 Matthias C. Rillig1,2*
Matthias C. Rillig1,2* Wolfgang Kiessling2,3,4
Wolfgang Kiessling2,3,4 Thomas Borsch2,5
Thomas Borsch2,5 Arthur Gessler2,6,7
Arthur Gessler2,6,7 Alex D. Greenwood2,8Heribert Hofer2,8
Alex D. Greenwood2,8Heribert Hofer2,8 Jasmin Joshi2,9
Jasmin Joshi2,9 Boris Schröder2,10
Boris Schröder2,10 Kirsten Thonicke2,11Klement Tockner2,12Karoline Weisshuhn1,2Florian Jeltsch2,13
Kirsten Thonicke2,11Klement Tockner2,12Karoline Weisshuhn1,2Florian Jeltsch2,13- 1Plant Ecology, Institute of Biology, Freie Universität Berlin, Berlin, Germany
- 2Berlin-Brandenburg Institute of Advanced Biodiversity Research (BBIB), Berlin, Germany
- 3GeoZentrum Nordbayern, Friedrich-Alexander Universität Erlangen-Nürnberg, Erlangen, Germany
- 4Museum für Naturkunde, Leibniz Institute for Research on Evolution and Biodiversity, Berlin, Germany
- 5Botanischer Garten und Botanisches Museum Berlin-Dahlem, Freie Universität Berlin, Berlin, Germany
- 6Forest Dynamics, Swiss Federal Research Institute WSL, Birmensdorf, Switzerland
- 7Leibniz Center for Agricultural Landscape Research (ZALF), Müncheberg, Germany
- 8Leibniz Institute for Zoo and Wildlife Research (IZW), Berlin, Germany
- 9Biodiversity Research/Systematic Botany, University of Potsdam, Potsdam, Germany
- 10Institute of Geoecology, Technische Universität Braunschweig, Braunschweig, Germany
- 11Potsdam Institute for Climate Impact Research (PIK), Potsdam, Germany
- 12Leibniz-Institute of Freshwater Ecology and Inland Fisheries (IGB), Berlin, Germany
- 13Plant Ecology and Conservation Biology, University of Potsdam, Potsdam, Germany
Policy Support and Biodiversity Assessment
Meet two famous researchers from the early days of biodiversity research: Charles Darwin and Alexander von Humboldt. Darwin developed a powerful theory, using a limited amount of data by modern standards. Humboldt, in contrast, compiled a “Cosmos” of data without developing a major theory, although some of Humboldt's observations on latitudinal biodiversity gradients (von Humboldt, 1808) were later used to develop theory. This tension between data and theory still persists today and is perhaps becoming more acute.
We are on the verge of a major biodiversity crisis (Pimm et al., 2014; Steffen et al., 2015), and biodiversity research is receiving increased attention and funding worldwide. National and international political frameworks are changing, resulting in an impressive increase in coordinated biodiversity assessment initiatives. There is also an increasing number of experimental studies on functional biodiversity research; many of them require massive infrastructure investments and are admirable logistical achievements of the biodiversity research community. Nevertheless, they are, by necessity, restricted in space, time, and structural complexity. The linkage between global assessment centers and experimental setups is weak or not as strong as it could be at present because, as we argue here, they lack a shared theoretical background. On the one hand, an enormous amount of data is gathered without explicit reference to theory, whereas on the other hand there are theories that would benefit from more empirical studies. Building on recent calls for guidelines for biodiversity research (Perrings et al., 2011) we propose a theory-guided development of biodiversity research and assessment strategies to help advance this field into a scientific discipline, which in turn may advance conservation and management strategies.
Data without Theory
Natural history museums around the world are crammed with primary biodiversity data, collected over centuries, usually without a specific theoretical question. Together with the huge number of observational data gathered by professionals and laymen, a decent picture of global biodiversity patterns emerges. These data are precious, especially because they are increasingly made publically available through initiatives such as GBIF and others (Table 1). Numerous publications currently use GBIF data, mostly targeting conservation issues. But the GBIF data are rarely used for hypothesis testing, let alone the development of theory.
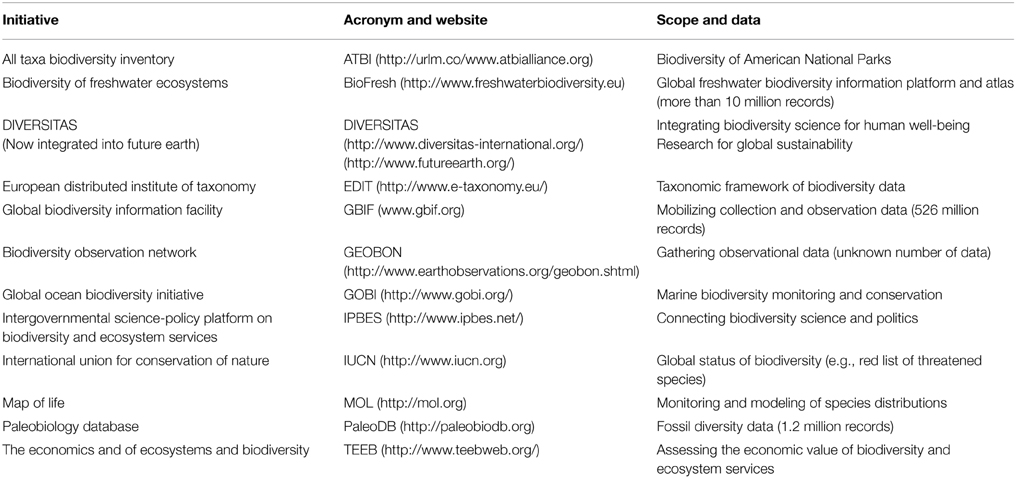
Table 1. Examples of biodiversity initiatives and their respective scopes (sorted alphabetically by acromyn).
We use “theory” here in the pluralistic sense for ecology (Pickett et al., 2007): a system of conceptual constructs, prone to continuous refinement, which is linked to observable phenomena via causal explanation, generalization, and testing within a specified domain. Theory should not be mistaken for (numerical) modeling, which is merely one component of theory-building.
We need theory because it is at the core of understanding in science (Pickett et al., 2007). Only a continuous iteration of testing data against theory-derived hypotheses will allow for a more in-depth and generalized view of biodiversity and ecosystem functioning. Owing to limited resources, such generalization will always be necessary, even with continuous development of high-throughput techniques (e.g., for metagenomics). In this sense, theory may enhance the speed of knowledge gain, help sharpen the profile of biodiversity research as a scientific discipline, and prevent biodiversity research to be used as a buzzword that runs the risk of vanishing once funding priorities shift. Some might argue we have already an over-abundance of theory in biodiversity research. Do we really need more concepts and theories? To the contrary, we do not envision more theory but a better integration of empirical research and theory directly aiming at solving pressing problems, for example in conservation biology.
Our call for theory goes beyond a call for hypothesis-driven research. Many, if not all, studies operating within the hypothesis-testing framework examine relatively narrow hypotheses, often aimed at falsification. These are not necessarily linked to larger theoretical concepts and are often too specific. Merely hypothesis-driven data collection approaches in complex systems might thus be prone to biases depending on the selection of the target variables (Kitsios and Zintzaras, 2009). The main criterion to distinguish theory-driven research and hypothesis-driven research is generalizability. Generalizability is a call for abstraction of factors and a move away from the notion of “uniqueness of place” (Beven, 2000). Sometimes this may be just a change in perspective for even the same given study: at parity of scientific quality of a study, this effort to generalize represents an added value when feedback to theory refinement is discussed.
Theory without Data
Ever since Hutchinson (1961) raised the paradox of diversity: “that the number of species in many systems greatly exceeds the number of limiting resources,” a significant number of theories tried to explain the coexistence of species in diverse systems. Theory development in community ecology has been so rapid in the past decade that empirical data, including tests of theory, are sorely needed (Agrawal et al., 2007).
There is a rich gradient of partly contrasting and partly overlapping biodiversity theories ranging from equilibrium or non-equilibrium concepts of niche separation (Chase and Leibold, 2003; Holt, 2009) to Hubbell's hotly debated “neutral theory” of biodiversity (Hubbell, 2001). In addition, the appropriate basic functional level of biodiversity is under debate: most current theoretical perspectives choose the individual scale or the level of species traits. Trait-based approaches, i.e., characterizing organisms in terms of their multiple biological attributes such as physiological, morphological, or life-history traits, promise to open new avenues in linking theories with data (Agrawal et al., 2007; Jeltsch et al., 2008) and are expected to strengthen theoretical linkages between community and ecosystem ecology (McGill et al., 2006; Reiss et al., 2009; Krause et al., 2014).
While there are several approaches for integrating and synthesizing some of the existing biodiversity theories there is currently no comprehensive “grand theory” which unifies all facets and fields of biodiversity research (McGill, 2010) This is not surprising given the relative youth of biodiversity research as a focal scientific topic. Theory maturation requires the continuous dialogue between observable phenomena and conceptual constructs (Pickett et al., 2007). Whereas some biodiversity theories or their components have been frequently tested against observational data (e.g., the “neutral theory” of biodiversity, Dornelas et al., 2006; Cardinale et al., 2009; Jabot and Chave, 2009), others still lack empirical testing, mainly due to logistic complexity. An example of the latter are theory components that require experiments at the landscape level, such as manipulations of dispersal kernels that permit the distinction between the diversity gains by species immigration (Loreau and Mouquet, 1999) and related losses by reduced species segregation at larger scales (Bolker and Pacala, 1997). Especially challenging are questions concerning the mechanisms affecting biodiversity at different spatiotemporal scales and how they depend on the overall species pool. Other examples include tests of the proposed relevance of facilitation and mutualism in shaping diversity pattern across scales (Bruno et al., 2003) or the broad application of biodiversity theory in microbial ecology (Prosser et al., 2007).
Current integrated initiatives in monitoring and experimental biodiversity research offer, in principle, the unique opportunity to fill such gaps in theory testing and refinement. These initiatives could build platforms for truly integrative biodiversity research that is oriented toward conceptual clarity, unification and generalization. However, this requires a more prominent role of theory in the earliest phases of program development and evaluation.
Conclusions
Biodiversity research has started as a rather unconnected combination of several disciplines (e.g., ecology, evolution, taxonomy, paleobiology) with separate subdisciplines, traditions and research foci. To successfully integrate this disciplinary diversity and to eventually become a self-contained discipline, biodiversity research needs to invest in the development of a mature body of theory. As welcome and needed as current biodiversity initiatives are, many lack this clear orientation toward linking empirical and theoretical approaches and theory-driven understanding.
More flexible frameworks for biodiversity monitoring and data collection schemes are needed that contribute to and integrate advances in theory development and that are driven by the search for deductive or causal laws and empirical generalizations. Beyond the search for empirical patterns, they should include data collection on functional features, integrate experiments and modeling efforts related to theory testing and refinement, and allow for flexible funding allocation and feedbacks between the combined approaches.
A giant leap toward an integration-oriented and theory-driven biodiversity science would be the critical evaluation of research proposals: panels should check whether research questions and hypotheses clearly relate to biodiversity theory, and, importantly, how the expected results will feed back into theory enhancement. Moreover, future funding frameworks in the field of biodiversity research should explicitly integrate or even focus on theory testing and development and on the education and training of young researchers in this field. A first obstacle to overcome here is the fact that some funding agencies may work with too simplistic concepts of what constitutes theory (May, 2010).
Theory, with its demand for conceptual clarity, its provision of models of how biological systems are put together and how they work, is likely to be the most promising tool to prepare biodiversity science for the challenges of the coming decades. There might be a general “theory-aversion” among biodiversity researchers, because theories are thought to oversimplify the real world, different rules seem to operate at different scales, and most biodiversity theories are stochastic rather than deterministic (McGill, 2010). To overcome this aversion we all need a bit more of the spirit of Darwin combined with Humboldt.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We thank Lord Robert May, Clive Jones, and Karl Flessa for constructive comments on earlier versions of this paper.
References
Agrawal, A. A., Ackerly, D. D., Adler, F., Arnold, A. E., Caceres, C., Doak, D. F., et al. (2007). Filling key gaps in population and community ecology. Front. Ecol. Environ. 5, 145–152. doi: 10.1890/1540-9295(2007)5[145:FKGIPA]2.0.CO;2
Beven, K. J. (2000). Uniqueness of place and process representations in hydrological modelling. Hydrol. Earth Syst. Sci. 4, 203–213. doi: 10.5194/hess-4-203-2000
Bolker, B., and Pacala, S. W. (1997). Using moment equations to understand stochastically driven spatial pattern formation in ecological systems. Theor. Popul. Biol. 52, 179–197. doi: 10.1006/tpbi.1997.1331
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Bruno, J. F., Stachowicz, J. J., and Bertness, M. D. (2003). Inclusion of facilitation into ecological theory. Trends Ecol. Evol. 18, 119–125. doi: 10.1016/S0169-5347(02)00045-9
Cardinale, B. J., Hillebrand, H., Harpole, W. S., Gross, K., and Ptacnik, R. (2009). Separating the influence of resource ‘availability’ from resource ‘imbalance’ on productivity-diversity relationships. Ecol. Lett. 12, 475–487. doi: 10.1111/j.1461-0248.2009.01317.x
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Chase, J., and Leibold, M. (2003). Ecological Niches. Linking Classical and Contemporary Apporaches. Chicago, IL: University of Chicago Press.
Dornelas, M., Connolly, S. R., and Hughes, T. P. (2006). Coral reef diversity refutes the neutral theory of biodiversity. Nature 440, 80–82. doi: 10.1038/nature04534
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Holt, R. D. (2009). Bringing the Hutchinsonian niche into the 21st century: ecological and evolutionary perspectives. Proc. Natl. Acad. Sci. U.S.A. 106, 19659–19665. doi: 10.1073/pnas.0905137106
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Hubbell, S. P. (2001). The Unified Theory of Biogeography and Biodiversity. Princeton, NJ: Princeton University Press.
Jabot, F., and Chave, J. (2009). Inferring the parameters of the neutral theory of biodiversity using phylogenetic information and implications for tropical forests. Ecol. Lett. 12, 239–248. doi: 10.1111/j.1461-0248.2008.01280.x
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Jeltsch, F., Moloney, K. A., Schurr, F. M., Kochy, M., and Schwager, M. (2008). The state of plant population modelling in light of environmental change. Perspect. Plant Ecol. Evol. Syst. 9, 171–189. doi: 10.1016/j.ppees.2007.11.004
Kitsios, G. D., and Zintzaras, E. (2009). Genome-wide association studies: hypothesis-“free” or “engaged”? Transl. Res. 154, 161–164. doi: 10.1016/j.trsl.2009.07.001
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Krause, S., Le Roux, X., Niklaus, P. A., Van Bodegom, P. M., Lennon, J. T., Bertilsson, S., et al. (2014). Trait-based approaches for understanding microbial biodiversity and ecosystem functioning. Front. Microbiol. 5:251. doi: 10.3389/fmicb.2014.00251
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Loreau, M., and Mouquet, N. (1999). Immigration and the maintenance of local species diversity. Am. Nat. 154, 427–440. doi: 10.1086/303252
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
May, R. M. (2010). Tropical arthropod species, more or less? Science 329, 41–42. doi: 10.1126/science.1191058
McGill, B. J. (2010). Towards a unification of unified theories of biodiversity. Ecol. Lett. 13, 627–642. doi: 10.1111/j.1461-0248.2010.01449.x
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
McGill, B. J., Enquist, B. J., Weiher, E., and Westoby, M. (2006). Rebuilding community ecology from functional traits. Trends Ecol. Evol. 21, 178–185. doi: 10.1016/j.tree.2006.02.002
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Perrings, C., Duraiappah, A., Larigauderie, A., and Mooney, H. (2011). The biodiversity and ecosystem services science-policy interface. Science 331, 1139–1140. doi: 10.1126/science.1202400
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Pickett, S. T. A., Kolasa, J., and Jones, C. G. (2007). Ecological Understanding: The Nature of Theory and the Theory of Nature, 2nd Edn. San Diego, CA: Academic Press.
Pimm, S. L., Jenkins, C. N., Abell, R., Brooks, T. M., Gittleman, J. L., Joppa, L. N., et al. (2014). The biodiversity of species and their rates of extinction, distribution, and protection. Science 344:1246752. doi: 10.1126/science.1246752
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Prosser, J. I., Bohannan, B. J. M., Curtis, T. P., Ellis, R. J., Firestone, M. K., Freckleton, R. P., et al. (2007). The role of ecological theory in microbial ecology. Nat. Rev. Microbiol. 5, 384–392. doi: 10.1038/nrmicro1643
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Reiss, J., Bridle, J. R., Montoya, J. M., and Woodward, G. (2009). Emerging horizons in biodiversity and ecosystem functioning research. Trends Ecol. Evol. 24, 505–514. doi: 10.1016/j.tree.2009.03.018
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Steffen, W., Richardson, K., Rockström, J., Cornell, S. E., Fetzer, I., Bennett, E. M., et al. (2015). Planetary boundaries: guiding human development on a changing planet. Science 347:1259855. doi: 10.1126/science.1259855
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Keywords: biodiversity, theory, policy, data bases, ecology, evolution, conservation
Citation: Rillig MC, Kiessling W, Borsch T, Gessler A, Greenwood AD, Hofer H, Joshi J, Schröder B, Thonicke K, Tockner K, Weisshuhn K and Jeltsch F (2015) Biodiversity research: data without theory—theory without data. Front. Ecol. Evol. 3:20. doi: 10.3389/fevo.2015.00020
Received: 03 February 2015; Accepted: 18 February 2015;
Published: 05 March 2015.
Edited by:
Gustavo A. B. Da Fonseca, Global Environment Facility, USAReviewed by:
G. Wilson Fernandes, Universidade Federal de Minas Gerais, BrazilCopyright © 2015 Rillig, Kiessling, Borsch, Gessler, Greenwood, Hofer, Joshi, Schröder, Thonicke, Tockner, Weisshuhn and Jeltsch. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Matthias C. Rillig,bWF0dGhpYXMucmlsbGlnQGZ1LWJlcmxpbi5kZQ==