 Sanjo Rose†
Sanjo Rose† Petra Sumasgutner
Petra Sumasgutner Arjun Amar
Arjun Amar- FitzPatrick Institute of African Ornithology, DST-NRF Centre of Excellence, University of Cape Town, Cape Town, South Africa
As human populations have increased globally, so too has the transformation of natural landscapes into more urban areas. Within Africa, population growth rates and urbanization rates are amongst the highest in the world, but the impacts of these processes on Africa's wildlife are largely un-explored. In this study, we focus on a recently established population of black sparrowhawks Accipiter melanoleucus in Cape Town, South Africa. Using long-term data (16 years), we investigate the relationship between the timing of breeding and breeding performance along an urban-rural habitat gradient. We found no evidence for a direct effect of urbanization on either timing of breeding or breeding performance. However, we did find some evidence for a differential seasonal decline in breeding performance dependent on the levels of urbanization. Thus, higher productivity was found in more urbanized habitats earlier in the season, but towards the end of the breeding season, birds in less urbanized habitats performed better. Our study represents one of the first to examine avian productivity in relation to urbanization in Africa, and also represents one of the first studies to suggest that seasonal declines in productivity may differ according to levels of urbanization. These results have considerable implications for potential changes in phenology or productivity for the regions' avifauna as African urbanization continues.
Introduction
Urbanization is a rapid and global process in which natural environments are transformed into new urban systems (Berry et al., 1998) which are characterized by an increase of unproductive sealed surfaces in highly disturbed fragmented landscapes (McDonnell and Pickett, 1990). By 2050, 66% of the global population is predicted to live in urban areas (United Nations, 2015). Urbanization significantly alters the physical, chemical, and biotic environment (McDonnell and Pickett, 1990). The resulting decrease of natural habitat forces ecosystems and species to adapt, avoid or tolerate these new conditions (Blair, 1996; Isaksson, 2015).
Key processes that contribute to the impact of urbanization on birds include vegetation changes, habitat fragmentation, introduction of exotic species, changes in food abundance and quality, and novel predator assemblages (Chace and Walsh, 2006). An urban gradient characterizes different levels of urban stressors, such as pollution (light, noise, and air), the abundance of novel predators like cats and corvids (e.g., Stracey, 2011), and of anthropogenic food resources that may be of lower quality (“urban junk food,” Pierotti and Annett, 2001; Schoech and Bowman, 2001). All of these factors can potentially affect demographic parameters of city-dwelling species (Isaksson, 2015). In order to anticipate the costs and benefits for birds living in urban areas, it is useful to quantify the relative breeding performance within urbanized areas compared to more natural habitats.
There is a growing body of literature comparing breeding performance of birds living in urban and rural habitats, with some general patterns emerging. The onset of breeding tends to be earlier in urban areas as a potential result of increased food availability (Chamberlain et al., 2009), the urban heat island effect (i.e., higher temperatures in the city center, Shochat et al., 2006; since the beginning of incubation can be temperature dependent, Visser et al., 2009) and light pollution (Spoelstra and Visser, 2014). Clutch sizes tend to be smaller and nestling conditions tend to be poorer in urban vs. natural habitats (reviewed by Chamberlain et al., 2009). However, these studies have mostly been conducted in the Northern Hemisphere with very different climatic conditions. Despite an increased focus on urban avian ecology, as far as we know no studies have yet focused on these questions for any bird species in Africa.
Many bird species show seasonal declines in breeding performance during the course of a breeding season (e.g., Both et al., 2004; Grüebler and Naef-Daenzer, 2010), and this has also been found for some urban nesting species (Chamberlain et al., 2009), including urban raptors (e.g., Sumasgutner et al., 2014a). The explanation for these patterns often focuses on declines in food availability that might also influence sibling competition (Gil et al., 2008), which may in theory vary across levels of urbanization. If so, it might be expected that the patterns of such seasonal declines might vary across an urban gradient. In the only study to explore this issue so far, Wawrzyniak et al. (2015) found seasonal declines in clutch size of blue tits (Parus major) nesting in urban habitats but not for those nesting in forests.
Because raptors belong to the upper trophic level, they can be particularly susceptible to changes in the environment (Newton, 1998). Despite this, some raptors are increasingly associated with urbanized areas (e.g., Falco peregrinus, Cade and Bird, 1990; F. columbarius, Sodhi et al., 1992; F. tinnunculus, Village, 1990; Charter et al., 2007; Accipiter cooperii, Boal and Mannan, 1999, 2000; A. gentilis, Rutz, 2008). This is potentially due to their relatively high mobility that could allow them to escape from some of the limitations of urban areas (Chace and Walsh, 2006), while still benefitting from the advantages, such as lower direct persecution (Rutz, 2008) and novel resources like higher abundance of avian prey or nesting sites (e.g., Palomino and Carrascal, 2007; Cavalli et al., 2014; Rullman and Marzluff, 2014; Sumasgutner et al., 2014b). However, some rodent specialists such as Barn owls Tyto alba or Eurasian kestrels F. tinnunculus show decreased fledgling success with increasing urbanization, probably due to decreased prey availability connected to sensory interference due to noise pollution for owls (Senzaki et al., 2016), or nocturnal rodents being harder to locate for a diurnal raptor in an urban environment (Hindmarch et al., 2014; Sumasgutner et al., 2014a). Dietary requirements may be an important factor in determining the success of an urban raptor, specifically the degree of specialization in their diet choice. Highly specialized raptors may have to spend more energy and time seeking preferred prey, whereas generalists may have more flexibility in coping with fluctuating food availability (see Terraube et al., 2011 for a natural setting).
In this study we focus on the black sparrowhawk Accipiter melanoleucus in South Africa, a raptor that colonized the city of Cape Town in the last two decades after expanding its distribution range from the North-East into the South-West of Southern Africa (Amar et al., 2014). Black sparrowhawks appear to have adapted well to urban areas, with more than 50 breeding pairs (Martin et al., 2014b) within our study area. Together with the colonization of the urban habitat, the species extended its breeding season toward earlier egg-laying, resulting in a breeding season of 9 months (Mar-Nov; Martin et al., 2014b). Potentially contributing to the success of the species in this area, the health of black sparrowhawks' chicks does not appear to be negatively influenced by levels of urbanization (Suri et al., 2017, but see van Velden et al., 2017 for a specific Knemidokoptes mite infection in adults). This might be linked to the high abundance of suitable prey species across all habitat types that may buffer any potential negative health effects of urban stressors (Suri et al., 2017). Building on these results, our study aims to explore how the breeding performance of urban black sparrowhawks varies with urbanization in Cape Town. Using a long-term study (16 years), we investigated the timing of breeding and breeding performance along an urban-rural habitat gradient. Additionally, we specifically explore whether the established relationship between timing of breeding and breeding performance (Martin et al., 2014b) differed depending on levels of urbanization. This question is particularly relevant to our study area, since it is unique within a South African context in having an extended 9 months long breeding season (Martin et al., 2014b). Given the results of several other studies exploring timing of breeding, we predicted the onset of breeding to be earlier in more urbanized areas. The expected relationship between urbanization and breeding performance is harder to predict, however, given that Suri et al. (2017) tended to find higher abundance of doves and pigeons in more urbanized habitats, we might expect higher productivity in more urban areas. Likewise the precise relationship between timing of breeding and breeding performance in relation to urbanization was difficult to predict, but if urbanized areas do have higher or more regular prey availability, the established seasonal declines in breeding performance may be somewhat buffered within more urbanized areas.
Methods
Study Site and Species
This study focused on the resident population of black sparrowhawks in Cape Town, Western Cape, South Africa (Figure 1). The study area covers various habitats over 595 km2 (Martin et al., 2014b), including heavily urbanized areas, plantations of exotic tree stands, patches of Afromontane forest and indigenous fynbos vegetation. Cape Town experiences a Mediterranean climate, with an average annual rainfall of 788 mm and average monthly temperatures ranging from 15 to 30°C. This population breeds mainly during the winter months, which coincides with the rainy season (Amar et al., 2014; Martin et al., 2014b). The population has been actively monitored since 2000 (Amar et al., 2014).
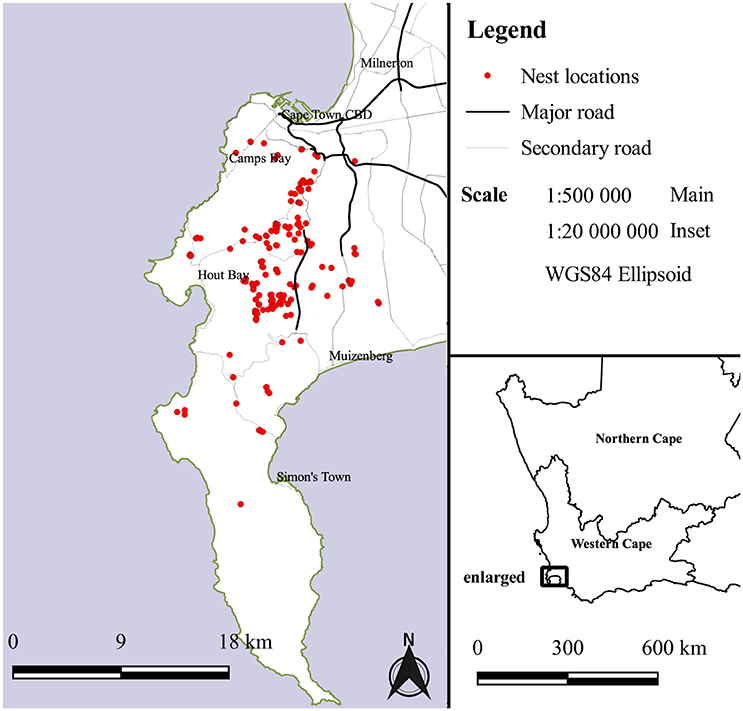
Figure 1. Distribution of black sparrowhawk nest localities on the Cape Peninsula over a 16-year monitoring period. Insert situates the Cape Peninsula in the South African context.
Systematic monitoring by a team of experienced researchers occurred during the breeding season from March to November. Territories were located by surveying suitable stands of trees. Occupancy was confirmed by activity indicators such as calling, whitewash, prey remains or nest structures. Following confirmation of breeding attempts, pairs are then monitored approximately weekly. About 90% of the population is individually color ringed, allowing individual identification of many breeding pairs (see Martin et al., 2014a for further details). Adults were trapped on territories using a bal-chatri trap baited with a live white “racing” pigeon, Columba livia (Berger and Mueller, 1959; see Ethical Note in Martin et al., 2014a for details). Chicks were ringed when they were 3.5–4.5 weeks old. Age of chicks was estimated visually by the extent of wing and tail feather growth (using reference photographs from nest cameras). This study uses data collected between 2000 and 2015.
Defining the Urban Gradient
Quantification of the urban gradient was based on a circular buffer with a radius of 2,000 meters (m) assigned to each nest; the chosen scale was based on previous work in which adult males with global positioning system (GPS) tags were tracked and their home range sizes during the breeding season where determined (Sumasgutner et al., 2016b). Urban cover was calculated based on land cover classes from the 2013–2014 South African National land-cover dataset produced by GEOTERRA (Department of Environmental Affairs, 2015). This land cover dataset compiled 72 land types at a 30 m resolution, from which sealed, unproductive areas of land were classified as “urban.” Percentage urban cover was derived for each nest buffer using the packages raster (Hijmans, 2015), sp (Pebesma and Bivand, 2005; Bivand et al., 2013), and rgdal (Bivand et al., 2016). Figure 2 illustrates the binary urban/non-urban land covers as compared with aerial photographs.
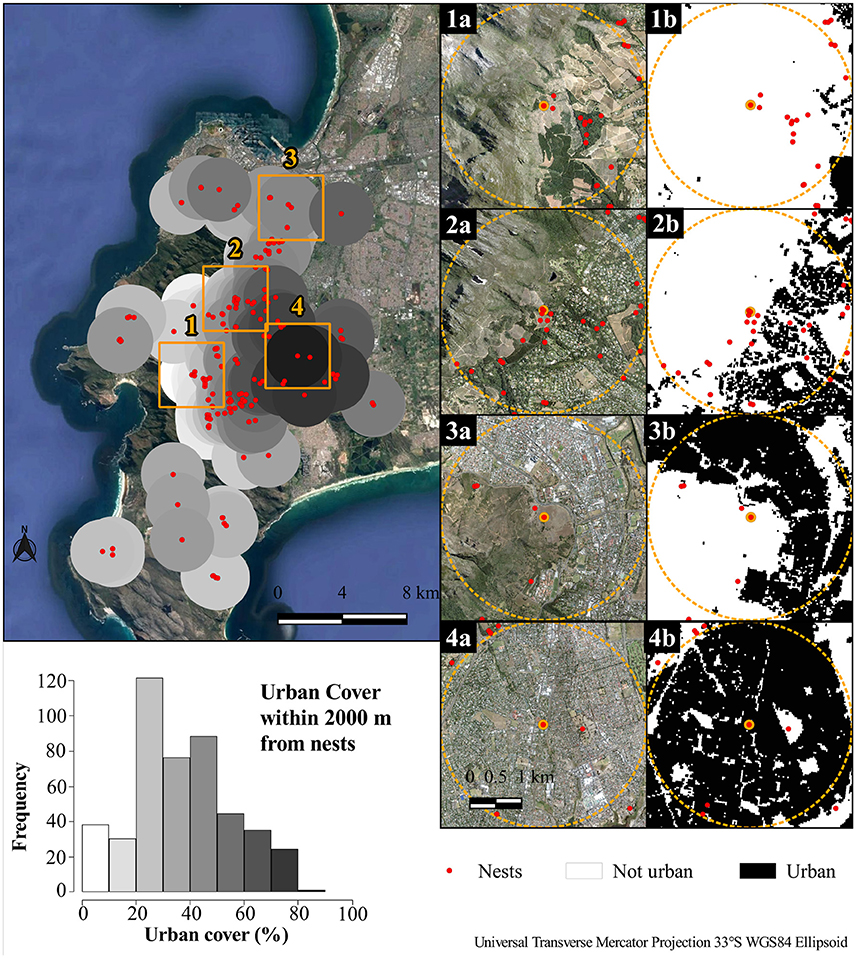
Figure 2. Percentage urbanization for each black sparrowhawk nest on the Cape Peninsula over a 16-year monitoring period (top left), frequency plot of the distribution of nests along an urban gradient (bottom left), illustrations of representative individual nest locations with aerial imagery (center column) and land cover reduced to urban/non-urban binary (right column). Urban cover is 1.4, 12.7, 47.1, and 87.0%, respectively, for nests 1, 2, 3, and 4. Data from Department of Department of Environmental Affairs (2015); Google Earth Pro (2016), and City of Cape Town (2016).
Statistical Analysis
The “lay month” (the only accurate measurement of the timing of breeding possible) for successful nests is backdated from the age of the chicks; we used the incubation behavior (female sitting low on nest) to time the onset of breeding for unsuccessful nests. “Breeding success” is a binomial variable distinguishing between successful and unsuccessful nests, “brood size” only includes successful nests (1-3 chicks that fledged) and “productivity” all active nests where 0–3 chicks fledged. All analyses were performed in relation to urbanization using Linear Mixed Models (LMMs) for the timing of breeding and Generalized Linear Mixed Models (GLMMs) for breeding performance, in R studio (R Core Team, 2016) with the packages nlme (Pinheiro et al., 2014), lme4 (Bates et al., 2015), MASS (Venables and Ripley, 2002), lattice (Deepayan, 2008), ggplots2 (Wickham, 2009) effects (Fox, 2003), and car (Fox and Weisberg, 2011). All quantitative variables were scaled in order to bring the variables to comparable dimensions to interpret effect sizes for interaction terms. We tested the dispersion of our response variables using the “qcc.overdispersion.test” in the qcc package (Scrucca, 2004). Residual distributions of the models were inspected visually to assess model fit.
Unless otherwise stated (when using successful nests only), the sample size in all following analyses was 456 broods surveyed over 16 years starting in 2000. Year, territory ID, and nest ID were included as random terms to account for pseudoreplication arising from same nests sampled repeatedly within and between years (nest ID), and breeding pairs occupying territories over several years (territory ID), and multiple measures from different nests within the same year (year). All analyses were conducted at the nest level since some pairs build multiple nests in different locations within and between seasons, resulting in varying urban scores for some individuals. The lay month (Gaussian distribution) was modeled as the response variable with the urban gradient as the explanatory variable. For the other analyses examining breeding performance, we accounted for the timing of breeding (using lay month) as a quadratic term and explored the interaction of timing of breeding with the urban gradient. We included the lay month as linear and quadratic term to investigate a potential polynomial relationship. Error structures for the GLMMs were: negative binomial (productivity), Poisson (brood size) and binomial (breeding success). Least squares means (post-hoc contrasts for mixed-models, Lenth and Hervé, 2015) are not yet implemented for interaction terms between continuous variables. Thus, to understand the nature of the relationship between the timing of breeding and levels of urbanization better, we performed additional tests whereby we selected the earlier 25% of nests (egg-laying Mar-May, n = 123) to see if there is a significant difference in the breeding performance in relation to the urban gradient. Then we repeated this analysis for the latest 25% of nests (egg-laying Sep-Nov, n = 120); see Tate et al. (2016) for a similar approach. Throughout, reported χ2 and p-values are based on an ANOVA Table of Deviance using Type II Wald χ2 tests for mixed models.
Ethical Note
The University of Cape Town's Science Faculty Animal Ethics Committee has approved all the procedures required for the long-term study of the black sparrowhawk population in Cape Town (permit number: 2012/V37/AA). Furthermore, all procedures were carried out under a Cape Nature Hunting Permit (No. 0035-AAA004-00428), a South African National Parks Permit and a SAFRING ringing permit (no. 1439).
Results
Urban Gradient
The urban gradient around each nest ranged from 87% to 1.4%. See Figure 2 for representations of (1) 1.4%, (2) 12.7%, (3) 47.1%, and (4) 87.0% urban cover, and the frequency of different urbanization scores for black sparrowhawk nests over the 16-year monitoring period.
Timing of Breeding and Breeding Performance along the Urban Gradient
Timing of breeding did not differ in relation to the urban gradient (Table 1, Figure 3), nor was there a direct relationship between breeding performance and the urbanization gradient term. However, an interesting interaction was noted between timing of breeding and urbanization in relation to breeding performance (Table 1, Figures 4A,C). Firstly, for pairs with nests in the most urbanized areas (Figure 4A, blue dotted line), we found that productivity declined strongly as the season progressed, whereas this decline was less steep for pairs in less urban areas (Figure 4A, black solid line), indicating that the timing of breeding had greatest influence on productivity in more urbanized areas. This interaction term shifts after approximately one third of the breeding season (June/July). A very similar relationship was also seen for breeding success (Figure 4C). However, the interaction term was statistically not significant, but suggested a tendency for differential seasonal declines in relation to urbanization (productivity GLMM: χ2 = 3.45, P = 0.063; breeding success GLMM: χ2 = 3.20, P = 0.074). This apparent relationship was not observed for brood size (Figure 4B, successful nests only, 1–3 chicks) where we found slightly more chicks fledging in urban areas throughout the season and less chicks fledging in more rural areas. Thus, our results suggest that the interaction between timing of breeding and urban gradient was due to nest failures, rather than the number of fledglings produced in successful nests. When splitting the data for early nests vs. late nests these observations were confirmed: we found slightly higher productivity in early nests in more urbanized areas (χ2 = 3.41, P = 0.065), but no such difference in late nests (χ2 = 2.20, P = 0.138). Additionally, we found significantly higher probability of nest failure in more urbanized areas later in the season (χ2 = 5.28, P = 0.022; vs. breeding success in early nests: χ2 = 2.74, P = 0.098).
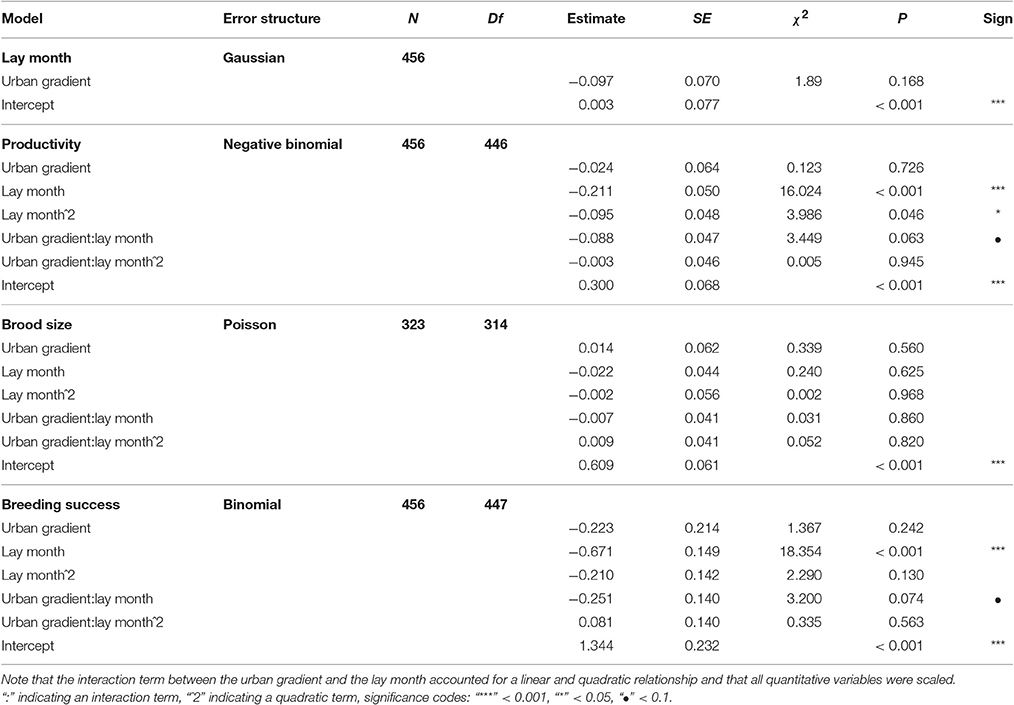
Table 1. LMMs (for lay month) and GLMMs investigating the relationship between breeding parameters (productivity, brood size, and breeding success) and the urban gradient.
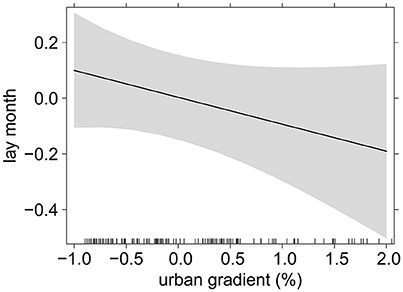
Figure 3. The relationship between the timing of breeding (lay month, scaled) and the urban gradient (% of urban cover, scaled), based on predicted values of LMM, 95% CIs in shaded gray; black bars on the x-axis represent sample size. Model details in Table 1 (χ2 = 1.89, P = 0.17).
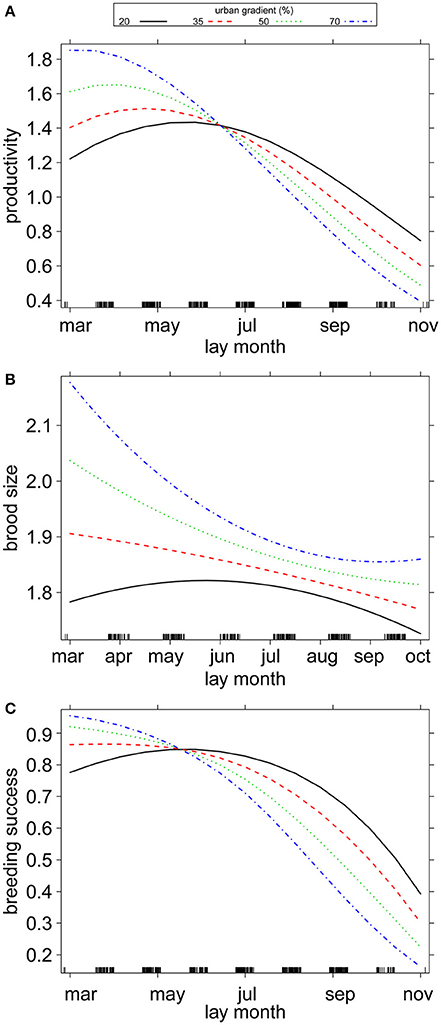
Figure 4. Plots showing the relationship between the urban gradient (% urban cover) and (A) productivity (χ2 = 3.45, P = 0.06); (B) brood size (χ2 = 0.03, P = 0.86); and (C) breeding success (χ2 = 3.20, P = 0.07), based on predicted values of GLMMs, black bars on the x-axis represent sample size. Note that all quantitative variables were scaled. Model details in Table 1.
Discussion
The most fundamental result from our analysis was that the extent of urbanization surrounding nest sites did not seem to have a strong influence on breeding performance. Thus, despite many additional stressors that may come from breeding within a more urbanized environment (Isaksson, 2015), black sparrowhawks appear able to cope with these challenges and produced similar number of chicks irrespective of the levels of urbanization around their nest sites. This result therefore adds to the findings of Suri et al. (2017), which explored influences of urbanization on chick health for this same black sparrowhawk population, and found no negative effect on a range of health parameters. Suri et al. (2017) suggested that the main prey species (doves and pigeons, Streptopelia semitorquata, and Columba livia) were plentiful across all habitats, and this abundance of prey may explain why there were no negative impacts of urbanization either on chick health and ultimately on the population's productivity. Thus, from these two studies it does seem that the species is able to cope well with the urban environment within the Cape Town region.
Contrary to our expectations, there was no direct relationship between the percentage of urban cover and timing of breeding. Many urban species are known to start breeding earlier compared to their rural counterparts (see reviews by Chamberlain et al., 2009), but these studies were located in the Northern hemisphere where the urban heat island effect might be beneficial (but see advanced incubation in Australian magpies Gymnorhina tibicen, Rollinson and Jones, 2002; and earlier first nest attempts for White-winged chough Corcorax melanorhamhos, Beck and Heinsohn, 2006). However, for black sparrowhawks in South Africa breeding coincides with the colder periods of the year, and within Cape Town with the rainy season (Martin et al., 2014a).
One of the most interesting findings from this study was the apparent difference in the degree of seasonal decline in breeding performance depending on the surrounding levels of urbanization. Declining productivity as breeding seasons progress is a well-known phenomenon, that has been documented in many bird species mostly in the Northern hemisphere (e.g., Both et al., 2004; Grüebler and Naef-Daenzer, 2010), but increasingly also in the Southern hemisphere (e.g., Garcia-Heras et al., 2016; Murgatroyd et al., 2016), including previously in our study system (Martin et al., 2014b). In addition to this pattern, however, we also found a convincing indication that the level of this decline differed depending on the extent of urbanization surrounding the nests. Thus, it appears that earlier on in the season productivity (the number of chicks produced) is highest for pairs breeding in more urban habitats, but later on this advantage disappears. In fact, when focussing on breeding success, we found significantly higher probability of nest failure in more urbanized areas later in the season.
Several other recent studies have documented differences in seasonal declines in breeding parameters between habitats, but usually on a larger geographical scale. For example, a similar result was noted for another South African raptor, the black harrier Circus maurus. Although the nests in that study were located in the different habitats, they were also often separated by large distances. The differences in seasonal declines in clutch size and to a lesser degree productivity that were found in that study were thought to be driven by differences in weather conditions between the areas (Garcia-Heras et al., 2016). Likewise Zárybnická et al. (2015) found different levels of seasonal declines between Tengmalm's owl Aegolius funereus breeding in either boreal or temperate forests. Here the differences were attributed to varying prey availability (small mammals) over the season influencing nestling mortality and failure rates. The only other study, as far as we know, to explore this issue in relation to urbanization, is the study by Wawrzyniak et al. (2015) on blue tits (Parus major). This study found that clutch size tended to decline during the breeding season in an urban setting but not within forest habitat. They also attributed this result largely to better prey availability within the forest habitat. Thus, in these other studies changes in either weather or prey were thought to be the drivers for differences in seasonal declines.
What might then be responsible for the relationship witnessed in our current study? It could be that food is more abundant in urban habitat earlier in the season. This could be the case if timing of breeding of avian prey species in urban areas follows the more usually trend seen for birds (Chamberlain et al., 2009). Thus prey may be more available, especially inexperienced juvenile prey, earlier on in the season, which may account for the higher productivity of earlier breeding black sparrowhawks within urban habitats compared to their rural conspecifics. Later on in the season, the situation may be reversed due to the later breeding of prey species in the more rural habitats. Unfortunately we have no information on the timing of breeding on any of the key prey species in the area in relation to urbanization nor are we aware of a seasonal variation in prey abundance (Suri et al., 2017). Additionally, we found that differences in productivity were principally driven by nest failures rather than differences in brood sizes of successful nests. Thus, the mechanism for the differential seasonal decline may not be food related, since prey abundance would be expected to impact brood sizes rather than to induce complete nest failures. Alternatively, we know from other studies that one of the main causes of nest failure in this black sparrowhawks is nest usurpation by Egyptian geese (Alopochen aegyptiaca, Curtis et al., 2007) and that this increases later in the breeding season (Sumasgutner et al., 2016a). Thus it may be that usurpation in more prevalent in more urbanized areas, since both species have recently expanded their South African range into the urban and suburban habitats of Cape Town (see Mangnall and Crowe, 2002 on Egyptian geese). This might further explain the significantly lower breeding success in more urbanized habitats later in the season, since the breeding season of Egyptian geese peaks later in the year (MacLean, 1997; Carboneras and Kirwan, 2016); when also the threat of nest usurpation increases (Sumasgutner et al., 2016a).
Conclusion
In this study we found no evidence that black sparrowhawks suffer from a decrease in breeding performance as a result of living in an urban environment, but timing of breeding was important, specifically in more urbanized areas. To summarize, black sparrowhawks appear to have successfully navigated various steps outlined by Møller (2014) of becoming an urban raptor species, which was likely facilitated by a reliable food supply (Suri et al., 2017) and abundant nesting opportunities in the form of alien tree species (Malan and Robinson, 1999).
Author Contributions
SR and PS contributed equally to this manuscript (shared first authorship). The original idea and study design came from PS and AA. AK performed most of the field work; help by others is accordingly acknowledged. SR and PS analyzed the data. The manuscript was prepared by SR, PS, and AA.
Funding
The Black Sparrowhawk project is funded by the DST-NRF Centre of Excellence. SR was supported by the NRF and PS by the Claude Leon Foundation.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We are grateful to the Black Sparrowhawk team—Johan Koeslag, Mark Cowen, Odette Curtis, Gareth Tate, Margaret MacIver and Sharon Pryce—for the long-term population monitoring, data collection and extensive support in the field.
References
Amar, A., Koeslag, A., Malan, G., Brown, M., and Wreford, E. (2014). Clinal variation in the morph ratio of Black Sparrowhawks Accipiter melanoleucus in South Africa and its correlation with environmental variables. Ibis 156, 627–638. doi: 10.1111/ibi.12157
Bates, D., Maechler, M., Bolker, B., and Walker, S. (2015). Fitting linear mixed-effects models using lme4. J. Stat. Softw. 67, 1–48. doi: 10.18637/jss.v067.i01
Beck, N. R., and Heinsohn, R. (2006). Group composition and reproductive success of cooperatively breeding white-winged choughs (Corcorax melanorhamphos) in urban and non-urban habitat. Austral Ecol. 31, 588–596. doi: 10.1111/j.1442-9993.2006.01589.x
Berger, D. D., and Mueller, H. C. (1959). The bal-chatri: a trap for the birds of prey. Bird-Banding 30:18e26. doi: 10.2307/4510726
Berry, M., Bock, C., and Haire, S. (1998). Abundance of diurnal raptors on open space grasslands in an urbanized landscape. Condor 100, 601–608. doi: 10.2307/1369742
Bivand, R. S., Pebesma, E., and Gomez-Rubio, V. (2013). Applied Spatial Data Analysis with R. 2nd Edn. New York, NY: Springer. doi: 10.1007/978-1-4614-7618-4
Bivand, R., Keitt, T., and Rowlingson, B. (2016). rgdal: Bindings for the Geospatial Data Abstraction Library. R package version 1.1–10.
Blair, R. B. (1996). Land use and avian species diversity along an urban gradient. Ecol. Appl. 6, 506–519. doi: 10.2307/2269387
Boal, C. W., and Mannan, R. W. (1999). Comparative breeding ecology of Cooper's hawks in urban and exurban areas of Southeastern Arizona. J. Wildl. Manag. 63, 77–84. doi: 10.2307/3802488
Boal, C. W., and Mannan, R. W. (2000). Cooper's hawks in urban and exurban areas: a reply. J. Wildl. Manag. 64, 601–604. doi: 10.2307/3803259
Both, C., Artemyev, A. V., Blaauw, B., Cowie, R. J., Dekhuijzen, A. J., Eeva, T., et al. (2004). Large–scale geographical variation confirms that climate change causes birds to lay earlier. Proc. R. Soc. B 271, 1657–1662. doi: 10.1098/rspb.2004.2770
Cade, T. J., and Bird, D. M. (1990). Peregrine falcons, Falco peregrinus, nesting in an urban environment: a review. Can. Field Nat. 104, 209–218.
Carboneras, C., and Kirwan, G. M. (2016). “Egyptian goose (Alopochen aegyptiaca),” in Handbook of the Birds of the World Alive, eds A. del Hoyo, J. Elliott, J. Sargatal, D. A. Christie, and E. de Juana (Barcelona: Lynx Edicions).
Cavalli, M., Baladron, A. V., Isacch, J. P., Martinez, G., and Bo, M. S. (2014). Prey selection and food habits of breeding Burrowing Owls (Athene cunicularia) in natural and modified habitats of Argentine pampas. Emu 114, 184–188. doi: 10.1071/MU13040
Chace, J. F., and Walsh, J. J. (2006). Urban effects on native avifauna: a review. Landscape Urban Plan. 74, 46–69. doi: 10.1016/j.landurbplan.2004.08.007
Chamberlain, D. E., Cannon, A. R., Toms, M. P., Leech, D. I., Hatchwell, B. J., and Gaston, K. J. (2009). Avian productivity in urban landscapes: a review and meta analysis. Ibis 151, 1–18. doi: 10.1111/j.1474-919X.2008.00899.x
Charter, M., Izhaki, I., Bouskila, A., Leshem, Y., and Penteriani, V. (2007). Breeding success of the Eurasian Kestrel (Falco tinnunculus) nesting on buildings in Israel. J. Raptor Res. 41, 139–143. doi: 10.3356/0892-1016(2007)41[139:BSOTEK]2.0.CO;2
City of Cape Town (2016). Aerial Photography February 2015. Open Data Portal. Available online at: https://web1.capetown.gov.za (accessed September 7, 2016).
Curtis, O. E., Hockey, P. A. R., and Koeslag, A. (2007). Competition with Egyptian Geese Alopochen aegyptiaca overrides environmental factors in determining productivity of Black Sparrowhawks Accipiter melanoleucus. Ibis 149, 502–508. doi: 10.1111/j.1474-919X.2007.00675.x
Department of Environmental Affairs (2015). DEA National Landcover (TIFF). 30 metre. Available online at: http://bgis.sanbi.org/SpatialDataset/Detail/496 (accessed October 9, 2016).
Fox, J. (2003). Effect displays in R for generalised linear models. J. Stat. Softw. 8, 1–27. doi: 10.18637/jss.v008.i15
Fox, J., and Weisberg, S. (2011). An R Companion to Applied Regression. 2nd Edn. Thousand Oaks, CA: Sage.
Garcia-Heras, M., Arroyo, B., Mougeot, F., Amar, A., and Simmons, R. E. (2016). Does timing of breeding matter less where the grass is greener? Seasonal declines in breeding performance differ between regions in an endangered endemic raptor. Nat. Conserv. 15, 23–45. doi: 10.3897/natureconservation.15.9800
Gil, D., Bulmer, E., Celis, P., and López-Rull, I. (2008). Adaptive developmental plasticity in growing nestlings: sibling competition induces differential gape growth. Proc. R. Soc. B. 275, 549–554. doi: 10.1098/rspb.2007.1360
Google Earth Pro (2016). Cape Town Region, South Africa. Available online at: http://www.google.com/earth/index.html (accessed September 7, 2016).
Grüebler, M. U., and Naef-Daenzer, B. (2010). Fitness consequences of timing of breeding in birds: date effects in the course of a reproductive episode. J. Avian Biol. 41, 282–291. doi: 10.1111/j.1600-048X.2009.04865.x
Hindmarch, S., Krebs, E. A., Elliott, J., and Green, D. J. (2014). Urban development reduces fledging success of Barn Owls in British Columbia, Canada. Condor 116, 507–517. doi: 10.1650/CONDOR-13-052.1
Isaksson, C. (2015). Urbanisation, oxidative stress and inflammation: a question of evolving, acclimatizing or coping with urban environmental stress. Funct. Ecol. 29, 913–923. doi: 10.1111/1365-2435.12477
MacLean, G. L. (1997). “Egyptian goose,” in The Atlas of Southern African Birds, eds J. A. Harrison and M. Cherry (Johannesburg: BirdLife South Africa), 122–3.
Malan, G., and Robinson, E. R. (1999). The diet of the Black Sparrowhawk Accipiter melanoleucus (Aves: Accipitridae) in South Africa: hunting columbids in man-altered environments. Durban Museum Novitates 24, 43–47.
Mangnall, M. J., and Crowe, T. M. (2002). Population dynamics and the physical and financial impacts to cereal crops of the Egyptian goose Alopochen aegyptiacus on the Agulhas Plain, Western Cape, South Africa. Agr. Ecosyst. Environ. 90, 231–246. doi: 10.1016/s0167-8809(01)00215-8
Martin, R. O., Koeslag, A., Curtis, O., and Amar, A. (2014a). Fidelity at the frontier: divorce and dispersal in a newly colonized raptor population. Anim. Behav. 93, 59–68. doi: 10.1016/j.anbehav.2014.04.018
Martin, R. O., Sebele, L., Koeslag, A., Curtis, O., Abadi, F., and Amar, A. (2014b). Phenological shifts assist colonisation of a novel environment in a range-expanding raptor. Oikos 123, 1457–1468. doi: 10.1111/oik.01058
McDonnell, M. J., and Pickett, S. T. A. (1990). Ecosystem structure and function along urbanrural gradients: an unexploited opportunity for ecology. Ecology 71, 1232–1237. doi: 10.2307/1938259
Møller, A. P. (2014). “Behavioural and ecological predictors of urbanization,” in Avian Urban Ecology, eds D. Gil and H. Brumm (Oxford: Oxford University Press), 54–68.
Murgatroyd, M., Underhill, L. G., Rodrigues, L., and Amar, A. (2016). The influence of agriculture transformation on the breeding performance of a top predator: Verreaux's Eagles in contrasting land use areas. Condor 118, 238–252. doi: 10.1650/CONDOR-15-142.1
Palomino, D., and Carrascal, L. M. (2007). Habitat associations of a raptor community in a mosaic landscape of Central Spain under urban development. Landsc. Urban Plan. 83, 268–274. doi: 10.1016/j.landurbplan.2007.04.011
Pierotti, R., and Annett, C. (2001). “The ecology of western gulls in habitats varying in degree of urban influence,” in Avian Ecology and Conservation in an Urbanizing World, eds J. M. Marzluff, R. Bowman, and R. Donnelly (Norwell, MA: Kluwer Academic Publishers), 307–329.
Pinheiro, J., Bates, D., DebRoy, S., and Sarkar, D. (2014). nlme: Linear and Nonlinear Mixed Effects Models. R package version 3.1–118.
R Core Team (2016). R: A Language and Environment for Statistical Computing. Vienna: R Foundation for Statistical Computing.
Rollinson, D. J., and Jones, D. N. (2002). Variation in breeding parameters of the Australian magpie Gymnorhina tibicen in suburban and rural environments. Urban Ecosyst. 6, 257–269. doi: 10.1023/B:UECO.0000004826.52945.ed
Rullman, S., and Marzluff, J. M. (2014). Raptor presence along an urban-wildland gradient: influences of prey abundance and land cover. J. Raptor Res. 48, 257272. doi: 10.3356/JRR-13-32.1
Rutz, C. (2008). The establishment of an urban bird population. J Anim Ecol. 77, 1008–1019. doi: 10.1111/j.1365-2656.2008.01420.x
Schoech, S. J., and Bowman, R. (2001). “Variation in the timing of breeding between suburban and wildland Florida scrub jays: do physiologic measures reflect different environments?,” in Avian Ecology and Conservation in an Urbanizing World, eds J. M. Marzluff, R. Bowman, and R. Donnelly (Norwell, MA: Kluwer Academic Publishers), 289–306.
Scrucca, L. (2004). qcc: An R package for Quality Control Charting and Statistical Process Control. R News 4/1, 11–17.
Senzaki, M., Yamaura, Y., Francis, C. D., and Nakamura, F. (2016). Traffic noise reduces foraging efficiency in wild owls. Sci. Rep. 6:30602. doi: 10.1038/srep30602
Shochat, E., Warren, P. S., Faeth, S. H., McIntyre, N. E., and Hope, D. (2006). From patterns to emerging processes in mechanistic urban ecology. Trends Ecol. Evol. 21, 187–191. doi: 10.1016/j.tree.2005.11.019
Sodhi, N. S., James, P. C., Warkentin, I. G., and Oliphant, L. W. (1992). Breeding ecology of urban merlins (Falco columbarius). Can. J. Zool. 70, 1477–1483. doi: 10.1139/z92-204
Spoelstra, K., and Visser, M. E. (2014). “The impact of artificial light on avian ecology,” in Avian Urban Ecology, 1st Edn., eds D. Gil and H. Brumm (Oxford: Oxford University Press), 20–28.
Sumasgutner, P., Nemeth, E., Tebb, G., Krenn, H. W., and Gamauf, A. (2014a). Hard times in the city - attractive nest sites but insufficient food supply lead to low reproduction rates in a bird of prey. Front. Zool. 11:48. doi: 10.1186/1742-9994-11-48
Sumasgutner, P., Schulze, C. H., Krenn, H. W., and Gamauf, A. (2014b). Conservation related conflicts in the nest-site selection of the Eurasian Kestrel (Falco tinnunculus) and the distribution of its avian prey. Landsc. Urban Plann. 127, 94–103. doi: 10.1016/j.landurbplan.2014.03.009
Sumasgutner, P., Millán, J., Curtis, O., Koelsag, A., and Amar, A. (2016a). Is multiple nest building an adequate strategy to cope with inter-species nest usurpation? BMC Evol. Biol. 16:97. doi: 10.1186/s12862-016-0671-7
Sumasgutner, P., Tate, G. J., Koeslag, A., and Amar, A. (2016b). Seasonal home ranges of Black Sparrowhawks (Accipiter melanoleucus) breeding in an urban environment. Bird Study 63, 430–435. doi: 10.1080/00063657.2016.1214814
Suri, J., Sumasgutner, P., Hellard, E., Koeslag, A., and Amar, A. (2017). Stability in prey abundance may buffer Black Sparrowhawks from health impacts of urbanisation. Ibis 159, 38–54. doi: 10.1111/ibi.12422
Stracey, C. M. (2011). Resolving the urban nest predator paradox: the role of alternative foods for nest predators. Biol. Conserv. 144, 1545–1552. doi: 10.1016/j.biocon.2011.01.022
Tate, G. J., Bishop, J. M., and Amar, A. (2016). Differential foraging success across a light level spectrum explains the maintenance and spatial structure of colour morphs in a polymorphic bird. Ecol. Lett. 19, 679–686. doi: 10.1111/ele.12606
Terraube, J., Arroyo, B., Madders, M., and Mougeot, F. (2011). Diet specialisation and foraging efficiency under fluctuating vole abundance: a comparison between generalist and specialist avian predators. Oikos 120, 234–244. doi: 10.1111/j.1600-0706.2010.18554.x
United Nations, A. (2015). World Urbanization Prospects: The 2014 Revision. New York, NY: United Nations.
van Velden, J. L., Koeslag, A., Curtis, O., Gous, T. A., and Amar, A. (2017). Negative Effect of Knemidokoptes Mite Infection on Reproductive Output in an African Raptor. The Auk. doi: 10.1642/auk-16-134.1
Venables, W. N., and Ripley, B. D. (2002). Modern Applied Statistics with R. 4th Edn. New York, NY: Springer. doi: 10.1007/978-0-387-21706-2
Visser, M. E., Holleman, L. J. M., and Caro, S. P. (2009). Temperature has a causal effect on avian timing of reproduction. Proc. R. Soc. B 276, 2323–2331. doi: 10.1098/rspb.2009.0213
Wickham, H. (2009). ggplot2: Elegant Graphics for Data Analysis. New York, NY: SpringerVerlag. doi: 10.1007/978-0-387-98141-3
Wawrzyniak, J., Kaliński, A., Glądalski, M., Bańbura, M., Markowski, M., Skwarska, J., et al. (2015). Long-term variation in laying date and clutch size of the great tit Parus major in central Poland: a comparison between urban parkland and deciduous forest. Ardeola 62, 311–322. doi: 10.13157/arla.62.2.2015.311
Keywords: urban ecology, timing of breeding, reproduction, South Africa, Accipiter melanoleucus, raptors
Citation: Rose S, Sumasgutner P, Koeslag A and Amar A (2017) Does Seasonal Decline in Breeding Performance Differ for an African Raptor across an Urbanization Gradient?. Front. Ecol. Evol. 5:47. doi: 10.3389/fevo.2017.00047
Received: 31 January 2017; Accepted: 28 April 2017;
Published: 16 May 2017.
Edited by:
Diego Gil, Consejo Superior de Investigaciones Científicas, SpainReviewed by:
Davide M. Dominoni, Netherlands Institute of Ecology (NIOO-KNAW), NetherlandsMeritxell Genovart, Mediterranean Institute for Advanced Studies (CSIC), Spain
Copyright © 2017 Rose, Sumasgutner, Koeslag and Amar. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Petra Sumasgutner, cGV0cmEuc3VtYXNndXRuZXJAdW5pdmllLmFjLmF0
†These authors have contributed equally to this work.