this alteration may affect Bcc pathogenicity (Lieberman et al., 2011; Maldonado et al., 2016; Silva et al., 2016; Hassan et al., 2017). An extensive study on an epidemic outbreak of Burkholderia dolosa during 16 years, involving 14 CF patients and 112 isolates, has shown that several late isolates produce an LPS exhibiting the OAg that was absent in the LPS of the ancestral strain; this fact was related with the appearance of two different mutations in the glycosyltransferase encoding gene wbaD (Lieberman et al., 2011). The genome analysis of clones and metagenomes of evolved B. cenocepacia biofilms, after more than one thousand generations, unveiled a mutation in the gene manC – a gene involved in mannose metabolism located within the LPS biosynthesis gene cluster-resulting in the loss of the LPS OAg (Traverse et al., 2013). Remarkably, mutations in the same manC gene were previously found to disrupt OAg biosynthesis in late B. cenocepacia (Saldias et al., 2009) or early B. dolosa (Lieberman et al., 2011) isolates retrieved during chronic infection. A comparative genomic analysis focused on sequential B. multivorans isolates obtained over 20 years of CF chronic infection showed that late isolates accumulate three different mutations in a locus homologous to the wbi gene cluster, involved in LPS OAg biosynthesis, leading to OAg loss (Silva et al., 2016). Another recent study involving eleven serial clonal variants of B. cenocepacia recA lineage IIIA obtained from a CF patient over a 3.5 year period, since the onset of infection until death with the cepacia syndrome, revealed that the OAg was only present in the early isolate but not in any of the late variants (Hassan et al., 2017). Genomic sequencing of these serial isolates enabled the identification of mutations, also within the OAg cluster, but none of these mutations could definitively be associated to OAg loss (Hassan et al., 2017).
Although the Bcc comprises 24 bacterial species (De Smet et al., 2015; Depoorter et al., 2016; Ong et al., 2016; Bach et al., 2017; Weber and King, 2017; Martina et al., 2018) and several were found to be involved in CF respiratory infections, only B. cenocepacia (Hassan et al., 2017), B. multivorans (Silva et al., 2016), and B. dolosa (Lieberman et al., 2011) were examined concerning the variation of the OAg presence over chronic infection. B. cenocepacia is recognized as the dominant species with high potential for inter-patient transmission (Drevinek and Mahenthiralingam, 2010), and B. multivorans has recently replaced B. cenocepacia in this first position in several countries (Lipuma, 2010). However, it is recognized that the less represented species may also be associated with poor clinical outcome, but it is not clear why different Bcc species/strains differ in their ability to persist in the CF lung and in their pathogenic potential (Woods et al., 2004; Kalish et al., 2006; Cunha et al., 2007; Moehring et al., 2014; Coutinho et al., 2015; Nunvar et al., 2016; Marquez et al., 2017; Roux et al., 2017). It also remains to be elucidated whether LPS OAg loss confers an advantage to the different Bcc species in particular and to Gram-negative bacteria, in general, over the course of infection.
The present study was performed to understand whether OAg loss in different Bcc species during respiratory infection in CF patients can be considered a general phenomenon that affects immune evasion favoring chronic infection. For this, a systematic retrospective and longitudinal screening was performed based on a collection of isolates, recovered from 1995 to 2016 from 19 CF patients under surveillance at the major Portuguese CF treatment center at Hospital de Santa Maria (HSM), in Lisbon, over the duration of chronic infection (ranging from 1.2 to 15.2 years). The 357 sequential isolates examined were molecularly identified at the species level and genotyped during this study or in previous studies and belong to B. cenocepacia, B. multivorans or to the more rare species in CF population worldwide, B. dolosa, B. stabilis, B. cepacia and B. contaminans (Cunha et al., 2003, 2007; Coutinho et al., 2011, 2015; Moreira et al., 2014). The abnormally high representation of the species B. cepacia and B. contaminans in our collection is considered related with a contamination of saline solutions for nasal application (Cunha et al., 2007; Coutinho et al., 2015). Information on the health condition of several patients at the time of isolation based on the forced expiratory values (FEV), an indicator of pulmonary function and, consequently, of oxygen availability in the patient's lung, is also available (Correia et al., 2008; Moreira et al., 2014). This systematic retrospective study allowed to answer the question of whether the described variation of the LPS OAg expression for a limited number of the Bcc species/strains and patients is extensive to the more rarely found Bcc species and can be considered a general phenomenon in the Bcc during long-term respiratory infections in CF patients.
Materials and Methods
Bacterial Isolates and Growth Conditions
The 357 Burkholderia cepacia complex (Bcc) isolates examined in this study were retrieved from the respiratory secretions of 19 chronically infected cystic fibrosis (CF) patients under surveillance at the major Portuguese CF treatment center at Hospital de Santa Maria (HSM), Centro Hospitalar Lisboa Norte EPE, Lisbon, from 1995 to 2016, during hospital routines. The isolates examined in this retrospective study were selected at random among the colonies isolated in selective Burkholderia cepacia Selectatab medium at the Hospital, at a specific date of isolation. Over the years, a significant part of the isolates were molecularly confirmed to belong to six Bcc species: Burkholderia cenocepacia, Burkholderia multivorans, Burkholderia cepacia, Burkholderia contaminans, Burkholderia dolosa, and Burkholderia stabilis and genotyped (Cunha et al., 2003, 2007; Coutinho et al., 2011, 2015) (Table 1 and Table S1). Bacterial cultures are stored at −80°C in 1:1 (v/v) glycerol. During this study, bacteria were grown in Lysogeny broth agar (LB agar; Conda, Pronadisa) at 37°C with shaking at 250 rpm or in LB agar plates.

Table 1. Schematic representation of the sequential isolates from various species chronically infecting the different CF patients examined in this study.
Species Identification and Genotyping of Bcc Isolates
The isolates examined in this study that had not been previously identified at the species level and genotyped (by ribotyping or recA gene PCR Restriction fragment length polymorphism–recA PCR RFLP–and species-specific recA direct PCR) were identified and genotyped in this study based on recA PCR RFLP analysis and Random Amplified Polymorphic DNA (RAPD) (Mahenthiralingam et al., 1996, 2000). Genomic DNA was extracted from cells of those isolates grown overnight in LB broth with orbital agitation at 37°C, using the Puregene DNA isolation kit (cell and tissue kit, Gentra Systems, Qiagen, Germany). The concentration of the resulting genomic DNA solutions was estimated using a ND-1000 spectrophotometer (NanoDrop Technologies). Species identification was based on the recA PCR RFLP analysis that was performed by amplification of the entire recA gene using specific primers for Bcc bacteria (BCR1-5′TGACCGCCGAGAAGAGCAA3′; BCR2-5′CTCTTCTTCGTCCATCGCCTC3′) (Mahenthiralingam et al., 2000). Bcc recA amplicons were digested with HaeIII (Amersham Biosciences) and the restriction fragments separated by electrophoresis in 2% (w/v) agarose gels. The RFLP patterns obtained were compared with those reported in the literature (Mahenthiralingam et al., 2000; Vermis et al., 2002; Cunha et al., 2003; Coutinho et al., 2015). Strain assignment was performed by RAPD (Mahenthiralingam et al., 1996). Genotyping of the isolates tested was performed using primer 270 (5′TGCGCGCGGG3′), followed by confirmation of the strain types established by this primer with primers 208 (5′ACGGCCGACC3′) and 272 (5′AGCGGGCCAA3′). RAPD profiles were visually compared by GelJ software (Heras et al., 2015). Similarity coefficients were calculated using the RAPD profiles obtained with each primer for the complete set of isolates examined using Pearson coefficient with Arithmetic Mean (UPGMA) linkage method.
LPS Extraction and SDS-PAGE Analysis
LPS was extracted from cell envelopes obtained from bacterial cells as described before (Marolda et al., 1990) with small modifications (Hassan et al., 2017). Briefly, Bcc cells were harvested from overnight liquid cultures by centrifugation for 1 min after OD640nm adjustment to 2.0 in 1 mL of phosphate buffer saline—PBS (at pH 7.4: 1.8 mM/L KH2PO4, 137 mM/L NaCl, 2.7 mM/L KCl and 10 mM/L Na2HPO4). These cells were suspended in 150 μL of lysis buffer, containing 2% SDS, 4% 2-β-mercaptoethanol, and 500 mM Tris-HCl (pH 6.8), and boiled for 10 min. Proteinase K (20 mg/mL) was added, and the samples were incubated at 60°C for 2 h. Finally, samples were mixed with a tracking dye solution (125 mM Tris-HCl [pH 6.8], 2% SDS, 20% [v/v] glycerol, 0.002% bromophenol blue, and 10% mercaptoethanol) and boiled for 5 min before loading the gels. LPS samples were resolved by electrophoresis – at 150 V for about 1 h 40 m – in 14% polyacrylamide gels with a Tricine-SDS system followed by silver staining to visualize the O-antigen (OAg) banding patterns (Marolda et al., 2006).
Ethics
The present retrospective work involves clinical Bcc isolates obtained as a part of the hospital routine and its use in clinical research was approved by the hospital ethics committee. Consent was obtained from the patient or legal representative for the use of these isolates in research. Patients' anonymity is preserved.
Results
During Chronic Infections, B. cenocepacia recA Lineage IIIA and B. multivorans Isolates Were More Prone to Lose the OAg Present in Early Isolates, Compared With B. cenocepacia recA Lineage IIIB
The 357 Burkholderia cepacia complex (Bcc) isolates tested in this study were examined with the goal of systematically compare the O-antigen (OAg) banding patterns when this LPS component is present and to demonstrate the eventual absence of the OAg in late isolates (Figure 1 and Table 1). It was found that the different strains tested and the corresponding clonal variants obtained during long-term-infection, as demonstrated by Multilocus sequence typing (MLST), gave rise to characteristic banding patterns of the OAg whenever this component is present (Figures 2A,B).

Figure 1. Schematic representation of the Bcc isolates examined in this study retrieved from different CF patients over 21 years (from 1995 to 2016) of epidemiological survey at the CF Center of Hospital de Santa Maria. Time zero marks the date of isolation of the first Bcc isolate from a specific patient with essentially a single Bcc species/strain (A) or co-infected with different Bcc species (B). The presence or absence of the LPS O-antigen is represented by closed or open symbols, respectively. The same symbol in a different line is used to indicate a different strain of the same species co-infecting the patient, as detailed in Table 1 and Table S1, as in the case for patients AU, B, H, O, and AF. Whenever available, the Forced Expiratory Values (%) in the first second (FEV1), before and during infection, are provided (Correia et al., 2008; Coutinho et al., 2011; Moreira et al., 2014). These values are an indication of the pulmonary function in the infected patients. * - Sporadic isolation of strains from other Bcc species (in Table S1): B. cenocepacia IIIB - , B. cepacia -
, B. cepacia - , B. multivorans -
, B. multivorans - , and B. dolosa -
, and B. dolosa - .
.
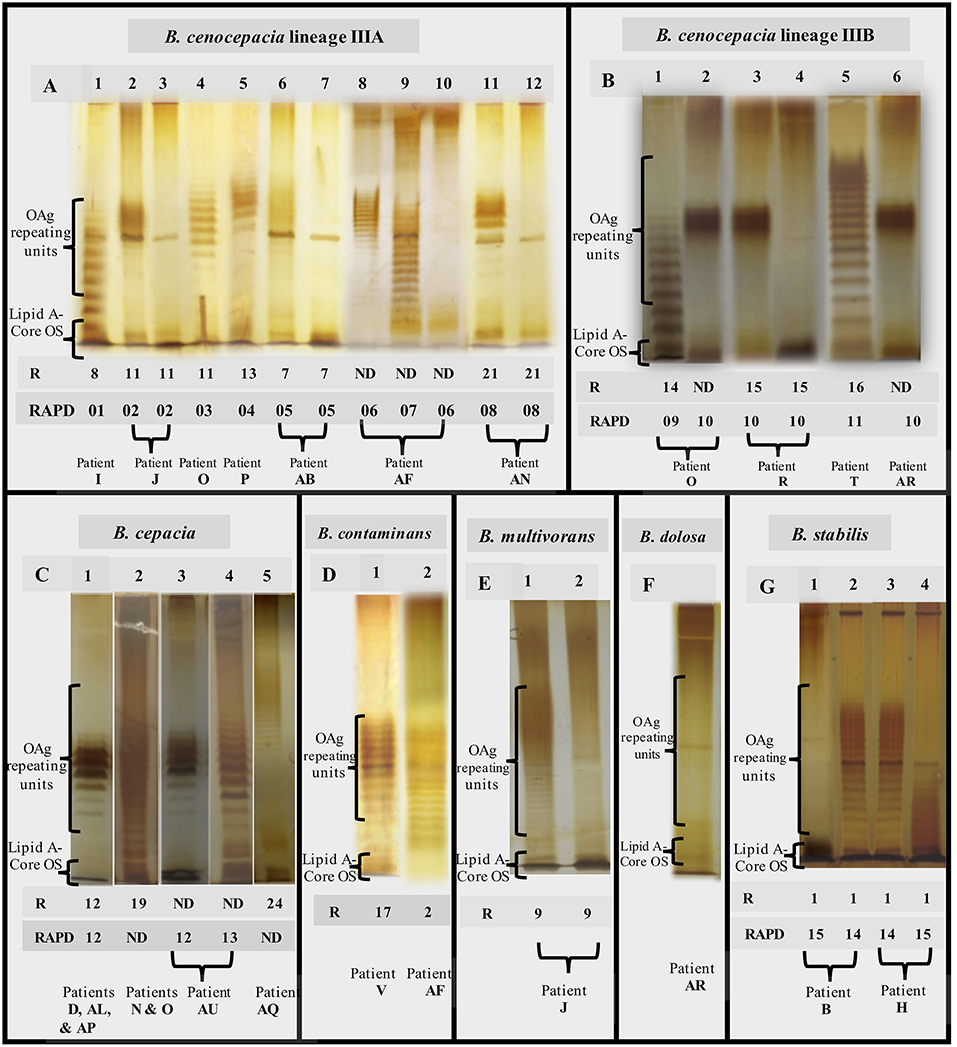
Figure 2. Representative SDS-PAGE gels showing the O-antigen banding patterns obtained (A1–A12, B1–B6, C1–C5, D1, D2, E1, E2, F, G1–G4) for the Bcc isolates examined. Genotyping data for the different strains tested are also shown: the ribopatterns were obtained before (Cunha et al., 2003, 2007; Coutinho et al., 2011) and the RAPD profiles were obtained during this study (Figure S1). R, Isolate ribopattern; RAPD, Isolate profile based on Random amplified polymorphism DNA profiles (Figure S1); ND, not determined. The isolates tested are (Table S1): A1–IST416, A2–IST439, A3–IST4103, A4–IST462, A5–IST432, A6–IST4121, A7–IST4893, A8–IST4240a, A9–IST4272, A10–IST4253a, A11–IST4197, A12–IST4386, B1–IST435, B2–IST4177, B3–IST438, B4–IST4155, B5–IST466, B6–IST4481 IIIB, C1–IST4152, C2–IST4128, C3–IST4283, C4–IST4546, C5–IST4198, D1–IST481, D2–IST4193, E1–IST419, E2–IST453, F–IST4208, G1–IST402, G2–IST409, G3–IST413, G4–IST412.
Concerning the 153 isolates of B. cenocepacia recA lineages IIIA (112 isolates from 7 patients) and IIIB (41 isolates from 3 patients) (Table 1) tested for the presence of OAg polysaccharide during lung infection (timelines shown in Figure 1), it was found that strains from B. cenocepacia lineage IIIA were more prone to lose the OAg than lineage IIIB strains.
There were two cases of extended chronic infection of patients AB and AN by B. cenocepacia lineage IIIA, with a duration of 14.7 and 6.8 years, respectively (Figure 1A and Table 1), where the late isolates retrieved after 4.5 or 2.5 years, respectively, were found to lack the OAg that was present in the early isolates (Figure 1 and Table S1). The loss of the OAg in late isolates was also previously reported for another B. cenocepacia lineage IIIA strain infecting patient J for a shorter period of 3.4 years (Hassan et al., 2017) and (Figure 1 and Table 1). Patient J was co-infected with a B. multivorans strain and the LPS of the majority of late isolates of this B. multivorans strain also lack the OAg present in the early isolates, as shown in the electrophoretic profiles of the extracted LPS (Figure 2, columns E1 and E2). However, one B. multivorans clonal isolate retrieved at the last isolation date exhibited again the OAg, consistent with the pattern observed before by others (Silva et al., 2016). Late isolates of B. cenocepacia and B. multivorans were retrieved from patient J during a period of oxygen depletion in the CF lung that accompanied disease progression and the deterioration of lung function. In fact, according to the hospital records, the forced expiratory values in the first second (FEV1) available are low during co-infection (22–34%) and no further values of FEV1 were registered during the later stages of the patient's life due to the highly severe deterioration of pulmonary function (Figure 1B; Correia et al., 2008).
Differently, the B. cenocepacia lineage IIIA strain that infected patient O for 3 year-infection period kept the LPS OAg and the same was found for the co-infecting Bcc strains of B. cenocepacia lineage IIIB and B. cepacia chronically infecting patient O for 3.4 and 5.6 years, respectively. Remarkably, the FEV1 values available were above 50% during the co-infection period and, after 9 years, the Burkholderia infection was eradicated (Correia et al., 2008; Figure 1, Table S1). When examining the 11 and 10 B. cenocepacia IIIA isolates obtained from patients' J and O, respectively, we were surprised by the fact that the electrophoretic profiles of the extracted LPS obtained by SDS-PAGE analysis and silver-staining produced two different profiles (Figure 2, columns A2 and A4, respectively) even though they were considered to share the same ribopattern 11 (Table S1; Coutinho et al., 2011). However, the RAPD analysis of the tested isolates revealed that they are clonal isolates but from two different ancestral strains, RAPD02 for patient J and RAPD03 for patient O (Table 1), in line with the different MLST sequence types, ST-218 and ST-280 at the MLST database [http://pubmlst.org/bcc/], obtained for the isolates infecting patient J and O, respectively.
For the B. cenocepacia IIIA strain co-infecting patient AF for 2.5 years, the loss of the OAg was also observed when the FEV1 values were close to 40%. However, as demonstrated by the different RAPD profiles obtained (Table 1), another B. cenocepacia IIIA strain was obtained at an isolation date after the loss of the OAg by the original infecting strain; this new strain exhibit an OAg with a distinct banding pattern from the early isolates of the series (Figure 2A, column A9), Remarkably, all the B. contaminans clonal isolates co-infecting this patient kept the OAg when, for identical isolation times, B. cenocepacia IIIA clonal isolates lack the OAg (Figure 1B).
Only for B. cenocepacia IIIA isolates chronically infecting patients I and P over a period of time below 3 years, the loss of OAg along infection was not observed (Figure 1A). Interestingly, after this shorter period of chronic infection, the Burkholderia strains were eradicated from both patients.
Concerning the sequential B. cenocepacia lineage IIIB isolates retrieved from patient T over 1.2 years of infection before eradication, all the sequential isolates exhibit the OAg even though the FEV1 value was in the low range of 25–30%. In the case of patient R, among the 25 sequential isolates of the B. cenocepacia lineage IIIB strain obtained during a very extended infection period (7.5 years) two isolates retrieved after the fifth year of infection were found to lack the OAg (Figures 1, 2, columns B3 and B4, and Table 1). Interestingly, the OAg banding pattern obtained for isolates with ribopattern 15 infecting patient R was identical to the OAg profile extracted from two isolates retrieved from patient O (Figure 2, columns B3 and B2) and identical to those from four B. cenocepacia recA lineage IIIB isolates also present in patient AR who was mainly infected with B. dolosa (Figure 2, columns B3 and B6). All these isolates producing the same banding pattern were confirmed to share the same RAPD profile (Table 1 and Figure S1). Consistent with these results the MLST database showed that the B. cenocepacia lineage IIIB isolates retrieved from patients R and AR also have the same MLST sequence type (ST-43) [http://pubmlst.org/bcc/].
The OAg Is Stably Expressed in B. cepacia, B. contaminans and B. stabilis
Regarding the usually less frequent species worldwide that were examined in this study, the B. cepacia isolates (a total of 87) retrieved from five patients (N, AL, AP, AQ an AU) infected with this species for a period of time ranging from 2.2 to 9.6 years, all exhibit the OAg even when the FEV1 value was as low as 30% (patient N). Patients AL, AP and AQ, eradicated the infection after the screening period. Interestingly, the OAg banding pattern obtained for isolates with ribopattern 19 that infected patients AL and AP was identical to the OAg profile extracted from isolates retrieved from patients AU and D (Figure 2, columns C1 and C3), the sole exception were the two isolates retrieved from patient AU during the third year of infection with a different RAPD profile (Figures 1, 2, columns C3 and C4, and Table 1). All these isolates producing the same banding pattern were confirmed to share the same RAPD profile (Table 1 and Figure S1).
Concerning the 13 B. contaminans clonal isolates retrieved from patient V infected for a very long period of 15.2 years (Cunha et al., 2003; Coutinho et al., 2011), all of them were found to express the OAg (Figure 2, column D1).
Regarding the 17 B. stabilis isolates retrieved from patients B and H for a period of 3.8 and 2.5 years of chronic infection, respectively (Figure 1 and Table 1), the two strains isolated from each patient exhibit the same RAPD profiles (Figure S1, RAPD14 and RAPD15, Table 1). Remarkably, a stable phenotype concerning the presence or absence of the OAg over the infection period was found. The four isolates with RAPD15 profile retrieved from patient B exhibit an LPS that lack the OAg, independently of being early or late isolates, while the 4 clonal isolates of the strain with the same RAPD profile obtained from patient H, all have the OAg (Table 1 and Figure 2, columns G1 and G4). Interestingly, patient H died from the cepacia syndrome while patient B death was not related with the cepacia syndrome. Isolates with RAPD14 profile do have an LPS that stably maintained the OAg. When present, the banding patterns of the OAg were different for the two strains but identical for the clonal variants of each strain obtained from both patients (Figure 2, columns G2-G4).
The LPS From Early and Late B. dolosa Isolates Lack the OAg
The analysis of 14 B. dolosa isolates obtained from a single patient (AR) chronically infected with this species for 5.5 years showed that this strain did not have an LPS with the OAg chain from the beginning of the infection until dead (Figure 2F). The very peculiar B. dolosa LPS chemical structure of the first isolate IST4208 obtained from patient AR (Table S1) was examined before and a novel complete structure of the lipooligosaccharide (LOS) lacking the OAg component was revealed (Lorenzo et al., 2013). This LOS was found to show a strong proinflammatory activity (Lorenzo et al., 2013).
Discussion
Different virulence factors have been described in Burkholderia cepacia complex (Bcc) bacteria (Loutet and Valvano, 2010; Sousa et al., 2017) but less is known concerning their adaptive traits during chronic infection, compared with P. aeruginosa, the most prevalent and studied CF pathogen (Valentini et al., 2018). Before the systematic retrospective-longitudinal-screening, carried out in the present study, a retrospective phenotypic longitudinal assessment of the variation of mucoid exopolysaccharide production (Zlosnik et al., 2008), the switch from mucoid to non-mucoid (Zlosnik et al., 2011) and the swimming motility (Zlosnik et al., 2014) was performed during chronic infections involving numerous Bcc isolates representing different Bcc species and collected from 100 CF patients in the Vancouver area. Conversion of the mucoid-to-non-mucoid phenotype during chronic infection strengthen the concept of the association of the non-mucoid phenotype with the severity of the disease and the mucoid phenotype with persistence (Zlosnik et al., 2008, 2011). This behavior is in contrast with the phenotypic switch from non-mucoid to mucoid in P. aeruginosa chronic infections as a well-established paradigm for infection disease severity and persistence in this species (Valentini et al., 2018). Moreover, upon establishment of chronic infection, subsequent P. aeruginosa isolates show a reduction in the swimming ability to be non-motile (Valentini et al., 2018), contrasting with the observed data in Bcc infections (Zlosnik et al., 2014). The understanding of the occurrence of these and other adaptive traits that have been well-described in P. aeruginosa chronic infections (Valentini et al., 2018), but are far less known in Bcc chronic infection, is instrumental for a better understanding of bacterial transformations in the CF lungs during long term infections and infection control.
The LPS forms the outer leaflet of the outer membrane and has roles in antigenicity, the inflammatory response and exclusion of external molecules (King et al., 2009). Although the lipid A can vary, the variability in the O-antigen (OAg) is greater influencing adherence, colonization and the ability to evade the host's defense mechanisms (Kintz and Goldberg, 2008; King et al., 2009; Saldias et al., 2009; Kotrange et al., 2011; Maldonado et al., 2016; Ranf, 2016; Kintz et al., 2017). The ability of Bcc species to evade the host immune response often leads to chronic infections that are associated with significant loss of lung function due to the hyperactive inflammatory response that ultimately can lead to fatalities. The systematic, comprehensive and retrospective longitudinal screening conducted in this work, covering multiple sequential isolates from different Bcc strains of species, frequently or rarely isolated from CF patients, to search for the eventual lack of OAg expression in the late-stage of infections, reinforce the relevance that has been attributed to this switch. This was particularly frequent and clear for B. cenocepacia recA lineage IIIA and B. multivorans, also occurring at lower frequency in B. cenocepacia recA lineage IIIB. However, in the case of B. multivorans, there is a late isolate obtained within the series of sequential cloned isolates lacking the OAg that was found to express it. This behavior was previously observed for B. multivorans by others (Silva et al., 2016).
For the first time, it was shown that B. cepacia and B. contaminans do not lose the OAg even during long term infections that last for periods of time as extended as 9.6 and 15.2 years, respectively. Among the less frequent species, B. contaminans is currently considered an emerging CF pathogen (Vanlaere et al., 2009; Martina et al., 2013; Coutinho et al., 2015; Medina-Pascual et al., 2015; Nunvar et al., 2016; Power et al., 2016). Concerning the other two rarely found species examined, B. stabilis exhibits in two infected patients a stable phenotype concerning the presence or absence of the OAg over the infection period, consistent with the description of B. stabilis sp. Nov, as having a relatively stable genome (Vandamme et al., 2000). For the single patient chronically infected for 5.5 years with the more distantly related Bcc species B. dolosa, the OAg chain was absent from the beginning of the infection until the patient death. The novel LPS structure of this B. dolosa strain was examined and its strong pro-inflammatory activity reported before (Lorenzo et al., 2013).
The conclusions from this work were in general supported by the results gathered and suggest that the observed switch frequency of the OAg expression appears to be species dependent and also dependent on the duration of the infection and the level of deterioration of the lung function, increasing with both. This observation is consistent with the fact that cystic fibrosis is a genetic disorder associated with inflammation, sub-optimal antioxidant protection and the continuous use of antimicrobial therapy, all resulting in marked oxidative stress (Galli et al., 2012) that causes an increase in mutation rate. Results obtained from co-infected patients with different Bcc species reinforce those conclusions as it is for example the case for patient AF who was infected with B. contaminans isolates that kept the OAg during 5.8 years of chronic infection while late clonal isolates of the co-infecting B. cenocepacia IIIA strain during a much shorter period of 2.5 loss the OAg.
It is interesting to note that the two Bcc species that showed a higher tendency to lose the OAg expression along chronic lung infections in CF (B. cenocepacia and B. multivorans) are also the two most prevalent worldwide and the most feared among the CF community (Jones et al., 2004; Mahenthiralingam et al., 2005). Remarkably, B. cenocepacia and B. multivorans cell lysates are more potent stimulators of pro-inflammatory cytokines compared with those from other Bcc species (De Soyza et al., 2004). However, even though the prevalence of B. multivorans has recently replaced B. cenocepacia in several countries (Baldwin et al., 2008; Lipuma, 2010; Medina-Pascual et al., 2012; Peeters et al., 2017), it is clear that B. cenocepacia recA lineage IIIA remains the most dominant species (Manno et al., 2004; Nunvar et al., 2017; Scoffone et al., 2017; Teri et al., 2018). Remarkably, the particularly destructive nature of B. cenocepacia recA group IIIA infections in CF patients when compared with other Bcc bacteria has been reported (Manno et al., 2004; Zlosnik et al., 2015).
Remarkably, the single clone of the more distantly related B. dolosa Bcc species tested did not exhibit the OAg chain from the beginning to the end of the 5.5 year-infection as reported before (Lorenzo et al., 2013). Comprehensive comparative genomic studies performed during chronic infection on the most feared B. cenocepacia (Nunvar et al., 2017) and B. multivorans (Silva et al., 2016) and on the far less distributed B. dolosa (Lieberman et al., 2011); revealed that the genes affected during adaptive evolution in B. cenocepacia or in B. multivorans are different from B. dolosa. This indicates that evolution in B. dolosa during chronic CF infection is driven by different selective forces presumably linked to host immune responses (Lieberman et al., 2014; Nunvar et al., 2017). Specifically, while the most mutated genes reported to undergo adaptive within-patient evolution in B. cenocepacia were associated with oxidative stress response and transition metal metabolism, the same was not observed either in B. dolosa or in Pseudomonas aeruginosa (Lieberman et al., 2014; Nunvar et al., 2017). These genes encode proteins required for protection against the reactive oxygen species (ROS) produced by leukocytes thus suggesting the involvement of the host immune system in driving B. cenocepacia evolution during chronic CF infection (Nunvar et al., 2017). Given that persistent inflammation and neutrophil infiltration often accompany chronic lung infections, the authors hypothesized that under increased stress encountered in CF macrophages, the global stress response might be activated by the evolved bacterial population and thus modulate the course of infection (Chua et al., 2016, 2017; Nunvar et al., 2017). Since the interactions between pathogens and the immune system are highly complex, affecting pathogen adaptation to different host immune system stimuli, further investigation into the roles of macrophages and defense mechanisms in chronic infection outcome is needed (Chua et al., 2016, 2017).
The presence/absence of the OAg at the Bcc bacteria surface is important for cell evasion from the host immune response and pathogenicity. The OAg is considered highly immunogenic and induces the production of antibodies that may activate the complement pathway, either through the classic pathway or an alternate pathway, which leads to cellular death or phagocytosis (Reyes et al., 2012). Certain modifications in the oligosaccharide chain of the OAg may alter the interaction of the complement pathway. Several OAg of pathogens are similar to host molecules and this facilitate invasion through mimicking in the host (Reyes et al., 2012). An OAg deficient B. cenocepacia strain was found to be more susceptible to phagocytic internalization (Saldias et al., 2009) while, the loss of OAg expression in P. aeruginosa during chronic pulmonary infection did not confer phagocytic resistance in vitro (Demirdjian et al., 2017). Given that Burkholderia is a non-obligate intracellular pathogen while P. aeruginosa is considered an extracellular pathogen, it was hypothesized that the loss of OAg may benefit the ability of Burkholderia to access an intracellular environment, having no similar benefits for P. aeruginosa (Saldias et al., 2009). Recent studies suggest that there is a high selective pressure on the OAg biosynthetic locus leading to alterations both at the structural, genetic and regulatory levels strongly suggestive of an adaptive mechanism that potentially contribute to evade the host immune defenses in several Gram-negative pathogens (Murray et al., 2003; Yang et al., 2011; Maldonado et al., 2016; McCarthy et al., 2017; Faure et al., 2018; Neiger et al., 2019). Moreover, the OAg absence was shown to increase Bcc survival in eukaryotic cells as amoebae, epithelial cells and human macrophages (Saldias and Valvano, 2009; Saldias et al., 2009; Kotrange et al., 2011; Maldonado et al., 2016), leading to increased internalization of B. cenocepacia into macrophages upon phagocytosis (Saldias et al., 2009; Kotrange et al., 2011) and to facilitate B. multivorans growth inside macrophage (Schmerk and Valvano, 2013). These results may suggest that OAg loss could promote Bcc persistence through intracellular survival. Remarkably, the comparison of the ability to subvert the host's immune function, assessed by internalization assays using human dendritic cells, of three of the B. cenocepacia isolates examined in this work and retrieved from CF patient J showed that the late variants, IST4113 and IST4134, were significantly more internalized exhibiting increased survival within dendritic cells than the early isolate IST439 (Cabral et al., 2017). This early isolate was the only one that expresses the OAg unit (Hassan et al., 2017) supporting the idea that the loss of the OAg is advantageous for Bcc persistence. In addition, the absence of the OAg in these same isolates was suggested to reduce B. cenocepacia virulence potential using the Galleria mellonella infection model (Moreira et al., 2017). Such decrease in the virulence potential of early B. cenocepacia clonal variants exhibiting the OAg compared with late clonal variants lacking the OAg was also observed for two other chronically infected patients (AB and AN) with B. cenocepacia (Moreira et al., 2017). Also, a Salmonella typhimurium mutant missing the entire OAg was found to be avirulent in G. mellonella infection model while the shortening of the OAg chain length reduced the pathogenic potential by one-half compared to wild-type strain (Bender et al., 2013) suggesting that the OAg length is also a key-determinants of virulence in G. mellonella. The OAg truncation has also been implicated in increased neutrophil-mediated killing, complement-mediated susceptibility and phagocytosis of Salmonella by macrophages (Murray et al., 2003).
It is essential to obtain comprehensive knowledge about the pathogenesis of the various Bcc species involved in acute or chronic infections in different patients in order to fight the associated infections. This work reinforces the relevance that has been attributed to the switch involved in the lack of expression of the OAg during chronic infection in the most feared pathogenic Bcc species B. cenocepacia (in particular the recA lineage IIIA) and B. multivorans. It also suggests that the evolution of different Bcc species in chronic CF infection also at the level of OAg expression may be driven by different selective forces presumably associated to host immune responses.
Data Availability
All datasets generated for this study are included in the manuscript/Supplementary Files.
Author Contributions
AH performed most of the experimental work and CC contributed to the species/lineage identification and genotyping of part of the isolates. IS-C designed and supervised the work and with AH analyzed the data and wrote the manuscript. All the authors approved it.
Funding
Funding received by iBB—Institute for Bioengineering and Biosciences from Portuguese Foundation for Science and Technology (FCT) (UID/BIO/04565/2019) is acknowledged. This work was also supported by FCT through PhD fellowship to AH (FCT-Ph.D. Programme BIOTECnico—PD/BD/113629/2015).
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
The contribution, over the last 2 decades of epidemiological survey of Bcc respiratory infections in CF patients receiving care at Hospital de Santa Maria, in Lisbon, of Luís Lito, Celeste Barreto, Pilar Azevedo, Susana Correia, Luísa Pereira, Maria José Salgado, and José Melo-Cristino, is gratefully acknowledged. We also acknowledge Miguel Valvano, Queen's University Belfast, UK, for having shared with us his knowledge and interest on LPS.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fcimb.2019.00273/full#supplementary-material
Figure S1. Representative profiles based on Random Amplified Polymorphic DNA (RAPD) analysis of the Bcc isolates examined in this study retrieved from different CF patients during several years of chronic infection. The polymorphisms shown are based on the amplification by RAPD primers 270 (A), 208 (B), and 272 (C) in the same sequential order. The polymorphisms originated were analyzed using the software GelJ to detect the banding patterns. These profiles correspond to the 15-different representative selected isolates of B. cenocepacia IIIA (RAPD 01-08), B. cenocepacia IIIB (RAPD 09-11), B. cepacia (RAPD 12 and 13), and B. stabilis (RAPD 14 and 15). RAPD profiles for isolates: 01–IST416, 02–IST439, 03–IST462, 04–IST432, 05–IST4121, 06–IST4240a, 07–IST4272, 08–IST4197, 09–IST435, 10–IST438, 11–IST466, 12–IST4152, 13–IST4546, 14–IST413, 15–IST412 and λ−1 Kb plus DNA ladder.
Table S1. Detailed description of the Burkholderia sequential isolates per patient examined in this study with their isolation dates. The presence or absence of the LPS OAg is presented by + or – symbols, respectively, and their corresponding profiles (Figure 2) is provided. Information on the species identification and genotyping for the different strains tested are also presented: the genotyping data and the ribopatterns were obtained before (Cunha et al., 2003, 2007; Coutinho et al., 2011, 2015; Moreira et al., 2014, 2017) and the RAPD profiles were obtained in this study (Figure S1) together with the sequence type, when present in the MLST database. The different letter (A, B, C and D) of the isolate signature (as ISTnumber A, B, C or D) was used to indicate different clone at the same isolation date. ND, Not determined.
Abbreviations
Bcc, Burkholderia cepacia complex; CF, cystic fibrosis; HSM, Hospital de Santa Maria, Centro Hospitalar Lisboa Norte; LPS, lipopolysaccharide; OAg, O-antigen; RAPD, random amplified polymorphism DNA; MLST, multilocus sequence typing; ST, MLST sequence type.
References
Bach, E., Sant'Anna, F. H., Magrich Dos Passos, J. F., Balsanelli, E., de Baura, V. A., Pedrosa, F. O., et al. (2017). Detection of misidentifications of species from the Burkholderia cepacia complex and description of a new member, the soil bacterium Burkholderia catarinensis sp. nov. Pathog Dis. 75:6. doi: 10.1093/femspd/ftx076
Baldwin, A., Mahenthiralingam, E., Drevinek, P., Pope, C., Waine, D. J., Henry, D. A., et al. (2008). Elucidating global epidemiology of Burkholderia multivorans in cases of cystic fibrosis by multilocus sequence typing. J. Clin. Microbiol. 46, 290–295. doi: 10.1128/JCM.01818-07
Bender, J. K., Wille, T., Blank, K., Lange, A., and Gerlach, R. G. (2013). LPS structure and PhoQ activity are important for Salmonella Typhimurium virulence in the Galleria mellonella infection model [corrected]. PLoS ONE 8:e73287. doi: 10.1371/journal.pone.0073287
Berry, M. C., McGhee, G. C., Zhao, Y., and Sundin, G. W. (2009). Effect of a waaL mutation on lipopolysaccharide composition, oxidative stress survival, and virulence in Erwinia amylovora. FEMS Microbiol. Lett. 291, 80–87. doi: 10.1111/j.1574-6968.2008.01438.x
Bowden, S. D., Hale, N., Chung, J. C., Hodgkinson, J. T., Spring, D. R., and Welch, M. (2013). Surface swarming motility by Pectobacterium atrosepticum is a latent phenotype that requires O antigen and is regulated by quorum sensing. Microbiology 159(Pt 11), 2375–2385. doi: 10.1099/mic.0.070748-0
Cabral, M., Pereira, M., Silva, Z., Iria, I., Coutinho, C., Lopes, A., et al. (2017). Using dendritic cells to evaluate how Burkholderia cenocepacia clonal isolates from a chronically infected cystic fibrosis patient subvert immune functions. Med. Microbiol. Immunol. 206, 111–123. doi: 10.1007/s00430-016-0488-4
Chua, S. L., Ding, Y., Liu, Y., Cai, Z., Zhou, J., Swarup, S., et al. (2016). Reactive oxygen species drive evolution of pro-biofilm variants in pathogens by modulating cyclic-di-GMP levels. Open Biol. 6:11. doi: 10.1098/rsob.160162
Chua, S. L., Ding, Y., Liu, Y., Cai, Z., Zhou, J., Swarup, S., et al. (2017). Correction to 'Reactive oxygen species drive evolution of pro-biofilm variants in pathogens by modulating cyclic-di-GMP levels'. Open Biol. 7:160162. doi: 10.1098/rsob.170197
Correia, S., Nascimento, C., Pereira, L., Cunha, M. V., Sá-Correia, I., and Barreto, C. (2008). Infecção respiratória por bactérias do complexo cepacia: evolução clínica em doentes com fibrose quística. Rev. Portuguesa Pneumol. 14, 5–26. doi: 10.1016/S0873-2159(15)30216-6
Coutinho, C. P., Barreto, C., Pereira, L., Lito, L., Melo Cristino, J., and Sá-Correia, I. (2015). Incidence of Burkholderia contaminans at a cystic fibrosis centre with an unusually high representation of Burkholderia cepacia during 15 years of epidemiological surveillance. J. Med. Microbiol. 64, 927–935. doi: 10.1099/jmm.0.000094
Coutinho, C. P., Dos Santos, S. C., Madeira, A., Mira, N. P., Moreira, A. S., and Sá-Correia, I. (2011). Long-term colonization of the cystic fibrosis lung by Burkholderia cepacia complex bacteria: epidemiology, clonal variation, and genome-wide expression alterations. Front. Cell. Infect. Microbiol. 1:12. doi: 10.3389/fcimb.2011.00012
Cullen, L., and McClean, S. (2015). Bacterial adaptation during chronic respiratory infections. Pathogens 4, 66–89. doi: 10.3390/pathogens4010066
Cunha, M. V., Leitão, J. H., Mahenthiralingam, E., Vandamme, P., Lito, L., Barreto, C., et al. (2003). Molecular analysis of Burkholderia cepacia complex isolates from a Portuguese cystic fibrosis center: a 7-year study. J. Clin. Microbiol. 41, 4113–4120. doi: 10.1128/JCM.41.9.4113-4120.2003
Cunha, M. V., Pinto-de-Oliveira, A., Meirinhos-Soares, L., Salgado, M. J., Melo-Cristino, J., Correia, S., et al. (2007). Exceptionally high representation of Burkholderia cepacia among B. cepacia complex isolates recovered from the major Portuguese cystic fibrosis center. J. Clin. Microbiol. 45, 1628–1633. doi: 10.1128/JCM.00234-07
De Smet, B., Mayo, M., Peeters, C., Zlosnik, J. E., Spilker, T., Hird, T. J., et al. (2015). Burkholderia stagnalis sp. nov. and Burkholderia territorii sp. nov., two novel Burkholderia cepacia complex species from environmental and human sources. Int. J. Syst. Evol. Microbiol. 65, 2265–2271. doi: 10.1099/ijs.0.000251
De Soyza, A., Ellis, C. D., Khan, C. M., Corris, P. A., and Demarco de Hormaeche, R. (2004). Burkholderia cenocepacia lipopolysaccharide, lipid A, and proinflammatory activity. Am. J. Resp. Crit. Care Med. 170, 70–77. doi: 10.1164/rccm.200304-592OC
Demirdjian, S., Schutz, K., Wargo, M. J., Lam, J. S., and Berwin, B. (2017). The effect of loss of O-antigen ligase on phagocytic susceptibility of motile and non-motile Pseudomonas aeruginosa. Mol. Immunol. 92, 106–115. doi: 10.1016/j.molimm.2017.10.015
Depoorter, E., Bull, M. J., Peeters, C., Coenye, T., Vandamme, P., and Mahenthiralingam, E. (2016). Burkholderia: an update on taxonomy and biotechnological potential as antibiotic producers. Appl. Microbiol. Biotechnol. 100, 5215–5229. doi: 10.1007/s00253-016-7520-x
Döring, G., Parameswaran, I. G., and Murphy, T. F. (2011). Differential adaptation of microbial pathogens to airways of patients with cystic fibrosis and chronic obstructive pulmonary disease. FEMS Microbiol. Rev. 35, 124–146. doi: 10.1111/j.1574-6976.2010.00237.x
Drevinek, P., and Mahenthiralingam, E. (2010). Burkholderia cenocepacia in cystic fibrosis: epidemiology and molecular mechanisms of virulence. Clin. Microbiol. Infect. 16, 821–830. doi: 10.1111/j.1469-0691.2010.03237.x
Faure, E., Kwong, K., and Nguyen, D. (2018). Pseudomonas aeruginosa in chronic lung infections: how to adapt within the host? Front. Immunol. 9:2416. doi: 10.3389/fimmu.2018.02416
Galli, F., Battistoni, A., Gambari, R., Pompella, A., Bragonzi, A., Pilolli, F., et al. (2012). Oxidative stress and antioxidant therapy in cystic fibrosis. Biochim. Biophys. Acta 1822, 690–713. doi: 10.1016/j.bbadis.2011.12.012
Hassan, A. A., Maldonado, R. F., dos Santos, S. C., Di Lorenzo, F., Silipo, A., Coutinho, C. P., et al. (2017). Structure of O-antigen and hybrid biosynthetic locus in Burkholderia cenocepacia clonal variants recovered from a cystic fibrosis patient. Front. Microbiol. 8:1027. doi: 10.3389/fmicb.2017.01027
Heras, J., Dominguez, C., Mata, E., Pascual, V., Lozano, C., Torres, C., et al. (2015). GelJ–a tool for analyzing DNA fingerprint gel images. BMC Bioinform. 16:270. doi: 10.1186/s12859-015-0703-0
Hogardt, M., and Heesemann, J. (2010). Adaptation of Pseudomonas aeruginosa during persistence in the cystic fibrosis lung. Int. J. Med. Microbiol. 300, 557–562. doi: 10.1016/j.ijmm.2010.08.008
Jones, A. M., Dodd, M. E., Govan, J. R., Barcus, V., Doherty, C. J., Morris, J., et al. (2004). Burkholderia cenocepacia and Burkholderia multivorans: influence on survival in cystic fibrosis. Thorax 59, 948–951. doi: 10.1136/thx.2003.017210
Kalish, L. A., Waltz, D. A., Dovey, M., Potter-Bynoe, G., McAdam, A. J., Lipuma, J. J., et al. (2006). Impact of Burkholderia dolosa on lung function and survival in cystic fibrosis. Am. J. Resp. Crit. Care Med. 173, 421–425. doi: 10.1164/rccm.200503-344OC
Kenna, D. T. D., Lilley, D., Coward, A., Martin, K., Perry, C., Pike, R., et al. (2017). Prevalence of Burkholderia species, including members of Burkholderia cepacia complex, among UK cystic and non-cystic fibrosis patients. J. Med. Microbiol. 66, 490–501. doi: 10.1099/jmm.0.000458
King, J. D., Kocincova, D., Westman, E. L., and Lam, J. S. (2009). Review: Lipopolysaccharide biosynthesis in Pseudomonas aeruginosa. Innate Immunity 15, 261–312. doi: 10.1177/1753425909106436
Kintz, E., and Goldberg, J. B. (2008). Regulation of lipopolysaccharide O antigen expression in Pseudomonas aeruginosa. Fut. Microbiol. 3, 191–203. doi: 10.2217/17460913.3.2.191
Kintz, E., Heiss, C., Black, I., Donohue, N., Brown, N., Davies, M. R., et al. (2017). Salmonella enterica Serovar Typhi Lipopolysaccharide O-antigen Modifications Impact on serum resistance and antibody recognition. Infect. Immunity. 85, e01021–16. doi: 10.1128/IAI.01021-16
Kotrange, S., Kopp, B., Akhter, A., Abdelaziz, D., Abu Khweek, A., Caution, K., et al. (2011). Burkholderia cenocepacia O polysaccharide chain contributes to caspase-1-dependent IL-1beta production in macrophages. J. Leukoc. Biol. 89, 481–488. doi: 10.1189/jlb.0910513
Lieberman, T. D., Flett, K. B., Yelin, I., Martin, T. R., McAdam, A. J., Priebe, G. P., et al. (2014). Genetic variation of a bacterial pathogen within individuals with cystic fibrosis provides a record of selective pressures. Nat. Genetics 46, 82–87. doi: 10.1038/ng.2848
Lieberman, T. D., Michel, J. B., Aingaran, M., Potter-Bynoe, G., Roux, D., Davis, M. R. Jr., et al. (2011). Parallel bacterial evolution within multiple patients identifies candidate pathogenicity genes. Nat. Genetics 43, 1275–1280. doi: 10.1038/ng.997
Lipuma, J. J. (2010). The changing microbial epidemiology in cystic fibrosis. Clin. Microbiol. Rev. 23, 299–323. doi: 10.1128/CMR.00068-09
Lorè, N. I., Cigana, C., De Fino, I., Riva, C., Juhas, M., Schwager, S., et al. (2012). Cystic fibrosis-niche adaptation of Pseudomonas aeruginosa reduces virulence in multiple infection hosts. PLoS ONE 7:e35648. doi: 10.1371/journal.pone.0035648
Lorenzo, F. D., Sturiale, L., Palmigiano, A., Lembo-Fazio, L., Paciello, I., Coutinho, C. P., et al. (2013). Chemistry and biology of the potent endotoxin from a Burkholderia dolosa clinical isolate from a cystic fibrosis patient. Chem. Biochem. 14, 1105–1115. doi: 10.1002/cbic.201300062
Loutet, S. A., and Valvano, M. A. (2010). A decade of Burkholderia cenocepacia virulence determinant research. Infect. Immunity 78, 4088–4100. doi: 10.1128/IAI.00212-10
Madeira, A., dos Santos, S. C., Santos, P. M., Coutinho, C. P., Tyrrell, J., McClean, S., et al. (2013). Proteomic profiling of Burkholderia cenocepacia clonal isolates with different virulence potential retrieved from a cystic fibrosis patient during chronic lung infection. PLoS ONE. 8:e83065. doi: 10.1371/journal.pone.0083065
Madeira, A., Santos, P. M., Coutinho, C. P., Pinto-de-Oliveira, A., and Sá-Correia, I. (2011). Quantitative proteomics (2-D DIGE) reveals molecular strategies employed by Burkholderia cenocepacia to adapt to the airways of cystic fibrosis patients under antimicrobial therapy. Proteomics 11, 1313–1328. doi: 10.1002/pmic.201000457
Mahenthiralingam, E., Bischof, J., Byrne, S. K., Radomski, C., Davies, J. E., Av-Gay, Y., et al. (2000). DNA-Based diagnostic approaches for identification of Burkholderia cepacia complex, Burkholderia vietnamiensis, Burkholderia multivorans, Burkholderia stabilis, and Burkholderia cepacia genomovars I and III. J. Clin. Microbiol. 38, 3165–3173.
Mahenthiralingam, E., Campbell, M. E., Henry, D. A., and Speert, D. P. (1996). Epidemiology of Burkholderia cepacia infection in patients with cystic fibrosis: analysis by randomly amplified polymorphic DNA fingerprinting. J. Clin. Microbiol. 34, 2914–2920.
Mahenthiralingam, E., Urban, T. A., and Goldberg, J. B. (2005). The multifarious, multireplicon Burkholderia cepacia complex. Nat. Rev. Microbiol. 3, 144–156. doi: 10.1038/nrmicro1085
Maldonado, R. F., Sá-Correia, I., and Valvano, M.A. (2016). Lipopolysaccharide modification in Gram-negative bacteria during chronic infection. FEMS Microbiol. Rev. 40, 480–493. doi: 10.1093/femsre/fuw007
Manno, G., Dalmastri, C., Tabacchioni, S., Vandamme, P., Lorini, R., Minicucci, L., et al. (2004). Epidemiology and clinical course of Burkholderia cepacia complex infections, particularly those caused by different Burkholderia cenocepacia strains, among patients attending an Italian Cystic Fibrosis Center. J. Clin. Microbiol. 42, 1491–1497. doi: 10.1128/JCM.42.4.1491-1497.2004
Markussen, T., Marvig, R. L., Gomez-Lozano, M., Aanaes, K., Burleigh, A. E., Hoiby, N., et al. (2014). Environmental heterogeneity drives within-host diversification and evolution of Pseudomonas aeruginosa. MBio 5, e01592–e01514. doi: 10.1128/mBio.01592-14
Marolda, C. L., Lahiry, P., Vines, E., Saldias, S., and Valvano, M. A. (2006). Micromethods for the characterization of lipid A-core and O-antigen lipopolysaccharide. Methods Mol. Biol. 347, 237–252. doi: 10.1385/1-59745-167-3:237
Marolda, C. L., Welsh, J., Dafoe, L., and Valvano, M. A. (1990). Genetic analysis of the O7-polysaccharide biosynthesis region from the Escherichia coli O7:K1 strain VW187. J. Bacteriol. 172, 3590–3599. doi: 10.1128/jb.172.7.3590-3599.1990
Marquez, L., Jones, K. N., Whaley, E. M., Koy, T. H., Revell, P. A., Taylor, R. S., et al. (2017). An Outbreak of Burkholderia cepacia complex infections associated with contaminated liquid docusate. Infect. Control Hospital Epidemiol. 38, 567–573. doi: 10.1017/ice.2017.11
Martina, P., Bettiol, M., Vescina, C., Montanaro, P., Mannino, M. C., Prieto, C. I., et al. (2013). Genetic diversity of Burkholderia contaminans isolates from cystic fibrosis patients in Argentina. J. Clin. Microbiol. 51, 339–344. doi: 10.1128/JCM.02500-12
Martina, P., Leguizamon, M., Prieto, C. I., Sousa, S. A., Montanaro, P., Draghi, W. O., et al. (2018). Burkholderia puraquae sp. nov., a novel species of the Burkholderia cepacia complex isolated from hospital settings and agricultural soils. Int. J. Syst. Evol. Microbiol. 68, 14–20. doi: 10.1099/ijsem.0.002293
Marvig, R. L., Johansen, H. K., Molin, S., and Jelsbak, L. (2013). Genome analysis of a transmissible lineage of Pseudomonas aeruginosa reveals pathoadaptive mutations and distinct evolutionary paths of hypermutators. PLoS Genetics 9:e1003741. doi: 10.1371/journal.pgen.1003741
McCarthy, R. R., Mazon-Moya, M. J., Moscoso, J. A., Hao, Y., Lam, J. S., Bordi, C., et al. (2017). Cyclic-di-GMP regulates lipopolysaccharide modification and contributes to Pseudomonas aeruginosa immune evasion. Nat. Microbiol. 2:17027. doi: 10.1038/nmicrobiol.2017.27
Medina-Pascual, M. J., Valdezate, S., Carrasco, G., Villalon, P., Garrido, N., and Saez-Nieto, J. A. (2015). Increase in isolation of Burkholderia contaminans from Spanish patients with cystic fibrosis. Clin. Microbiol. Infection 21, 150–156. doi: 10.1016/j.cmi.2014.07.014
Medina-Pascual, M. J., Valdezate, S., Villalon, P., Garrido, N., Rubio, V., and Saez-Nieto, J. A. (2012). Identification, molecular characterisation and antimicrobial susceptibility of genomovars of the Burkholderia cepacia complex in Spain. Eur. J. Clin. Microbiol. Infect. Dis. 31, 3385–3396. doi: 10.1007/s10096-012-1707-6
Moehring, R. W., Lewis, S. S., Isaacs, P. J., Schell, W. A., Thomann, W. R., Althaus, M. M., et al. (2014). Outbreak of bacteremia due to Burkholderia contaminans linked to intravenous fentanyl from an institutional compounding pharmacy. JAMA Internal Med. 174, 606–612. doi: 10.1001/jamainternmed.2013.13768
Moreira, A. S., Coutinho, C. P., Azevedo, P., Lito, L., Melo-Cristino, J., and and, Sá-Correia, I. (2014). Burkholderia dolosa phenotypic variation during the decline in lung function of a cystic fibrosis patient during 5.5 years of chronic colonization. J. Med. Microbiol. 63(Pt 4), 594–601. doi: 10.1099/jmm.0.069849-0
Moreira, A. S., Mil-Homens, D., Sousa, S. A., Coutinho, C. P., Pinto-de-Oliveira, A., Ramos, C. G., et al. (2017). Variation of Burkholderia cenocepacia virulence potential during cystic fibrosis chronic lung infection. Virulence 8, 782–796. doi: 10.1080/21505594.2016.1237334
Murray, G. L., Attridge, S. R., and Morona, R. (2003). Regulation of Salmonella typhimurium lipopolysaccharide O antigen chain length is required for virulence; identification of FepE as a second Wzz. Mol. Microbiol. 47, 1395–1406. doi: 10.1046/j.1365-2958.2003.03383.x
Neiger, M. R., Gonzalez, J. F., Gonzalez-Escobedo, G., Kuck, H., White, P., and Gunn, J. S. (2019). Pathoadaptive alteration of Salmonella biofilm formation in response to the gallbladder environment. J. Bacteriol. 201, e00774–18. doi: 10.1128/JB.00774-18
Nunvar, J., Capek, V., Fiser, K., Fila, L., and Drevinek, P. (2017). What matters in chronic Burkholderia cenocepacia infection in cystic fibrosis: insights from comparative genomics. PLOS Pathog. 13:e1006762. doi: 10.1371/journal.ppat.1006762
Nunvar, J., Kalferstova, L., Bloodworth, R. A., Kolar, M., Degrossi, J., Lubovich, S., et al. (2016). Understanding the Pathogenicity of Burkholderia contaminans, an Emerging Pathogen in Cystic Fibrosis. PLoS ONE. 11:e0160975. doi: 10.1371/journal.pone.0160975
Ong, K. S., Aw, Y. K., Lee, L. H., Yule, C. M., Cheow, Y. L., and Lee, S. M. (2016). Burkholderia paludis sp nov., an antibiotic-siderophore producing novel Burkholderia cepacia complex species, isolated from malaysian tropical peat swamp soil. Front. Microbiol. 7:2046. doi: 10.3389/fmicb.2016.02046
Peeters, C., Cooper, V. S., Hatcher, P. J., Verheyde, B., Carlier, A., and Vandamme, P. (2017). Comparative genomics of Burkholderia multivorans, a ubiquitous pathogen with a highly conserved genomic structure. PLoS ONE 12:e0176191. doi: 10.1371/journal.pone.0176191
Pier, G. B. (2007). Pseudomonas aeruginosa lipopolysaccharide: a major virulence factor, initiator of inflammation and target for effective immunity. Int. J. Med. Microbiol. 297, 277–295. doi: 10.1016/j.ijmm.2007.03.012
Power, R. F., Linnane, B., Martin, R., Power, N., Harnett, P., Casserly, B., et al. (2016). The first reported case of Burkholderia contaminans in patients with cystic fibrosis in Ireland: from the Sargasso Sea to Irish Children. BMC Pulmonary Med. 16:57. doi: 10.1186/s12890-016-0219-z
Ranf, S. (2016). Immune Sensing of Lipopolysaccharide in plants and animals: same but different. PLoS Pathog. 12:e1005596. doi: 10.1371/journal.ppat.1005596
Reyes, R. E., González, C. R., Jiménez, R. C., Herrera, M. O., and Andrade, A. A. (2012). “Mechanisms of O-Antigen Structural Variation of Bacterial Lipopolysaccharide (LPS),” in The Complex World of Polysaccharides, ed. D. N. Karunaratne (Rijeka: InTech), 71–98.
Roux, D., Weatherholt, M., Clark, B., Gadjeva, M., Renaud, D., Scott, D., et al. (2017). Immune recognition of the epidemic cystic fibrosis pathogen Burkholderia Dolosa. Infect. Immunity e00765–16. doi: 10.1128/IAI.00765-16
Saldias, M. S., Ortega, X., and Valvano, M. A. (2009). Burkholderia cenocepacia O antigen lipopolysaccharide prevents phagocytosis by macrophages and adhesion to epithelial cells. J. Med. Microbiol. 58(Pt 12), 1542–1548. doi: 10.1099/jmm.0.013235-0
Saldias, M. S., and Valvano, M. A. (2009). Interactions of Burkholderia cenocepacia and other Burkholderia cepacia complex bacteria with epithelial and phagocytic cells. Microbiology 155(Pt 9), 2809–2817. doi: 10.1099/mic.0.031344-0
Schmerk, C. L., and Valvano, M. A. (2013). Burkholderia multivorans survival and trafficking within macrophages. J. Med. Microbiol. 62(Pt 2), 173–184. doi: 10.1099/jmm.0.051243-0
Scoffone, V. C., Chiarelli, L. R., Trespidi, G., Mentasti, M., Riccardi, G., and Buroni, S. (2017). Burkholderia cenocepacia Infections in cystic fibrosis patients: drug resistance and therapeutic approaches. Front. Microbiol. 8:1592. doi: 10.3389/fmicb.2017.01592
Silva, I. N., Ferreira, A. S., Becker, J. D., Zlosnik, J. E., Speert, D. P., He, J., et al. (2011). Mucoid morphotype variation of Burkholderia multivorans during chronic cystic fibrosis lung infection is correlated with changes in metabolism, motility, biofilm formation and virulence. Microbiology 157(Pt 11), 3124–3137. doi: 10.1099/mic.0.050989-0
Silva, I. N., Santos, P. M., Santos, M. R., Zlosnik, J. E., Speert, D. P., Buskirk, S. W., et al. (2016). Long-term evolution of Burkholderia multivorans during a chronic cystic fibrosis infection reveals shifting forces of selection. mSystems 1, e00029–e00016. doi: 10.1128/mSystems.00029-16
Sousa, S. A., Feliciano, J. R., Pita, T., Guerreiro, S. I., and Leitao, J. H. (2017). Burkholderia cepacia complex regulation of virulence gene expression: A Review. Genes 8:43. doi: 10.3390/genes8010043
Teri, A., Sottotetti, S., Biffi, A., Girelli, D., D'Accico, M., Arghittu, M., et al. (2018). Molecular typing of Burkholderia cepacia complex isolated from patients attending an Italian Cystic Fibrosis Centre. New Microbiol. 41, 141–144.
Toguchi, A., Siano, M., Burkart, M., and Harshey, R. M. (2000). Genetics of swarming motility in Salmonella enterica serovar typhimurium: critical role for lipopolysaccharide. J. Bacteriol. 182, 6308–6321. doi: 10.1128/JB.182.22.6308-6321.2000
Traverse, C. C., Mayo-Smith, L. M., Poltak, S. R., and Cooper, V. S. (2013). Tangled bank of experimentally evolved Burkholderia biofilms reflects selection during chronic infections. Proc. Natl. Acad. Sci. U.S.A. 110, E250–259. doi: 10.1073/pnas.1207025110
Valentini, M., Gonzalez, D., Mavridou, D. A., and Filloux, A. (2018). Lifestyle transitions and adaptive pathogenesis of Pseudomonas aeruginosa. Curr. Opin. Microbiol. 41, 15–20. doi: 10.1016/j.mib.2017.11.006
Vandamme, P., Mahenthiralingam, E., Holmes, B., Coenye, T., Hoste, B., De Vos, P., et al. (2000). Identification and population structure of Burkholderia stabilis sp. nov. (formerly Burkholderia cepacia genomovar IV). J. Clin. Microbiol. 38, 1042–1047.
Vanlaere, E., Baldwin, A., Gevers, D., Henry, D., De Brandt, E., LiPuma, J. J., et al. (2009). Taxon K, a complex within the Burkholderia cepacia complex, comprises at least two novel species, Burkholderia contaminans sp. nov. and Burkholderia lata sp. nov. Int. J. Syst. Evol. Microbiol. 59(Pt 1), 102–111. doi: 10.1099/ijs.0.001123-0
Vermis, K., Coenye, T., Mahenthiralingam, E., Nelis, H. J., and Vandamme, P. (2002). Evaluation of species-specific recA-based PCR tests for genomovar level identification within the Burkholderia cepacia complex. J. Med. Microbiol. 51, 937–940. doi: 10.1099/0022-1317-51-11-937
Vinion-Dubiel, A. D., and Goldberg, J. B. (2003). Lipopolysaccharide of Burkholderia cepacia complex. J. Endotoxin Res. 9, 201–213. doi: 10.1179/096805103225001404
Weber, C. F., and King, G. M. (2017). Volcanic soils as sources of novel CO-oxidizing paraburkholderia and burkholderia: Paraburkholderia hiiakae sp. nov., Paraburkholderia metrosideri sp. nov., Paraburkholderia paradisi sp. nov., Paraburkholderia peleae sp. nov., and Burkholderia alpina sp. nov. a Member of the Burkholderia cepacia Complex. Front. Microbiol. 8:207. doi: 10.3389/fmicb.2017.00207
Winstanley, C., O'Brien, S., and Brockhurst, M. A. (2016). Pseudomonas aeruginosa evolutionary adaptation and diversification in cystic fibrosis chronic lung infections. Trends Microbiol. 24, 327–337. doi: 10.1016/j.tim.2016.01.008
Woods, C. W., Bressler, A. M., LiPuma, J. J., Alexander, B. D., Clements, D. A., Weber, D. J., et al. (2004). Virulence associated with outbreak-related strains of Burkholderia cepacia complex among a cohort of patients with bacteremia. Clin. Infect. Dis. 38, 1243–1250. doi: 10.1086/383313
Yang, L., Jelsbak, L., Marvig, R. L., Damkiaer, S., Workman, C. T., Rau, M. H., et al. (2011). Evolutionary dynamics of bacteria in a human host environment. Proc. Natl. Acad. Sci. U.S.A. 108, 7481–7486. doi: 10.1073/pnas.1018249108
Zlosnik, J. E., Costa, P. S., Brant, R., Mori, P. Y., Hird, T. J., Fraenkel, M. C., et al. (2011). Mucoid and nonmucoid Burkholderia cepacia complex bacteria in cystic fibrosis infections. Am. J. Respir. Crit. Care Med. 183, 67–72. doi: 10.1164/rccm.201002-0203OC
Zlosnik, J. E., Hird, T. J., Fraenkel, M. C., Moreira, L. M., Henry, D. A., and Speert, D. P. (2008). Differential mucoid exopolysaccharide production by members of the Burkholderia cepacia complex. J. Clin. Microbiol. 46, 1470–1473. doi: 10.1128/JCM.02273-07
Zlosnik, J. E., Mori, P. Y., To, D., Leung, J., Hird, T. J., and Speert, D. P. (2014). Swimming motility in a longitudinal collection of clinical isolates of Burkholderia cepacia complex bacteria from people with cystic fibrosis. PLoS ONE. 9:e106428. doi: 10.1371/journal.pone.0106428
Keywords: Burkholderia cepacia complex species, cystic fibrosis, chronic pulmonary infections, lipopolysaccharide O-antigen, within-host clonal variation
Citation: Hassan AA, Coutinho CP and Sá-Correia I (2019) Burkholderia cepacia Complex Species Differ in the Frequency of Variation of the Lipopolysaccharide O-Antigen Expression During Cystic Fibrosis Chronic Respiratory Infection. Front. Cell. Infect. Microbiol. 9:273. doi: 10.3389/fcimb.2019.00273
Received: 15 May 2019; Accepted: 17 July 2019;
Published: 31 July 2019.
Edited by:
Eric Ghigo, IHU Mediterranee Infection, FranceReviewed by:
Evguenii Vinogradov, National Research Council Canada, CanadaPaul Race, University of Bristol, United Kingdom
Copyright © 2019 Hassan, Coutinho and Sá-Correia. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Isabel Sá-Correia, aXNhY29ycmVpYUB0ZWNuaWNvLnVsaXNib2EucHQ=