 Sandra Sánchez-Urtaza1,2†
Sandra Sánchez-Urtaza1,2† Laura Alfonso-Alarcón1†
Laura Alfonso-Alarcón1† Rocío Arazo del Pino2,3
Rocío Arazo del Pino2,3 Tessa Burgwinkel2,3
Tessa Burgwinkel2,3 Alain Ocampo-Sosa4
Alain Ocampo-Sosa4 Ruth Gonzalez5
Ruth Gonzalez5 Kyriaki Xanthopoulou2,3
Kyriaki Xanthopoulou2,3 Paul G. Higgins2,3*
Paul G. Higgins2,3* Itziar Alkorta6
Itziar Alkorta6 Lucia Gallego1*
Lucia Gallego1*- 1Laboratory of Antibiotics and Molecular Bacteriology, Department of Immunology, Microbiology and Parasitology, Faculty of Medicine and Nursing, University of the Basque Country, Leioa, Spain
- 2Institute for Medical Microbiology, Immunology and Hygiene, Faculty of Medicine and University Hospital Cologne, University of Cologne, Cologne, Germany
- 3German Center for Infection Research (DZIF), Partner Site Bonn-Cologne, Cologne, Germany
- 4Microbiology Service, University Hospital Marqués de Valdecilla, Health Research Institute (Instituto de Investigación Valdecilla), Santander, Spain and CIBERINFEC, Instituto de Salud Carlos III, Madrid, Spain
- 5National Hospital of Itaugua, Itaugua, Paraguay
- 6Department of Biochemistry and Molecular Biology, Faculty of Science and Technology, University of the Basque Country, Leioa, Spain
Acinetobacter baumannii is a clinically important pathogen capable of causing serious nosocomial infections and acquiring resistance to antimicrobials, particularly carbapenems, making treatment difficult and prolonging hospital stays. In Latin America, high carbapenem-resistance rates have been described among A. baumannii isolates, however, Paraguay is one of the countries with limited data in this regard. Therefore, we aimed to investigate resistance rates of A. baumannii isolates from the National Hospital of Itaugua (NHI), Paraguay, from their database of 2022, and from December 2023 to February 2024, and to study in detail a representative group of multidrug-resistant clinical isolates. For this purpose, data were analyzed considering diagnostic, sample type and antimicrobial susceptibility. Eight A. baumannii isolates recovered from patients in six separate ICUs in 2024 were then selected and subjected to susceptibility testing using VITEK® and to short- and long-read sequencing, and clonality, resistome, virulome and plasmidome of the isolates were investigated. IC2 (ST2 Pasteur, ST1816/195 Oxford and ST872 Oxford) was the predominant clone among the Paraguayan isolates, and a single isolate belonging to clone IC5 (ST79 Pasteur and ST1283 Oxford) was also identified. The carbapenemase gene blaOXA-23 was located in transposons Tn2006 and Tn2008. Additionally, other antibiotic resistance genes conferring resistance to aminoglycosides, macrolides, sulfonamides, chloramphenicol, tetracyclines and trimethoprim were identified, and were found embedded in genetic environments containing mobile genetic elements. Multiple virulence genes were also detected, mainly promoting biofilm formation and immune system modulation. Plasmid analysis showed the presence of plasmids ranging in size from 2.27 to 10.74 Kb. This work describes the dissemination of the emerging clone IC2 in Paraguay and offers a detailed analysis of the resistome, virulome and plasmidome of carbapenem-resistant A. baumannii strains. The results obtained highlight the importance of correctly characterizing these multidrug-resistant pathogens to develop infection prevention and control strategies at hospital level.
1 Introduction
Acinetobacter baumannii is a highly clinically relevant pathogen that can lead to fatal nosocomial infections affecting mainly compromised patients or patients with underlying diseases. Infections caused by A. baumannii include pneumonia due to mechanical ventilation, catheter associated infections, bacteremia, urinary tract and bone infections, skin and wound infections. A. baumannii has a propensity to cause outbreaks at Intensive Care Units (ICU), and is strongly linked to the multidrug resistance phenotype, particularly carbapenem-resistance. Its clinical success is mainly due to its high virulence and the ability to survive in adverse conditions such as inanimate surfaces, combined with its desiccation and biocide tolerance, but also its capacity to acquire antibiotic resistance genes that are often coded by mobile genetic elements facilitating their spread (Jeon et al., 2023; Lee et al., 2017). Especially worrying is resistance to last resort antibiotics such as carbapenems, that usually makes infections difficult to treat, prolonging the hospital stay and making it necessary the use of more harmful and expensive antibiotics, leading sometimes to the death of the patients (Castanheira et al., 2023; Hamidian and Nigro, 2019; Kyriakidis et al., 2021; Ramirez et al., 2020; Vijayakumar et al., 2019).
During the COVID-19 pandemic, carbapenem resistant A. baumannii (CRAB) infections increased due to the massive admission of patients into hospitals, globalization, migration and the inappropriate use of antimicrobials (Castanheira et al., 2023; Montrucchio et al., 2022; Müller et al., 2023). In 2017 the World Health Organization published a list of critical pathogens for which new antibiotics were needed, with CRAB as the number one critical priority pathogen, and is still heading the list in the updated list published in 2024 (Tacconelli et al., 2018; World Health Organization, 2024). These traits contribute to its dissemination and contribute to its propensity for nosocomial outbreaks, especially where there are gaps in the infection-prevention and control regimes making it a critical pathogen threatening global health (Castanheira et al., 2023; Hamidian and Nigro, 2019; Montrucchio et al., 2022; Rodríguez et al., 2018).
According to Rodríguez et al. analyzing data obtained from 2000 to 2013, only 60% of the Latin-American countries had reported CRAB, mainly in Argentina and Brazil (Rodríguez et al., 2018). Resistance to carbapenems (meropenem and imipenem) in Latin-America is especially alarming considering it is ranging from 80 to 90%. The data obtained after the COVID-19 pandemic raised concern about the health system since that trend was increased in several countries such as Peru and Argentina, but also in Paraguay, Guatemala, Bolivia, El Salvador, Nicaragua and Ecuador (Manobanda Nata and Jaramillo Ruales, 2023). Most of the carbapenemase-encoding genes circulating in Latin-America belong to the oxacillinase families blaOXA-23, blaOXA-58, blaOXA-24/40 (blaOXA-72), blaOXA-143 (blaOXA-253) and blaOXA-214 (blaOXA-215, blaOXA-264, blaOXA-265, blaOXA-575) (Manobanda Nata and Jaramillo Ruales, 2023; Naas et al., 2017). However, less frequently, metallo-beta-lactamase genes such as blaNDM, blaVIM and blaIMP are also reported (Manobanda Nata and Jaramillo Ruales, 2023). Although Paraguay is one of the Latin-American countries with limited accessible data regarding antibiotic resistance and epidemiology of the circulating A. baumannii isolates, the most recent reports showed that 70% of the isolates are carbapenem-resistant (Melgarejo-Touchet et al., 2021) usually carrying blaOXA-23-like and blaNDM-like genes, while coexistence of blaNDM-like and blaOXA-58-like genes was reported in a 2% of isolates (Melgarejo-Touchet et al., 2021). International clone (IC) 5 and IC7 are the most common lineages found in the country (Rodríguez et al., 2016).
The aim of this work was the analysis of the resistance to antibiotics during the years 2022, 2023 and 2024, and the molecular characterization by whole genome sequencing of a representative series of carbapenem-resistant A. baumannii isolates recovered from the National Hospital of Itaugua (NHI), Paraguay, in 2024.
2 Materials and methods
2.1 Hospital database analysis and selection of bacterial isolates
The NHI is a specialized hospital affiliated to the Ministry of Public Health and Social Wellness, with more than 750 beds, from which 115 belong to the Intensive Care Units (ICUs). It is a reference center giving service to the Paraguayan population with the aim of providing preventive and curative health care. The data from 41 ICU isolates identified as A. baumannii complex and recovered between December 2023 and February 2024 were analyzed. For this study, within this last group, the first isolate recovered from each ICU (eight isolates in total) were selected for further analysis (Table 1). Furthermore, the evolution of antimicrobial resistance rates was analyzed based on the last official report of the NHI from 2022, which included information about diagnostics, type of clinical sample, species and antimicrobial susceptibility testing.
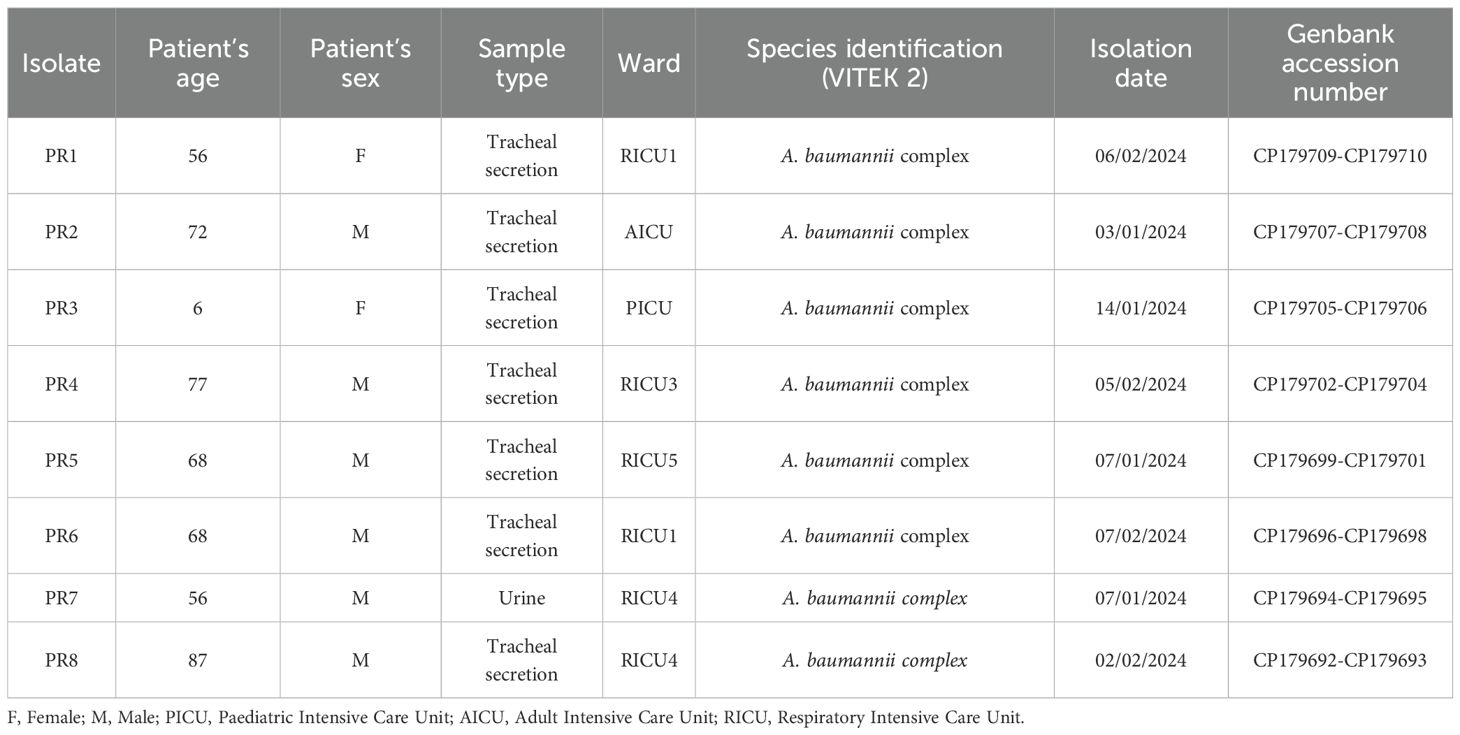
Table 1. Overview of the selected A. baumannii complex isolates recovered from National Hospital of Itaugua of Paraguay.
2.2 Species identification and susceptibility testing
Species identification and antibiotic susceptibility testing were initially performed at the hospital using the automated method VITEK 2® Compact (BioMérieux, Marcy l´Étoile, Francia) to determine the Minimum Inhibitory Concentrations (MIC) following the guidance of the Clinical and Laboratory Standards Institute (CLSI) (Lewis and Clinical and Laboratory Standards Institute, 2024). The species were later confirmed by gyrB multiplex PCR and PCR of the intrinsic blaOXA-51 gene, as previously described (Higgins et al., 2010b; Turton et al., 2006) and by whole genome sequencing. Isolates were determined to be MDR (Multidrug-Resistant) if they were at least resistant to one agent in three or more antimicrobial categories as previously described (Magiorakos et al., 2012).
2.3 Preliminary screening of carbapenemase genes
To investigate the presence of carbapenem-resistance genes, multiplex PCR experiments targeting blaOXA-51, blaOXA-23, blaOXA-24/40, blaOXA-58, blaOXA-143 and blaOXA-235 genes were first performed as previously described (Woodford et al., 2006; Higgins et al., 2010a, 2013). Additionally, two multiplex PCR were also carried out to detect the presence of other carbapenemases such as blaVIM, blaKPC, blaNDM, blaOXA-48, blaIMI, blaGES, blaGIM and blaIMP (Cerezales et al., 2021). The presence of these genes was further confirmed by whole genome sequencing.
2.4 Whole genome sequencing
For short-read sequencing, total DNA was purified with the DNeasy UltraClean Microbial Kit (Qiagen, Hilden, Germany) and libraries were prepared using the NEBNext® Ultra™ II FS DNA Library Prep Kit (New England Biolabs, Ipswich, Massachusetts, USA) for a 250 bp paired-end sequencing on an Illumina® MiSeq (Illumina, Inc., San Diego, California, USA). For long-read sequencing, genomic DNA was extracted using the NEB® Monarch genomic DNA purification kit and DNA concentrations were measured with the Qubit fluorimeter (Thermo Fisher Scientific) using the Qubit™ 1X dsDNA Broad-Range Assay-Kit (Waltham, Massachusetts, USA). Libraries were prepared with the Rapid Barcoding Kit 24 V14 (SQK-RBK114.24) and sequenced on a MinION Mk1b device using a R10.4.1 (FLO-MIN114) flow-cell (Oxford Nanopore Technologies, Cambridge, UK). Hybrid genome assemblies using short- and long-reads were performed with Unicycler v0.5.1 (Wick et al., 2017) and genome annotation was assessed with Bakta web v1.9.1; DB:5.0.0 (Schwengers et al., 2021) and manually curated. The closed genome assemblies of the isolates generated for this study were deposited and can be found in GenBank under the BioProject ID PRJNA1197744 and the accession numbers of each isolate are detailed in Table 1.
2.4.1 Analysis of the molecular epidemiology
Clonal relatedness was investigated by Multi Locus Sequence Typing (MLST) following Pasteur and Oxford schemes using the PubMLST website (Jolley et al., 2018), and analyzing the core genome MLST (cgMLST) using the Ridom SeqSphere+ software version 10.0.5 (Ridom GmbH, Münster, Germany) based on a core genome of 2390 alleles (Jünemann et al., 2013). A minimum spanning tree was also generated using Ridom SeqSphere+ to visualize the results.
2.4.2 Investigation of antibiotic resistance and virulence genes
Antimicrobial resistance genes were detected using ResFinder v4.5.0 (Zankari et al., 2012) and the BetaLactamase DataBase (BLDB) (Naas et al., 2017), virulence factors were screened using Virulence Factor Database (VFDB) search tool (Liu et al., 2018) and Kaptive v2.0.1 (Cahill et al., 2022) while BLAST search against NCBI database (Camacho et al., 2009) was used for confirmation. Assembled genomes were analyzed in detail, edited and visualized using SnapGene Viewer 7.2.1 (www.snapgene.com) to look for insertion sequences and transposons associated with the antibiotic resistance genes using ISFinder version 2024-06-13 (http://www-is.biotoul.fr) and TnCentral: a Prokaryotic Transposable Element Database (Ross et al., 2021), respectively.
2.4.3 Plasmid analysis
To investigate the plasmid content, plasmid extractions were first performed using the GeneJET Plasmid Miniprep Kit (ThermoFisher Scientific, Waltham, Massachusetts, USA) following the manufacturer’s instructions, and then loaded onto a 0.7% agarose gel. For further analysis, determination on the presence and classification of replicase genes was assessed by performing A. baumannii PCR-Based Replicon Typing, as previously described (Bertini et al., 2010), and doing a BLAST of the genomes against the Plasmid Typing Database developed by Lam et al (Lam et al., 2023).
2.5 Phenotypic analysis of virulence factors
2.5.1 Biofilm formation experiments
Biofilm production was tested in biological triplicates, on three independent days following the protocol described by O' Toole and Kolter with slight modifications (O’Toole and Kolter, 1998). Briefly, overnight cultures were adjusted to 0.5 McFarland, and 100 μL of each culture were placed on a 24-well flat-bottom plate (Sarstedt®, Nümbrecht, Germany) with 900 μL of Mueller Hinton broth, and incubated overnight at 37°C. Culture media were then discarded, washed twice with 1 mL of distilled water and air-dried for 20 min. The staining was done by adding 1 mL of 0.7% crystal violet solution and incubating for 12 min at room temperature, and then, it was washed and air-dried as previously described. Stained biofilms were solubilized using 1 mL of 33% acetic acid solution and incubated with agitation for 5 min. Finally, 100 μL of each well were placed into a 96 well flat bottom plates to measure the absorbances at 600 nm on a Tecan Infinite M200 Pro Microplate Reader (Tecan Group Ltd., Männedorf, Switzerland). The absorbance value for crystal violet bound to uninoculated Müller Hinton Broth control wells was subtracted in order to correct the results for background staining. The results were expressed as the mean ± standard deviation of the replicates. Escherichia coli J53 and Pseudomonas aeruginosa PAO1 were used as negative and positive controls, respectively. The statistical analysis was performed using GraphPad Prism v8.0.2. Differences between the isolates and the control strains were analyzed using one-way ANOVA, followed by a Tukey post hoc test for multiple comparisons and statistically significant differences were defined as those with p < 0.05.
2.5.2 Surface-motility assays
To investigate the motility of the isolates, overnight cultures were adjusted to 0.5 McFarland and 1 μL was placed in the center of the Motility Test Medium (Condalab, Madrid, Spain) plates. After 5 days of incubation at 37°C, plates were observed and compared to the positive and negative controls, A. baumannii ATCC 19606 and A. baumannii ATCC 17978, respectively.
3 Results
3.1 Hospital database analysis, characteristics and susceptibility testing of the isolates recovered from the NHI
In 2022, 11% of the microorganisms isolated in the hospital were A. baumannii, precisely, 84 isolates were recovered per month making a total of 1002 isolates, of which, 40% were isolated from female patients. Half of the isolates were recovered from respiratory samples and 18% from blood cultures (Supplementary Figure S1A). Analyzing the antimicrobial susceptibility rates of the year 2022, there was low susceptibility to imipenem and meropenem (15%), ceftazidime, ampicillin/sulbactam and ciprofloxacin (16%), gentamicin (37%) and amikacin (40%) (Supplementary Figure S1B). It is worth mentioning that those isolates resistant to carbapenems were also resistant to fluoroquinolones. In contrast, from December 2023 to February 2024, 88% of the A. baumannii isolates were recovered from respiratory samples and 3% from blood cultures (Supplementary Figure S2A). Antibiotic susceptibility analysis showed that 10% of isolates were susceptible to imipenem, meropenem, ceftazidime, ciprofloxacin and piperacillin/tazobactam; 24% were susceptible to amikacin and 100% were susceptible to colistin (Supplementary Figure S2B).
3.2 Species identification and preliminary screening of carbapenemase genes
The eight isolates selected for further analysis were identified as A. baumannii by amplification of the gyrB and blaOXA-51 genes. Investigation of carbapenemase genes showed that all the isolates were positive for blaOXA-23. No other carbapenemase-encoding genes were detected.
3.3 Analysis of the molecular epidemiology
Seven isolates belonged to International Clone 2 (IC2) and were assigned to ST2 according to the Pasteur scheme. Based on the Oxford scheme, isolates PR1, PR2, PR3 and PR8 were assigned to ST1816/ST195, while PR4, PR5, and PR6 were assigned to ST872. Notably, the assignment of two Oxford STs to some isolates was due to a duplication of the gdhB gene. Based on the cgMLST analysis, these seven isolates were grouped into two clusters (cluster A and cluster B), separated by 106 alleles (corresponding to ST872 and ST1816/ST195, respectively) (Figure 1). Within each cluster, isolates differed by five or fewer alleles. In cluster A, PR5 and PR6 differed from PR4 at a single locus, ACICU_RS04400, encoding an acetylglutamate kinase. In cluster B, isolates PR1 and PR3 differed from PR2 and PR8 at four loci: ACICU_RS01055 (lytic transglycosylase domain-containing protein), ACICU_RS07275 (hybrid sensor histidine kinase/response regulator), ACICU_RS08455 (RND transporter), and ACICU_RS11490 (alpha/beta hydrolase). Additionally, PR2 differed from PR1, PR3, and PR8 at ACICU_RS13890 (hypothetical protein), while PR8 differed from the other cluster B isolates at four loci: ACICU_RS02340 (lysine transporter LysE), ACICU_RS03900 (hypothetical protein), ACICU_RS13905 (aspartate aminotransferase), and ACICU_RS14920 (metal-binding protein) (Supplementary Table S1). A singleton isolate, PR7, belonged to IC5 and was assigned to ST79 (Pasteur) and ST1283 (Oxford).
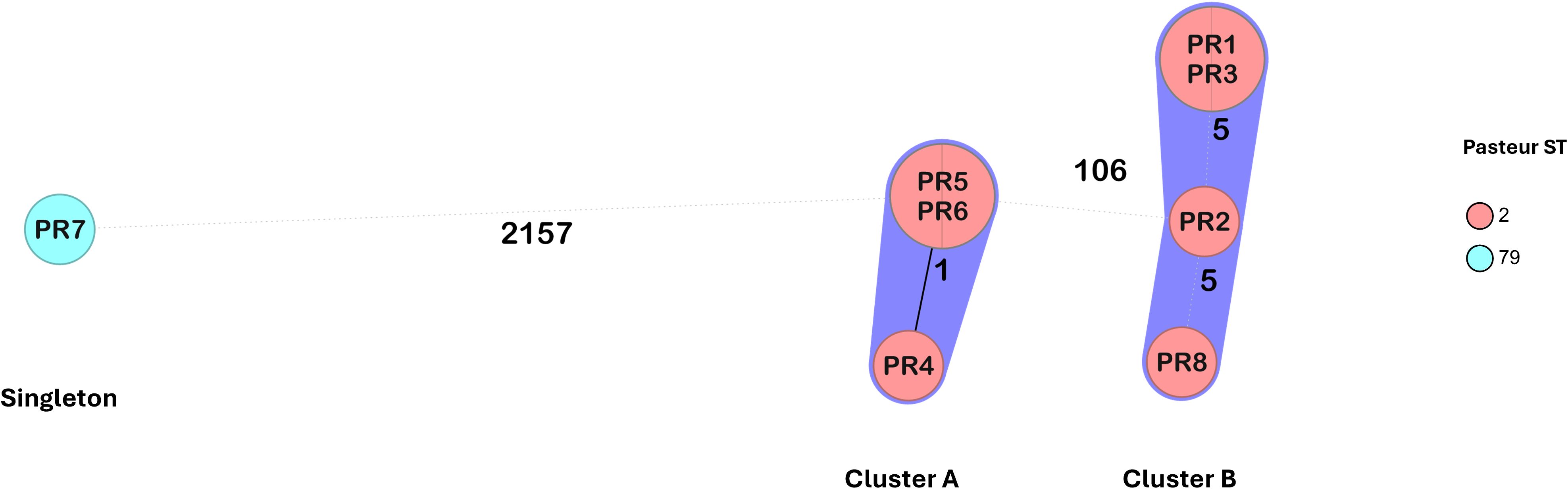
Figure 1. Minimum spanning tree of the A. baumannii isolates from the National Hospital of Itaugua (Paraguay) based on a core genome of 2390 alleles generated using Ridom SeqSphere+. The study IDs of the isolates are shown within the nodes. Isolates are colored by the assigned Pasteur STs.
3.4 Investigation of antibiotic resistance and virulence genes
By whole genome sequencing the β-lactamase variants were confirmed (Table 2). Two variants of the blaOXA-51-like gene were identified: blaOXA-66 (IC2) in seven isolates and blaOXA-65 (IC5) in isolate PR7. Two copies of the blaOXA-23 gene were also found in isolates PR1-PR6 and PR8. Regarding the chromosomal cephalosporinase genes, blaADC-73 was identified in all the IC2 isolates, while blaADC-5 was identified in isolate PR7, and the broad spectrum β-lactamase gene blaTEM-1 was also detected in the latter isolate.
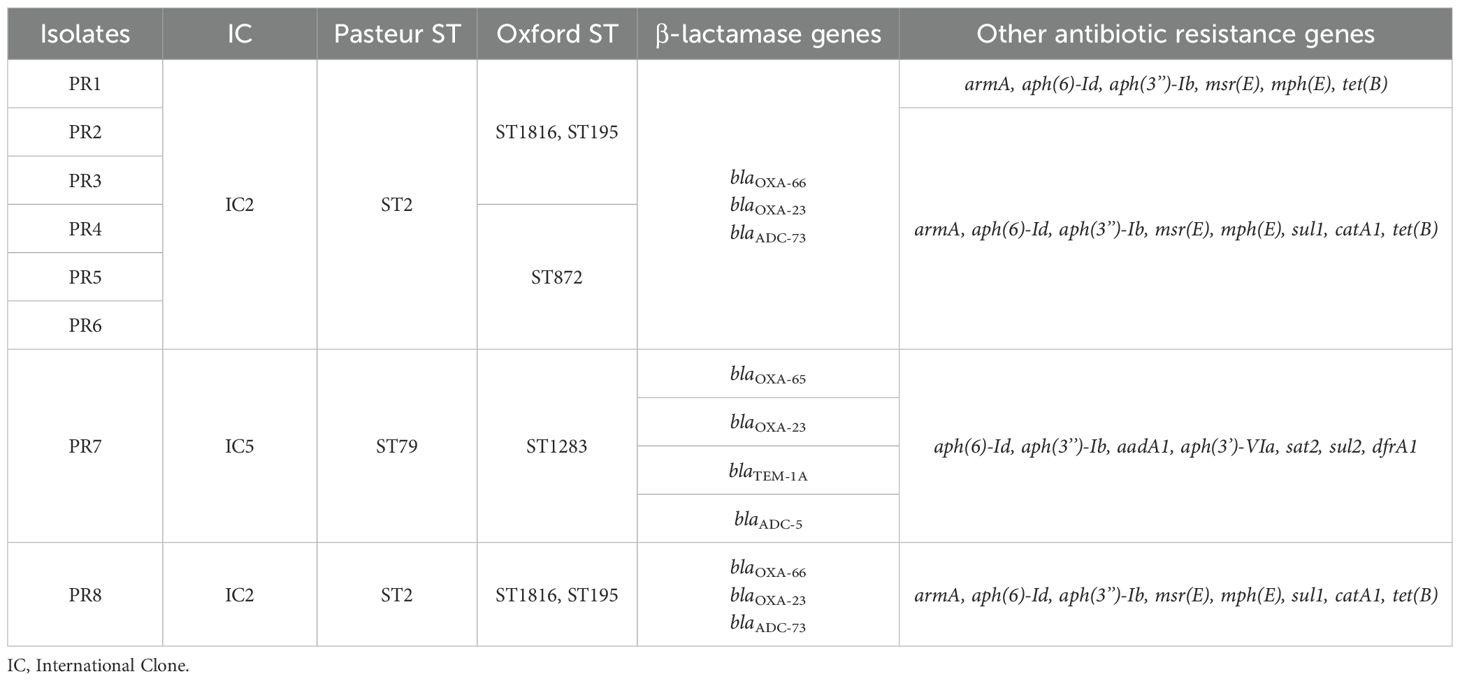
Table 2. Overview of clonal lineages, sequence types and resistome of the A. baumannii isolates from the National Hospital of Itaugua.
Other antibiotic resistance genes conferring resistance to aminoglycosides (armA, aph(6)-Id, aph(3’’)-Ib, aph(3’)-Via, aadA1 and sat2), macrolides (msr(E) and mph(E)), sulfonamides (sul1 and sul2), chloramphenicol (catA1), tetracyclines (tet(B)) and trimethoprim (dfrA1) were also identified (Table 2).
Regarding the virulome, all the isolates harbored a wide variety of genes involved in biofilm production, immune system modulation, secretion systems, acinetobactins and exotoxins (Supplementary Figure S3). Furthermore, capsule typing of the isolates revealed two different OC locus OCL1 in those isolates belonging to IC2, and OCL10 in the isolate belonging to IC5. Regarding the K locus, isolates within the IC2 cluster were subdivided into KL3 and KL81, and the IC5 isolate was categorized as KL9 (Supplementary Figure S3).
3.4.1 Genetic environments of the β-lactamase genes
The analysis and annotation of the genetic environment of the β-lactamase genes revealed that blaOXA-23 gene was located within different structures. In those isolates belonging to IC2 (PR1–PR6 and PR8), the gene was located within the transposon Tn2006, present in two identical copies per genome (Figure 2A). In the isolate belonging to IC5 it was harbored by the transposon Tn2008 (Figure 2B). These transposons were structurally conserved and matched the canonical configurations in all the isolates. No insertion sequences were detected immediately upstream of the blaOXA-51 genes (Figure 2C, D). Regarding the intrinsic cephalosporinase genes, ISAba1 was upstream of blaADC-73 gene (Figure 2E), while no mobile genetic element was found upstream of blaADC-5 (Figure 2F). The blaTEM-1 gene was encoded in transposon Tn3 (Figure 2G).
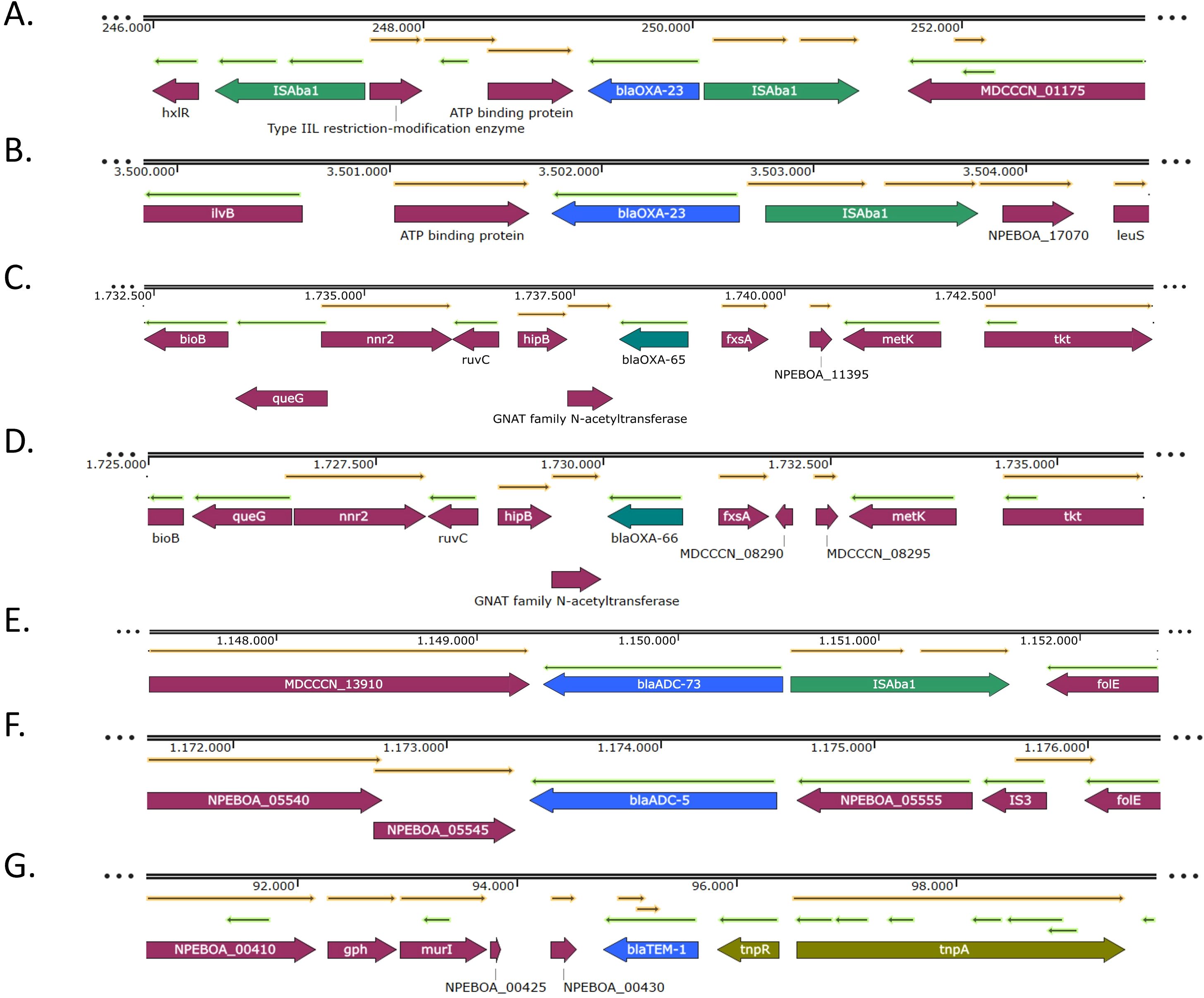
Figure 2. Genetic environments of the β-lactamase genes blaOXA-23 (A, B), blaOXA-51-like (C, D), blaADC-like (E, F) and blaTEM-1 (G) genes. Arrows represent open reading frames, with colors indicating gene function: resistance genes in blue, mobile genetic elements in green, and other genes in purple. Arrow direction indicates transcriptional orientation.
3.4.2 Genetic environments of non-β-lactam antibiotic resistance genes
The rest of the genes involved in resistance to other classes of antibiotics were also associated with mobile genetic elements. The armA, mph(E) and msr(E) genes were located on a Tn6180-derived fragment of the resistance island AbGRI3, flanked by partial sequences of ISEc28 and IS26. ISEc28 was positioned upstream of armA, followed by a permease gene, an ISEc29 inserted upstream of the macrolide resistance genes mph(E) and msr(E), and an ISAba24 located downstream of those genes (Figure 3A). This structure was identical in isolates PR1 (three copies), PR2, PR3 (two copies), PR4 and PR8. In isolates PR5 and PR6, a single copy of the same structure was present, except that the IS26 element was complete (Figure 3B). The aminoglycoside resistance genes aph(3’’)-Ib and aph(6)-Id, together with the tetracycline resistance gene tet(B), were found in close proximity to the transposon Tn2006 in all isolates belonging to IC2, presenting an AbGRI1-like resistance island structure. The insertion sequence ISVsa3 was identified downstream of aph(3’’)-Ib and aph(6)-Id, and upstream of tet(B). Additionally, two copies of the tnsE gene, typically associated with Tn7-like transposons, were also detected downstream the tet(B) gene and upstream of the aminoglycoside resistance genes (Figure 3C).
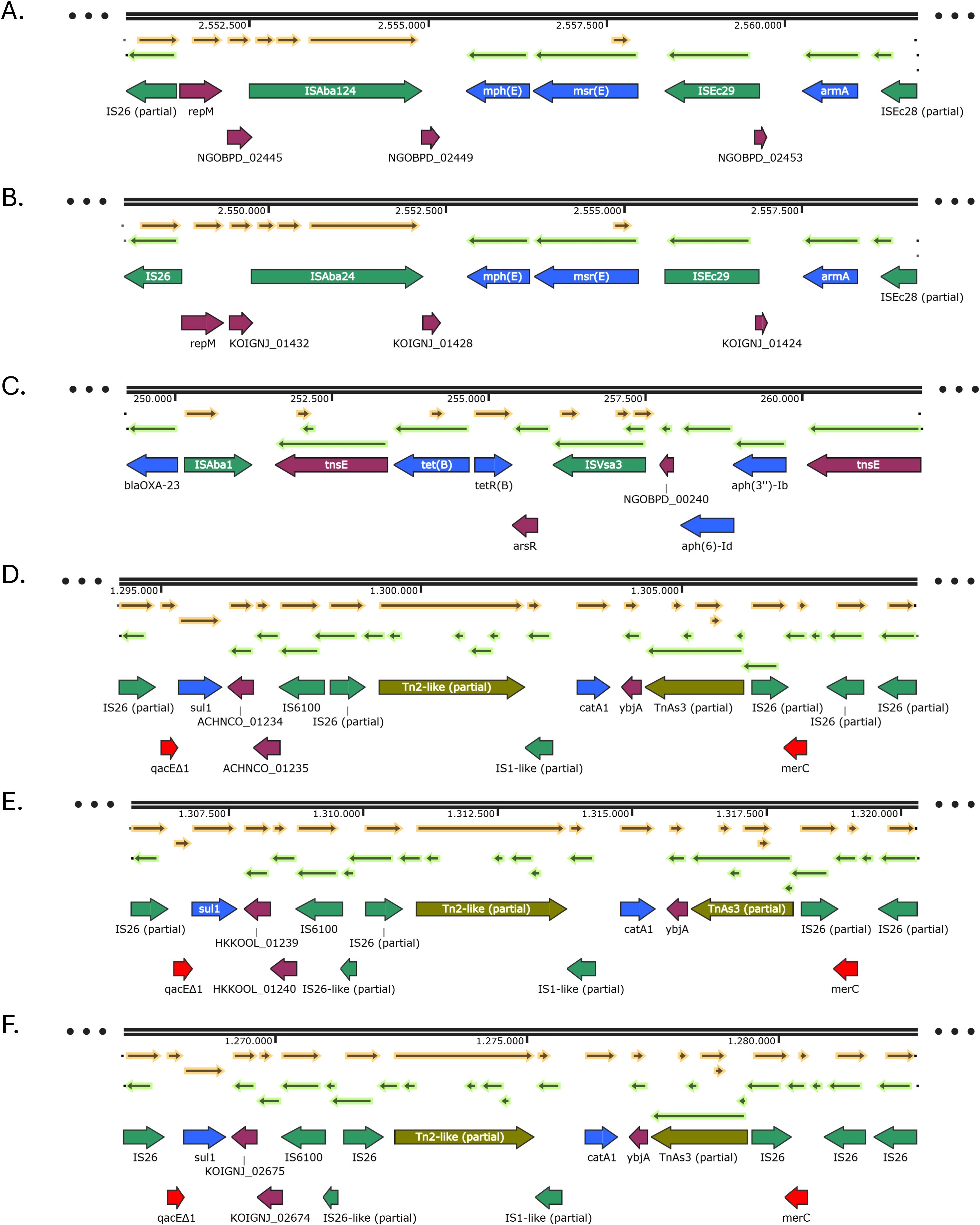
Figure 3. Genetic environments of other antibiotic resistance genes found in the IC2 isolates: armA, msr(E) and mph(E) (A, B); aph(3’’)-Ib, aph(6)-Id and tet(B) (C); sul1 and catA1 (D-F). Arrows represent open reading frames, with colors indicating gene function: resistance genes in blue, mobile genetic elements in green, and other genes in purple. Arrow direction indicates transcriptional orientation.
A complex genetic environment harboring the resistance genes sul1 and catA1 was identified in the IC2 isolates (Figures 3D-F). In isolates PR2 and PR8, a ~15 kb resistance island was observed, including the genes sul1, catA1, qacEΔ1 and merC, embedded within a mosaic of mobile genetic elements. The sul1 gene was located downstream of the antiseptic resistance gene qacEΔ1, which in turn was preceded by a partial IS26 element. Downstream of sul1, an IS6100 element and another partial IS26 element were present, followed by a partial Tn2-like transposon and a partial IS1-like insertion sequence. Further downstream and in the opposite orientation, the catA1 gene was found, followed by ybjA, a partial TnAs3 transposase gene, a partial IS26 element, the mercury resistance gene merC, and two additional IS26 partial sequences (Figure 3D). In isolates PR3 and PR4, a similar arrangement was identified, with a key difference, the IS6100 element was interrupted by a partial IS26-like sequence, and one of the downstream IS26 elements upstream of merC was absent (Figure 3E). In isolates PR5 and PR6, the gene arrangement was a combination of the two previously described contexts. In these isolates, the IS26 sequences were complete, and IS6100 was again interrupted by a partial IS26-like element. Additionally, two IS26 copies were found upstream of the merC gene (Figure 3F).
In isolate PR7, the aph(6)-Id and aph(3’’)-Ib genes were located within the transposon Tn5393c, which includes an ISAba1 element upstream of the aminoglycoside resistance genes. This transposon was embedded within a Tn6250-like structure that also carries the sul2 gene. Notably, this Tn6250 variant harbors an additional ISAba125 element inserted between the traA and traD genes, which is not present in the canonical Tn6250 structure (Figure 4A). Regarding the aminoglycoside resistance gene aph(3’’)-VIa, it was only present in isolate PR7, and was bracketed by two ISAba125 forming a transposon named TnaphA6 (Figure 4B). In addition, dfrA1, sat2 and aadA1 genes were found in a typical class 2 integron structure, located between the Tn7 transposition module (tnsABCDE) and the integrase gene intI2. Simultaneously, the Tn3 transposon harboring the blaTEM-1 gene was embedded in the Tn7 transposon forming a composite Tn7::Tn3 structure (Figure 4C).
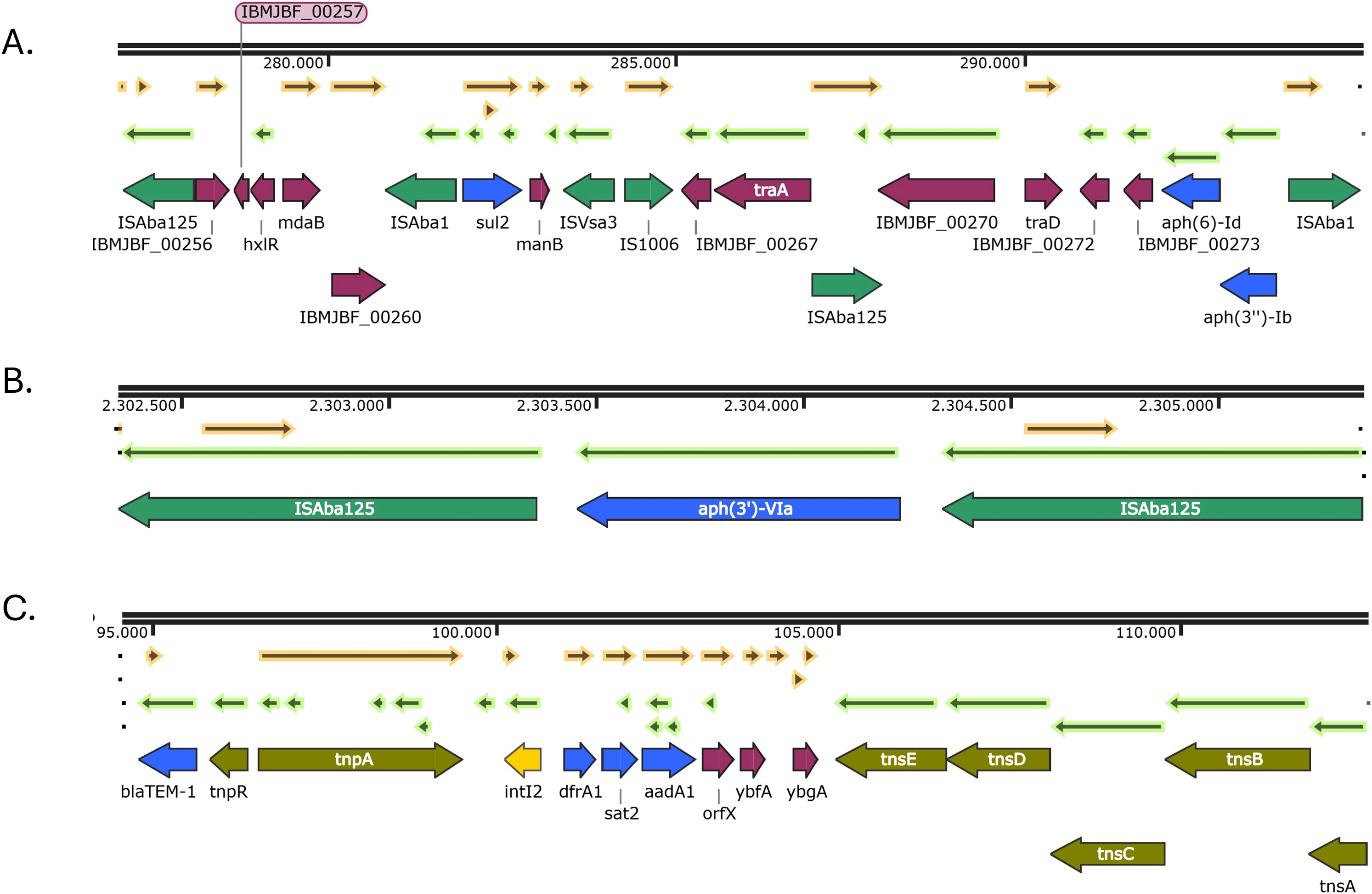
Figure 4. Genetic environments of other antibiotic resistance genes found in the IC5 isolate: aph(6)-Id, aph(3’’)-Ib and sul2 (A); aph(3’)-VIa (B); dfrA1, sat2 and aadA1 (C). Arrows represent open reading frames, with colors indicating gene function: resistance genes in blue, insertion sequences and transposons in green, integrons in yellow and other genes in purple. Arrow direction indicates transcriptional orientation.
3.5 Plasmid analysis and location of the resistance genes
Conventional lysis experiments showed plasmid profiles including plasmids ranging in size from 2 to around 13.6 Kb (Figure 5). Sequencing further confirmed the presence of a plasmid of 8.73 Kb in all the isolates except PR7, an additional plasmid of 2.28 Kb in isolates PR4, PR5 and PR6; and a plasmid of 10.85 Kb in isolate PR7 were also detected.
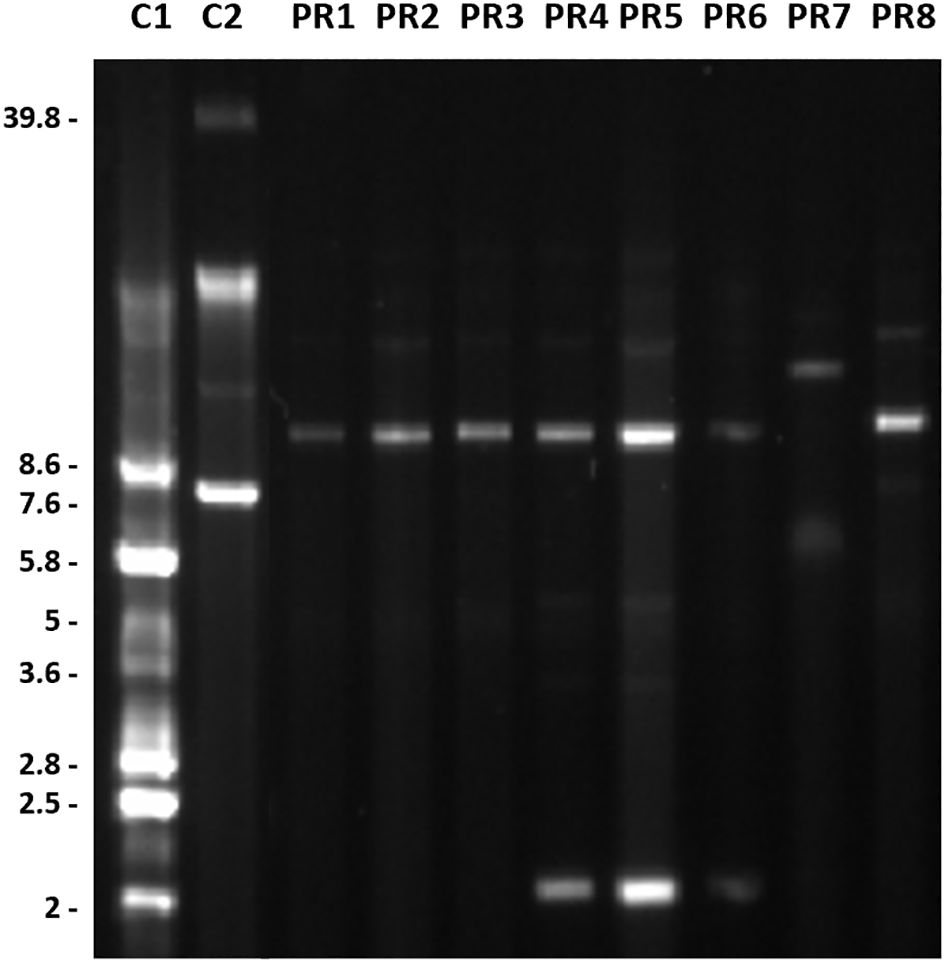
Figure 5. Plasmid profiles of the A. baumannii isolates recovered from the National Hospital of Itaugua, Paraguay. Molecular weights are expressed in Kb. Isolates C1, E. coli CECT 678 (NCTC 50193) and C2, E. coli CECT 679 (NCTC 50191) were used as molecular weight markers.
PCR-based replicon typing experiments revealed the presence of replicases of group GR2 (Aci1/Aci2) in all the isolates except PR7, which was positive for a group GR4 (Aci4) replicase. Additionally, PR1-PR7 isolates were also positive for a replicase of group GR19 (rep135040). Replicon typing using the Acinetobacter Plasmid Typing database allowed us to type the 8.73 Kb plasmids as r3-T1 (Aci1) and the 2.27 Kb plasmids as r3-T15. The 10.84 Kb plasmid of isolate PR7 was typed as r3-T14. Additionally, we also detected a r3-T60 replicase gene in the contigs belonging to the chromosomes of the eight isolates, and that region had a higher read coverage than the rest of the chromosome. In order to confirm the integration of that plasmid in the chromosome and to exclude the possibility of an assembly error, long-reads were aligned to the hybrid assemblies to find a read long enough to analyze the genes flanking the replicase gene. The analysis of those genes showed that they were involved in metabolic processes that are usually coded by the chromosome in the reference strains ATCC 19606 and ATCC 17978, which allowed us to conclude the assembly was correct. Although the isolates were carrying at least one plasmid, no resistant genes were located within those structures, but toxin-antitoxin systems (brnT/brnA), septicolysin and TonB-dependent receptor znuD2 genes were found.
3.6 Phenotypic analysis of virulence factors: biofilm production and motility
Statistically significant differences in biofilm production were observed among the isolates (One-way ANOVA, p < 0.0001), as shown in Supplementary Figure S4. All isolates except PR6 (p = 0.9932, Tukey’s post hoc test) exhibited significantly higher biofilm production compared to the negative control (p < 0.05). Notably, isolates PR1, PR4, and PR7 showed significantly greater biofilm production than the positive control (p < 0.05), indicating an enhanced biofilm-forming capacity.
All the isolates showed the same motility phenotype, more precisely, morphotype D according to the classification method described by Cosgaya et al (Cosgaya et al., 2019).
4 Discussion
Paraguay, like other Latin-American countries, has published little information on the incidence and epidemiology of A. baumannii. During 2022, an average of 743 microorganisms (bacteria and fungi) were recovered in the NHI, including isolates recovered upon hospital admission or and during hospitalization, of which 11% were identified as A. baumannii. Considering only the isolates recovered during hospitalization, A. baumannii is the most commonly isolated bacteria in the hospital. Thus, A. baumannii is of great concern to the healthcare community and it poses a challenge in the treatment of infections due to its high level of resistance to antibiotics (Rodríguez et al., 2018; Manobanda Nata and Jaramillo Ruales, 2023).
Carbapenem resistance rates of the A. baumannii isolates recovered in NHI in 2022 were above 80%. However, data obtained from the first two months of 2024 suggests that the carbapenem resistance rates are increasing up to 90% which is in accordance with data previously reported in the region (Melgarejo-Touchet et al., 2021; Manobanda Nata and Jaramillo Ruales, 2023). In the present study the majority of the A. baumannii isolates from the ICU patients were recovered from respiratory samples, which has also been reported in other studies, where isolates were recovered mainly from ICU patients that were suffering from pneumonia (Montrucchio et al., 2022; Brizuela Centurión et al., 2023; Castanheira et al., 2023).
Clonal relatedness analysis by MLST and cgMLST allowed us to identify the international clonal lineage to which the investigated isolates belong. A single isolate (PR7) was assigned to IC5 (ST79 Pasteur scheme), the so-called Pan-American clone that is characteristic and commonly described in CRAB isolates in Latin America, often along with clone IC7 (ST25) carrying the blaOXA-23 gene, confirming its circulation in the area (Rodríguez et al., 2016; Nodari et al., 2020). It should be noted that the characteristics of this clone are consistent with those previously described by our group in strains from the neighboring country, Bolivia (Cerezales González, 2018). Most isolates were assigned to IC2, a lineage known for its global distribution, although its presence in Latin-American countries is rarely reported (Castillo-Ramirez, 2023; Morgado et al., 2023), as in Brazil in 2008 and 2015-2016 (Rodríguez et al., 2016; Camargo et al., 2022). Within the isolates belonging to IC2, ST2 (Pasteur scheme) was the most prevalent sub-lineage, which is in accordance with the reported by other authors worldwide (Hamidian and Nigro, 2019). The most recent region-wide study also including isolates from Paraguay between July 2013 and June 2014, showed that the circulating clones belonged to IC5 (ST79 Pasteur) and IC7 (ST25 Pasteur), but no IC2 isolates were detected (Rodríguez et al., 2016). The presence of IC2 isolates carrying the blaOXA-23 gene in this study confirms the regional distribution of clone IC2, as described by other authors who identified this clone in South America (Levy-Blitchtein et al., 2018; Camargo et al., 2022; Martins-Gonçalves et al., 2024). During the COVID-19 pandemic, factors such as the massive influx of patients into ICUs and the increased use of antimicrobial agents intensified the selective pressure for multidrug-resistant pathogens. This led to outbreaks worldwide and also, in neighboring countries to Paraguay such as Brazil, where outbreaks caused by ST2 isolates, carrying blaOXA-23, were identified between 2020 and 2021 (Camargo et al., 2022).
The detailed analysis of the antibiotic resistance genes showed the presence of two blaOXA-51 variants, blaOXA-66 and blaOXA-65, with blaOXA-66 as the characteristic variant of the IC2 clone and in line with its emergence at a regional level (Camargo et al., 2022). As previously mentioned, all the isolates were carrying the carbapenemase gene blaOXA-23, a higher prevalence than the previously reported by Melgarejo et al (Melgarejo-Touchet et al., 2021). It is worth mentioning that IC2 isolates carried two copies of the gene, a frequent phenomenon among clinical A. baumannii isolates, according to the literature (Hua et al., 2016). These genes were located within transposons Tn2006 and Tn2008, a commonly described structure that plays a key role in the overexpression and mobilization of the gene (Hamidian and Nigro, 2019; Sánchez-Urtaza et al., 2023). Additionally, two different variants of the blaADC gene were identified: blaADC-73 and blaADC-5. The isolates belonging to IC2 were carrying blaADC-73 gene with ISAba1 upstream of it, a mechanism of overexpression that is commonly reported in clinical isolates (Nodari et al., 2020; Sánchez-Urtaza et al., 2023; Hamed et al., 2023). However, the blaADC-5 gene that was identified in the isolate belonging to IC5 was accompanied by IS3 upstream of it, which differs from the previously reported in other isolates from clone IC5 (Lopes et al., 2013).
Additional genes conferring resistance to other classes of antibiotics were also identified in isolates belonging to clone IC2: armA, aph(6)-Id, aph(3’’)-Ib, msr(E), mph(E), sul1, catA1 and tet(B); genes that have been reported recently in isolates that caused an outbreak in Brazil (Camargo et al., 2022). It is worth mentioning that in those countries where the IC2 isolates carrying the armA gene was detected and was predominant, they exhibited high resistance rates to aminoglycosides such as amikacin, gentamicin and tobramycin (Camargo et al., 2022; Martins-Gonçalves et al., 2024). In this study, the armA, mph(E) and msr(E) genes were identified in the chromosome within a Tn6180-derived fragment of the resistance island AbGRI3. This arrangement is globally spread and has been previously described in other CRAb isolates assigned to ST2 (Pasteur Scheme) and also ST195/1816 (Oxford Scheme), which is in accordance with the genetic contexts identified in our isolates (Wiradiputra et al., 2023; Wei et al., 2024). Although chromosomal integration may reduce the immediate risk of horizontal transfer compared to plasmid-borne elements, it can promote stable maintenance of these resistance genes within bacterial populations, and also, the presence of these genes in a Tn6180-derived structure suggests the potential for mobilization under selective pressure. This highlights the importance of monitoring chromosomal resistance islands as reservoirs of clinically significant resistance determinants. The aph genes (aph(6)-Id, aph(3’’)-Ib) together with tet(B) were found close to the carbapenemase gene blaOXA-23 in an AbGRI1-like resistance island in all the isolates belonging to IC2. This island has been repeatedly documented in clinical IC2 isolates across different countries and contributes significantly to their multidrug-resistant phenotype (Cerezales et al., 2020; Vijayakumar et al., 2022; Wiradiputra et al., 2023; Naderi et al., 2023). All the studied isolates belonging to IC2 were also harboring sul1 and catA1 together with qacEΔ1 (antiseptic resistance) and merC (mercury resistance) genes in a complex structure of ~15 kb involving multiple mobile genetic elements such as IS26, IS6100 and Tn2-like elements, that closely resembles mosaic resistance regions previously described in Acinetobacter genomic islands (Hamidian and Hall, 2018b). Moreover, IC2 isolates have been shown to carry AbGRI variants with class 1 integron backbones containing sul1 and qacEΔ1 together with IS26-mediated rearrangements (Harmer et al., 2023). These findings support the hypothesis that this region represents a chromosomally integrated resistance island related to AbaR or AbGRI structures that emerged through IS26‐mediated horizontal gene capture.
The resistome of the IC5 isolate also included a variety of genes conferring resistance to non-β-lactam antibiotics such as aminoglycosides (armA, aph(6)-Id, aph(3’’)-Ib, aph(3’)-VIa, sat2 and aadA1), sulfonamides (sul2) and trimethoprim (dfrA1), which are commonly described in CRAb strains belonging to this clone (Nodari et al., 2020). In this isolate the aph(6)-Id, aph(3’’)-Ib and sul2 genes were identified within a non-canonical variant of the transposon Tn6250, characterized by the insertion of an ISAba125 element between the traA and traD genes. This transposon has been previously described by other authors (Brito et al., 2022), however, the presence of the ISAba125 was not previously reported. From a clinical perspective, such rearrangements may influence the mobility and transfer efficiency of resistance genes, potentially enhancing their spread across bacterial populations. The aph(3’)-VIa gene, is commonly associated with resistance to amikacin and is typically found within the TnAphA6 transposon, a composite element bracketed by ISAba125 insertion sequences (Khongfak et al., 2022; Brito et al., 2022). This transposon has been frequently identified in A. baumannii isolates, particularly those belonging to IC5, suggesting a strong association between this mobile element and the clonal lineage (Brito et al., 2022; Jouybari et al., 2021; Morgado et al., 2023). Moreover, TnAphA6 has been found both on conjugative plasmids and integrated into the chromosome, often in multidrug-resistant backgrounds (Khongfak et al., 2022; Brito et al., 2022). The mobility conferred by IS26 likely facilitates its dissemination not only through clonal expansion but also via horizontal gene transfer. The presence of aph(3’)-VIa within the TnAphA6 transposon in our IC5 isolate further supports its role in the global spread of aminoglycoside resistance within this lineage. In addition, dfrA1, sat2 and aadA1 genes were also identified together in isolate PR7 within a class 2 integron structure involving the blaTEM-1 gene. However, the additional insertion of a Tn3 transposon harboring blaTEM-1 within the Tn7 backbone, forming a Tn7::Tn3 composite transposon, suggests active modular recombination events. This unusual arrangement may enhance the mobility and plasticity of Tn7 by incorporating new resistance determinants. Similar nested transposition events have been reported for instance in ST79 A. baumannii isolates from Chile and Bolivia (Brito et al., 2022; Cerezales et al., 2020) but are relatively rare, and their presence highlights the role of Tn7 as a key element for the acquisition of multiple resistance genes, particularly in multidrug-resistant A. baumannii.
Regarding the virulome of the isolates, most of the genes were involved in biofilm production, which is key for survival and invasion of host’s cells (Kumar et al., 2021). All the isolates were harboring genes coding for Csu fimbriae, BfmRS system, Bap protein, extracellular polysaccharide PNAG, OmpA, Ata transporter and AbaI/AbaR proteins, which according to the literature, are linked with the ability to form biofilm (Ghasemi et al., 2018). Another important virulence factor is the capsule polysaccharide K locus (KL) and outer core OC locus (OCL) (Müller et al., 2023). Capsular polysaccharide genes vary between isolates within the same species, and recent studies associate certain types of capsular polysaccharides or their alterations with increased virulence and biofilm production (Cahill et al., 2022). In this work, different capsule types were identified: OCL1/KL3, OCL1/KL81 and OCL10/KL9. Isolates belonging to IC2 had the OCL1 gene cluster, the most predominant among IC1 and IC2 according to the literature (Kenyon et al., 2014). Among the IC2 isolates, it is common to find a wide variety of different K loci, with KL3 gene cluster predominant according to other authors (Müller et al., 2023; Camargo et al., 2022; Cahill et al., 2022), which is in accordance with our results, where 4 out of 7 IC2 isolates were KL3. The OCL10 gene cluster is predominantly found in isolates belonging to IC5, as well as K locus KL9 (Müller et al., 2023), which is in accordance with our findings. Despite sharing identical virulence gene content and OCL/KL loci as other biofilm producing strains, isolate PR6 exhibited significantly reduced biofilm formation. A possible explanation is that this phenotype results from differences in the expression of biofilm-related genes, which are regulated by the BfmRS two-component system and the csu operon—both essential for pili-mediated surface attachment. Disruption of either bfmR or bfmS can lead to reduced csu expression, thereby preventing biofilm formation (Gao et al., 2019).
Plasmid analysis showed the presence of a RepAci1 plasmid of 8.73 Kb in all the isolates belonging to IC2. This plasmid has been described previously in isolates belonging to IC1, IC2 and IC3, encoding a toxin-antitoxin system (brnT/brnA), septicolysin and TonB-dependent receptor ZnuD2 (Aranzamendi et al., 2024; Hamidian and Hall, 2018a), but unlike some of those isolates, the Paraguayan isolates were not harboring any carbapenemase gene in their plasmids. In fact, some variants of r3-T1 plasmids carrying a carbapenemase gene appear to be limited to North America and Europe (Lam and Hamidian, 2024). Replicon typing experiments by PCR were positive for replicase Aci4, however, typing using Acinetobacter plasmid typing database published by Lam et al. (2023) did not show the presence of that replicase, instead, r3-T14 was identified, which is very similar to the r3-T11 (Aci4) and may explain the positive result in the PCR experiment (Lam et al., 2023).
The results obtained in this study allowed us to update and get an approximate picture of the current antibiotic resistance rates, resistome, virulome, plasmid profiles and also the circulating clones of A. baumannii in Paraguay. This type of study is crucial not only for gaining knowledge about the characteristics of the clones circulating in the country, but also to design and implement control measures to prevent outbreaks.
Data availability statement
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found in the article/Supplementary Material.
Ethics statement
The study was conducted in accordance with the Declaration of Helsinki and approved by the Ethics Committee of the University of the Basque Country CEIAB-UPV/EHU, M30/2024/140).
Author contributions
SS-U: Conceptualization, Data curation, Formal analysis, Investigation, Methodology, Software, Validation, Visualization, Writing – original draft, Writing – review & editing. LA-A: Formal Analysis, Visualization, Methodology, Writing – original draft, Investigation. RAP: Methodology, Writing – review & editing. TB: Methodology, Writing – review & editing. AO-S: Methodology, Writing – review & editing. RG: Resources, Writing – review & editing, Investigation. KX: Methodology, Supervision, Validation, Writing – review & editing. PH: Supervision, Investigation, Writing – review & editing, Software, Methodology, Resources, Visualization, Funding acquisition, Validation, Formal Analysis. IA: Project administration, Writing – review & editing, Funding acquisition. LG: Visualization, Conceptualization, Resources, Writing – review & editing, Formal Analysis, Supervision, Writing – original draft, Validation.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. This research was funded by the Department of Education of the Basque Government (Research Groups of the Basque University System 2021 grant number Group IT1578-22, GIC21/18), the Ministry of Science and Innovation of the Spanish Government (Grant N. PID2020-116495RB-I00). Dr. Sandra Sánchez-Urtaza is a postdoctoral researcher awarded with the Basque Government’s Postdoctoral Fellowship (2023–2026).
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fcimb.2025.1620479/full#supplementary-material
References
Aranzamendi, M., Xanthopoulou, K., Sánchez-Urtaza, S., Burgwinkel, T., Arazo Del Pino, R., Lucaßen, K., et al. (2024). Genomic surveillance uncovers a 10-year persistence of an OXA-24/40 acinetobacter baumannii clone in a tertiary hospital in Northern Spain. Int. J. Mol. Sci. 25, 2333. doi: 10.3390/ijms25042333
Bertini, A., Poirel, L., Mugnier, P. D., Villa, L., Nordmann, P., and Carattoli, A. (2010). Characterization and PCR-based replicon typing of resistance plasmids in acinetobacter baumannii. Antimicrobial Agents Chemother. 54, 4168–4177. doi: 10.1128/AAC.00542-10
Brito, B. P., Koong, J., Wozniak, A., Opazo-Capurro, A., To, J., Garcia, P., et al. (2022). Genomic analysis of carbapenem-Resistant acinetobacter baumannii strains recovered from Chilean hospitals reveals lineages specific to south america and multiple routes for acquisition of antibiotic resistance genes. Microbiol. Spectr. 10, 2463. doi: 10.1128/spectrum.02463-22
Brizuela Centurión, A. M., Martí Ruiz, A., Fusillo Montanaro, J. D., Oleynick Goncalves, A. F., and Ayala Careaga, J. B. (2023). Aislamiento de gérmenes oportunistas en pacientes internados en Unidad de Cuidados Intensivos. Revista de salud publica del Paraguay 1 (13), 35–39. doi: 10.18004/rspp.2023.abril.05
Cahill, S. M., Hall, R. M., and Kenyon, J. J. (2022). An update to the database for Acinetobacter baumannii capsular polysaccharide locus typing extends the extensive and diverse repertoire of genes found at and outside the K locus. Microbial. Genomics 8 (10), mgen000878. doi: 10.1099/mgen.0.000878
Camacho, C., Coulouris, G., Avagyan, V., Ma, N., Papadopoulos, J., Bealer, K., et al. (2009). BLAST+: architecture and applications. BMC Bioinf. 10, 421–421. doi: 10.1186/1471-2105-10-421
Camargo, C. H., Yamada, A. Y., Nagamori, F. O., de Souza, A. R., Tiba-Casas, M. R., de Moraes França, F. A., et al. (2022). Clonal spread of ArmA- and OXA-23-coproducing Acinetobacter baumannii International Clone 2 in Brazil during the first wave of the COVID-19 pandemic. J. Med. Microbiol. 71 (4). doi: 10.1099/jmm.0.001509
Castanheira, M., Mendes, R. E., and Gales, A. C. (2023). Global epidemiology and mechanisms of resistance of acinetobacter baumannii-calcoaceticus complex. Clin. Infect. Dis.: Off. Publ. Infect. Dis. Soc. America 76, S166–S178. doi: 10.1093/cid/ciad109
Castillo-Ramirez, S. (2023). Genomic epidemiology of Acinetobacter baumannii goes global. mBio 14, e0252023–23. doi: 10.1128/mbio.02520-23
Cerezales, M., Biniossek, L., Gerson, S., Xanthopoulou, K., Wille, J., Wohlfarth, E., et al. (2021). Novel multiplex PCRs for detection of the most prevalent carbapenemase genes in Gram-negative bacteria within Germany. J. Med. Microbiol. 70 (3). doi: 10.1099/jmm.0.001310
Cerezales, M., Xanthopoulou, K., Wille, J., Krut, O., Seifert, H., Gallego, L., et al. (2020). Mobile genetic elements harboring antibiotic resistance determinants in acinetobacter baumannii isolates from Bolivia. Front. Microbiol. 11. doi: 10.3389/fmicb.2020.00919
Cerezales González, M. (2018). Molecular characterization of Acinetobacter spp. and their antibiotic resistance mobilome from clinical isolates collected in Bolivia (Leioa, Spain: University of the Basque Country).
Cosgaya, C., Ratia, C., Marí-Almirall, M., Rubio, L., Higgins, P. G., Seifert, H., et al. (2019). In vitro and in vivo Virulence Potential of the Emergent Species of the Acinetobacter baumannii (Ab) Group. Front. Microbiol. 10. doi: 10.3389/fmicb.2019.02429
Gao, Q., Meng, X., Gu, H., Chen, X., Yang, H., Qiao, Y., et al. (2019). Two phenotype-differentiated acinetobacter baumannii mutants that survived in a meropenem selection display large differences in their transcription profiles. Front. Microbiol. 10. doi: 10.3389/fmicb.2019.02308
Ghasemi, E., Ghalavand, Z., Goudarzi, H., Yeganeh, F., Hashemi, A., Dabiri, H., et al. (2018). Phenotypic and genotypic investigation of biofilm formation in clinical and environmental isolates of acinetobacter baumannii. Arch. Clin. Infect. Dis. 13 (4), e12914. doi: 10.5812/archcid.12914
Hamed, S. M., Elkhatib, W. F., Brangsch, H., Gesraha, A. S., Moustafa, S., Khater, D. F., et al. (2023). Acinetobacter baumannii global clone-specific resistomes explored in clinical isolates recovered from Egypt. Antibiotics 12, 1149. doi: 10.3390/antibiotics12071149
Hamidian, M. and Hall, R. M. (2018a). Genetic structure of four plasmids found in Acinetobacter baumannii isolate D36 belonging to lineage 2 of global clone 1. PloS One 13, e0204357. doi: 10.1371/journal.pone.0204357
Hamidian, M. and Hall, R. M. (2018b). The AbaR antibiotic resistance islands found in Acinetobacter baumannii global clone 1 – Structure, origin and evolution. Drug Resistance Updates 41, 26–39. doi: 10.1016/j.drup.2018.10.003
Hamidian, M. and Nigro, S. J. (2019). Emergence, molecular mechanisms and global spread of carbapenem-resistant Acinetobacter baumannii. Microbial. Genomics 5 (10), e000306. doi: 10.1099/mgen.0.000306
Harmer, C. J., Nigro, S. J., and Hall, R. M. (2023). Acinetobacter baumannii GC2 Sublineage Carrying the aac(6′)-Im Amikacin, Netilmicin, and Tobramycin Resistance Gene Cassette. Microbiol. Spectr. 11, 1204. doi: 10.1128/spectrum.01204-23
Higgins, P. G., Lehmann, M., and Seifert, H. (2010a). Inclusion of OXA-143 primers in a multiplex polymerase chain reaction (PCR) for genes encoding prevalent OXA carbapenemases in Acinetobacter spp. Int. J. Antimicrobial Agents 35, 305. doi: 10.1016/j.ijantimicag.2009.10.014
Higgins, P. G., Lehmann, M., Wisplinghoff, H., and Seifert, H. (2010b). gyrB multiplex PCR to differentiate between acinetobacter calcoaceticus and acinetobacter genomic species 3. J. Clin. Microbiol. 48, 4592–4594. doi: 10.1128/JCM.01765-10
Higgins, P. G., Pérez-Llarena, F. J., Zander, E., Fernández, A., Bou, G., and Seifert, H. (2013). OXA-235, a novel class D β-lactamase involved in resistance to carbapenems in acinetobacter baumannii. Antimicrobial Agents Chemother. 57, 2121–2126. doi: 10.1128/AAC.02413-12
Hua, X., Shu, J., Ruan, Z., Yu, Y., and Feng, Y. (2016). Multiplication of blaoxa-23 is common in clinical Acinetobacter baumannii, but does not enhance carbapenem resistance. J. Antimicrobial Chemother. 71, 3381–3385. doi: 10.1093/jac/dkw310
Jeon, J. H., Jang, K., Lee, J. H., Kang, L., and Lee, S. H. (2023). Transmission of antibiotic resistance genes through mobile genetic elements in Acinetobacter baumannii and gene-transfer prevention. Sci. Total Environ. 857, 159497. doi: 10.1016/j.scitotenv.2022.159497
Jolley, K. A., Bray, J. E., and Maiden, M. C. J. (2018). Open-access bacterial population genomics: BIGSdb software, the PubMLST.org website and their applications. Wellcome Open Res. 3, 124. doi: 10.12688/wellcomeopenres.14826.1
Jouybari, M. A., Ahanjan, M., Mirzaei, B., and Goli, H. R. (2021). Role of aminoglycoside-modifying enzymes and 16S rRNA methylase (ArmA) in resistance of Acinetobacter baumannii clinical isolates against aminoglycosides. Rev. Sociedade Bras. Med. Trop. 54, e05992020–2020. doi: 10.1590/0037-8682-0599-2020
Jünemann, S., Sedlazeck, F. J., Prior, K., Albersmeier, A., John, U., Kalinowski, J., et al. (2013). Updating benchtop sequencing performance comparison. Nat. Biotechnol. 31, 294–296. doi: 10.1038/nbt.2522
Kenyon, J. J., Nigro, S. J., and Hall, R. M. (2014). Variation in the OC locus of acinetobacter baumannii genomes predicts extensive structural diversity in the lipooligosaccharide. PloS One 9, e107833. doi: 10.1371/journal.pone.0107833
Khongfak, S., Thummeepak, R., Leungtongkam, U., Tasanapak, K., Thanwisai, A., and Sitthisak, S. (2022). Insights into mobile genetic elements and the role of conjugative plasmid in transferring aminoglycoside resistance in extensively drug-resistant Acinetobacter baumannii AB329. PeerJ 10, e13718. doi: 10.7717/peerj.13718
Kumar, S., Anwer, R., and Azzi, A. (2021). Virulence potential and treatment options of multidrug-resistant (MDR) acinetobacter baumannii. Microorganisms 9, 2104. doi: 10.3390/microorganisms9102104
Kyriakidis, I., Vasileiou, E., Pana, Z. D., and Tragiannidis, A. (2021). Acinetobacter baumannii antibiotic resistance mechanisms. Pathogens 10, 373. doi: 10.3390/pathogens10030373
Lam, M. M. C. and Hamidian, M. (2024). Examining the role of Acinetobacter baumannii plasmid types in disseminating antimicrobial resistance. NPJ Antimicrobials Resistance 2, 1. doi: 10.1038/s44259-023-00019-y
Lam, M. M. C., Koong, J., Holt, K. E., Hall, R. M., and Hamidian, M. (2023). Detection and Typing of Plasmids in Acinetobacter baumannii Using rep Genes Encoding Replication Initiation Proteins. Microbiol. Spectr. 11, e0247822. doi: 10.1128/spectrum.02478-22
Lee, C., Lee, J. H., Park, M., Park, K. S., Bae, I. K., Kim, Y. B., et al. (2017). Biology of acinetobacter baumannii: pathogenesis, antibiotic resistance mechanisms, and prospective treatment options. Front. Cell. Infect. Microbiol. 7. doi: 10.3389/fcimb.2017.00055
Levy-Blitchtein, S., Roca, I., Plasencia-Rebata, S., Vicente-Taboada, W., Velásquez-Pomar, J., Muñoz, L., et al. (2018). Emergence and spread of carbapenem-resistant Acinetobacter baumannii international clones II and III in Lima, Peru. Emerg. Microbes Infect. 7, 1–9. doi: 10.1038/s41426-018-0127-9
Lewis, J. S., II and Clinical and Laboratory Standards Institute (2024). Performance standards for antimicrobial susceptibility testing. 34th (Wayne, PA: Clinical and Laboratory Standards Institute).
Liu, B., Zheng, D., Jin, Q., Chen, L., and Yang, J. (2018). VFDB 2019: a comparative pathogenomic platform with an interactive web interface. Nucleic Acids Res. 47, 687. doi: 10.1093/nar/gky1080
Lopes, B. S., Gallego, L., and Amyes, S. G. B. (2013). Multi-drug resistance profiles and the genetic features of Acinetobacter baumannii isolates from Bolivia. J. Infect. Developing Countries 7, 323–328. doi: 10.3855/jidc.2711
Magiorakos, A. P., Srinivasan, A., Carey, R. B., Carmeli, Y., Falagas, M. E., Giske, C. G., et al. (2012). Multidrug-resistant, extensively drug-resistant and pandrug-resistant bacteria: an international expert proposal for interim standard definitions for acquired resistance. Clin. Microbiol. Infect. 18, 268–281. doi: 10.1111/j.1469-0691.2011.03570.x
Manobanda Nata, C. I. and Jaramillo Ruales, E. K. (2023). Carbapenem-resistant Acinetobacter baumannii complex: a review in Latin America. Salud Ciencia Tecnol. 3, 479. doi: 10.56294/saludcyt2023479
Martins-Gonçalves, T., Pimenta, J. S., Fontana, H., Esposito, F., Melocco, G., Dantas, K., et al. (2024). Acinetobacter baumannii international clone 2 co-producing OXA-23, NDM-1, and ArmA emerging in South America. Antimicrobial Agents Chemother. 68, e0029824. doi: 10.1128/aac.00298-24
Melgarejo-Touchet, N., Brítez, C. M., Busignani, S., Falcón, M., López, E., Laconich, M., et al. (2021). Caracterización molecular de carbapenemasas en bacilos gramnegativos circulantes en hospitales de Paraguay. Primer cuatrimestre 2021. Memorias Del Instituto Investigaciones En Cienc. La Salud 19, 49–58. doi: 10.18004/mem.iics/1812-9528/2021.019.02.49
Montrucchio, G., Corcione, S., Lupia, T., Shbaklo, N., Olivieri, C., Poggioli, M., et al. (2022). The burden of carbapenem-resistant acinetobacter baumannii in ICU COVID-19 patients: A regional experience. J. Clin. Med. 11, 5208. doi: 10.3390/jcm11175208
Morgado, S. M., Fonseca, ÉL., Freitas, F. S., Bighi, N. S., Oliveira, P. P. C., Monteiro, P. M., et al. (2023). Outbreak of high-risk XDR CRAB of international clone 2 (IC2) in Rio Janeiro, Brazil. J. Global Antimicrobial Resistance 34, 91–98. doi: 10.1016/j.jgar.2023.06.011
Müller, C., Reuter, S., Wille, J., Xanthopoulou, K., Stefanik, D., Grundmann, H., et al. (2023). A global view on carbapenem-resistant Acinetobacter baumannii. MBio 14, e0226023. doi: 10.1128/mbio.02260-23
Naas, T., Oueslati, S., Bonnin, R. A., Dabos, M. L., Zavala, A., Dortet, L., et al. (2017). Beta-lactamase database (BLDB) – structure and function. J. Enzyme Inhibition Med. Chem. 32, 917–919. doi: 10.1080/14756366.2017.1344235
Naderi, G., Asadian, M., Khaki, P. A., Salehi, M., Abdollahi, A., and Douraghi, M. (2023). Occurrence of Acinetobacter baumannii genomic resistance islands (AbGRIs) in Acinetobacter baumannii strains belonging to global clone 2 obtained from COVID-19 patients. BMC Microbiol. 23, 234. doi: 10.1186/s12866-023-02961-3
Nodari, C. S., Cayô, R., Streling, A. P., Lei, F., Wille, J., Almeida, M. S., et al. (2020). Genomic analysis of carbapenem-resistant acinetobacter baumannii isolates belonging to major endemic clones in South America. Front. Microbiol. 11. doi: 10.3389/fmicb.2020.584603
O’Toole, G. A. and Kolter, R. (1998). Initiation of biofilm formation in Pseudomonas fluorescens WCS365 proceeds via multiple, convergent signalling pathways: a genetic analysis. Mol. Microbiol. 28, 449–461. doi: 10.1046/j.1365-2958.1998.00797.x
Ramirez, M. S., Bonomo, R. A., and Tolmasky, M. E. (2020). Carbapenemases: Transforming Acinetobacter baumannii into a Yet More Dangerous Menace. Biomolecules 10, 720. doi: 10.3390/biom10050720
Rodríguez, C. H., Balderrama Yarhui, N., Nastro, M., Nuñez Quezada, T., Castro Cañarte, G., Magne Ventura, R., et al. (2016). Molecular epidemiology of carbapenem-resistant Acinetobacter baumannii in South America. J. Med. Microbiol. 65, 1088–1091. doi: 10.1099/jmm.0.000328
Rodríguez, C. H., Nastro, M., and Famiglietti, A. (2018). Carbapenemases in Acinetobacter baumannii. Review of their dissemination in Latin America. Rev. Argent. Microbiol. 50, 327–333. doi: 10.1016/j.ram.2017.10.006
Ross, K., Varani, A. M., Snesrud, E., Huang, H., Alvarenga, D. O., Zhang, J., et al. (2021). TnCentral: a prokaryotic transposable element database and web portal for transposon analysis. mBio 12 (5), e0206021. doi: 10.1128/mbio.02060-21
Sánchez-Urtaza, S., Ocampo-Sosa, A., Molins-Bengoetxea, A., El-Kholy, M. A., Hernandez, M., Abad, D., et al. (2023). Molecular characterization of multidrug resistant Acinetobacter baumannii clinical isolates from Alexandria, Egypt. Front. Cell. Infect. Microbiol. 13. doi: 10.3389/fcimb.2023.1208046
Schwengers, O., Jelonek, L., Dieckmann, M. A., Beyvers, S., Blom, J., and Goesmann, A. (2021). Bakta: Rapid and standardized annotation of bacterial genomes via alignment-free sequence identification. Microbial. Genomics 7 (11), 000685. doi: 10.1099/mgen.0.000685
Tacconelli, E., Carrara, E., Savoldi, A., Harbarth, S., Mendelson, M., Monnet, D. L., et al. (2018). Discovery, research, and development of new antibiotics: the WHO priority list of antibiotic-resistant bacteria and tuberculosis. Lancet Infect. Dis. 18, 318–327. doi: 10.1016/S1473-3099(17)30753-3
Turton, J. F., Woodford, N., Glover, J., Yarde, S., Kaufmann, M. E., and Pitt, T. L. (2006). Identification of Acinetobacter baumannii by Detection of the blaOXA-51-like Carbapenemase Gene Intrinsic to This Species. J. Clin. Microbiol. 44, 2974–2976. doi: 10.1128/JCM.01021-06
Vijayakumar, S., Biswas, I., and Veeraraghavan, B. (2019). Accurate identification of clinically important acinetobacter spp.: an update. Future Sci. OA 5, FSO395. doi: 10.2144/fsoa-2018-0127
Vijayakumar, S., Jacob, J. J., Vasudevan, K., Mathur, P., Ray, P., Neeravi, A., et al. (2022). Genomic characterization of mobile genetic elements associated with carbapenem resistance of acinetobacter baumannii from India. Front. Microbiol. 13. doi: 10.3389/fmicb.2022.869653
Wei, W., Jiahui, W., Jie, W., Qinghuan, Z., Yu, Z., Yanju, H., et al. (2024). Whole genome sequencing insight into carbapenem-resistant and multidrug-resistant Acinetobacter baumannii harboring chromosome-borne blaOXA-23. Microbiol. Spectr. 12, 501. doi: 10.1128/spectrum.00501-24
Wick, R. R., Judd, L. M., Gorrie, C. L., and Holt, K. E. (2017). Unicycler: Resolving bacterial genome assemblies from short and long sequencing reads. PloS Comput. Biol. 13, e1–e1005595. doi: 10.1371/journal.pcbi.1005595
Wiradiputra, M. R. D., Thirapanmethee, K., Khuntayaporn, P., Wanapaisan, P., and Chomnawang, M. T. (2023). Comparative genotypic characterization related to antibiotic resistance phenotypes of clinical carbapenem-resistant Acinetobacter baumannii MTC1106 (ST2) and MTC0619 (ST25). BMC Genomics 24, 689. doi: 10.1186/s12864-023-09734-2
Woodford, N., Ellington, M., Coelho, J., Turton, J., Ward, M., Brown, S., et al. (2006). Multiplex PCR for genes encoding prevalent OXA carbapenemases in Acinetobacter spp. Int. J. Antimicrobial Agents 27, 351–353. doi: 10.1016/j.ijantimicag.2006.01.004
World Health Organization (2024). WHO bacterial priority pathogens list 2024: bacterial pathogens of public health importance, to guide research, development and strategies to prevent and control antimicrobial resistance. Available online at: https://iris.who.int/bitstream/handle/10665/376776/9789240093461-eng.pdf (Accessed Jan 24, 2025).
Keywords: Acinetobacter baumannii, Paraguay, whole genome sequencing, epidemiology, antibiotic resistance, plasmids
Citation: Sánchez-Urtaza S, Alfonso-Alarcón L, Arazo del Pino R, Burgwinkel T, Ocampo-Sosa A, Gonzalez R, Xanthopoulou K, Higgins PG, Alkorta I and Gallego L (2025) Genomic characterization of multidrug-resistant clinical Acinetobacter baumannii isolates from a hospital in Paraguay. Front. Cell. Infect. Microbiol. 15:1620479. doi: 10.3389/fcimb.2025.1620479
Received: 29 April 2025; Accepted: 18 July 2025;
Published: 31 July 2025.
Edited by:
Jinxin Zhao, Monash University, AustraliaReviewed by:
Andres Felipe Opazo-Capurro, University of Concepcion, ChilePiyali Chatterjee, United States Department of Veterans Affairs, United States
German Matias Traglia, Universidad de La Republica, Salto, Uruguay, Uruguay
Copyright © 2025 Sánchez-Urtaza, Alfonso-Alarcón, Arazo del Pino, Burgwinkel, Ocampo-Sosa, Gonzalez, Xanthopoulou, Higgins, Alkorta and Gallego. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Paul G. Higgins, cGF1bC5oaWdnaW5zQHVuaS1rb2Vsbi5kZQ==; Lucia Gallego, bHVjaWEuZ2FsbGVnb0BlaHUuZXVz
†These authors have contributed equally to this work