 Jenélle L. Dowling
Jenélle L. Dowling Diane Colombelli-Négrel
Diane Colombelli-Négrel Michael S. Webster1
Michael S. Webster1- 1Cornell Lab of Ornithology, Department of Neurobiology and Behaviour, Cornell University, Ithaca, NY, USA
- 2School of Biological Sciences, Flinders University, Adelaide, SA, Australia
Many vocal animals recognize kin using vocal cues, in territorial contexts and in rearing young, but little is known about the developmental and evolutionary mechanisms that produce vocal kin recognition systems. In the cooperatively breeding red-backed fairy-wren (Malurus melanocephalus), females give specific “in-nest calls” while incubating their eggs. Elements from these calls are incorporated into chicks' begging calls, and appear to be used by parents for recognition. This is likely a result of an embryo's ability to learn the call elements in the egg. Here, we explore the idea that maternal vocal elements may be incorporated into offspring's adult songs, and serve as signatures of kinship, which would aid in kin recognition and benefit signaler and receiver. To investigate this hypothesis, we tested for similarities between maternal vocal elements (in-nest calls and songs) and songs of their adult offspring. We then determined whether offspring songs were more similar to the maternal vocalizations they heard only as embryos (in-nest calls), or maternal songs they heard throughout development, but mostly post-fledge (mothers' full songs). We used dynamic time warping to compare maternal vocal elements with elements in their offspring's adult songs. The elements of each offspring were more similar to the elements of their own mother than to the elements of any other female (but only for the average similarity score), suggesting they may serve as kin signatures. We also found that offspring song elements were more similar to their mother's song elements than to their mother's in-nest call elements. In addition, female in-nest call elements were more similar to their own song elements than to the song elements of any other female. Offspring that produced song elements highly similar to their mother's song elements also had song elements highly similar to their mother's in-nest call elements. Signals of kinship may function critically for survival and successful reproduction, by allowing an individual to allocate care and defense to kin and to avoid mating with genetic relatives. Understanding the processes that facilitate effective kin recognition, including development of kin signatures, helps us to understand how these crucial signaling systems may have evolved.
Introduction
Kin recognition can provide adaptive benefits to both senders and receivers of kinship signals. Receivers of such signals can provide care and feeding to related young, alert kin to resources and danger, and avoid mating with genetic relatives (Holmes and Sherman, 1983; Akcay et al., 2013). Senders can also benefit from reliably identifying themselves as kin, if this allows them to obtain nepotistic benefits and avoid inbreeding (Beecher, 1988). When senders benefit from being recognized, selection is expected to act directly on signal properties to enhance the reliability of kin signatures (Beecher, 1988).
Kin recognition systems have been described in different signaling modalities and across taxa (Porter, 1986; Hurst and Beynon, 2010). Despite this, we have a limited understanding of how kin signatures develop, or the mechanisms that underlie similarity among kin. Therefore, studies that elucidate how, when and from whom individuals acquire signatures that may be used for kin recognition are needed (Sharp et al., 2005). This information is an important piece in the evolutionary puzzle of how kin recognize one another, since senders and receivers can only benefit from recognition if individuals accurately acquire and produce signals produced by their kin (Waldman, 1987). We expect selection for ontogenetic mechanisms that minimize acquisition mistakes. Investigating these questions allows us to better understand the evolutionary underpinnings of kin recognition, a process that is critical for survival and successful reproduction across the majority of social animal species (Holmes and Sherman, 1983; Akcay et al., 2013).
In vertebrates, receivers recognize kin signatures most commonly by learning through direct association, rather than through innate recognition (Halpin, 1991; Caspers et al., 2013). In birds specifically, associative learning of kin signatures appears to be the primary kin recognition mechanism, especially in cooperatively breeding species (Sharp et al., 2005; Caspers et al., 2013; Amo et al., 2014), and vocalizations are the most common communication modality used for avian kin recognition (Beecher, 1988; Riehl and Stern, 2015). Vocal kin signatures have been described in many taxonomic groups (Price, 1998; McDonald and Wright, 2011; Knörnschild et al., 2012) and vocal kin recognition has been experimentally supported in several mammal (Rendall et al., 1996) and bird species (Price, 1999; Sharp et al., 2005; McDonald, 2012; Akcay et al., 2013). Although these avian studies have laid excellent groundwork, our understanding of the occurrence and extent of vocal kin recognition across avian groups remains incomplete. In addition, we know little about how young birds acquire vocal kin signatures and how these ontogenetic processes are shaped by selection. Gaining a more comprehensive understanding of vocal kin recognition systems, including their development and evolution, from the perspective of both senders and receivers, helps us to elucidate the signaling behaviors that can evolve via inclusive fitness benefits.
Here, we tested for the presence of kin signatures in the adult songs of the red-backed fairy-wren (Malurus melanocephalus) and investigated a potential mechanism of kin signature acquisition, as well as the timing of acquisition. In this species, females call to their eggs and nestlings while inside the nest using a particular “in-nest” call that they do not produce in other contexts (Colombelli-Négrel et al., 2016, Figure 1). Mothers begin calling at the onset of incubation and continue to call until day 5–6 post-hatching (Colombelli-Négrel et al., 2016). After hatching, young reproduce elements from these maternal calls, incorporating them into their begging calls (Colombelli-Négrel et al., 2016). This is likely a result of the embryos' ability to learn the elements from within the egg, which has been shown in the closely related superb fairy-wren (Malurus cyaneus) (Colombelli-Négrel et al., 2012, 2014). While feeding chicks at the nest, red-backed fairy-wren parents seem to recognize young that can imitate their maternal elements better and feed them preferentially (Colombelli-Négrel et al., 2016). This behavior is thought to have evolved in superb fairy-wrens because it decreases the cost of inter-specific brood parasitism (Colombelli-Négrel et al., 2012), an evolutionary pressure that red-backed fairy-wrens are also subject to (albeit less so, in our population, Colombelli-Négrel et al., 2016). Despite our understanding of kin signature use and recognition in red-backed fairy-wrens within the context of the nest, we do not yet know whether young incorporate maternal vocal elements (including both in-nest call and full song elements) into other vocalizations, such as their adult songs.
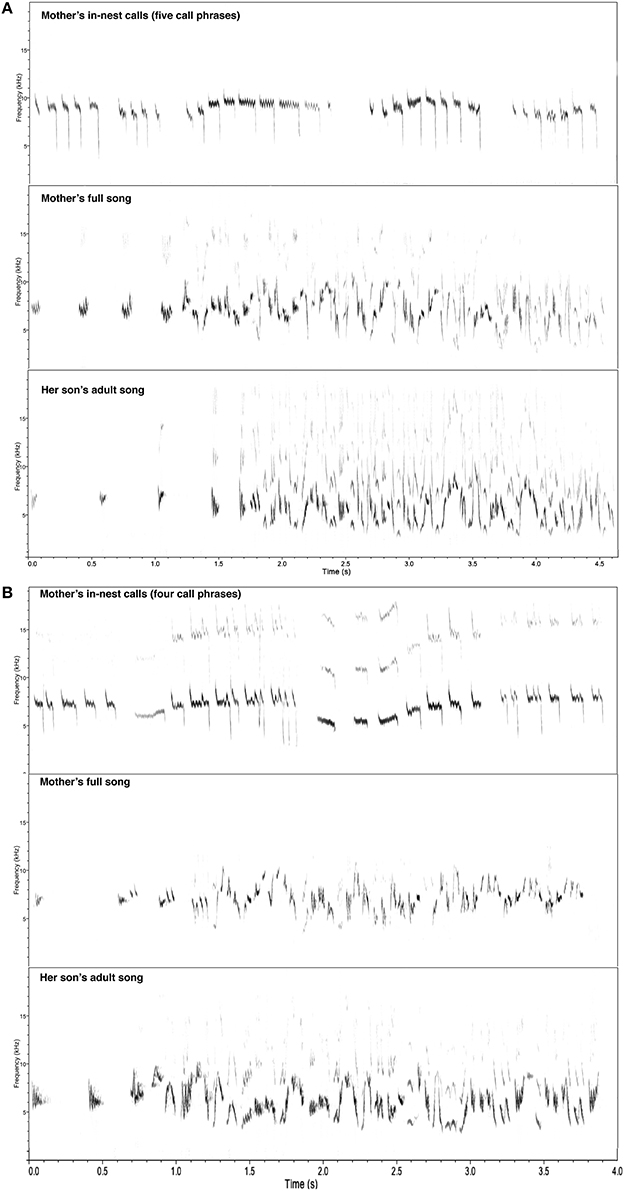
Figure 1. Example spectrograms of female in nest calls (top panels), her full songs (center panels), and her son's adult songs (bottom panels). (A) First mother-son pair example from our population. (B) Second mother-son pair example from our population.
To investigate this, we compared maternal in-nest call and full song elements to elements present in the adult songs of their offspring. In addition, we determined if adult songs of offspring were more similar to the maternal vocalizations they heard only as embryos/nestlings (in-nest calls) or to the maternal songs heard throughout development and after fledging (mothers' full songs) in order to help us to understand when kin signatures may be acquired. We predicted that if kin signatures were present in red-backed fairy-wren songs, then offspring's song elements would be more similar to the elements of their own mother than to the elements of any other female. We additionally predicted that if this species used song elements to recognize kin, then the kin signatures individuals used for recognition as nestlings might be preserved in their adult songs, which might also suggest that kin signatures may be learned in ovo and/or soon after hatching.
Methods
Field Methods
We studied a population of color-banded red-backed fairy-wrens near Herberton, Queensland, Australia (145°25′E, 17°23′S), which has been monitored continuously since 2003. The study site is located in open dry sclerophyl forest with tall eucalypt overstory and grass understory. Red-backed fairy-wrens breed cooperatively, with 34% of 1-year-old males remaining on their natal territory as auxiliary helpers (Varian-Ramos et al., 2010). A red-backed fairy-wren breeding group consists of a breeding male, breeding female and 0–3 auxiliary helpers (Varian-Ramos et al., 2010).
Females build domed nests low in the grassy understory and lay two to four (usually three) eggs per clutch. Females are solely responsible for building the nest and incubating the eggs, but all members of the group participate in feeding nestlings and fledglings (Varian-Ramos et al., 2010). Eggs usually hatch after 12 days of incubation and chicks fledge after 12 days in the nest. Fledglings remain dependent for approximately 40 days post-fledging and often remain on the territory through the winter with females, with breeding fledglings dispersing in the early spring prior to breeding. The reproductive biology of this non-migratory species is described in further detail by Webster et al. (2010).
In our study, group compositions, territory boundaries, and nest status were determined through daily observations of each breeding group and nest checks every other day. We monitored all breeding attempts by each group on the field site. On the sixth day after hatching, nestlings were banded with an aluminum band bearing a unique identifying number. We used this information to determine each bird's natal group and determined relatedness by linking offspring to the female that attended the nest, and constructing a social pedigree.
We recorded in-nest vocalizations from females in 7 nests, during the 2011-2012 breeding season (September–January). Vocalizations were recorded for 2 continuous hours per nest between days 10 and 12 of incubation. We recorded all vocalizations produced at the nest using either (1) an Olympus linear LS-10 handheld PCM recorder or (2) a Wildlife Acoustics Song Meter SM2 Autonomous recording unit (Wildlife Acoustics Inc., Concorde, MA, USA), both with a sample rate of 24 KHz in 16 bit PCM format. Both recorders had integrated microphones and were placed directly under the nest as close as possible without causing disturbance (usually 20–30 cm). All recordings were done in the morning between 0500 and 1100 h.
Songs of adult offspring were recorded in the breeding seasons of 2012-2013 and 2013-2014 during the pre-dawn chorus (Dowling and Webster, 2016). Each adult offspring was at least 1 year old and breeding independently at the time of recording. Independent breeders had established their own mate and a breeding territory; independently breeding females typically disperse several territories away from their natal territory, whereas males typically settle close to their natal territory (Varian-Ramos et al., 2010). Songs of adult offspring and full songs of mothers were recorded with a Marantz PMD 661 solid-state digital recorder (D&M Professional, Itasca, IL, U.S.A.) at 96 kHz sampling rate, 24-bit depth, connected to a K6 power module and a ME67 shotgun microphone capsule (Sennheiser Electronic Corporation, Old Lyme, CT, U.S.A.) with a Rycote softie windshield and mount. Individuals were recorded mostly during the pre-dawn chorus (Dowling and Webster, 2016). In the first comparison, we compared in-nest call elements of 7 mothers to adult song elements of 8 of their adult offspring (6 male, 2 female). Our sample of adult offspring included one offspring each for 6 mothers (5 male, 1 female), and two offspring for one of the mothers (1 male, 1 female). In the second comparison, we compared song elements of 5 mothers to adult song elements of 5 of their adult offspring (one offspring for each mother, 4 male, 1 female). In the third comparison, we compared 5 mother's in-nest call elements to full song elements from those same 5 mothers.
We transferred all recordings onto an Apple MacBook Pro (Apple Corporation, U.S.A) and spectrograms were created in RavenPro 1.5 (Cornell Laboratory of Ornithology, Ithaca, NY, USA, 16-bit sample format; discrete Fourier transform (DFT) = 512 samples; frequency resolution = 124 Hz; time resolution = 11.6 ms; frame overlap = 50%). Recordings of female in-nest calls were band-pass filtered so that only the spectral area between 1000 and 17,800 Hz remained, which removed background noise but left the signal intact. Recordings of songs of adult offspring and mothers were band-pass filtered so that only the spectral area between 2400 and 17800 Hz remained, again removing background noise but leaving the signal intact.
Element Comparisons
We selected one exemplar of each element type in each individual's repertoire for maternal in-nest calls, maternal songs, and their offspring's adult songs. Element types were classified for maternal songs and offspring adult songs using element classification methods described in Schwabl et al. (2015). In short, we used JMP 11.0 (SAS Institute Inc., Cary, NC, U.S.A.) to classify elements into groups of element types using normal mixtures clustering. From this, we identified an optimal number of clusters (i.e., element types) using AICc values (Burnham and Anderson, 2002) as the fit statistic to determine the optimal number of clusters for the element data set within the current study (i.e., we set up a new classification for the current sample rather than using element type classification from a previous study of red-backed fairy-wrens, Schwabl et al., 2015). This yielded 25 element types, which were then visually inspected to confirm that they were classified appropriately.
In-nest call elements were classified visually using the element comparison interface in Luscinia (Lachlan, 2015). This yielded 25 element types per individual, which is similar to the number of element types per song for this species (Schwabl et al., 2015, current study). Since in-nest calls typically have a smaller number of elements overall than songs (Figure 1), 25 representative elements for each female were chosen from a sample of five in-nest calls.
This generated a sample of representative exemplar elements, 175 maternal in-nest call elements (7 mothers × 25 elements each) and 200 offspring song elements (8 offspring × 25 elements each) for the comparison between maternal in-nest call elements and their offspring's adult song elements. For the comparison between maternal song elements and their offspring's adult song elements, we used a sample of 125 maternal song elements (5 mothers whose offspring's adult songs were also recorded × 25 elements each) and 125 offspring song elements (5 offspring whose mother's full songs were also recorded × 25 elements each). For the comparison between maternal song elements and maternal in-nest call elements, 125 maternal in-nest call elements (5 mothers × 25 elements each) were compared with 125 full song elements (5 mothers × 25 elements each) from those same mothers.
In each separate comparison (maternal in-nest call vs. offspring adult songs, maternal songs vs. offspring adult songs, and maternal songs vs. maternal in-nest calls), we made pair-wise comparisons of every element in the sample (i.e., each maternal element was compared to each offspring element in the sample, not just her own offspring). We used the dynamic time-warping (DTW) algorithm in the software package Luscinia (Lachlan, 2015). This algorithm searches for an optimal alignment between two time series on the basis of the Euclidean distance between acoustic features. This method is more appropriate, in this context, than other automated measurements of song similarity (e.g., spectrographic cross-correlation, Clark et al., 1987), because we do not expect offspring to produce near-exact copies of tutor's song elements, but rather to produce variations of tutor elements that have similar, but modified shape and may differ in frequency and time characteristics.
In our analysis, the acoustic features we used were spectrograph measures calculated from tracing a contour over each element, and included: time, relative position, peak frequency, mean frequency, median frequency, peak frequency change, mean frequency change, median frequency change, and frequency bandwidth (additional parameter settings in Luscinia: compression factor: 0.001, minimum element length: 10, SD ratio: 0.5, maximum warp 60%, cost for stitching syllables: 1, cost for alignment error: 1, syllable comparison by individual element, with frequencies log transformed). In Luscinia (Lachlan, 2015), frequency characteristics are measured by analyzing many spectral slices of the signal, and the actual measurement of each slice's frequency is made from a power spectrum, rather than from the original spectrogram.
We checked that DTW similarity assignments matched the subjective assessments of human observers using the element dendrogram visual display in Luscinia. The output of the DTW analysis is a dissimilarity score between each pair of elements in our sample. Therefore, we created three matrices of dissimilarity comparing: (1) maternal in-nest call elements and offspring's adult song elements (n = 8), (2) maternal song elements and offspring's adult song elements (n = 5), and (3) maternal song elements and maternal in-nest call elements (n = 5).
Each matrix of dissimilarity was analyzed separately using principal coordinate analysis (PCoA) with the R-package software for Multivariate and Spatial analysis version 4 (Casgrain and Legendre, 2001). PCoA is a data reduction and ordination technique commonly used for cluster analysis (Legendre and Legendre, 1998) that ordinates distances between objects (in this study, elements) in multi-dimensional space, while preserving Euclidean distances between those elements. It is conceptually similar to principal components analysis (PCA); however, PCoA conserves distances generated from any (dis)similarity value allowing a more flexible analysis of the complexity of the data. In addition, the orthogonal nature of PCoA has the advantage of satisfying assumptions of independence when testing statistical hypotheses.
From the matrix of dissimilarity, PCoA creates a set of independent axes that position the objects (in this study, elements) in reduced space based on their similarity between each other (Legendre and Legendre, 1998), where objects ordinated closer to one another are more similar than those ordinated further away. Each axis has an eigenvalue whose magnitude indicates the amount of variation captured in that axis and the first five eigenvalues explain most of the variation in the data set (Baker and Logue, 2003). The first five PCoA coordinate values represent our five similarity scores per element, which we then averaged to create an average similarity score. In this analysis, smaller scores represented higher similarity. For more details on the methods see Colombelli-Négrel et al. (2012) and Colombelli-Négrel et al. (2016).
We then performed statistical analyses with SPSS 18.0 for Windows (SPSS Inc., Chicago, USA). We used the five similarity scores and the average score in a MANOVA and then in a discriminant function analysis (DFA) to test for acoustical similarity in elements between individuals. The percentage of correct classification by chance for the DFA was calculated by dividing 100% by the number of individuals used in each analysis. To determine if offpring song elements were more similar to maternal in-nest call elements or maternal song elements, we compared the average similarity scores per individual using a paired t-test. We tested for the relationship between offspring's element similarity to their mother's in-nest call elements and to their mother's song elements using a linear regression.
Ethics Approval Statement
All work was conducted with approval from appropriate animal ethics and permitting agencies (Cornell University Animal Care and Use Committee approval 2009-0105, James Cook University Ethics approval number A1340, and Queensland Government Environmental Protection Agency Scientific Purposes Permit number WISP07773610).
Results
Similarity between Maternal In-Nest Calls and Offspring Adult Songs
The first eigenvalues extracted by PCoA accounted for 74.72% of the variance in the data (with 93.15% accounted for by the first five eigenvalues). We found that the element pair comparison (maternal in-nest call vs. offspring song elements) differed significantly between individuals for one of the five similarity scores and for the average score (Table 1A), suggesting that the song elements of each offspring were more similar to the in-nest call elements of their own mother than to the elements of any other female (see also Figure 2). Cross-validated DFA classified 24.5% of elements to the correct mother-offspring pair (n = 8), which was higher than the percentage of correct classification by chance (12.5%).
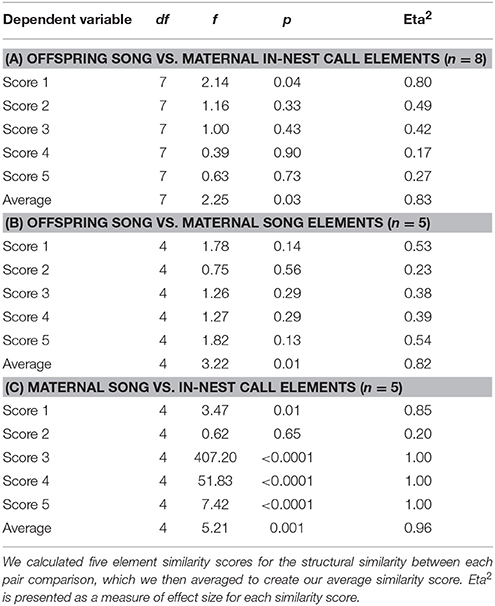
Table 1. Results of dynamic time warping comparison and principal coordinate analysis (PCoA).
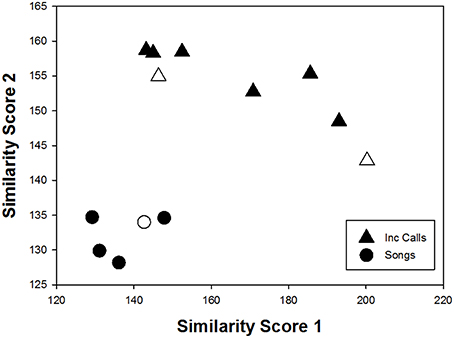
Figure 2. Scatterplot of similarity score 1 vs. 2 showing the distribution in space of offspring song elements (averaged per individual) in relation to their maternal in-nest call elements (triangles; n = 8) and maternal song elements (circles; n = 5): offspring song elements were more similar to maternal song elements than maternal in-nest call elements (smaller scores on z and y represented higher similarity). The black circles and triangles represent the male offspring and white circles and triangles represent the female offspring.
Similarity between Maternal Songs and Offspring Adult Songs
The first eigenvalues extracted by PCoA accounted for 76.75% of the variance in the data (with 94.72% accounted for by the first five eigenvalues). We found that the element pair comparison (maternal song elements vs. offspring song elements) did not differ significantly between individuals for five similarity scores but differed for the average score (Table 1B; see also Figure 2), again suggesting that the average song elements of each offspring were more similar to the song elements of their own mother than to the elements of any other female. Cross-validated DFA classified 32% of elements to the correct mother-offspring pair (n = 5), which was higher than the percentage of correct classification by chance (20%).
Offspring Song Similarity to Maternal In-Nest Calls vs. Maternal Songs
We next determined if adult songs of offspring were more similar to the maternal vocalizations they heard as embryos (in-nest calls) or maternal songs (mother's full songs). Comparisons of the averaged similarity scores per individual showed that offspring song elements were more similar to maternal song elements than maternal in-nest call elements (Paired t-test: df = 4; t = 20.376; p < 0.0001; Figure 2).
Similarity between Maternal Songs and Maternal In-Nest Calls
The first eigenvalues extracted by PCoA accounted for 53.89% of the variance in the data (with 86.27% accounted for by the first five eigenvalues). We found that female song elements were more similar to their own in-nest call elements than to the elements of any other female: the element pair comparison (maternal in-nest call vs. maternal song elements) differed significantly between individuals for four of the five similarity scores and for the average score (Table 1C). Cross-validated DFA classified 79.2% of elements to the correct female (n = 5), which was higher than the percentage of correct classification by chance (20%). In addition, offspring that produced song elements highly similar to their mother's song elements also had song elements highly similar to their mother's in-nest call elements (Linear regression: t = 6.277, β = 0.964, p = 0.008; Figure 3).
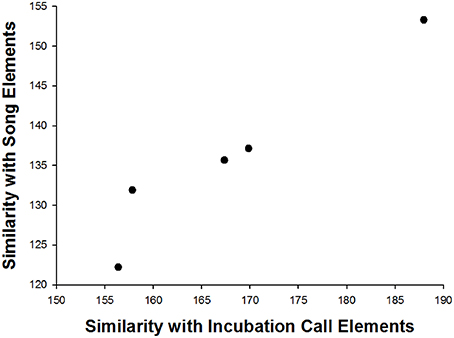
Figure 3. A positive correlation between offspring element similarity to their mothers' in-nest call elements and to their mothers' song elements: offspring that produced song elements highly similar to their mothers' song elements also had song elements highly similar to their mothers' in-nest call elements. Elements with the smallest similarity values were more similar to their mothers' elements than those with highest values.
Discussion
We found that adult birds produced song elements similar to the maternal in-nest call elements they heard as embryos in the nest as well as to their mother's full songs. However, offspring song elements were more similar to their mother's song elements than to the maternal in-nest call elements. Though we do not rule out genetic inheritance of song traits in our study, cultural transmission is extremely well supported across songbirds and likely also occurs in our study species (Slater, 1986; Podos et al., 2004). As such, our results, combined with previous studies of vocal kin recognition in red-backed fairy-wrens (Colombelli-Négrel et al., 2016) and embryonic vocal learning in superb fairy-wrens (Colombelli-Négrel et al., 2012, 2014), provide evidence that red-backed fairy-wrens incorporated both types of maternal elements into their crystallized songs, where they could serve as kin signatures. These elements were likely learned from their mothers throughout development, but perhaps also from within the egg. These signatures could be used for kin recognition, and as such, may increase the survival and reproduction of both sender and receiver. We also found that offspring that produced song elements highly similar to their mother's song elements had song elements highly similar to their mother's in-nest call elements, which suggests there is variation between young birds in how well they learn to produce elements similar to their maternal tutor in both contexts.
We hypothesized that if red-backed fairy-wrens use song to recognize kin, then the kin signatures individuals used as nestlings may be preserved in their adult songs, as this would facilitate kin recognition. Our result that offspring adult songs are similar to their mother's in-nest calls provides some indirect support for this hypothesis. However, it should be noted that the DFA assigned only 24.5% (in-nest calls) and 32% (songs) of elements to the correct mother-offspring pair, suggesting some overlap between individuals. These results may be explained by low variation between the individuals (possibly due to low sample size) or because there is some natural vocal variation within individuals over time (see Ellis, 2008). For example, Matrosova et al. (2009, 2010) showed that alarm call structures were unstable over time. In our case, a young bird's song may differ from its mother's vocalizations because it has changed over time, even though the bird learned the song from its mother originally. This may be expected if the vocalization converges on a group signature (Radford, 2005), if it encodes local dialect (Nelson and Poesel, 2007), and/or if other information influences vocalization structure (such as dominance status, Mathevon et al., 2010). Another potential explanation could be that young bird's songs differ from their mothers' because they learn vocal elements from multiple tutors. They may learn from their social father, which has been shown in several closely related species (Greig et al., 2012; Evans and Kleindorfer, 2016), but also possibly from auxiliary helpers and other nearby adults.
It is also important to note that further testing is required to confirm that this species is indeed using vocal kin signatures to recognize genetic relatives (e.g., with playback experiments, sensu Akcay et al., 2013). In addition, since offspring songs were more similar to maternal full songs, and there is high similarity between a mother's in-nest calls and her full songs, the similarity between offspring adult songs and maternal in-nest calls may only emerge due to multicollinearity. In other words, offspring may learn their adult song elements only from the songs of tutors they hear late in development (e.g., their mother's full songs), and not from maternal in-nest calls, meaning the similarity between offspring adult songs and maternal in-nest calls may exist only due to similarity between maternal in-nest calls and maternal full songs.
An alternative interpretation is that offspring learn the elements in their full songs from both their mother's in-nest calls and their mother's full songs, which would suggest that song learning occurs both within the egg/soon after hatch and later in development. It was discovered very recently that birds can discriminate between individual's calls and songs as embryos (Colombelli-Négrel et al., 2014; Colombelli-Négrel and Kleindorfer, unpublished data). Our results may provide further evidence to suggest that embryonic chicks may not only hear and respond to vocalizations while in the egg, but also may retain elements heard prenatally from those vocalizations (Colombelli-Négrel et al., 2012, 2014, 2016), and produce them later in life. Although future studies are needed (e.g., cross-fostering experiments that test whether embryos learn acoustic elements, controlling for genetic inheritance), our results may shed light on how birds acquire elements in their crystallized songs, suggesting that in some species, they may be acquired at an earlier developmental stage than previously believed. This illustrates a need for further investigation of the timing of avian vocal development, since classic and widely accepted models of song learning in birds suggest that the sensitive period for song acquisition begins much later, and propose that exposure to songs before 10 days post-hatch has no effect on learning (Marler, 1970).
Timing and length of the sensitive period for song acquisition vary widely across bird species (Marler, 1991; Brainard and Doupe, 2002). Both appear to be flexible, and tend to coincide with social and environmental contexts that are most ideal for appropriate song acquisition (Kroodsma and Pickert, 1980; Slater et al., 1992). So why would the sensitive period extend to such an early developmental stage in some species? For red-backed and other fairy-wrens (Maluridae), the sensitive period may include an extraordinarily early developmental stage because these species may experience particularly strong selective pressure for accurate kin signature acquisition. This could be due to the critical importance of accurate kin recognition in a nest provisioning context, since there is a strong threat of brood parasitism from many cuckoo species (Cuculinae spp.; Langmore et al., 2003; Colombelli-Négrel et al., 2012), and in a cooperative breeding context, since auxiliary helpers must ensure they assist adult kin (i.e., parents) and care only for siblings that are genetically related (McDonald and Wright, 2011; McDonald, 2012).
Our study helps to address how kin signatures are acquired within an individual's lifetime. Our results suggest that kin signatures are acquired from the mother (though may additionally be acquired from other adults present, including the social father and helpers, Greig et al., 2012) with a potential role of both prenatal and post-hatching learning. When senders benefit from being recognized, selection is expected to act directly on their signal properties to enhance the reliability of their kin signatures (Beecher, 1988). Early developmental learning of kin signatures could benefit both individuals that produce vocal kin signatures and those that recognize and respond to them, by minimizing mistakes in signature acquisition through limiting possible tutors to those that sing frequently within close range of the nest.
Conclusions
Currently, our understanding of vocal kin recognition systems remains incomplete (Rendall et al., 1996; Knörnschild et al., 2012; Akcay et al., 2013). General explorations of whether genetic relatives share vocal kin signatures, such as the present study, are needed across taxa to understand the occurrence and extent of vocal kin recognition. In addition, studies like ours help to further our understanding of how vocal kin recognition systems may evolve, by helping to elucidate developmental timing of kin signature acquisition, and general adaptive benefits of vocal kin recognition systems for maximizing indirect fitness benefits and minimizing costs of inbreeding. Gaining a more comprehensive understanding of kin recognition signaling systems, including the developmental and evolutionary mechanisms that produce them, helps us to better understand how adaptive signaling systems like these come to be, both within the span of an individual's lifetime and over evolutionary time.
Author Contributions
JD, MW, and DC all contributed to the conception and design of the work. JD and DC both contributed to the acquisition, analysis, and interpretation of data for the paper. JD drafted the paper. JD, MW, and DC all contributed to revising the paper critically for important intellectual content.
Funding
Research was supported by a National Science Foundation (USA) grant to MW (grant number 0818962). The National Science Foundation Graduate Research Fellowship Program and the Cornell University Student Research Grant in Animal Behavior provided additional grant support.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We thank S. Carouso, C. Mathers-Winn, and anonymous reviewers for helpful comments on previous versions of this manuscript and J. Kapoor for input on research design and acoustic analysis. R. Lachlan provided substantial technical support with acoustic analyses using software package Luscinia (Lachlan, 2015). T. Daniel and B. Congdon provided logistic support in the field, and a large number of excellent field assistants were instrumental in data collection. C. Giuliano, J. Houck, C. Mathers-Winn, L. Nietmann, and A. Potticary in particular helped with recording of in-nest calls and songs in the field. We also thank Frontiers in Ecology and Evolution for hosting this special issue and M. Hall and N. Langmore for organizing the issue and inviting us to contribute.
References
Akcay, C., Swift, R. J., Reed, V. A., and Dickinson, J. L. (2013). Vocal kin recognition in kin neighborhoods of western bluebirds. Behav. Ecol. 24, 898–905. doi: 10.1093/beheco/art018
Amo, L., Tomás, G., Parejo, D., and Avilés, J. M. (2014). Are female starlings able to recognize the scent of their offspring? PLoS ONE 9:e109505. doi: 10.1371/journal.pone.0109505
Baker, M. C., and Logue, D. M. (2003). Population differentiation in a complex bird sound: a comparison of three bioacoustical analysis procedures. Ethology 109, 223–242. doi: 10.1046/j.1439-0310.2003.00866.x
Brainard, M. S., and Doupe, A. J. (2002). What songbirds teach us about learning. Nature 417, 351–358. doi: 10.1038/417351a
Burnham, K. P., and Anderson, D. R. (2002). Model Selection and Multimodel Inference: A Practical Information-Theoretic Approach, 2nd Edn. New York, NY: Pringer-Verlag.
Casgrain, P., and Legendre, P. (2001). The R Package for Multivariate and Spatial Analysis, Version 4.0 d6 User's Manual. Département de Sciences Biologiques, Université de Montréal. Available online at: http://www.fas.umontreal.ca/BIOL/legendre
Caspers, B. A., Hoffman, J. I., Kohlmeier, P., Krüger, O., and Krause, E. T. (2013). Olfactory imprinting as a mechanism for nest odour recognition in zebra finches. Anim. Behav. 86, 85–90. doi: 10.1016/j.anbehav.2013.04.015
Clark, C. W., Marler, P., and Beaman, K. (1987). Quantitative analysis of animal vocal phonology: an application to swamp sparrow song. Ethology 76, 101–115. doi: 10.1111/j.1439-0310.1987.tb00676.x
Colombelli-Négrel, D., Hauber, M. E., and Kleindorfer, S. (2014). Prenatal learning in an Australian songbird: habituation and individual discrimination in superb fairy-wren embryos. Proc. Biol. Sci. 281:20141154. doi: 10.1098/rspb.2014.1154
Colombelli-Négrel, D., Hauber, M. E., Robertson, J., Sulloway, F. J., Hoi, H., Griggio, M., et al. (2012). Embryonic learning of vocal passwords in superb fairy-wrens reveals intruder cuckoo nestlings. Curr. Biol. 22, 2155–2160. doi: 10.1016/j.cub.2012.09.025
Colombelli-Négrel, D., Webster, M. S., Dowling, J. L., Hauber, M. E., and Kleindorfer, S. (2016). Vocal imitation of mother's calls by begging Red-backed Fairy-wren nestlings increases parental provisioning. Auk 133, 273–285. doi: 10.1642/AUK-15-162.1
Dowling, J., and Webster, M. S. (2016). An experimental test of duet function in a fairy-wren (Malurus) with moderate cuckoldry rates. Behav. Ecol. 2, 228–236. doi: 10.1093/beheco/arv144
Ellis, J. M. S. (2008). Decay of apparent individual distinctiveness in the begging calls of adult female white-throated magpie-jays. Condor 110, 648–657. doi: 10.1525/cond.2008.8592
Evans, C., and Kleindorfer, S. (2016). Superb fairy-wren (Malurus cyaneus) sons and daughters acquire song elements of mothers and social fathers. Front. Ecol. Evol. 4:9. doi: 10.3389/fevo.2016.00009
Greig, E. I., Taft, B. N., and Pruett-Jones, S. (2012). Sons learn songs from their social fathers in a cooperatively breeding bird. Proc. R. Soc. B Biol. Sci. 279, 3154–3160. doi: 10.1098/rspb.2011.2582
Halpin, Z. T. (1991). “Kin recognition cues of vertebrates,” in Kin Recognition, 1st Edn., ed P. G. Hepper (Cambridge: Cambridge University Press), 220–258. doi: 10.1017/CBO9780511525414.010
Holmes, W. G., and Sherman, P. W. (1983). Kin recognition in animals: the prevalence of nepotism among animals raises basic questions about how and why they distinguish relatives from unrelated individuals. Am. Sci. 71, 46–55.
Hurst, J. L., and Beynon, R. J. (2010). Making progress in genetic kin recognition among vertebrates. J. Biol. 9, 13. doi: 10.1186/jbiol221
Knörnschild, M., Nagy, M., Metz, M., Mayer, F., and von Helversen, O. (2012). Learned vocal group signatures in the polygynous bat Saccopteryx bilineata. Anim. Behav. 84, 761–769. doi: 10.1016/j.anbehav.2012.06.029
Kroodsma, D. E., and Pickert, R. (1980). Environmentally dependent sensitive periods for avian vocal learning. Nature 288, 477–479.
Lachlan, R. F. (2015). Luscinia: A Bioacoustics Analysis Computer Program. Version 2.0 [Computer program]. Available onlinde at: https://github.com/rflachlan/Luscinia/releases on 9th September 2015.
Langmore, N. E., Hunt, S., and Kilner, R. M. (2003). Escalation of a coevolutionary arms race through host rejection of brood parasitic young. Nature 422, 157–160. doi: 10.1038/nature01460
Marler, P. (1970). A comparative approach to vocal learning: song development in Zonotrichia leucophrys nuttalli. J. Comp. Physiol. Monogr. 71, 157–160
Marler, P. (1991). Song-learning behavior: the interface with neuroethology. Trends Neurosci. 14, 199–206. doi: 10.1016/0166-2236(91)90106-5
Mathevon, N., Koralek, A., Weldele, M., Glickman, S. E., and Theunissen, F. (2010). What the hyena's laugh tells: sex, age, dominance and individual signature in the giggling call of Crocuta crocuta. BMC Ecol. 10:9. doi: 10.1186/1472-6785-10-9
Matrosova, V. A., Volodin, I. A., and Volodina, E. V. (2009). Short-term and long-term individuality in speckled ground squirrel alarm calls. J. Mammal. 90, 158–166. doi: 10.1644/08-MAMM-A-032.1
Matrosova, V. A., Volodin, I. A., Volodina, E. V., Vasilieva, N. A., and Kochetkova, A. A. (2010). Between-year stability of individual alarm calls in the yellow ground squirrel Spermophilus fulvus. J. Mammal. 91, 620–627. doi: 10.1644/09-MAMM-A-143.1
McDonald, P. G. (2012). Cooperative bird differentiates between the calls of different individuals, even when vocalizations were from completely unfamiliar individuals. Biol. Lett. 8, 365–368. doi: 10.1098/rsbl.2011.1118
McDonald, P. G., and Wright, J. (2011). Bell miner provisioning calls are more similar among relatives and are used by helpers at the nest to bias their effort towards kin. Proc. R. Soc. B Biol. Sci. 278, 3403–3411. doi: 10.1098/rspb.2011.0307
Nelson, D. A., and Poesel, A. (2007). Segregation of information in a complex acoustic signal: individual and dialect identity in white-crowned sparrow song. Anim. Behav. 74, 1073–1084. doi: 10.1016/j.anbehav.2007.01.018
Podos, J., Huber, S. K., and Taft, B. (2004). BIRD SONG: the interface of evolution and mechanism. Annu. Rev. Ecol. Evol. Syst. 35, 55–87. doi: 10.1146/annurev.ecolsys.35.021103.105719
Porter, R. H. (1986). Chemical signals and kin recognition in spiny mice (Acomys cahirinus). Chem. Signals Vertebrates 4, 397–411. doi: 10.1007/978-1-4613-2235-1_29
Price, J. (1999). Recognition of family-specific calls in stripe-backed wrens. Anim. Behav. 57, 483–492. doi: 10.1006/anbe.1998.1018
Price, J. J. (1998). Family- and sex-specific vocal traditions in a cooperatively breeding songbird. Proc. Biol. Sci. 265, 497–502.
Radford, A. N. (2005). Group-specific vocal signatures and neighbour-stranger discrimination in the cooperatively breeding green woodhoopoe. Anim. Behav. 70, 1227–1234. doi: 10.1016/j.anbehav.2005.04.002
Rendall, D., Rodman, P. S., and Emond, R. E. (1996). Vocal recognition of individuals and kin in free-ranging rhesus monkeys. Anim. Behav. 51, 1007–1015. doi: 10.1006/anbe.1996.0103
Riehl, C., and Stern, C. A. (2015). How cooperatively breeding birds identify relatives and avoid incest: new insights into dispersal and kin recognition. Bioessays 37, 1303–1308. doi: 10.1002/bies.201500120
Schwabl, H., Dowling, J., Baldassarre, D. T., Gahr, M., Lindsay, W. R., and Webster, M. S. (2015). Variation in song system anatomy and androgen levels does not correspond to song characteristics in a tropical songbird. Anim. Behav. 104, 39–50. doi: 10.1016/j.anbehav.2015.03.006
Sharp, S. P., McGowan, A., Wood, M. J., and Hatchwell, B. J. (2005). Learned kin recognition cues in a social bird. Nature 434, 1127–1130. doi: 10.1038/nature03522
Slater, P. (1986). The cultural transmission of bird song. Trends Ecol. Evol. 1, 94–97. doi: 10.1016/0169-5347(86)90032-7
Slater, P. J. B., Jones, A., and Cate, T. C. (1992). Can lack of experience delay the end of the sensitive phase for song learning? Netherlands J. Zool. 43, 80–90.
Varian-Ramos, C. W., Karubian, J., Talbott, V., Tapia, I., and Webster, M. S. (2010). Offspring sex ratios reflect lack of repayment by auxiliary males in a cooperatively breeding passerine. Behav. Ecol. Sociobiol. 64, 967–977. doi: 10.1007/s00265-010-0912-5
Waldman, B. (1987). Mechanisms of kin recognition. J. Theor. Biol. 128, 159–185. doi: 10.1016/S0022-5193(87)80167-4
Keywords: kin signatures, female song, in-nest calls, embryonic learning, songbirds, Malurus melanocephalus
Citation: Dowling JL, Colombelli-Négrel D and Webster MS (2016) Kin Signatures Learned in the Egg? Red-Backed Fairy-Wren Songs Are Similar to Their Mother's In-Nest Calls and Songs. Front. Ecol. Evol. 4:48. doi: 10.3389/fevo.2016.00048
Received: 26 November 2015; Accepted: 18 April 2016;
Published: 06 May 2016.
Edited by:
Michelle L. Hall, University of Melbourne, AustraliaReviewed by:
Marion Linda East, Leibniz Institute for Zoo and Wildlife Research, GermanyKatharina Riebel, Leiden University, Netherlands
Copyright © 2016 Dowling, Colombelli-Négrel and Webster. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Jenélle L. Dowling, amxkMjc2QGNvcm5lbGwuZWR1