 Elena G. Salina1†
Elena G. Salina1† Artem Grigorov2†
Artem Grigorov2† Yulia Skvortsova2
Yulia Skvortsova2 Konstantin Majorov3
Konstantin Majorov3 Oksana Bychenko2
Oksana Bychenko2 Albina Ostrik1
Albina Ostrik1 Nadezhda Logunova3
Nadezhda Logunova3 Dmitriy Ignatov2‡
Dmitriy Ignatov2‡ Arseny Kaprelyants1
Arseny Kaprelyants1 Alexander Apt3
Alexander Apt3 Tatyana Azhikina2*
Tatyana Azhikina2*- 1Laboratory of Biochemistry of Stresses in Microorganisms, Bach Institute of Biochemistry, Research Center of Biotechnology, Moscow, Russia
- 2Laboratory of Regulatory Transcriptomics, Department of Genomics and Postgenomic Technologies, Shemyakin and Ovchinnikov Institute of Bioorganic Chemistry, Moscow, Russia
- 3Laboratory for Immunogenetics, Department of Immunology, Central Institute for Tuberculosis, Moscow, Russia
Small non-coding RNAs play a significant role in bacterial adaptation to changing environmental conditions. We investigated the dynamics of expression of MTS1338, a small non-coding RNA of Mycobacterium tuberculosis, in the mouse model in vivo, regulation of its expression in the infected macrophages, and the consequences of its overexpression in bacterial cultures. Here we demonstrate that MTS1338 significantly contributes to host-pathogen interactions. Activation of the host immune system triggered NO-inducible up-regulation of MTS1338 in macrophage-engulfed mycobacteria. Constitutive overexpression of MTS1338 in cultured mycobacteria improved their survival in vitro under low pH conditions. MTS1338 up-regulation launched a spectrum of shifts in the transcriptome profile similar to those reported for M. tuberculosis adaptation to hostile intra-macrophage environment. Using the RNA-seq approach, we demonstrate that gene expression changes accompanying MTS1338 overexpression indicate reduction in translational activity and bacterial growth. These changes indicate mycobacteria entering the dormant state. Taken together, our results suggest a direct involvement of this sRNA in the interplay between mycobacteria and the host immune system during infectious process.
Introduction
M. tuberculosis persistence in the infected host involves several stages and may have different manifestations: initial infection followed by semi-acute or chronic diseases; latent infection characterized by the presence of viable bacteria with slow-to-no level of replication and the lack of clinical manifestations; and transition from the latent state to reactivation processes (Stewart et al., 2003; Russell, 2007). The spectrum of the disease manifestations depends upon a dynamic balance between protective host responses and defensive strategies of M. tuberculosis. Identification of molecular mechanisms of M. tuberculosis adaptation to the host immune defense during its persistence within macrophages is an important scientific and medical problem.
Long co-evolution of M. tuberculosis and its human host allowed the pathogen to develop strategies that can effectively combat host defense systems. Regulatory proteins, non-coding RNAs and their targets constitute complex adaptive metabolic networks that allow the pathogen to resist host response at different stages of infection. Bacterial sRNAs participate in regulation of transcription and translation by affecting the level of gene expression and mRNA stability. Mostly, sRNAs are expressed in response to the external factors, helping bacteria to adaptively react to the changing environmental conditions and regulate the key stages of pathogenesis (Holmqvist and Wagner, 2017; Dutta and Srivastava, 2018; Hör et al., 2018).
Application of the high throughput sequencing and computer algorithm approaches allowed identification of dozens of sRNAs in mycobacterial species (Haning et al., 2014; Schwenk and Arnvig, 2018; Taneja and Dutta, 2019). Several in vitro studies have elucidated the functioning of sRNAs in M. tuberculosis (Arnvig et al., 2011; Solans et al., 2014; Moores et al., 2017; Gerrick et al., 2018; Mai et al., 2019). However, dissecting the role of a particular sRNA in mycobacterial physiology appeared to be difficult, especially in in vivo settings.
One of such RNAs, MTS1338 (DosR-associated sRNA, ncRv11733), is highly expressed during the stationary phase of growth (Arnvig and Young, 2012), and the dormancy state (Ignatov et al., 2015). This sRNA is present only in genomes of highly pathogenic mycobacteria and is very conservative. In vitro experiments demonstrated that its transcription is controlled by the transcriptional regulator DosR and is activated under hypoxic and NO-induced stresses (Moores et al., 2017), suggesting that MTS1338 may play a role during the stable phase of infection, when host responses confront mycobacterial multiplication more or less successfully. Indeed, we and others demonstrated a striking increase in the MTS1338 transcription in animal models of chronic infection (Arnvig and Young, 2012; Ignatov et al., 2014). Thus, it seems likely that MTS1338 triggers adaptive biochemical cascades for intracellular persistence.
Here, we characterize the dynamic changes in the MTS1338 expression in mycobacteria obtained from the lungs of genetically susceptible and resistant TB-infected mice, and provide a direct evidence that the level of expression is regulated by the IFN-γ-dependent NO production. Using high-throughput technologies, we describe changes in the genome transcription profile that accompany an increased MTS1338 transcription. Overexpression of MTS1338 leads to transcriptional shifts consistent with decreased bacterial metabolism, cell division and adaptation to host immune responses experienced by mycobacteria residing within host macrophages. Taken together, our results demonstrate that the small non-coding MTS1338 RNA regulates molecular mechanisms providing M. tuberculosis inter-macrophage survival.
Materials and Methods
Bacterial Strains, Media, and Growth Conditions
For in vitro experiments, M. tuberculosis H37Rv (WT), pMV (empty plasmid control) and OVER (MTS1338 overexpressing) M. tuberculosis strains were initially grown from frozen stocks for 10 days in Sauton medium. Medium content (per liter): 0.5 g KH2PO4, 1.4 g MgSO4×7H2O, 4 g L-asparagine, 60 ml glycerol, 0.05 g ferric ammonium citrate, 2 g sodium citrate, 0.1 ml 1% ZnSO4, pH 7.0 (adjusted with 1M NaOH). Supplements: ADC growth supplement (Connell, 1994), 0.05% Tween 80 and 50 μg/ml kanamycin (Sigma-Aldrich, USA). Growth conditions: 37°C with agitation (200 rpm). The starter cultures were inoculated into fresh medium (the same composition) and grown up to stationary phase for RNA-seq experiments and stress survival experiments.
For cloning procedures, Escherichia coli DH5α was grown in Luria Bertani (LB) broth and LB-agar. When required, antibiotics were added at the following concentrations: kanamycin (Sigma-Aldrich), 50 μg/ml (M. tuberculosis); ampicillin (Invitrogen, USA), 100 μg/ml (E. coli).
M. tuberculosis Over and pMV (Control) Strains Establishment
The MTS1338 gene-containing vector was constructed on the basis of the pMV261 (Stover et al., 1991) as described by Ignatov et al. (2015). The plasmid was transferred into mycobacteria by electroporation. MTS1338 overexpression was confirmed by quantitative PCR. The control strain was produced using an empty pMV261 vector.
Growth Inhibition in vitro by NO, H2O2, and Low pH
Bacterial cultures were grown up to the stationary phase, washed up with PBS and diluted to OD600 = 0.2 (107 CFU/ml) by (i) Sauton medium (pH 5.5) with ADC growth supplement and 0.05% Tween for low pH stress; (ii) by the culture supernatant to study inhibitory effects of NO (provided by the DETA/NO donor, 0.5 mM) and H2O2 (10 mM). Cell viability after 24 h and 48 h of stresses exposure were measured by incorporation of [3H]-uracil label. Two microliters of 5,6,-[3H]-uracil (2 μCi) were added to 1-ml culture samples and incubated at 37°C with agitation for 20 h. Two hundred microliters of culture were put in 3 ml 7% ice-cold CCl3COOH, incubated at 0° C for 15 min and filtered through glass microfiber filters (Whatman, USA). Precipitated cells were washed with 3 ml 7% CCl3COOH and 3 ml 96% ethanol. Filters were put in 10 ml of scintillation mixture; CPM were determined by LS analyser (Beckman Instruments Inc., USA).
RNA Extraction From Cultured Mycobacteria
Bacterial cultures were grown up to the stationary phase, rapidly cooled on ice, centrifuged, and total RNA was isolated by phenol-chloroform extraction after cell disruption by Bead Beater with 0.1 mm zirconia beads (BioSpec Products, USA) as previously described (Rustad et al., 2009). After isolation, RNA was treated with Turbo DNase (Life Technologies, USA) to remove traces of genomic DNA, and purified with the RNeasy mini kit (Qiagen, Netherlands). Amounts and purity of RNA were determined spectrophotometrically; integrity of RNA was assessed in 1% agarose gel.
Libraries for RNA-Seq and RNA-Seq Data Analyses
RNA samples were depleted of 16S and 23S rRNA using RiboMinus™ Transcriptome Isolation Kit, bacteria (Invitrogen, USA). Sequencing libraries were generated using the resulting ribosomal transcript-depleted RNA and NEBNext Ultra II Directional RNA Library Prep Kit (NEB, USA) according to the manufacturers' protocol. Sequencing was performed using the Illumina NovaSeq as the single-ended 100 nt-long reads. Experiments were performed in triplicates.
After quality control evaluation and trimming of bad qualitative reads the reads were mapped on the reference M. tuberculosis genome (AL123456.3, http://www.ncbi.nlm.nih.gov/) by Bowtie2 (Langmead and Salzberg, 2012). The alignment was performed with the “-local” option, which allows leaving 5′ and 3′ ends uncharted. Calculation of the mapped reads for all genes was performed using functions of the featureCounts package (Liao et al., 2014) built into the author's script. Resulting statistics were visualized as transcription profiles using the Artemis genome browser (Carver et al., 2012).
Differentially expressed genes were identified by the software package DESeq2 (Love et al., 2014). The genes were considered to be differentially expressed, if the p-value was less than 0.05, the expected measure of false deviations (FDR) was not higher than 0.1, and the expression change module (FC, Fold change) was not less than 3. Further distribution of genes according functional categories was performed using the Mycobrowser database (https://mycobrowser.epfl.ch/).
Quantitative Reverse Transcription-PCR (qRT-PCR)
One microgram of total RNA was used for cDNA synthesis with random hexanucleotides and SuperScript III reverse transcriptase (Life Technologies, USA). Quantitative PCR was performed using qPCRmix-HS SYBR (Evrogen, Russia) and the Light Cycler 480 real-time PCR system (Roche, Switzerland); cycling conditions were as follows: 95°C for 20 s, 61°C for 20 s, 72°C for 30 s, repeat 40 times; primers are listed in Supplementary Table 1. In the end of amplification, a dissociation curve was plotted to confirm specificity of the product. All real-time experiments were repeated in triplicate. The results were normalized against the 16S rRNA gene. Calculations were performed according to (Ganger et al., 2017) for the relative expression ratio.
Infections of Peritoneal Macrophages and in vivo
Mycobacteria
For infection of mice and macrophage cultures, M. tuberculosis H37Rv (substrain Pasteur) from the collection of CIT were used. Mycobacteria were prepared to infect mice and macrophages as described previously (Lyadova et al., 2000). Briefly, to obtain log-phase bacteria for challenge, 50 μl from a thawed aliquot was added to 30 ml of Dubos broth (BD Bioscience, USA) supplemented with 0.5% Fatty Acid-Poor BSA (Calbiochem-Behring Corp., USA) and incubated for 2 weeks at 37°C. The resulting suspension was washed two times at 3,000 g, 20 min, 4°C with Ca2+- and Mg2+-free PBS containing 0,2 mM EDTA and 0,025% Tween 80. Cultures were filtered through a 5 μm-pore-size filter (Millipore, USA) to remove clumps. To estimate the CFU content in the filtrate, 20 μl from each 5-fold serial dilution was plated onto Dubos agar (BD), and the total number of micro-colonies in the spot was calculated under an inverted microscope (200x magnification) after being cultured for 3 days at 37°C. The bulk of the filtered culture was stored at 4°C, and it was found that no change in the CFU content occurred during this storage period.
Mice
C57BL/6Ycit (B6) and I/StSnEgYCit strain (I/St) mice were kept under conventional, non-SPF conditions in the Animal Facilities of the Central Research Institute of Tuberculosis (CIT, Moscow, Russia) in accordance with the guidelines from the Russian Ministry of Health # 755, and under the NIH Office of Laboratory Animal Welfare (OLAW) Assurance #A5502-11. Female mice aged 2.5–3.0 months were used. All experimental procedures were approved by the Bioethics Committee of the Central Research Institute of Tuberculosis (IACUC), protocols # 2, 3, 7, 8, 11 approved on March 6, 2016.
Infection of Mice, RNA Extraction
To infect mice, mycobacteria were re-suspended in supplemented PBS. Mice were infected via respiratory tract with ~100 viable CFU/mouse using an Inhalation Exposure System (Glas-Col, USA), as described in Radaeva et al. (2005, 2008). The size of challenging dose was confirmed in preliminary experiments by plating serial 2-fold dilutions of 2-ml homogenates of the whole lungs obtained from B6 and I/St females at 2 h post-exposure onto Dubos agar and counting colonies after 3-wk incubation at 37°C. To assess CFU counts, lungs from individual mice were homogenized in 2.0 ml of sterile saline, and 10-fold serial dilutions were plated on Dubos agar and incubated at 37°C for 20–22 days.
To extract total RNA, lungs of infected mice were homogenized in Trizol reagent (Life Technologies, USA), and RNA was isolated according to a standard protocol using BeadBeater with 0.1 mm zirconia beads (BioSpec Products, USA) (Rustad et al., 2009).
Infection of Peritoneal Macrophages, iNOS Activation, RNA Extraction
To obtain peritoneal macrophages, B6 mice were injected intra-peritoneally with 3% peptone (Sigma-Aldrich) in saline. Five days later, peritoneal exudate cells (PEC) were eluted from the peritoneal cavities with Ca2+- and Mg2+-free PBS supplemented with 2% FCS and 10 U/ml heparin, washed twice with PBS, and resuspended in RPMI 1640 containing 5% FCS, 10 mM HEPES and 2 mM L-glutamine. The content of nonspecific esterase-positive cells in PEC exceeded 85%. PEC were plated onto 90 mm Petri dishes (Costar, Corning Inc., USA) at 10 × 106 cells/dish in 10 ml of RPMI-1640 containing 5% FCS, 10 mM HEPES and 2 mM L-glutamine to obtain macrophage monolayers. The cells were allowed to adhere for 2 h at 37°C, 5% CO2 before mycobacteria were added in 10 ml of supplemented RPMI-1640 at MOI = 20, 15, and 5 for 2-, 4- and 24-h incubation, respectively. Macrophage-free mycobacterial cultures served as controls.
To activate macrophages, monolayers were treated with murine rIFN-γ (100 U/ml, Sigma) for 14 h before adding mycobacteria. To block iNOS, 100 μM L-NIL (Sigma) was added 1 h before rIFN-γ administration.
To extract RNA, dishes with cell monolayers were gently shaken, culture medium was completely aspirated and macrophages were lysed with 5 ml/dish of Trizol (Invitrogen) as recommended by the manufacturer. Mycobacteria alone in control cultures were suspended by pipetting and centrifuged at 3,000 g, 20 min, 4°C. Pellets were suspended in 1 ml of Trizol. RNA was isolated by bacterial cell disruption and phenol-chloroform extraction as described in Rustad et al. (2009).
Statistics
Statistical analysis was performed using ANOVA test and t-test by GraphPad Prism6.0 software (GraphPad Software, San Diego, CA, USA). P < 0.05 was considered statistically significant.
Results
MTS1338 Expression in TB-Infected Mice
Earlier it was demonstrated that several M. tuberculosis non-coding RNAs, including MTS1338, are highly transcribed in vivo (Arnvig et al., 2011; Ignatov et al., 2014). Here, we investigated the dynamical transcription profile of MTS1338 in mycobacteria persisting in the mouse lungs from initial to terminal phases of infection. Aerosol infection with low doses of WT strain leads to a chronic and temporary effectively controlled infection in genetically resistant B6 mice, whilst in susceptible I/St mice fatal pulmonary pathology develops relatively rapidly (Kondratieva et al., 2010). Differences in mycobacterial lung CFU counts between I/St and B6 mice reached about 1.2 logs during the first 2 months post challenge and remained stable until I/St mice succumbed to infection (Figure 1A). We isolated total RNA from the mouse lungs, and profiled the MTS1338 expression in the lung mycobacterial population by quantitative real-time PCR (Figure 1B). The highest level of expression was observed at week 10 post-challenge. In B6 mice, it remained high throughout the experiment, although slowly decreased at the very late phase of infection. At week 10 of infection, when I/St mice start to lose control of the disease progression, the level of MTS1338 expression in their lung mycobacterial population was significantly higher (P < 0.01) than that in more resistant B6 mice (Figure 1B). Overall, at the stage of flourishing infection, the MTS1338 expression level in the lung-residing bacteria was more then 1,000-fold higher compared to its expression level during stationary phase of growth in vitro (Ignatov et al., 2014).
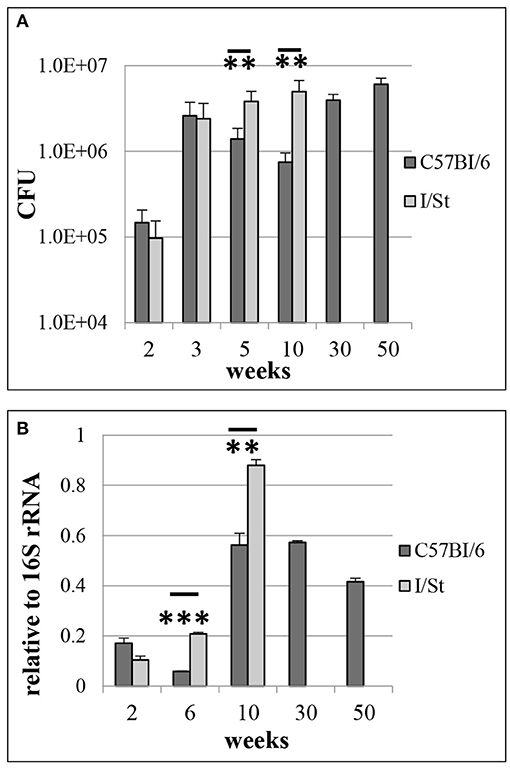
Figure 1. M. tuberculosis WT infection in resistant B6 and susceptible I/St mice. (A) Lung CFU counts along the disease progression (**P < 0.01 at 5- and 10-weeks post challenge, ANOVA). (B) MTS1338 expression levels of at different time points (**P < 0.01 and ***P < 0.001, unpaired t-test). At indicated time points, samples of total RNA were analyzed by quantitative real-time PCR, and the MTS1338 expression levels were normalized to those of 16S rRNA (ΔCt data). The data are presented as the mean ± SD of three independent experiments.
The Expression of MTS1338 Is Regulated by iNOS
Our in vivo experiments demonstrated that the level of MTS1338 expression peaks at the stage of fully developed adaptive immune response against mycobacteria. At this stage, B6 mice display significantly higher levels of IFN-γ production compare to their I/St counterparts (Eruslanov et al., 2004; Radaeva et al., 2005; Logunova et al., 2015). Since IFN-γ is the key cytokine activating macrophages for intracellular mycobacterial killing (Cooper, 2009), we compared MTS1338 expression levels in peritoneal B6 macrophages, infected WT, either activated by the external IFN-γ, or not. The level of MTS1338 expression was assessed in dynamics at 2, 4, and 24 hours of macrophage infection (Figure 2A). In IFN-γ-activated macrophages, MTS1338 expression was significantly (P < 0.001, unpaired t-test) higher than in control macrophages at every time point, and the difference reached more than 10-fold at 24 hours post infection. Thus, pre-activation of macrophages with IFN-γ induced up-regulation of the MTS1338 expression in engulfed mycobacteria. Given that the efficacy of mycobacterial killing by peritoneal macrophages significantly increases in the presence of IFN-γ (Majorov et al., 2003), this result suggests that the level of MTS1338 expression correlates with the level of pressure emanating from macrophage antibacterial systems.
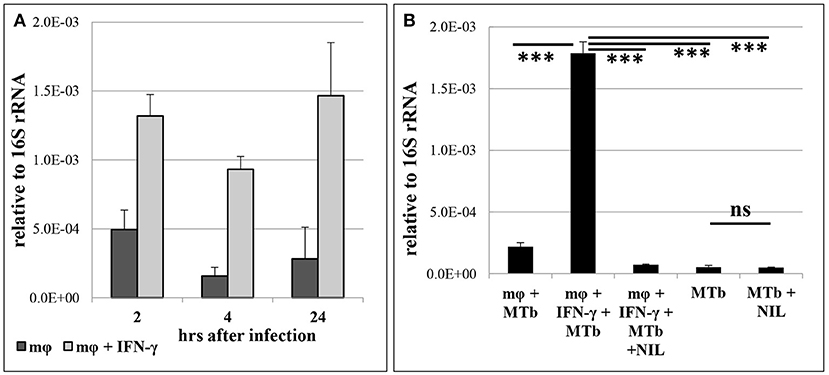
Figure 2. MTS1338 transcription is NO-dependent and correlates with activation of infected macrophages. (A) The MTS1338 transcription dynamics in peritoneal macrophages of B6 mice infected by WT strain. (B) The level of MTS1338 transcription at 24 h post infection: control (mφ +MTb), IFN-γ-activated (mφ +MTb + INF-γ), IFN-γ-activated and L-NIL treated (mφ +MTb + INF-γ + NIL). The levels of MTS1338 transcription in pure WT cultures (MTb) and L-NIL-treated cultures (MTb + NIL) serve as controls for the assessment of possible L-NIL influence onto cultured mycobacteria. The data calculated as ΔCt are presented as the mean ± SD of three independent experiments; ***P <0.005, ns – not significant, unpaired t-test).
Since the active nitrogen oxidative derivatives serve as the major trigger of MTS1338 transcription activation in vitro (Moores et al., 2017), we decided to test whether this is true for the infected macrophage system. Nitrogen oxidative derivatives production in macrophages depends upon inducible NO-synthase (iNOS2), thus we compared mycobacteria-infected IFN-γ-activated and control macrophages cultured for 24 hours in the presence or absence of L-NIL [N6-(1-iminoethyl)-L-lysine hydrochloride]—a selective inhibitor of iNOS2. Inhibition of NO production in IFN-γ-activated macrophages completely abrogated elevation in the MTS1338 expression. L-NIL itself did not affect MTS1338 expression in pure WT cultures (Figure 2B). Thus, in macrophages, nitrogen oxidative derivatives are an important trigger of MTS1338 expression.
Survival Under in vitro Stresses
To check whether elevated transcription of MTS1338 protects M. tuberculosis against hostile stressful environment, we compared survival of the OVER and pMV strains in cultures subjected to different type of stresses: low pH or elevated levels of NO and H2O2. Inhibitory effects of external NO, H2O2 and pH = 5.5 on mycobacteria were estimated by the level of incorporation of [3H]-uracil after 24 and 48 h of stress exposure (Figure 3). Uracil incorporation directly and strongly (ρ > 99) correlates with CFU counts in mycobacterial cultures (Hu et al., 2000; Majorov et al., 2003). In the absence of stress, the OVER strain grew slightly slower than the pMV one (P < 0.01 at the 48-h time point), which is consistent with earlier observations (Arnvig et al., 2011; Ignatov et al., 2015). Treatment of cultures with external NO or H2O2 had marginal to no effect on mycobacterial growth. However, overexpression of MTS1338 provided significant level of protection against acidic conditions: at pH = 5.5, uracil incorporation by the OVER strain was significantly higher both at 24 h (P < 0.05), and 48 h (P < 0.01) of culturing.
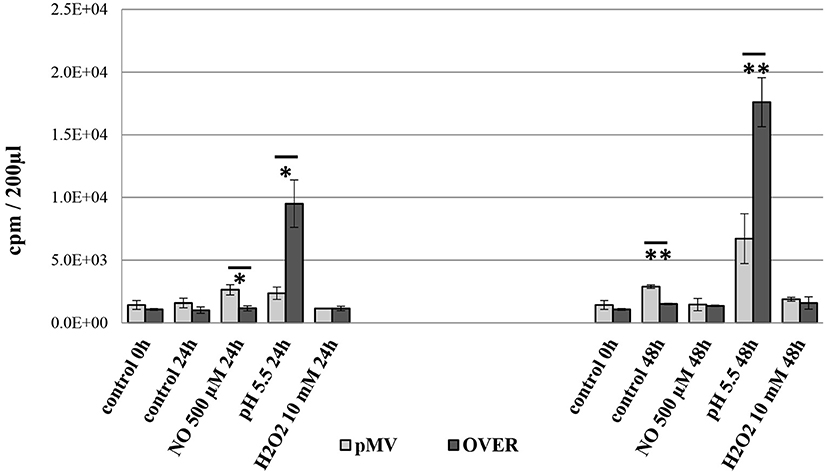
Figure 3. Viability of the OVER and pMV M. tuberculosis strains under stressful conditions in vitro. Stationary phase mycobacteria were subjected to pH = 5.5 or elevated levels of NO and H2O2 in 24- and 48-h cultures. The effect of stresses was measured by [3H]-uracil incorporation in three independent experiments and expresses as mean CPM ± SD. *P < 0.05, **P < 0.01, unpaired t-test). The data are presented as the mean ± SD of three independent experiments.
Transcriptome Changes Induced by the MTS1338 Overexpression Are Consistent With Mycobacterial Adaptation to Persistence
To assess how overexpression of MTS1338 influences mycobacterial adaptation, we compared transcriptomes of the OVER and pMV strains at the phase of stationary growth in liquid culture using RNA-seq approaches. The MTS1338 expression level in the OVER strain in these experiments was more than 10-fold higher compared to the pMV strain as confirmed by qRT-PCR (Supplementary Figure 1A).
Mapping the processed reads against the reference M. tuberculosis H37Rv genome (AL123456.3, http://www.ncbi.nlm.nih.gov/), provided the following numbers of mapped reads: 18940746 (96.91%), 19931958 (97.27%) and 20528207 (97.68%) for the OVER strains and 18671706 (97.04%), 16003598 (96.36%) and 15540058 (96.85%) for the pMV strain. The percentage of the protein-encoding part of the genome deduced from all mapped reads comprised 13.97% (2609191), 20.04% (3207752) and 15.52% (2411509) for pMV, and 10.9% (2065273), 15.39% (3067710), and 20.11% (4127293) for OVER (12.8 × 106 reads).
Using the software package DESeq2 (Love et al., 2014), we identified genes the expression of which differed between the two strains. Unexpectedly, only 28 genes were found to change their expression more then 1.5-fold under the MTS1338 overexpression conditions, with 15 genes demonstrating a decreased and 13 genes an increased expression. Further ascribing of genes to functional categories was performed using the Mycobrowser database. The list of differentially expressed genes (DEGs) is displayed in Table 1. Complete data on RNA-seq are displayed in Supplementary Table 2. Possible functional consequences of particular shifts in gene expression profiles are provided in the Discussion section. Differential expression of six randomly chosen genes was confirmed by the quantitative RT-PCR (Supplementary Figure 1B).
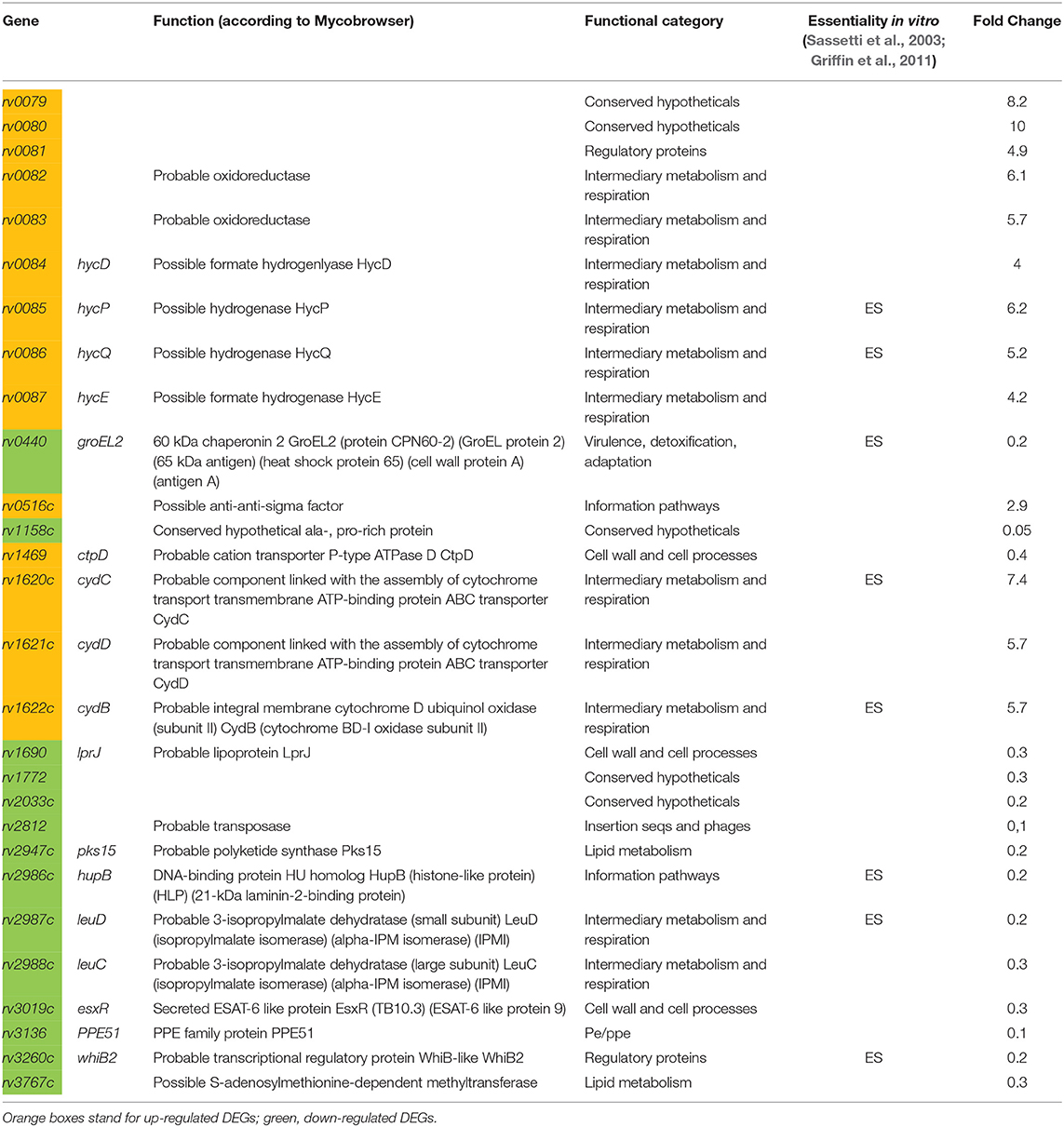
Table 1. List of genes, differentially expressed in OVER strain vs. pMV strain.
Discussion
In mycobacteria, sRNAs have been discovered much later then in many other bacterial species (Haning et al., 2014), and their functions mostly remain unknown. However, recent high-throughput transcriptional profiling of cultured M. tuberculosis exposed to relevant stresses identified a pool of both known and novel mycobacterial sRNAs involved in response to stress conditions in vitro (Gerrick et al., 2018).
Here, we present functional characteristics of the sRNA MTS1338, one of highly expressed in M. tuberculosis during the stationary growth phase (Arnvig et al., 2011) and at dormancy (Ignatov et al., 2015), suggesting its role in the maintenance of M. tuberculosis survival under unfavorable conditions. Since these observations suggest that high levels of MTS1338 expression are required for its functional activity, we constructed the M. tuberculosis strain overexpressing MTS1338 for identification of its in vitro phenotype, as well as transcriptional changes triggered by this small RNA. Earlier it was demonstrated that MTS1338 expression is NO-inducible and is activated by transcriptional regulator DosR under hypoxic cultural conditions (Moores et al., 2017), as well as under starvation, oxidative and low pH stresses (Gerrick et al., 2018). Our experiments demonstrate that the strain constitutively overexpressing MTS1338 is more resistant to low pH than the control (pMV) strain (Figure 3).
Overexpression of MTS1338 dramatically changes the bacterial growth rate (Arnvig et al., 2011; Ignatov et al., 2015). Our experiments with M. tuberculosis dormancy and resuscitation in vitro demonstrated that MTS1338 participates in entering dormancy (Ignatov et al., 2015), but is not involved in the resuscitation process (Salina et al., 2019). In vivo, high levels of MTS1338 transcription were reported for M. tuberculosis residing in chronically infected mouse lungs (Arnvig et al., 2011; Ignatov et al., 2014). In the present work, using a mouse model of infection, we demonstrate that MTS1338 up-regulation strictly follows activation of iNOS in macrophages. Importantly, at the stage of advanced infection the level of expression was significantly higher in genetically TB-susceptible I/St mice compared to more resistant B6 animals. This may reflect an attempt of mycobacteria residing in the I/St lungs to rapidly turn down metabolism, facing severe functional failure in the surrounding tissue, providing aggressive, highly hypoxic and necrotic conditions to a large proportion of mycobacterial population (Kondratieva et al., 2010). We anticipated that an abundant expression of MTS1338 leads to shifts of the whole genome transcriptional profile toward preparation of mycobacteria to stress-induced metabolic slowdown, thereby helping survival in hostile intra-macrophage surrounding.
However, RNA-seq transcriptome evaluation demonstrated that the number of DEGs in MTS1338-overexpressing and pMV strains is relatively small. MTS1338 overexpression resulted in elevated expression of three operons—rv0079-rv0081, rv0082-rv0087, and rv1620c-rv1622c. rv0079-0081 genes belong to the DosR regulon which activates under hypoxic conditions (Voskuil et al., 2003). rv0079 expression was shown to be regulated by rv0081 (Chauhan et al., 2011). In E. coli and M. bovis, homologous protein significantly inhibits cell growth, apparently interacting with the 30S ribosome subunit and inhibiting translation—the phenotype typical for transition to dormancy (Kumar et al., 2012). rv0080 encodes a conservative hypothetical protein with unknown functions. It contains a domain of pyridoxine 5′-phosphate (PNP) oxidase-like (PNPOx-like) superfamily, which catalyze flavin mononucleotide-mediated redox reactions. rv0081 is one of two key transcriptional factors mediating early response to hypoxia (Galagan et al., 2013). As an important “metabolic hub” working in concert with other transcription regulators, rv0081 is associated with the processes of lipid metabolism, protein degradation, and cholesterol biosynthesis (Rodriguez et al., 2014; Aguilar-Ayala et al., 2017).
The genes of other two operons encode proteins of the functional category “Intermediary metabolism and respiration.” rv0082-rv0087 genes are also regulated by rv0081 (He et al., 2011), but not included in the DosR regulon. The rv0082–rv0087 locus in M. tuberculosis encodes a putative [NiFe]-hydrogenase complex (Berney et al., 2014). In E. coli, homologous proteins are involved in the conversion of formate to CO2 and H2 under conditions of anaerobic respiration in the absence of an external terminal electron acceptor (Leonhartsberger et al., 2002). Facultative H2 metabolism is central for mycobacterial persistence. Mycobacteria enhance long-term survival by up-regulating hydrogenases during energy and oxygen limitations (Greening and Cook, 2014).
rv1620c-rv1622c (cydC, D, B respectively) encode proteins, which are involved in the cytochrome biogenesis and active transport across the membrane of components involved in the assembly of cytochrome. The expression of CydDC is linked to the incorporation of heme cofactors into a variety of periplasmic cytochromes, as well as the bd-type respiratory oxidases. CydB is the component of the aerobic respiratory chain that is supposedly predominant when cells are grown at low aeration, and is up-regulated under low pH (Baker et al., 2014). It has been reported that the presence of bd-type oxidases is correlated with bacterial virulence. For example, growth of mycobacteria at low oxygen tensions enhances both the expression of a bd-type oxidase and cell invasion (Bermudez et al., 1997).
Down-regulated genes belong to different functional categories. Among them, three genes with chaperone functions attract special attention. All these genes are essential for M. tuberculosis growth in vitro (Sassetti et al., 2003; Griffin et al., 2011). rv0440 encodes GroEL2, the chaperone belonging to the HSP60 family. Its chaperone-like functions provide resistance to stress (Qamra et al., 2004) and modulate host immune responses (Lewthwaite et al., 2007; Naffin-Olivos et al., 2014). GroEL2 is highly induced in response to environmental cues during infection like heat shock, oxidative stress, growth in macrophages, and hypoxia (Qamra et al., 2005). The HupB protein encoded by rv2986c belongs to the histone-like family of prokaryotic DNA-binding proteins capable of wrapping DNA to stabilize it, and prevent DNA denaturation under extreme environmental conditions (Kumar et al., 2010). It is involved in controlling the transfer of mycolic acids to sugars by the Ag85 complex (Katsube et al., 2007), as well as siderophore biosynthesis, and is essential for mycobacteria growth in macrophages (Pandey et al., 2014).
WhiB2 encoded by rv3260c belongs to the WhiB family of transcriptional regulators. Its apo-form displays a chaperone activity, preventing aggregation and providing correct refolding of proteins; this activity does not require ATP and is independent of its own oxidized or reduced status and co-chaperones (Konar et al., 2012). The homolog of whiB2 in M. smegmatis, WhmD, participates in septa formation during cell division. WhmD overexpression decreases the linear size of M. smegmatis cells (Raghunand and Bishai, 2006). In addition, the newly formed cell walls are more susceptible to lysis (Gomez and Bishai, 2000). It was suggested that WhiB2 is involved in the assembly and stabilization of the FtsZ ring around the cell septum during division (Huang et al., 2013).
Two other down-regulated genes, rv2987c and rv2988c, encode subunits of putative 3-isopropylmalate dehydratase and are involved in leucine biosynthesis. In vivo and in vitro studies demonstrate that leucine-auxotrophic M. tuberculosis strains do not replicate inside host cells (Hondalus et al., 2000).
Overall, up-regulated genes fall into “intermediate metabolism and respiration” functional category, and either belong to the DosR regulon directly (rv0079-rv0081), or are connected through DosR-regulated transcriptional factor rv0081 (rv0082-rv0087). All up-regulated genes are thought to be involved into mycobacterial survival inside macrophages.
The list of down-regulated genes is functionally more diverse. Of interest is a decreased of genes encoding chaperone proteins, such as GroEL2, HupB, WhiB2. Theoretically, their expression should be rather increased under unfavorable conditions. To find an explanation of this paradox, we studied TB databases (http://genome.tbdb.org), concentrating on groups of genes co-expressed with these chaperones genes. It appeared that the majority of DEGs, including groEL2, hupB, and whiB2, are co-expressed with genes of the functional category J: Translation, ribosomal structure and biogenesis (Clusters of Orthologous Groups, COG, Tatusov et al., 2000). In all cases, irrespective to up- or down-regulation of DEGs, there was reversed correlation with genes of the J category (Table 2). These data suggest that the MTS1338 overexpression leads to transcriptional changes that correlate with a translation slowdown.

Table 2. DEGs, and their correlation with expression of different functional categories (according to COG).
Summarizing, our results suggest an important potential role of MTS1338 in pathogenesis of mycobacteria-triggered diseases. An increase in MTS1338 production during infection in vivo and in activated macrophages, changes in the expression of genes important for mycobacterial metabolism and a better survival under low pH accompanying MTS1338 overexpression—all suggest that this sRNA may well contribute to successful persistence of M. tuberculosis within host cells.
Data Availability Statement
The data sets supporting the results of this article are available in the GEO data repository under the accession number GSE137857.
Ethics Statement
The animal study was reviewed and approved by Bioethics Committee of the Central Research Institute of Tuberculosis (IACUC).
Author Contributions
ES, DI, AK, AA, and TA conceived the idea and designed the experiments. ES, AG, YS, KM, OB, AO, and NL performed the experiments. ES, AK, AA, and TA analyzed the data and wrote the paper.
Funding
This work was supported by the Russian Science Foundation grant no. 18-15-00332 to TA (new M. tuberculosis strains, RNA-seq experiments and analyses, stresses in vitro, infection analyses); grant no. 18-45-04015 to AA (in vivo and macrophages infection experiments).
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fcimb.2019.00405/full#supplementary-material
Supplementary Table 1. Oligonucleotides used in the study.
Supplementary Table 2. RNA-seq data.
Supplementary Figure 1. (A) MTS1338 transcription level in samples used for RNA-seq. pMV stands for control M. tuberculosis strains. Cl2, cl8, cl4—OVER strains, independent clones. (B) Validation of RNA-seq data by qRT-PCR.
References
Aguilar-Ayala, D. A., Palomino, J. C., Vandamme, P., Martin, A., and Gonzalez-Y-Merchand, Y. A. (2017). Genetic regulation of Mycobacterium tuberculosis in a lipid-rich environment. Infect. Genet. Evol. 55, 392–402. doi: 10.1016/j.meegid.2016.10.015
Arnvig, K., and Young, D. (2012). Non-coding RNA and its potential role in Mycobacterium tuberculosis pathogenesis. RNA Biol. 9, 427–436. doi: 10.4161/rna.20105
Arnvig, K. B., Comas, I., Thomson, N. R., Houghton, J., Boshoff, H. I., Croucher, N. J., et al. (2011). Sequence-based analysis uncovers an abundance of non-coding RNA in the total transcriptome of Mycobacterium tuberculosis. PLoS Pathog. 7:e1002342. doi: 10.1371/journal.ppat.1002342
Baker, J. J., Johnson, B. K., and Abramovitch, R. B. (2014). Slow growth of Mycobacterium tuberculosis at acidic pH is regulated by phoPR and host-associated carbon sources. Mol. Microbiol. 94, 56–69. doi: 10.1111/mmi.12688
Bermudez, L. E., Petrofsky, M., and Goodman, J. (1997). Exposure to low oxygen tension and increased osmolarity enhance the ability of Mycobacterium avium to enter intestinal epithelial (HT-29) cells. Infect. Immun. 65, 3768–3773.
Berney, M., Greening, C., Hards, K., Collins, D., and Cook, G. M. (2014). Three different [NiFe] hydrogenases confer metabolic flexibility in the obligate aerobe Mycobacterium smegmatis. Environ. Microbiol. 16, 318–330. doi: 10.1111/1462-2920.12320
Carver, T., Harris, S. R., Berriman, M., Parkhill, J., and McQuillan, J. A. (2012). Artemis: an integrated platform for visualization and analysis of high-throughput sequence-based experimental data. Bioinformatics 28, 464–469. doi: 10.1093/bioinformatics/btr703
Chauhan, S., Sharma, D., Singh, A., Surolia, A., and Tyagi, J. S. (2011). Comprehensive insights into Mycobacterium tuberculosis DevR (DosR) regulon activation switch. Nucleic Acids Res. 39, 7400–7414. doi: 10.1093/nar/gkr375
Connell, N. D. (1994). Mycobacterium: isolation, maintenance, transformation, and mutant selection. Methods Cell Biol. 45, 107–125. doi: 10.1016/S0091-679X(08)61848-8
Cooper, A. M. (2009). Cell-mediated immune responses in tuberculosis. Annu. Rev. Immunol. 27, 393–422. doi: 10.1146/annurev.immunol.021908.132703
Dutta, T., and Srivastava, S. (2018). Small RNA-mediated regulation in bacteria: a growing palette of diverse mechanisms. Gene 656, 60–72. doi: 10.1016/j.gene.2018.02.068
Eruslanov, E. B., Majorov, K. B., Orlova, M. O., Mischenko, V. V., Kondratieva, T. K., Apt, A. S., et al. (2004). Lung cell responses to M. tuberculosis in genetically susceptible and resistant mice following intratracheal challenge. Clin. Exp. Immunol. 135, 19–28. doi: 10.1111/j.1365-2249.2004.02328.x
Galagan, J. E., Minch, K., Peterson, M., Lyubetskaya, A., Azizi, E., Sweet, L., et al. (2013). The Mycobacterium tuberculosis regulatory network and hypoxia. Nature 499, 178–183. doi: 10.1038/nature12337
Ganger, M. T., Dietz, G. D., and Ewing, S. J. (2017). A common base method for analysis of qPCR data and the application of simple blocking in qPCR experiments. BMC Bioinformatics 18:534. doi: 10.1186/s12859-017-1949-5
Gerrick, E. R., Barbier, T., Chase, M. R., Xu, R., François, J., Lin, V. H., et al. (2018). Small RNA profiling in Mycobacterium tuberculosis identifies MrsI as necessary for an anticipatory iron sparing response. Proc. Natl. Acad. Sci. U.S.A. 115, 6464–6469. doi: 10.1073/pnas.1718003115
Gomez, J. E., and Bishai, W. R. (2000). whmD is an essential mycobacterial gene required for proper septation and cell division. Proc. Natl. Acad. Sci. U.S.A. 97, 8554–8559. doi: 10.1073/pnas.140225297
Greening, C., and Cook, G. M. (2014). Integration of hydrogenase expression and hydrogen sensing in bacterial cell physiology. Curr. Opin. Microbiol. 18, 30–38. doi: 10.1016/j.mib.2014.02.001
Griffin, J. E., Gawronski, J. D., DeJesus, M. A., Ioerger, T. R., Akerley, B. J., and Sassetti, C. M. (2011). High-resolution phenotypic profiling defines genes essential for mycobacterial growth and cholesterol catabolism. PLoS Pathog. 7:e1002251. doi: 10.1371/journal.ppat.1002251
Haning, K., Cho, S. H., and Contreras, L. M. (2014). Small RNAs in mycobacteria: an unfolding story. Front. Cell Infect. Microbiol. 4:96. doi: 10.3389/fcimb.2014.00096
He, H., Bretl, D. J., Penoske, R. M., Anderson, D. M., and Zahrt, T. C. (2011). Components of the Rv0081-Rv0088 locus, which encodes a predicted formate hydrogenlyase complex, are coregulated by Rv0081, MprA, and DosR in Mycobacterium tuberculosis. J. Bacteriol. 193, 5105–5118. doi: 10.1128/JB.05562-11
Holmqvist, E., and Wagner, E. G. H. (2017). Impact of bacterial sRNAs in stress responses. Biochem. Soc. Trans. 45, 1203–1212. doi: 10.1042/BST20160363
Hondalus, M. K., Bardarov, S., Russell, R., Chan, J., Jacobs, W. R., and Bloom, B. R. (2000). Attenuation of and protection induced by a leucine auxotroph of Mycobacterium tuberculosis. Infect. Immun. 68, 2888–2898. doi: 10.1128/IAI.68.5.2888-2898.2000
Hör, J., Gorski, S. A., and Vogel, J. (2018). Bacterial RNA biology on a genome scale. Mol. Cell 70, 785–799. doi: 10.1016/j.molcel.2017.12.023
Hu, Y., Mangan, J. A., Dhillon, J., Sole, K. M., Mitchison, D. A., Butcher, P. D., et al. (2000). Detection of mRNA transcripts and active transcription in persistent Mycobacterium tuberculosis induced by exposure to rifampin or pyrazinamide. J. Bacteriol. 182, 6358–6365. doi: 10.1128/JB.182.22.6358-6365.2000
Huang, K. H., Durand-Heredia, J., and Janakiraman, A. (2013). FtsZ ring stability: of bundles, tubules, crosslinks, and curves. J. Bacteriol. 195, 1859–1868. doi: 10.1128/JB.02157-12
Ignatov, D. V., Salina, E. G., Fursov, M. V., Skvortsov, T. A., Azhikina, T. L., and Kaprelyants, A. S. (2015). Dormant non-culturable Mycobacterium tuberculosis retains stable low-abundant mRNA. BMC Genomics 16:954. doi: 10.1186/s12864-015-2197-6
Ignatov, D. V., Timoshina, O. Y., Logunova, N. N., Skvortsov, T. A., and Azhikina, T. L. (2014). Expression of small RNAs of Mycobacterium tuberculosis in murine models of Tuberculosis infection. Russ. J. Bioorg. Chem. 40, 233–235. doi: 10.1134/S1068162014020058
Katsube, T., Matsumoto, S., Takatsuka, M., Okuyama, M., Ozeki, Y., Naito, M., et al. (2007). Control of cell wall assembly by a histone-like protein in Mycobacteria. J. Bacteriol. 189, 8241–8249. doi: 10.1128/JB.00550-07
Konar, M., Alam, M. S., Arora, C., and Agrawal, P. (2012). WhiB2/Rv3260c, a cell division-associated protein of Mycobacterium tuberculosis H37Rv, has properties of a chaperone. FEBS J. 279, 2781–2792. doi: 10.1111/j.1742-4658.2012.08662.x
Kondratieva, E., Logunova, N., Majorov, K., Averbakh, M., and Apt, A. (2010). Host genetics in granuloma formation: human-like lung pathology in mice with reciprocal genetic susceptibility to M. tuberculosis and M. avium. PLoS ONE 5:e10515. doi: 10.1371/journal.pone.0010515
Kumar, A., Majid, M., Kunisch, R., Rani, P. S., Qureshi, I. A., Lewin, A., et al. (2012). Mycobacterium tuberculosis DosR regulon gene Rv0079 encodes a putative, ‘dormancy associated translation inhibitor (DATIN)’. PLoS ONE 7:e38709. doi: 10.1371/journal.pone.0038709
Kumar, S., Sardesai, A. A., Basu, D., Muniyappa, K., and Hasnain, S. E. (2010). DNA clasping by mycobacterial HU: the C-terminal region of HupB mediates increased specificity of DNA binding. PLoS One 5:e12551. doi: 10.1371/journal.pone.0012551
Langmead, B., and Salzberg, S. L. (2012). Fast gapped-read alignment with bowtie 2. Nat. Methods 9, 357–359. doi: 10.1038/nmeth.1923
Leonhartsberger, S., Korsa, I., and Böck, A. (2002). The molecular biology of formate metabolism in enterobacteria. J. Mol. Microbiol. Biotechnol. 4, 269–276.
Lewthwaite, J. C., Clarkin, C. E., Coates, A. R., Poole, S., Lawrence, R. A., Wheeler-Jones, C. P., et al. (2007). Highly homologous Mycobacterium tuberculosis chaperonin 60 proteins with differential CD14 dependencies stimulate cytokine production by human monocytes through cooperative activation of p38 and ERK1/2 mitogen-activated protein kinases. Int. Immunopharmacol. 7, 230–240. doi: 10.1016/j.intimp.2006.10.005
Liao, Y., Smyth, G. K., and Shi, W. (2014). Featurecounts: an efficient general purpose program for assigning sequence reads to genomic features. Bioinformatics 30, 923–930. doi: 10.1093/bioinformatics/btt656
Logunova, N., Korotetskaya, M., Polshakov, V., and Apt, A. (2015). The QTL within the H2 complex involved in the control of Tuberculosis infection in mice is the classical class II H2-Ab1 Gene. PLoS Genet. 11:e1005672. doi: 10.1371/journal.pgen.1005672
Love, M. I., Huber, W., and Anders, S. (2014). Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 15:550. doi: 10.1186/s13059-014-0550-8
Lyadova, I. V., Eruslanov, E. B., Khaidukov, S. V., Yeremeev, V. V., Majorov, K. B., Pichugin, A. V., et al. (2000). Comparative analysis of T lymphocytes recovered from the lungs of mice genetically susceptible, resistant, and hyperresistant to Mycobacterium tuberculosis-triggered disease. J. Immunol. 165, 5921–5931. doi: 10.4049/jimmunol.165.10.5921
Mai, J., Rao, C., Watt, J., Sun, X., Lin, C., Zhang, L., et al. (2019). Mycobacterium tuberculosis 6C sRNA binds multiple mRNA targets via C-rich loops independent of RNA chaperones. Nucleic Acids Res. 47, 4292–4307. doi: 10.1093/nar/gkz149
Majorov, K. B., Lyadova, I. V., Kondratieva, T. K., Eruslanov, E. B., Rubakova, E. I., Orlova, M. O., et al. (2003). Different innate ability of I/St and A/Sn mice to combat virulent Mycobacterium tuberculosis: phenotypes expressed in lung and extrapulmonary macrophages. Infect. Immun. 71, 697–707. doi: 10.1128/IAI.71.2.697-707.2003
Moores, A., Riesco, A. B., Schwenk, S., and Arnvig, K. B. (2017). Expression, maturation and turnover of DrrS, an unusually stable, DosR regulated small RNA in Mycobacterium tuberculosis. PLoS ONE 12:e0174079. doi: 10.1371/journal.pone.0174079
Naffin-Olivos, J. L., Georgieva, M., Goldfarb, N., Madan-Lala, R., Dong, L., Bizzell, E., et al. (2014). Mycobacterium tuberculosis Hip1 modulates macrophage responses through proteolysis of GroEL2. PLoS Pathog. 10:e1004132. doi: 10.1371/journal.ppat.1004132
Pandey, S. D., Choudhury, M., Yousuf, S., Wheeler, P. R., Gordon, S. V., Ranjan, A., et al. (2014). Iron-regulated protein HupB of Mycobacterium tuberculosis positively regulates siderophore biosynthesis and is essential for growth in macrophages. J. Bacteriol. 196, 1853–1865. doi: 10.1128/JB.01483-13
Qamra, R., Mande, S. C., Coates, A. R., and Henderson, B. (2005). The unusual chaperonins of Mycobacterium tuberculosis. Tuberculosis 85, 385–394. doi: 10.1016/j.tube.2005.08.014
Qamra, R., Srinivas, V., and Mande, S. C. (2004). Mycobacterium tuberculosis GroEL homologues unusually exist as lower oligomers and retain the ability to suppress aggregation of substrate proteins. J. Mol. Biol. 342, 605–617. doi: 10.1016/j.jmb.2004.07.066
Radaeva, T. V., Kondratieva, E. V., Sosunov, V. V., Majorov, K. B., and Apt, A. (2008). A human-like TB in genetically susceptible mice followed by the true dormancy in a Cornell-like model. Tuberculosis 88, 576–585. doi: 10.1016/j.tube.2008.05.003
Radaeva, T. V., Nikonenko, B. V., Mischenko, V. V., Averbakh, M. M. Jr., and Apt, A. S. (2005). Direct comparison of low-dose and Cornell-like models of chronic and reactivation tuberculosis in genetically susceptible I/St and resistant B6 mice. Tuberculosis 85, 65–72. doi: 10.1016/j.tube.2004.09.014
Raghunand, T. R., and Bishai, W. R. (2006). Mycobacterium smegmatis whmD and its homologue Mycobacterium tuberculosis whiB2 are functionally equivalent. Microbiol. 152, 2735–2747. doi: 10.1099/mic.0.28911-0
Rodriguez, J. G., Hernández, A. C., Helguera-Repetto, C., Aguilar Ayala, D., Guadarrama-Medina, R., Anzóla, J. M., et al. (2014). Global adaptation to a lipid environment triggers the dormancy-related phenotype of Mycobacterium tuberculosis. MBio 5:e01125–14. doi: 10.1128/mBio.01125-14
Russell, D. G. (2007). Who puts the tubercle in tuberculosis? Nat. Rev. Microbiol. 5, 39–47. doi: 10.1038/nrmicro1538
Rustad, T. R., Roberts, D. M., Liao, R. P., and Sherman, D. R. (2009). Isolation of mycobacterial RNA. Methods Mol. Biol. 465, 13–21. doi: 10.1007/978-1-59745-207-6_2
Salina, E. G., Grigorov, A., Bychenko, O., Skvortsova, Y., Mamedov, I., Azhikina, T., et al. (2019). Resuscitation of dormant “Non-culturable” mycobacterium tuberculosis is characterized by immediate transcriptional burst. Front. Cell Infect. Microbiol. 9:272. doi: 10.3389/fcimb.2019.00272
Sassetti, C. M., Boyd, D. H., and Rubin, E. J. (2003). Genes required for mycobacterial growth defined by high density mutagenesis. Mol. Microbiol. 48, 77–84. doi: 10.1046/j.1365-2958.2003.03425.x
Schwenk, S., and Arnvig, K. B. (2018). Regulatory RNA in Mycobacterium tuberculosis, back to basics. Pathog. Dis. 76:fty035. doi: 10.1093/femspd/fty035
Solans, L., Gonzalo-Asensio, J., Sala, C., Benjak, A., Uplekar, S., Rougemont, J., et al. (2014). The PhoP-dependent ncRNA Mcr7 modulates the TAT secretion system in Mycobacterium tuberculosis. PLoS Pathog. 10:e1004183. doi: 10.1371/journal.ppat.1004183
Stewart, G. R., Robertson, B. D., and Young, D. B. (2003). Tuberculosis: a problem with persistence. Nat. Rev. Microbiol. 1, 97–105. doi: 10.1038/nrmicro749
Stover, C. K., de la Cruz, V. F., Fuerst, T. R., Burlein, J. E., Benson, L. A., Bennett, L. T., et al. (1991). New use of BCG for recombinant vaccines. Nature 351, 456–60. doi: 10.1038/351456a0
Taneja, S., and Dutta, T. (2019). On a stake-out: Mycobacterial small RNA identification and regulation. Non coding RNA Res. 4, 86–95. doi: 10.1016/j.ncrna.2019.05.001
Tatusov, R. L., Galperin, M. Y., Natale, D. A., and Koonin, E. V. (2000). The COG database: a tool for genome-scale analysis of protein functions and evolution. Nucleic Acids Res. 28, 33–6. doi: 10.1093/nar/28.1.33
Keywords: Mycobacterium tuberculosis, tuberculosis, small RNA MTS1338, RNA-seq, infection
Citation: Salina EG, Grigorov A, Skvortsova Y, Majorov K, Bychenko O, Ostrik A, Logunova N, Ignatov D, Kaprelyants A, Apt A and Azhikina T (2019) MTS1338, A Small Mycobacterium tuberculosis RNA, Regulates Transcriptional Shifts Consistent With Bacterial Adaptation for Entering Into Dormancy and Survival Within Host Macrophages. Front. Cell. Infect. Microbiol. 9:405. doi: 10.3389/fcimb.2019.00405
Received: 03 October 2019; Accepted: 12 November 2019;
Published: 26 November 2019.
Edited by:
Anthony Baughn, University of Minnesota Twin Cities, United StatesReviewed by:
Subramanian Dhandayuthapani, Texas Tech University Health Sciences Center, United StatesEvgeniya V. Nazarova, Genentech, United States
Copyright © 2019 Salina, Grigorov, Skvortsova, Majorov, Bychenko, Ostrik, Logunova, Ignatov, Kaprelyants, Apt and Azhikina. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Tatyana Azhikina, dGF0YXpoaWtAaWJjaC5ydQ==
†These authors have contributed equally to this work
‡Present address: Dmitriy Ignatov, Max Planck Unit for the Science of Pathogens, Berlin, Germany