 Sandra Sánchez-Urtaza1
Sandra Sánchez-Urtaza1 Alain Ocampo-Sosa2Ainhoa Molins-Bengoetxea1
Alain Ocampo-Sosa2Ainhoa Molins-Bengoetxea1 Mohammed A. El-Kholy3
Mohammed A. El-Kholy3 Marta Hernandez4David Abad4
Marta Hernandez4David Abad4 Sherine M. Shawky5
Sherine M. Shawky5 Itziar Alkorta6
Itziar Alkorta6 Lucia Gallego1*
Lucia Gallego1*- 1Laboratory of Antibiotics and Molecular Bacteriology, Department of Immunology, Microbiology and Parasitology, Faculty of Medicine and Nursing, University of the Basque Country, Leioa, Spain
- 2Microbiology Service, University Hospital Marqués de Valdecilla, Health Research Institute (Instituto de Investigación Valdecilla), Santander, Spain and CIBERINFEC, Instituto de Salud Carlos III, Madrid, Spain
- 3Department of Microbiology and Biotechnology, Division of Clinical and Biological Sciences, College of Pharmacy, Arab Academy for Science, Technology & Maritime Transport (AASTMT), Alexandria, Egypt
- 4Laboratory of Molecular Biology and Microbiology, One Health, Agricultural Technological, Institute of Castile and Leon (ITACyL), Valladolid, Spain
- 5Medical Research Institute, Alexandria University, Alexandria, Egypt
- 6Department of Biochemistry and Molecular Biology, Faculty of Science and Technology, University of the Basque Country, Leioa, Spain
Carbapenem resistant Acinetobacter baumannii is a major global concern, especially in countries of the Middle East and North Africa, where the antibiotic resistance rates are on the rise. The aim of this study was to study the genomic characteristics and antimicrobial susceptibility profile of thirty-six multidrug resistant A. baumannii clinical isolates obtained in hospitals from Alexandria, Egypt. Antibiotic resistance rates were estimated by determination of Minimum Inhibitory Concentrations. Carbapenemase genes, other antibiotic resistance genes and virulence factors were then screened by the use of Whole Genome Sequencing. Isolates were also subjected to Multi Locus Sequence Typing (MLST) using the Pasteur Scheme and to core genome MLST to study their clonal relatedness. In addition, plasmid analysis was performed by the use of a commercial kit and S1- Pulsed Field Gel Electrophoresis, and Hybridization experiments with DIG-labeled DNA probes for blaNDM-1, blaPER-7 and blaGES-like were performed to locate these genes. The majority of isolates were resistant to β-lactams (including carbapenems), fluoroquinolones, aminoglycosides and trimethoprim; and some showed resistance to cefiderocol and minocycline. We identified 8 different blaOXA-51-like variants including blaOXA-51, blaOXA-64, blaOXA-65, blaOXA-66, blaOXA-68, blaOXA-91, blaOXA-94 and blaOXA-336; blaOXA-23, blaNDM-1, blaPER-7, blaGES-like and blaADC-like and other antibiotic resistance genes, some of these genes were within transposons or class 1 integrons. Multiple virulence factors responsible for adherence, biofilm production, type II and type VI secretion systems, exotoxins, exoenzymes, immune modulation and iron uptake were observed and 34 out of 36 isolates showed motility. Thirty-five out of 36 isolates clustered with International Clones 2, 4, 5, 7, 8 and 9; and 9 STs were identified including ST570, ST2, ST600, ST15, ST113, ST613, ST85, ST158, ST164. Plasmids ranging in size from 1.7 to 70 kb were found; blaNDM-1 and blaPER-7 genes were located in the chromosome and blaGES-like genes were simultaneously located in the chromosome and in a plasmid of 70kb. In conclusion, this study revealed a wide spectrum of antibiotic resistance genes and a variety of lineages among A. baumannii isolated in hospitals from Alexandria, and highlights the importance of investigating the molecular epidemiology to control the spread of multi-drug resistant isolates.
1 Introduction
Carbapenem resistant Acinetobacter baumannii (CRAB) is one of the major global threats for healthcare settings worldwide, as there are only few antibiotics effective to treat the infections caused by these isolates due to its high rates of antimicrobial resistance (Ejaz et al., 2021). This pathogen is responsible for multiple nosocomial infections such as bloodstream infections, urinary tract infections, wound infections, ventilator-associated pneumonia and other respiratory tract infections, meningitis and bacteremia (Kurihara et al., 2020). As a consequence, A. baumannii is heading the World Head Organization’s (WHO) list of critical pathogens for which new antibiotics are urgently needed (El-Kholy et al., 2021). Many other global institutions such as the European Centre for Disease Prevention and Control (ECDC), Infectious Diseases Society of America (IDSA) and the Center for Disease Control and Prevention of America (CDC) have also declared it an urgent threat (Mea et al., 2021).
Rates of mortality and disability caused by A. baumannii infections are increasing. Retrospective studies showed that the mortality rates associated with A. baumannii infections are ranging from 22.8% to 49.6% in the United States (US), and from 29% to 71.6% in Europe (Patel et al., 2019). According to autores, mortality associated with A. baumannii causing hospital-acquired and ventilator-associated pneumonia was higher in Western Asia (56.2%), Southern Europe (55.7%) and Northern Africa (53.3%). Countries of the Mediterranean area, such as Greece (68.2%), Turkey (61.4) or Egypt (53.3%) were the ones with the highest reported mortality rates (Mohd Sazlly Lim et al., 2019). Indeed, in Egypt 30–100% of A. baumannii isolates are considered as Multidrug-Resistant (MDR), and carbapenem resistance was reported in 26.6–100% of A. baumannii isolates (El-Kholy et al., 2021). Furthermore, many reports showed that the COVID-19 pandemic increased CRAB infection rates, for instance in the US, the rates in hospitals increased a 78% and overall by 35% in 2020 compared with 2019 (Centers for Disease Control and Prevention, 2022). In Egypt, MDR A. baumannii was the second most common cause of infection (27.4%) among mechanically ventilated patients during the second wave of COVID-19 (Elwakil et al., 2023). The economic impact is also remarkable in CRAB infections, for instance in the United States, the CDC reported that treatment costs were around $281 million in 2019 (Ejaz et al., 2021).
Different resistance mechanisms are commonly found in A. baumannii including antibiotic inactivation enzymes, alteration of target sites, overexpression of efflux pumps and loss of porins (Chakravarty, 2020). Carbapenemases are the main carbapenem resistance mechanism in A. baumannii, being carbapenem-hydrolyzing oxacilinases the most important ones. However, during the last years, class B metallo-β-lactamases such as New Delhi Metallo-Beta-lactamases (NDM) are on the rise (Xanthopoulou et al., 2020).
Nine major International Clones (IC1-9) of A. baumannii, have been described up to now, being IC2/CC92 with the acquired blaOXA-23 gene the most disseminated lineage worldwide (Al-Hassan et al., 2019; Al-Hassan et al., 2021); and IC1 and IC2 in Europe (Muthuirulandi Sethuvel et al., 2019). In the Middle East and North Africa, A. baumannii clinical outbreaks, caused by MDR isolates endemically producing carbapenemases NDM-1, NDM-2 and OXA-23, are usually poly-clonal but with dominance of IC2 lineage (Hassan et al., 2021). Although IC2 is the most disseminated clone in Northern Africa, isolates belonging to IC5 and IC9 are also reported (Al-Hassan et al., 2021; Hamed et al., 2022). From 2013 to date, isolates belonging to Oxford ST208 (IC2), are commonly reported in Egyptian hospitals (Al-Hassan et al., 2019). However, in spite of the alarming situation, there is little information about A. baumannii in Egypt and more studies are needed (Hassan et al., 2021). Nevertheless, the limited resources for research in low- and middle-income countries such as Egypt, makes it difficult to obtain epidemiological data. It is of high concern to investigate the molecular epidemiology to control the dissemination of these clinically important isolates.
The aim of the present study was to characterize the genetic features, to study the molecular epidemiology and to identify the antimicrobial susceptibility profiles of thirty-six carbapenem resistant A. baumannii clinical isolates obtained from hospitals from Alexandria, Egypt.
2 Materials and methods
2.1 Bacterial isolates, species identification and antimicrobial susceptibility testing
Thirty-six A. baumannii isolates were collected from August 2020 to February 2021 in hospitals from Alexandria, Egypt and processed and identified in the Medical Research Institute of Alexandria. The bacterial isolates were from 21 male and 15 female patients. Clinical samples were obtained from the following sources: bronchoalveolar lavage (14), swab (8), blood (5), aspirate (3), sputum (3), endotracheal tube (1), urine (1) and tissue (1).
Species identification was assessed by VITEK 2® automated system (Biomérieux, Marcy-l’Étoile, France) and gyrB multiplex PCR (Higgins et al., 2010b). Minimum Inhibitory Concentrations (MICs) to ticarcillin, ticarcillin/clavulanic acid, piperacillin, piperacillin/tazobactam, ceftazidime, cefepime, amikacin, gentamicin, tobramycin, minocycline, ciprofloxacin, trimethoprim/sulfamethoxazole, imipenem, meropenem and colistin were determined by the use of VITEK 2® automated system. Antimicrobial activity of cefiderocol was determined by disk diffusion method using 30 µg cefiderocol discs (ThermoFisher Scientific, Waltham, United States) following the EUCAST guidelines and clinical breakpoints (Versions 10.0 and 12.0, January 2022). Escherichia coli ATCC 25922 and Pseudomonas aeruginosa ATCC 27853 were used as control strains.
2.2 Detection of carbapenemase genes
Carbapenemase-encoding genes were analyzed by multiplex PCR including primers for: blaOXA-23-like, −40-like, −51-like, −58-like, −143-like, and −235-like (Woodford et al., 2006; Higgins et al., 2010a; Higgins et al., 2013). Two additional multiplex PCR were performed to investigate the presence of blaVIM, blaKPC, blaNDM, blaOXA-48, blaIMI, blaGES, blaGIM, blaIMP and ISAba-1/blaOXA-51-like (Cerezales et al., 2021).
2.3 Whole Genome Sequencing, genome annotation, analysis and visualization and virulence factors analysis
Total DNA was purified with the DNeasy Blood and Tissue Kit (Qiagen, Hilden, Germany) and sequenced on a MiSeq device using reagents kit v3 for 2×300 paired-end libraries (Illumina) as previously described (Hernández et al., 2017). Raw reads from the sequencing platform were directly analyzed using the in-house bioinformatics pipeline TORMES® (Quijada et al., 2019). A. baumannii ATCC 17978 was used as reference strain. The options used in this study included quality control and filtering of the reads by using Trimmomatic (Bolger et al., 2014), Prinseq (Schmieder and Edwards, 2011) and Kraken (Wood and Salzberg, 2014). Genome assembly was performed with SPAdes (Bankevich et al., 2012) and Quast (Gurevich et al., 2013) and genome annotation with Prokka software tool (Seemann, 2014). The whole-genome shotgun sequences of the isolates generated for this study were deposited and can be found in GenBank under the BioProject accession number PRJNA856145 and the accession numbers of each isolate are detailed in Supplementary Table 1. Search of antibiotic resistance genes was done using BLAST (Camacho et al., 2009) and ABRicate (https://github.com/tseemann/abricate (accessed on October 2021)) against ResFinder database (Zankari et al., 2012). Genome was edited and visualized by the use of SnapGene Viewer 6.0.5. Virulence factors were screened using Virulence Factors Database (VFDB) search tool (Liu et al., 2019) and Ridom SeqSphere+ software version 8.5.1 (Ridom GmbH,Münster, Germany).
2.4 Surface-associated motility
Motility assay was performed on Motility Test Medium (Condalab, Madrid, Spain) inoculated on the surface and incubated overnight at 37°C following manufacturer instructions.
2.5 Biofilm formation assays
Biofilm production was evaluated using the crystal violet staining assay described by O’Toole and Kolter as described before (O'Toole and Kolter, 1998) with slight modifications. Briefly, A. baumannii overnight cultures were adjusted to a 0.5 McFarland turbidity in 0.85% saline solution. Biofilms were developed in 24-well flat-bottom plates (Sarstedt®, Nümbrecht, Germany). First, bacterial suspensions were incubated at 37°C for 24 h. Then, biofilms were washed, air-dried and stained with 1 mL/well of 0.7% crystal violet solution (Sigma-Aldrich). Finally, stained biofilms were solubilized with 1mL/well of 33% acetic acid solution (Sigma-Aldrich).Biofilm production was determined at 600 nm using the Tecan Infinite M200 Pro Microplate Reader (Tecan Group Ltd., Männedorf, Suiza). Results were corrected for background staining by subtracting the value for crystal violet bound to uninoculated Müller Hinton Broth control wells. Isolates E. coli J53 and P. aeruginosa PAO1 were used as negative and positive controls, respectively. The experiments were performed in triplicate and repeated in three different days with similar results.
2.6 Molecular typing
Multi-Locus Sequence Typing (MLST) was performed using an open-source tool (MLST, T. Seemann, https://github.com/tseemann/mlst (accessed on October 2021) following Pasteur typing scheme. The blaOXA-51-like variant combined with the Sequence Type (ST) were used to assign the isolate to an International Clone (IC). Core genome MLST (cgMLST) based on a core genome of 2390 alleles was also carried out to study clonal relatedness by the use of Ridom SeqSphere+ software version 8.5.1 (Ridom GmbH) and a minimum spanning tree was generated.
2.7 Plasmid analysis and carbapenemase genes localization
Plasmid extractions were carried out by the use of GeneJET Plasmid Miniprep Kit following manufacturer indications (ThermoFisher Scientific, Waltham, Massachusetts, USA) and S1-Pulsed-Field Gel Electrophoresis (PFGE). Bacterial DNA embedded in agarose plugs was digested using 14 units of S1-nuclease (Takara Bio, Kusatsu, Japan) per plug followed by PFGE. Samples were run on a CHEF-DR III system (Bio-Rad, Munich, Germany) for 20 h at 6 V/cm and 14°C. CHEF DNA Size Standard Lambda Ladder (Bio-Rad) was used as molecular weight marker. Southern blot hybridizations were performed to locate blaNDM-1, blaPER-7 and blaGES-like genes with specific digoxigenin-labeled DNA probes (Roche, Mannheim, Germany). Signal detection was performed using DIG Nucleic Acid Detection Kit (Roche). Determination of the presence and classification of replicase genes was conducted using A. baumannii PCR-Based Replicon Typing as previously described (Bertini et al., 2010).
3 Results
3.1 Bacterial isolates, species identification and antimicrobial susceptibility testing
The thirty-six bacterial isolates were identified by VITEK 2® and gyrB multiplex PCR as A. baumannii (Supplementary Figure 1). Fifty-eight percent of the isolates were from male patients vs. forty-two percent from female patients. Data regarding to collection date, sex and type of sample are shown in Supplementary Table 1.
All isolates were resistant to ticarcillin, ticarcillin/clavulanic acid, piperacillin, piperacillin/tazobactam and ciprofloxacin. High resistance rates were also found for both imipenem and meropenem (94.4%). Resistance to gentamicin was observed in 80.5% of the isolates, whereas 75% of the isolates were resistant to tobramycin and trimethoprim/sulfamethoxazole. It is worth mentioning that cefiderocol resistance was found in 22.2% of the isolates and 16.7% were resistant to minocycline. No colistin-resistant isolates were found (Supplementary Table 2).
3.2 Antibiotic resistance genes identification by Whole Genome Sequencing
Sequencing results showed 8 different blaOXA-51-like variants including blaOXA-51, blaOXA-64, blaOXA-65, blaOXA-66, blaOXA-68, blaOXA-91, blaOXA-94 and blaOXA-336 (Table 1). Genes blaOXA-66 and blaOXA-65 were found in 16 and 10 isolates, respectively. Thirty-four isolates contained the blaOXA-23 gene, but the isolate Ale28 showed a 4bp deletion at position 203 of the gene leading to a premature stop codon at position 203 causing a lack of the protein expression. The presence of the blaNDM-1 carbapenemase gene was detected in ten isolates. Other β-lactamase genes such as blaGES-35 (6), blaGES-11 (3), blaPER-7 (4), blaADC-73 (15), blaADC-117 (9), blaADC-211 (1), blaADC-143 (2), blaADC-263 (2), blaADC-80 (3), blaADC-25 (1), blaADC-259 (1), blaADC-57 (1), blaADC-52 (1), blaADC-199 (1) and blaTEM-1 (15) were detected.
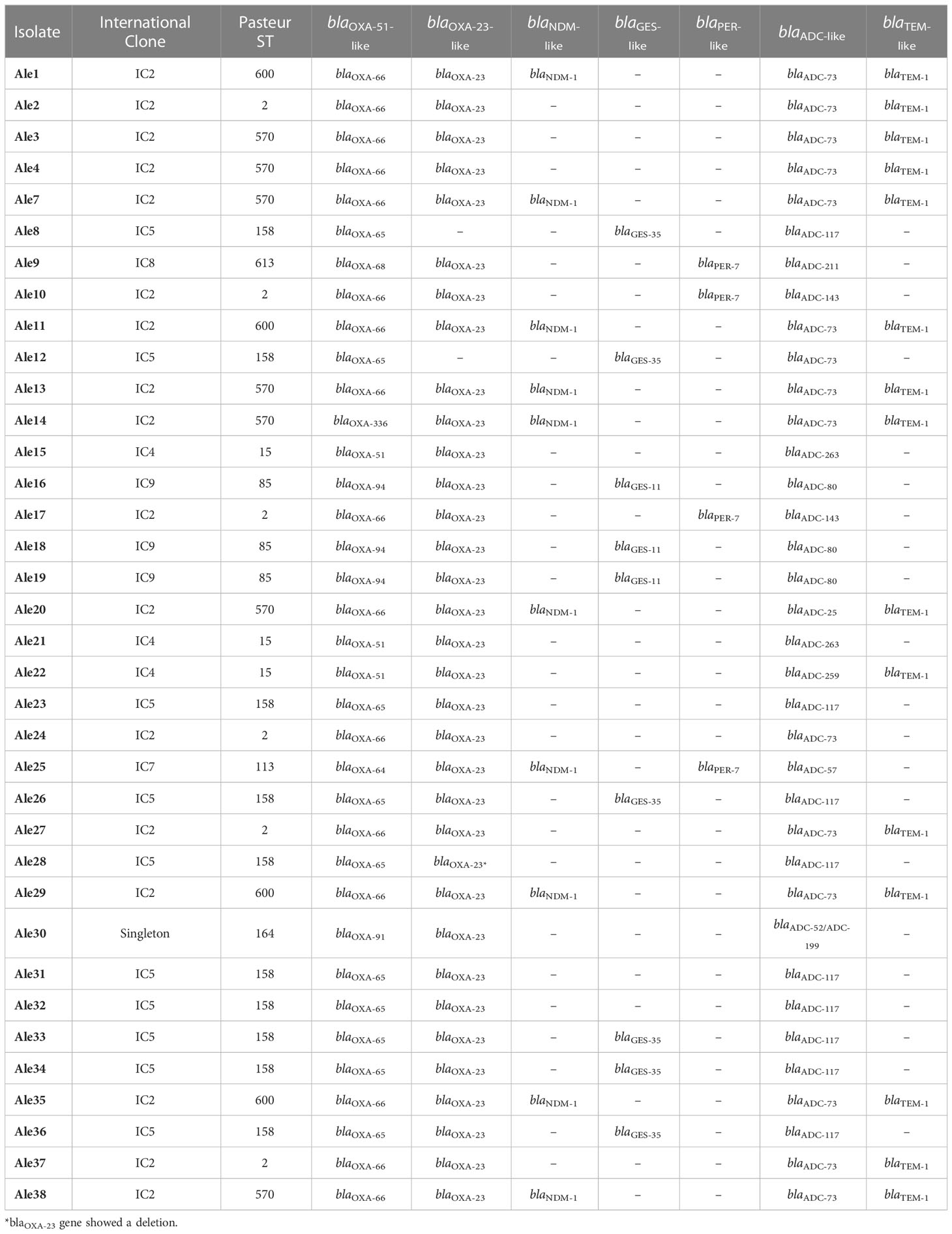
Table 1 Clonal lineages (Pasteur Sequence Type (ST) and International Clone (IC)) and β-lactamase genes identified through sequencing experiments.
Other antibiotic resistance genes were also detected including genes conferring resistance to trimethoprim (dfrA7), tetracyclines (tet(B), tet(39)), sulfonamides (sul1, sul2), aminoglycosides (armA, strA, strB, aph(3’)-Ia, aph(3’)-VI, aph(3’)-VIa, aac(6’)-Ib, ant(3’’)-II, ant(3’’)-IIa, aadA1-pm), macrolides (mph(E), msr(E)), rifamycin (arr-2) and chloramphenicol (cmlA5, catB8, catA1) (Table 2). Genes conferring resistance to aminoglycosides were especially abundant and present in the majority of the isolates. Genes coding for efflux pumps and their regulators were also detected including abeM (MATE family); abeS (SMR family); amvA, abaF and abaQ (MFS family); adeA, adeB, adeC, adeF, adeG, adeH, adeI, adeJ,adeK, adeL, adeN, adeR and adeS (RND family).
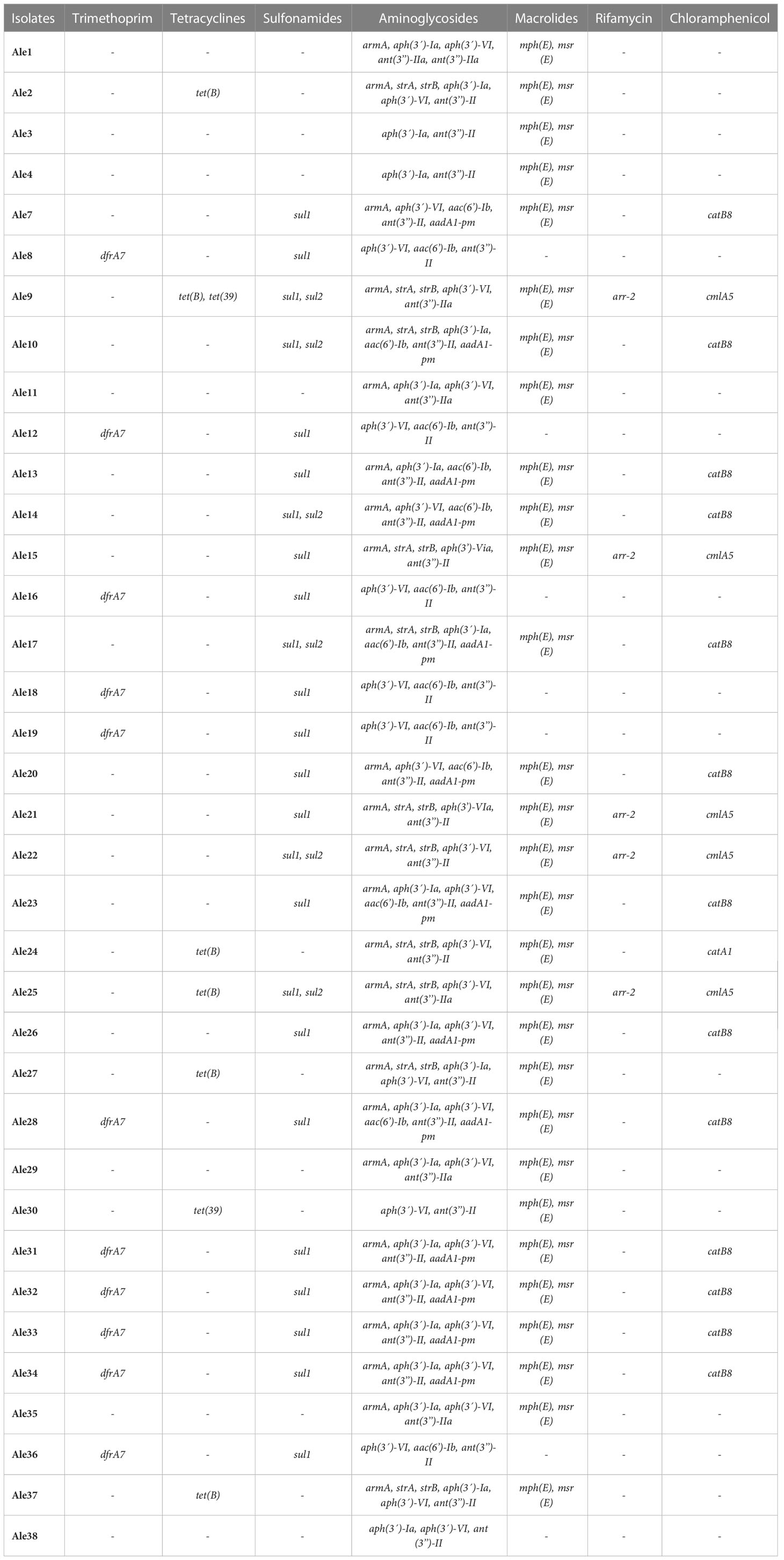
Table 2 Additional antibiotic resistance genes detected by Whole Genome Sequencing.
3.3 Genetic surroundings of β-lactamase genes
The genetic contexts of the β-lactamase coding genes are shown in Figure 1. Regarding to blaOXA-23, it was located within Tn2006 in the singleton and isolates belonging to IC2, IC4 and IC7; and within Tn2008 transposons in isolates belonging to IC9 and IC5 (Figure 1B). In all the isolates harboring blaNDM-1, the gene was within the truncated isoform of transposon Tn125 (ΔTn125) (Figure 1C). We found the blaPER-7 gene located within a complex structure of ISCR1 element and class 1 integron with part of IS26 upstream the integron in all the isolates carrying the gene (Figure 1D). The blaGES-like genes were located within a class 1 integron accompanied by other resistance genes such as aac(6´)-Ib, dfrA7 and sul1 (Figure 1E). We did not find insertion sequences upstream any of the blaOXA-51-like variants (Figure 1A).
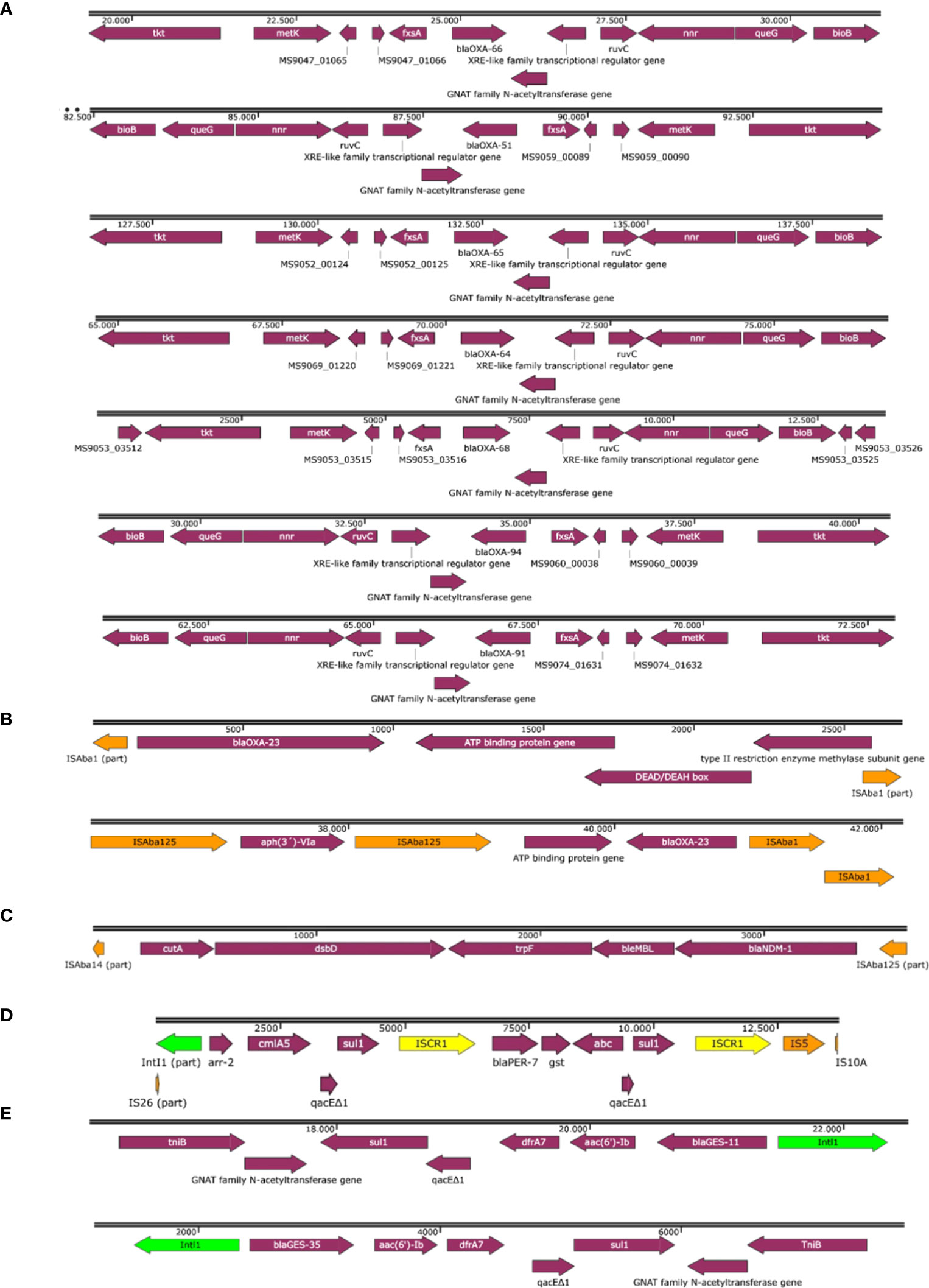
Figure 1 Genetic contexts of blaOXA-51-like variants (A), blaOXA-23 (B), blaNDM-1 (C), blaPER-7 (D) and blaGES-like variants (E).
3.4 Molecular typing and clonal relatedness
Isolates were assigned to nine Pasteur STs and clustered in six different International Clones, including IC2, IC4, IC5, IC7, IC8 and IC9 (Figure 2). One isolate assigned to ST164 was not related to any described IC.
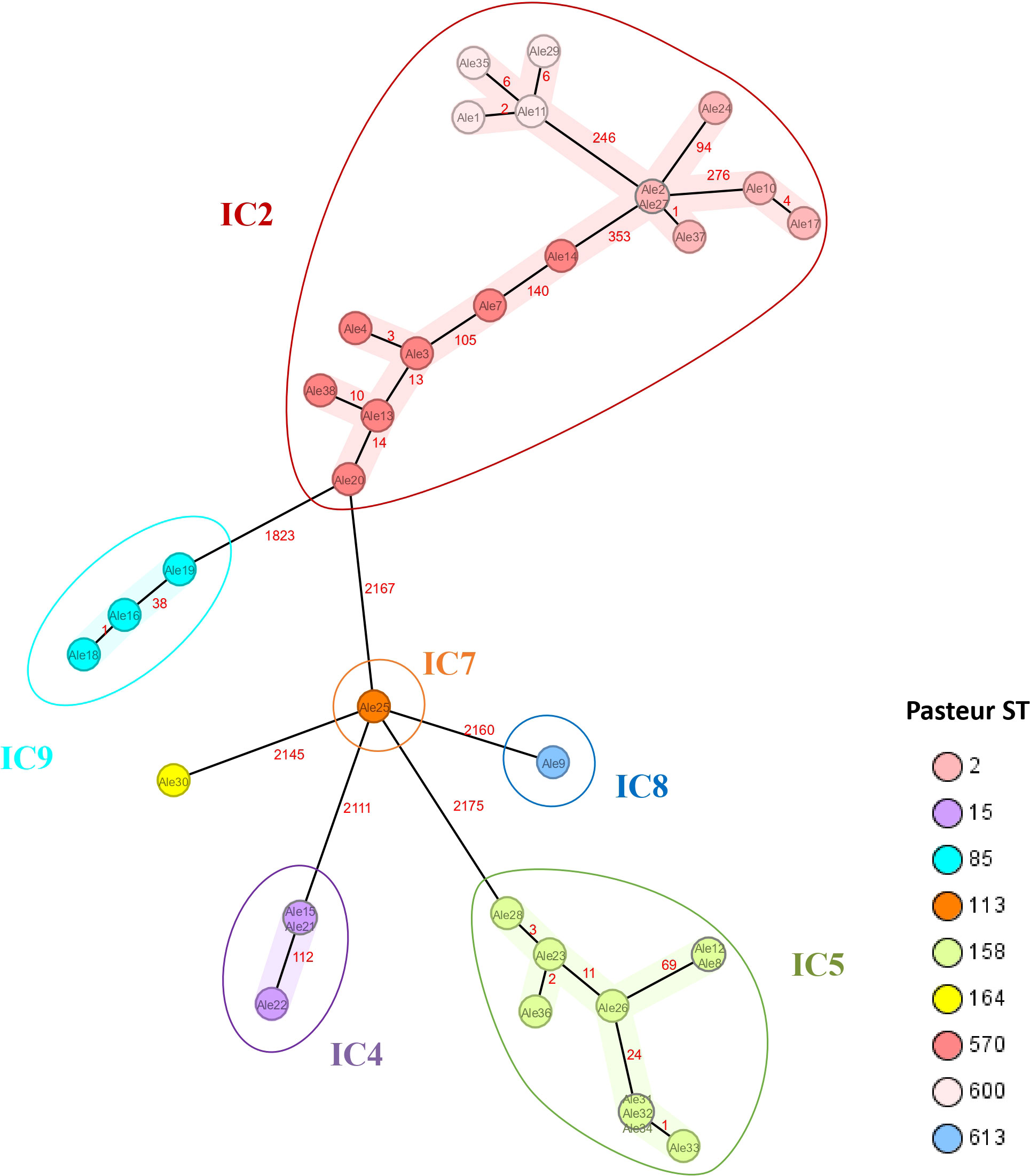
Figure 2 Minimum spanning tree of 36 A. baumannii isolates based on 2390 target alleles (core genome) generated using Ridom SeqSphere+ software. The study IDs of the isolates are shown within the nodes. Isolates are colored based on the assigned Pasteur STs and clustered by the International Clones they belong to.
3.5 Virulence factors analysis
Virulome analysis of each International Clone and the singleton showed the presence of multiple virulence factors responsible for adherence (Acinetobacter trimeric autotransporter ATA and type IV pili), biofilm production (AdeFGH efflux pump, biofilm associated protein BAP, Csu fimbriae, Poly-N-acetyl-D-glucosamine, biofilm-controlling response regulator and quorum sensing), type II and type VI secretion systems, exotoxins (phospholipases C and D), exoenzymes (coagulation targeting metallo-endopeptidase CpaA), immune modulation (capsule, lipopolysaccharide, outer membrane protein OmpA and penicillin-binding Protein G) and iron uptake (acinetobactin and HemO cluster). The gene coding for the coagulation targeting metallo-endopeptidase (cpaA) was just observed in IC8 and IC9, and ata gene coding for Acinetobacter trimeric autotransporter was present in isolates from IC2, IC4, IC5 and in the singleton. The bap gene was detected in the isolates belonging to IC2, IC4, IC5 and IC8.
3.6 Motility phenotypes
Different motility phenotypes were observed in all isolates, except to Ale36 and Ale1. A cloud-like morphology with well-defined edges was observed in the majority of isolates, although some isolates radiated uniformly from the inoculation point presenting a positive control-like morphology.
3.7 Biofilm formation assays
Varying degrees of biofilm production were observed among the isolates (Figure 3). It was observed that stronger biofilm-producers belonged to IC2, IC4 and IC7 and were isolated from BAL/miniBAL and swabs (Ale30, Ale21, Ale25, Ale38, Ale27 and Ale1). Especially remarkable was the strong biofilm production capacity of the singleton Ale30, which showed even a higher biofilm production than the positive control.
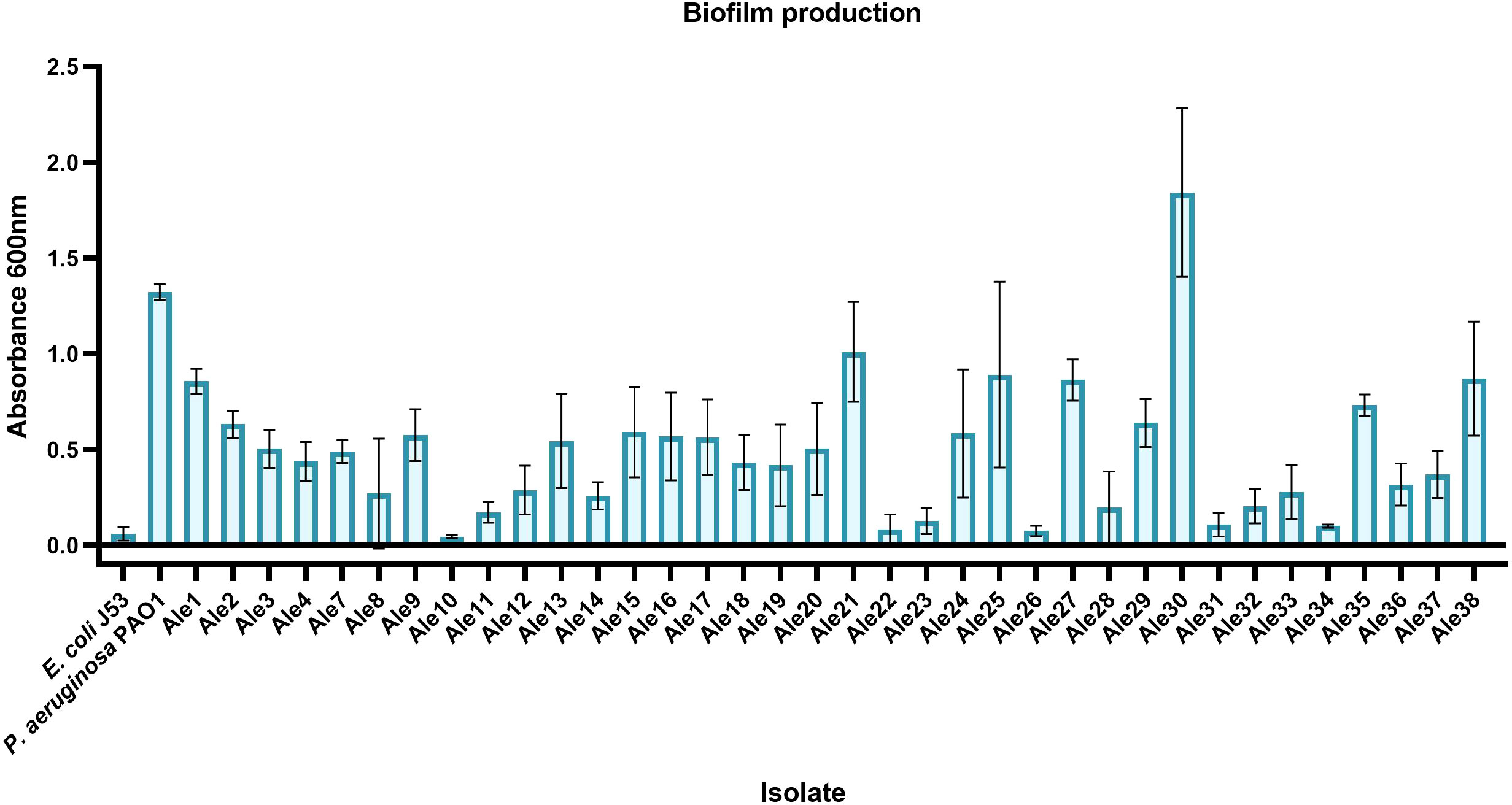
Figure 3 Biofilm production means ± standard deviations of the A. baumannii isolates. E. coli J53 and P. aeruginosa PAO1 were used as negative and positive controls, respectively.
3.8 Plasmid analysis and genetic localization of blaNDM-1, blaPER-7 and blaGES-like genes
Plasmids ranging in size from 1.7 to 70 kb were observed (Figure 4A). Hybridization experiments located the blaNDM-1 gene in the chromosome (Figure 4B). Chromosomic localization of blaPER-7 gene was also confirmed (Figure 4C). The blaGES-like genes were located in the chromosome in all the isolates, and simultaneously located in a plasmid of approximately 70kb in five isolates (Figure 4D). Replicon typing experiments showed the presence of genes coding for previously described replicases Aci1/Aci2, Aci4, Aci6, Aci8/Aci9, p2S1 and pAB49 pertaining to homology groups 2, 4, 6, 8, 12 and 16, respectively, in all the isolates except to Ale4. Twenty-one isolates showed a combination of two replicases: Aci1/Aci2 + Aci6 (6); Aci1/Aci2 + p2S1 (1); p2S1+Aci6 (10); Aci8/9 + Aci6 (1); Aci4+Aci8/Aci9 (3).
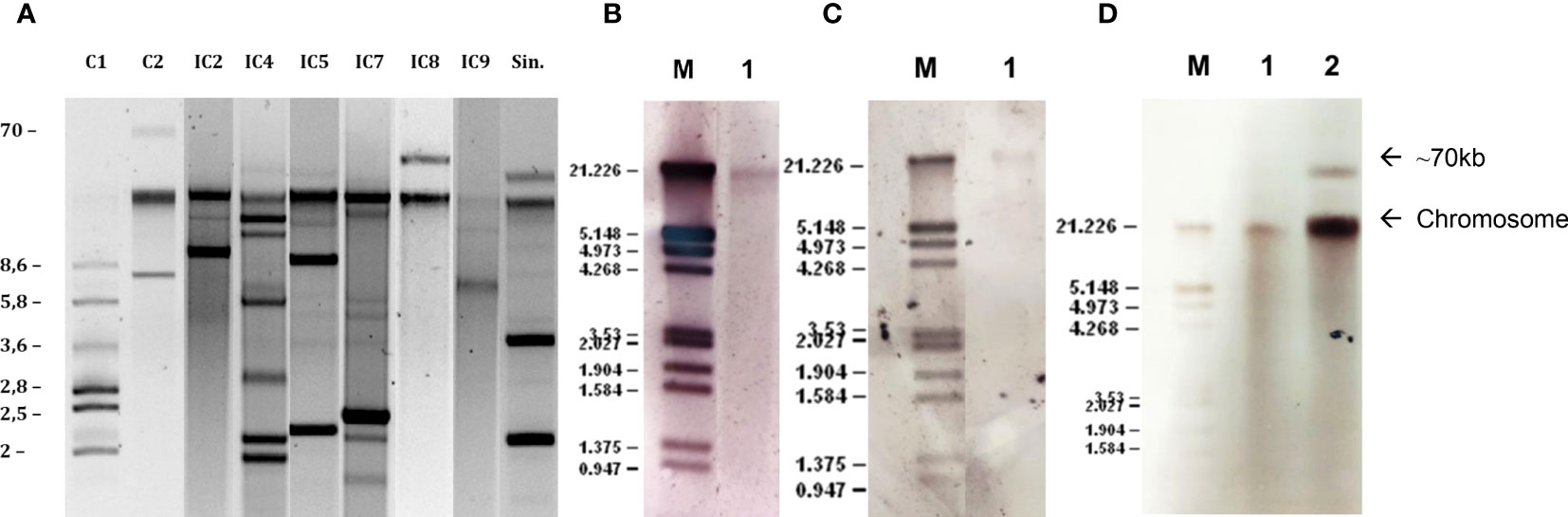
Figure 4 Plasmid profiles and hybridization experiments of the A. baumannii isolates. (A) plasmid profiles of the six identified International Clones and the singleton (Sin.); (B) hybridization signal in the chromosome with the blaNDM-1 probe; (C) hybridization signal in the chromosome with the blaPER-7 probe; (D) hybridization signal in the chromosome with the blaGES-like probe in lane 1 and hybridization signal in a 70kb plasmid in lane 2.
4 Discussion
It is said that the lack of regulation and the abuse in the use of antibiotics in Egypt are the main cause of the acceleration in the emergence of resistant isolates, and also responsible for exporting resistance to other countries (Elwakil et al., 2023). In the present study, 94.4% of carbapenem, 100% of fluoroquinolone and 86.11% of aminoglycoside resistance was observed, which is consistent with previous studies reporting a carbapenem resistance of 98% with elevated levels of resistance to quinolones and aminoglycosides in Mansoura, Egypt (Said et al., 2018). In fact, our isolates showed higher aminoglycoside resistance ratios than the resistance reported in recent studies from Egypt (up to 82% and 67%) (ELsheredy et al., 2021; Kishk et al., 2021). To our knowledge, up to date, no cefiderocol resistant isolates have been reported in Egypt apart from the eight resistant isolates analyzed in our study. Fortunately, isolates remained susceptible to colistin, although colistin resistance was reported in up to 53% of A. baumannii Egyptian isolates according to studies published in 2020 (Fam et al., 2020; Makharita et al., 2020).
WGS analysis showed blaOXA-66 and blaOXA-65 as the most frequent blaOXA-51-like variants among the isolates, which is consistent with the high prevalence of blaOXA-66 in North Africa and the Middle East (Muthuirulandi Sethuvel et al., 2019) but not with the reported prevalence of blaOXA-65 in Egypt (Elwakil et al., 2023). A higher prevalence of blaOXA-51 has been reported in Egypt during the last years, while other variants described in this study have been reported less frequently in Egypt (Elwakil et al., 2023). The genetic contexts of the blaOXA-51-like variants found in our isolates, were similar to those found in previously described isolates (Vijayakumar et al., 2022). Although ISAba is a commonly found element upstream these genes, not all the variants harbor it necessarily. The most common acquired carbapenemase gene found in these isolates was blaOXA-23, which is usually found in 90–100% of carbapenem-resistant A. baumannii isolates in Egypt (Abouelfetouh et al., 2019; Hassan et al., 2021). This gene was located within Tn2006 or Tn2008 transposons as previously described, which is crucial for the overexpression and mobilization of blaOXA-23 (Hamidian and Nigro, 2019). Regarding to blaGES-like genes, these type of Extended-Spectrum β-Lactamase genes have been increasingly reported during the last years, and some of its variants can possess carbapenemase activity (Bonnin Rémy et al., 2013). These type of β-Lactamase genes are frequently reported in the Mediterranean Area and the Middle East countries such as Turkey, Tunisia, or Kuwait (Bonnin Rémy et al., 2013; Cicek et al., 2014; Chihi et al., 2016). However, in our study, these genes were identified in 25% of the isolates, half of the frequency reported in other studies in Egypt (50%) (Ramadan et al., 2018). As previously described in isolates from Pakistan, blaGES-like genes were part of a class 1 integron co-harboring dfrA7, sul1 and aac(6’)-Ib resistance genes (Karah et al., 2020). Furthermore, blaOXA-23 and blaGES-11 genes have been described encoded in a large conjugative plasmid named pK50a (79.6kb) which is a member of the Aci6 group (Wibberg et al., 2018). This is approximately of the same size as the plasmid in which we identified the blaGES-11 gene, and it is worth mentioning that all the isolates harboring blaGES-like genes were positive for replicase Aci6 too. The blaNDM-1 gene was detected in 27.8% of isolates, in concordance with the literature where NDM-type carbapenemases are commonly reported in Egyptian isolates with a prevalence of 0–39.3% (El-Kholy et al., 2021). This gene was found within the truncated isoform of Tn125 (ΔTn125) in our isolates with the characteristic ISAba125 upstream of the blaNDM-1 gene which appears to enhance its expression (Wang et al., 2012). In all the isolates, blaNDM-1 gene was chromosome-borne, which is the most frequent localization (Fernández-Cuenca et al., 2020). The blaPER-7 gene was located within a complex structure connecting ISCR1 element and class 1 integron, which is a previously described structure closely related to multidrug resistant bacteria (Cheng et al., 2016). However, another ISCR1 element, IS5 and part of IS10A were identified downstream of the 3’-CS which differs from the described structure. NDM-type carbapenemases and PER-type β-lactamases seem to be involved in cefiderocol resistance, the recently developed last resort antibiotic (Naeimi Mazraeh et al., 2021; Poirel et al., 2021).
Analysis of the sequenced genomes showed that seventeen isolates belonged to IC2 with the characteristic blaOXA-66 gene, which is the dominant clone circulating worldwide and in Egypt (Hassan et al., 2021). Among this lineage, ST570 was the most abundant clade, an ST which has four entries in the PubMLST database, submitted from Vietnam (1) and Egypt (3) indicating that this lineage is circulating in Egyptian health settings at least, since 2017. Regarding to ST2, among 1319 isolates submitted to the database, two entries from Egypt between 2013 and 2015, and eight entries from Jordan between 2019 and 2020 were found. A recent study showed six out of seven Syrian strains pertaining to ST2 (Higgins et al., 2021), demonstrating that ST2 is commonly circulating between countries of the Mediterranean area. The less abundant clade was ST600, which has five entries in the PubMLST database isolated from Jordan (4) and Libya (1), between 2014 and 2020, suggesting a possible transmission between these two nearby countries during these years. Ten isolates belonged to IC5 (ST158) harbouring the characteristic blaOXA-65 gene. Four isolates were identified in the PubMLST database isolated from Iraq, Turkey, Russia and Egypt; consistent with a study published in 2020 in Egypt, where the majority of the isolates belonged to ST158, indicating that this clone is circulating in the Mediterranean and Middle East area (Elwakil et al., 2023). Three isolates belonged to IC4 (ST15), a ST which is predominantly found in Latin American countries, although it has also been described in countries of the Mediterranean are such as Turkey (Di Popolo et al., 2011). Three isolates were related to IC9 (ST85), which have been recently reported in Libya (Higgins et al., 2021). A search in PubMLST also showed four entries in Jordan during 2020 and one in Egypt in 2017 demonstrating the presence of this clade in Egypt and the Middle East area. Just one isolate belonging to IC7(ST113) was identified, a sequence type frequently reported in South America (Kurihara et al., 2020) although, it has also been described in Cairo in 5 isolates from 2018 to 2020 according to PubMLST. A single isolate pertaining to IC8(ST613) was detected, this is a linage with little presence in the Middle East and North Africa, with just one isolate reported in Alexandria in 2013 according to PubMLST. A singleton assigned to ST164 was identified among our isolates, this ST have been identified in Germany in 2021 (Wareth et al., 2021) and in Turkey in 2016. There are no public records of isolates belonging to this linage in Egypt up to date.
In conclusion, this study showed a high clonal diversity among CRAB isolates collected from hospitals in Alexandria, and highlights the emergence of not frequently reported lineages in Egypt. The high incidence of blaOXA-23 carbapenemase as well as blaNDM-1 is of concern as they are key in carbapenem resistance and to many other antibiotics. This work also puts the spotlight in the emergence of cefiderocol resistant isolates in Egyptian hospitals. It becomes necessary to harden infection control measures and to increase epidemiologic studies in Egypt to limit the development of new clones with highly resistant genes.
Data availability statement
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found in the article/Supplementary Material.
Author contributions
SS-U, LG and IA contributed to the design of the experiments and were responsible for the project funding. SS-U, AM-B, AO-S, MH and DA performed the experiments. SS-U, AO-S and LG analyzed and interpreted the data. SS-U and LG wrote the manuscript. SMS and ME-K were responsible for the collection, identification and determination of the resistance profile of the isolates. All authors contributed to the article and approved the submitted version.
Funding
This study was financially supported by the Ministry of Science and Innovation (MCIN/AEI/10.13039/501100011033) [Grant PID2020-116495RB-I00]; the Department of Education, Basque Government (Research Groups of the Basque University System 2021 [Group IT1578-22, GIC21/18]; and the Arab Academy for Science, Technology and Maritime Transport [Grant Number 2072].
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fcimb.2023.1208046/full#supplementary-material
References
Abouelfetouh, A., Torky, A. S., Aboulmagd, E. (2019). Phenotypic and genotypic characterization of carbapenem-resistant Acinetobacter baumannii isolates from Egypt. Antimicrobial resistance infection control 8, 185–186. doi: 10.1186/s13756-019-0611-6
Al-Hassan, L., Elbadawi, H., Osman, E., Ali, S., Elhag, K., Cantillon, D., et al. (2021). Molecular epidemiology of carbapenem-resistant Acinetobacter baumannii from Khartoum State, Sudan. Front. Microbiol. 12. doi: 10.3389/fmicb.2021.628736
Al-Hassan, L., Zafer, M. M., El-Mahallawy, H. (2019). Multiple sequence types responsible for healthcare-associated Acinetobacter baumannii dissemination in a single centre in Egypt. BMC Infect. Dis. 19 (1), 829. doi: 10.1186/s12879-019-4433-1
Bankevich, A., Nurk, S., Antipov, D., Gurevich, A. A., Dvorkin, M., Kulikov, A. S., et al. (2012). SPAdes: a new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. 19 (5), 455–477. doi: 10.1089/cmb.2012.0021
Bertini, A., Poirel, L., Mugnier, P. D., Villa, L., Nordmann, P., Carattoli, A. (2010). Characterization and PCR-based replicon typing of resistance plasmids in Acinetobacter baumannii. Antimicrobial Agents Chemotherapy 54 (10), 4168–4177. doi: 10.1128/AAC.00542-10
Bolger, A. M., Lohse, M., Usadel, B. (2014). Trimmomatic: a flexible trimmer for illumina sequence data. Bioinformatics 30 (15), 2114–2120. doi: 10.1093/bioinformatics/btu170
Bonnin Rémy, A., Rotimi Vincent, O., Al, H. M., Elise, G., Al, S. N., Patrice, N., et al. (2013). Wide dissemination of GES-type carbapenemases in Acinetobacter baumannii isolates in Kuwait. Antimicrobial Agents Chemotherapy 57 (1), 183–188. doi: 10.1128/aac.01384-12
Camacho, C., Coulouris, G., Avagyan, V., Ma, N., Papadopoulos, J., Bealer, K., et al. (2009). BLAST+: architecture and applications. BMC Bioinf. 10 (1), 421. doi: 10.1186/1471-2105-10-421
Centers for Disease Control and Prevention (2022) COVID-19: U.S. impact on antimicrobial resistance, special report 2022 (Hyattsville, MD). Available at: https://stacks.cdc.gov/view/cdc/117915 (Accessed 16th of March,2023).
Cerezales, M., Biniossek, L., Gerson, S., Xanthopoulou, K., Wille, J., Wohlfarth, E., et al. (2021). Novel multiplex PCRs for detection of the most prevalent carbapenemase genes in gram-negative bacteria within Germany. J. Med. Microbiol. 70 (3), 1310. doi: 10.1099/jmm.0.001310
Chakravarty, B. (2020). Genetic mechanisms of antibiotic resistance and virulence in Acinetobacter baumannii: background, challenges and future prospects. Mol. Biol. Rep. 47 (5), 4037–4046. doi: 10.1007/s11033-020-05389-4
Cheng, C., Sun, J., Zheng, F., Lu, W., Yang, Q., Rui, Y. (2016). New structures simultaneously harboring class 1 integron and ISCR1-linked resistance genes in multidrug-resistant gram-negative bacteria. BMC Microbiol. 16 (1), 71. doi: 10.1186/s12866-016-0683-x
Chihi, H., Bonnin, R. A., Bourouis, A., Mahrouki, S., Besbes, S., Moussa, M. B., et al. (2016). GES-11-producing Acinetobacter baumannii clinical isolates from Tunisian hospitals: long-term dissemination of GES-type carbapenemases in north Africa. J. Global Antimicrobial Resistance 5, 47–50. doi: 10.1016/j.jgar.2016.03.005
Cicek, A. C., Saral, A., Iraz, M., Ceylan, A., Duzgun, A. O., Peleg, A. Y., et al. (2014). OXA- and GES-type β-lactamases predominate in extensively drug-resistant Acinetobacter baumannii isolates from a Turkish University Hospital. Clin. Microbiol. infection 20 (5), 410–415. doi: 10.1111/1469-0691.12338
Di Popolo, A., Giannouli, M., Triassi, M., Brisse, S., Zarrilli, R. (2011). Molecular epidemiological investigation of multidrug-resistant Acinetobacter baumannii strains in four Mediterranean countries with a multilocus sequence typing scheme. Clin. Microbiol. Infection 17 (2), 197–201. doi: 10.1111/j.1469-0691.2010.03254.x
Ejaz, H., Ahmad, M., Younas, S., Junaid, K., Abosalif, K. O. A., Abdalla, A. E., et al. (2021). Molecular epidemiology of extensively-drug resistant Acinetobacter baumannii sequence type 2 Co-harboring blaNDM and blaOXA from clinical origin. Infection Drug Resistance 14, 1931–1939. doi: 10.2147/IDR.S310478
El-Kholy, A., El-Mahallawy, H. A., Elsharnouby, N., Abdel Aziz, M., Helmy, A. M., Kotb, R. (2021). Landscape of multidrug-resistant gram-negative infections in Egypt: survey and literature review. Infection Drug Resistance 14, 1905–1920. doi: 10.2147/IDR.S298920
ELsheredy, A., Yousif, Z., Elghazzawi, E., Elmenshawy, A., Ghazal, A. (2021). Prevalence of genes encoding aminoglycoside-modifying enzymes and armA among Acinetobacter baumannii clinical isolates in Alexandria, Egypt. Infect. Disord. Drug Targets 21 (8), e300821191828. doi: 10.2174/1871526521666210225113041
Elwakil, W. H., Rizk, S. S., El-Halawany, A., Rateb, M. E., Attia, A. S. (2023). Multidrug-resistant Acinetobacter baumannii infections in the United Kingdom versus Egypt: trends and potential natural products solutions. Antibiotics 12 (1), 77. doi: 10.3390/antibiotics12010077
Fam, N. S., Gamal, D., Mohamed, S. H., Wasfy, R. M., Soliman, M. S., El-Kholy, A. A., et al. (2020). Molecular characterization of Carbapenem/Colistin-resistant Acinetobacter baumannii clinical isolates from Egypt by whole-genome sequencing. Infection Drug resistance 13, 4487–4493. doi: 10.2147/IDR.S288865
Fernández-Cuenca, F., Pérez-Palacios, P., Galán-Sánchez, F., López-Cerero, L., López-Hernández, I., López Rojas, R., et al. (2020). First identification of blaNDM-1 carbapenemase in blaOXA-94-producing Acinetobacter baumannii ST85 in Spain. Enfermedades Infecciosas y Microbiología Clínica 38 (1), 11–15. doi: 10.1016/j.eimc.2019.03.008
Gurevich, A., Saveliev, V., Vyahhi, N., Tesler, G. (2013). QUAST: quality assessment tool for genome assemblies. Bioinf. (Oxford England) 29 (8), 1072–1075. doi: 10.1093/bioinformatics/btt086
Hamed, S. M., Hussein, A. F. A., Al-Agamy, M., Radwan, H. H., Zafer, M. M. (2022). Genetic configuration of genomic resistance islands in Acinetobacter baumannii clinical isolates from Egypt. Front. Microbiol. 13. doi: 10.3389/fmicb.2022.878912
Hamidian, M., Nigro, S. J. (2019). Emergence, molecular mechanisms and global spread of carbapenem-resistant Acinetobacter baumannii. Microbial Genomics 5 (10), e000306. doi: 10.1099/mgen.0.000306
Hassan, R. M., Salem, S. T., Hassan, S. I. M., Hegab, A. S., Elkholy, Y. S. (2021). Molecular characterization of carbapenem-resistant Acinetobacter baumannii clinical isolates from Egyptian patients. PloS One 16 (6), e0251508. doi: 10.1371/journal.pone.0251508
Hernández, M., Iglesias, M. R., Rodríguez-Lázaro, D., Gallardo, A., Quijada, N., Miguela-Villoldo, P., et al. (2017). Co-Occurrence of colistin-resistance genes mcr-1 and mcr-3 among multidrug-resistant Escherichia coli isolated from cattle, Spain, September 2015. Euro Surveillance: Bull. Europeen Sur Les Maladies Transmissibles = Eur. Communicable Dis. Bull. 22 (31), 77. doi: 10.2807/1560-7917.ES.2017.22.31.30586
Higgins, P. G., Hagen, R. M., Kreikemeyer, B., Warnke, P., Podbielski, A., Frickmann, H., et al. (2021). Molecular epidemiology of carbapenem-resistant Acinetobacter baumannii isolates from Northern Africa and the Middle East. Antibiotics (Basel Switzerland) 10 (3), 291. doi: 10.3390/antibiotics10030291
Higgins, P. G., Lehmann, M., Seifert, H. (2010a). Inclusion of OXA-143 primers in a multiplex polymerase chain reaction (PCR) for genes encoding prevalent OXA carbapenemases in Acinetobacter spp. Int. J. Antimicrobial Agents 35 (3), 305. doi: 10.1016/j.ijantimicag.2009.10.014
Higgins, P. G., Lehmann, M., Wisplinghoff, H., Seifert, H. (2010b). gyrB multiplex PCR to differentiate between Acinetobacter calcoaceticus and Acinetobacter genomic species 3. J. Clin. Microbiol. 48 (12), 4592–4594. doi: 10.1128/JCM.01765-10
Higgins, P. G., Pérez-Llarena, F. J., Zander, E., Fernández, A., Bou, G., Seifert, H. (2013). OXA-235, a novel class d β-lactamase involved in resistance to carbapenems in Acinetobacter baumannii. Antimicrobial Agents Chemotherapy 57 (5), 2121–2126. doi: 10.1128/AAC.02413-12
Karah, N., Khalid, F., Wai, S. N., Uhlin, B. E., Ahmad, I. (2020). Molecular epidemiology and antimicrobial resistance features of Acinetobacter baumannii clinical isolates from Pakistan. Ann. Clin. Microbiol. Antimicrobials 19 (1), 2. doi: 10.1186/s12941-019-0344-7
Kishk, R., Soliman, N., Nemr, N., Eldesouki, R., Mahrous, N., Gobouri, A., et al. (2021). Prevalence of aminoglycoside resistance and aminoglycoside modifying enzymes in Acinetobacter baumannii among intensive care unit patients, Ismailia, Egypt. Infection Drug resistance 14, 143–150. doi: 10.2147/IDR.S290584
Kurihara, M. N. L., de Sales, R. O., da Silva, K. E., Maciel, W. G., Simionatto, S. (2020). Multidrug-resistant Acinetobacter baumannii outbreaks: a global problem in healthcare settings. Rev. da Sociedade Bras. Medicina Trop. 53, e20200248. doi: 10.1590/0037-8682-0248-2020
Liu, B., Zheng, D., Jin, Q., Chen, L., Yang, J. (2019). VFDB 2019: a comparative pathogenomic platform with an interactive web interface. Nucleic Acids Res. 47 (D1), D687–D692. doi: 10.1093/nar/gky1080
Makharita, R. R., El-Kholy, I., Hetta, H. F., Abdelaziz, M. H., Hagagy, F. I., Ahmed, A. A., et al. (2020). Antibiogram and genetic characterization of carbapenem-resistant gram-negative pathogens incriminated in healthcare-associated infections. Infection Drug resistance 13, 3991–4002. doi: 10.2147/IDR.S276975
Mea, H. J., Yong, P. V. C., Wong, E. H. (2021). An overview of Acinetobacter baumannii pathogenesis: motility, adherence and biofilm formation. Microbiological Res. 247, 126722. doi: 10.1016/j.micres.2021.126722
Mohd Sazlly Lim, S., Zainal Abidin, A., Liew, S. M., Roberts, J. A., Sime, F. B. (2019). The global prevalence of multidrug-resistance among Acinetobacter baumannii causing hospital-acquired and ventilator-associated pneumonia and its associated mortality: a systematic review and meta-analysis. J. Infection 79 (6), 593–600. doi: 10.1016/j.jinf.2019.09.012
Muthuirulandi Sethuvel, D. P., Devanga Ragupathi, N. K., Bakthavatchalam, Y. D., Vijayakumar, S., Varghese, R., Shankar, C., et al. (2019). Current strategy for local- to global-level molecular epidemiological characterisation of global antimicrobial resistance surveillance system pathogens. Indian J. Med. Microbiol. 37 (2), 147–162. doi: 10.4103/ijmm.IJMM_19_396
Naeimi Mazraeh, F., Hasani, A., Sadeghi, J., Samadi Kafil, H., Soroush Barhaghi, M. H., Yeganeh Sefidan, F., et al. (2021). High frequency of blaPER-1 gene in clinical strains of Acinetobacter baumannii and its association with quorum sensing and virulence factors. Gene Rep. 24, 101232. doi: 10.1016/j.genrep.2021.101232
O'Toole, G. A., Kolter, R. (1998). Initiation of biofilm formation in Pseudomonas fluorescens WCS365 proceeds via multiple, convergent signalling pathways: a genetic analysis. Mol. Microbiol. 28 (3), 449–461. doi: 10.1046/j.1365-2958.1998.00797.x
Patel, R. V., Shah, J. S., Revathi, G., Siika, W., Shah, R. (2019). Acinetobacter infections: a retrospective study to determine inhospital mortality rate and clinical factors associated with mortality. Infection Prev. Pract. 1 (2), 100010. doi: 10.1016/j.infpip.2019.100010
Poirel, L., Sadek, M., Nordmann, P. (2021). Contribution of PER-type and NDM-type β-lactamases to cefiderocol resistance in Acinetobacter baumannii. Antimicrobial Agents Chemotherapy 65 (10), e0087721. doi: 10.1128/AAC.00877-21
Quijada, N. M., Rodríguez-Lázaro, D., Eiros, J. M., Hernández, M. (2019). TORMES: an automated pipeline for whole bacterial genome analysis. Bioinformatics 35 (21), 4207–4212. doi: 10.1093/bioinformatics/btz220
Ramadan, R. A., Gebriel, M. G., Kadry, H. M., Mosallem, A. (2018). Carbapenem-resistant Acinetobacter baumannii and Pseudomonas aeruginosa: characterization of carbapenemase genes and e-test evaluation of colistin-based combinations. Infection Drug resistance 11, 1261–1269. doi: 10.2147/IDR.S170233
Said, H. S., Benmahmod, A. B., Ibrahim, R. H. (2018). Co-Production of AmpC and extended spectrum beta-lactamases in cephalosporin-resistant Acinetobacter baumannii in Egypt. World J. Microbiol. Biotechnol. 34 (12), 189. doi: 10.1007/s11274-018-2571-z
Schmieder, R., Edwards, R. (2011). Quality control and preprocessing of metagenomic datasets. Bioinformatics 27 (6), 863–864. doi: 10.1093/bioinformatics/btr026
Seemann, T. (2014). Prokka: rapid prokaryotic genome annotation. Bioinformatics 30 (14), 2068–2069. doi: 10.1093/bioinformatics/btu153
Vijayakumar, S., Jacob, J. J., Vasudevan, K., Mathur, P., Ray, P., Neeravi, A., et al. (2022). Genomic characterization of mobile genetic elements associated with carbapenem resistance of Acinetobacter baumannii from India. Front. Microbiol. 13. doi: 10.3389/fmicb.2022.869653
Wang, Y., Wu, C., Zhang, Q., Qi, J., Liu, H., Wang, Y., et al. (2012). Identification of new Delhi metallo-β-lactamase 1 in Acinetobacter lwoffii of food animal origin. PloS One 7 (5), e37152. doi: 10.1371/journal.pone.0037152
Wareth, G., Linde, J., Hammer, P., Splettstoesser, W. D., Pletz, M. W., Neubauer, H., et al. (2021). Molecular characterization of German Acinetobacter baumannii isolates and multilocus sequence typing (MLST) analysis based on WGS reveals novel STs. Pathog. (Basel Switzerland) 10 (6), 690. doi: 10.3390/pathogens10060690
Wibberg, D., Salto, I. P., Eikmeyer, F. G., Maus, I., Winkler, A., Nordmann, P., et al. (2018). Complete genome sequencing of Acinetobacter baumannii strain K50 discloses the Large conjugative plasmid pK50a encoding carbapenemase OXA-23 and extended-spectrum β-lactamase GES-11. Antimicrob. Agents Chemother. 62 (5), e00212–e00218. doi: 10.1128/AAC.00212-18
Wood, D. E., Salzberg, S. L. (2014). Kraken: ultrafast metagenomic sequence classification using exact alignments. Genome Biol. 15 (3), R46. doi: 10.1186/gb-2014-15-3-r46
Woodford, N., Ellington, M. J., Coelho, J. M., Turton, J. F., Ward, M. E., Brown, S., et al. (2006). Multiplex PCR for genes encoding prevalent OXA carbapenemases in Acinetobacter spp. Int. J. Antimicrobial Agents 27 (4), 351–353. doi: 10.1016/j.ijantimicag.2006.01.004
Xanthopoulou, K., Urrutikoetxea-Gutiérrez, M., Vidal-Garcia, M., Diaz de Tuesta del Arco, J.-L., Sánchez-Urtaza, S., Wille, J., et al. (2020). First report of new Delhi metallo-β-Lactamase-6 (NDM-6) in a clinical Acinetobacter baumannii isolate from Northern Spain. Front. Microbiol. 11 (589253). doi: 10.3389/fmicb.2020.589253
Keywords: Acinetobacter baumannii, carbapenemase genes, antibiotic resistance, molecular epidemiology, whole genome sequencing
Citation: Sánchez-Urtaza S, Ocampo-Sosa A, Molins-Bengoetxea A, El-Kholy MA, Hernandez M, Abad D, Shawky SM, Alkorta I and Gallego L (2023) Molecular characterization of multidrug resistant Acinetobacter baumannii clinical isolates from Alexandria, Egypt. Front. Cell. Infect. Microbiol. 13:1208046. doi: 10.3389/fcimb.2023.1208046
Received: 18 April 2023; Accepted: 03 July 2023;
Published: 20 July 2023.
Edited by:
Henan Li, Peking University People’s Hospital, ChinaReviewed by:
Bogdan M. Benin, Northeast Ohio Medical University, United StatesAunchalee Thanwisai, Naresuan University, Thailand
Vittoria Mattioni Marchetti, Charles University, Czechia
Carlos Henrique Camargo, Adolfo Lutz Institute, Brazil
Copyright © 2023 Sánchez-Urtaza, Ocampo-Sosa, Molins-Bengoetxea, El-Kholy, Hernandez, Abad, Shawky, Alkorta and Gallego. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Lucia Gallego, bHVjaWEuZ2FsbGVnb0BlaHUuZXM=