 Juliana Chacón
Juliana Chacón Federico Luebert
Federico Luebert Maximilian Weigend
Maximilian Weigend- 1Nees-Institut für Biodiversität der Pflanzen, Fachgruppe Biologie, Universität Bonn, Bonn, Germany
- 2Departamento de Silvicultura y Conservación de la Naturaleza, Universidad de Chile, Santiago, Chile
Long-distance dispersal seems to be the main biogeographic event responsible for intriguing distribution patterns in plant groups in which sister taxa are separated by thousands of kilometers of distance across oceans and continents. The biotic and abiotic mechanisms behind such dispersal events are poorly understood and many attempts have been made to explain how plants can manage to disperse and survive these long journeys. The biogeographic history of Boraginaceae, a subcosmopolitan plant family with many disjunct clades, is here addressed and analyzed in the context of the different dispersal modes exhibited by the species. The lack of a clear pattern between the main dispersal events in Boraginaceae and the phylogenetic distribution of the dispersal modes indicates that no single dispersal mechanism can be associated with the events of dispersal in the family. Moreover, adaptations to different dispersal agents and unassisted dispersal modes in some clades might have promoted the diversification of Boraginaceae in various habitats across several continents. Our study reveals that long-distance dispersal is a very complex process that needs to be analyzed in the context of climatic and environmental changes and the response of plants and their dispersal vectors to these variable conditions.
Introduction
Disjunctive distribution patterns in many angiosperms are the result of rare long-distance dispersal (LDD) events that have taken place many times during evolution (e.g., Renner et al., 2000; Christenhusz and Chase, 2013; Chacón and Renner, 2014; Berger et al., 2016). The unpredictability and complexity of such events, which imply not only the efficient dispersal of diaspores (seeds, fruits, plant parts) by means of biotic or abiotic vectors, but also successful establishment and survival, depending on a range of factors, makes them very difficult to test (Nathan, 2006; Gillespie et al., 2012). In spite of this difficulty, LDD has become the most plausible explanation to many disjunctive plant distributions, especially for clades that have diversified after the Cretaceous continental break-up (de Queiroz, 2005; McGlone, 2005; Sanmartín et al., 2007). The identification of the mechanisms leading to the successful transport of diaspores over long distances is key for disentangling the biological basis behind the geographic disjunctions in angiosperms. There are many studies on the subject some of them suggesting that animal mediated dispersal (broadly epizoochory and endozoochory) is more effective than wind or water dispersal, because the likelihood of reaching a comparable suitable habitat is larger (Van der Pijl, 1982; Herrera, 1989). Although this might be true for short-distance dispersals taking place at a local scale, this has been considered as the evidence of a selection process toward long-distance dispersal (Ronce, 2007). At a larger geographical and temporal scale it has been proposed that “nonstandard dispersal vectors” (i.e., vectors different than the ones inferred from the morphological characteristics of the plants) can dominate LDD, even if only small fraction of all seeds are dispersed that way (Berg, 1983; Higgins et al., 2003; Heleno and Vargas, 2015). Moreover, angiosperm groups with both biotic and abiotic dispersal modes (here referred to as polychory) appear to have more species than those with only one type of dispersal (Ricklefs and Renner, 1994). A recent study showed that the shifts between abiotic and biotic dispersal modes were apparently responsible for the differences in geographic range size and species richness in the Fagales (Larson-Johnson, 2016). On the other hand, unspecialized diaspores lacking dispersal syndromes have apparently been successful in colonizing the Galápagos Islands (Vargas et al., 2012). Although this kind of relationship has been shown before (e.g., Edwards and Westoby, 1996) it is still not well understood if the dispersal modes in angiosperms could be related to the incidence of long-distance dispersal or to the biogeographic patterns of a given clade.
The family Boraginaceae is a suitable study group to address this topic, because the phylogeny is well resolved and the biology of different life history traits is very well documented. Boraginaceae has ca. 1,700 species distributed worldwide and is mostly diverse in temperate regions of the northern hemisphere (Weigend et al., 2013, 2016; Chacón et al., 2016). Initial diversification occurred about 50 million years ago (Luebert et al., 2017). Intercontinental geographic disjunctions probably resulting from LDD events are observed in almost all Boraginaceae clades (Chacón et al., 2016; Weigend et al., 2016 and references therein). Interestingly, a wide range of biotic and abiotic dispersal modes, which may be related to the geographic distribution patterns of some clades (Cohen, 2014), are present in the family. The typical borage fruit consist of four mericarpids or nutlets that are dispersed separately. However, in some species the complete fruit or even parts of the plant can function as dispersal unit (Weigend et al., 2013). At least seven dispersal modes have been reported for Boraginaceae: Animal mediated dispersal, with seeds or fruits carried on the outer surface of animals (epizoochory), inside the gizzard and stomach of animals after ingestion (endozoochory), or carried by ants (myrmecochory); wind-mediated dispersal (anemochory); water-mediated dispersal (hydrochory); and unassisted dispersal in which diaspores fall just because of their weight (barochory; Van der Pijl, 1982). Atelochory or the avoidance of dispersal (therefore also known as “achory,” Van der Pijl, 1982) has also been reported in some species as explained below. Most Boraginaceae species are biotically dispersed and epizoochory is probably the single most important mechanism, especially in the Cynoglossoideae, which have glochidiate nutlets that can easily attach to the feathers and fur of animals (Selvi et al., 2011; Weigend et al., 2016). Some further examples of epizoochory are found in the genera Lappula (Ridley, 1930) and Cryptantha, the latter possessing small nutlets and rough sculpturing, which are easily detached from the plant (Carlquist, 1983; Grau, 1983; Hasenstab-Lehman and Simpson, 2012). Endozoochory is typical in Lithospermum L., whose stone-like nutlets are swallowed by granivorous birds, passing intact through their guts and germinating successfully later on (Van der Pijl, 1982; Best and Smartt, 1986; Bricker et al., 2010; Selvi et al., 2011; Weigend et al., 2016). In this genus the occurrence of barochory has also been shown (Selvi et al., 2011). Adaptations to myrmecochory have presumably played an important role in speciation of the Boragineae, where the species are characterized by the presence of oil-rich appendages (elaiosomes) that are nutritive for the ants (Van der Pijl, 1982; Pemberton and Irving, 1990; Quilichini and Debussche, 2001; Selvi et al., 2011). Both endozoochory and epizoochory have been reported in Echium, and even anemochorous and myrmecochorous species have been observed (Ridley, 1930; Parsons and Cuthbertson, 2001). Wind-dispersed nutlets are known in Paracaryum, Rindera, and Omphalodes (Selvi et al., 2011; Weigend et al., 2016). According to Van der Pijl (1982). Mysotis scorpioides L. is a typical nautohydrochorous as the nutlets are waterproofed and can float on the surface of water. Atelechory is presumably the dispersal mode characterizing the species of Craniospermum, which grow in dry steppes with poor soils in Eastern Asia (Ovchinnikova and Korolyuk, 2016). A detailed list of the main dispersal modes in Boraginaceae is presented in the Supplementary Table 1.
We aim to test the hypothesis that LDD events in Boraginaceae have been more frequent in clades exhibiting both biotic and abiotic dispersal modes, following the premise that LDD is expected to have occurred in clades with polychory. In order to study the biogeographic history of Boraginaceae the node ages and ancestral ranges of the major clades were inferred. The main biogeographic dispersal events that occurred during the evolution of the family were estimated using current biogeographic models that take into account the time-calibrated phylogeny and the geographic distribution of species. Ancestral dispersal modes and shifts in dispersal mode in Boraginaceae are inferred and compared with the biogeographic results. The role that different diaspore dispersal mechanisms could have played in the diversification of Boraginaceae is discussed.
Materials and Methods
Taxon Sampling and DNA Sequencing
The dating analyses were conducted for a dataset consisting of 284 species (287 taxa) including 183 species of Boraginaceae representing 72 of 90 genera from all tribes and subtribes proposed in Chacón et al. (2016). Our sampling includes a subset of the taxa analyzed in that study as multiple accessions of the same species were excluded, except in three cases in which the species where non-monophyletic [i.e., Bothriospermum tenellum (Hornem.) Fisch. and C.A.Mey., Caccinia macranthera (Banks and Sol.) Brand, and Cynoglossum montanum L.]. The ingroup sampling was also expanded to better represent the geographic range of widely distributed taxa or taxa that exhibit geographic disjunctions. Also clades in which the fossil constraints were placed were expanded (see next section). Therefore, the 16 following species were newly sequenced: Antiphytum cf. heliotropioides DC., Lithospermum californicum A.Gray, L. caroliniense (Walt. ex J.F. Gmel.) MacM., L. cinereum DC., L. cobrense E.L. Greene, L. erythrorhizon Siebold and Zucc., L. gayanum (Wedd.) I.M. Johnst., L. incisum Lehm., L. papillosum Thunb. L. parksii I.M.Johnst. L. scabrum Thunb., and L. virginianum L., Moritzia dasyantha Fresen., M. dusenii I.M. Johnst., and M. tetraquetra Brand, and Ogastemma pusillum (Coss. and Dur. ex Bonnet and Barratte) Brummitt. The outgroup consisted of 60 species belonging to the Boraginales families Codonaceae (the two species currently recognized), Cordiaceae s.l. (5 out of ca. 350 species), Ehretiaceae s.l. (17 out of ca. 170 species), Heliotropiaceae (27 out of ca. 450 species), Hydrophyllaceae (6 out of 250 species), Lennoaceae (1 out of 4 species), and Wellstediaceae (2 out of 6 species), following Weigend et al. (2016) and Luebert et al. (2016). Additional 41 species of the asterid orders Apiales, Aquifoliales, Asterales, Cornales, Dipsacales, Ericales, Gentianales, Lamiales, Solanales, and Vahliales were also added to the outgroup following Stevens (2001 onward) and Weigend et al. (2014).
All DNA sequences were obtained in the molecular lab of the Nees Institute for Biodiversity of Plants, University of Bonn, except for most asterid outgroups, which were downloaded from GenBank. The DNA extraction and amplification was done using standard protocols and universal primers as explained in Chacón et al. (2016). The chloroplast DNA regions tRNA-Leu (trnL) gene and trnL-trnF intergenic spacer (trnL-F), rps16 and trnS-trnG (trnS-G) were sequenced. All material sampled, including GenBank numbers and voucher information are listed in Supplementary Table 2.
Molecular Clock Analyses
The DNA sequences were assembled in Geneious v. 8.1.2 (Kearse et al., 2012) and aligned in MAFFT v. 7 (Katoh and Standley, 2013) using the FFT-NS-i option, which is a highly accurate iterative refinement method for fast multiple sequence alignment (Katoh et al., 2002). The resulting matrices were used to infer the ages of the main Boraginaceae clades by means of the Bayesian Markov chain Monte Carlo (MCMC) method implemented in the program BEAST v1.8.3 (Drummond et al., 2012). A partitioned analysis was specified in BEAUti v.1.8.3 (Drummond et al., 2012) by unlinking the substitution models and the clock models for each individual data partition. The GTR+G site model was used for each DNA partition according to the results of FindModel (available at: http://www.hiv.lanl.gov/content/sequence/findmodel/findmodel.html), which is a web implementation of the program Modeltest, using the Akaike information criterion (Posada and Crandall, 1998). A lognormal relaxed clock model using a random starting tree and a speciation Yule Process tree prior were selected in BEAUti. Our calibration scheme is based on the results of Luebert et al. (2017), who found better support for the chronograms obtained with seven fossil priors analyzed simultaneously. Therefore, we used the same priors to calibrate (i) two nodes in Boraginaceae (Cryptantha Lehm. ex Fisch. and C.A.Mey and Ogastemma Brummitt), one in Heliotropiaceae (Tournefortia L.), (ii) one node in Ehretiaceae (Ehretia P.Brow), and (iii) three nodes in other asterid clades (Ericales, Cornales, and Asterales). Additional fossil nutlets of Lappula Moench, Lithospermum, and Moritzia DC. ex Meisn were also included (see below). In summary, our dating scheme consisted of 10 calibration points that were applied to the following nodes of the phylogeny, using lognormal prior distributions for all the fossil constraints:
1. Boraginaceae (Cynoglossoideae, subtribe Amsinckiinae): The stem node of Cryptantha (incl. Cryptantha spp., Plagiobothrys Fisch., and C.A.Mey. spp. and Amsinckia Lehm. spp.) was set a minimum age of 10.3 Ma [lognormal mean (logMean): 0.5, lognormal standard deviation (logStdev): 1.0, median age: 11.95 Ma]. These settings are based on the oldest age of Cryptantha-like fossils reported for the upper Miocene of the Ogallala formation in the United States (ca. 13.6–10.3 Ma; Segal, 1966a; Thomasson, 1987; Gabel et al., 1998). Morphological affinities of this fossil with the genus Oreocarya Greene have been suggested (M. Simpson, personal communication, May, 2016). Our fossil placement is conservative, because the relationships between Oreocarya and the Amsinskiinae genera Plagiobothrys, Amsinckia, Dasynotus I.M.Johnst., and Pectocarya DC. Ex Meisn. are not fully resolved (Hasenstab-Lehman and Simpson, 2012; Ripma et al., 2014) and we therefore use the fossil to date the entire crown-clade.
2. Boraginaceae (Cynoglossoideae, subtribe Eritrichiinae): The crown node of Lappula (incl. Lappula and Rochelia spp.) was determined exactly in the same way as described for Cryptantha, based on fossil nutlets of Prolappula verrucosa Thomasson, that were found in the same Miocene macroflora of the Ogalalla formation (Thomasson, 1979; Gabel et al., 1998).
3. Boraginaceae (Boraginoideae, tribe Lithospermeae): Fossils fruits of Biorbia Elias, Prolithospermum Elias, Eliasiana Thomasson, and Lithospermum have been reported for the upper Miocene of the Ogallala formation in United States (e.g., Segal, 1966b; Thomasson, 1977, 1979, 1987; Gabel, 1987; Gabel et al., 1998) and can be assigned to the crown node of the latter genus. The oldest fossils of this group have been assigned to the Clarendonian (ca. 12–9 Ma; Thomasson, 1987; Gabel et al., 1998); therefore the crown node of Lithospermum was calibrated in the same way as described for the previous two nodes.
4. Boraginaceae (Boraginoideae, tribe Boragineae): The stem node of Moritzia was set the same priors as described for Cryptantha as these fossils were also reported in the Ogalalla formation (Thomasson, 1979; Gabel et al., 1998). In this case the stem node of Moritzia was constrained following Weigend et al. (2010), who suggest the close resemblance of the fossils with the Boragineae genera Moritzia and Thaumatocaryon Baill.
5. Boraginaceae (Echiochiloideae): The stem node of Ogastemma (incl. Antiphytum DC. Ex Meisn., Echiochilon Desf. and Ogastemma) was set to a minimum age of 41.2 Ma (logMean: 2.0, logStdev: 1.0, median age: 48.6 Ma), based on the recently discovered fossil nutlets assignable to Ogastemma for the Eocene (Ypresian-Lutetian, 56–41.2 Ma) of SW Algeria (Hammouda et al., 2016).
6. Ehretiaceae: The stem node of Ehretia II was set to a minimum age of 47.8 Ma (logMean: 1.4, logStdev: 1.0, median age: 51.86 Ma) based on fossils fruits of Ehretia P. Browne reported from the lower Eocene (Ypresian, 56–47.8 Ma) of the London Clay (Chandler, 1961, 1962, 1964), which are the oldest fossils assigned to this family.
7. Heliotropiaceae: The stem node of Tournefortia L. sect. Tournefortia was set to a minimum age of 28.1 Ma (logMean: 1.1, logStdev: 1.0, median age: 31.1 Ma). This is based on fossil pollen grains recorded by Graham and Jarzen (1969) for the lower Oligocene (ca. 33.9–28.1 Ma) of Puerto Rico.
8. Asteraceae: The stem node of Asteraceae was set to a minimum age of 47.5 Ma (logMean: 1.5, logStdev: 1.0, median age: 51.98 Ma) based on fossils reported for the Ypresian of Patagonia (Barreda et al., 2010, 2012). The recently reported pollen fossil of Asteraceae from the Cretaceous of Antarctica (Barreda et al., 2015) was not employed here due to the controversy regarding its assignment to a specific clade (Barreda et al., 2016; Panero, 2016).
9. Ericales: The crown node of Ericales was set to a minimum age of 89.8 Ma (logMean: 0.7, logStdev: 1.0, median age: 91.8 Ma) based on fossils related to the extant genus Enkianthus Lour. (Ericaceae) from the Turonian of New Jersey (Nixon and Crepet, 1993).
10. Cornales: The crown node of Cornales was set an offset of 86.3 Ma, logMean: 0.8, logStdev: 1.0, median age: 88.53 Ma, based on fossils from the Coniacian of Japan (Takahashi et al., 2002).
Finally, the root height was set to 103 Ma using a normal distribution with a standard deviation of 7.5. This is a secondary calibration is based on Magallón et al. (2015) and corresponds to the crown node of the Asteridae. The MCMC was run for 200 million generations sampling every 20,000 trees and a burnin of 10%. The XML file generated in BEAUti was run using the web resources available in the CIPRESS Science Gateway (Miller et al., 2010).
Biogeographical Analyses
The biogeographical analyses were conducted for a reduced dataset in which only the 190 taxa of Boraginaceae and the closest relatives were included [i.e., 183 Boraginaceae ingroup species plus two species of each Codonaceae and Wellstediaceae. The remaining taxa from the Boraginales II-clade (Ehretiaceae and Heliotropiaceae) plus other distantly related outgroups (Asteraceae, Cornales, and Ericales) were excluded as they are outside the geographical scope of this study]. Seven geographic areas were defined following the treatment of Luebert et al. (2017) for Boraginales and the floristic kingdoms of Takhtajan (1986) and Cox (2001). A subdivision of Eurasia in three sub-regions was necessary, as more than 80% of all Boraginaceae genera are differentially distributed in that area (Weigend et al., 2016). Additionally, the Indo-Pacific region (see Cox, 2001) was not included in this study because only one accession in our data set could have been assigned to it [Trichodesma zeylanicum (Burm. Fil.) R.Br., see Retief and van Wyk, 2002]. The designation of areas for each species was based on geographic information derived from herbarium vouchers, floras and taxonomic revisions (see references in Weigend et al., 2016). The following geographic areas were defined: A: South America, where the genus Moritzia is endemic and several species of Lithospermum (Lithospermeae) as well as Cryptantha, Plagiobothrys, and other Amsinckiinae occur. B: North America, with seven endemic genera (Adelinia J. I. Cohen, Andersonglossum J. I. Cohen, Amsinckia, Antiphytum, Cryptantha, Dasynotus, Pectocarya). C: Western Europe (including the Balkan region and the Mediterranean zone comprised between southern Europe and northern Africa), this is the main distribution center of the Lithospermeae as well as many taxa in Boraginineae and Cynoglossoideae, like the large genera Myosotis L. or Cynoglossum L. (the latter also with species in the Americas, Australia and New Zealand). D: Africa (southwards to the Mediterranean zone, including Macaronesia and the Arabian Peninsula), with endemic genera such as Afrotysonia Rauschert, Cystostemon Balf. F., Ogastemma, and species of Echiochilon Desf. and Echium L. E: Irano-Turanian region in the sense of Takhtajan (1986) including the western Himalayas. Is the center of endemism of Caccinia Savi, Craniospermum Lehm., Heterocaryum A.DC., Suchtelenia Karel. Ex meisn., among others. F: Australia and New Zealand, with many species showing disjunct distributions with Eurasia and the Americas (e.g., Hackelia Opiz, Cynoglossum, or Myosotis) and only one endemic genus in New Zealand (Myosotidium Hook.). G: East Asia (including the eastern Himalayas, China, Mongolia, Taiwan, Japan, and eastern Russia), where the Lasiocaryeae are endemic (Chionocharis I.M.Johnst., Lasiocaryum I.M.Johnst., and Microcaryum I.M.Johnst.; see Chacón et al., 2017) as well as many genera such as Decalepidanthus Riedl, Nihon A.Otero, Jim.Mejías, Valcárcel, and P. Vargas, Trigonotis Steven, among others.
The maximum clade credibility (MCC) tree (chronogram) calculated in BEAST was used to reconstruct the ancestral geographic ranges in Boraginaceae. The analyses were carried out in the R package BioGeoBEARS (Matzke, 2013) using the Dispersal Extinction Cladogenesis model (DEC; Ree and Smith, 2008) and its extended version DEC+J (Matzke, 2014). This is a maximum likelihood-based model of geographic range evolution in which the distribution ranges of species are allowed to change along the branches of a phylogenetic tree by means of two main types of events: “dispersal,” or range expansion, and “extinction” or range reduction. In BioGeoBEARS a jump dispersal parameter (“J”) is added to the model allowing for the inference of founder-event speciation (Matzke, 2014). Through this method the likelihood of the ancestral states at cladogenesis (i.e., range inheritance scenarios) are estimated (Ree and Smith, 2008). BioGeoBEARS infers ancestral areas based on the global maximum likelihood model.
The maximum number of areas was set to three as most Boraginaceae clades have very restricted geographic ranges, occurring currently in no more than 3 areas. We ran unconstrained and constrained analyses for both the DEC and DEC+J models. In the constrained analyses all non-adjacent areas were excluded, therefore the following ranges were forbidden: AC, AE, AG, BD, BE, BF, CF, CG, DF, EF, ACE, ACG, AGE, BCF, BDE, BDF, BEF, CEF, and CFG. To assess how well the four models (DEC unconstrained, DEC constrained, DEC+J unconstrained, DEC+J constrained) fit our data (i.e., the geographic areas and the MCC-BEAST tree), their likelihood scores were compared my means of the Akaike Information Criterion (AIC) and the Akaike weights implemented in the Optimx function of BioGeoBEARS. These tests allow for model comparison and assessment for model selection uncertainty even if the models are not nested (Posada and Buckley, 2004). To evaluate and quantify the relative importance of dispersal, vicariance and sympatric speciation in the evolution of Boraginaceae we obtained the overall probabilities of the anagenetic and cladogenetic events conditional on the geographic distributions, the phylogeny, and the best-fitting model, from 1,000 stochastic biogeographic mappings (Matzke, 2016).
Dispersal Modes in Boraginaceae
In order to understand the role of the different dispersal modes in the diversification of Boraginaceae we used the previously compiled list of the mechanisms reported in the literature for all the species of Boraginaceae and the related families Wellstediaceae and Codonaceae sampled in this study (Supplementary Table 1). In the cases where no information for a particular species was found the most plausible dispersal mode was assigned based on own observations of fruit and/or nutlet phenotypes and descriptions of related species reported in the literature.
Ancestral dispersal modes in Boraginaceae were inferred using the likelihood-based stochastic mapping method of ancestral character reconstruction implemented in SIMMAP (Bollback, 2006). Seven dispersal mechanisms were coded: Anemochory, atelechory, chamaeochory, endozoochory, epizoochory, myrmecochory, and nautochory (incl. hydrochory). To evaluate shifts in dispersal mode of those clades for which transoceanic or continental LDD was inferred in the biogeographical analysis, the results obtained in SIMMAP and those obtained in BioGeoBEARS were visually compared.
Results
Divergence Times in Boraginaceae
The three chloroplast markers were successfully sequenced for all taxa except for three outgroup species for which either trnS-G or rps16 failed to amplify (i.e., trnS-G sequences are missing for Coldenia procumbens L. and Pholisma arenarium Nutt., and rps16 for Chuquiraga jussieui J.F.Gmel.). Also in Lactuca sativa L. the sequence of trnS-G could not be obtained due to two inversions in the chloroplast genome between the trnG-UCC and trnE-UUC genes and upstream of the trnS-GCU gene (Kim et al., 2005). Therefore, the alignments analyzed in BEAST have almost no missing data. The rps16, trnL-F, and trnS-G matrices consisted of 1,734, 1,667, and 2,794 aligned nucleotides, respectively. Effective sample sizes (ESS) were checked in Tracer 1.6 (Rambaut et al., 2014) and were all well above 200. The MCC tree obtained in BEAST was well supported, with posterior probability values ≥0.98 for most clades (Supplementary Figure 1), and inferred ages within the 95% high posterior density interval (HPD) for most nodes (Table 1 and Supplementary Figure 2). The resulting tree confirms the position of the newly sequenced taxa Antiphytum cf. heliotropioides, Ogastemma pusillum (Echiochiloideae), the Brazilian Moritzia spp. (Boraginoideae), and the American Lithospermum spp. (Lithospermeae) based on the phylogenetic studies of Chacón et al. (2016) and Cohen (2014). The age of the most recent common ancestor (MRCA) of Boraginales was dated to the Cenomanian stage in the Upper Cretaceous, ca. 96.2 million years ago (Ma; 95% HPD = 85.0–107.4 Ma), which is congruent with the age reported in Luebert et al. (2017) as for the crown node of Boraginales (i.e., 91.1 Ma, 95% HPD = 81.8–100 Ma). The split between Wellstediaceae and Boraginaceae occurred in the Upper Cretaceous, ca. 79 Ma (95% HPD = 68.4–89.2 Ma) and the diversification Boraginaceae began during the Palaeocene, ca. 62.6 Ma (95% HPD = 54.6–71.7 Ma). The crown nodes of the three Boraginaceae subfamilies Boraginoideae, Cynoglossoideae and Echiochiloideae were all dated within the Eocene (ca. 55.5 Ma, 52.3 Ma and 45 Ma, respectively). Divergence times for the nodes of interest in Boraginaceae are shown in Table 1.
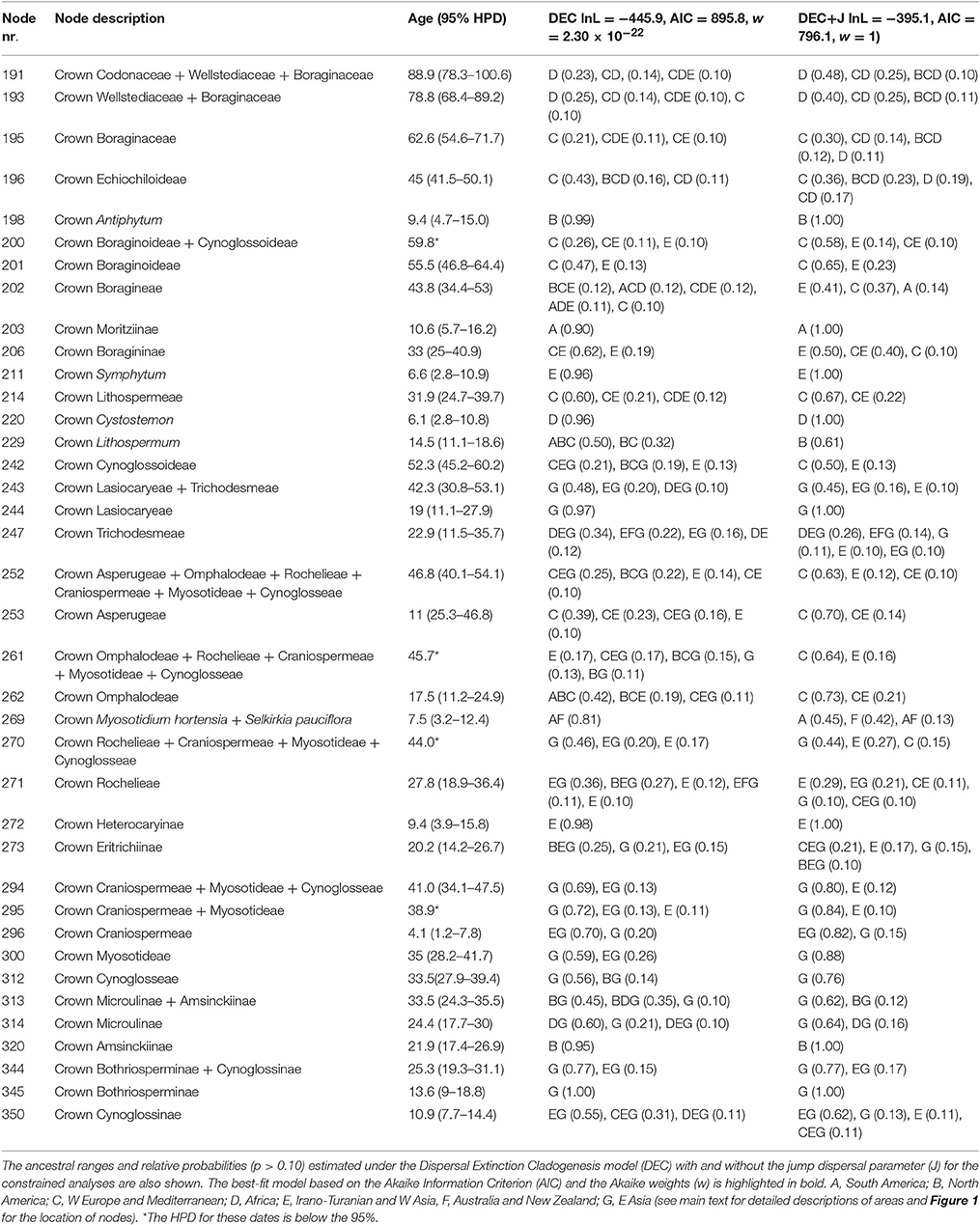
Table 1. Mean node ages (millions of years before present) and 95% high posterior density intervals (HPD) for the nodes of interest in Boraginaceae.
Biogeography of Boraginaceae
The AIC test and the Akaike weights indicated that the constrained analyses are preferred over the unconstrained ones for both the DEC and the DEC+J models. Between these two models the DEC+J was chosen as the best-fitting model given our data. A summary of the results of the AIC test and the Akaike weights, as well as the ancestral ranges inferred for the nodes of interest under the four models are presented in Table 1 (constrained analyses) and in Supplementary Table 3 (unconstrained analyses). The results obtained under the best-fitting model (i.e., DEC+J using constrained ranges) are shown in Supplementary Table 4 (for all nodes of the Boraginaceae tree) and also illustrated in Figure 1. Based on that model the ancestral range of the MRCA of Boraginaceae + Wellstediaceae was in Africa with low probability (p = 0.41, Table 1 and Figure 1). The ancestral range of Boraginaceae was inferred in the Mediterranean and Western Europe (area C, p = 0.30). For some major nodes in Boraginaceae with wide geographic distributions the reconstructions were ambiguous. This was the case of the crown node of Boragininae [Western Europe and the Irano-Turanian region inferred with similar probabilities: C (p = 0.35) and E (p = 0.43), respectively], and the crown node of Heterocaryinae [the composite range comprising the whole Eurasian continent and the Irano-Turanian region inferred with similar probabilities: CEG (p = 0.20) and E (p = 0.18)].
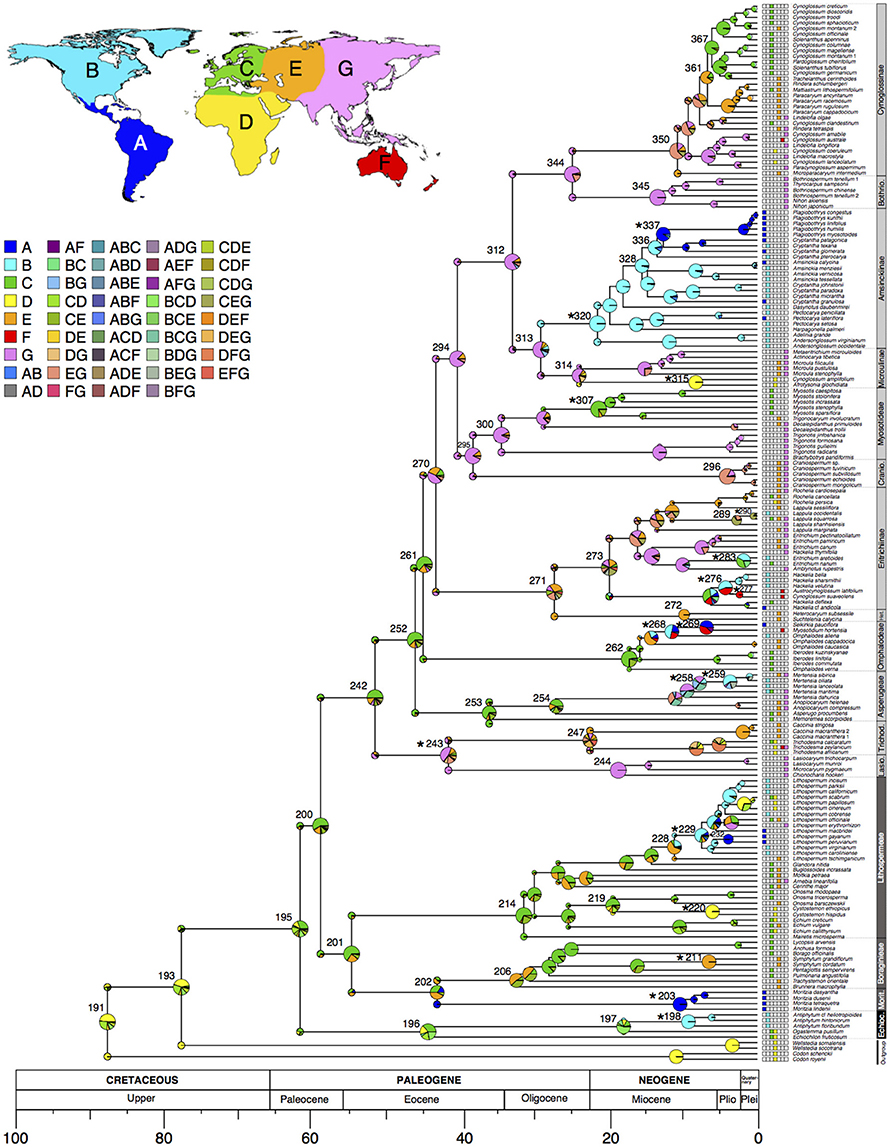
Figure 1. Dated phylogeny of Boraginaceae and ancestral ranges inferred under the DEC+J constrained model. The main infrafamilial subdivisions are shown in front of each clade following Chacón et al. (2016). Relative probabilities of each ancestral reconstruction are indicated with pie charts at each node following the color-coding described in the inset. The nodes numbers for the clades of interest are shown next to each pie chart (numbers for all the remaining nodes are shown in Supplementary Figure 4). Areas included in the biogeography analyses are depicted in the map (see main text for detailed descriptions). Dispersal events at cladogenesis are marked with an asterisk. The scale at the bottom corresponds to the geological time in millions of years before present.
The most frequent modes of range inheritance inferred from 1,000 biogeography stochastic maps in Boraginaceae were narrow sympatry (range copying within the same area; see Matzke, 2014) in 47.03% of all events (110,672 out of 235,290 total events), and dispersals in 38.9% of all events (91,456 out of 235,290 total events; see Table 2 and Supplementary Figure 3). Founder dispersal accounted for 49.4% of all dispersals (45,166 out of 91,456 of all dispersals) while range expansion (anagenetic dispersal) accounted for 50.6% (46,290 out of 91,456 of all dispersals). Among all the events at cladogenesis vicariance represented only 5.3% of these events (10,044 out of 189,000 cladogenetic events). Most dispersal events at anagensis occurred within Eurasia (Table 3), especially between Eastern Asia and the Irano-Turanian region (from G to E and viceversa), and between the Irano-Turanian region and Western Europe (from E to C and viceversa). Among all cladogenetic dispersals (Table 4) the most frequent were from the Mediterranean and Western Europe into the Irano-Turanian region (C to E) and from North America to South America (B to A). Eleven transoceanic dispersals were inferred, most of them from the Mediterranean and Western Europe to North America (C to B). The most frequent cladogenetic and anagenetic dispersal events (inferred with p ≥ 0.80) are illustrated in Figure 2.

Table 2. Summary of the speciation events inferred in BioGeoBEARS based on 1,000 biogeography stochastic mappings.
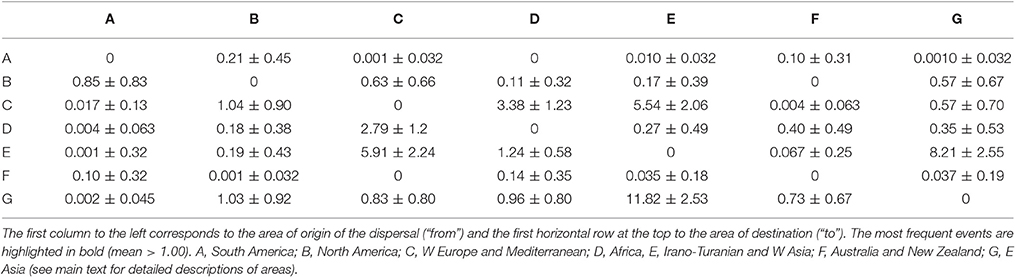
Table 3. Range expansion events (mean and standard deviation of all observed anagenetic dispersals) inferred in BioGeoBEARS across 1,000 biogeography stochastic maps.

Table 4. Cladogenetic dispersals (mean and standard deviation of all observed jump dispersals) inferred in BioGeoBEARS across 1,000 biogeography stochastic maps.
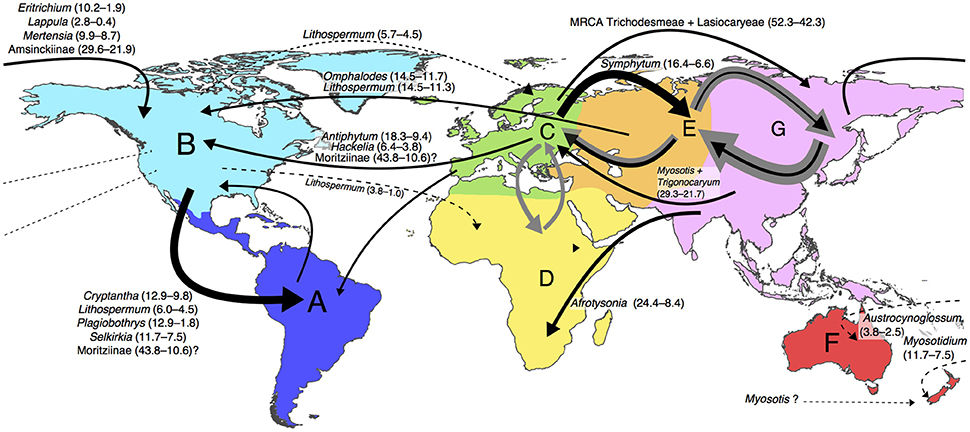
Figure 2. Main cladogenetic and anagenetic dispersal events in Boraginaceae according to the results of the biogeography stochastic mapping. The direction of each event is indicated with an arrow. In black are the cladogenetic dispersals and in gray the anagenetic dispersals. The frequency of each event is proportional to the thickness of the arrow according to the values shown in Tables 3, 4. Some examples of Boraginaceae clades where one or more cladogenetic dispersals have taken place are shown next to the corresponding arrow. The time interval in which the dispersal event might have occurred (i.e., the interval between the stem node and the crown node in millions of years before present) is indicated in parenthesis. Dotted arrows indicate a low probability of the inferred event (p < 0.80). In the case of Myosotis the dotted arrow represents a hypothetical dispersal, as no New Zealand species were included in this study. MRCA: the most recent common ancestor (see Section Materials and Methods for detailed area descriptions).
Dispersal Modes in Boraginaceae
The results obtained in SIMMAP (Figure 3) showed that the ancestral dispersal mode in Boraginaceae was epizoochory and that shifts to other dispersal modes occurred at least 12 times during the evolution of Boraginaceae. Sister group comparisons in Figure 3 reveal that dispersal mode of clades for which LDD can be inferred (marked with an asterisk in Figure 1) rarely differ from the dispersal mode of its sister clade (Figure 3). That is the case of Lithospermum (node 229 of Figure 1 and Supplementary Figure 4), Omphalodes (node 268), Hackelia (node 276), Austrocynoglossum (node 277), Eritrichium (node 283), Lappula (node 290), Amsinckiinae (node 320), Cryptantha (node 328), Symphytum (node 211), and Cystostemon (node 220). Cases in which dispersal mode differ between sister groups (Figure 3) are in endozoochorous Antiphytum (node 198), whose sister group has a bimodal dispersal mode; epizoochorous Moritzia (node 203) whose sister clade is myrmecochorous; endozoochorous Mertensia (node 259), whose sister clade (Mertensia maritima) is nautochorous; epizoochorous Selkirkia (node 269), sister to anemochorous Myosotidium; epi- and endozoochorous (bimodal-1), as well as in the sister clades Trichodesmeae and Lasiocaryeae (node 243), with atelechory and endozoochory, respectively, and Myosotis and Trigonocaryum, (node 307) with epizoochory and myrmecochory, respectively. After visual comparison of Figures 1, 3 no correlation between a particular LDD event and a shift in dispersal mode was found. Moreover, the clades in which LDD was observed are characterized by more than one type of dispersal (i.e., polychorous), such as the case of node 337, with endozoochorous and epizoochorous species; node 320, including endo- and epizoochorous as well as atelochorous species; node 307, with epizoochorous and myrmecochorous species; node 258, with endozoochorous and nautochorous species; nodes 268 and 269, with anemochorous, epizoochorous, and nautochorous species; node 258, with endozoochorous and nautochorous species, and node 243, with a predominance of atelochory but also with anemochorous and endozoochorous species (see Figure 3).
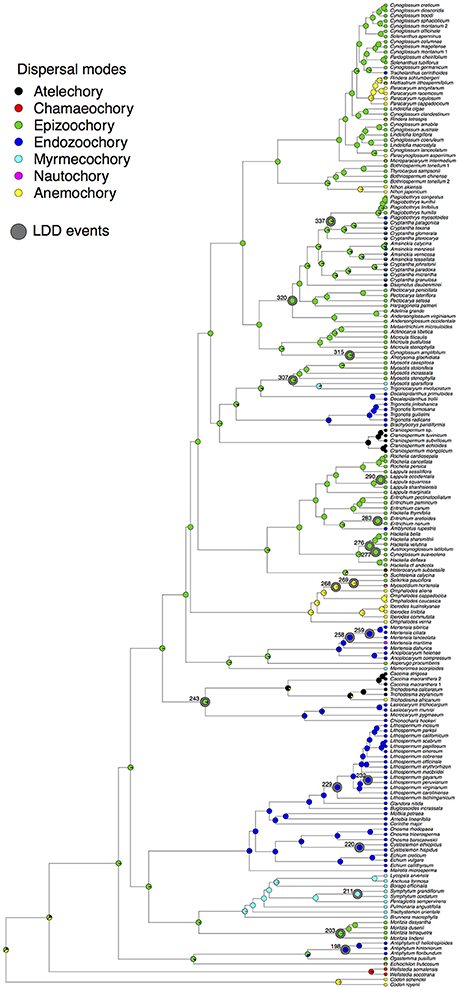
Figure 3. Ancestral dispersal modes in Boraginaceae and the closest relatives Codonaceae and Wellstediaceae. Pies at nodes indicate the reconstructed ancestral states inferred in SIMMAP. Pies at tips correspond to the primary dispersal modes assigned to each species according to the color-coding in the inset and the information compiled in the Supplementary Table 1. The nodes in which a long-distance dispersal (LDD) event took place are surrounded with a gray circle.
Discussion
Our comprehensive taxon sampling, which covers all tribes and 80% of all Boraginaceae genera from throughout the distribution range, the low percentage of missing data in the alignments (0.46%, only 4 out of 861 sequences could not be obtained) and the robust calibration scheme with multiple fossils from Boraginaceae and related families, provided a well-supported chronogram for further biogeographic analyses (Figure 1 and Supplementary Figure 2). The dated phylogeny allowed us to infer the main biogeographic events and distribution shifts that took place during the evolution of Boraginaceae, thus providing a solid historical scenario that allowed us to analyze the relationship with different dispersal modes in this clade.
The biogeographic history of the crown Boraginaceae dates back to Paleocene ca. 55–72 Ma (Figure 1 and Table 1). This is older than the age reported in Luebert et al. (2017), who dated the crown node of Boraginaceae to 53.2 Ma (95% HPD = 46.7–59.6 Ma). The discrepancies with respect to that study, which was focused on the biogeography of the whole Boraginales clade, are due to differences in the taxon sampling and the fossil calibration scheme used for the dating (i.e., 20 Boraginaceae species instead of 183 and eight calibration points instead of ten, see Materials and Methods Section). Initial diversification of Boraginaceae occurred in Western Europe, a region that by this time was about 10 times closer to Greenland than it is today (Ellis and Stoker, 2014). The global warming that affected the climate worldwide during the Paleocene-Eocene Thermal maximum (Zachos et al., 2001) had a tremendous impact on the plant distribution patterns, with temperate floras and biota becoming restricted to higher latitudes, and tropical taxa expanding polewards (Graham, 2011). Long branches near the backbone of Boraginaceae suggest that the extinction of lineages in the Paleogene also took place, for instance in the stem branch of Lithospermeae (between 55.5 and 31.9 Ma; Figure 1 and Table 1). This could have been associated with the climatic deterioration and increased aridification in some parts of Eurasia, as evidenced by the fossil record of xerophytes in the area of Kazakhstan and western Central Asia (Akhmetiev and Beniamovski, 2009). Our age estimates suggest that the early diversification of most clades of Boraginaceae (Table 1, Figure 1 and Supplementary Figure 2) occurred in Western Europe during the maximum warming period of the Cenozoic, which started ca. 59–50 Ma and peaked 52–50 Ma (i.e., the Paleocene-Eocene thermal maximum; Zachos et al., 2001). The considerably warmer conditions during this period are evidenced by the appearance of fossil remains of thermophilic angiosperms such as Liquidambar sp., Magnolia sp., Arecaceae, Myrtaceae, Loranthaceae, Moraceae, Magnoliaceae, among others (Akhmetiev and Beniamovski, 2009 and references therein) in localities polewards of 70°N.
The BioGeoBEARS results inferred that the main cladogenetic events in Boraginaceae were range copying within the same area (here interpreted as the speciation events that occurred within the same large continental area) and dispersal (Matzke, 2014; Table 2). Previous phylogenetic and biogeographic studies have already suggested the prevalence of LDD in Boraginaceae (Weigend et al., 2013; Luebert et al., 2017). The Paleocene diversification of Cynoglossoideae, the most species rich group Boraginaceae (Weigend et al., 2013, 2016; Chacón et al., 2016), perhaps exemplifies both of these events, as the initial divergence of this clade was followed by range expansions (or LDD) from Western Europe to Eastern Asia (Figure 2); once during the Lutetian (47.8–41.3 Ma; see the geological time scale in Cohen et al., 2013) in the MRCA of the clade (Rochelieae + Craniospermeae + Myosotideae + Cynoglosseae) clade, and once in the MRCA of the Lasiocaryeae + Trichodesmeae clade (Figure 1 and Table 1). During this period, parts of Western Asia and the Irano-Turanian region where covered by the Tethys Ocean, whose closure started about 50 Ma after the final collision of India with the Eurasian plate (Zhang et al., 2012). This allowed the posterior dispersal of plants between Europe and Eastern Asia, both across and from the Irano-Turanian region (e.g., Manafzadeh et al., 2014).
The biogeographic events that were inferred for the Cynoglossoideae (Figure 2 and Table 2) involved many Oligocene and Miocene dispersals, back and forth into the Irano-Turanian region (e.g., in the Trichodesmeae, Heterocariineae, Eritrichiineae, Craniospermeae, and Cynoglossineae) and Western Europe (Cynoglossum, Hackelia, Myosotideae, Omphalodeae). Long-distance dispersals from Eastern Asia to North America were estimated for Mertensia, in agreement with the study of Nazaire et al. (2014). Further dispersals from North America to South America (MRCA of Amsinckiinae and Mertensia) as well as from Eastern Asia to Australia (Austrocynoglossum), and from North America to New Zealand (Myosotidium) also occurred between the Miocene and the Pliocene (Figures 1, 2). Myosotis is an interesting group that also dispersed from Eurasia into the southern hemisphere, with a species radiation in New Zealand (40 species, Meudt et al., 2015). Although our sampling did not permit to infer the timing and direction of these events, the study of Meudt et al. (2015) showed that the diversification of Myosotis in New Zealand, Chile and New Guinea goes back to the Pleistocene, apparently from a northern hemisphere ancestor. Outside of the Cynoglossoideae older occurrences of LDD events are perhaps in the Moritziinae (Figure 1 and Table 1). In our analyses, only Brazilian accessions of Moritzia are included, but the current geographic distribution of the sister genus Thaumatocaryon in South America indicates that this clade reached South America after a LDD event originating from Western Europe (Figures 1, 2). The abundance of fossils assignable to this clade in Miocene deposits of Kansas suggests that the MRCA ancestor of Moritziinae dispersed first to North America and from here to South America, going extinct in North America afterwards (Weigend et al., 2010). Without samples of Thaumatocaryon the exact time of this dispersal was, however, not possible to infer. Nevertheless, it is known that an extensive interchange of tropical plant lineages across the North Atlantic land bridge into North America took place during the Eocene (Lavin and Luckow, 1993; Tiffney and Manchester, 2001; Donoghue, 2008). Toward the end of the Miocene the severe cooling conditions and the formation of a northern-hemisphere ice sheet (Zachos et al., 2001) interrupted these migrations (Lavin and Luckow, 1993). Between the middle Miocene and the Pleistocene many dispersal events of Cynoglossoideae clades either from Western and Central Europe (Irano-Turanian region; e.g., in Omphalodes, Lithospermum and Hackelia) or from Eastern Asia (e.g., in Eritrichium, Lappula, Mertensia) into North America also took place (Figures 1, 2). The rich fossil record of Boraginaceae found in Miocene deposits of central North America (see Materials and Methods) probably represents evidence for “temporal (and ecological) continuity between Eurasian-North American disjuncts of warm and humid climates and those of warm and dry climates” (Kadereit and Baldwin, 2012: p. 14). The dispersal of Antiphytum into North America might have originated either from the northern end of Africa or from Eurasia, based on the Echiochiloideae fossil record of Algeria, France and India (Taugourdeau-Lantz and Rosset, 1966; Mathur and Mathur, 1983; Berger et al., 2013; Hammouda et al., 2016).
Relationships between Long-Distance Dispersal and Diaspore Dispersal Modes in Boraginaceae
The Boraginaceae groups illustrated in Figure 2 represent some examples of lineages in which the occurrence of LDD can be analyzed in the context of life history traits such as the diaspore dispersal modes. Based on the results of the stochastic character mapping conducted in SIMMAP (Figure 3) no shifts in dispersal modes could be associated with the events of long-distance dispersal estimated in the biogeography analyses. The only exception is perhaps Antiphytum, in which a shift from epizoochory to endozoochory could be associated with the Miocene long-distance dispersal from Western Europe to North America (Figures 2, 3). In genera such as Lithospermum, which has dispersed multiple times across oceans and continents since the middle Miocene (Figure 2), the endozoochory seems to be a preadaptation to LDD, as it is the primary dispersal mode of all other the members of the Lithospermeae. Anemochory was inferred as the ancestral dispersal mode in the Omphalodeae (Figure 3) in which a LDD event from the Irano-Turanian region into North America during the Miocene was inferred for Omphalodes (Figure 2). In summary, we did not find a single dispersal mode common to all the examples of LDD documented in this study. However, the absence of LDD was associated with the avoidance of any dispersal (atelechory) in the case of genera adapted to extreme habitat conditions such as Craniospermum or the Lasiocaryeae (Figures 2, 3).
The data set here presented has its limitations with regards to sampling and constraints inherent to the statistical methods employed, but our results are congruent with the idea that LDD appears to be associated with dispersal vectors other than those inferred from the plant phenotypes (Berg, 1983; Higgins et al., 2003). Moreover, the hypothesis that LDD tends to be more frequent in polychorous clades seems to be supported from our data for Boraginaceae, with at least seven events of LDD inferred at nodes in the phylogeny that include taxa with both biotic and abiotic dispersal modes (Figure 3). The incidence of LDD in plant groups lacking dispersal syndromes and the successful establishment in island ecosystems evidences the complexity of this phenomenon (Vargas et al., 2012; Heleno and Vargas, 2015). The idea that different dispersal agents might be playing differential roles depending on the plant responses to environmental changes (Nathan et al., 2008 and references therein) can explain the lack of a correlation between one particular dispersal mode and the occurrence of LDD in Boraginaceae. The profound climatic and environmental changes that took place during the Paleogene and Neogene evolution of this clade might have had great impacts on the species life history traits and the dispersal capabilities of the species, which today are almost impossible to track.
Conclusions
The initial Paleocene divergence of Boraginaceae in Western Eurasia was followed by range expansions into the Irano-Turanian region and Eastern Asia, including multiple events of LDD into the Americas, Australia, and New Zealand. There is no hard evidence for any back-dispersal from the Americas to Africa or Eurasia, but some evidence for dispersal from the Americas to Australia/New Zealand. The data further indicate that, at genus level, dispersal and establishment between the Americas was exclusively North to South. A denser sampling in Amsinckiinae would probably confirm the presence of repeated back-dispersal to North America in this clade (Hasenstab-Lehman and Simpson, 2012). The extreme climatic changes that took place in Eurasia during the Paleogene and Neogene likely shaped the biogeographic patterns of Boraginaceae, influencing at the same time complex interactions between the environmental change and the differential effectiveness of the dispersal vectors. The association between LDD and polychorous dispersal modes in some clades suggests that the diversification of Boraginaceae was likely driven by adaptations to different dispersal agents and to unassisted diaspore dispersal, allowing the species to colonize new habitats and remote geographic regions worldwide. A complete overview over ca. 60 million years of evolutionary history in the Boraginaceae is certainly impossible. However, the data here presented tentatively suggest that different dispersal modes rather unpredictably led to LDD in this family, with no single dispersal mode responsible for a majority of successful dispersal events.
Author Contributions
JC, FL, MW conceived and design the study; JC, FL, MW acquired information on dispersal modes; JC, FL analyzed the data; JC drafted the manuscripts; FL, MW critically revised and contributed content to the manuscript. All authors agree to be accountable for all aspects of the work.
Funding
This study was supported by research funds of the University of Bonn.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
The authors thank R. Trevisan for material of Moritzia collected in Brazil, F. Selvi for samples of Lithospermum, S. Ovczinnikova for samples of Craniospermum, W. Lobin and T. Joßberger for material collected in Morocco and specimens from the Bonn Botanical Gardens. Thanks to N. Schmandt for assistance in the molecular lab, H. H. Hilger and all other participants of the III International Boraginales Meeting that took place in Bonn in September 2016 for valuable feedback and discussion.
Supplementary Material
The Supplementary Material for this article can be found online at: http://journal.frontiersin.org/article/10.3389/fevo.2017.00026/full#supplementary-material
References
Akhmetiev, M. A., and Beniamovski, V. N. (2009). Paleogene floral assemblages around epicontinental seas and straits in Northern Central Eurasia: proxies for climatic and paleogeographic evolution. Geol. Acta 7, 297–309. doi: 10.1344/105.000000278
Barreda, V. D., Palazzesi, L., Katinas, L., Crisci, J. V., Tellería, M. C., Bremer, K., et al. (2012). An extinct eocene taxon of the daisy family (Asteraceae): evolutionary, ecological and biogeographical implications. Ann. Bot. 109, 127–134. doi: 10.1093/aob/mcr240
Barreda, V. D., Palazzesi, L., Tellería, M. C., Katinas, L., Crisci, J. V., Bremer, K., et al. (2010). Eocene patagonia fossils of the daisy family. Science 329:1621. doi: 10.1126/science.1193108
Barreda, V. D., Palazzesi, L., Tellería, M. C., Olivero, E. B., Raine, J. I., and Forest, F. (2015). Early evolution of the angiosperm clade asteraceae in the cretaceous of antarctica. Proc. Natl. Acad. Sci. U.S.A. 112, 10989–10994. doi: 10.1073/pnas.1423653112
Barreda, V. D., Palazzesi, L., Tellería, M. C., Olivero, E. B., Raine, J. I., and Forest, F. (2016). Reply to panero: robust phylogenetic placement of fossil pollen grains: the case of Asteraceae. Proc. Natl. Acad. Sci. U.S.A. 113, E412–E412. doi: 10.1073/pnas.1521642113
Berger, B. A., Kriebel, R., Spalink, D., and Sytsma, K. J. (2016). Divergence times, historical biogeography, and shifts in speciation rates of Myrtales. Mol. Phylogenet. Evol. 95, 116–136. doi: 10.1016/j.ympev.2015.10.001
Berger, J. P., Collinson, M. E., and Weidmann, M. (2013). Un curieux micro- fossile de la Molasse oligocène de Suisse occidentale et de Haute-Savoie (France). Swiss J. Geosci. 106, 125–133. doi: 10.1007/s00015-013-0124-x
Berg, R. Y. (1983). “Plant distribution as seen from plant dispersal: General principles and basic modes of plant dispersal,” in Dispersal and Distribution, ed K. Kubitzki (Hamburg: Paul Parey), 13–36.
Best, T. L., and Smartt, R. A. (1986). Feeding ecology of mourning doves (Zenaida macroura) in southeastern New Mexico. Southwest. Nat. 31, 33–38. doi: 10.2307/3670957
Bollback, J. P. (2006). SIMMAP: stochastic character mapping of discrete traits on phylogenies. BMC Bioinformatics 7:88. doi: 10.1186/1471-2105-7-88
Bricker, M., Pearson, D., and Maron, J. (2010). Small-mammal seed predation limits the recruitment and abundance of two perennial grassland forbs. Ecology 91, 85–92. doi: 10.1890/08-1773.1
Carlquist, S. (1983). “Intercontinental dispersal,” in Dispersal and Distribution: An International Symposium, (Hamburg), 37–47.
Chacón, J., Hilger, H. H., Luebert, F., Ovchinnikova, S., Selvi, F., Cecchi, L., et al. (2016). The borage family (Boraginaceae s.s.): a revised infrafamilial classification based on new phylogenetic evidence, with emphasis on the placement of some enigmatic genera. Taxon 65, 523–546. doi: 10.12705/653.6
Chacón, J., and Renner, S. S. (2014). Assessing model sensitivity in ancestral area reconstruction using Lagrange: a case study using the Colchicaceae family. J. Biogeogr. 41, 1414–1427. doi: 10.1111/jbi.12301
Chacón, J., Siwakoti, M., Hilger, H. H., and Weigend, M. (2017). Dwarves on the roof of the world: a taxonomic revision of the Himalayan Lasiocaryeae Weigend (Boraginaceae). Phytotaxa 297, 001–014. doi: 10.11646/phytotaxa.297.1.1
Chandler, M. E. J. (1961). The Lower Tertiary Floras of Southern England. I. Paleocene Floras. London Clay Flora. London: The British Museum.
Chandler, M. E. J. (1962). The Lower Tertiary Floras of Southern England. II. Flora of the Pipe-Clay of Dorset (Lower Basshot). London: The British Museum.
Chandler, M. E. J. (1964). The Lower Tertiary Floras of Southern England. IV. A Summary and Survey of Findings in the Light of Recent Botanical Observations. London: The British Museum.
Christenhusz, M. J. M., and Chase, M. W. (2013). Biogeographical patterns of plants in the Neotropics – dispersal rather than plate tectonics is most explanatory. Bot. J. Linn. Soc. 171, 277–286. doi: 10.1111/j.1095-8339.2012.01301.x
Cohen, J. I. (2014). A phylogenetic analysis of morphological and molecular characters of Boraginaceae: evolutionary relationships, taxonomy, and patterns of character evolution. Cladistics 30, 139–169. doi: 10.1111/cla.12036
Cohen, K. M., Finney, S. C., Gibbard, P. L., and Fan, J.-X. (2013). The ICS international chronostratigraphic chart. Episodes 36, 199–204.
Cox, B. (2001). The biogeographic regions reconsidered. J. Biogeogr. 28, 511–523. doi: 10.1046/j.1365-2699.2001.00566.x
de Queiroz, A. (2005). The resurrection of oceanic dispersal in historical biogeography. Trends Ecol. Evol. 20, 68–73. doi: 10.1016/j.tree.2004.11.006
Donoghue, M. J. (2008). A phylogenetic perspective on the distribution of plant diversity. Proc. Natl. Acad. Sci. U.S.A. 105, 11549–11555. doi: 10.1073/pnas.0801962105
Drummond, A. J., Suchard, M. A., Xie, D., and Rambaut, A. (2012). Bayesian phylogenetics with BEAUti and the BEAST 1.7. Mol. Biol. Evol. 29, 1969–1973. doi: 10.1093/molbev/mss075
Edwards, W., and Westoby, M. (1996). Reserve mass and dispersal investment in relation to geographic range of plant species: phylogenetically independent contrasts. J. Biogeogr. 23, 329–338. doi: 10.1046/j.1365-2699.1996.00034.x
Ellis, D., and Stoker, M. S. (2014). The Faroe–Shetland Basin: a regional perspective from the Paleocene to the present day and its relationship to the opening of the North Atlantic Ocean. Geol. Soc. Spec. Publ. 397, 11–31. doi: 10.1144/SP397.1
Gabel, M. L. (1987). A fossil Lithospermum (Boraginaceae) from the Tertiary of South Dakota. Am. J. Bot. 74, 1690–1693. doi: 10.2307/2444138
Gabel, M. L., Backlund, D. C., and Haffner, J. (1998). The Miocene macroflora of the northern Ogallala group, northern Nebraska and southern South Dakota. J. Paleontol. 72, 388–397. doi: 10.1017/S0022336000036362
Gillespie, R. G., Baldwin, B. G., Waters, J. M., Fraser, C. I., Nikula, R., and Roderick, G. K. (2012). Long-distance dispersal: a framework for hypothesis testing. Trends Ecol. Evol. 27, 47–56. doi: 10.1016/j.tree.2011.08.009
Graham, A. (2011). The age and diversification of terrestrial new world ecosystems through Cretaceous and Cenozoic time. Am. J. Bot. 98, 336–351. doi: 10.3732/ajb.1000353
Graham, A., and Jarzen, D. M. (1969). Studies in Neotropical paleobotany. I. The oligocene communities of puerto rico. Ann. Mo. Bot. Gard. 56, 308–357. doi: 10.2307/2394849
Grau, J. (1983). “Life form, reproductive biology and distribution of the Californian/Chilean genus Cryptantha,” in Dispersal and Distribution, ed K. Kubitzki (Hamburg: Sonderbd. Naturwiss. Ver), 231–240.
Hammouda, S. A., Weigend, M., Mebrouk, F., Chacón, J., Bensalah, M., Ensikat, H. J., et al. (2016). Fossil nutlets of Boraginaceae from the continental Eocene of Hamada of Méridja (southwestern Algeria): the first fossil of the borage family in Africa. Am. J. Bot. 102, 2108–2115. doi: 10.3732/ajb.1500350
Hasenstab-Lehman, K. E., and Simpson, M. G. (2012). Cat's eyes and popcorn flowers: phylogenetic systematics of the genus Cryptantha s.l. (Boraginaceae). Syst. Bot. 37, 738–757. doi: 10.1600/036364412X648706
Heleno, R., and Vargas, P. (2015). How do islands become green? Glob. Ecol. Biogeogr. 24, 518–526. doi: 10.1111/geb.12273
Herrera, C. M. (1989). Seed dispersal by animals: a role in angiosperm diversification? Am. Nat. 133, 309–322. doi: 10.1086/284921
Higgins, S. I., Nathan, R., and Cain, M. L. (2003). Are long-distance dispersal events in plants usually caused by nonstandard means of dispersal? Ecology 84, 1945–1956. doi: 10.1890/01-0616
Kadereit, J. W., and Baldwin, B. G. (2012). Western Eurasian–western North American disjunct plant taxa: The dry-adapted ends of formerly widespread north temperate mesic lineages—and examples of long-distance dispersal. Taxon 61, 3–17.
Katoh, K., Misawa, K., Kuma, K., and Miyata, T. (2002). MAFFT: a novel method for rapid multiple sequence alignment based on fast Fourier transformation. Nucleic Acids Res. 30, 3059–3066. doi: 10.1093/nar/gkf436
Katoh, T., and Standley, D. M. (2013). MAFFT multiple sequence alignment software version 7: improvements in performance and usability. Mol. Biol. Evol. 30, 772–780. doi: 10.1093/molbev/mst010
Kearse, M., Moir, R., Wilson, A., Stones-Havas, S., Cheung, M., Sturrock, S., et al. (2012). Geneious basic: an integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 28, 1647–1649. doi: 10.1093/bioinformatics/bts199
Kim, K. J., Choi, K. S., and Jansen, R. K. (2005). Two chloroplast DNA inversions originated simultaneously during the early evolution of the sunflower family (Asteraceae). Mol. Biol. Evol. 22, 1783–1792. doi: 10.1093/molbev/msi174
Larson-Johnson, K. (2016). Phylogenetic investigation of the complex evolutionary history of dispersal mode and diversification rates across living and fossil Fagales. New Phytol. 209, 418–435. doi: 10.1111/nph.13570
Lavin, M., and Luckow, M. (1993). Origins and relationships of tropical North America in the context of the boreotropics hypothesis. Am. J. Bot. 80, 1–14. doi: 10.2307/2445114
Luebert, F., Cecchi, L., Frohlich, M. W., Gottschling, M., Guilliams, C. M., Hilger, H. H., et al. (2016). Familial classification of Boraginales. Taxon 65, 502–522. doi: 10.12705/653.5
Luebert, F., Couvreur, T. L. P., Gottschling, M., Hilger, H. H., Miller, J. S., and Weigend, M. (2017). Historical biogeography of Boraginales: west gondwanan vicariance followed by long-distance dispersal? J. Biogeogr. 44, 158–169. doi: 10.1111/jbi.12841
Magallón, S., Gómez-Acevedo, S., Sánchez-Reyes, L. L., and Hernández-Hernández, T. (2015). A metacalibrated time- tree documents the early rise of flowering plant phylogenetic diversity. New Phytol. 207, 437–453. doi: 10.1111/nph.13264
Manafzadeh, S., Salvo, G., and Conti, E. (2014). A tale of migrations from east to west: the Irano-Turanian floristic region as a source of Mediterranean xerophytes. J. Biogeogr. 41, 366–379. doi: 10.1111/jbi.12185
Mathur, A. K., and Mathur, U. B. (1983). Boraginaceae (Angiosperm) seeds and their bearing on the age of Lameta beds of Gujarat. Curr. Sci. India 54, 1070–1071.
Matzke, N. J. (2013). Probabilistic historical biogeography: new models for founder-event speciation, imperfect detection, and fossils allow improved accuracy and model-testing. Front. Biogeogr. 5, 242–248. Available online at: http://escholarship.org/uc/item/44j7n141
Matzke, N. J. (2014). Model selection in historical biogeography reveals that founder-event speciation is a crucial process in island clades. Syst. Biol. 63, 951–970. doi: 10.1093/sysbio/syu056
Matzke, N. J. (2016). Stochastic Mapping under Biogeographical Models. PhyloWiki BioGeoBEARS. Available Online at: http://phylo.wikidot.com/biogeobears#stochastic_mapping./journalabbrev (Accessed November, 2016).
McGlone, M. S. (2005). Goodbye Gondwana. J. Biogeogr. 32, 739–740. doi: 10.1111/j.1365-2699.2005.01278.x
Meudt, H. M., Prebble, J. M., and Lehnebach, C. A. (2015). Native New Zealand forget-me-nots (Myosotis, Boraginaceae) comprise a Pleistocene species radiation with very low genetic divergence. Plant Syst. Evol. 301, 1455–1471. doi: 10.1007/s00606-014-1166-x
Miller, M. A., Pfeiffer, W., and Schwartz, T. (2010). “Creating the CIPRES Science Gateway for inference of large phylogenetic trees,” in Proceedings of the Gateway Computing Environments Workshop (GCE), (New Orleans, LA), 1–8.
Nathan, R. (2006). Long-distance dispersal of plants. Science 313, 786–788. doi: 10.1126/science.1124975
Nathan, R., Schurr, F. M., Spiegel, O., Steinitz, O., Trakhtenbrot, A., and Tsoar, A. (2008). Mechanisms of long-distance seed dispersal. Trends Ecol. Evol. 23, 638–647. doi: 10.1016/j.tree.2008.08.003
Nazaire, M., Wang, X. Q., and Hufford, L. (2014). Geographic origins and patterns of radiation of Mertensia (Boraginaceae). Am. J. Bot. 101, 104–118. doi: 10.3732/ajb.1300320
Nixon, K. C., and Crepet, W. L. (1993). Late Cretaceous fossil flowers of ericalean affinity. Am. J. Bot. 80, 616–623. doi: 10.2307/2445430
Ovchinnikova, S. V., and Korolyuk, A. Y. (2016). New species of the genus Craniospermum (Boraginaceae) from Mongolia. Plant Life Asian Russia 3, 33–40.
Panero, J. L. (2016). Phylogenetic uncertainty and fossil calibration of Asteraceae chronograms. Proc. Natl. Acad. Sci. U.S.A. 113, E411. doi: 10.1073/pnas.1517649113
Parsons, W. T., and Cuthbertson, E. G. (2001). Noxious weeds of Australia, 2nd Edn. Collingwood, ON: CSIRO Publishing.
Pemberton, R. W., and Irving, D. W. (1990). Elaiosomes on weed seeds and the potential for myrmecochory in naturalized plants. Weed Sci. 38, 615–619.
Posada, D., and Buckley, T. R. (2004). Model selection and model averaging in phylogenetics: advantages of akaike information criterion and bayesian approaches over likelihood ratio tests. Syst. Biol. 53, 793–808. doi: 10.1080/10635150490522304
Posada, D., and Crandall, K. A. (1998). MODELTEST: testing the model of DNA substitution. Bioinformatics 14, 817–818. doi: 10.1093/bioinformatics/14.9.817
Quilichini, A., and Debussche, M. (2001). Seed dispersal and germination patterns in a rare Mediterranean island endemic (Anchusa crispa Viv. Boraginaceae). Acta Oecol. 21, 303–313. doi: 10.1016/S1146-609X(00)01089-4
Rambaut, A., Suchard, M. A., Xie, D., and Drummond, A. J. (2014). Tracer v1.6. Available online at: http://beast.bio.ed.ac.uk/Tracer
Ree, R. H., and Smith, S. A. (2008). Maximum likelihood inference of geographic range evolution by dispersal, local extinction, and cladogenesis. Syst. Biol. 57, 4–14. doi: 10.1080/10635150701883881
Renner, S. S., Foreman, D. B., and Murray, D. (2000). Timing transantarctic disjunctions in the Atherospermataceae (Laurales): evidence from coding and noncoding chloroplast sequences. Syst. Biol. 49, 579–591. doi: 10.1080/10635159950127402
Retief, E., and van Wyk, A. E. (2002). The genus Trichodesma (Boraginaceae: Boraginoideae) in southern Africa. Bothalia 32, 151–166. doi: 10.4102/abc.v32i2.479
Ricklefs, R. E., and Renner, S. S. (1994). Species richness within families of flowering plants. Evolution 48, 1619–1636. doi: 10.2307/2410252
Ridley, H. N. (1930). The Dispersal of Plants throughout the World. Ashford: Reeve & Company, Limited.
Ripma, L. A., Simpson, M. G., and Hasenstab-Lehman, K. (2014). Geneious! Simplified genome skimming methods for phylogenetic systematic studies: a case study in Oreocarya (Boraginaceae). Appl. Plant Sci. 2:1400062. doi: 10.3732/apps.1400062
Ronce, O. (2007). How does it feel to be like a rolling stone? Ten questions about dispersal evolution. Annu. Rev. Ecol. Evol. Syst. 38, 231–253. doi: 10.1146/annurev.ecolsys.38.091206.095611
Sanmartín, I., Wanntorp, L., and Winkworth, R. C. (2007). West wind drift revisited: testing for directional dispersal in the Southern Hemisphere using event-based tree fitting. J. Biogeogr. 34, 398–416. doi: 10.1111/j.1365-2699.2006.01655.x
Segal, R. (1966a). Taxonomic study of the fossil species of the genus Cryptantha (Boraginaceae). Southwest. Nat. 11, 205–210. doi: 10.2307/3669642
Segal, R. (1966b). Biorbia (Boraginaceae) in the central U. S. Pliocene. Univ. Kans. Sci. Bull. 46, 495–508.
Selvi, F., Coppi, A., and Cecchi, L. (2011). High epizoochorous specialization and low DNA sequence divergence in Mediterranean Cynoglossum (Boraginaceae): Evidence from fruit traits and ITS region. Taxon 60, 969–985.
Stevens, P. F. (2001 onward). Angiosperm Phylogeny Website, Version. Available online at: http://www.mobot.org/MOBOT/research/APWeb/ (Accessed February 28 2017).
Takahashi, M., Crane, P. R., and Manchester, S. R. (2002). Hironoia fusiformis gen. et sp. nov.; a cornalean fruit from the Kamikitaba locality (Upper Cretaceous, Lower Coniacian) in north- eastern Japan. J. Plant Res. 115, 463–473. doi: 10.1007/s10265-002-0062-6
Takhtajan, A. (1986). Floristic Regions of the World. Berkeley, CA: The University of California Press.
Taugourdeau-Lantz, J., and Rosset, C. (1966). Sur un nouveau microfossile incertae sedis de l'Oligocéne du bassin de Narbonne. Rev. Micropaleontol. 9, 186–191.
Thomasson, J. R. (1977). Late Cenozoic Grasses, Borages, and Hackberries from Southwestern Nebraska. Contributions to Geology from the University of Wyoming.
Thomasson, J. R. (1979). Late cenozoic grasses and other angiosperms from kansas, nebraska and colorado: biostratigraphy and relationships to living taxa. Kansas Geol. Surv. Bull. 218, 1–68.
Thomasson, J. R. (1987). Late miocene plants from northeastern nebraska. J. Palaeontol. 61, 1065–1079. doi: 10.1017/S0022336000029413
Tiffney, B. H., and Manchester, S. R. (2001). The use of geological and paleontological evidence in evaluating plant phylogeographic hypotheses in the northern hemisphere Tertiary. Int. J. Plant Sci. 162, S3–S17. doi: 10.1086/323880
Van der Pijl, L. (1982). Principles of Dispersal in Higher Plants, 3rd Edn. Berlin and New York, NY: Springer-Verlag.
Vargas, P., Heleno, R., Traveset, A., and Nogales, M. (2012). Colonization of the Galápagos Island by plants with no specific syndromes for long-distance dispersal: a new perspective. Ecography 35, 33–43. doi: 10.1111/j.1600-0587.2011.06980.x
Weigend, M., Gottschling, M., Selvi, F., and Hilger, H. H. (2010). Fossil and extant Western Hemisphere Boragineae, and the polyphyly of “Trigonotideae” Riedl (Boraginaceae: Boraginoideae). Syst. Bot. 35, 409–419. doi: 10.1600/036364410791638423
Weigend, M., Luebert, F., Gottschling, M., Couvreur, T. L. P., Hilger, H. H., and Miller, J. S. (2014). From capsules to nutlets – Phylogenetic relationships in the Boraginales. Cladistics 30, 508–518. doi: 10.1111/cla.12061
Weigend, M., Luebert, F., Selvi, F., Brokamp, G., and Hilger, H. H. (2013). Multiple origins for Hounds tongues (Cynoglossum, L.) and Navel seeds (Omphalodes Mill.) – the phylogeny of the borage family (Boraginaceae s.str.). Mol. Phylogenet. Evol. 68, 604–618. doi: 10.1016/j.ympev.2013.04.009
Weigend, M., Selvi, F., Thomas, D. C., and Hilger, H. H. (2016). “Boraginaceae,” in The Families and Genera of Vascular Plants, vol. 14. Flowering Plants: Eudicots; Aquifoliales, Boraginales, Bruniales, Dipsacales, Escalloniales, Garryales, Paracryphiales, Solanales (Except Convolvulaceae), Icacinaceae, Metteniusaceae, Vahliaceae, eds J. W. Kadereit and V. Bittrich (Heidelberg: Springer-Verlag GmbH), 41–102.
Zachos, J., Pagani, M., Sloan, L., Thomas, E., and Billups, K. (2001). Trends, rhythms, and aberrations in global climate 65 Ma to present. Science 292, 686–693. doi: 10.1126/science.1059412
Keywords: anemochory, atelechory, stochastic mapping methods, endozoochory, epizoochory, nautochory, long-distance dispersal, paleocene-eocene thermal maximum
Citation: Chacón J, Luebert F and Weigend M (2017) Biogeographic Events Are Not Correlated with Diaspore Dispersal Modes in Boraginaceae. Front. Ecol. Evol. 5:26. doi: 10.3389/fevo.2017.00026
Received: 21 December 2016; Accepted: 21 March 2017;
Published: 06 April 2017.
Edited by:
Marco A. Molina-Montenegro, University of Talca, ChileReviewed by:
Katy Morgan, University of New Orleans, USAAna M. C. Santos, University of Alcalá, Spain
Copyright © 2017 Chacón, Luebert and Weigend. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Juliana Chacón, amNoYWNvbnBAdW5pLWJvbm4uZGU=