 Juliette Bailly1*
Juliette Bailly1* Bruno Faivre1
Bruno Faivre1 Nadine Bernard2
Nadine Bernard2 Mickaël Sage3Nadia Crini2Vincent Driget2
Mickaël Sage3Nadia Crini2Vincent Driget2 Stéphane Garnier1Dominique Rieffel2
Stéphane Garnier1Dominique Rieffel2 Renaud Scheifler2
Renaud Scheifler2- 1BioGéoSciences, UMR 6282, Université Bourgogne Franche-Comté/Centre National de la Recherche Scientifique, Dijon, France
- 2Chrono-Environment, UMR 6249 Université Bourgogne Franche-Comté/Centre National de la Recherche Scientifique Usc Institut National de la Recherche Agronomique, Besançon, France
- 3Wildlife, Environment, and Expertises, Besançon, France
Urbanization is a growing phenomenon characterized by a complete restructuring of natural areas. In urban bird populations, a reduced offspring survival and body condition and an overall lower breeding success are often observed compared to populations inhabiting more natural habitats. Higher pollution levels and poorer quality of natural resources in cities are two environmental factors frequently mentioned in the literature to explain the differences between urban and non-urban populations. Pollution and poor quality of food may lead to an excess of non-essential elements such as Pb or Cd or to deficiencies of essential elements such as Cu or Zn, which may explain some of the impacts, notably on immunity, observed in urbanization studies. The present study compared the breeding parameters, brood body mass and condition, and haptoglobin levels, a marker of inflammatory immunity, in two urban and two forest populations of Great tits in Eastern France, together with a multi-element analysis (25 non-essential and essential trace elements) of blood samples from 13-day-old nestlings from the four populations. The concentration of NO2, a gaseous pollutant typical of urban pollution, was also measured. The NO2 concentrations were significantly higher in the urban areas, but no association with biological variables was detected. Non-essential metals were undetectable in the plasma of the birds from both habitats, except Pb, whose concentrations, however, did not differ between the urban and forest birds. A positive relationship was found between the plasmatic richness in essential elements (as assessed from the coordinates of the first axis of a PCA including 12 elements) and the average brood body mass and condition. We suggest that lower quality resources or/and a higher metabolic demand may be a causal mechanism for the reduced body condition often observed in urban bird nestlings. Finally, our exploratory study could promote more mechanistic experiments (e.g., supplementation) to explain the negative effect of urban conditions on bird populations.
Introduction
Urbanization is increasing, and 54% of the world population now lives in urban areas (United Nations, 2014). Urban habitats have been colonized by many wild species, including birds (McKinney and Lockwood, 1999), which can reach high densities (Chace and Walsh, 2006). Many studies have suggested that urban conditions exert strong constraints on bird fitness (Chamberlain et al., 2009; Vaugoyeau et al., 2016). A number of breeding parameters are affected, with the most consistent patterns being earlier laying dates, lower clutch size, lower nestling weight or condition, and an overall lower productivity per nesting attempt in urban areas (Chamberlain et al., 2009) even if inter specific variations between those life history traits and the intensity of urbanization have recently been demonstrated (Vaugoyeau et al., 2016). Other individual parameters can be modified in urban bird populations compared to rural ones. For example, urban birds are often—but not always—smaller and in poorer condition (Liker et al., 2008; Evans et al., 2009a) and show higher oxidative stress and antioxidant activity than birds from more natural habitats (Isaksson et al., 2005).
Urban and natural environments differ strongly in many factors (Grimm et al., 2008). Temperature, predator and parasite communities, artificial noise and light, availability and/or quality of resources, and pollution are among important features that contrast between cities and natural habitat (Collier, 2006; Evans et al., 2009b; Slabbekoorn, 2013). Cities are frequently described as resource-poor habitats for birds during the breeding season, and several works have shown that prey required for chicks are less abundant, smaller and poorer in key nutrients for growth and survival, such as carotenoids (Isaksson and Andersson, 2007; Seress et al., 2012). Those descriptive studies provided some data that allowed the design of experimental supplementation aimed at testing whether the quantity (energy intake) or the quality of resources (e.g., carotenoids) could explain the contrast of breeding performance between urban and non-urban populations (Peach et al., 2014; Giraudeau et al., 2015). Several other key resources, such as essential elements, could be involved in the differences of growth, body condition, and/or breeding performance between rural and urban nestlings. For instance, calcium (Ca) is an essential micronutrient during egg formation and skeletal development of nestlings (Patten, 2007; Reynolds and Perrins, 2010). Supplementation experiments in wild populations of Great tits (Parus major) reported that Ca-supplemented birds produced larger brood and raised more nestlings of better body condition compared to control groups (Tilgar and Reynolds, 2005; Espín et al., 2016a). Other essential elements such as zinc (Zn) or copper (Cu) are also of great interest in the bird physiology and behavior. They affect enzymatic activity and antioxidant defenses (Powell, 2000; Sahin et al., 2005) and play an important role in embryo development and nestling growth (Park et al., 2004). Their concentration can exceptionally raise toxicity thresholds (Sundaresan et al., 2008), but, more likely for vertebrates that are very tolerant to these elements, they may impact physiological performances if their concentrations do not meet homeostatic requirements (deficiencies) (Keen et al., 1998). Despite the crucial role of essential elements in nestling growth, and in bird physiology more generally, very few studies have focused on the potential deficiencies in these elements in urbanization studies. Therefore, quantifying these elements in birds might allow the discrimination of urban and rural populations and the identification of deficiencies in urban individuals.
Similarly, the impact of pollution has received surprisingly little attention in urbanization studies, although cities are characterized by the circulation of many different pollutants in air, water, and soils. The air pollutants that draw the most concerns for ecosystem and human health include particulate matter (PM), ozone (O3), carbon monoxide (CO), sulfur dioxide (SO2), nitrogen dioxide (NO2), lead (Pb), volatile organic compounds (VOC), and polycyclic aromatic hydrocarbons (PAHs) (Mayer, 1999; Berback et al., 2001; Marr et al., 2004). Very few prior studies were dedicated to the effects of air pollutants on birds in the context of urbanization, while studies in (generally heavily) industrially contaminated sites are numerous. To our knowledge, only one study found that reduced reproductive success of House sparrows (Passer domesticus) along an urban gradient was associated with air pollution from traffic as assessed by NO2 concentrations in the air (Peach et al., 2008). Studies dealing with pollution by non-essential metal elements are more numerous, but most of them only report biomonitoring data. For instance, studies found higher concentrations of Pb in urban birds than in their rural counterparts with individuals exhibiting blood concentrations higher than the benchmark value related to subclinical and physiological effects in birds (Scheifler et al., 2006; Roux and Marra, 2007). Clearly, descriptive studies assessing elements in rural and urban birds may provide data about possible excesses of non-essential and potentially toxic elements such as Pb, arsenic (As), cadmium (Cd), or mercury (Hg) in urban birds compared to rural individuals, and may facilitate the design of more mechanistic experiments about the impacts of urbanization on birds.
Pollutants may interact with the immune system depending on the nature of the contamination (element or molecule involved, chronic vs. acute, high vs. low dose) with consequences on host immune profile. Components of inflammatory response showed both down-regulated or up-regulated activities in vertebrates exposed to contaminants (Tersago et al., 2004; Schiraldi and Monestier, 2009) with expected consequences on host-parasite interactions and host fitness (Sorci et al., 2013). However, how urban pollution may affect host immunity has been neglected, and the assessment of markers such as inflammatory markers can provide information on immune status of urban individuals. Recently, we observed contrasts in the production of haptoglobin, an acute protein associated to the inflammatory process, between urban and forest nestlings (Bailly et al., 2016a). Therefore, it could be of interest to explore how this inflammatory marker is affected by urban pollution.
In this study, we measured the concentrations of 25 non-essential and essential trace elements (TEs) in nestling plasma together with the breeding parameters, nestling body condition, and the baseline plasma levels of an inflammatory marker (haptoglobin) in Great tits from two medium-sized cities of Eastern France (Besançon and Dijon) and two forests from the surroundings of each city (Forêt de Chaux and Forêt d'Auxonne, respectively). The atmospheric concentrations of NO2, one of the main traffic-related pollutants and precursor forming photochemical smog (Han and Naeher, 2006), were also measured with passive air samplers in the studied areas. We hypothesized (i) contrasts between urban and rural populations in circulating TEs (with lower levels of essential elements and higher levels of non-essential elements in urban birds), breeding parameters (clutch size, hatching success, nestling survival, brood body mass and condition) and immunological marker (baseline level of circulating haptoglobin) with harsher conditions for urban birds, and (ii) correlations suggesting negative effects of non-essential TEs and positive effects of essential TEs. Comparison with rural organisms may bring crucial insights about the mechanisms underlying the urbanization process and open the route for more mechanistic and experimental approaches to test them.
Materials and Methods
Study Sites
Two cities (Besançon, 47°25 N, 6°03 E; Dijon, 47°32 N, 5°02 E) and two forests (Forêt de Chaux, 47°09 N, 5°68 E; Forêt d'Auxonne, 47°10 N, 5°26 E) were studied in two regions of Eastern France, Franche-Comté, and Burgundy. Besançon and Dijon are two middle-sized cities (176,764 and 244,577 inhabitants in 2008, respectively, and 432.3 and 219.3 km2, respectively) whose economies are dominated by tertiary activities (http://www.insee.fr). None of those cities have important industrial sources of metals like metallurgic plants for instance. The Forêt d'Auxonne and the Forêt de Chaux are deciduous forests of 7,800 and 20,493 ha, respectively. The most abundant tree species are oaks (Quercus petrae and Q. robur), beechwood (Fagus sylvatica) and charm (Carpinus sp.). Nest boxes were installed in medium- (50 years old) to old- (100 years old) growth stands at least 1 km from the edge of the forests. Each site received approximatively 150 artificial nestboxes (Nest box 1B with protective front panel, 12 cm diameter and 20 cm height, Schwegler, Germany). The nest boxes were hung from 0.5 m long steel tubes to protect the nests from predation. The nest boxes in the cities were dispersed in parks, squares, and along streets, with each patch containing one to 25 nest boxes separated by at least 50 m from each other. The vegetation of most of the area of the parks has been formed artificially. The tree cover is patchy with tree-free areas mainly covered by grass. The tree patches are composed of deciduous and coniferous species of different stands, including both native and exotic species. Each study site covers an average area measuring 5.6 km long and 2.8 km wide. In the forests, the nest boxes were placed along pathways to facilitate access, with the same minimal distance between two nest boxes as in the cities.
Reproductive Parameters and Blood Sampling
The reproduction of the Great tits was monitored from March to June 2013. Only the first breeding attempts were included in the study, second broods were not studied due to logistical constraints. The nests were inspected once a week from March 20th to record the laying date of the first egg. When more than one egg was found in the nest, the laying date of the first egg was back calculated, assuming that one egg was laid per day. Incubation was assumed to begin 1 day before clutch completion, and we did not observe asynchronous hatching (except sometimes for one egg). The nests were not visited again until the estimated hatching date (13 days after the beginning of incubation), and any eggs that had not hatched were then checked every 2 days until hatching. The number of nestlings was noted on day 1 (D1), 7 (D7), and 13 (D13) post-hatching. The body mass and tarsus length of the 13-day-old nestlings were measured using a Pesola spring balance (±0.05 g), and an electronic caliper (±0.1 mm). We defined clutch size as the number of eggs laid and hatching success was the proportion of eggs that hatched in the clutches where at least one egg hatched. We defined nestling survival as the proportion of hatched nestlings that were still alive on D13 in nests where at least one nestling survived until D13. When nestlings were 7 days old, they were fitted with individually numbered metal leg rings (Museum National d'Histoire Naturelle, CRBPO, Paris, France). Blood was sampled (50–100 μl) from one brachial vein of 13-day-old nestlings using sterile needles and heparinized capillary tubes. The blood samples were stored in a cool box until their return to the laboratory within 4 h of collection. Then, the blood was centrifuged (10 min, 4,000 rpm, 4°C), and the plasma was separated from the pellet and stored at −80°C until analysis.
Haptoglobin Assay
Haptoglobin (Hp) belongs to the category of acute phase proteins and is used as a marker to measure the intensity of the inflammatory response in vertebrates, including birds (Matson et al., 2006, 2012). The production of this protein by the liver is stimulated by activated inflammatory cytokines (Dobryszycka, 1997), so as the amount of plasma Hp increases, the inflammatory reaction becomes stronger. Here, we quantified the baseline level of this protein to assess the inflammatory status of the nestlings. Hp was measured in the plasma of 13-day-old nestlings using a commercial assay kit (TP-801, Tridelta Development LTD, Ireland). First, 7.5 μl of each sample were randomly aliquoted on a flat-bottomed, 96-well plate. Samples were distributed randomly among the plates. Standard curves were included (in duplicate) on each plate (n = 16) containing the plasma samples, and the curves consisted of dilutions of an initial standard of 2.5 mg/ml diluted five times to 0 mg/ml. One hundred microliters of a stock solution of hemoglobin (catalyser) and 140 μl of a stock solution of chromogen (coloration) were added to all wells. The plates were agitated to ensure the mixing of the samples with hemoglobin and chromogen and left to incubate for 5 min at room temperature. The absorbance of each well was read at 630 nm, and the intra-assay and inter-plate variations were, respectively, 2.4 and 2.99%. The samples above the detection threshold were diluted and assayed again.
NO2 Measurements
Passive NO2 samplers were put on 6–58 nest boxes in the four study sites during the nestling stage (6 and 7 in the two forests, 38 in Dijon, and 58 in Besançon). Not all nestboxes were equipped for both financial and logistical reasons. The samplers were set up vertically on the middle of the tube from which the nest boxes were hung, allowing air to circulate freely around them. Passive samplers (Palmes et al., 1977) are calibrated tubes, 7 cm long, with an inside diameter of 1 cm, in which gases move only by molecular diffusion (Gradko International, Winchester, Great Britain). A triethanolamine solution, which was deposited on the grid at one end of the tube, fixed the NO2. The other end of the tube remained open for gas diffusion during the sampling period. Each passive sampler allowed a measurement of the NO2 pollution level over a period of 14 days, optimal duration of sampling (Bernard et al., 1997). At 21.1°C and at a pressure of 1 atmosphere, the diffusion coefficient for NO2 is 0.154 cm2/s, which means that the collection rate for NO2 passive samplers could be calculated at 72 cm3/h. Mean hourly concentration of NO2 (expressed in μg/m3; 1 ppb = 1.91 μg/m3 at 21.1°C) in the air sample was calculated on the basis of the amount of pollutant collected, exposure time, and gas collection rate in the passive sampler. Absorbed NO2 was measured by spectrophotometry using a variant of the Griess-Saltzman method (Atkins, 1990). In an earlier study, NO2 measurements with passive samplers were validated on chemiluminescence analysers, equipment used by the French network of air quality monitoring and advocated by European legislation (Bernard et al., 1998).
Multi-Element Analysis
The concentrations of 25 TEs [aluminum (Al), antimony (Sb), As, Cd, Ca, chromium (Cr), cobalt (Co), Cu, tin (Sn), iron (Fe), magnesium (Mg), manganese (Mn), Hg, molybdenum (Mo), phosphorus (P), Pb, potassium (K), nickel (Ni), selenium (Se), silicon (Si), sodium (Na), strontium (Sr), titanium (Ti), thallium (Tl), and Zn] were determined in 93 composite (one sample per family) plasma samples. Four samples had volumes lower than 50 μl and could not been analyzed. Eight samples had a volume ranging from 55 to 85 μl. For the other samples, 100 μl were used. The samples were acid digested in 700 μl of nitric acid (HNO3, 65%, analytical quality, Optima) for half a day in open tubes under hood, and then for 60 h in closed tubes at 60°C in an oven. After digestion, samples were diluted to 30 ml by the addition of ultra-pure water (18.2 MΩ/cm2). Blanks (acid + ultra-pure water) and Certified Reference Materials (CRMs; lobster hepatopancreas, TORT-2, National Research Council Canada, and drinking water, ERM®-CA011b, European Reference Materials) were prepared and analyzed using the same methods as the samples. Elements were analyzed by Inductively Coupled Plasma—Atomic Emission Spectrometry (ICP-AES, ThermoFisher Scientific iCAP 6,000) or by Inductively Coupled Plasma—Mass Spectrometry (ICP-MS, ThermoFischer Scientific XSeries 2). Over the 25 elements measured, only 12 had a satisfactory proportion of values above quantification limits (Table S1), allowing for statistical analyses. For the values under quantification limits for these 12 elements, the value was replaced by the quantification limit divided by square root of two for statistical analysis (Helsel, 2010). Average recoveries of the CRMs ranged from 73 (for Fe) to 135% (for Pb) for the TORT-2, and from 78 (Fe) to 124% (for Se) for the ERM®-CA011b (Table S1). The concentrations of the elements in the plasma are expressed as μg/dl.
Statistical Analysis
Levels of atmospheric NO2, plasmatic TEs concentrations, and biological parameters were compared among the four study sites using the non-parametric Kruskal–Wallis test because the distribution of most of the data were skewed. When a significant effect was detected, post-hoc Tukey HSD tests were applied to detect differences among sites. Urban vs. forest differences were analyzed using Wilcoxon Mann–Whitney test. All NO2 data were used to compare NO2 levels among the four study sites while only the data related to nestboxes hosting successful reproduction were used to study the relationships between NO2 levels and biological parameters. Moreover, because NO2 concentrations in the forests were, as expected, very low and exhibited low variations, only data from the two cities were analyzed to test the relationships between the NO2 concentrations and biological parameters (clutch size, hatching success, nestling survival, brood body mass and condition, baseline level of circulating haptoglobin). Correlations among TEs concentrations and NO2 levels were studied using Spearman's correlations due to skewed distribution. A Principal Component Analysis (PCA) was conducted to investigate the relationships among TEs concentrations. The first axis of the PCA (PC1) was used as a plasma synthetic index describing richness in the essential elements and was used for further analysis with trace elements (see Section Results). Because we have only one measure of NO2 and TEs concentrations per brood, the average brood mass, tarsus length, and baseline levels of Hp per brood were used for further analysis. An average brood condition at D13 was estimated as the residuals of the linear regression between the log-transformed average brood mass and the log-transformed average brood tarsus. The correlations between richness in essential elements as assessed by PC1 and brood mass and condition were studied using Spearman's correlations because of the skewed distribution of the data.
Clutch size, hatching success, nestling survival, and brood mass, body condition, and Hp levels were analyzed using habitat and NO2 levels or PC1 (as an estimate of the plasmatic richness in essential elements) as explanatory variables, and some co-variates when needed (see below). The variation in clutch size was analyzed using generalized linear models (GLMs) with Poisson error distributions (Zuur et al., 2009). For hatching success and nestling survival, GLMs with binomial distributions and logit link functions were used (Warton and Hui, 2011), and the first egg-laying date (standardized as the number of days between March 1st and the first egg-laying date) was added as a covariate (Cresswell and McCleery, 2003; Blondel, 2007). Because overdispersion was detected, the standard errors were corrected using a quasi-GLM model (Zuur et al., 2009). Linear models (LMs) were used for brood mass and body condition at D13 with the first egg-laying date and brood size included as covariates. Because body mass may affect nestling immunity (Alonso-Alvarez and Tella, 2001), it was included as a covariate for the Hp analysis.
Each analysis was performed using the full model with software R v.3.15.1 (R Development Core Team, 2014). Statistical significance was set at p < 0.05 for all results.
Results
Reproduction Parameters and Immune Marker
Laying dates differed significantly among all groups except between Besançon and Dijon (KW Chi2 = 61.5, df = 3, p < 0.001, Table S2) and urban Great tits laid earlier than forest individuals, in average of 4.3 days (W = 6699, p < 0.001). The number of eggs laid did not differ neither between the two cities nor between the two forests (KW Chi2 = 90.0, df = 3, p < 0.001, Table S2). Urban Great tits, however, laid in average 2.7 eggs less than forest tits (W = 8153, p < 0.001). Nestlings were significantly heavier in the Forêt d'Auxonne than in the two cities but did not differ from the nestlings from the Forêt de Chaux (KW Chi2 = 43.3, df = 3, p < 0.001, Table S2). Urban broods were significantly and in average of 1.8 g lighter than forest broods (W = 2298, p < 0.001). Nestling body condition was the highest in the Forêt d'Auxonne and did not differ between the two cities (KW Chi2 = 56.9, df = 3, p < 0.001, Table S2). The condition was better for forest individuals than for urban ones (W = 2312, p < 0.001). Hatching success and nestling survival did not differ not among sites nor between the two habitats (Table S2). The baseline levels of circulating haptoglobin was higher in the Forêt de Chaux than in the other forest and in Dijon (KW Chi2 = 12.9, df = 3, p < 0.005, Table S2). There was no difference of haptoglobin levels between urban and forest individuals (W = 817, p = 0.3).
Relationships between NO2 and Biological Parameters
Nitrogen dioxide concentrations in those both middle-size urban areas were close to 20 μg/m3 (mean ± standard deviation: 19.6 ± 4.6 and 23.4 ± 10.5 μg/m3 in Dijon and Besançon, respectively, Figure 1) and those of forests were 7- to 10-fold lower (4.0 ± 0.9 and 1.8 ± 0.8 μg/m3 in the Forêt d'Auxonne and the Forêt de Chaux, respectively). Nitrogen dioxide concentrations did not differ neither between the two cities nor between the two forests (KW Chi2 = 33.6, df = 3, p < 0.001) while urban NO2 concentrations were significantly higher than the ones measured in the forests (W = 13, p < 0.001). No significant correlation was detected between the NO2 in urban areas and any of the biological parameters measured.
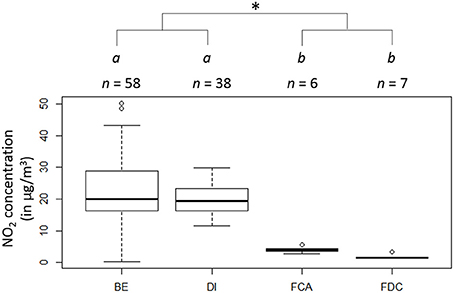
Figure 1. Atmospheric concentrations of NO2 (μg/m3) in the different study sites: Besançon (BE), Dijon (DI), the Forêt d'Auxonne (FCA), and the Forêt de Chaux (FDC). Sites with different letters are significantly different. The asterisk indicates a difference between urban and forest levels.
Relationships between Element Concentrations and Biological Parameters
Over the 25 elements analyzed in this study, 13 were detected in very few or even no birds, including the non-essential As, Cd, and Hg. Over the 12 remaining elements, Spearman's correlation showed significant but not very strong correlations (r < 0.5) among metals (Figure S1). The plasma concentrations of Ca (W = 1297, p = 0.002), Cu (W = 1416, p < 0.001), Fe (W = 1301, p = 0.018), P (W = 1332, p < 0.001), Si (W = 1355, p < 0.001), and Zn (W = 1173, p = 0.045) were lower in urban nestlings than in forest individuals, whereas the Mo concentrations were higher (W = 86, p < 0.001) in urban nestlings than in forest ones (Table S3). K, Mg, Na, Pb, and Se concentrations did not differ between the urban and forest populations of Great tits (ps > Table S3).
PCA analysis found that 74.4% of the variation was driven by five elements (Ca, Na, Mg, P, and Si, Figure S2). The first and second axes of the PCA explained 27 and 18% of the variation, respectively. Based on the results of the PCA, we used PC1 as an index of the richness in essential elements, with negative values indicating high concentrations of essential elements, and positive values indicating low concentrations. The richness of the essential elements of Great tits did not differ neither between the two cities nor between the two forest sites, but was significantly higher in the forests than in the cities (W = 265.5, p < 0.001, Figure 2). Brood body mass and body condition were negatively correlated with PC1 (Figure 3, Table 1), therefore positively correlated to the richness in essential elements.
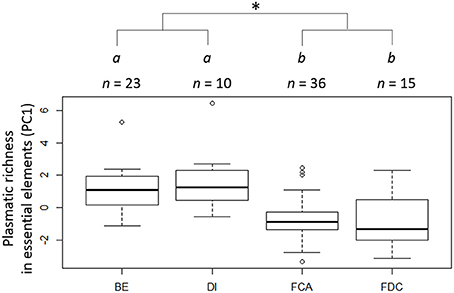
Figure 2. Richness in essential elements (as assessed by the coordinates on the first axis of a PCA: PC1, with negative values indicating high concentrations of essential elements, and positive values indicating low concentrations) in the plasma of Great tit 13-day old nestlings from the four study sites: Besançon (BE), Dijon (DI), the Forêt d'Auxonne (FCA), and the Forêt de Chaux (FDC). Sites with different letters are significantly different. The asterisk indicates a difference between urban and forest individuals.
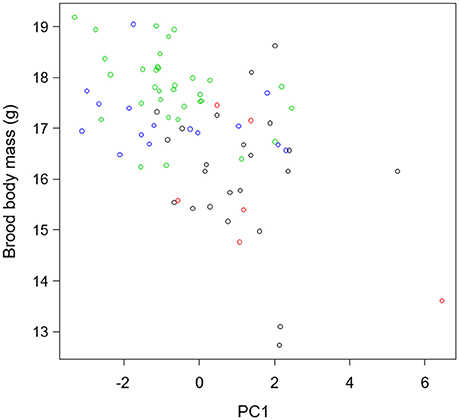
Figure 3. Relationship between richness in essential elements (PC1) and brood body mass of 13-day old nestlings of Great tit. Black circles: Besançon, red circles: Dijon, green circles: Forêt d'Auxonne, blue circles: Forêt de Chaux.
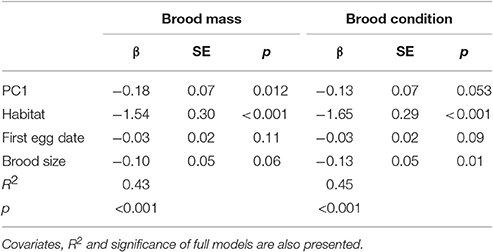
Table 1. Relationship between richness in essential elements (as assessed by the coordinates on the first axis of a PCA: PC1) and average brood mass and body condition of 13-day old nestlings of Great tits according to their habitats (urban vs. forest).
Discussion
Our results showed that several components of Great tit reproductive output were negatively affected in the urban habitats. The clutch size, nestling body mass, and nestling body condition were lower in urban sites than in forests. However, we observed no difference in the circulating haptoglobin. The essential elements richness was lower in urban sites and, interestingly, it appeared to be positively correlated with the nestling mass and body condition. Our results are consistent with those reported in previous studies (Hõrak, 1993; Chamberlain et al., 2009; Bailly et al., 2016b) on the components of reproductive output, and suggest mechanisms to explain the negative effect of urbanization on the reproductive performance of Great tits.
Among ecological factors that might explain the observed contrast in clutch size, brood mass and condition, food availability has frequently been cited in the literature (see for instance Chamberlain et al., 2009). The quantity and the quality of trophic resources strongly influence female investment in the clutch through its own condition (Marzal et al., 2005; Sofaer et al., 2012) as well as nestling growth and survival (Naef-Daenzer and Keller, 1999). The energetic requirement during growth is very high for tissue formation (for instance muscle, bone marrow) and the maintenance of those already established (Starck and Ricklefs, 1998). Several nutrients and elements are also essential for nestling development, and organisms have to obtain some of them from their food because they cannot synthetize these elements. For instance, Isaksson and Andersson (2007) showed that carotenoids, crucial compounds for several functions, are less abundant in prey consumed by urban than by forest Great tits. Similarly, in the surroundings of a metal-processing factory in Harjavalta, SW Finland, both the intensity of the yellow color in the plumage of P. major nestlings and caterpillar abundance increased with increasing distant from the pollution source, suggesting a deficiency of carotenoids in birds breeding close to the factory (Eeva et al., 1998). Here, the richness of essential elements was positively correlated with average brood mass and body condition, suggesting that deficiency in essential elements may be one of the mechanisms underlying the lower body condition often observed in urban nestlings compared to forest ones. According to the PCA, the variability of the dataset is mainly driven by the concentrations of 5 elements, namely Ca, Na, Mg, P, and Si. Ca is particularly important for the structural strength of the avian skeleton, plays vital roles in many biochemical reactions and allows birds to lay large megalecithal eggs (Dacke, 2000). Because of its fundamental role in bird physiology, Ca has received relatively higher attention in environmental studies than the other elements studied here. Indeed, among the abundant literature dealing with the effects of acidification or naturally base-poor habitats on bird reproduction (see for instance Graveland et al., 1994; Graveland, 1996). Mänd et al. (2000) found a positive effect of Ca supplementation on the tarsus length and body mass in fledgling Great tits growing in base-poor pine forests. This positive effect was observed only in the most unfavorable year (in terms of weather conditions) over the 3 years studied (Mänd et al., 2000). A similar positive effect of Ca supplementation was observed in P. major nestlings in a Ca-poor area associated with metal pollution in Harjavalta, SW Finland (Espín et al., 2016a). Eeva and Lehikoinen (2004) found reduced eggshell thickness, egg size, and hatchability, and delayed ossification of nestling leg and wing bones in Pied flycatchers (Ficedula hypoleuca) living close to a copper smelter in Finland where Ca availability is poor (Eeva and Lehikoinen, 2004). However, this effect was not found in the control Great tits studied in the same area. In our study, Ca was one the main essential elements explaining the variability of the TEs dataset and circulated at lower levels in urban nestling plasma, suggesting that urban birds suffer from reduced Ca-rich resources compared to the forest populations. Phosphorus deficiencies can lead to poor bone mineralization or even abnormal bone development such as chondrodystrophy (deformity of the leg bones) (Driver et al., 2006), and Na is an essential nutrient known to influence several aspects of normal animal growth. Sodium deficiency has been shown to reduce growth and food consumption, impair feed conversion water intake, acid-base balance, and basal metabolism (Vieira et al., 2003). Magnesium and Si are also considered as bone minerals even if the role of Si in bone health is still unclear (Jugdaohsingh, 2007). We are not aware of any study reporting Mg, Na, P, or Si deficiency effects on wild birds, but our results showed their lower levels in urban nestling plasma (two of them, P and Si being significantly less concentrated in urban nestling plasma).
Taken together, these results concerning so-called bone minerals, and the positive relationship between essential elements richness and body mass and condition, could explain the reduced growth found in urban nestlings. This may be linked to a lower quality, in terms of essential elements, of the resources that birds can forage in the cities compared to what is available in forest areas. An alternative hypothesis is that whatever the availability of these elements in urban habitat, the metabolic requirement of urban nestlings might be higher because of higher element consumption by organs. Indeed, stressful environmental conditions, as that prevailing in towns, are known to increase the organism demand for energetic and non-energetic resources.
Our study also shows that non-essential elements that are often considered as potentially toxic in urban or industrial areas are almost undetectable in the plasma of Great tits from the two middle size cities studied here. This may be linked to the fact that several metals have been shown to be mainly associated to erythrocytes rather than to plasma, as analyzed in the present work (Coeurdassier et al., 2012), but the lack of differences of the concentrations in the plasma of urban and forest birds suggests a low exposure to those elements in the two habitats. Pb, one of the non-essential elements for which there has been a great concern for wildlife conservation in urban and industrial areas, was detected in 78% of the broods studied but its concentrations did not differ between urban and forest birds. This suggests that this element, which has been banned from gasoline in most industrialized countries, tends to be less at risk for wildlife than it has been in the past for this category of middle-size and tertiary towns (see for instance Scheifler et al., 2006).
Atmospheric NO2 is a pollutant that is particularly relevant in the context of urbanization as it is emitted from the burning of fossil fuels for traffic and heating. All the average NO2 concentrations in the urban and forest study sites are below the thresholds fixed by French legislation (Code de l'environnement—Titre R22, 2013). They are also below the European hourly and annual average for urban background concentrations (European Environmental Agency, 2016), and below the values advised for health protection by the World Health Organization (2006). NO2 concentrations in Besançon and Dijon are in accordance with atmospheric levels of NO2 measured and modeled in middle-sized cities by Tenailleau et al. (2015). NO2 levels in both of these middle-sized urban areas are low as compared to those observed in large cities (Lebret et al., 2000). Therefore, these values indicate that Besançon and Dijon are moderately polluted area.
Urban populations of Great tits were exposed to higher levels of atmospheric NO2 than forest ones, but these concentrations were not linked with any component of the reproductive success measured in this study. This is not in agreement with the work of Peach et al. (2008) on the House sparrow, showing (for quite similar NO2 concentrations ranging from 15 to 35 μg/m3 in their study compared to 10–50 μg/m3 in the present work) that the brood body condition of 2- to 6-day-old nestlings and the brood tarsus length and body mass of 10- to 12-day-old nestlings were strongly and negatively correlated to local summer NO2 concentrations. The discrepancy between our study and this previous work may be due to a number of factors linked with environmental conditions, such as weather, and food availability, or with species-specific sensitivity to NO2.
Conclusion
Overall, our study brings new data about plasmatic concentrations of various essential elements in birds from urban and forest habitats and their potential effects on nestling body mass and condition. In our opinion, this is a supplementary clue that suggests food availability and/or quality may indeed be a causal mechanism for reduced growth of nestling birds in urban areas, an issue that needs further investigation. More specifically, the simultaneous assessment of circulating essential and non-essential elements allows for (i) design supplementation experiments to test how the availability of specific essential elements or blends of elements explains the reduced performances of birds in urban habitat or/and (ii) identify non-essential elements that may alter biological functions in urban birds. Our case study suggests that supplementation experiments with Ca, Cu, Fe, P, or Si should be relevant as a next step to analyze the negative consequences of urban habitat on the reproduction of Great tits. Several Ca-supplementation experiments have already been performed, in particular in the surroundings of the Cu smelter of Harjavalta, SW Finland (Espín et al., 2016a,b), but to our knowledge not in a context of urbanization without heavy industrial pollution sources. However, even though the present study did not show an effect of low concentrations of NO2 on any of the parameters studied and the concentrations of non-essential elements did not differ in the plasma of birds from the two habitats, the ecotoxicological dimension of the impacts of urbanization on birds deserves further attention because many common pollutants, and their interactions, have not yet been studied.
Ethics Statement
This study conforms to the legal requirements of France. The experiment has received the agreement of the Animal Care and Ethical Committee of the Université de Bourgogne, Dijon (protocol # 8112) and of the Préfectures from Côte d'Or, Doubs, and Jura (Arrêté # 448).
Author Contributions
BF, RS, and JB conceived the design of the study. JB, BF, MS, VD, SG, DR, and RS conducted the field work. JB, NB, and NC conducted the lab analyses. JB, BF, and RS analyzed the data and wrote the manuscript.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
The Regional Council of Burgundy and the Center for Research and Higher Education provided financial support for this work. This study was also funded by the Universities of Burgundy and Franche-Comté (“2012 BQR-PRES project n°38”) and the Ville de Dijon, de Besançon, d'Auxonne and the Office National des Forêts provided permissions to install nest boxes. The study site in Franche-Comté belongs to the CNRS long-term ecological research site “Zone Atelier Arc Jurassien” (http://zaaj.univ-fcomte.fr/?lang=en). We thank M. Coeurdassier who helped with the fieldwork.
Supplementary Material
The Supplementary Material for this article can be found online at: http://journal.frontiersin.org/article/10.3389/fevo.2017.00006/full#supplementary-material
References
Alonso-Alvarez, C., and Tella, J. L. (2001). Effects of experimental food restriction and body-mass changes on the avian T-cell-mediated immune response. Can. J. Zool. 79, 101–105. doi: 10.1139/z00-190
Atkins, D. H. F. (1990). “A passive diffusion tube sampler for the measurement of atmospheric NO2. A new approach,” in Physico-Chemical Behaviour of Atmospheric Pollutants: Air Pollution Research Reports, eds G. Restelli and G. Angeletti (Dordrecht: Springer Netherlands), 18–26. doi: 10.1007/978-94-009-0567-2_3
Bailly, J., Scheifler, R., Belvalette, M., Garnier, S., Boissier, E., Clément-Demange, V.-A., et al. (2016a). Negative impact of urban habitat on immunity in the Great tit Parus major. Oecologia 182, 1053–1062. doi: 10.1007/s00442-016-3730-2
Bailly, J., Scheifler, R., Berthe, S., Clément-Demange, V.-A., Leblond, M., Pasteur, B., et al. (2016b). From eggs to fledging: negative impact of urban habitat on reproduction in two tit species. J. Ornithol. 157, 377–392. doi: 10.1007/s10336-015-1293-3
Berback, B., Johansson, K., and Mohlander, U. (2001). Urban metal flows- A case study of Stockholm. Review and conclusions. Water Air Soil Pollut. 1, 3–24. doi: 10.1023/A:1017531532576
Bernard, N., Astre, C., Vuillot, B., Saintot, M., and Gerber, M. (1997). Measurement of background urban nitrogen dioxide pollution levels with passive samplers in Montpellier, France. J. Expo. Anal. Environ. Epidemiol. 7, 165–117.
Bernard, N., Saintot, M., Astre, C., and Gerber, M. (1998). Personal exposure to nitrogen dioxide pollution and effect on plasma antioxidants. Arch. Environ. Health 53, 122–128. doi: 10.1080/00039896.1998.10545973
Blondel, J. (2007). Coping with habitat heterogeneity: the story of Mediterranean Blue tits. J. Ornithol. 148, 3–15. doi: 10.1007/s10336-007-0161-1
Chace, J. F., and Walsh, J. J. (2006). Urban effects on native avifauna: a review. Landsc. Urban Plan. 74, 46–69. doi: 10.1016/j.landurbplan.2004.08.007
Chamberlain, D. E., Cannon, A. R., Toms, M. P., Leech, D. I., Hatchwell, B. J., and Gaston, K. J. (2009). Avian productivity in urban landscapes: a review and meta-analysis. Ibis 151, 1–18. doi: 10.1111/j.1474-919X.2008.00899.x
Coeurdassier, M., Fritsch, C., Faivre, B., Crini, N., and Scheifler, R. (2012). Partitioning of Cd and Pb in the blood of European Blackbirds (Turdus merula) from a smelter contaminated site and use for biomonitoring. Chemosphere 87, 1368–1373. doi: 10.1016/j.chemosphere.2012.02.031
Collier, C. G. (2006). The impact of urban areas on weather. Q. J. R. Meteorol. Soc. 132, 1–25. doi: 10.1256/qj.05.199
Cresswell, W., and McCleery, R. H. (2003). How great tits maintain synchronization of their hatch date with food supply in response to long-term variability in temperature. J. Anim. Ecol. 72, 356–366. doi: 10.1046/j.1365-2656.2003.00701.x
Dacke, G. C. (2000). “The parathyroids, calcitonium, and citamin D,” in Sturkie's Physiology, ed G. C. Whittow (San Diego, CA: Academinc Press), 472–485.
Dobryszycka, W. (1997). Biological functions of haptoglobin - new pieces to an old puzzle. Eur. J. Clin. Chem. Clin. Biochem. 35, 647–654.
Driver, J. P., Pesti, G. M., Bakalli, R. I., and Edwards, H. M. (2006). The effect of feeding calcium-and phosphorus-deficient diets to broiler chickens during the starting and growing-finishing phases on carcass quality. Poult. Sci. 85, 1939–1946. doi: 10.1093/ps/85.11.1939
Eeva, T., and Lehikoinen, E. (2004). Rich calcium availability diminishes heavy metal toxicity. Funct. Ecol. 18, 548–553. doi: 10.1111/j.0269-8463.2004.00875.x
Eeva, T., Lehikoinen, E., and Rönkä, M. (1998). Air pollution fades the plumage of the Great tit. Funct. Ecol. 12, 607–612. doi: 10.1046/j.1365-2435.1998.00221.x
Espín, S., Ruiz, S., Sánchez-virosta, P., and Eeva, T. (2016a). Effects of calcium supplementation on growth and biochemistry in two passerine species breeding in a Ca-poor and metal-polluted area. Environ. Sci. Pollut. Res. 23, 9809–9821. doi: 10.1007/s11356-016-6219-y
Espín, S., Ruiz, S., Sanchez-Virosta, P., Salminen, J.-P., and Eeva, T. (2016b). Effects of experimental calcium availability and anthropogenic metal pollution on eggshell characteristics and yolk carotenoid and vitamin levels in two passerine birds. Chemosphere 151, 189–201. doi: 10.1016/j.chemosphere.2016.02.074
European Environmental Agency (2016). Air Quality in Europe - 2016 Report. Denmark. doi: 10.2800/413142
Evans, K. L., Gaston, K. J., Sharp, S. P., McGowan, A., and Hatchwell, B. J. (2009a). The effect of urbanisation on avian morphology and latitudinal gradients in body size. Oikos 118, 251–259. doi: 10.1111/j.1600-0706.2008.17092.x
Evans, K. L., Gaston, K. J., Sharp, S. P., McGowan, A., Simeoni, M., and Hatchwell, B. J. (2009b). Effects of urbanisation on disease prevalence and age structure in Blackbird Turdus merula populations. Oikos 118, 774–782. doi: 10.1111/j.1600-0706.2008.17226.x
Giraudeau, M., Chavez, A., Toomey, M. B., and McGraw, K. J. (2015). Effects of carotenoid supplementation and oxidative challenges on physiological parameters and carotenoid-based coloration in an urbanization context. Behav. Ecol. Sociobiol. 69, 957–970. doi: 10.1007/s00265-015-1908-y
Graveland, J. (1996). Avian eggshell formation in calcium-rich and calcium-poor habitats: importance of snail shells and anthropogenic calcium sources. Can. J. Zool. 74, 1035–1044. doi: 10.1139/z96-115
Graveland, J., van der Wal, R., van Balen, J. H., and van Noordwijk, A. J. (1994). Poor reproduction in forest passerine from decline of snail abundance on acidified soils. Nature 368, 446–448. doi: 10.1038/368446a0
Grimm, N. B., Faeth, S. H., Golubiewski, N. E., Redman, C. L., Wu, J., Bai, X., et al. (2008). Global change and the ecology of cities. Science 319, 756–760. doi: 10.1126/science.1150195
Han, X., and Naeher, L. P. (2006). A review of traffic-related air pollution exposure assessment studies in the developing world. Environ. Int. 32, 106–120. doi: 10.1016/j.envint.2005.05.020
Helsel, D. R. (2010). Summing nondetects: incorporating low-level contaminants in risk assessment. Integr. Environ. Assess. Manag. 6, 361–366. doi: 10.1002/ieam.31
Isaksson, C., and Andersson, S. (2007). Carotenoid diet and nestling provisioning in urban and rural Great tits Parus major. J. Avian Biol. 38, 564–572. doi: 10.1111/j.0908-8857.2007.04030.x
Isaksson, C., Örnborg, J., Stephensen, E., and Andersson, S. (2005). Plasma glutathione and carotenoid coloration as potential biomarkers of environmental stress in Great tits. Ecohealth 2, 138–146. doi: 10.1007/s10393-005-3869-5
Keen, C. L., Uriu-Hare, J. Y., Hawk, S. N., Jankowski, M. A., Daston, G. P., Kwik-Uribe, C. L., et al. (1998). Effect of copper deficiency on prenatal development and pregnancy outcome. Am. J. Clin. Nutr. 67, 1003S–1011S.
Lebret, E., Briggs, D., Van Reeuwijk, H., Fischer, P., Smallbone, K., Harssema, H., et al. (2000). Small area variations in ambient NO2 concentrations in four European areas. Atmos. Environ. 34, 177–185. doi: 10.1016/S1352-2310(99)00292-7
Liker, A., Papp, Z., Bókony, V., and Lendvai, A. Z. (2008). Lean birds in the city: body size and condition of House sparrows along the urbanization gradient. J. Anim. Ecol. 77, 789–795. doi: 10.1111/j.1365-2656.2008.01402.x
Mänd, R., Tilgar, V., and Leivits, A. (2000). Reproductive response of Great tits, Parus major, in a naturally base-poor forest habitat to calcium supplementation. Can. J. Zool. 78, 689–695. doi: 10.1139/z99-255
Marr, L. C., Grogan, L. A., Wohrnschimmel, H., Molina, L. T., Molina, M. J., Smith, T. J., et al. (2004). Vehicle traffic as a source of particulate polycyclic aromatic hydrocarbon exposure in the Mexico city metropolitan area. Environ. Sci. Technol. 38, 2584–2592. doi: 10.1021/es034962s
Marzal, A., de Lope, F., Navarro, C., and Møller, A. P. (2005). Malarial parasites decrease reproductive success: an experimental study in a passerine bird. Oecologia 142, 541–545. doi: 10.1007/s00442-004-1757-2
Matson, K. D., Cohen, A. A., Klasing, K. C., Ricklefs, R. E., and Scheuerlein, A. (2006). No simple answers for ecological immunology: relationships among immune indices at the individual level break down at the species level in waterfowl. Proc. R. Soc. B Biol. Sci. 273, 815–822. doi: 10.1098/rspb.2005.3376
Matson, K. D., Horrocks, N. P. C., Versteegh, M. A., and Tieleman, B. I. (2012). Baseline haptoglobin concentrations are repeatable and predictive of certain aspects of a subsequent experimentally-induced inflammatory response. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 162, 7–15. doi: 10.1016/j.cbpa.2012.01.010
Mayer, H. (1999). Air pollution in cities. Atmos. Environ. 33, 4029–4037. doi: 10.1016/S1352-2310(99)00144-2
McKinney, M., and Lockwood, J. (1999). Biotic homogenization: a few winners replacing many losers in the next mass extinction. Trends Ecol. Evol. 14, 450–453. doi: 10.1016/S0169-5347(99)01679-1
Naef-Daenzer, B., and Keller, L. F. (1999). The foraging performance of Great and Blue tits (Parus major and P. caeruleus) in relation to caterpillar development, and its consequences for nestling growth and fledging weight. J. Anim. Ecol. 68, 708–718. doi: 10.1046/j.1365-2656.1999.00318.x
Palmes, E. D., Tomczyk, C., and DiMattio, J. (1977). Average NO2 concentrations in dwellings with gas or electric stoves. Atmos. Environ. (1967) 11, 869–872. doi: 10.1016/0004-6981(77)90061-0
Park, S. Y., Birkhold, S. G., Kubena, L. F., Nisbet, D. J., and Ricke, S. C. (2004). Review on the role of dietary zinc in poultry nutrition, immunity, and reproduction. Biol. Trace Elem. Res. 101, 147–163. doi: 10.1385/BTER:101:2:147
Patten, M. A. (2007). Geographic variation in calcium and clutch size. J. Avian Biol. 38, 637–643. doi: 10.1111/j.2007.0908-8857.04203.x
Peach, W. J., Sheehan, D. K., and Kirby, W. B. (2014). Supplementary feeding of mealworms enhances reproductive success in garden nesting House sparrows Passer domesticus. Bird Study 61, 378–385. doi: 10.1080/00063657.2014.918577
Peach, W. J., Vincent, K. E., Fowler, J. A., and Grice, P. V. (2008). Reproductive success of House sparrows along an urban gradient. Anim. Conserv. 11, 493–503. doi: 10.1111/j.1469-1795.2008.00209.x
R Development Core Team (2014). R: A Language and Environment for Statistical Computing. Vienna: R Foundation for Statistical Computing. Available online at: http://www.R-project.org
Reynolds, S. J., and Perrins, C. M. (2010). “Dietary calcium availability and reproduction in birds,” in Current Ornithology, ed C. F. Thompson (New York, NY: Springer), 31–74.
Roux, K. E., and Marra, P. P. (2007). The presence and impact of environmental lead in passerine birds along an urban to rural land use gradient. Arch. Environ. Contam. Toxicol. 53, 261–268. doi: 10.1007/s00244-006-0174-4
Sahin, K., Smith, M. O., Onderci, M., Sahin, N., Gursu, M. F., and Kucuk, O. (2005). Supplementation of zinc from organic or inorganic source improves performance and antioxidant status of heat-distressed quail. Poult. Sci. 84, 882–887. doi: 10.1093/ps/84.6.882
Scheifler, R., Curdassier, M., Morilhat, C., Bernard, N., Faivre, B., Flicoteaux, P., et al. (2006). Lead concentrations in feathers and blood of common Blackbirds (Turdus merula) and in earthworms inhabiting unpolluted and moderately polluted urban areas. Sci. Total Environ. 371, 197–205. doi: 10.1016/j.scitotenv.2006.09.011
Schiraldi, M., and Monestier, M. (2009). How can a chemical element elicit complex immunopathology ? Lessons from mercury-induced autoimmunity. Trends Immunol. 732, 1–8. doi: 10.1016/j.it.2009.07.005
Seress, G., Bókony, V., Pipoly, I., Szép, T., Nagy, K., and Liker, A. (2012). Urbanization, nestling growth and reproductive success in a moderately declining House sparrow population. J. Avian Biol. 43, 403–414. doi: 10.1111/j.1600-048X.2012.05527.x
Slabbekoorn, H. (2013). Songs of the city: noise-dependent spectral plasticity in the acoustic phenotype of urban birds. Anim. Behav. 85, 1089–1099. doi: 10.1016/j.anbehav.2013.01.021
Sofaer, H. R., Sillett, T. S., Peluc, S. I., Morrison, S. A., and Ghalambor, C. K. (2012). Differential effects of food availability and nest predation risk on avian reproductive strategies. Behav. Ecol. 24, 698–707. doi: 10.1093/beheco/ars212
Sorci, G., Cornet, S., and Faivre, B. (2013). Immunity and the emergence of virulent pathogens. Infect. Genet. Evol. 16, 441–446. doi: 10.1016/j.meegid.2012.12.031
Starck, J. M., and Ricklefs, R. E. (1998). Avian Growth and Development: Evolution within the Altricial-Precocial Spectrum. New York, NY: Oxford University Press.
Sundaresan, N. R., Anish, D., Sastry, K. V. H., Saxena, V. K., Nagarajan, K., Subramani, J., et al. (2008). High doses of dietary zinc induce cytokines, chemokines, and apoptosis in reproductive tissues during regression. Cell Tissue Res. 332, 543–554. doi: 10.1007/s00441-008-0599-3
Tenailleau, Q. M., Mauny, F., Joly, D., François, S., and Bernard, N. (2015). Air pollution in moderately polluted urban areas: how does the definition of ‘neighborhood’ impact exposure assessment? Environ. Pollut. 206, 437–448. doi: 10.1016/j.envpol.2015.07.021
Tersago, K., Coen, W., De Scheirs, J., Vermeulen, K., Blust, R., and Verhagen, R. (2004). Immunotoxicology in wood mice along a heavy metal pollution gradient. Environ. Pollut. 132, 385–394. doi: 10.1016/j.envpol.2004.05.029
Tilgar, V., and Reynolds, S. J. (2005). Chick development in free-living Great tits Parus major in relation to calcium availability and egg composition. Physiol. Biochem. Zool. 78, 590–598. doi: 10.1086/430690
United Nations (2014). World Urbanization Prospects: The 2014 Revision, Highlights (ST/ESA/SER.A/352). Department of Economic and Social Affairs, Population Division (2014).
Vaugoyeau, M., Adriaensen, F., Artemyev, A., Banbura, J., Barba, E., Biard, C., et al. (2016). Interspecific variation in the relationship between clutch size, laying date and intensity of urbanization in four species of hole-nesting birds. Ecol. Evol. 6, 5907–5920. doi: 10.1002/ece3.2335
Vieira, S. L., Penz, A. M., Pophal, S., and de Almeida, J. G. (2003). Sodium requirements for the first seven days in broiler chicks. J. Appl. Poult. Res. 12, 362–370. doi: 10.1093/japr/12.3.362
Warton, D., and Hui, F. (2011). The arcsine is asinine: the analysis of proportions in ecology. Ecology 92, 3–10. doi: 10.1890/10-0340.1
Keywords: urban ecology, pollution, Parus major, body condition, NO2
Citation: Bailly J, Faivre B, Bernard N, Sage M, Crini N, Driget V, Garnier S, Rieffel D and Scheifler R (2017) Multi-Element Analysis of Blood Samples in a Passerine Species: Excesses and Deficiencies of Trace Elements in an Urbanization Study. Front. Ecol. Evol. 5:6. doi: 10.3389/fevo.2017.00006
Received: 22 October 2016; Accepted: 03 February 2017;
Published: 24 February 2017.
Edited by:
Caroline Isaksson, Lund University, SwedenReviewed by:
Rafael Mateo, Instituto de Investigación en Recursos Cinegéticos, (CSIC-UCLM), SpainTapio Eeva, University of Turku, Finland
Copyright © 2017 Bailly, Faivre, Bernard, Sage, Crini, Driget, Garnier, Rieffel and Scheifler. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Juliette Bailly, YmFpbGx5anVsaWV0dGVAZnJlZS5mcg==