 Margaret J. Couvillon*
Margaret J. Couvillon* Francis L. W. Ratnieks
Francis L. W. Ratnieks- Laboratory of Apiculture and Social Insects, School of Life Sciences, University of Sussex, Brighton, UK
Here we explore how waggle dance decoding may be applied as a tool for ecology by evaluating the benefits and limitations of the methodology compared to other existing ways to evaluate the honey bees' use of the landscape. The honey bee foragers sample and “report” back on large areas (c. 100 km2). Because honey bees perform dances only for the most profitable resources, these data provide spatial information about the availability of good quality forage for any given time. We argue that dance decoding may inform on a range of ecological, conservation, and land management issues. In this way, one species and methodology gives us a novel measure of a landscape's profitability, or “health,” that may be widely relevant, not just for honey bees, but for other flower-visiting insects as well.
Introduction
Wilhem Nylander, a Finnish-born botanist, liked to stroll around Paris, finding perhaps that the more verdant parts of the city reminded him of his Helsinki origins. It was during one of these walks, through the Jardin du Luxembourg, that the idea of bioindication was born (Skye, 1979). For years, Nylander had studied lichens, which occur worldwide from the humid tropics to the arctic and can even colonize bare rock. However, they are very sensitive to air pollution. Nylander noticed that there were more lichen species growing in the Luxembourg Gardens than in other parts of Paris. He hypothesized that lichens were susceptible to atmospheric pollutants and therefore failed to thrive in much of Paris, which was very polluted at that time (Nylander, 1866).
Today, lichens are still routinely used as an assessment of air quality (Pinho et al., 2004). In the years since Nylander, strides have been made to clean up the air, and the lichens have responded by returning to Paris (Skye, 1979). Pollution, however, is not the only challenge humans have introduced to the earth. In the last century, man-made landscape changes, such as agricultural intensification, have had a large, negative impact on biodiversity. For example, insect pollinators that depend on the presence of flowers and flower-rich habitats have been recently challenged by the conversion of those areas to intensively farmed land (Matson et al., 1997; Tilman et al., 2001; Robinson and Sutherland, 2002; Carvell et al., 2006; Winfree et al., 2011; Wright and Wimberly, 2013). The link between landscape changes and pollinator declines has generated intense interest from both government and private sectors (Berenbaum et al., 2007; Kluser et al., 2011), such as a recent announcement by the United States government that they are giving $8 million to increase available forage for Midwestern honey bees (USDA, 2014). However, it is difficult to know not only when and where supplementary forage would be most beneficial, but also to know if such policies, once in place, are having the intended effect.
The answer may lie with the honey bee, an organism that itself would benefit from a healthy landscape. Honey bees, with their unique waggle dance communication, may be an untapped, biologically relevant resource to provide cost-effective bioindication by surveying, monitoring, and reporting on a landscape's “health,” specifically in regards to floral abundance.
What Information is Available from Waggle Dance Decoding?
Honey bees, Apis species, possess a remarkable and unique behavior. A successful forager communicates to her nestmates the vector (distance and direction) from the hive to the nectar or pollen (von Frisch, 1946, 1967; Grüter and Farina, 2009; Couvillon, 2012) by making waggle dances. The vector information, which the bee repeats many times within a single dance and which is encoded in the waggle phase portion of the dance, can be decoded by researchers (Visscher and Seeley, 1982; Beekman and Ratnieks, 2000; Steffan-Dewenter and Kuhn, 2003; Couvillon et al., 2012b; Schürch et al., 2013). By decoding many dances, a map of where the colony as a whole (or indeed several colonies) is foraging can be made. Importantly, honey bee foragers only dance for profitable resources, which means that, on average, observed dances are for the “best” forage that is available at any given time (von Frisch, 1967; Seeley, 1994) after the forager has weighed the benefit of the forage against the costs, including the distance she must fly (von Frisch, 1967; Seeley, 1994; Couvillon et al., 2014b,c). The dance allows the honey bee colony to exploit either particularly good resources or resources when availability is low, and it therefore contributes to the fitness of the colony (Seeley, 1995; Sherman and Visscher, 2002; Dornhaus et al., 2006; Donaldson-Matasci and Dornhaus, 2012; Schürch and Grüter, 2014).
Dancing foragers also give direct and indirect information about the forage quality, which could include quantity and the molarity of the nectar. Directly, the number of repeated circuits within a single dance per return to the hive indicates the forager's assessment of quality: when resource quality increases, the forager makes more waggle runs per dance, and this results in the recruitment of more nestmates to the advertised location (von Frisch, 1967; Seeley, 1995; Seeley et al., 2000). Indirectly, a particularly good location may be indicated by multiple dances by the same or different bees, as more and more bees become recruited to the location that they indicate themselves in subsequent dances. When such multiple dances are plotted, “hotspots” of good quality forage become visible (Beekman and Ratnieks, 2000; Steffan-Dewenter and Kuhn, 2003; Couvillon et al., 2014b).
Lastly, it is also possible to know if a particular dance is for a pollen or a non-pollen, usually nectar, source because the pollen is visible in the corbiculae (Couvillon et al., 2014a). Although it is quite difficult to obtain the pollen from a dancing bee for identification, it is possible for pollen traps to be placed over the entrance to collect pollen from all returning foragers (e.g., colony-level information about pollen collection). These data on plant type composition, particularly a high representation of a single plant type (Garbuzov and Ratnieks, 2014), can then be correlated to the forage location information obtained by dance decoding (Table 1). Knowing the breadth of the plant community composition gives indication of the landscape's biodiversity.
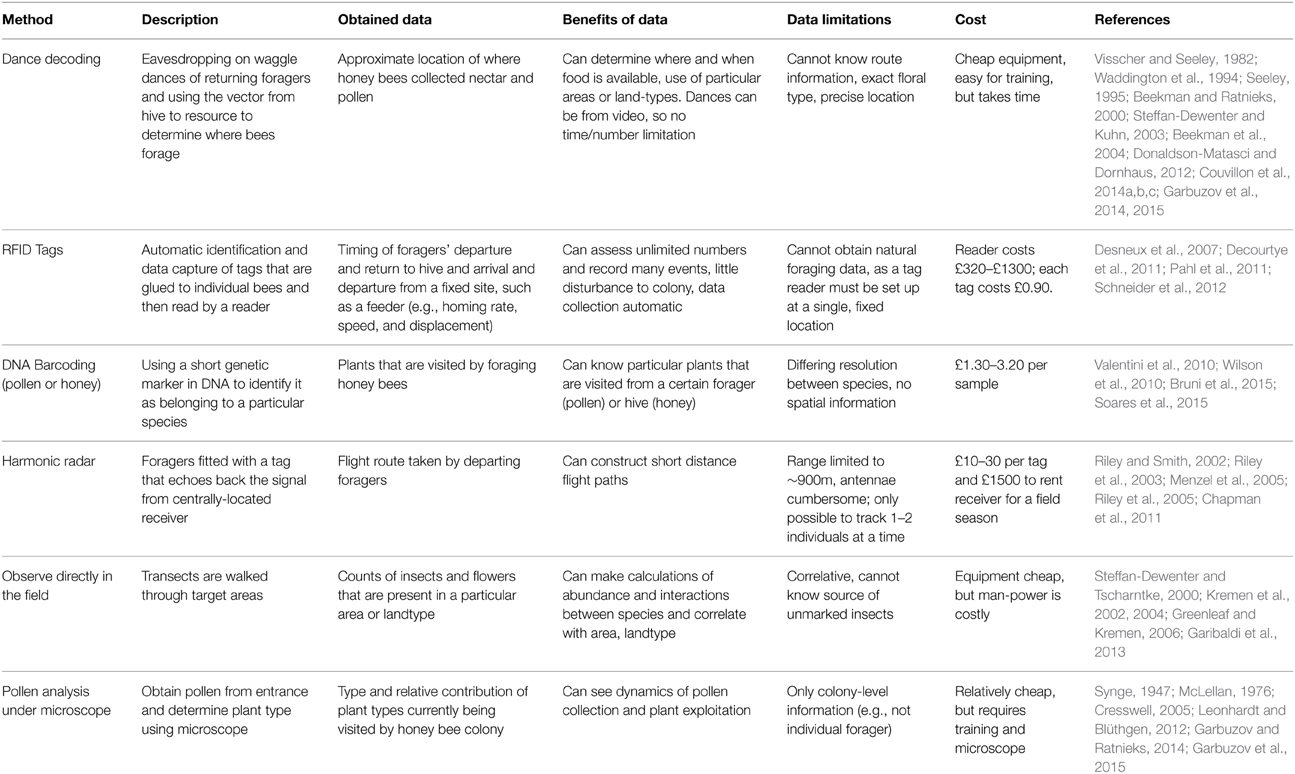
Table 1. Comparison of methodologies to investigate honey bee foraging.
What Information is Not Available from Waggle Dance Decoding?
Firstly, it is not possible to know the route, including obstacle avoidance, that a forager took from the hive to the resource (von Frisch, 1967). Route information can be partially reconstructed if dance decoding is combined with harmonic radar (Table 1), where a radar signal is received by a transponder on an insect and re-emitted such that the flight path of a honey bee is tracked (Riley et al., 2003, 2005; Menzel et al., 2005). However, the antenna for harmonic radar can only be applied after a dancing A. mellifera forager exits the hive, when it would actually be better to obtain the path information from the flight that precedes the dance. Additionally, harmonic radar is limited by a range of approximately 900 m, or the first hedge, for signal reception (Riley and Smith, 2002; Chapman et al., 2011). This would not cover the long-distance foraging of many kilometers that honey bees are capable of performing (von Frisch, 1967; Visscher and Seeley, 1982; Beekman and Ratnieks, 2000; Couvillon et al., 2014c).
It is equally challenging to know exactly what flowers were visited by a dancing bee. As previously mentioned, pollen samples can be obtained from the pollen traps at the entrance, which gives colony-level information on the plants that have been visited, but individual dances cannot then be linked to the samples. It is potentially possible to collect a pollen forager mid-dance (Hart and Ratnieks, 2001), once she has danced long enough for the data to be extracted (Couvillon et al., 2012b), but this would involve using a cover (e.g., polyurethane and not glass) that can be easily cut through, which may be intractable for large scale analyses. The pollen then could be identified, either by shape (Synge, 1947; McLellan, 1976; Cresswell, 2011; Garbuzov and Ratnieks, 2014), or by DNA markers, such as barcoding (Table 1, Bruni et al., 2015).
Lastly, because the honey bee is not a very precise dancer, decoding dances does not give exact location information. An individual honey bee often forages in a small area of approximately 10 × 10 m (Ribbands, 1953); however, the waggle dance made by her at different times or made by other bees working the same patch will display considerable scatter in both vector components of distance and direction (e.g., inter-dance variation, Schweiger, 1958; von Frisch and Lindauer, 1961). The variation found within a dance between successive, repeated circuit (intra-dance variation) (Beekman et al., 2005; De Marco et al., 2008; Tanner and Visscher, 2010a,b; Couvillon et al., 2012b; Schürch and Couvillon, 2013; Schürch et al., 2013; Preece and Beekman, 2014) also makes the signal challenging to understand. Certainly it is simply a limitation in dance accuracy that we cannot pinpoint a foraging location to, for example, a hedge.
One way to combat the issue of dance imprecision when we want to know where a bee foraged is to incorporate the variability from within the dance into the mapping of the dance itself. In Schürch et al. (2013), a Bayesian approach combined with simulations allowed for the production of a spatial probability distribution for a forage location. Instead of plotting a dance as a point, which over-represents our certainty about a location, dances were plotted as a heat map (Schürch et al., 2013; Couvillon et al., 2014c). Additionally, when multiple dances, either from different bees going to the same location or the same bee dancing multiple times, are rasterized together, a more accurate estimation of a “hot spot” can be achieved (Couvillon et al., 2014b; Garbuzov et al., 2015).
It should be noted that for many specific investigations, we do not need to know a precise location to obtain biologically-relevant information about honey bee foraging ecology. For example, because distance is such a relevant cost in a bees' decision to recruit and because honey bees have evolved exceptional sensitivity to relative energetic reward, communicated distance is a proxy for forage availability: the further a dancer indicates in her recruitment, the relatively less available forage is nearby (Couvillon et al., 2014c). Because dance duration translates into flight distance, examining the average dance duration per month provides important information about in what months forage is relatively less or more available (Beekman and Ratnieks, 2000; Couvillon et al., 2014c) or how relative distances (durations) vary between urban and rural and suburban settings (Waddington et al., 1994; Garbuzov et al., 2014) or across differently structured landscapes (Steffan-Dewenter and Kuhn, 2003).
Why is the Honey Bee a Useful Bioindicator of Landscape Floral Profitability?
Even given the above limitations, honey bees possess great potential for monitoring the landscape for floral resources. One reason is because the foraging range of the honey bee is so great, probably greater than other bees. Honey bees routinely forage at a distance of a few kilometers and have been known to fly 10–12 km to collect food when it is less available closer to the hive (von Frisch, 1967; Waddington et al., 1994; Beekman and Ratnieks, 2000; Steffan-Dewenter and Kuhn, 2003). In Couvillon et al. (2014b), by statistically correcting for distance, authors were able to assess honey bee foraging preferences across 94 km2, which represents the “surveying power” of three glass-walled observation hives from one location (Couvillon et al., 2014b). A second reason why the honey bee is an ideal bioindicator is because they are generalists in their foraging preferences (Biesmeijer and Slaa, 2006). The flower species visited by honey bees for nectar and pollen will also be visited by other pollinators, which makes information about honey bee foraging preferences broadly relevant to a range of flower-visiting insects. As a proof of concept to the general nature of honey bee foraging, the bees indicated that the highest visited area in the 94 km2 was a National Nature Reserve that is an area well known as being good for butterflies (Ellis et al., 2012; Couvillon et al., 2014b).
Thirdly, the honey bees possess the waggle dance, which is the only known recruitment behavior that also communicates a distance and direction, information available to eavesdropping researchers. A honey bee forager, in her decision to perform a waggle dance, integrates all relevant costs of flight distance, potential competition with other flower-visiting insects, and nectar and pollen availability, and if her assessment comes out in the positive, she performs a dance. Imagine how difficult it would be to obtain those data directly on the availability of good forage across a landscape-wide area. An ecologist would need to walk transects to count competing flower-visiting insects and flowers and determine forage availability. If an ecologist walks a straight-line transect at 1 m/s and can see a meter to the left and to the right, s/he will cover about 200 m2 in 5 min. Even at this speedy clip and if s/he worked 24 h/day, it would take that ecologist 1632 days to cover 94 km2 one time. Therefore, it is safe to say that the data obtained from decoding waggle dances is not available by surveying.
What are the Potential Subject Areas that would Benefit from Waggle Dance Decoding Data?
In addition to the applied application of landscape evaluation, data obtained from waggle dance decoding could be relevant in a variety of yet-unexplored contexts (Härtel and Steffan-Dewenter, 2014). Honey bees are important pollinators for many crops (Aizen and Harder, 2009), contributing £27 billion annually to the world economy from this ecological service. Combining waggle dance information with the colony-level pollen collection will help farmers better manage pollination services: pollen analysis would allow growers to determine if a target crop was visited or not, and dance information would allow growers to determine which field areas were visited (Kremen et al., 2002; Duan et al., 2008). These data would then work together to help manage the process (e.g., to fine-tune the bloom time such that bees are not attracted elsewhere). Additionally, dance information can also give other information, such as the time taken for a colony's foraging to recover following relocation (Riddell Pearce et al., 2013), which also helps for optimal management of pollination services.
In recent decades, there has been increasingly intense interest in mitigating the harmful effects of anthropogenic landscape changes through government incentives to encourage more wildlife-friendly farming. These stewardship schemes are required for all EU-member states and carry an impressive price-tag of €41 billion in the past 20 years. And yet there are little data evaluating the efficacy of such schemes, probably because it is hard to survey at the landscape scale. However, by monitoring the dances of honey bees foraging naturally in the landscape, Couvillon et al. (2014b) determined that some management stewardship schemes, such as the ones that encouraged set-asides or field margins in High Level Stewardship, may be more beneficial to honey bees than others, such as Organic Entry Level Stewardship that require the regular mowing of grass, including wildflowers (Couvillon et al., 2014b). The study provided biologically relevant data that may directly impact land management and existing efforts at nature conservation for honey bees and other flower-visiting insects and adds to a growing body of work linking pollinator health to landscape composition/“health” (Kremen et al., 2004; Klein et al., 2007; Brittain et al., 2013).
Because the honey bees are collecting their food in the landscape, they are potentially exposed to man-made toxins and may therefore provide useful information in environmental risk assessment. For example, in 2013 the European Union imposed a moratorium on the use of neonicotinoid insecticides as seed treatments for bee-attractive crops, such as oilseed rape (canola). However, one potentially missing element in the assessment of pesticide exposure is a better understanding of just how much foraging bees carry out on the treated crops. Recently it was demonstrated that in a typical European landscape that contains the mass-flowering crop of oilseed rape, which is a potential source of neonicotinoids, honey bees visit the nearby fields between 0 and 23% of the foraging time, depending on distance from hive and time of year (Garbuzov et al., 2015). By decoding waggle dances, researchers will better understand the dynamics of exposure (Garbuzov et al., 2015).
The waggle dance is one of the few Nobel Prize winning discoveries that can be seen with the naked eye in real time. The finding was exciting and, at times, controversial (Gould, 1976; Munz, 2005; Couvillon, 2012). “Language,” as von Frisch diplomatically called the behavior, was thought to be in the exclusive domain of humans. In the waggle dance, not only a non-human but an invertebrate is able to communicate symbolically, using the sun and gravity as a reference, where she has collected good forage. Decoding waggle dances therefore is also a wonderful gateway to biology, providing an observable, interesting behavior to get the public engaged in science. Dance decoding also may be easily demonstrated in the classroom and to the larger, non-scientific community. In fact, the interest and amazement that the public express about the waggle dance and dance decoding may also represent a powerful opportunity for citizen science. It is possible to instruct anyone, even someone without a science background, into the fundamentals of waggle dance decoding, such that video recordings of dancing bees, uploaded to videos, can be decoded by community volunteers. There is great interest at the moment in helping bees, and tapping into the lay community may represent a powerful workforce that in turn are afforded an opportunity to help the honey bee in a very real way by contributing to our understanding of how and where they collect their food.
Lastly, although this article has mostly underscored the applied uses of the honey bee waggle dance, it is worth noting that studies involving decoded waggle dances also generate significant gains for basic biology, specifically behavioral ecology, foraging ecology, and neuroscience. Karl von Frisch, the Austrian-born ethologist who shared the 1973 Nobel Prize for discovering the waggle dance communication, famously remarked that the honey bee “is like a magic well,” in that the discoveries never end. Certainly this applies to the waggle dance. In recent years, for example, we have witnessed many exciting discoveries about the nature of the honey bee waggle dance. These discoveries include the honey bee dance stop signal, which is made by foragers when they encounter a nestmate dancing for a location that is dangerous (Nieh, 2010), a discovery that helps our understanding of positive and negative feedback loops in self-organizing systems; the flexible use of dance information vs. memory by honey bees (Grüter et al., 2008; Grüter and Ratnieks, 2011); the effect of gravity on the angular intra-dance (im)precision (Couvillon et al., 2012a), which demonstrates how a biological entity manages to communicate in the presence of noise; the increased benefit of the dance for larger-sized colonies (Donaldson-Matasci et al., 2013), which aids in our understanding of how to optimize collective exploitation; and the ability of flying bees to obtain and to signal compass information purely from polarized light (Evangelista et al., 2014), which provides important insight into the capabilities of invertebrate visual navigation. The list is in no way exhaustive and only serves to demonstrate that there are still exciting and unknown features to be uncovered about this amazing behavior.
Conclusion: Is It Worth It?
Dance decoding is a relatively easy task. No specialist scientific training is needed and a person can be instructed on how to do it within a few hours. However, the method is time-consuming, especially if one wishes to decode hundreds or thousands of dances. After training, a decoder can decode a dance in under 5 min, but this estimate does not include the time spent watching a video to find a bee that is dancing. In all, it may take a trained worker several hours to locate and decode 20 dances from 1 h of video of the dance floor area of an observation hive. Therefore, it is worth considering a discussion on whether or not dance decoding is worth it? Perhaps decoding large numbers of dances, which would take the contributions of many people over many months, would not be worthwhile for basic biology alone. However, the applied benefits that may be gained that can be used to help honey bees and other insect pollinators could outweigh the costs.
Inherent in the discussion of “Is dance decoding worth it” is a comparison between dance decoding with other methods that may be used to assess where and when honey bees are collecting nectar and pollen (Table 1). Across all the methods, dance decoding is the only way to obtain, at the level of the individual, location information about where nectar and pollen has been collected, and it remains the superior method to use when investigating honey bee use of the landscape for forage (e.g., to assess WHERE bees are foraging; Table 1). Tagging foragers with RFID tags or harmonic radar can provide data on the timing of departure and return from a fixed site (e.g., the hive) or a short-distance flight path; however, neither can be applied realistically to field foraging conditions to determine where bees are collecting nectar and pollen. Tagging foragers generates information on HOW bees are foraging (Table 1). Likewise, methods that analyze the forage itself, such as DNA barcoding or pollen analysis, while providing important information about WHAT the bees are visiting, are limited in that neither provides location information, and both are challenging to perform at the level of the individual (Table 1). Lastly, field observations remain correlative, as it would be difficult to mark insects to identify their colony of origin.
Because each method carries its own unique benefits and costs, interesting results can be obtained if methods are combined. For example, because RFID tags require a tag reader, they can be used in conjunction with feeder experiments, where tagged foragers are trained to collect sugar water from a set feeder location. These bees, which may carry an additional identifying paint mark, may then analyzed for their dance behavior. In this instance, the actual vector information in the dance is less interesting, as it will communicate the feeder; however, the other interesting questions that require individual-level information regarding dancing and recruitment behavior may be investigated. Secondly, dance decoding can be effectively combined with forage analysis (barcoding or pollen microscopy) in circumstances where the bee may be foraging upon a particular target crop that is growing in known locations to generate powerful individual and colony level information about food collection and pollination services (Garbuzov et al., 2015).
Currently, there is no reliable method to automate the process, despite decades' worth of effort and even with recent advances in machine vision and automated video analysis for other organisms (Mersch et al., 2013). The goal has always been to create a system that can detect and decode waggle runs reliably. Although some advancement has taken place in the latter (Kimura et al., 2011; Landgraf et al., 2011), the process of detecting dances remains problematic. The dancing bee is just one moving part of a larger, very busy, and also moving background of her nestmates, and attempts to detect dances automatically leads to a large number of false positives [e.g., 199 true dances detected and 25 false positives (Rau, 2014)]. Also, no one has a real sense of the number of false negatives (waggle dances that fail to be detected). However, recently the field appears to be right on the cusp of observing significant advances in this area (see Landgraf et al. in this special issue).
In conclusion, it is our opinion that, given the time cost of dance decoding by hand, if the data obtained by the methodology were relevant only from a basic honey bee behavioral ecology perspective, it would not be worth to undertake large-scale projects. However, given that these data are unique and not possible to obtain any other way, and given that it may provide biologically-relevant information that could be used to help honey bees and other pollinators and to use resources wisely, we say yes. In dance decoding, we let the honey bees do the hard work to survey huge areas of land. Our job is only to listen to the bees.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We would like to thank all the people over the years who have worked on Project 2 of the Sussex Plan, “Decoding waggle dances to see where bees forage” and to all LASI members, past and present. MJC was funded by a grant from The Nineveh Charitable Trust and from charitable donations from Rowse Honey and Waitrose Ltd.
References
Aizen, M. A., and Harder, L. D. (2009). The global stock of domesticated honey bees is growing slower than agricultural demand for pollination. Curr. Biol. 19, 915–918. doi: 10.1016/j.cub.2009.03.071
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Beekman, M., Doyen, L., and Oldroyd, B. (2005). Increase in dance imprecision with decreasing foraging distance in the honey bee Apis mellifera L. is partly explained by physical constraints. J. Comp. Physiol. A 191, 1107–1113. doi: 10.1007/s00359-005-0034-0
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Beekman, M., and Ratnieks, F. L. W. (2000). Long-range foraging by the honey-bee, Apis mellifera L. Funct. Ecol. 14, 490–496. doi: 10.1046/j.1365-2435.2000.00443.x
Beekman, M., Sumpter, D. J. T., Seraphides, N., and Ratnieks, F. L. W. (2004). Comparing foraging behaviour of small and large honey-bee colonies by decoding waggle dances made by foragers. Funct. Ecol. 18, 829–835. doi: 10.1111/j.0269-8463.2004.00924.x
Berenbaum, M., Bernhardt, P., Buchmann, S., Calderone, N. W., Goldstein, P., Inouye, D. W., et al. (2007). Status of Pollinators in North America. Washington, DC: The National Academies Press.
Biesmeijer, J. C., and Slaa, J. E. (2006). The structure of eusocial bee assemblages in Brazil. Apidologie 37, 240–258. doi: 10.1051/apido:2006014
Brittain, C., Williams, N., Kremen, C., and Klein, A. M. (2013). Synergistic effects of non-Apis bees and honey bees for pollination services. Proc. R. Soc. B.. 280:20122767. doi: 10.1098/rspb.2012.2767
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Bruni, I., Galimberti, A., Caridi, L., Scaccabarozzi, D., De Mattia, F., Casiraghi, M., et al. (2015). A DNA barcoding approach to identify plant species in multiflower honey. Food Chem. 170, 308–315. doi: 10.1016/j.foodchem.2014.08.060
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Carvell, C., Roy, D. B., Smart, S. M., Pywell, R. F., Preston, C. D., and Goulson, D. (2006). Declines in forage availability for bumblebees at a national scale. Biol. Conserv. 132, 481–489. doi: 10.1016/j.biocon.2006.05.008
Chapman, J. W., Drake, V. A., and Reynolds, D. R. (2011). Recent insights from radar studies of insect flight. Annu. Rev. Entomol. 56, 337–356. doi: 10.1146/annurev-ento-120709-144820
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Couvillon, M. J. (2012). The dance legacy of Karl von Frisch. Insectes Soc. 59, 297–306. doi: 10.1007/s00040-012-0224-z
Couvillon, M. J., Phillipps, H. L. F., Schürch, R., and Ratnieks, F. L. W. (2012a). Working against gravity: horizontal honeybee waggle runs have greater angular scatter than vertical waggle runs. Biol. Lett. 8, 540–543. doi: 10.1098/rsbl.2012.0182
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Couvillon, M. J., Riddell Pearce, F. C., Accleton, C., Fensome, K. A., Quah, S. K. L., Taylor, E. L., et al. (2014a). Honey bee foraging distance depends on month and forage type. Apidologie 1–10. doi: 10.1007/s13592-014-0302-5
Couvillon, M. J., Riddell Pearce, F. C., Harris-Jones, E. L., Kuepfer, A. M., Mackenzie-Smith, S. J., Rozario, L. A., et al. (2012b). Intra-dance variation among waggle runs and the design of efficient protocols for honey bee dance decoding. Biol. Open 1, 467–472. doi: 10.1242/bio.20121099
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Couvillon, M. J., Schürch, R., and Ratnieks, F. L. W. (2014b). Dancing bees communicate a foraging preference for rural lands under High Level Agri-Environment Schemes. Curr. Biol. 24, 1212–1215. doi: 10.1016/j.cub.2014.03.072
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Couvillon, M. J., Schürch, R., and Ratnieks, F. L. W. (2014c). Waggle dance distances as integrative indicators of seasonal foraging challenges. PLoS ONE 9:e93495. doi: 10.1371/journal.pone.0093495
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Cresswell, J. E. (2005). Accurate theoretical prediction of pollinator-mediated gene dispersal. Ecology 86, 574–578. doi: 10.1890/04-1109
Cresswell, J. E. (2011). A meta-analysis of experiments testing the effects of a neonicotinoid insecticide (imidacloprid) on honey bees. Ecotoxicology 20, 149–157. doi: 10.1007/s10646-010-0566-0
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Decourtye, A., Devillers, J., Aupinel, P., Brun, F., Bagnis, C., Fourrier, J., et al. (2011). Honeybee tracking with microchips: a new methodology to measure the effects of pesticides. Ecotoxicology 20, 429–437. doi: 10.1007/s10646-011-0594-4
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
De Marco, R. J., Gurevitz, J. M., and Menzel, R. (2008). Variability in the encoding of spatial information by dancing bees. J. Exp. Biol. 211, 1635–1644. doi: 10.1242/jeb.013425
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Desneux, N., Decourtye, A., and Delpuech, J.-M. (2007). The sublethal effects of pesticides on beneficial arthropods. Annu. Rev. Entomol. 52, 81–106. doi: 10.1146/annurev.ento.52.110405.091440
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Donaldson-Matasci, M. C., Degrandi-Hoffman, G., and Dornhaus, A. (2013). Bigger is better: honeybee colonies as distributed information-gathering systems. Anim. Behav. 85, 585–592. doi: 10.1016/j.anbehav.2012.12.020
Donaldson-Matasci, M., and Dornhaus, A. (2012). How habitat affects the benefits of communication in collectively foraging honey bees. Behav. Ecol. Sociobiol. 66, 583–592. doi: 10.1007/s00265-011-1306-z
Dornhaus, A., Klügl, F., Oechslein, C., Puppe, F., and Chittka, L. (2006). Benefits of recruitment in honey bees: effects of ecology and colony size in an individual-based model. Behav. Ecol. 17, 336–344. doi: 10.1093/beheco/arj036
Duan, J. J., Marvier, M., Huesing, J., Dively, G., and Huang, Z. Y. (2008). A meta-analysis of effects of Bt crops on honey bees (Hymenoptera: Apidae). PLoS ONE 3:e1415. doi: 10.1371/journal.pone.0001415
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Ellis, S., Bourn, N., and Bulman, C. (2012). Landscape-scale Conservation for Butterflies and Moths: Lessons from the UK. Wareham: Butterfly Conservation.
Evangelista, C., Kraft, P., Dacke, M., Labhart, T., and Srinivasan, M. V. (2014). Honeybee navigation: critically examining the role of the polarization compass. Philos. Trans. R. Soc. Lond. B Biol. Sci. 369:20130037. doi: 10.1098/rstb.2013.0037
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Garbuzov, M., Couvillon, M. J., Schürch, R., and Ratnieks, F. L. W. (2015). Honey bee dance decoding shows limited foraging on oilseed rape - a potential source of neonicotinoid contamination. Agric. Ecosystems Environ. 203, 62–68. doi: 10.1016/j.agee.2014.12.009
Garbuzov, M., and Ratnieks, F. L. W. (2014). Ivy: an underappreciated key resource to flower visiting insects in autumn. Insect. Conserv. Diver. 7, 91–102. doi: 10.1111/icad.12033
Garbuzov, M., Schürch, R., and Ratnieks, F. L. W. (2014). Eating locally: dance decoding demonstrates that urban honey bees in Brighton, UK, forage mainly in the surrounding urban area. Urban Ecosystems 7, 91–102. doi: 10.1007/s11252-014-0403-y
Garibaldi, L. A., Steffan-Dewenter, I., Winfree, R., Aizen, M. A., Bommarco, R., Cunningham, S. A., et al. (2013). Wild pollinators enhance fruit set of crops regardless of honey bee abundance. Science. 339, 1608–1611. doi: 10.1126/science.1230200
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Gould, J. L. (1976). The dance-language controversy. Q. Rev. Biol. 51, 211–244. doi: 10.1086/409309
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Greenleaf, S. S., and Kremen, C. (2006). Wild bees enhance honey bees' pollination of hybrid sunflower. Proc. Natl. Acad. Sci. U.S.A. 103, 13890–13895. doi: 10.1073/pnas.0600929103
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Grüter, C., Balbuena, M. S., and Farina, W. M. (2008). Informational conflicts created by the waggle dance. Proc. Biol. Sci. 275, 1321–1327. doi: 10.1098/rspb.2008.0186
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Grüter, C., and Farina, W. M. (2009). The honeybee waggle dance: can we follow the steps? Trends Ecol. Evol. 24, 242–247. doi: 10.1016/j.tree.2008.12.007
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Grüter, C., and Ratnieks, F. L. W. (2011). Honeybee foragers increase the use of waggle dance information when private information becomes unrewarding. Anim. Behav. 81, 949–954. doi: 10.1016/j.anbehav.2011.01.014
Hart, A. G., and Ratnieks, F. L. W. (2001). Why do honey-bee (Apis millifera) foragers transfer nectar to several receivers? Information improvement through multiple sampling in a biological system. Behav. Ecol. Sociobiol. 49, 244–250. doi: 10.1007/s002650000306
Härtel, S., and Steffan-Dewenter, I. (2014). Ecology: honey bee foraging in human-modified landscapes. Curr. Biol. 24, R524–R526. doi: 10.1016/j.cub.2014.04.052
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Kimura, T., Ohashi, M., Okada, R., and Ikeno, H. (2011). A new approach for the simultaneous tracking of multiple honeybees for analysis of hive behavior. Apidologie 42, 607–617. doi: 10.1007/s13592-011-0060-6
Klein, A.-M., Vaissière, B. E., Cane, J. H., Steffan-Dewenter, I., Cunningham, S. A., Kremen, C., et al. (2007). Importance of pollinators in changing landscapes for world crops. Proc. Biol. Sci. 274, 303–313. doi: 10.1098/rspb.2006.3721
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Kluser, S., Neumann, P., Chauzat, M.-P., and Pettis, J. S. (2011). UNEP Emerging Issues: Global Honey Bee Colony Disorder and Other Threats to Insect Pollinators. Geneva: United Nations Environment Programme.
Kremen, C., Williams, N. M., Bugg, R. L., Fay, J. P., and Thorp, R. W. (2004). The area requirements of an ecosystem service: crop pollination by native bee communities in California. Ecol. Lett. 7, 1109–1119. doi: 10.1111/j.1461-0248.2004.00662.x
Kremen, C., Williams, N. M., and Thorp, R. W. (2002). Crop pollination from native bees at risk from agricultural intensification. Proc. Natl. Acad. Sci. U.S.A. 99, 16812–16816. doi: 10.1073/pnas.262413599
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Landgraf, T., Rojas, R., Nguyen, H., Kriegel, F., and Stettin, K. (2011). Analysis of the waggle dance motion of honeybees for the design of a biomimetic honeybee robot. PLoS ONE 6:e21354. doi: 10.1371/journal.pone.0021354
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Leonhardt, S. D., and Blüthgen, N. (2012). The same, but different: pollen foraging in honeybee and bumblebee colonies. Apidologie 43, 449–464. doi: 10.1007/s13592-011-0112-y
Matson, P. A., Parton, W. J., Power, A. G., and Swift, M. J. (1997). Agricultural intensification and ecosystem properties. Science 277, 504–509. doi: 10.1126/science.277.5325.504
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
McLellan, A. R. (1976). Factors affecting pollen harvesting by the honeybee. J. Appl.Ecol. 13, 801–811. doi: 10.2307/2402256
Menzel, R., Greggers, U., Smith, A., Berger, S., Brandt, R., Brunke, S., et al. (2005). Honey bees navigate according to a map-like spatial memory. Proc. Natl. Acad. Sci. U.S.A. 102, 3040–3045. doi: 10.1073/pnas.0408550102
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Mersch, D. P., Crespi, A., and Keller, L. (2013). Tracking individuals shows spatial fidelity is a key regulator of ant social organization. Science 340, 1090–1093. doi: 10.1126/science.1234316
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Munz, T. (2005). The bee battles: Karl von Frisch, Adrian Wenner and the honey bee dance language controversy. J. Hist. Biol. 38, 535–570. doi: 10.1007/s10739-005-0552-1
Nieh, J. C. (2010). A negative feedback signal that is triggered by peril curbs honey bee recruitment. Curr. Biol. 20, 310–315. doi: 10.1016/j.cub.2009.12.060
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Nylander, M. W. (1866). Les lichens du Jardin du Luxembourg. Bull. Soc. Bot. France 13, 364–371. doi: 10.1080/00378941.1866.10827433
Pahl, M., Zhu, H., Tautz, J., and Zhang, S. (2011). Large scale homing in honeybees. PLoS ONE 6:e19669. doi: 10.1371/journal.pone.0019669
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Pinho, P., Augusto, S., Branquinho, C., Bio, A., Pereira, M. J., Soares, A., et al. (2004). Mapping Lichen diversity as a first step for air quality assessment. J. Atmos. Chem. 49, 377–389. doi: 10.1007/s10874-004-1253-4
Preece, K., and Beekman, M. (2014). Honeybee waggle dance error: adaption or constraint? Unravelling the complex dance language of honeybees. Anim. Behav. 94, 19–26. doi: 10.1016/j.anbehav.2014.05.016
Ribbands, C. R. (1953). The Behaviour and Social Life of Honeybees. London: Bee Research Association Limited.
Riddell Pearce, F. C., Couvillon, M. J., and Ratnieks, F. L. W. (2013). Hive relocation does not adversely affect honey bee (Hymenoptera: Apidae) foraging. Psyche 2013, 8. doi: 10.1155/2013/693856
Riley, J. R., Greggers, U., Smith, A. D., Reynolds, D. R., and Menzel, R. (2005). The flight paths of honeybees recruited by the waggle dance. Nature 435, 205–207. doi: 10.1038/nature03526
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Riley, J. R., Greggers, U., Smith, A. D., Stach, S., Reynolds, D. R., Stollhoff, N., et al. (2003). The automatic pilot of honeybees. Proc. R. Soc. Lond. B Biol. Sci. 270, 2421–2424. doi: 10.1098/rspb.2003.2542
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Riley, J. R., and Smith, A. D. (2002). Design considerations for an harmonic radar to investigate the flight of insects at low altitude. Comput. Electron. Agric. 35, 151–169. doi: 10.1016/S0168-1699(02)00016-9
Robinson, R. A., and Sutherland, W. J. (2002). Post-war changes in arable farming and biodiversity in Great Britain. J. Appl. Ecol. 39, 157–176. doi: 10.1046/j.1365-2664.2002.00695.x
Schneider, C. W., Tautz, J., Grünewald, B., and Fuchs, S. (2012). RFID tracking of sublethal effects of two neonicotinoid insecticides on the foraging behavior of Apis mellifera. PLoS ONE 7:e30023. doi: 10.1371/journal.pone.0030023
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Schürch, R., and Couvillon, M. J. (2013). Too much noise on the dance floor: intra- and inter-dance angular error in honey bee waggle dances. Commun. Integr. Biol. 6:e22298. doi: 10.4161/cib.22298
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Schürch, R., Couvillon, M. J., Burns, D. D. R., Tasman, K., Waxman, D., and Ratnieks, F. L. W. (2013). Incorporating variability in honey bee waggle dance decoding improves the mapping of communicated resource locations. J. Comp. Physiol. A Neuroethol. Sens. Neural Behav. Physiol. 199, 1143–1152. doi: 10.1007/s00359-013-0860-4
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Schürch, R., and Grüter, C. (2014). Dancing bees improve colony foraging success as long-term benefits outweigh short-term costs. PLoS ONE 9:e104660. doi: 10.1371/journal.pone.0104660
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Schweiger, E. M. (1958). Über individuelle unterschiede in der entfernungs- und richtungsangabe bei den tänzen der bienen. Z. Vergl. Physiol. 41, 272–299. doi: 10.1007/BF00365323
Seeley, T. D. (1994). Honey bee foragers as sensory units of their colonies. Behav. Ecol. Sociobiol. 34, 51–62. doi: 10.1007/BF00175458
Seeley, T. D., Mikheyev, A. S., and Pagano, G. J. (2000). Dancing bees tune both duration and rate of waggle-run production in relation to nectar-source profitability. J. Comp. Physiol. A 186, 813–819. doi: 10.1007/s003590000134
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Sherman, G., and Visscher, P. K. (2002). Honeybee colonies achieve fitness through dancing. Nature 419, 920–922. doi: 10.1038/nature01127
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Skye, E. (1979). Lichens as biological indicators of air pollution. Annu. Rev. Phytopathol. 17, 325–341. doi: 10.1146/annurev.py.17.090179.001545
Soares, S., Amaral, J. S., Oliveira, M. B. P. P., and Mafra, I. (2015). Improving DNA isolation from honey for the botanical origin identification. Food Control 48, 130–136. doi: 10.1016/j.foodcont.2014.02.035
Steffan-Dewenter, I., and Kuhn, A. (2003). Honeybee foraging in differentially structured landscapes. Proc. Biol. Sci. 270, 569–575. doi: 10.1098/rspb.2002.2292
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Steffan-Dewenter, I., and Tscharntke, T. (2000). Resource overlap and possible competition between honey bees and wild bees in central Europe. Oecologia 122, 288–296. doi: 10.1007/s004420050034
Synge, A. D. (1947). Pollen collection by honeybees (Apis mellifera). J. Anim. Ecol. 16, 122–138. doi: 10.2307/1492
Tanner, D. A., and Visscher, P. K. (2010a). Adaptation or constraint? Reference-dependent scatter in honey bee dances. Behav. Ecol. Sociobiol. 64, 1081–1086. doi: 10.1007/s00265-010-0922-3
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Tanner, D. A., and Visscher, P. K. (2010b). Does imprecision in the waggle dance fit patterns predicted by the tuned-error hypothesis? J. Insect Behav. 23, 180–188. doi: 10.1007/s10905-010-9204-1
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Tilman, D., Fargione, J., Wolff, B., D'antonio, C., Dobson, A., Howarth, R., et al. (2001). Forecasting agriculturally driven global environmental change. Science 292, 281–284. doi: 10.1126/science.1057544
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
USDA. (2014). “USDA provides $8 million to help boost declining honey bee populations,” in U. S. D. O. Agriculture (Washington, DC: USDA). Available online at: http://www.fsa.usda.gov/FSA/newsReleases?area=newsroom&subject=landing&topic=ner&newstype=newsrel&type=detail&item=nr_20140620_rel_0130.html
Valentini, A., Miquel, C., and Taberlet, P. (2010). DNA barcoding for honey biodiversity. Diversity 2, 610–617. doi: 10.3390/d2040610
Visscher, P. K., and Seeley, T. D. (1982). Foraging strategy of honeybee colonies in a temperate deciduous forest. Ecology 63, 1790–1801. doi: 10.2307/1940121
von Frisch, K. (1967). The Dance Language and Orientation of Bees. Cambridge, MA: Harvard University Press.
von Frisch, K., and Lindauer, M. (1961). Über die “Mißweisung” bei den richtungsweisenden tänzen der bienen. Naturwissenschaften 48, 585–594. doi: 10.1007/BF00601984
Waddington, K. D., Herbert, T. J., Visscher, P. K., and Richter, M. R. (1994). Comparisons of forager distributions from matched honey bee colonies in suburban environments. Behav. Ecol. Sociobiol. 35, 423–429. doi: 10.1007/BF00165845
Wilson, E. E., Sidhu, C. S., Levan, K. E., and Holway, D. A. (2010). Pollen foraging behaviour of solitary Hawaiian bees revealed through molecular pollen analysis. Mol. Ecol. 19, 4823–4829. doi: 10.1111/j.1365-294X.2010.04849.x
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Winfree, R., Bartomeus, I., and Cariveau, D. P. (2011). Native pollinators in anthropogenic habitats. Annu. Rev. Ecol. Evol. System. 42, 1–22. doi: 10.1146/annurev-ecolsys-102710-145042
Wright, C. K., and Wimberly, M. C. (2013). Recent land use change in the Western Corn Belt threatens grasslands and wetlands. Proc. Natl. Acad. Sci. U.S.A. 110, 4134–4139. doi: 10.1073/pnas.1215404110
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Keywords: waggle dance, Apis mellifera, dance decoding, bee foraging, environmental monitoring
Citation: Couvillon MJ and Ratnieks FLW (2015) Environmental consultancy: dancing bee bioindicators to evaluate landscape “health.” Front. Ecol. Evol. 3:44. doi: 10.3389/fevo.2015.00044
Received: 05 December 2014; Accepted: 17 April 2015;
Published: 01 May 2015.
Edited by:
Madeleine Beekman, The University of Sydney, AustraliaReviewed by:
James C. Nieh, University of California San Diego, USAMatina Donaldson-Matasci, Harvey Mudd College, USA
Copyright © 2015 Couvillon and Ratnieks. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Margaret J. Couvillon, Laboratory of Apiculture and Social Insects, School of Life Sciences, University of Sussex, John Maynard Smith Building, Brighton, BN1 9QG, UK,bWFnZ2llamFuZWNAZ21haWwuY29t