 Marialetizia Palomba1†
Marialetizia Palomba1† Aurelia Rughetti2†
Aurelia Rughetti2† Tiziana Castrignanò1*
Tiziana Castrignanò1* Chiara Napoletano2
Chiara Napoletano2 Xavier Roca-Geronès3
Xavier Roca-Geronès3 Valentina Pinna1
Valentina Pinna1 Franco Liberati1
Franco Liberati1 Daniele Canestrelli1
Daniele Canestrelli1 Simonetta Mattiucci4*
Simonetta Mattiucci4*- 1Department of Ecological and Biological Sciences, “Tuscia University”, Viterbo, Italy
- 2Department of Experimental Medicine, Sapienza University of Rome, Rome, Italy
- 3Department of Biology, Health and Environment, Section of Parasitology, Faculty of Pharmacy and Food Sciences, University of Barcelona, Barcelona, Spain
- 4Department of Public Health and Infectious Diseases, University Hospital “Policlinico Umberto I”, Sapienza-University of Rome, Rome, Italy
Anisakis pegreffii is a zoonotic marine nematode whose life-cycle involves marine organisms–small crustaceans, fish and squids as intermediate/paratenic hosts, and marine mammals, mainly cetaceans–as definitive ones. When its third-stage larvae (L3) are accidentally ingested by humans with the consumption of raw or undercooked parasitized fish and/or squids, the parasite fails to complete its life cycle, leading to human anisakiasis. Despite increasing interest in understanding the molecular basis of pathogenic effects in human anisakiasis, the transcriptomic response of A. pegreffii L3 to interaction with human immune cells, remains poorly understood. Thus, in this study, the transcriptomic profile of A. pegreffii L3 larvae under in vitro exposure to human dendritic cells (DCs) was performed for the first time. A total of 3914 differentially expressed genes (DEGs) were identified in A. pegreffii L3 after exposure to immature DCs (iDCs), by RNA-seq, allowing to detect 1868 upregulated and 2046 downregulated transcripts. Upregulated genes were significantly enriched in pathways related to energy metabolism, oxidative stress response and structural maintenance, suggesting active metabolic and structural adaptation to immune-induced stress. Conversely, genes involved in cytoskeletal organization and intracellular trafficking were downregulated, potentially reflecting the parasite’s developmental arrest in an unsuitable host such as humans. These findings provide novel insights into the molecular response pathways activated by this zoonotic parasite during the early stages of interaction with the human immune system.
Introduction
Anisakis pegreffii is a zoonotic nematode within the A. simplex (s. l.) complex, with an indirect life cycle involving cetaceans as definitive hosts, and small crustaceans, teleost fish, and cephalopods as intermediate/paratenic hosts (Mattiucci et al., 2018; 2022). Humans become accidental hosts by consuming raw or undercooked fish or squids infected with live third stage larvae (L3) of A. pegreffii (Mattiucci et al., 2018; Guardone et al., 2018). In the human, however, the parasite is unable to complete its development (Trumbić et al., 2021) but it can trigger inflammation leading to acute gastrointestinal symptoms and, if not promptly removed, marked eosinophilic infiltration, with abscess and granuloma formation (Nieuwenhuizen and Lopata, 2013; Mattiucci et al., 2013; Trumbić et al., 2021). Exposure to Anisakis antigens may also elicit IgE hypersensitization, provoking allergic reactions (Nieuwenhuizen and Lopata, 2013; Mattiucci et al., 2013; Brusca et al., 2023; EFSA, 2024). The pathogenicity of A. pegreffii is likely driven by a combination of factors, involving both mechanical invasive capacity and the release of excretory/secretory products (ESPs) (Mehrdana et al., 2017)—some of which are transported via extracellular vesicles (EVs) (Cavallero et al., 2022; Palomba et al., 2023) and miRNA (Cavallero et al., 2022; Bellini et al., 2025) —as well as complex interactions with host immune cells (Maizels and McSorley, 2016). Recent transcriptomic studies in a rodent model have shown that A. pegreffii L3 larvae upregulate genes associated with ribosomal function and oxidative phosphorylation, potentially indicating a failed attempt to molt (Trumbić et al., 2021). In contrast, in natural heterothermic fish hosts, autophagy-related pathways are activated, suggesting a dormancy-like state, highlighting the parasite’s ability to modulate its gene expression in response to diverse host environments (Trumbić et al., 2021).
However, very little is known about how A. pegreffii responds to specific components of the human immune system. Dendritic cells (DCs), as antigen-presenting cells (APCs), play a pivotal role in initiating cellular immunity (Maizels and McSorley, 2016; Lundie et al., 2016; Peng et al., 2022). Generally speaking, upon exposure to helminth-derived products, DCs undergo phenotypic changes that promote the triggering of a Th2-polarized immune response, contributing to immune tolerance; this response is frequently associated with elevated IgE levels, eosinophilia, and mast cell activation (Sher et al., 2003). Previous in vitro studies have shown that co-cultures of DCs with A. pegreffii L3 can impair DC function by reducing their maturation and immunogenic potential (Napoletano et al., 2018). Similarly, antigens from A. simplex (s. s.) larvae have been shown to suppress the expression of costimulatory molecules on DCs while promoting Treg expansion and modulating IL - 10 and IFN-γ production in a host-genotype-dependent manner (Zamora et al., 2019; 2021). However, the reciprocal interaction between the parasite and DCs remains poorly understood—both in terms of how DCs are modulated by the parasite and how the parasite’s gene expression responds to this key immune cell type during their interaction.
Therefore, this study aims to characterize the transcriptomic response of A. pegreffii L3 larvae during in vitro exposure to human DCs, in order to better understand early parasites’ molecular interactions with components of the human immune system.
Materials and methods
Anisakis L3 sampling
Anisakis L3 larvae were carefully removed from the body cavity of European hake (Merluccius merluccius) specimens, caught approximately 12h before, from the Adriatic Sea (off San Benedetto del Tronto coast)–a fishing area with a known high prevalence of Anisakis infection (Cipriani et al., 2018). To control for variability and minimize possible host-related effects, larvae were collected from three different M. merluccius individuals. After their removal, L3 were checked for their integrity and viability under a dissecting microscope. A viable larva in this context was considered physically intact and motile, as measured in terms of its ability to move spontaneously or by stimulation with tweezers and a needle (EFSA, 2024). Alive and not disrupted larvae were washed in a sterile 1X phosphate-buffered saline solution (PBS, Sigma, St Louis, MO) several times and then treated for 1 min with 4% acetic acid (Carlo Erba, Cornaredo, Italy) to inhibit bacterial contamination.
Molecular identification of Anisakis spp.
Anisakis L3 were identified by using a multilocus molecular approach. The mitochondrial cytochrome c oxidase 2 (mtDNA cox2) gene locus was amplified using the primers 211F (forward; 5′-TTTTCTAGTTATATAGATTGRTTYAT-3′) and 210R (reverse; 5′- CACCAACTCTTAAAATTATC - 3′) (Valentini et al., 2006; Mattiucci et al., 2014). The successful PCR products were purified, and Sanger sequenced through an Automated Capillary Electrophoresis Sequencer 3730 DNA Analyzer (Applied Biosystems), using the BigDye® Terminator v3.1 Cycle Sequencing Kit (Life Technologies). Additionally, a direct genotyping determination of the nuclear metallopeptidase 10 gene locus (nas10 nDNA) was performed by the amplification-refractory mutations system (ARMS) PCR assay at nas10 nDNA by the combined use of OUT-F1 (forward; 5’- TATGGCAAATATTATTATCGTA - 3’), OUT-R1 (reverse; 5’-TATTTCCGACAGCAAACAA-3’), INN-F1 (forward; 5’-GCATTGTACACTTCGTATATT-3’), INN-R1 (reverse; 5’-ATTTCTYCAGCAATCGTAAG-3’), following the procedures reported in Palomba et al., 2020. PCR products were separated by electrophoresis using agarose gel (1.5%) stained with GelRed. The distinct banding patterns were detected using ultraviolet transillumination.
Exposure of A. pegreffii L3 to iDCs
For exposure of A. pegreffii L3 to DCs, immature DCs (iDCs) were generated by immunoselected CD14+ monocytes cells obtained from healthy donors, in presence of GM-CSF and IL - 4 as previously described (Dionisi et al., 2018; Napoletano et al., 2007). Each iDCs culture underwent quality control assessment before being used with A. pegreffii (File S1, Supplementary Figure S1). iDCs-L3 larvae co-cultures were established, as previously described (Napoletano et al., 2018). iDCs and L3 larvae were co-cultured in transwell plate (C24 Transwell plate, Corning Costar): L3 larvae were placed in the upper chambers (7 larvae/mL), while iDCs were seeded in the lower chamber (6x105 cells/mL). In detail, a total of 63 L3 larvae were used for the co-culture condition, distributed across nine upper chambers, with 7 larvae per each chamber. Each upper chamber was paired with a corresponding lower chamber containing iDCs at a density of 6×105 cells/mL. To account for biological variability, every three chambers shared iDCs from the same healthy donor, resulting in three experimental groups: Chambers 1-3: Paired with iDCs from Donor 1; Chambers 4-6: Paired with iDCs from Donor 2; Chambers 7-9: Paired with iDCs from Donor 3. As a control, an additional set of nine chambers was prepared under the same conditions but without iDCs, maintaining 7 L3 larvae per chamber. After 24 h, the surviving Anisakis L3 were collected, washed in PBS (twice), and stored in RNA later at -80°C for DNA and RNA extraction.
Ethics statement
All research was conducted in accordance with relevant guidelines and regulations. The study protocol was approved by the Ethics Committee of University Hospital “Policlinico Umberto I” ‘Sapienza’ University of Rome (Protocol nr. 4212). Peripheral blood mononuclear cells (PBMCs) were isolated by Ficoll-Hypaque gradient (1.077 g/mL; Pharmacia LKB, Uppsala, Sweden) from buffy coats of healthy donors obtained from the Transfusion Center, Policlinico Umberto I. Formal written informed consent was obtained from all the healthy blood donors, prior to blood sample collection. Research involving human participants was conducted in compliance with the Declaration of Helsinki.
DNA/RNA extraction
Both DNA and RNA were extracted from each Anisakis larva using TRIzol reagent (Invitrogen, Carlsbad, CA, USA), as previously described (Palomba et al., 2019). Specifically, one larva was collected from each chamber for extraction. This resulted in 9 larvae from the co-culture condition with iDCs (3 larvae for each donor, from different chambers), 9 larvae from the control condition without iDCs (1 larva for each chamber). DNA obtained from each larva was used for species-level identification, while RNA was treated with DNase (DNase I, Invitrogen) to remove any genomic DNA contamination. Then, treated RNA from three larvae (each obtained from a separate chamber with iDCs from different donators) was pooled together. Pooling of larvae was performed to reduce the impact of individual-level transcriptional noise and to obtain a representative average expression profile of A. pegreffii larvae under each experimental condition. This process was repeated three times (3L3/pool, 3 pools in total) to provide triplicate samples for RNA sequencing. RNA concentration, purity and integrity were verified and measured on agarose gel (2%) and by a Bioanalyzer 2100 (Agilent Technologies, Waldbronn, Germany).
Library preparation and RNA sequencing
Following the manufacturer’s protocol, the cDNA library was prepared using a TruSeq Stranded mRNA kit (Illumina, San Diego, USA). In brief, polyadenylated (PolyA+) RNA was purified from 10 μg of total RNA of A. pegreffii L3 using Sera-Mag oligo (dT) beads, fragmented to a length of 100 - 500 nucleotides and reverse transcribed to cDNA using random hexamers. The size-fractionated cDNA was end-repaired and adaptor-ligated according to the manufacturer’s protocol (Illumina). Ligated products of 200 bp were excised from agarose gels and PCR amplified. Products were cleaned using a MinElute PCR purification kit (Qiagen Hilden, Germany) and single-end sequenced on an Illumina HiSeq 2000, according to the manufacturer’s protocol.
Transcript quantification
The raw sequences obtained were processed using Salmon software (v. 1.5.1) (Patro et al., 2017) for mapping against the A. pegreffii de novo reference transcriptome, available online at figshare (https://figshare.com/articles/online_resource/AP_-_Unigenes/18301772) (Palomba et al., 2022). This process was conducted in two distinct phases: 1) the index command of Salmon was used to generate an index of the reference transcriptome; 2) the quant command was used to quantify the abundance of transcripts in the analyzed samples. Both steps were carried out using the software’s default parameters. The transcript quantification phase was performed on a High-Performance Computing Cluster (Castrignanò et al., 2020; Flati et al., 2020). We applied the TransDecoder tool to the Corset-assembled transcriptome, to predict Open Reading Frames, generating peptide (AnisakisPegr:Longest_ORFs_PEP.fasta) and coding sequence (AnisakisPegr:Longest_ORFs_CDS.fasta) files.
Differential gene expression analysis
Differentially expressed genes (DEGs) were identified using IGUANER (Pinna et al., 2024), a software based on DESeq2algorithm (Love et al., 2014). DEGs were defined based on adjusted p-values (padj< 0.05) and log2 fold change thresholds. For the functional annotation of DEGs identified with IGUANER, we adopted two complementary approaches. Initially, we used DIAMOND software (v. 2.0.11) (Buchfink et al., 2014) to conduct sequence comparisons against the Nr, SwissProt, and TrEMBL databases. This step allowed us to obtain highly accurate and relevant homology-based annotations for each analyzed sequence. As second approach, we used Eggnog Mapper (v. 2) (Cantalapiedra et al., 2021) to perform comparisons of sequences with a broad range of homologous groups catalogued in the EggNOG database. This process enabled the acquisition of detailed functional annotations on the basis of the information contained in Gene Ontology (GO) database. A heatmap was generated to visualize the expression patterns of genes based on GO enrichment analysis. Briefly, DEGs from the GO enrichment analysis were filtered to remove duplicates, categorized into up- and down-regulated genes, and merged into a single dataset. The expression data were extracted from the annotated transcriptome file, filtered for significant DEGs (padj< 0.05, |log2FC| > 1), and visualized after eliminating redundant entries.
The complete bioinformatics pipeline is reported in Supplementary Figure S2. All results obtained from the bioinformatics analyses were deposited on Figshare (Table 1). From the complete list of DEGs (padj< 0.05, |log2FC| > 1), genes were selected based on their upregulation and relevance to detoxificant/antioxidant processes as well as chaperone and allergenic properties. This selection was performed manually, guided by existing literature and known gene functions.

Table 1. Overview of produced data files and their access on Figshare.
Quantitative real-time PCR validation
To validate the results of RNA-seq, six differentially expressed genes were randomly selected for quantitative real-time PCR (RT-qPCR). The RNA samples used for the RT-qPCR assay were the same as those used for RNA-seq. The cDNA was synthesized for the RT-qPCR using the high-capacity cDNA Reverse Transcription Kit (Thermo Fisher Scientific, Wilmington, DE, USA) following the manufacturer’s instructions. The RT-qPCR was performed using SYBR Green PCR Master Mix, according to the manufacturer’s instructions. Amplifications were conducted for p-glycoprotein 2, tetraspanin, carboxypeptidase, ras-related protein Rab, galectin, aspartic protease 6, and elongation factor (EF) as reference gene. Specific primer pairs were designed (Supplementary Table S1), and standard curves were generated. All reactions were performed in triplicate in the StepOnePlus Real-Time PCR Detection System (Applied Biosystems) and relative quantification was carried out with the ΔΔCT method (Livak and Schmittgen, 2001) using the abundance of EF mRNA as endogenous housekeeping control. The relative transcription levels as obtained by RT-qPCR analyses were compared with abundance levels detected by RNA-seq. Values from replicate experiments were averaged. Finally, to obtain values suitable for statistical comparisons, the fold change (FC) value was calculated. These values (plotted after conversion in log2 numbers) were used to evaluate the correlation between RNA-seq and RT-qPCR methods, applying statistical evaluation using the Pearson test (in Prism GraphPad software).
Results
Molecular identification of A. pegreffii L3
A multilocus genotyping approach, combining mitochondrial and nuclear markers, was applied to ensure species identification (Mattiucci et al., 2018, 2025). BLAST analysis of the 21 sequences obtained from the Anisakis L3 at the mtDNA cox2 gene locus (~600 bp) retrieved a percentage of identity of 99 - 100% with the sequences of A. pegreffii previously deposited (accession numbers, KY565564-KY565562). Additionally, ARMS-PCR analysis targeting the nas10 locus generated a species-specific 117 bp band, diagnostic of the C-allele, further confirming the identification of A. pegreffii (Palomba et al., 2020).
Sequencing statistics
The quality of extracted RNA from L3 reached the standard of sequencing with RIN values >7, and the concentration of each sample was higher than 50 ng/μL. The total number of raw reads obtained by sequencing was 154047207, while the total number of clean reads after filtration was 153572782 (Table 2). The total number of clean read bases was 9580666822. Q30 was greater than 97.13%, and GC content was 43.02-46.81% (Table 2).
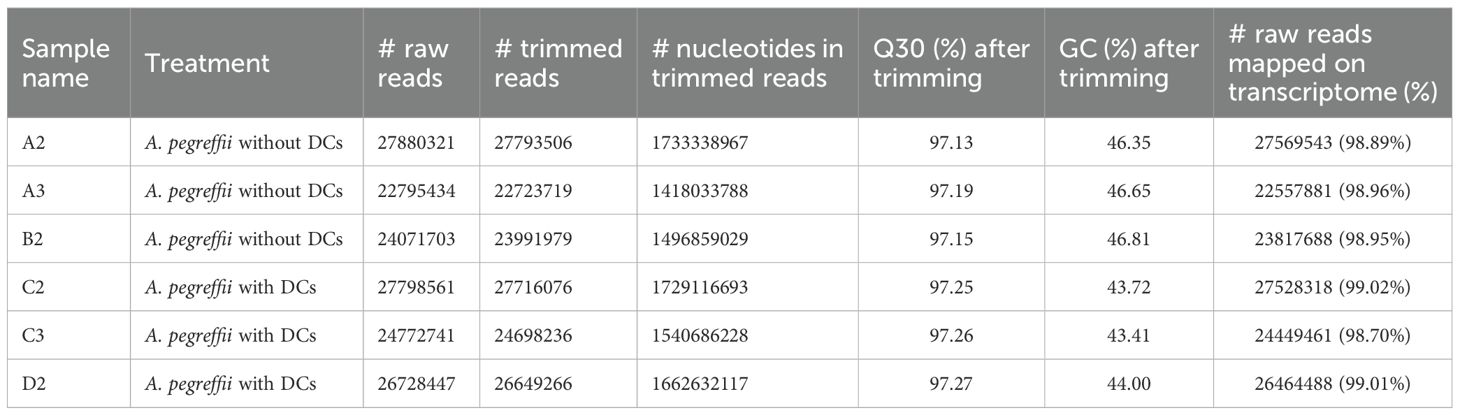
Table 2. Summary of data output quality of various libraries.
The raw data of all samples reported in this study were deposited in the NCBI (National Center for Biotechnology Information) under the accession number PRJNA752284.
Differential gene expression in A. pegreffii following iDC exposure
A total of 3914 differentially expressed genes (DEGs) with|log2FC| > 1 and p.adj< 0.05 were detected in the A. pegreffii L3 larvae treatment group (L3 co-cultured with iDCs) compared to the control group (L3 alone) (Supplementary Table S2). Results from DEG counts are reported in Table 3. Among DEGs, 1868 genes were up-regulated in L3 plus iDC compared to L3 alone, while 2046 genes were down-regulated (Figure 1). The most significantly modulated genes, characterized by the highest log2 fold change values, are highlighted in the plot (Figure 1). Notably, these included: glutathione peroxidase, SH3 domain-containing protein, calponin-like protein, and collagen cuticle N domain-containing protein, which exhibit strong differential expression, i.e., 22.00, 22.04, 28.55, 27.39 log2 fold change, respectively.
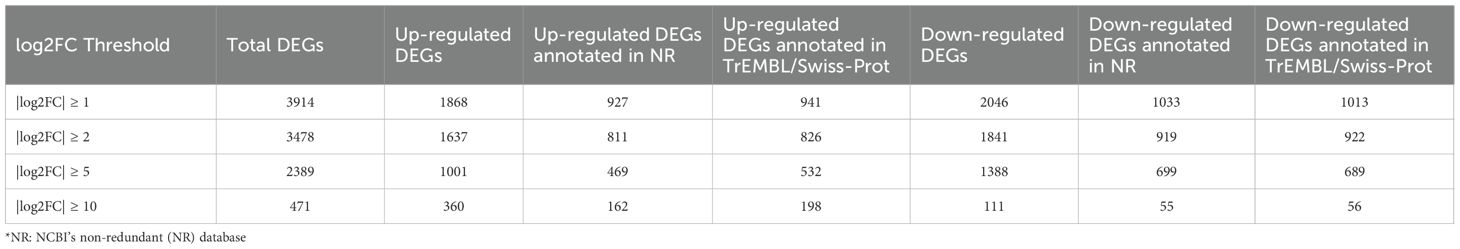
Table 3. Overview of differentially expressed gene counts across log2FC thresholds, padj ≤ 0.05.
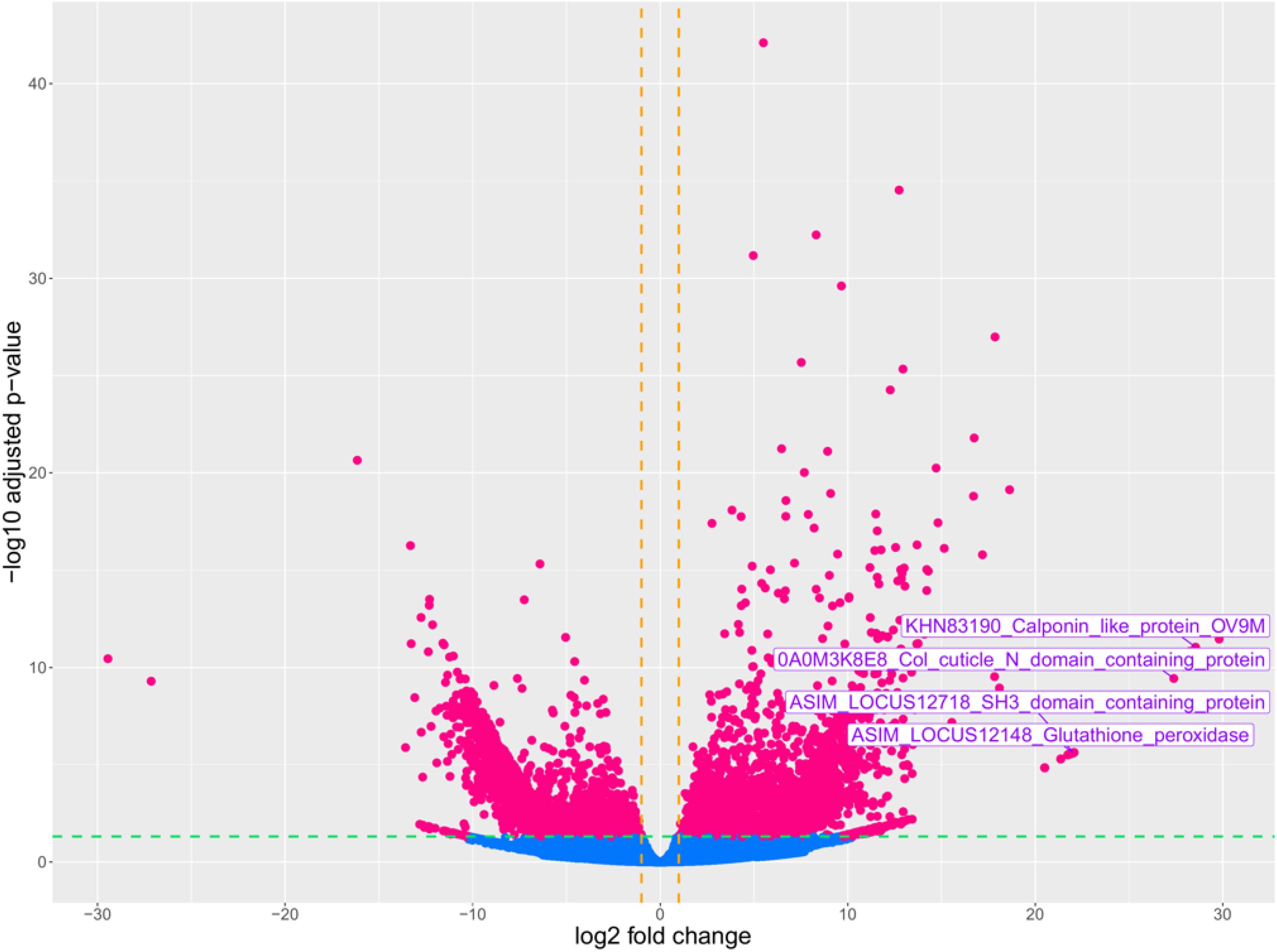
Figure 1. Volcano plot of DEGs between the treatment group (A. pegreffii L3 in the presence of iDCs) and control group (A. pegreffii in the absence of iDCs). The plot represents gene expression levels, with log2 Fold Change displayed on the x-axis and log10 (p.adjust) on the y-axis. The vertical and horizontal dashed lines indicate the fold-change cut-off = ± 1 and the p-value = 0.05, respectively. Pink dots indicate up and down-regulated genes, and light blue dots represent genes without significant changes in expression.
Functional enrichment and biological relevance of DEGs
To better understand the functional implications of the transcriptional changes, we conducted GO enrichment analysis on up- and down-regulated DEGs. GO enrichment results revealed significantly enriched categories (p.adj< 0.05) (Figure 2, Supplementary Table S3). Up-regulated genes were mostly involved in collagen and cuticulin-based cuticle development (BP), dipeptidyl-peptidase activity (MF), and anchored component of membrane (CC) (Figure 2A, Supplementary Table S3). Conversely, down-regulated genes were enriched in actin filament bundle organization (BP), nuclear receptor activity in (MF), and stereocilium bundle (CC) (Figure 2B, Supplementary Table S3). The heatmap (Figure 3) visualizes the expression patterns of the GO-annotated genes identified in the enrichment analysis. In addition, we specifically examined the list of DEGs (Supplementary Table S2) to identify genes of relevance. Further analysis identified the upregulation of genes associated with detoxification and antioxidant processes (e.g., glutathione peroxidase, thioredoxin reductase 1, peroxidase mlt-7, and superoxide dismutase), allergenic responses (e.g., heat shock protein 70, anis12, anis11, and anis10), and chaperone-related functions (e.g., calponin, SH3 domain-containing protein, HSP70, and HSP60) (Figure 4). Comparison of the DEGs identified with those previously reported in the extracellular vesicles (EVs) of A. pegreffii (Palomba et al., 2022) revealed 21 overlapping genes (Supplementary Table S4).
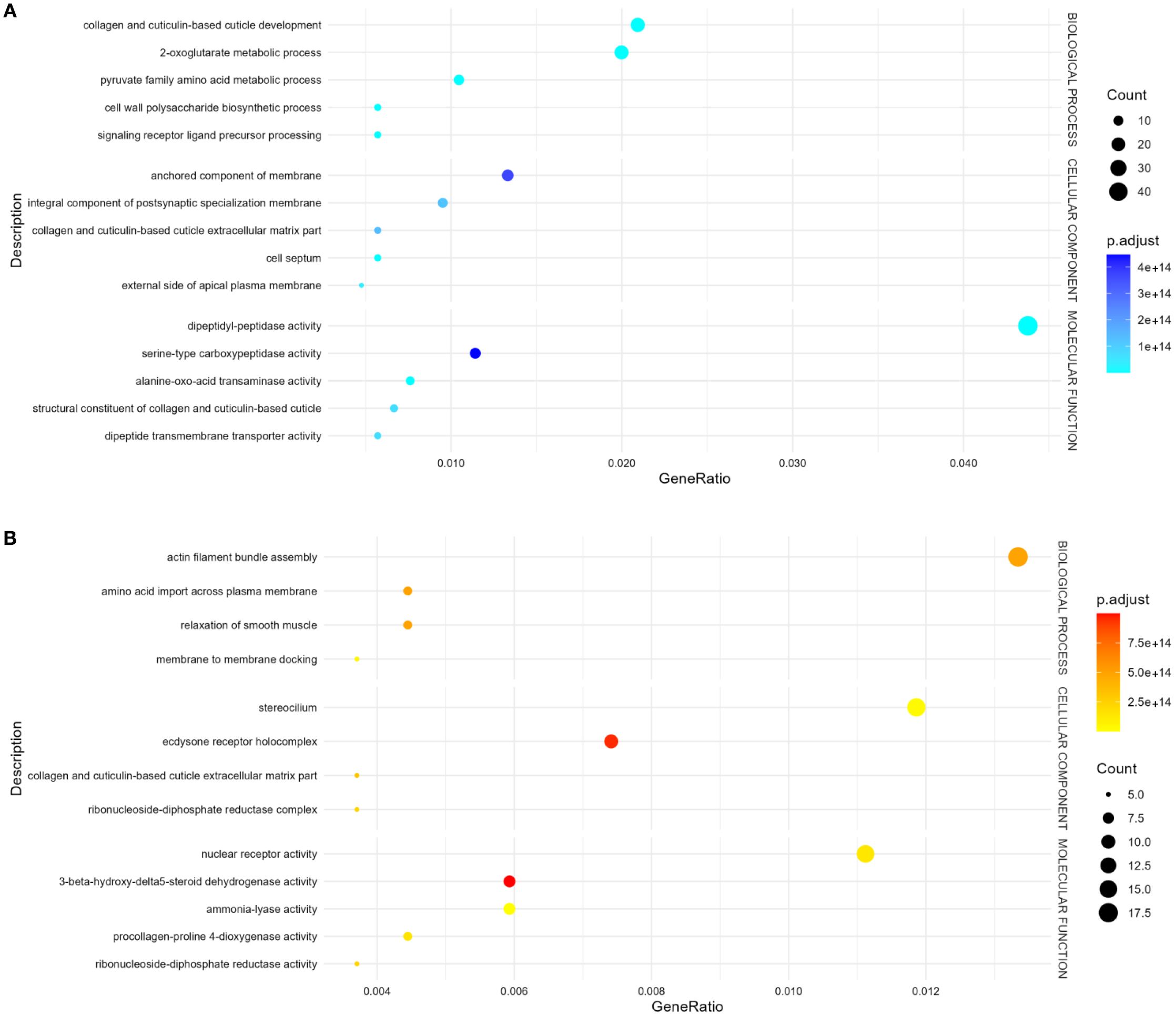
Figure 2. Bubble plot of GO terms enrichment analysis of DEGs. “Count” indicates the number of DEGs enriched in the pathway; “GeneRatio” indicates the ratio of enriched DEGs to background genes; “p.adj”, shown by color, indicates a significant level of enrichment results. (A) Up-regulated genes, (B) Down-regulated genes.
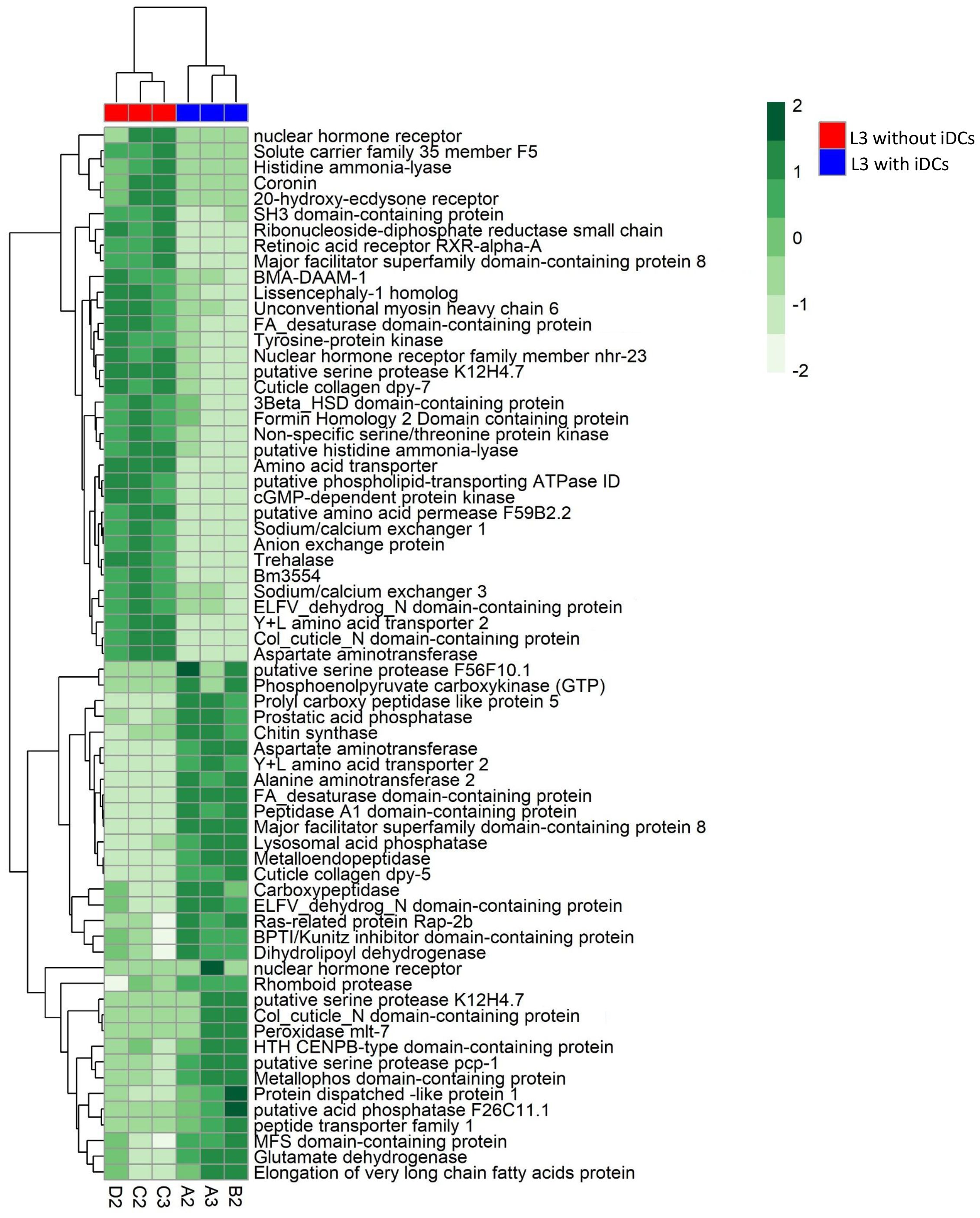
Figure 3. Heat map showing the log2 fold change of genes enriched in GO ontology categories in A. pegreffii L3.
![The image displays three categories of genes: Detoxificant/Antioxidant, Chaperone, and Allergenic genes. Each category lists specific genes with corresponding numerical values shown in color-coded boxes. Detoxificant/Antioxidant genes include Glutathione peroxidase (22), Peroxidase mlt-7 (13.44), Thioredoxin reductase 1 (11.77), and Superoxide dismutase [Cu-Zn] (11.55). Chaperone genes feature Calponin-like protein OV9M (28.55), SH3 domain-containing protein (22.04), Heat shock 70 kDa protein (11.70), and Chaperonin-like protein Hsp-60 (1.83). Allergenic genes display Heat shock 70 kDa (11.77), Ani s 12 allergen (6.80), Ani s 11 allergen (3.49), and Ani s 10 allergen (1.44).](https://www.frontiersin.org/files/Articles/1646537/fcimb-15-1646537-HTML/image_m/fcimb-15-1646537-g004.jpg)
Figure 4. Heat map showing the log2 fold change of upregulated genes in A. pegreffii L3 within selected specific categories.
Correlation between RNA-seq and RT-qPCR results
The abundance of transcripts emerging from transcriptomic analyses was confirmed by RT-qPCR by examining six transcripts differentially modulated in A. pegreffii L3 in the presence/absence of iDCs i.e. p-glycoprotein 2, tetraspanin, carboxypeptidase, ras-related protein Rab, galectin, and aspartic protease 6. The validation analysis was performed using triplicate RNA samples. Correlation analysis indicated significant linear relationships (R2 = 0.70; p< 0.03) between RNA-seq and RT-qPCR results (Table 4, Supplementary Figure S3).
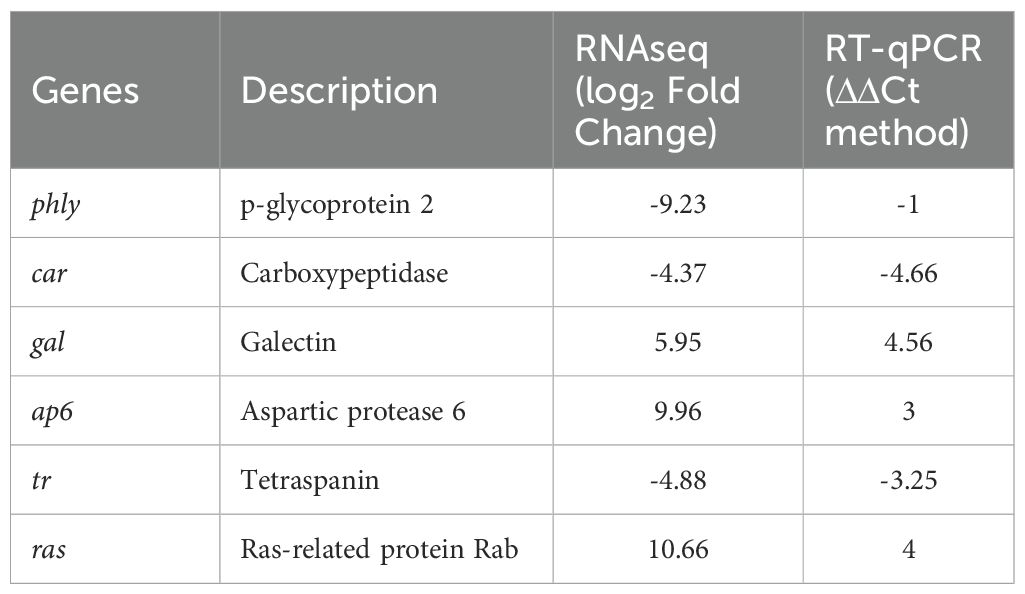
Table 4. Gene expression levels obtained by RNAseq and RT-qPCR.
Discussion
Throughout their life cycle, helminth parasites are exposed to diverse microhabitat conditions within their hosts, including fluctuations in temperature, pH, nutrient availability, and host immune pressures. Among these, the interaction with the host immune system is particularly critical for parasite survival, especially in accidental hosts, such as humans, which represent an evolutionary dead-end and an unsuitable ecological niche for A. pegreffii (Trumbić et al., 2021), placing the parasite at a disadvantageous position. In this case, the host immune system rapidly recognizes the parasite as a threat, activating local effector mechanisms that may lead to sustained inflammation (Nieuwenhuizen, 2016). The transcriptomic analysis, here described, revealed that the A. pegreffii larvae respond to the presence of iDCs primarily through an initial transcriptional pattern, likely reflecting an early attempt to cope with human derived immune-mediated stress. Notably, A. pegreffii is able to regulate a complex network of metabolic pathways, strategically investing energy in activating and upregulating genes associated with energy production (i.e., cGMP-dependent protein kinase, sodium/calcium exchangers, trehalase), as well as nitrogen and amino acid metabolism (e.g., aspartate aminotransferase, histidine ammonia-lyase, amino acid transporters). This metabolic shift likely provides the necessary resources to sustain vital functions under host-induced stress (Stryiński et al., 2024). However, this response is energetically costly and may not be sustainable in the long term. The sustained metabolic demand may deplete internal energy reserves leading to the accumulation of toxic metabolic waste products, and ultimately resulting in the death of the invading Anisakis larva.
Interestingly, this upregulation of metabolic genes appears closely linked to the enhanced expression of genes involved in cuticle maintenance. The synthesis of its components is an energy-intensive process that requires a steady supply of amino acids and efficient protein biosynthesis mechanisms (Johnstone, 1994). The modulation of structural genes (e.g., cuticle collagen dpy-7, forming homology 2, myosin heavy chain 6) is crucial for cuticle formation (Johnstone, 2000). This serves multiple functions, including acting as a protective barrier between the nematode and its environment, maintaining its body shape (Johnstone et al., 1992), and facilitating movement through connections with muscle tissue (Francis and Waterston, 1991; Hresko et al., 1999). These processes are pivotal for nematode survival, enabling interactions with hostile environments, when trying to penetrate the gastric and intestinal submucosa (Johnstone, 1994; Page et al., 2014; Sandhu et al., 2021). Over time, the expression of genes encoding these structural proteins has likely evolved to support both developmental processes and defense mechanisms (Kim et al., 2018). For instance, Kim et al. (2018) reported that collagen-related genes were abundantly transcribed in the Anisakis L3, reflecting the active molting process. Similarly, Trumbić et al. (2021) observed that the genes involved in molting, such as structural constituents of the cuticle, collagen, and cuticulin, were upregulated in A. pegreffii L3 infecting rats, suggesting that molting-related gene expression can be influenced by host-specific factors. In this context, in the presence of iDCs, A. pegreffii L3 stage larvae may perceive them as potential threats, triggering a defense response characterized by the coordinated activation of metabolic, antioxidant, and structural pathways. This upregulation may represent a protective strategy aimed at shielding the nematode from host immune effectors, potentially enhancing its survival in an accidental host. However, the direct involvement of structural gene modulation —particularly collagen and cuticulin genes— remains speculative. Further functional validation is required to confirm their role in human host–parasite interactions.
Interestingly, alongside the upregulation of structural genes involved in cuticle formation, we observed the downregulation of genes associated with procollagen-proline 4-dioxygenase activity, critical for the hydroxylation of proline residues during collagen maturation (Page and Johnstone, 2007). The simultaneous upregulation of cuticle-related genes and downregulation of procollagen-proline 4-dioxygenase activity might suggest a complex regulatory mechanism in which A. pegreffii actively synthesizes structural precursors but limits their full post-translational maturation. However, this interpretation remains speculative, and further molecular and functional analyses are required to confirm the regulatory dynamics involved.
Moreover, the downregulation of genes involved in cytoskeletal organization (e.g., actin filament, bundle assembly) and membrane dynamics (e.g., membrane-to-membrane docking) suggests a strategic suppression of processes related to cellular motility and intracellular trafficking. This downregulation may reflect an attempt to conserve energy by limiting metabolically expensive activities that are not immediately critical for the parasite development.
Alongside this metabolic shift, genes involved in oxidative stress responses and those linked to ion homeostasis (sodium/calcium exchangers, anion exchange proteins), may play a role in modulating the host immune response (Nawaratna et al., 2018). This mechanism could contribute to mitigating the effects of reactive oxygen species (ROS) generated during the immune response, potentially enhancing parasite survival within the host. This suggests that A. pegreffii not only adjusts its metabolic activity, but also engages calcium-dependent pathways and antioxidant-related mechanisms to counteract host-induced oxidative stress. The modulation of antioxidant enzymes is a key strategy for parasites to defend themselves against the host immune system, specifically targeting ROS generated by immune cells such as macrophages, neutrophils and eosinophils. Indeed, our previous studies have shown that iDCs strongly increase ROS production as part of their immune defense against A. pegreffii (Napoletano et al., 2018). In addition, the increased transcription of genes implicated in the detoxification of ROS and bolstering antioxidant defense, the upregulation of glutathione peroxidase, thioredoxin reductase 1, peroxidase mlt-7 and superoxide dismutase suggest a metabolic pathway of A. pegreffii to shield itself from ROS produced by host tissue-resident cells, besides iDCs. For example, glutathione peroxidase plays a critical role in removing harmful ROS by catalyzing the reduction of hydrogen peroxide and organic hydroperoxides in A. pegreffii. Instead, superoxidase dismutase has been shown to be exploited in resistance to radiation (Seo et al., 2006) and xenobiotics (Stryiński et al., 2022, 2024), suggesting a versatile role of antioxidant enzymes in the parasite’s defense strategy against various environmental stressors. In the case of thioredoxin reductase, recent studies in filariasis have shown that inhibiting this enzyme induces apoptotic death of the parasite (Sen et al., 2023), highlighting its crucial role in the parasite’s survival and, consequently, in the disease pathogenesis and progression. Additionally, it is interesting to note that the thioredoxin reductase exerts an immunosuppressive role by downregulating the “inflammasome” pathways in macrophages exposed to the parasite (Joardar et al., 2021).
In addition to antioxidant enzymes, the modulation of molecular chaperones represents another critical mechanism employed by A. pegreffii to manage iDC-induced stress. Among these, HSP70 plays a key role in stabilizing proteins during stress (Hartl, 1996). Beyond its role in protein maintenance, HSP70 is involved in differentiation, protection against host-induced damage (including oxidative stress from free radicals), and promoting virulence (Polla, 1991; Cruz-Laufer et al., 2025). These functions allow parasites to adapt to various host niches, endure immune responses, and respond to environmental changes. In this context, HSP70 also emerges as a potential virulence factor of A. pegreffii. The up-regulation of the Hsp70 gene in A. pegreffii co-cultured with iDCs can be understood as an adaptation to changing environments and host-parasite interactions over time. Additionally, it is noteworthy that HSP70 was detected as being carried by EVs released by L3 larvae, along with other proteins involved in metabolism and immune modulation (Palomba et al., 2023). Other specific genes were also found in EVs of A. pegreffii to be differentially regulated following exposure to iDCs (Supplementary Table S4). Beyond their role in stress responses and parasite adaptation, HSP70s are also recognized allergens in various arthropods, including mites, midges, flies, and cockroaches. Similarly, HSPs (including HSP β-1, HSP - 12.2, HSP70, HSP 75KDa, and HSP 90-α), along with superoxide dismutase and galectin, have been identified in the larval extract of the ascarid Toxocara canis (de Silva et al., 2018). In the larva migrans of T. canis, HSPs serve as key immunomodulatory molecules, playing essential roles in the parasite survival, even during accidental human infection (Pérez-Morales and Espinoza, 2015).
Finally, Anis10, Anis11, and Anis12—previously described as allergens—were detected among the differentially expressed genes (Audicana and Kennedy, 2008; Mattiucci et al., 2017). However, their specific roles remain unclear, although they are likely closely interconnected with the host’s immune response, and future investigations are essential to elucidate the precise functions of these genes and their coded proteins (Castrignanò et al., 2007).
Conclusions
This study provides the first comprehensive transcriptomic overview of A. pegreffii L3 larvae during in vitro exposure to human iDCs, revealing for the first time a coordinated activation of antioxidant enzymes, molecular chaperones, and structural genes in response to early immune recognition. The responses triggered by the parasite likely reflect a short-term survival strategy against immune-mediated stress in a non-permissive host environment. Notably, the upregulation of detoxification enzymes and molecular chaperones, along with the modulation of cuticle-related genes, highlights the parasite’s attempt to mitigate host-induced oxidative stress and maintain structural integrity. However, the concurrent downregulation of genes necessary for collagen maturation and cellular motility suggests that while A. pegreffii initiates defense mechanisms, it may be unable to sustain the energetic cost of adaptation in the human host.
Overall, the molecular characterization of the A. pegreffii L3 stage larvae, interacting with iDCs here described, appear to be consistent with the parasite’s inability to develop. At the same time, our data show that the larva activates a transcriptional response likely aimed at counteracting the immune pressure exerted by iDCs — an early, but ultimately unsuccessful, attempt to ensure survival.
While previous work by Napoletano et al. (2018) has shown that A. pegreffii L3 can impair DC function by inducing apoptosis and blocking their differentiation, maturation, and migration to lymph nodes, our study focuses on the parasite’s point of view — specifically, how the larva itself responds to contact with human iDCs at the transcriptomic level. The expression patterns observed suggest that the parasite attempts to resist immune stress by activating genes involved in energy metabolism, oxidative stress response, and structural maintenance. This may allow temporary adaptation to the host environment and could contribute indirectly to shaping the local immune response. In particular, the modulation of DC activity by the parasite — as reported in our previous studies (Napoletano et al., 2018) — may influence Th1/Th2 polarization, potentially favoring a shift toward a Th2-biased environment, typical of chronic helminth infections (Sher et al., 2003).
Further studies are needed to explore whether the upregulated genes identified in this analysis influence the host immune response. For instance, specific antigens derived from these genes could be tested in vitro to assess their immunomodulatory potential, as already done in other parasite models, such as Trichinella spiralis (Yu et al., 2025). In this context, human organoid systems represent a promising experimental platform to investigate the host–parasite interaction in a physiologically relevant setting. These 3D models can mimic the structure and function of the human gastrointestinal epithelium and allow co-culture with immune cells (Perez et al., 2025). They could be used to assess barrier integrity, local cytokine responses, and immune cell activation, as well as to explore whether A. pegreffii antigens contribute to chronic inflammation or tumor-promoting conditions, particularly in the context of prolonged or repeated exposure (Bellini et al., 2024).
Data availability statement
The datasets generated for this study can be found in the NCBI (National Center for Biotechnology Information) under the accession number PRJNA752284 and in Figshare (see Table 3).
Ethics statement
The studies involving humans were approved by Ethics Committee of University Hospital “Policlinico Umberto I” ‘Sapienza’ University of Rome (Protocol nr. 4212). The studies were conducted in accordance with the local legislation and institutional requirements. The participants provided their written informed consent to participate in this study.
Author contributions
MP: Writing - review & editing, Conceptualization, Methodology, Writing - original draft, Investigation, Resources. AR: Writing - original draft, Resources, Methodology, Writing - review & editing, Investigation, Conceptualization. TC: Investigation, Writing - original draft, Software, Validation, Supervision, Methodology, Writing - review & editing. CN: Writing - review & editing, Writing - original draft, Methodology. XR-G: Writing - review & editing, Writing - original draft, Methodology. VP: Validation, Writing - original draft, Software, Writing - review & editing. FL: Writing - original draft, Software, Writing - review & editing, Validation. DC: Writing - review & editing, Writing - original draft. SM: Writing - original draft, Conceptualization, Funding acquisition, Resources, Project administration, Methodology, Writing - review & editing, Supervision, Investigation.
Funding
The author(s) declare financial support was received for the research and/or publication of this article. This study was supported by the Italian Ministry of Health (RF) 2018 - 12367986, title “Innovative approaches and parameters in the diagnosis and epidemiological surveillance of the Anisakisrelated human diseases in Italy”.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The author(s) declared that they were an editorial board member of Frontiers, at the time of submission. This had no impact on the peer review process and the final decision.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fcimb.2025.1646537/full#supplementary-material
References
Audicana, M. T. and Kennedy, M. W. (2008). Anisakis simplex: from obscure infectious worm to inducer of immune hypersensitivity. Clin. Microbiol. Rev. 21, 360–379. doi: 10.1128/CMR.00012-07
Bellini, I., Scribano, D., Ambrosi, C., Chiovoloni, C., Rodon, S., Pronio, A., et al. (2025). Anisakis extracellular vesicles elicit immunomodulatory and potentially tumorigenic outcomes on human intestinal organoids. Parasitol. Vectors. 17, 393. doi: 10.1186/s13071-024-06471-7
Brusca, I., Barrale, M., Zarcone, M., Fruscione, S., Onida, R., De Bella, D. D., et al. (2023). Basophil activation test in the diagnosis of Anisakis allergy: An observational study from an area of high seafood consumption in Italy. Pathogens 12, 777. doi: 10.3390/pathogens12060777
Buchfink, B., Xie, C., and Huson, D. H. (2014). Fast and sensitive protein alignment using DIAMOND. Nat. Methods 12, 59–60. doi: 10.1038/nmeth.3176
Cantalapiedra, C. P., Hernandez-Plaza, A., Letunic, I., Bork, P., and Huerta-Cepas, J. (2021). eggNOG-mapper v2: Functional annotation, orthology assignments, and domain prediction at the metagenomic scale. Mol. Biol. Evol. 38, 5825–5829. doi: 10.1093/molbev/msab293
Castrignanò, T., De Meo, P. D. O., Carrabino, D., Orsini, M., Floris, M., and Tramontano, A. (2007). The MEPS server for identifying protein conformational epitopes. BMC Bioinform. 8, S6. doi: 10.1186/1471-2105-8-S1-S6
Castrignanò, T., Gioiosa, S., Flati, T., Cestari, M., Picardi, E., Chiara, M., et al. (2020). ELIXIR-IT HPC @ CINECA: High performance computing resources for the bioinformatics community. BMC Bioinform. 21, 352. doi: 10.1186/s12859-020-03565-8
Cavallero, S., Bellini, I., Pizzarelli, A., Arca, B., and D’Amelio, S. (2022). A miRNAs catalogue from third-stage larvae and extracellular vesicles of Anisakis pegreffii provides new clues for host-parasite interplay. Sci. Rep. 12, 9667. doi: 10.1038/s41598-022-13594-3
Cipriani, P., Sbaraglia, G. L., Paoletti, M., Giulietti, L., Bellisario, B., Palomba, M., et al. (2018). The Mediterranean European hake, Merluccius merluccius: Detecting drivers influencing the Anisakis spp. larvae distribution. Fish. Res. 202, 79–89. doi: 10.1016/j.fishres.2017.07.010
Cruz-Laufer, A. J., Vanhove, M. P. M., Bachmann, L., Barson, M., Bassirou, H., Bitja Nyom, A. R., et al. (2025). Adaptive evolution of stress response genes in parasites aligns with host niche diversity. BMC Biol. 23, 10. doi: 10.1186/s12915-024-02091-w
da Silva, M. B., Oviedo, Y., Cooper, P. J., Pacheco, L. G., Pinheiro, C. S., Ferreira, F., et al. (2018). The somatic proteins of Toxocara canis larvae and excretory-secretory products revealed by proteomics. Vet. Parasitol. 259, 25–34. doi: 10.1016/j.vetpar.2018.06.015
Dionisi, M., De Archangelis, C., Battisti, F., Rahimi Koshkaki, H., Belleudi, F., Zizzari, I. G., et al. (2018). Tumor-derived microvesicles enhance cross-processing ability of clinical grade dendritic cells. Front. Immunol. 9. doi: 10.3389/fimmu.2018.02481
EFSA Panel on Biological Hazards, Koutsoumanis, K., Allende, A., Alvarez-ordóñez, A., Bover-cid, S., Chemaly, M., et al. (2024). Re-evaluation of certain aspects of the EFSA Scientific Opinion of April 2010 on risk assessment of parasites in fishery products, based on new scientific data. EFSA J22, e9090. doi: 10.2903/j.efsa.2024.9090
Flati, T., Gioiosa, S., Spallanzani, N., Tagliaferri, I., Diroma, M. A., Pesole, G., et al. (2020). HPC-REDItools: A novel HPC-aware tool for improved large-scale RNA-editing analysis. BMC Bioinform. 21, 353. doi: 10.1186/s12859-020-03562-x
Francis, R. and Waterston, R. H. (1991). Muscle cell attachment in Caenorhabditis elegans. J. Cell. Biol. 14, 465–479. doi: 10.1083/jcb.114.3.465
Guardone, L., Armani, A., Nucera, D., Costanzo, F., Mattiucci, S., and Bruschi, F. (2018). Human anisakiasis in Italy: a retrospective epidemiological study over two decades. Parasite 25, 41. doi: 10.1051/parasite/2018034
Hartl, F. U. (1996). Molecular chaperones in cellular protein folding. Nature 381, 571–579. doi: 10.1038/381571a0
Hresko, M. C., Schriefer, L. A., Shrimankar, P., and Waterston, R. H. (1999). Myotactin, a novel hypodermal protein involved in muscle-cell adhesion in Caenorhabditis elegans. J. Cell. Biol. 146, 659–672. doi: 10.1083/jcb.146.3.659
Joardar, N., Bhattacharya, R., Halder, S., Sen, A., Biswas, S. R., Jana, K., et al. (2021). Filarial thioredoxin reductase exerts anti-inflammatory effects upon lipopolysaccharide induced inflammation in macrophages. Int. J. Biol. Macromol. 193, 1379–1390. doi: 10.1016/j.ijbiomac.2021.10.200
Johnstone, I. L. (1994). The cuticle of the nematode Caenorhabditis elegans: a complex collagen structure. Bioessays 16, 171–178. doi: 10.1002/bies.950160307
Johnstone, I. L. (2000). Cuticle collagen genes expression in Caenorhabditis elegans. Trends Genet. 16, 21–27. doi: 10.1016/s0168-9525(99)01857-0
Johnstone, I. L., Shafi, Y., and Barry, J. D. (1992). Molecular analysis of mutations in the Caenorhabditis elegans collagen gene dpy-7. EMBO J. 1, 3857–3863. doi: 10.1002/j.1460-2075.1992.tb05478.x
Kim, J. H., Kim, J. O., Jeon, C. H., Nam, U. H., Subramaniyam, S., Yoo, S. I., et al. (2018). Comparative transcriptome analyses of the third and fourth stage larvae of Anisakis simplex (Nematoda: Anisakidae). Mol. Biochem. Parasitol. 226, 24–33. doi: 10.1016/j.molbiopara.2018.10.002
Livak, K. J. and Schmittgen, T. D. (2001). Analysis of relative gene expression data using real-time quantitative PCR and the 2– ΔΔCT method. methods 25, 402–408. doi: 10.1006/meth.2001.1262
Love, M. I., Huber, W., and Anders, S. (2014). Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 15, 550. doi: 10.1186/s13059-014-0550-8
Lundie, R. J., Webb, L. M., Marley, A. K., Phythian-Adams, A. T., Cook, P. C., Jackson-Jones, L. H., et al. (2016). A central role for hepatic conventional dendritic cells in supporting Th2 responses during helminth infection. Immunol. Cell. Biol. 94, 400–410. doi: 10.1038/icb.2015.114
Maizels, R. M. and McSorley, H. J. (2016). Regulation of the host immune system by helminth parasites. J. Allergy Clin. Immunol. 138, 666–675. doi: 10.1016/j.jaci.2016.07.007
Mattiucci, S., Cipriani, P., Levsen, A., Paoletti, M., and Nascetti, G. (2018). Molecular epidemiology of Anisakis and Anisakiasis: an ecological and evolutionary road map. Adv. Parasitol. 99, 93–263. doi: 10.1016/bs.apar.2017.12.001
Mattiucci, S., Colantoni, A., Crisafi, B., Mori-Ubaldini, F., Caponi, L., Fazii, P., et al. (2017). IgE sensitization to Anisakis pegreffii in Italy: Comparison of two methods for the diagnosis of allergic anisakiasis. Parasite Immunol. 39, 1–10. doi: 10.1111/pim.12440
Mattiucci, S., Fazii, P., De Rosa, A., Paoletti, M., Megna, A. S., Glielmo, A., et al. (2013). Anisakiasis and gastroallergic reactions associated with Anisakis pegreffii infection, Italy. Emerg. Infect. Dis. 19, 496–499. doi: 10.3201/eid1903.121017
Mattiucci, S., Palomba, M., Belli, B., Aco-Alburqueque, R., Cipriani, P., Roca-Gerones, X., et al. (2025). Hybridization and introgression of mitochondrial genome between the two species Anisakis pegreffii and A. simplex (s. s.) using a wide genotyping approach: evolutionary and ecological implications. Parasitology. 152, 293–313 doi: 10.1017/S0031182025000228
Mattiucci, S., Palomba, M., and Nascetti, G. (2022). “Anisakis.” Encyclopedia of infection and immunity. 2, 408–423. doi: 10.1016/B978-0-12-818731-9.00075-6
Mehrdana, F., Kania, P. W., Nazemi, S., and Buchmann, K. (2017). Immunomodulatory effects of excretory/secretory compounds from Contracaecum osculatum larvae in a zebrafish inflammation model. PloS One 12, e0181277. doi: 10.1371/journal.pone.0181277
Napoletano, C., Mattiucci, S., Colantoni, A., Battisti, F., Zizzari, I. G., Rahimi, H., et al. (2018). Anisakis pegreffii impacts differentiation and function of human dendritic cells. Parasite Immunol. 40, e12527. doi: 10.1111/pim.12527
Napoletano, C., Pinto, D., Bellati, F., Taurino, F., Rahimi, H., Tomao, F., et al. (2007). A comparative analysis of serum and serum-free media for generation of clinical grade DCs. J. Immunother. 30, 567–576. doi: 10.1097/CJI.0b013e318046f396
Nawaratna, S. S. K., You, H., Jones, M. K., McManus, D. P., and Gobert, G. N. (2018). Calcium and Ca 2+/Calmodulin-dependent kinase II as targets for helminth parasite control. Biochem. Soc Trans. 46, 1743-1751. doi: 10.1042/BST20180480
Nieuwenhuizen, N. E. (2016). Anisakis–immunology of a foodborne parasitosis. Parasite Immunol. 38, 548–557. doi: 10.1111/pim.12349
Nieuwenhuizen, N. E. and Lopata, A. L. (2013). Anisakis-a food-borne parasite that triggers allergic host defences. Int. J. Parasitol. 43, 1047–1057. doi: 10.1016/j.ijpara.2013.08.001
Page, A. P., Stepek, G., Winter, A. D., and Pertab, D. (2014). Enzymology of the nematode cuticle: a potential drug target? Int. J. Parasitol. Drugs Drug Resist. 4, 133–141. doi: 10.1016/j.ijpddr.2014.05.003
Palomba, M., Libro, P., Di Martino, J., Rughetti, A., Santoro, M., Mattiucci, S., et al. (2022). De novo transcriptome assembly and annotation of the third stage larvae of the zoonotic parasite Anisakis pegreffii. BMC Res. Notes 15, 223. doi: 10.1186/s13104-022-06099-9
Palomba, M., Paoletti, M., Colantoni, A., Rughetti, A., Nascetti, G., and Mattiucci, S. (2019). Gene expression profiles of antigenic proteins of third stage larvae of the zoonotic nematode Anisakis pegreffii in response to temperature conditions. Parasite 26, 52. doi: 10.1051/parasite/2019055
Palomba, M., Paoletti, M., Colantoni, A., Rughetti, A., Nascetti, G., and Mattiucci, S. (2023). Proteomic characterization of extracellular vesicles released by third stage larvae of the zoonotic parasite Anisakis pegreffii (Nematoda: Anisakidae). Front. Cell. Infect. Microbiol. 13. doi: 10.3389/fcimb.2023.1079991
Palomba, M., Paoletti, M., Webb, S. C., Nascetti, G., and Mattiucci, S. (2020). A novel nuclear marker and development of an ARMS-PCR assay targeting the metallopeptidase 10 (nas 10) locus to identify the species of the Anisakis simplex (s. l.) complex (Nematoda, Anisakidae). Parasite 27, 39. doi: 10.1051/parasite/2020033
Patro, R., Duggal, G., Love, M. I., Irizarry, R. A., and Kingsford, C. (2017). Salmon provides fast and bias-aware quantification of transcript expression. Nat. Methods 14, 417–419. doi: 10.1038/nmeth.4197
Peng, J., Federman, H. G., Hernandez, C. M., and Siracusa, M. C. (2022). Communication is key: Innate immune cells regulate host protection to helminths. Front. Immunol. 13. doi: 10.3389/fimmu.2022.995432
Perez, M. G., Coman, D., Neves, J. F., and Britton, C. (2025). Organoids in parasitology: a game-changer for studying host-nematode interactions. Parasitology. 1–44 doi: 10.1017/S0031182025100620
Pérez-Morales, D. and Espinoza, B. (2015). The role of small heat shock proteins in parasites. Cell. Stress Chaperones 20, 767–780. doi: 10.1007/s12192-015-0607-y
Pinna, V., Di Martino, J., Liberati, F., Bottoni, P., and Castrignanò, T. (2024). “IGUANER - differential gene expression and fUnctionAlaNalyzER,” in Lecture notes in computer science, vol. 14516 . Eds. Sachdeva, S. and Watanobe, Y.. (Switzerland: Springer Nature), 78–93. doi: 10.1007/978-3-031-58502-9_5
Sandhu, A., Badal, D., Sheokand, R., Tyagi, S., and Singh, V. (2021). Specific collagens maintain the cuticle permeability barrier in Caenorhabditis elegans. Genetics 217, iyaa047. doi: 10.1093/genetics/iya047
Sen, A., Dewaker, V., Debnath, U., Jana, K., Rath, J., Joardar, N., et al. (2023). In silico exploration and in vitro validation of the filarial thioredoxin reductase inhibitory activity of Scytonemin and its derivatives. J. Biomol. Struct. Dyn 21, 1–13. doi: 10.1080/07391102.2023.2283876
Seo, M., Kho, B.-M., Guk, S.-M., Lee, S.-H., and Chai, J.-Y. (2006). Radioresistance of Anisakis simplex third-stage larvae and the possible role of superoxide dismutase. J. Parasitol. 92, 416–418. doi: 10.1645/GE-610R.1
Sher, A., Pearce, E., and Kaye, P. (2003). Shaping the immune response to parasites: role of dendritic cells. Curr. Opin. Immunol. 15, 421–429.
Stryiński, R., Mateos, J., Carrera, M., Jastrzębski, J. P., Bogacka, I., and Łopieńska-Biernat, E. (2022). Tandem mass tagging (TMT) reveals tissue-specific proteome of L4 larvae of Anisakis simplex ss: Enzymes of energy and/or carbohydrate metabolism as potential drug targets in anisakiasis. Int. J. Mol. Sci. 23, 4336. doi: 10.3390/ijms23084336
Stryiński, R., Polak, I., Gawryluk, A., Rosa, P., and Łopieńska-Biernat, E. (2024). The response of Anisakis simplex (s. s.) to anthelmintics - Specific changes in xenobiotic metabolic processes. Exp. Parasitol., 261, 108751. doi: 10.1016/j.exppara.2024.108751
Trumbić, Ž., Hrabar, J., Palevich, N., Carbone, V., and Mladineo, I. (2021). Molecular and evolutionary basis for survival, its failure, and virulence factors of the zoonotic nematode Anisakis pegreffii. Genomics 113, 2891–2905. doi: 10.1016/j.ygeno.2021.06.032
Valentini, A., Mattiucci, S., Bondanelli, P., Webb, S. C., Mignucci-Giannone, A. A., Colom-Llavina, M. M., et al. (2006). Genetic relationships among Anisakis species (Nematoda: Anisakidae) inferred from mitochondrial cox2 sequences, and comparison with allozyme data. J. Parasitol. 92, 156–166. doi: 10.1645/GE-3504.1(2006
Yu, W., Yuan, X., Zhai, P., Li, X., and Han, C. (2025). Effects of Trichinella spiralis excretory-secretory antigens on expression of indoleamine 2, 3-dioxygenase on dendritic cells in vitro. Parasite 32, 26. doi: 10.1051/parasite/2025018
Zamora, V., Andreu-Ballester, J. C., Rodero, M., and Cuéllar, C. (2021). Anisakis simplex: Immunomodulatory effects of larval antigens on the activation of Toll like Receptors. International. Immunopharmacol. 100, 108120. doi: 10.1016/j.intimp.2021.108120
Keywords: human anisakiasis, Anisakis pegreffii, human dendritic cells, RNA-seq, gene expression, oxidative stress, energy metabolism
Citation: Palomba M, Rughetti A, Castrignanò T, Napoletano C, Roca-Geronès X, Pinna V, Liberati F, Canestrelli D and Mattiucci S (2025) Transcriptomic profile of the zoonotic parasite Anisakis pegreffii upon in vitro exposure to human dendritic cells. Front. Cell. Infect. Microbiol. 15:1646537. doi: 10.3389/fcimb.2025.1646537
Received: 13 June 2025; Accepted: 26 August 2025;
Published: 15 September 2025.
Edited by:
Collette Britton, University of Glasgow, United KingdomReviewed by:
Daniel A. Abugri, Alabama State University, United StatesSaurabh Gupta, GLA University, India
Copyright © 2025 Palomba, Rughetti, Castrignanò, Napoletano, Roca-Geronès, Pinna, Liberati, Canestrelli and Mattiucci. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Simonetta Mattiucci, c2ltb25ldHRhLm1hdHRpdWNjaUB1bmlyb21hMS5pdA==; Tiziana Castrignanò, dGl6aWFuYS5jYXN0cmlnbmFub0B1bml0dXMuaXQ=
†These authors have contributed equally to this work