Abstract
High Neotropical mountains are among the most threatened ecosystems by climate change and this problem could be accentuated in cities where temperatures are higher. However, there are few data of long-term avifaunal changes in Neotropical cities, and the potential impact of climate change has yet to be addressed. Using data from 26 years of Audubon's Christmas Bird Counts (CBCs) in the Bogotá area (ca. 2,600–3,100 m in Colombia's eastern Andes), we analyze long-term changes of the avifauna using linear regressions corrected for species' habitat preferences and compared urban and rural sites. For the CBC we followed the National Audubon Society's methodology in which birds are counted over a 24-h period within a fixed circle 24 km in diameter. We recorded 235 species in the Bogotá circle with an average of 122 spp/year, including 46 boreal migrants, three endemic species, and four endemic subspecies, two globally and four locally threatened species. Species richness was higher in rural than in urban areas and most species were associated with native forest and scrub and wetlands. Among the species that were analyzed for changes in time 31% increased, 20% decreased, and 49% did not change. Strong fluctuations or changes in abundance were more frequent in urban than rural environments. Many of the species that increased or became established during the CBC interval came from lower elevations or the warmer, drier parts of the region beyond the count circle. By contrast, the lower elevational limits moved to higher elevations in several species that decreased. Climate change with its related effects represented the factor associated with the most changes in abundance. Other potential causes were direct human actions, mostly associated with urbanization, and “natural” successional changes in vegetation; predation by feral dogs and cowbird parasitism also affected several species negatively. Observations indicate that these factors will continue into the future, and the effects of climate change and urbanization in particular will probably intensify. The Bogotá CBC has provided much valuable information for Neotropical urban ornithology and its practice should be encouraged in other Latin American cities and should definitely continue in Bogotá for many years to come.
Introduction
Climate change is presently a growing threat to biodiversity (Root et al., 2003). It is affecting a variety of organisms by shifting elevational distributions (Chen et al., 2011), causing decoupling of phenological relationships (Visser and Both, 2005), changing times of reproductive events (Both et al., 2004), and changing composition and structure of ecosystems (Grimm et al., 2013), among others. High mountain neotropical ecosystems such as those of the Andes are among the most threatened, because of the progressively smaller areas available as elevation increases (Sekercioglu et al., 2008; Chen et al., 2011; Chaverri et al., 2016). Organisms that shift their altitudinal limits upwards, as predicted by global warming, will therefore suffer reductions in their populations and eventually may be driven to extinction at the highest elevations (Larsen et al., 2011; Feeley et al., 2012); tropical mountain systems are particularly vulnerable to climate change because organisms are adapted to more stable climates than temperate organisms at similar elevations and many populations may not have the ability to withstand temperatures not previously experienced (Janzen, 1967; Ghalambor et al., 2006). Moreover, such species often occupy narrow elevational ranges (Anderson et al., 2011). Birds represent an ideal study group for monitoring elevational shifts because they are more easily identifiable in the field than other vertebrate groups and often exhibit strong habitat preferences, especially in montane areas. There are few long-term studies along elevation transects that repeat surveys made decades earlier to evaluate altitudinal movements. One by Forero-Medina et al. (2011) in Perú and another by Freeman and Class-Freeman (2014) in New Guinea detected upward shifts in dozens of bird species. Another approach is to monitor species composition of birds over several decades at a single locality. Pounds et al. (1999) compared data over nearly two decades for a mid-elevation site in Costa Rica, and found that the bird community changed due to an increase of species moving from lower elevations.
There are still relatively few studies of urban birds in Latin America (Ortega-Álvarez and MacGregor-Fors, 2011; Delgado-V and Correa-H, 2013), and these focus mainly on urban gradients, ecology and community composition but few have been developed over a long-term and none have focused on the effects of climate change on birds. Climate change in cities is of particular interest due to their increased tendency to warming, forming “heat islands” due to high carbon dioxide emissions and aerial contamination (Foley et al., 2005; Wilby and Perry, 2006). The city of Bogotá has a population of nearly eight million and is located on an Andean plateau (the Sabana de Bogotá (Montañez et al., 1994) at an elevation of 2,600 m with adjacent mountain ridges reaching over 3,000 m. The city contains a heat island over most of its extension with a temperature 3°C above the perimeter and surrounding areas (Angel et al., 2010). This combination of features makes Bogotá an ideal site for studying the combined effects of climate change and urbanization on birds at high elevations because it is an expanding city at a high altitude, and because few—if any—Andean sites with these characteristics have long-term observations similar to those we report for Bogotá.
Long-term studies of bird communities and populations have a long history in Europe and North America and have contributed greatly to knowledge in science and ecology (e.g., Holmes, 2011; Perrins and Gosler, 2011), but have mostly concentrated on particular forest sites. Another type of long-term monitoring is the annual North American Audubon's Christmas Bird Counts (CBCs), involving the participation of professionals and amateur observers that count birds during 1 day in 24 km diameter circles chosen by compilers. The CBCs in North America now involve thousands of bird watchers and more than 1,286 count circles monitored over periods of up to a century or more (National Audubon Society, 2017; http://www.audubon.org/conservation/science/christmas-bird-count) and have provided fundamental information on changes in distribution and abundance of birds (e.g., Johnsgard, 1967, 2015; Bonney, 1979; Monahan and Koenig, 2006; Soykan et al., 2016). However, such long-term bird monitoring is in its early stages in Latin America due to the lack of a tradition of popular bird watching, largely reflecting the lack of well-illustrated bird guides until the 1980s. In Colombia the authors started a CBC circle in the Sabana de Bogotá in 1986 which became established in 1989 with the foundation of the Bogotá Ornithological Association (ABO). This CBC has continued uninterrupted for more than 25 years, becoming the longest monitoring of any vertebrate group in the country. Our objective here is to use the data of the CBC from 1989 to 2014 to evaluate the changes in the bird community of the city of Bogotá and its periphery and their potential causes, including their possible association with climate change.
Prior to the initiation of this CBC, there was no systematic program of bird observation in Bogotá and no reliable, updated list of the birds of the Sabana. However, the important work by Olivares (1969) permits an earlier summary of the birds of this area, updated through the 1960's. Despite being based nearly exclusively on specimen records (sometimes with vague localities like “around Bogotá”), this work provides a basis for evaluating changes in the two decades prior to the CBC; especially useful in this regard are Olivares's comments on the local abundances of a number of species. Also providing data that help in interpreting the changes in the avifauna of the CBC is a concurrent study by the authors (Rosselli et al., in press) of an area of high Andean forest at 2,800–3,300 m ca. 25 km east of the CBC circle and involving two inventories of its birds in 1991–1992 and 2015–2016.
Methods
Study area
The area of the count circle includes the northern part of the city of Bogotá and adjacent rural areas, as well as several mountain ridges reaching elevations between 2,800 and 3,100 m. Its center at 4°48′N, 73°11′W was chosen to include a variety of habitats in an area that we predicted would be affected by the expansion of urbanization in future decades (Figure 1).
Figure 1
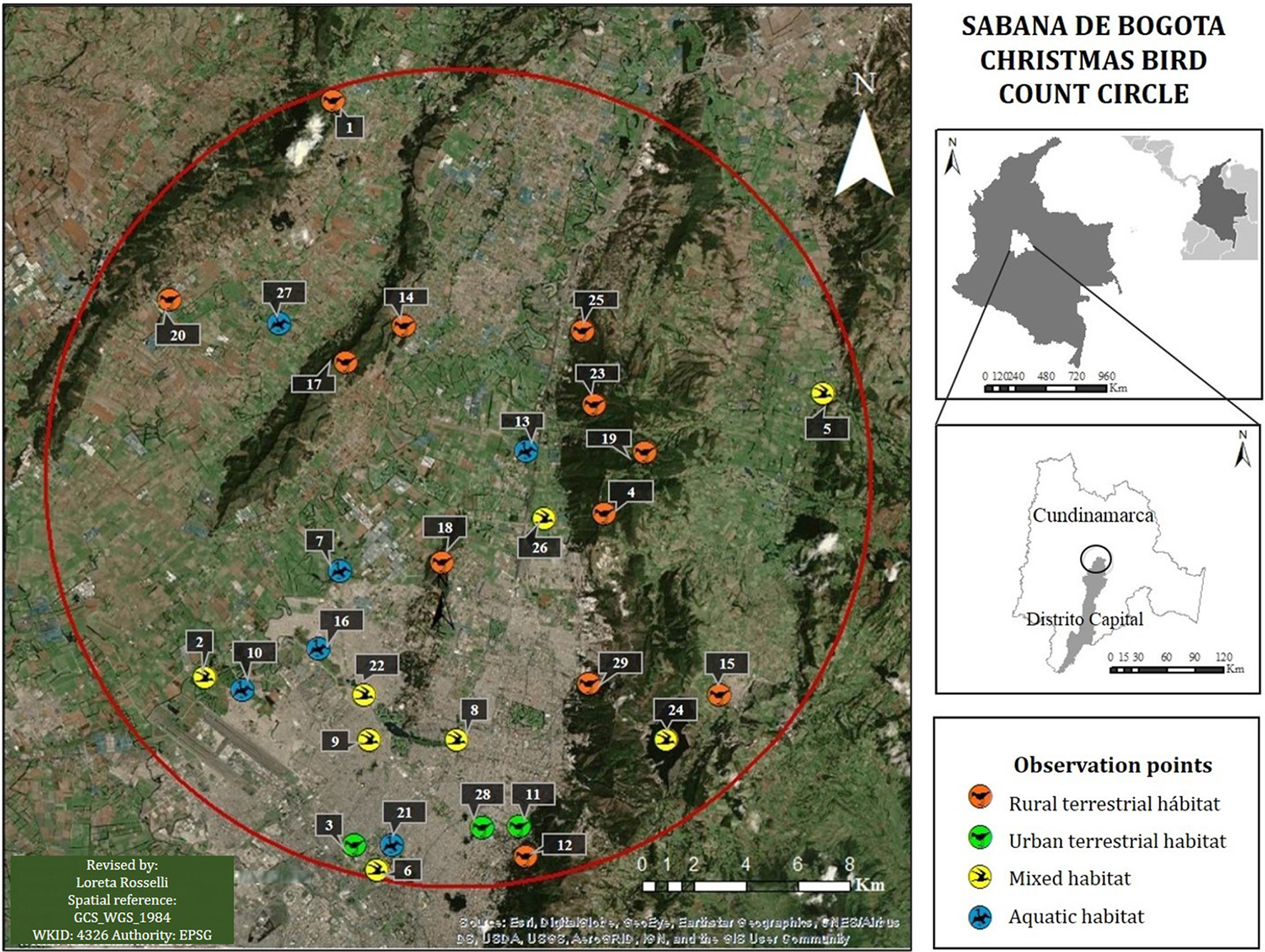
Study area. Location of the Sabana de Bogotá Audubon's Christmas Bird Count circle and the 29 locations visited between 1989 and 2014. Habitat types were assigned according to dominant land covers in each place (more details about habitats in Section Audubon's Christmas Bird Count Methods and Circle).
Mean annual temperature in the city ranges from 12 to 16°C and mean annual rainfall shows a gradient from 800 mm in the south to more than 1,000 mm on the northeastern border (Angel et al., 2010). The rainfall pattern of the count area is bimodal, with the CBCs occurring in the transition between the October–November rainy season and the January–March dry season. There has been an increase of 0.2–0.4°C per decade in the mean maximum temperature in the region (Rojas et al., 2010) and a particularly high increase in recent decades in the city itself due to the formation of a heat island that is expanding toward the northwest (Angel et al., 2010), and now includes 9 of the 11 CBC localities within the urban matrix of Bogotá.
The flat Sabana was the bottom of a large lake that was naturally drained and the original habitats of the now dry Sabana were probably native grasslands, wetlands that varied according to rainfall and patches of forest along streams and around wetlands. Most of the city of Bogotá lies on the flat part of the Sabana, which beyond the urban areas includes mostly pastures and croplands. The Sabana is bordered eastward by a branch of the Colombia's eastern Andes reaching elevations of up to 3,200 m, parts of which are included in the count circle, as are several small chains of mountains with elevations between 2,800 and 3,100 m that run in a north-south direction within the Sabana. The slopes of the mountains support native forests and scrub as well as portions of exotic forest plantations. The Teusacá River drains into the Sabana through a gap in the eastern mountains; part of the flat lower valley includes pastures, croplands and small artificial wetlands, which are also included within the count circle.
The transformation of the natural habitats of the Sabana began over 10,000 years ago with the arrival of indigenous peoples who cultivated areas near the wetlands and rivers. Transformation of terrestrial habitats to urban areas, pastures and croplands became more intense during the colonial period. The draining of the wetlands increased especially following the independence from Spain in the nineteenth century and reached a peak in the latter half of the twentieth century (van der Hammen et al., 2008), by the end of which only 3% of the original wetlands remained (Andrade, 1998). The growth of the city of Bogotá also produced the repeated deforestation of most of the adjacent mountains for construction and firewood. Coal and more recently, oil and gas replaced firewood for cooking in the latter part of the twentieth century leading to succession of native forest and growth of exotic tree plantations (mostly Eucalyptus globulus) on most of the mountain slopes. The accelerated displacement of rural pastures and croplands by the city's growth over the flat areas of the Sabana continues through the present day. The introduction of the exotic kikuyu grass in the early twentieth century to improve pastures (Pérez-Arbeláez, 1935) resulted in the eradication of native grasses and herbs over most of the Sabana. Between 1995 and 2015 Bogota's population increased by 38% to nearly 7,878,783 (Secretaría Distrital de Planeación, 2016), and its urban area by 33.8% to ca. 38,430 ha between 1993 and 2010, with this growth concentrated on the northern, western and southern edges of the city (Romero, 2010) including some of the CBC points. The reduced remaining urbanizable area in Bogotá has also pushed urbanization to the neighboring municipalities west and north of the city (Martínez-Herrera, 2015) where some of the CBC observation points are located. To evaluate the local changes in our observation points we estimated the changes of 1 km radius circles around the center of each locality using available high resolution historic images from Google Earth (mostly from 1999 to 2005 to the present). We also made comparisons of the amount of urbanized and rural areas in the complete CBC circle comparing such images dating from 1980, 2005, and 2016.
The status of the wetlands is of special concern, due to the number of endemic and threatened bird species they support. Although, in 2000 the city's authorities declared 12 wetlands as ecological parks for conservation, the pressure from urbanization continues, including increased pollution, encroachment of the borders and further reduction of wetland areas, and increasing presence of domestic dogs (Rosselli and Stiles, 2012a).
Audubon's Christmas bird count methods and circle
The methodology of the CBC was standardized in North America in the 1980s (Arbib, 1981, 1982). For a period of 24 h between approximately December 14 and January 5, parties (groups) of bird watchers identify and count all birds they can detect in a 15 mi diameter circle (24.1 km) with a fixed center. Each party counts birds in one or more specific sites representing some of the habitats in the circle. The distance traveled is recorded (by foot, car, etc.), as are the starting and ending times and the weather conditions. Party coordinators need to have the knowledge and ability to ensure that the identifications of the reported species are correct. Every group's data is revised and compiled by the circle's coordinator. To correct for differences in sampling effort all data for the circle are divided by a global measurement such as party-hours or distance traveled. The method is designed to ensure as far as possible that the data gathered by different groups in different years are comparable and enable assessment of long-term changes of species and populations in the circles. Bock and Root (1981), Butcher and McCulloch (1990), Dunn et al. (2005) and others have addressed the standardization and comparability of data since observers in the circle may change from year to year. Some of these difficulties include identification capabilities of individuals, number of bird watchers in the groups, weather and times of observation, travel distances, hours of observation, extent coverage of different sites and habitats, and use of bird song recordings to attract the birds.
The CBCs have proven reliable to register presence-absence of species and abundance tendencies and have shown correlations with various other surveys in North America (Dunn et al., 2005; Soykan et al., 2016). Over the 26 years of the Sabana de Bogotá counts, group coordinators were familiar with their areas and birds, and as far as possible coordinated in the same locations year after year to reduce variations due to human factors. Since 2000 we designated 13 representative sites with at least 8 years of data as high priority to be counted every year to reduce variability of coverage and to be able to compare the sites year after year. The center and diameter of the Bogotá circle have remained constant over the 26 years, thus ensuring comparability between years.
The CBC circle
The circle includes a variety of habitats including urban parks, rural areas with crops and pastures, natural and artificial wetlands of different sizes and surroundings, natural and planted forests (mostly Eucalyptus) and secondary scrub. We did not intend to count birds in completely urban areas without vegetation due to the limited presence of birds in them; neither did we include the only naturalized species in the area: the rock dove (Columba livia); its status as a wild bird is uncertain outside of urban areas because many farmers keep a pigeons that fly freely during the day but sleep and are mostly fed in dovecotes.
We used a simple system to classify the four principal kinds of vegetation cover found in the circle:
NFS- native forests and secondary scrub (locally called matorral; Figure 2a): because such scrub represents natural succession to forest and there is a gradation in the ages and heights of the shrubbery, it was better to combine them into a single category. We include exotic forests in this category because their avifauna is basically a subset of those adjacent or intermixed native forests.
Figure 2

Main habitats of Audubon's Christmas Bird Counts in the Sabana de Bogotá from 1989 to 2015. (a) Native forests and secondary scrub (NFS). (b) Open rural areas (ORA). (c) Wetlands (WET). (d) Urban forested parks (UPT).
ORA- open rural areas: pastures, crops, some scrub and living fences of different types that divide or define a property, farm buildings, and constructions (Figure 2b).
WET: Wetlands of various types, from marshes and natural lagoons to artificial ponds, the Bogotá and Teusacá rivers, and seasonal puddles (Figure 2c). Species considered as wetland birds include those that depend on the water of the wetland and not vegetation from surrounding areas with different types of terrestrial vegetation.
UPT: Urban parks with trees, with an arboreal stratum of exotic or native trees and underneath areas of lawns and gardens, which sometimes may include patches of dense vegetation such as ornamental shrubs or natural vegetation at the edges (Figure 2d). Such parks may be within the urban matrix or adjacent to some wetlands within the city. It is common for parks to attract birds from adjacent urban areas, as well as a greater variety of species that depend on the greater complexity of their vegetation.
TER: A general term for terrestrial vegetation, including NFS, ORA, and UPT, when we make comparisons with wetland birds and vegetation (WET).
URB, RUR: general terms for urban (within Bogotá's urban perimeter) or rural (outside the city) for other comparisons (see Figure 1).
Counts were made in a total of 29 sites in the circle during the 26 years analyzed (Table 1). Some localities were changed in subsequent years in favor of others more representative or accessible. Sometimes permission was no longer granted to visit a site and we tried to replace it with a similar one to maintain continuity of the habitats censused. Beginning in 2001, 13 localities, identified as prioritary, were visited every year. Additional localities were added to these 13 as coordinators and participants were available.
Table 1
| A. Numbers of years a site was censused: in parentheses, the number of sites designated as priority (*) and the number of sites requiring substitution due to problems of access (†). | |||||
|---|---|---|---|---|---|
| No. years | 1–5 | 6–10 | 11–15 | 16–20 | 21–26 |
| No. sites | 7 | 9 (2*, 2†) | 3 (1*,1†) | 4 (4) | 6 (6) |
| B. Numbers of sites in which the four principal habitats were represented by 20% or more of the area in a given site. | ||||
|---|---|---|---|---|
| Numbers of habitats | 1 | 2 | 3 | 4 |
| Numbers of sites | 10 | 18 | 1 | 0 |
| C. Numbers of sites with the four principal habitats represented (see below for abbreviations of habitats). | ||||
|---|---|---|---|---|
| Habitats | NFS | ORA | UPT | WET |
| As principal habitat: 40% or more of site area (in parentheses, two equally represented habitats) | 9(3) | 2 | 6(2) | 10(2) |
| Habitats represented by 20% or more of area of site | 12 | 14 | 10 | 13 |
| D. Numbers of sites with different combinations of habitats represented by 20% or more of the site areas. | ||||||
|---|---|---|---|---|---|---|
| Combinations of habitats | NFS-ORA | WET-UPT | WET-ORA | NFS-WET | ORA-UPT | WET-UPT-ORA |
| Numbers of sites | 8 | 4 | 4 | 1 | 1 | 1 |
Characteristics of the 29 sites censused during the 26 years of the Sabana de Bogotá Audubon's Christmas Bird Count (1989–2014).
NFS, native forests and scrub; ORA, open rural areas; WET, Wetlands; UPT, Urban forested parks.
Analysis
To analyze the structure and composition of the birds registered in the 26 years of the CBC, we first defined the status of all recorded species (Table 2) as residents or visitors; for visitors, we determined their most probable origin in terms of elevation and region. This classification was based on the count data and on our experience of decades in the Sabana and in other parts of the eastern Andes region. We then identified four important parameters for each species: (a) regularity, in terms of number of years in which it was recorded; (b) its distribution in terms of the number of sites in which it was recorded; (c) its abundance calculated as the average of individuals counted in the years it was registered; and (d) changes in abundance and status of the species over the 26 years, here correcting for habitat preferences (see below). To simplify analysis, we divided these numerical parameters into a smaller number of classes (Table 2). Initially, we compiled all general data from the counts: number of sites, party-hours, kilometers/party, number of species, and individuals for each species and site. We obtained the species accumulation curve and analyzed the frequency distributions of species according to their regularity, distribution, and abundance using the numbers of species in each status class of these parameters.
Table 2
| A. SITUATION IN THE CIRCLE | |
| PR | Permanent Resident. Present all year long, reproduces in the circle or nearby áreas. Rare or difficult to detect species are classified as residents if there is no evidence of migration or regular movements away from the circle for part of the year. Includes species that were residents at the beginning of the study but eventually disappeared from the counts and others that became established as residents and reproduced in the years included in the analysis. |
| SR | Seasonal Resident. Present for several months around the date of the CBC although not necessarily all years; almost all these species are boreal migrants that spend the northern winter in the circle. At this time of year austral migrants are almost never found. |
| MV | Mountain Visitors. Residents from other areas in Colombia's Eastern Andes or “Cerros Orientales” but not within the circle where they appear in low numbers and not regularly in some years. |
| HV | Highland Visitors. Visitors from higher elevations that appear sporadically in the circle, mostly in low numbers and for which there is no evidence of reproduction in the circle. |
| LV | Lowland Visitors. Visitors from lower elevations that appeared in a few years but also in low numbers without evidence of settling. |
| RV | Reproductive Visitors. Visitors in some years with evidence of reproduction attempts but that eventually disappeared without settling. |
| TV | Transient Visitors. Visitors that pass through the circle, generally flying, in the course of migration or regional movements. Includes a record of a late austral migrant and a large flock of another species that disappeared after the count. |
| IV | Introduced Visitor. Resident in distant areas of the country that may have been set free or has escaped from captivity. |
| B. REGULARITY IN THE CIRCLE* | |
| 7 | Recorded in 24–26 years |
| 6 | Recorded in 19–23 years |
| 5 | Recorded in 13–18 years |
| 4 | Recorded in 8–12 years |
| 3 | Recorded in 4–7 years |
| 2 | Recorded in 2–3 years |
| 1 | Recorded only in 1 year |
| C. DISTRIBUTION IN THE CIRCLE* | |
| 6 | Recorded in 19–26 localities |
| 5 | Recorded in 11–18 localities |
| 4 | Recorded in 7–10 localities |
| 3 | Recorded in 4–6 localities |
| 2 | Recorded in 2–3 localities |
| 1 | Recorded in only one locality |
| D. ABUNDANCE IN THE CIRCLE** | |
| 8 | Average ≥250.00 |
| 7 | Average 100.00–249.99 |
| 6 | Average 50.00–99.99 |
| 5 | Average 25.00–49.99 |
| 4 | Average 12.50–24.99 |
| 3 | Average 5.50–12.49 |
| 2 | Average 2.50–5.49 |
| 1 | Average 1.00–2.49 |
| E. ABUNDANCE TENDENCIES IN THE CIRCLE*** | |
| Significant increase | Regression coefficient (slope) positive, p ≤ 0.05 |
| Slight increase | Regression coefficient (slope) positive 0.10 ≤ p ≥ 0.05 |
| No change | Regression coefficient positive or negative but p > 0.10 |
| Slight decrease | Regression coefficient (slope) negative, 0.10 ≤ p ≥ 0.05 |
| Significant decrease | Regression coefficient (slope) negative, p ≤ 0.05 |
| X | Insufficient data to be able to determine tendency |
Classification of species registered in the Audubon's Christmas Bird Counts in the Sabana de Bogotá circle, 1989–2014.
In categories B and C, we arbitrarily established classes according to the distribution of numbers of species trying to leave similar numbers of species in each class to make comparison between species easier in the analysis.
Since the averages of numbers recorded for each species ranged over three orders of magnitude and most species have small abundances, for category D we established classes in an approximately exponential scale, with similar numbers in the majority of classes. We did not include the years in which the species were not recorded in the averages
To evaluate the tendencies in Category E, we used linear regressions, especially for species recorded during 10 or more years and some with less (but always above five) that became established or went extinct during the period of the CBCs. Classes were defined according to the p value of the regression. Each number was divided by the party-hours spent in the species preferred habitat to correct for differences in sampling intensity between years.
We standardized sampling effort by dividing the abundances of every species by party-hours in the preferred habitat of each one to take into account habitat preferences and yield a more precise assessment of long-term population changes; this is an adaptation of the most commonly used effort adjustment (Raynor, 1975). To begin with, we arbitrarily calculated linear regressions of the corrected numbers/count for all species recorded in 10 or more years but examining the data we found that clear tendencies were evident for some species with 6–9 records that were also analyzed. We excluded from the analysis all species with 5 or fewer records because we considered them insufficient to permit reliable analysis. We also analyzed data for a few species with 6–9 years (data points) where these appeared to show a clear tendency and did not have many years without data between the data points to evaluate tendencies in populations. For several species we calculated regressions with and without outliers (atypical high or low counts in a year, Gotelli and Ellison, 2004) because these can have a disproportionate influence on the regression results, especially when occurring at the beginning or end of the interval of time considered. We considered that a species had become established in the circle if it was present for at least 10 consecutive or nearly consecutive (with few isolated missing years in the series) years, preceded by 5 or more years without records; extinction of a species in the circle was inferred if no record of its presence was obtained through the end of the study period following at least 5 continuous years in which it was present. We accepted as valid changes in the numbers of a species when the p-value for the regression was ≤0.05; if p was between 0.05 and 0.10, we considered changes as slight increases or decreases and we considered regressions with p > 0.10 as indicators of stable populations. We compared the number of the species in rural vs. urban areas, and terrestrial localities (TER) vs. wetland localities (WET) using Chi-square goodness of fit tests.
We used a non-parametrical Mann–Kendall Trend Test to evaluate possible temperature increases with the Sen method for slope calculation. This method is appropriate for time series with missing values and outliers and is oriented to detect and calculate tendencies in time series of climate data (Salmi et al., 2002; He and Zhang, 2005).
Results
Land cover and temperature changes
There were relatively few pronounced changes in the CBC observation points: more than half of the sites showed no change or <5% increase in urbanization (Figure 3). These sites with little or no change were mainly (75%) rural and native forest and secondary scrub in which a few houses were added. Three were parks embedded within the city. Management efforts by the city's authorities increased open water habitat in some of the wetlands (Figure 3) whilst in a rural locality a 3.5 ha pond was destroyed. The highest percentages in urbanization (10–25%) occurred on the borders of the city and in one of the sites in a neighboring municipality. In some of the urban parks that were established at the beginning of the CBC counts, the trees grew and densified in 5–12% of the area. Therefore, changes at the local level in the observation points were not large but reflect general changes at a broader scale. The urban and conservation areas that were present in late eighties have changed little; the main changes have occurred along the borders of the city in and around the CBC circle and mostly consist of replacement of pastures with housing and commercial developments as well as floriculture greenhouses. In the circle as a whole, we found that the proportion of rural areas, especially pastures, had been decreased by 30–40% by 2005, and by 60–75% by 2016; two neighboring towns have also reduced pastures and crop fields by a similar amount in the last 10–15 years (Supplementary Image 1).
Figure 3
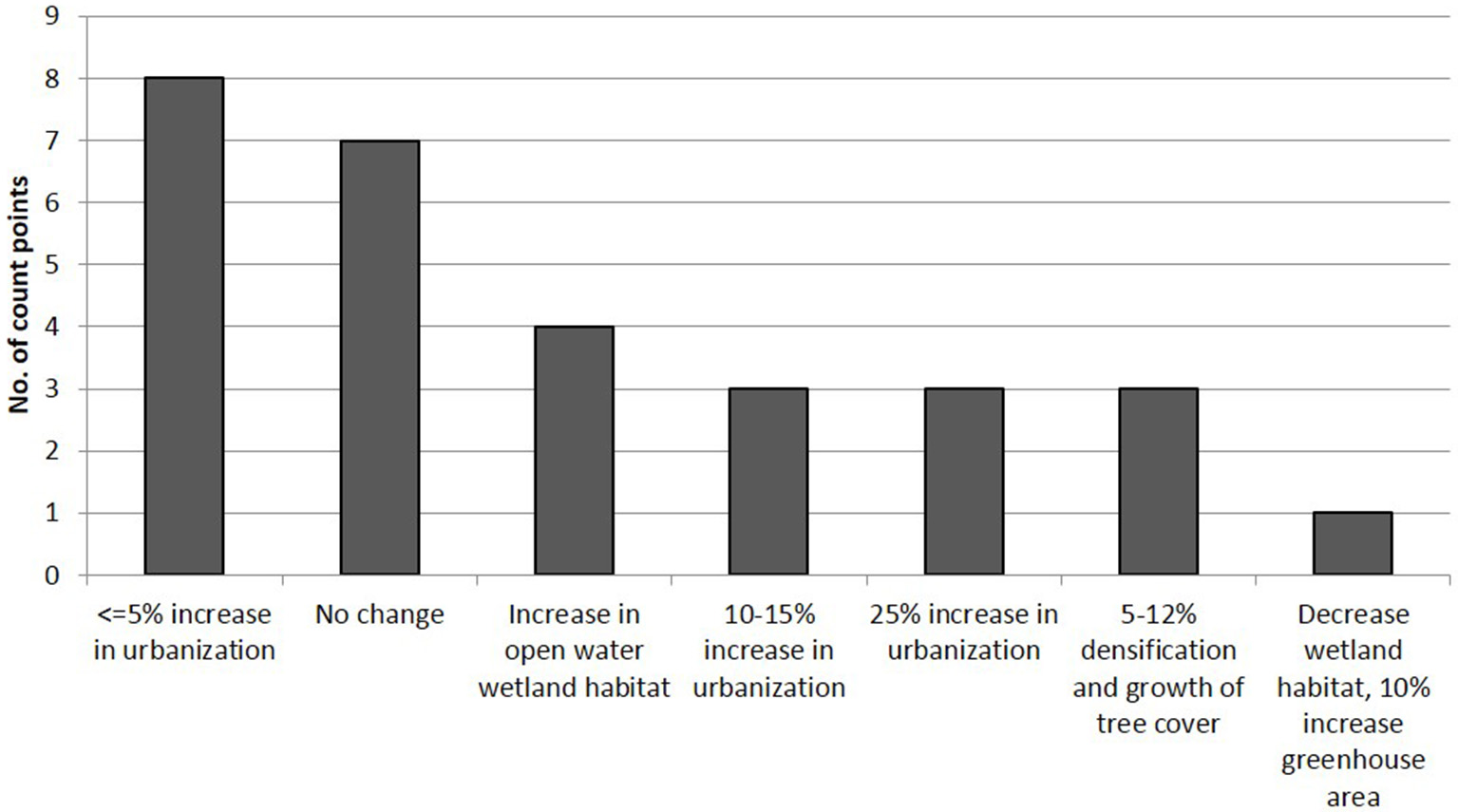
Land cover changes in the last ~16 years in 1 km radius circles around the 29 count points in the Sabana de Bogotá Audubon's Christmas Bird Count circle.
There was a significant trend of temperature increase at the Eldorado airport between 1962 and 2015 in mean maximum, minimum and mean annual temperatures (Figure 4). The strongest tendency occurred in the minimum annual mean (Kendall's Tau = 0.62, p < 0.0001) with an increase of 0.043°C/year (Table 3). The increase rate in temperature was higher and significative for the minimum and mean annual means during the CBC years, but not for the maximum mean (Table 3).
Figure 4
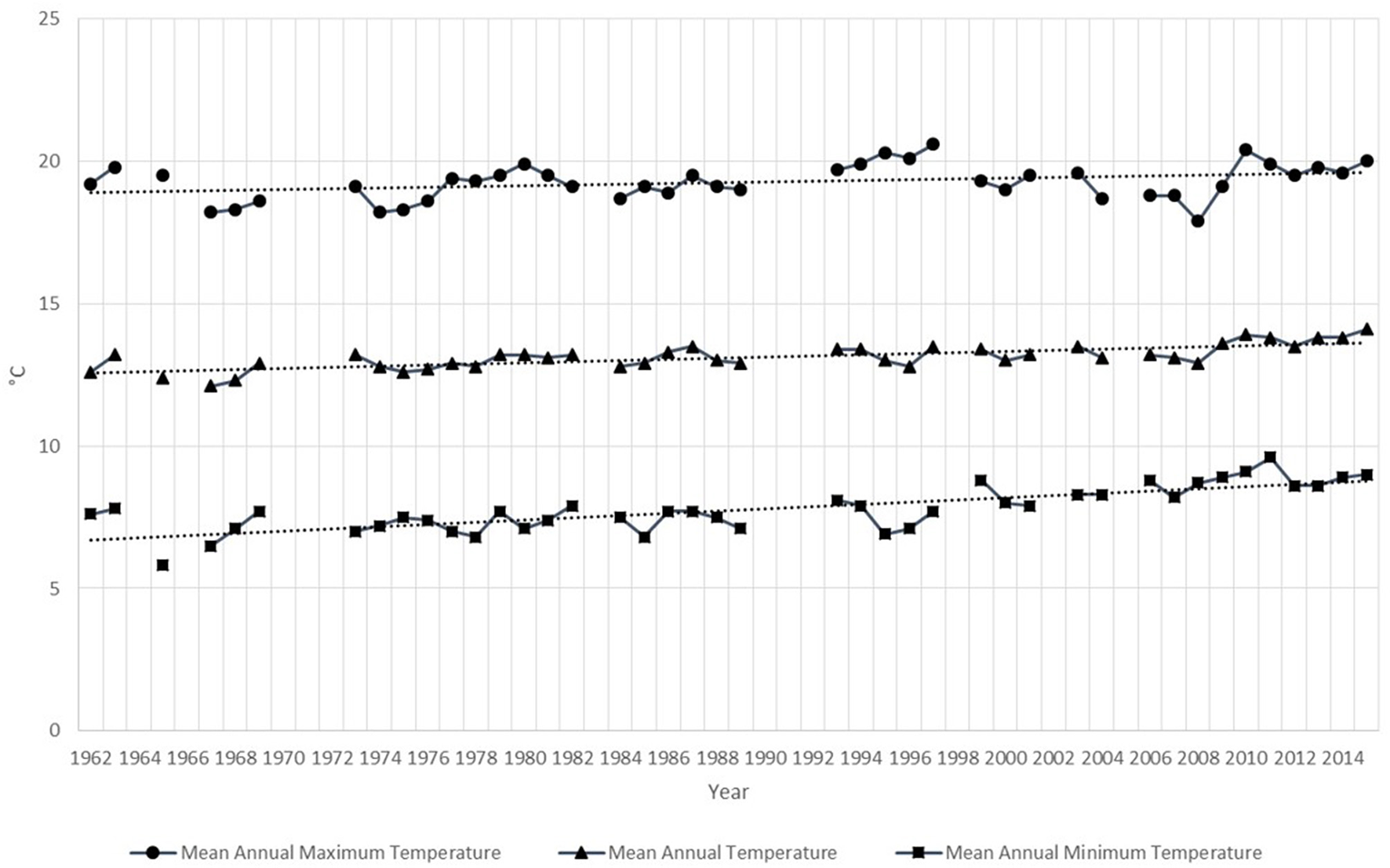
Mean annual maximum, mean and minimum temperatures, and tendency lines between 1962 and 2015. Data from Eldorado airport located in the CBC circle.
Table 3
| 1962–2015 | 1989–2014 | |||||
|---|---|---|---|---|---|---|
| Minimum mean annual temperature | Mean annual temperature | Maximum mean annual temperature | Minimum mean annual temperature | Mean annual temperature | Maximum mean annual temperature | |
| Kendall's Tau | 0.622 | 0.547 | 0.266 | 0.582 | 0.370 | −0.148 |
| p | <0.0001 | <0.0001 | 0.017 | 0.001 | 0.034 | 0.400 |
| Temperature (°C) increase rate/year | 0.043 | 0.02 | 0.017 | 0.075 | 0.025 | |
Results of the non-parametrical Mann–Kendall trend test for temperature increase in Bogotá's airport between 1962 and 2015 and during the 26 years of Audubon's Christmas Bird Counts (1989–2014).
General ornithological results
In the 26 years of counts, we recorded 235 species of birds in the Sabana de Bogotá Audubon's Christmas Bird Count circle (Table S1). The accumulated number of species increased notably in the first 16 years of counts (1989–2003) from 103 to 215 species and stabilized thereafter but always increasing, reaching the maximum in 2014 (Figure 5). The average number of species/count was 122.1 ± 12.4 s.d. increasing from 103 in 1989 to a maximum (142) in 2001 and remaining more or less constant in the following years (Figure 5).
Figure 5

Species accumulation curve and number of species/year registered at the Sabana de Bogotá Audubon's Christmas Bird Count circle between 1989 and 2014.
Families with most species recorded in the counts were Thraupidae (33), Tyrannidae (28), Trochilidae (18), and Parulidae (15). Fifteen of the families are represented by only one species (Table S1). Permanent residents comprised the largest category, with 187 species. Forty-six of the species were wintering boreal migrants, mainly Parulidae and Tyrannidae (Table S1). In terms of geographical distribution, three species and four subspecies are endemic to the northern part of Colombia's eastern Andes (Rallus semiplumbeus, Synallaxis subpudica, Cistothorus apolinari, Ixobrychus exilis bogotensis, Porphyriops melanops bogotensis, Eremophila alpestris peregrina, and Chrysomus icterocephalus bogotensis; Table S1). Seven species are Range Restricted according to BirdLife International's criteria (global breeding range below 50,000 km2; Stattersfield et al., 1998). R. semiplumbeus and C. apolinari are globally threatened (IUCN, 2017) and four other species are locally threatened in Colombia (Oxyura jamaicensis, Porphyriops melanops, Pseudocolopteryx acutipennis, and Eremophila alpestris; Renjifo et al., 2014, 2016; Table S1).
Most species (84) were associated with native forest and scrub, or wetlands (56) whereas there were only 15 species exclusive to open rural areas (Figure 6). Nearly all of the species (91%) that were restricted to urban parks with trees (UPT) correspond either to visitors from lower elevations (57%) or wintering boreal migrants (34%). Some of the species from lower elevations could also have been cage birds escaped from captivity (e.g., Pteroglossus castanotis, Cyanocorax affinis, Icterus icterus) while others had more likely shifted from lower surrounding areas (e.g., established species such as Elaenia flavogaster, Pitangus sulphuratus, and Thraupis palmarum, or visitors like Todirostrum cinereum, Columbina talpacoti, and Saltator striatipectus; Table S1).
Figure 6
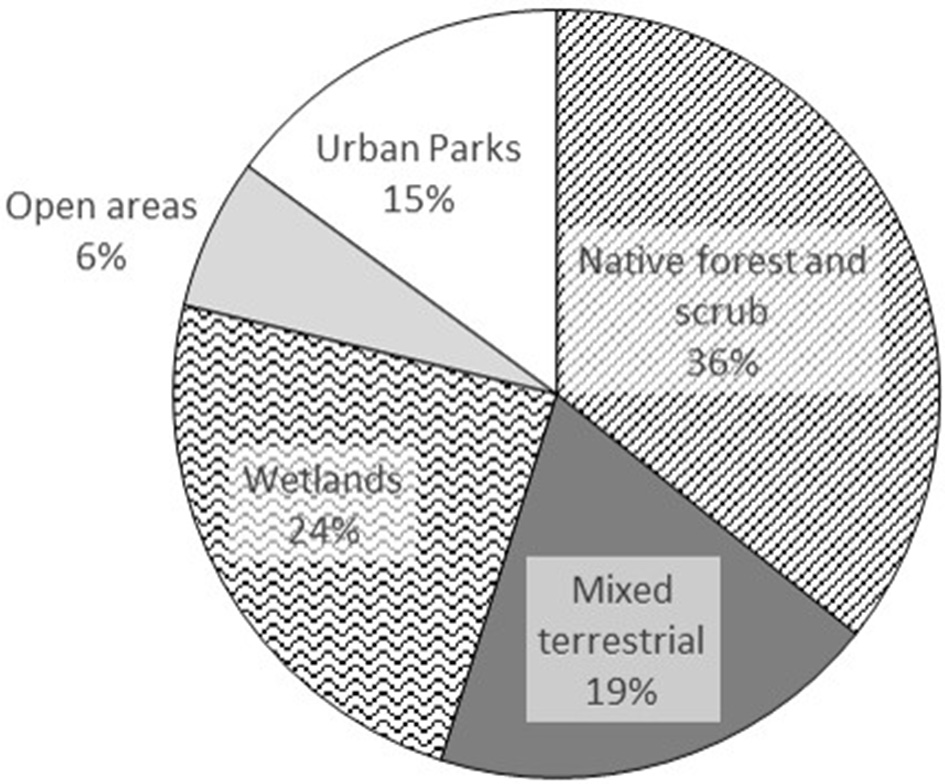
Percentages of habitats preferred by the 235 bird species recorded in the Sabana de Bogotá Audubon's Christmas Bird Count circle between 1989 and 2014.
Urban vs. rural areas
The total number of species registered was significantly higher in rural than in urban areas (209 vs. 167 species, χ2 = 4.691, p = 0.03). Sixty-eight species were exclusive to rural areas, 26 were exclusive to urban areas and 141 were registered in both urban- and rural-areas (Sorensen similarity Index = 0.7446, Table S2). The number of species in terrestrial localities (TER) was higher for rural areas than for urban areas (χ2 = 5.442, p = 0.02), and there was no significant difference in the total number of aquatic species between rural and urban areas for wetland localities (WET; Table S2).
Forty-two species that were registered both in rural and urban areas had a significant difference in corrected abundance. Eighteen species were more abundant in the urban areas, among them Gallinula galeata, Phimosus infuscatus, Vanellus chilensis, Zenaida auriculata, Colibri coruscans, Molothrus bonariensis, Tyrannus melancholicus, Thraupis episcopus and the boreal migrants Tringa melanoleuca, T. solitaria, T. flavipes, Leiothlypis peregrina, Piranga rubra, and Parkesia noveboracensis. Among the species that were more abundant in the rural areas were O. jamaicensis, Ardea alba, Bubulcus ibis, and Gallinula melanops and NFS species like Penelope montagnii, Patagioenas fasciata, Grallaria ruficapilla, Scytalopus griseicollis, S. subpudica, Elaenia frantzii, Mecocerculus leucophrys, and 4 species of hummingbirds. The only boreal migrant more abundant in rural areas was Setophaga fusca (Table S2).
Twenty-six species were registered only in urban areas, among them residents or visitors that came from the warmer lowlands, like P. sulphuratus, C. affinis, T. palmarum, Cacicus cela, and Gymnomystax mexicanus. Most of the 68 species that were registered only in rural areas were found only in natural habitats (NFS) (Table S2).
Long-term changes in birds in the bogotá area
Sixty-six percent (155) of the species were analyzed for changes over the 26 year interval; of these 31% increased, 20% decreased, and 50 did not change significantly (Table 4). Thirty-six permanent residents increased their abundance and 22 decreased at least slightly (p ≤ 0.10). Two species that did not reach the minimum number of records for the regression analysis (I. exilis and E. alpestris) should be added to the latter since they did change but disappeared from the counts in the initial years; this would add a total of 60 (47%) permanent residents that showed changes along the study period; 53 species did not change their abundance (42%) and the remaining 14 not analyzed or without evident tendencies added only 11% (Table S3). Among species with significant increases (p ≤ 0.05), six arrived in the circle area, started to reproduce and continue to increase: P. infuscatus, V. chilensis, Rupornis magnirostris, Elanus leucurus, I. icterus, Quiscalus lugubris; Asio clamator became established but did not increase thereafter. Several other species showed significant increases: P. montagnii, Coragyps atratus, C. coruscans, two species of Grallaria, S. subpudica, T. melancholicus, M. leucophrys, Troglodytes aedon, Turdus fuscater, Hemispingus superciliaris, two species of Diglossa, Atlapetes schistaceus, Cacicus chrysonotus, and Icterus chrysater. On the other hand, the regression results for several species of permanent residents showed significant decreases: Butorides striata, Tyto alba, Eriocnemis cupreoventris, Ochthoeca fumicolor, C. apolinari, Diglossa lafresnayii, two species of Catamenia and Spinus spinescens (Figure 7). The species that either increased or decreased significantly or slightly (0.05 < p < 0.10; Table S3) were ecologically and taxonomically diverse, indicating multiple factors involved in these changes.
Table 4
| Situation | Types of abundance changes | Total | |||
|---|---|---|---|---|---|
| I | D | NC | NA | ||
| PR-Permanent resident | 36 | 24† | 53 | 14 | 127 |
| SR-Seasonal resident | 11 | 3 | 18 | 10 | 42 |
| MV-Mountain Visitors | 0 | 0 | 0 | 8 | 8 |
| HV-Highland Visitors | 1 | 3 | 3 | 8 | 15 |
| LV-Lowland Visitors | 0 | 0 | 1 | 30 | 31 |
| RV-Reproductive Visitors | 0 | 0 | 1 | 2 | 3 |
| TV-Transient Visitors | 0 | 0 | 0 | 4 | 4 |
| IV-Introduced Visitor | 0 | 0 | 1 | 4 | 5 |
| Total | 48 | 30 | 77 | 80 | 235 |
Summary of abundance changes of bird species due to the different factors affecting them in the Sabana de Bogotá Audubon's Christmas Bird Counts circle in 26 years (1989–2014).
I, increase; D, decrease; NC, no change; NA, not analyzed (see text).
Includes two species (Ixobrychus exilis, Eremophila alpestris) that disappeared from the circle in the first years of the CBC without having accumulated 5 years of records; they were not analyzed with regressions but were included as decreases.
Figure 7
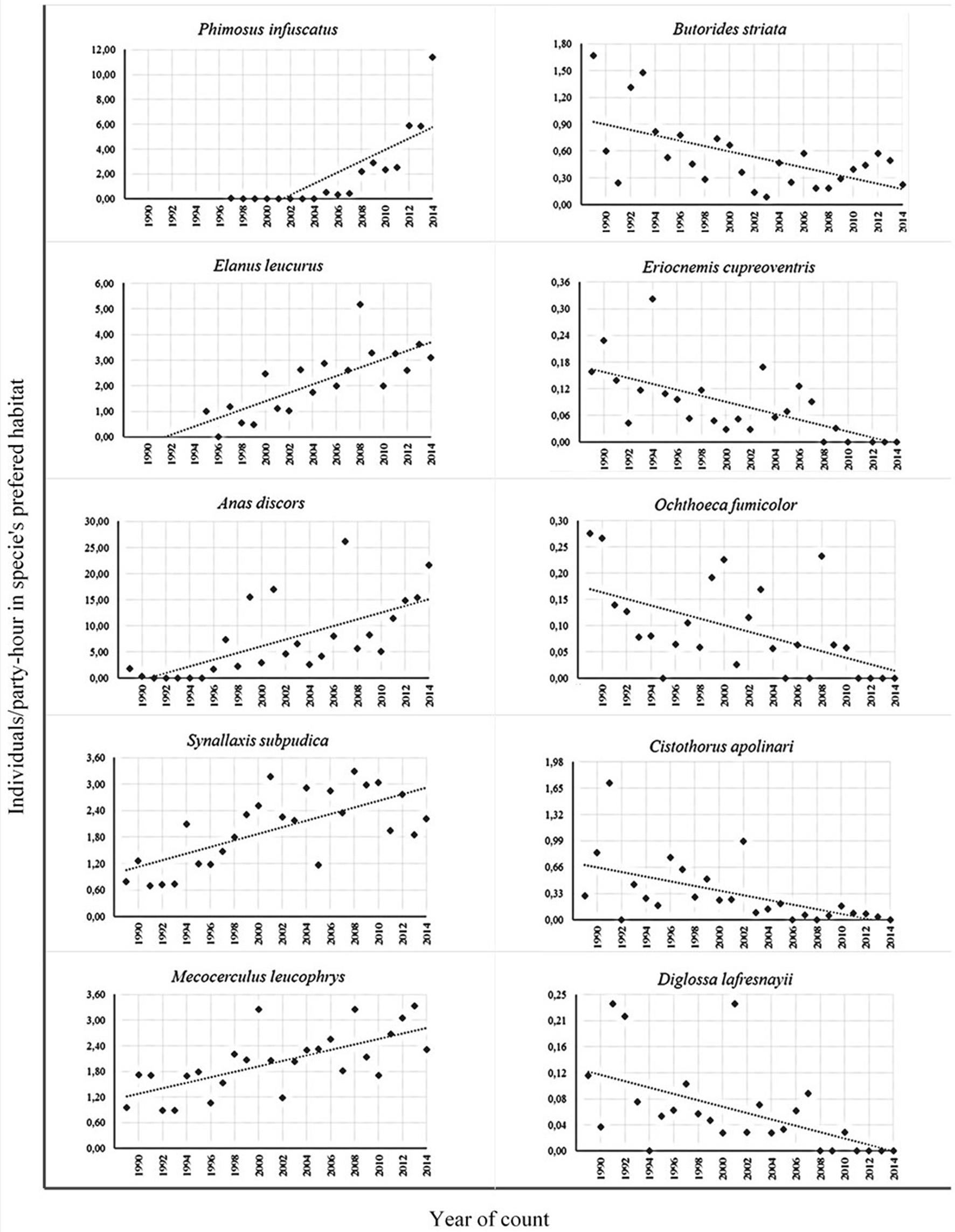
Examples of bird species that increased or decreased significantly between 1989 and 2014 in the Sabana de Bogotá Audubon's Christmas Bird Count circle.
In addition to these unidirectional changes, in seven species of permanent residents the direction of changes varied within the count period. For most, a period of 10 years of increases (B. ibis, Z. auriculata, T. episcopus, and palmarum) or stability (Elaenia frantzii, Zonotrichia capensis) was followed by several years of decreases (Table 5). M. bonariensis was an exception: its abundance increased significantly at the beginning of the study (not including two “outliers” that corresponded to flocks overflying the area), then was followed by several years with a marginally slight increase (Table 5, Figure 8).
Table 5
| Species | Years | Slope | p | Interpretation | Possible cause and comments |
|---|---|---|---|---|---|
| Bubulcus ibis | 1989–1999 | 3.237 | 0.043 | Significant increase | Reproduction increase in circle |
| 2000–2014 | −1.633 | 0.019 | Significant decrease | Reduction in pasture areas because of urbanization, drainage of a wetland site | |
| Zenaida auriculata | 1989–2003 | 3.729 | 0.001 | Significant increase | Climate change and adaptation to urban environment? |
| 2004–2014 | −2.606 | 0.009 | Significant decrease | Increase in mortality due to predation, diseases? | |
| Elaenia frantzii | 1989–1998 | 0.182 | 0.165 | No change | |
| 1999–2014 | −0.045 | 0.006 | Significant decrease | Unknown | |
| Thraupis episcopus | 1989–2005 | 0.055 | 0.010 | Significant increase | Climate change and adaptation to urban environment? |
| 2006–2014 | −0.122 | 0.062 | Slight decrease | Brood parasitism by Molothrus bonariensis? | |
| Thraupis palmarum | 1989–2006 | 0.092 | 0.001 | Significant increase | Climate change and adaptation to urban environment? |
| 2007–2014 | −0.080 | 0.062 | Slight decrease | Brood parasitism by Molothrus bonariensis? | |
| Zonotrichia capensis | 1991–2009 | −0.273 | 0.186 | No change | Two atypical outliers removed (see text) |
| 2010–2009 | −2.707 | 0.044 | Significant decrease | Brood parasitism by Molothrus bonariensis? Changes in urbanization? | |
| Molothrus bonariensis | 1989–1998 | 1.086 | 0.014 | Significant increase | Reproduction increase in circle |
| 2000–2014 | 0.168 | 0.102 | No change |
Complex changes in abundance trends in seven resident species throughout the years of the Christmas Bird Counts of the Sabana of Bogotá (1989–2014).
Figure 8
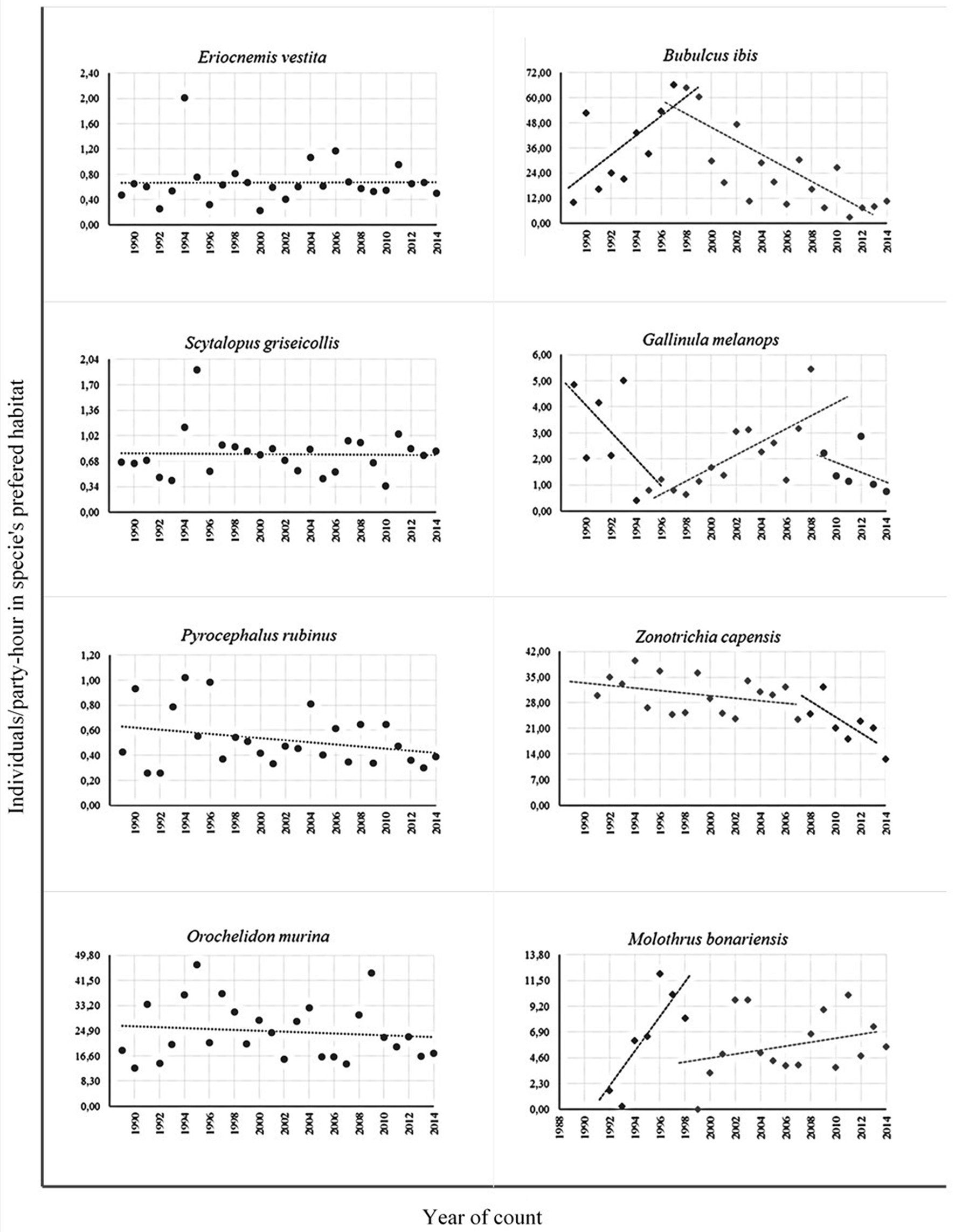
Examples of bird species that were stable or had complex tendencies in which the direction of change varied along the period between 1989 and 2014 in the Sabana de Bogotá Audubon's Christmas Bird Count circle.
Among the 46 seasonal residents (wintering boreal migrants), 11 increased their abundance at least slightly (26%) whereas only three (7%) decreased and 18 (43%) did not change according to regression results (Table 4). Since the species that increased were mainly known as passage migrants, it is evident that several of them were becoming winter residents during this period. In fact, regressions could be underestimating this change; when we compared the number of years in which several of these species were recorded in the first vs. the second half of the study period, five showed more frequent records in the last 13 year even though their regressions did not detect numeric changes in abundances (e.g., Cardellina canadensis and Contopus cooperi, Table S3); three additional species not analyzed with regressions were also recorded more frequently during the second half of the study (Ardea herodias, Butorides virescens, and Myiarchus crinitus). We may conclude therefore that 19 species (45%) increased as winter residents during the study (Table S3), reinforcing this unidirectional tendency of changes. The records of Gallinago delicata and Hirundo rustica decreased during the second half of the study. By contrast, among the visitors from higher elevations one species apparently increased in abundance (Myiotheretes striaticollis) but three species of hummingbirds decreased; for three more, regressions did not show significant changes and the other eight species did not have enough years of records to perform the analysis. For the other types of visitors very few species (7%) could be analyzed with regressions; in all such cases (one lowland visitor, one reproductive visitor, and one introduced visitor) there were no changes; the remaining 93% were not analyzed due to insufficient years of records (Table 4, Table S3).
The abundance tendencies between rural and urban localities for two of the most common species in the study (Z. auriculata and T. fuscater) were very different: in both cases the situation outside the city was very stable (slope = −0.0003 for Z. auriculata, p = 0.99 and 0.0044 for T. fuscater, p = 0.81) whilst in the urban localities the slope was 2.365 (p = 0.005) between 1986 and 2003 and −1.915 (p = 0.078) between 1994 and 2014 for Z. auriculata and 0.105 (p = 0.203) for T. fuscater (Figure 9).
Figure 9
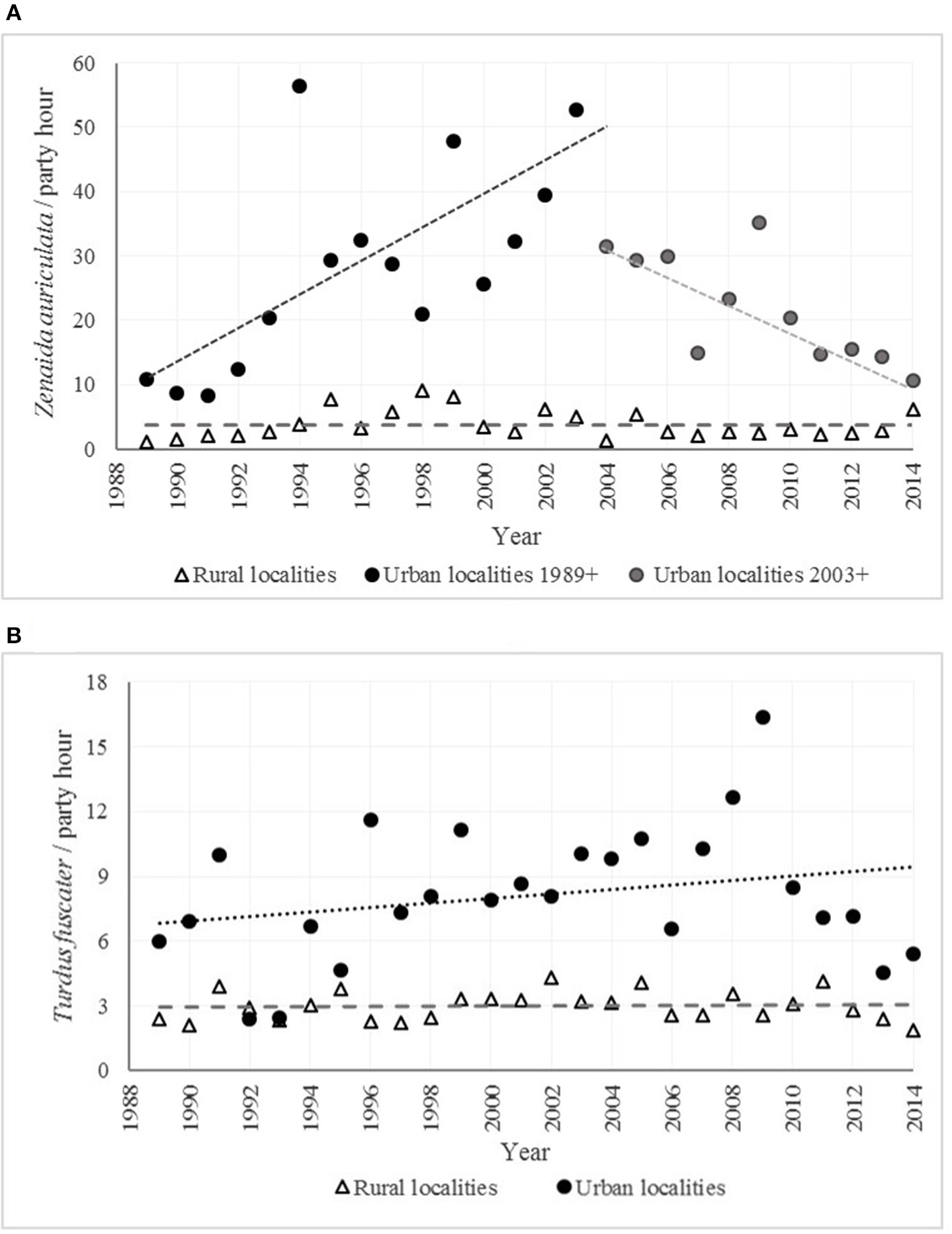
Abundance tendencies in rural and urban localities for two of the most common species in the Sabana de Bogotá Audubon's Christmas Bird Counts. (A)Zenaida auriculata (Eared Dove). (B)Turdus fuscater (Great Thrush).
Discussion
The most difficult and important aspect of this study is to assign possible causes to the abundance changes found in the 85 species (55%) analyzed with regressions or frequency of occurrence. This task is made more difficult given that many of these changes could have been due to a combination of factors that operate simultaneously and that the same type of factor could benefit some species and affect negatively others. To begin, we classify the factors that could cause the changes observed in bird abundance in five general groups and then analyze their effects in further detail.
Lack of continuity in observations in particular sites
This factor has been a problem throughout the study period, largely reflecting the representation of different habitats in the final results. The most evident example is the representation of well-preserved forest in the mountain slopes, the habitat with the highest species richness, in which many species occur naturally in low densities. Only one locality with this kind of forest had continual observations over almost all the study period but in 2000, we added a second site. The duplication of forest sites may have caused the apparent increases in regressions of some forest species (e. g., Glaucidium jardini, Margarornis squamiger, Coeligena helianthea, C. chrysonotus), which therefore might not indicate real increases in their populations. Also, the increase in some nocturnal species (e.g., Megascops choliba) may reflect more time invested in nocturnal auditory surveys in the last years, more than real changes. Finally, during these years the availability of recordings of several forest species in particular helped in their detection and identification. Nevertheless, when we checked the data we did not find major differences in the amount of auditory vs. visual detections along the period.
Direct human actions
The reduction in hunting of P. montagnii in several forest localities could have well caused its increase. The reduction also of the intense hunting of Z. auriculata reported by Olivares (1969) could explain partially its marked increase in the first half of the study. Another positive change could have been the diminution of trapping of I. chrysater as a cage bird, reported as common by Olivares in the past; we recorded only one or two cases of this species inside a cage in the first years of count. On the other hand, there was also disruption of a nesting colony of B. ibis and Nycticorax nycticorax in 2000, apparently caused by “hunters” in one of the wetland localities (Vargas, personal communication).
However, the most important human actions during the study period were those that caused changes in bird habitats. There were three types of interventions: changes in several wetlands, urbanization and tree planting in several urban parks and avenues, and some wetland boundaries and reduction of open pastures. During several years Bogotá's water company (EAAB) performed major works in the city's wetlands, some beneficial and some harmful. New open water areas were opened and bulrush (Schoenoplectus californicus) areas increased in some wetlands, benefiting some aquatic species. However, the water supply of some other wetlands was reduced or eliminated by flood control measures, and the interruption of the constant and necessary removal of water hyacinth eliminated open water habitat in one of the most important wetlands, eradicating the largest population of the endemic and endangered subspecies of P. melanops. Another population of this species was eliminated with the drainage of a rural pond between 2009 and 2011, which also affected other species such as O. jamaicensis and Podilymbus podiceps. Most of the remaining population of P. melanops in the circle survives in some artificial ponds and along the Bogotá River.
The effect of urbanization was noted mainly with the construction of commercial establishments, greenhouses for flower production and housing projects over areas formerly covered by pastures and crops, removing habitat for several species including B. ibis, T. alba, Sturnella magna, Sicalis luteola, and Catamenia analis and hence the decrease of their populations. Another effect of urbanization in the northern portion of the city was the replacement of single family houses with gardens and trees by apartment blocks with few green areas, possibly affecting some urban birds such as Diglossa humeralis, Z. capensis and possibly the urban population of T. fuscater, which has apparently declined since ca. 2010; however, Z. auriculata could apparently adapt to this change.
There have been two types of tree planting programs: the planting of exotic plants including some with fleshy fruits (especially Syzygium paniculatum) in parks and city avenues, which seem to have benefited several migrants and T. fuscater in particular: its population has increased in the study period in the city but not in natural areas with shrubs and forest. Instead, tree planting in the boundaries of some wetlands was done with a predominance of native species and favored the increase of the urban populations of several species such as M. leucophrys, T. aedon, and possibly Arremon assimilis and the wintering populations of some migrant species. The establishment or improvement of gardens in some parks could also have favored species such as C. coruscans and Diglossa sittoides; the latter seems to have expanded its local distribution during the counts.
“Natural” changes in some habitats
Even though these types of changes may be the secondary effects of human actions, they were not produced deliberately. An example is the secondary succession of native scrub toward secondary forest in several sites after deforestation or fires in previous years. In one of the forest localities that was visited continuously for 24 years, scrub vegetation has grown from a height of 1–3 m in 1991 to 3–6 m at present with more trees in several places. To a greater or lesser degree a similar situation has occurred in other localities with long-term observations. The increase in observations of species such as C. helianthea, Grallaria squamigera and ruficapilla, Atlapetes schistacea and H. superciliaris are possibly related to these changes. Another change that has occurred in several wetlands is related to increase in water pollution: the filling of water bodies with macrophytes reducing open water areas and eliminating exposed mud on the edges with shallow waters. This is related to the increase in some species such as G. galeata and decrease in probers like Gallinago and Tringa spp. (Rosselli and Stiles, 2012a). The increase in the shrub vegetation around the edges of some wetlands has probably induced the increment in S. subpudica and perhaps also T. aedon. The disappearance of the endemic and endangered subspecies Eremophila alpestris peregrina is probably related to the coverage with introduced kikuyu grass in a newly established park at the southern end of the circle that had bunch grasses and areas of bare soil, an important habitat for this species, at the beginning of the study.
Interactions with other species
An interesting case is the increase in the abundance of Buteo platypterus as a wintering migrant, which is probably related to the significant increase of its favorite prey, the dove Z. auriculata, during most of the study period. The reduction of Z. auriculata in the last years (even though it is still the most common species in the circle) could be associated partially with this predation, although its high reproductive rate (nests are present all year long) would not be enough to explain its recent decline. Another possibility could be the increase in parasites or diseases in the population. Until recently we have observed dormitories of hundreds of birds in dense trees in city parks and such congregations are favorable for their transmission (Delgado-V and French, 2011). The recent occurrence of the snail specialist Chondrohierax uncinatus apparently reflected the appearance of the Giant African Snail (Achatina fulica) in the Sabana de Bogotá and nearby mountain localities; this species is a rapidly increasing agricultural pest and health threat (Linares et al., 2013).
T. fuscater regularly predates on nestlings and fledglings of Z. capensis but this is unlikely to have caused the decrease of the sparrow in recent years; the data show that the Z. capensis population was stable for many years during which the T. fuscater population increased. Another more worrying example of predation is the hunting of aquatic birds by dogs (domestic dogs let loose by neighboring owners or feral dogs that reproduce in the wetlands). This hunting almost certainly has caused the local extinction of the endemic subspecies of I. exilis and possibly the decreases of B. striata and the endemic and endangered R. semiplumbeus in the circle. Although, cats are widely known to have significant effects on bird and other wildlife populations, especially in the United States and on some islands previously lacking mammalian predators (Medina et al., 2001; Nogales et al., 2013; Seress and Liker, 2015; Doherty et al., 2016), they do not appear to have had a significant impact on the birds of the Bogotá area. Contrary to the high number of feral dogs ranging in the wetlands, cats were rarely seen in the circle sites. Campos et al. (2007) compared the distribution and analyzed the diet of feral cats and dogs in Brazil and found that both were more abundant in suburban than in rural areas and their diet consisted mostly of invertebrates, followed by mammals and birds. We have found no other published studies of cat predation on birds in Latin America, but such studies are badly needed.
Nevertheless, the most worrisome interaction has been the increase in the brood parasitism by the cowbird M. bonariensis on various species. The recent decrease of Z. capensis may rather be the result of cowbird parasitism and in a lesser degree, to urbanization. An informal count on the Universidad Nacional campus in 2013–2014 (Stiles, personal observation) recorded more pairs of this species feeding fledgling cowbirds than those of their own species. This parasitism is the most probable cause of the dramatic decline of the endemic species C. apolinari, now at the brink of extinction in the city's wetlands despite the protection and improvement of its bulrush habitat (S. californicus) in several wetlands (Rosselli and Stiles, 2012b). Velásquez-Tibatá et al. (2000) first noted this parasitism and later it was observed in other localities; a nest collected in one wetland in the circle had more M. bonariensis eggs than those of its host (Castro et al., 2007). Villaneda and Rosselli (2011) found that M. bonariensis is more abundant in urban wetlands than in rural ones. The decline of T. episcopus, T. palmarum, and Ramphocelus dimidiatus in the urban areas could also be related to this parasitism; these species and others in their genera have been reported as hosts of M. bonariensis by Friedmann and Kiff (1985). However, another species also reported as a M. bonariensis host, Chrysomus icterocephalus (Naranjo, 1995), maintained a stable population during the time of the study. Villaneda and Rosselli (2011) noted aggressions from this species against the parasite, and it apparently may defend its nests more effectively.
Climate change
During the CBC period the mean annual temperature in Bogotá's airport increased by 0.65°C and the mean minimal annual temperature has risen by almost 2°C (Figure 4, Table 3). Several changes recorded in the avifauna in the CBCs match the predictions of transformations made by some theoretical models developed to assess such phenomena (Velásquez-Tibatá et al., 2013; see also the revision by Herzog et al., 2011). The clearest indication of the effect of global warming on birds is the change in the lower or upper elevational limits of species. The counts have documented the displacement of the lower limits of E. cupreoventris, O. fumicolor, Catamenia inornata, and D. lafresnayii from ca. 2,600 m at the level of the flat area of the Sabana to at least 2,850 m in the surrounding mountains. Several species have become established as residents in the flat part of the Sabana at a higher elevation than that reported for reproduction according to Hilty and Brown (1986), including P. infuscatus, R. magnirostris, A. clamator, Q. lugubris, and I. icterus (although the origin of this population is probably from escaped cage birds, it has been reproducing and it seems to be increasing). Both I. icterus and Q. lugubris were exclusive to urban areas suggesting the influence of the city's “heat island”; otherwise they could have been registered in areas outside the city where exotic as well as native vegetation can be found and would provide resources to these species. Other species known mainly or exclusively from the southwestern drier and warmer parts of the Sabana have expanded their distributions as residents widely toward the north and east (E. leucurus, V. chilensis, Mimus gilvus and probably M. bonariensis and B. ibis). It is possible that the “heat island” in most of the city also has facilitated this expansion. The activity and abundance of heterothermic animals including insects may be related to temperature (Garcia et al., 2008; Coutinho-Silva et al., 2017); lower temperatures can cause longer biological cycles, reduced fertility and even total interruption of development (Hodkinson, 2005; Nava et al., 2007). Moreover, upslope displacements of various insect groups in response to climate change have been recorded (e.g., for butterflies see Forister et al., 2010 and Chen et al., 2011). Hence, the warming of the Sabana could have produced an increment in prey for insectivorous birds such as T. melancholicus in addition to the various insectivorous boreal migrants that increased their abundance and frequency during the study period. In fact, it appears that climate change and the heat island might have acted synergistically in such cases, particularly favoring the establishment of those species now found mainly in the flat parts of the Sabana. However, the upslope movements of some forest species in parts of the circle beyond the heat island are more likely to reflect climate change.
Climate change may be related to observed changes in the elevational movements of some species, hummingbirds in particular. During the first years of the study hummingbirds from higher elevations were fairly frequently recorded in the counts in scrub vegetation on lower mountain slopes around the Sabana; these included Aglaeactis cupripennis, Pterophanes cyanoptera, and Lesbia victoriae. Examining specimens from the ornithological collection of the Universidad Nacional de Colombia, the time of the counts seems to match the molting season for these species: effectively this period represented a “molt migration.” All of them declined in frequency or ceased to appear during the CBC period. The contrary apparently happened with C. coruscans, which at the beginning became much scarcer between December and March–April, also in the molting period. However, presently many more individuals now remain in the Sabana during that season, possibly reflecting in part the planting of trees and increase in gardens, particularly in the urban areas.
Finally, we recorded changes in a few species that do not match easily with the main categories. The increase of C. atratus could be related simply to the increase of human population (and the consequent increase in dumps and waste); we have no explanation for the reduction of Contopus fumigatus and for others (e.g., T. aedon), we are not sure that the suggested factors as causes may explain the magnitudes of the observed changes.
Summarizing the results (Table 6) we find that climate change, possibly abetted by the city's heat island, and their related effects seem to be the most important factor associated with the changes in abundance in permanent and seasonal residents in the Sabana. The pattern of the population changes is consistent with climate change for more than a third of the cases (ca. 38%). Next in importance are changes resulting from direct human actions (ca. 19%) and those resulting from “natural” changes (ca. 18%): combining these two results in the diverse changes in habitats in the circle having a similar magnitude to climate change. The lack of continuity could partially explain 12% of the observed changes, although possibly not reflecting real changes in populations in several cases; interactions between species appears to be related to ca. 11%, with a small residual of 1–2% without clear explanations.
Table 6
| Factor | Number of species |
|---|---|
| 1. Lack of continuity | 12–15 |
| a. Changes in localities and habitat representation | 11+ |
| b. Changes in observation time in some localities | 1+ |
| c. Changes in methodologies (using more vocalizations) | 2–3 |
| 2. Direct human actions | 18–21 |
| a. Changes in certain habitats | |
| i. Wetlands intervention | 5+ |
| ii. Reforestation actions in some parks and other urban areas | 4–6 |
| iii. Urbanization: Building of housing projects, warehouses and commercial buildings on pastures and crop areas; replacement of houses that had front and backyard gardens with apartment buildings with little green areas | 6–8 |
| b. Hunting and other disturbances | 2 |
| 3. Habitat changes with no human intervention | 17–20 |
| a. Secondary succession of native thickets to next stages (higher scrub, secondary forest) or bare soil changing to lawn | 10+ |
| b. Filling of water bodies with macrophytes reducing open water areas and eliminating narrow mudflats along the water edges | 7 |
| c. Increase in the shrub vegetation around some wetlands or from the edges inside the wetland | 3–5 |
| 4. Interactions with other species | 10–12 |
| a. Predation | |
| i. Predation among bird species | 1–2 |
| ii. Predation by dogs in wetlands | 3+ |
| b. Parasitism | |
| i. Parasitism by other birds, mainly Molothrus bonariensis | 4–5 |
| ii. Parasitism by parasites, microbes, etc. and disease spreading | 1? |
| 5. Climate change because of local and global warming | 37–42+ |
| a. Changes in the lower or upper elevational limits of species | 8+ |
| b. Easier movement of some species inside the Sabana | 7–9 |
| c. Changes in the altitude movement patterns | 4–5 |
| d. Changes in status or abundance of species already registered in the circle | 18–20 |
Number of species with changes in status or abundance recorded in the Audubon's Christmas Bird Counts of the Sabana of Bogotá over 26 years (1989–2014), for each of the five factors that could have caused these changes (positive or negative).
The number of species is greater than the registered changes number (see Table S1) because many changes reflect the effects of a combination of factors.
The recent study by Rosselli et al. (in press) in high Andean forest and subpáramo between 2,800 and 3,300 m complements the results we present here. The work was done in high Andean forest, lower paramo and paramo areas between 2,800 and 3,300 m to replicate a previous study (Stiles and Rosselli, 1998) that comprised a period of time comparable to the counts although it was not continuous. Among the species that the counts were found moving toward those elevations were R. magnirostris, E. frantzii, and T. melancholicus. The increase of E. frantzii concurred with its decline in the Sabana, as did the increase of O. fumicolor in the paramo, so the abundance changes of these species could have well been the result of climate change.
A look at the past
Despite differences in methods and scope, the study by Olivares (1969) provides a basis for comparison with our results comprising the period up to the 1960's and for evaluating some changes that occurred during the two decades prior to the initiation of the CBC. Among the differences, the areas are not exactly the same: Olivares included larger areas both south and west from our CBC circle with habitats and species not found in it; on the other hand the CBC included high locations on the eastern mountains and a river valley not mentioned by Olivares, who apparently only studied places on the western base of the mountains. However, his comments on the abundances of a considerable number of are applicable to the CBC circle and will be discussed here.
Olivares (1969) recorded ca. 180 species in his more broadly defined Sabana de Bogotá. At least 15 of these species, including V. chilensis and R. magnirostris, were recorded only in the drier and warmer parts of the Sabana south and west of the CBC circle, within which they have since become established. In addition, several species that Olivares considered accidental in the Sabana had apparently become established in the two decades between 1969 and the initiation of the CBC, all originating from lower elevations in the Magdalena valley to the west, including E. flavogaster, P. sulphuratus, T. episcopus, T. palmarum, R. dimidiatus, Sicalis flaveola, and Icterus nigrogularis. The origins of these species could suggest that the effects of global warming were already being felt in the Sabana and the temperature records from the Eldorado airport since 1960 support this, although the rate of temperature increase and the intensity of the city's heat island have evidently augmented through the CBC period. On the other hand, two species recorded by Olivares as common (E. alpestris and Asio flammeus) had become very rare by 1969, and several other species of agricultural areas had decreased during this interval (e.g., Colinus cristatus, S. luteola, and Sporophila luctuosa), which coincides with the replacement of croplands by pastures over much of the Sabana. The CBC counts also recorded at least 20 species from the highest forests in the circle that apparently were not visited by Olivares or his collaborators. Very few of the species recorded in the counts as visitors from lower elevations were mentioned by Olivares, as well as none of those recorded as visitors in transit (TV) or introduced (IV), and the CBCs did not record several visitors from mountain forests (MV) (Pharomachrus antisianus, Geotrygon montana, Claravis mondetoura) mentioned by Olivares. Some transient migrants considered as common in the Sabana are at best rare now, including Progne subis; Antrostomus carolinensis, and Charadrius vociferus, which he considered to be common winter residents, were not recorded in the CBC although there are a few recent records from beyond the circle.
The situation of the aquatic birds is even more dramatic, and the information gathered by Fjeldså (1985) and van der Hammen et al. (2008) supplements the records of Olivares (1969) for the pre-CBC period, during which the most drastic reduction of the wetland areas occurred. Populations of several waterbird species were extirpated during this period, including Podiceps andinus, Anas spinicauda niceforoi, A. cyanoptera borreroi, Circus cinereus, Botaurus pinnatus, and Netta erythrophthalmus, the first three of which were endemic. Also, several waterbird migrants considered regular by Olivares were not recorded during the CBC counts, including Anas acuta, A. americana, A. clypeata, Aythya affinis, Pluvialis squatarola y P. dominica, Charadius vociferus y Numenius phaeopus. Moreover, ca. 10 species of large waterbirds mentioned as accidental by Olivares have not been recorded since.
Clearly the avifauna of the Sabana de Bogotá has been highly dynamic at least since the middle of the twentieth century. Prior to the CBC period, direct human actions including wetland drainage, expansion of urbanization, changes in rural land use appear to have been the most important factors affecting the avifauna, diminishing aquatic bird diversity in particular. The present study has documented the impacts of urbanization in rural areas, human intervention and succession in wetlands and forest succession on some mountain slopes as explanations for many avifaunal changes over the last 26 years. However, it has also added climate change, perhaps abetted by the city's heat island, as a possible driver of avifaunal changes.
A glimpse into the future
All indications are that the changes in Bogotá's avifauna during the CBC will continue; the effects of climate change and urbanization may well intensify in some of the sites, especially if the present administration's intentions to urbanize the largest remaining green area close to the northern city limits (Reserva Forestal Van der Hammen, CAR, 2011) materialize. We may therefore expect that the frequency and variety of boreal migrants remaining to winter in the Sabana will probably increase and that some native species now considered as visitors from lower elevations may become established. On the other hand, we could expect that some species whose lower elevation limits have moved upward might disappear from the count circle, as almost surely will the endemic and threatened wren species C. apolinari due to cowbird parasitism; it will be important to monitor the abundances of other species known or suspected to be suffering from this brood parasite. The plans of the district government to urbanize much of the rural area remaining in the circle could, if implemented, affect the populations of several open country species. Although, La Sorte et al. (2009) reported delays of up to 35 years in the latitudinal response to climate change of some North American species, data are still scarce for altitudinal changes by Andean species to climate change (Forero-Medina et al., 2011; Velásquez-Tibatá et al., 2013) and the Sabana de Bogotá could continue to contribute important data on this phenomenon. The most general conclusion is that the Sabana de Bogotá CBC has supplied much important information about its changing avifauna, and highlights the importance of continuing it for many more years.
The present study in the context of urban ornithology in latin america
When the Sabana de Bogotá CBC was begun, urban ornithology was still in its infancy in Neotropical countries although it was gaining recognition as a major field of study in North America and Europe (e.g., Chace and Walsh, 2006; Ortega-Álvarez and MacGregor-Fors, 2011; Marzluff, 2016). Because the Neotropics host the world's richest avifauna, biodiversity loss through urbanization was the main concern of most publications. Major reviews of this field in Latin America (Ortega-Álvarez and MacGregor-Fors, 2011) and Colombia (Delgado-V and Correa-H, 2013) found two main themes of most publications: compilation of species lists and comparisons of avian communities varying from natural vegetation through rural, periurban and urban areas, mostly around major cities (e.g., Fandiño et al., 2010; Biamonte et al., 2011; Lessi et al., 2016). In particular, many studies attempted to evaluate the importance of urban green islands like urban parks and university campuses with different proportions of native and exotic vegetation for conservation of native avian biodiversity (e.g., Muñoz et al., 2007; Vásquez-Muñoz and Castaño-Villa, 2008; Lessi et al., 2016). However, the temporal dynamics of avifaunal change due to urbanization over periods of decades were rarely documented because data for most studies were obtained over short periods, usually 2 years or less. That the importance of urban green islands cannot be appreciated over such short periods is demonstrated by the results for the Bogotá Botanical Garden: the mean number of species counted per year was ca. 32, but over the 26 years of the CBC, three times that number of species have been recorded. An exception was a qualitative description of avifaunal changes over a 20-year period as coffee plantations were converted to suburbs in Costa Rica (Stiles, 1990). The few studies that attempted to include a temporal component involved a comparison of historical data with current inventories 50 or more years later (e.g., Castaño-V and Castaño-V, 2000; Donegan et al., 2009).
The present study thus helps to fill a gap in Neotropical urban ecology and is the first to explicitly address the importance of climate change in the context of urban-periurban-rural-natural vegetation involving a range of elevations. However, several other CBCs have been conducted in Colombia since the 1990s in similar situations, notably in the cities of Medellín and Manizales (e.g., Cuadros, 1992), but the results of these have yet to be synthesized and analyzed. We hope that the present study will stimulate such analyses, particularly with respect to the possible impacts of climate change. Although, not a primary objective, our study has produced 26-year species lists for sites with different degrees of habitat alteration that could be analyzed from our data. However, the present study also is incomplete as regards the latter stages of urbanization in that it included no sites in completely urban or suburban situations without such green islands, which should be considered for the future.
Other aspects requiring more study, as recommended by Delgado-V and Correa-H (2013) deal with comparisons of the ecology of species successful vs. unsuccessful in occupying urban habitats, the importance of specific factors favoring (e.g., additional food resources, possible reductions of predation, especially of nests) vs. inhibiting (predation by cats, collisions with structures and windows, parasites and diseases, lack of specific food types such as insects in exotic vegetation) for the occupation of urban areas by native species. A further area needing study is the human-bird interaction itself, involving the attitudes of local residents toward birds and methods of environmental education to permit increased citizen involvement in biodiversity conservation. At least in Colombia, a number of such programs have been instituted but their results need to be evaluated and published. Finally, an aspect emphasized by Marzluff and Rodewald (2008) is the importance of involving city planners and managers in designing and improving green areas as urbanization proceeds, a critical need for the future of a rapidly expanding city like Bogotá.
Statements
Author contributions
FS coordinated the Bogotá CBC in 25 of the 26 years included in this article, collected and curated the observers' field data. FS, LR and SD initiated the CBC circle in Bogotá, participated in the counts, digitized and analyzed the data, and wrote and approved the manuscript.
Funding
The Universidad Nacional de Colombia and the Universidad de Ciencias Aplicadas y Ambientales U.D.C.A. granted time to FS and LR for the writing of the paper. The Science and International Alliances from National Audubon Society provided funding that made possible the publication of this article.
Acknowledgments
The authors express their gratitude to landowners and institutions that have permitted the entrance of CBC participants to many localities for more than 20 years. The counts could not have been made without the hundreds of bird watchers and the locality compilers that with enthusiasm have risen early and have enjoyed the birds. This type of participation is the essence of citizen science. We thank Bogotá's Ornithological Association (ABO) for its continuous support and coordination of the CBC. J. Niño and J. Pinzón from the Universidad de Ciencias Aplicadas y Ambientales (U.D.C.A.) offered valuable help with data organization. J. Cifuentes and S. Velásquez-Licona greatly helped with data analysis and preparation of the figures, and J. A. Vargas-González helped prepare Figure 1. For many years the National Audubon Society waived registration fees for the Bogotá CBC count participants. We especially thank Matthew Jeffery, Gary Langham, and Geoffrey S. LeBaron from the National Audubon Society for their help in obtaining funds to assure the publication of this article.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Supplementary material
The Supplementary Material for this article can be found online at: http://journal.frontiersin.org/article/10.3389/fevo.2017.00058/full#supplementary-material
References
1
AndersonE. P.MarengoJ.VillalbaR.HalloyS.YoungB.Corderoet al. (2011). Consequences of climate change in the tropical Andes, in Climate Change and Biodiversity in the Tropical Andes, eds HerzogS. K.MartínezR.JørgensenP. M.TiessenH. (Sao Jose dos Campos: Inter-American Institute for Global Change Research and Scientific Committee on Problems of the Environment), 1–18.
2
AndradeG. (1998). Los humedales del altiplano de Cundinamarca y Boyacá. Ecosistemas en peligro de desaparecer, in Una Aproximación a los Humedales de Colombia, ed GuerreroE. (Bogotá: Fondo FEN), 59–72.
3
AngelL.RamírezA.DomínguezE. (2010). Isla de calor y cambios espacio-temporales de la temperatura en la ciudad de Bogotá. Rev. Acad. Colomb. Cienc.34, 173–183.
4
ArbibR. S. (1981). The christmas bird count: constructing an ideal model, in Estimating Numbers of Terrestrial Birds, Studies Avian Biology, Vol. 6, eds RalphC. J.ScottJ. M. (Camarillo, CA: Cooper Ornithological Society), 30–33.
5
ArbibR. S. (1982). “Ideal model” Christmas Bird Counts: a start in 1982–83. Am. Birds36, 146–148.
6
BiamonteE.SandovalL.ChacoE.BarrantesG. (2011). Effect of urbanization on the avifauna in a tropical metropolitan area. Landsc. Ecol.26, 183–194. 10.1007/s10980-010-9564-0
7
BockC. E.RootT. L. (1981). The christmas bird count and avian ecology, in Estimating Numbers of Terrestrial Birds, Studies Avian Biology, Vol. 6, eds RalphC. J.ScottJ. M. (Camarillo, CA: Cooper Ornithological Society), 17–23.
8
BonneyR. E. (1979). Wintering Peregrine Falcon populations in the eastern United States 1940-1975: a Christmas Count analysis. Am. Birds33, 695–697.
9
BothC.ArtemyevA. V.BlaauwB.CowieR. J.DekhuijzenA. J.EevaT. (2004). Large-scale geographical variation confirms that climate change causes birds to lay earlier. Proc. R. Soc. Lond. B.271, 1657–1662. 10.1098/rspb.2004.2770
10
ButcherG. S.McCullochC. E. (1990). Influence of observer effort on the number of individual birds recorded on Christmas Bird Counts, in Survey Designs and Statistical Methods for the Estimation of Avian Population Trends, U.S. Fish and Wildlife Service Biology Report, Vol. 9, eds SauerJ. R.DroegeS.120–129.
11
CamposC. B.EstevesC. F.FerrazK. M. P. M. B.CrawshawP. G.Jr.VerdadeL. M. (2007). Diet of free-ranging cats and dogs in a suburban and rural environment, south-eastern Brazil. J. Zool.273, 14–20. 10.1111/j.1469-7998.2007.00291.x
12
Corporación Autónoma Regional de Cundinamarca (CAR) (2011). Acuerdo 11 de 2011 “Por medio del Cual se Declara la Reserva Forestal Regional Productora del Norte de Bogotá D.C.Bogotá: “Thomas van der Hammen”, se Adoptan unas Determinantes Ambientales para su Manejo, y se Dictan Otras Disposiciones.
13
Castaño-VG. J.Castaño-VJ. C. (2000). Cambios en la composición de la avifauna en Santa Helena durante el siglo XX. Crónica Forestal y del Medio Ambiente15, 137–161.
14
CastroJ. A.BenítezH. D.MoralesJ. E.CamposE. (2007). Primer registro de parasitismo por parte del “chamón” al Cucarachero de Pantano (Cistothorus apolinari) en el humedal de La Conejera, Bogotá. Restauración Ecológica, Humedal de La Conejera30–33.
15
ChaceJ. F.WalshJ. J. (2006). Urban effects on native avifauna: a review. Landsc. Urban Plan.74, 46–69. 10.1016/j.landurbplan.2004.08.007
16
ChaverriG.GarinI.AlberdiA.JimenezL.Castillo-SalazarC.AihartzaJ. (2016). Unveiling the hidden bat diversity of a neotropical montane forest. PLoS ONE11:e0162712. 10.1371/journal.pone.0162712
17
ChenI.-C.HillJ. K.OhlemüllerR.RoyD. B.ThomasC. D. (2011). Rapid range shifts of species associated with high levels of climate warming. Science333, 1024–1026. 10.1126/science.1206432
18
Coutinho-SilvaR. D.MontesM. A.OliveiraG. F.de Carvalho-NetoF. G.RohdeC.GarciaA. C. L. (2017). Effects of seasonality on drosophilids (Insecta, Diptera) in the northern part of the Atlantic Forest, Brazil. Bull. Entomol. Res. [Epub ahead of print]. 10.1017/S0007485317000190
19
CuadrosT. (1992). Resultados del conteo anual de aves en Medellín. Boletín SAO3, 6–11.
20
Delgado-VC. A.Correa-HJ. C. (2013). Estudios ornitológicos urbanos en Colombia: revisión de literatura. Ingeniería y Ciencia9, 215–236. 10.17230/ingciecia.9.18.12
21
Delgado-VC. A.FrenchK. (2011). Parasite-bird interactions in urban areas: current evidence and emerging questions. Landsc. Urban Plan.105, 5–14. 10.1016/j.landurbplan.2011.12.019
22
DohertyT. S.AlistairS. G.NimmoD. G.RitchieE. G.DickmanC. R. (2016). Invasive predators and global biodiversity loss. Proc. Natl. Acad. Sci. U.S.A.113, 11261–11265. 10.1073/pnas.1602480113
23
DoneganT.AvendañoJ. E.HuertasB.FlórezP. (2009). Avifauna de San Pedro de los Milagros, Antioquia: una comparación entre colecciones antiguas y evaluaciones rápidas. Bol. Cient Mus. Hist. Nat.13, 63–72.
24
DunnE. H.FrancisC. M.BlancherP. J.DrennanS. R.HoweM. A.LepageD. (2005). Enhancing the scientific value of the Christmas Bird Count. Auk122, 338–346. 10.1642/0004-8038(2005)122[0338:ETSVOT]2.0.CO;2
25
FandiñoB.BerducA. J.BeltzerA. H. (2010). Ensambles de aves en bosques nativos y exóticos en la estación reproductiva de un área protegida en el espinal de Entre Ríos, Argentina. Ornitol. Neotrop.21, 1–16.
26
FeeleyK. J.RehmE. M.MachovinaB. (2012). The responses of tropical forest species to global climate change: acclimate, adapt, migrate, or go extinct?Front. Biogeogr.4, 69–84.
27
FjeldsåJ. (1985). Origin, evolution, and status of the avifauna of Andean wetlands in Neotropical Ornithology, Vol. 36, eds BuckleyP. A.FosterM. S.MortonE. S.RidgelyR. S.BuckleyF. G. (Washington, DC: Ornithological Monographs), 85–112.
28
FoleyJ. A.DeFriesR.AsnerG. P.BarfordC.BonanG.CarpenterS. R. (2005). Global consequences of land use. Science309, 570–574. 10.1126/science.1111772
29
Forero-MedinaG.TerborghJ.SocolarS. J.PimmS. L. (2011). Elevational ranges of birds on a tropical montane gradient lag behind warming temperatures. PLoS ONE6:e28535. 10.1371/journal.pone.0028535
30
ForisterM. L.McCallA. C.SandersN. J.FordyceJ. A.ThorneJ. A.O'BrienJ.et al. (2010). Compounded effects of climate change and habitat alteration shift patterns of butterfly diversity. Proc. Natl. Acad. Sci. U.S.A.107, 2088–2097. 10.1073/pnas.0909686107
31
FreemanB. G.Class-FreemanA. M. (2014). Rapid upslope shifts in New Guinean birds illustrate strong distributional responses of tropical montane species to global warming. Proc. Natl. Acad. Sci. U.S.A.111, 4490–4494. 10.1073/pnas.1318190111
32
FriedmannH.KiffL. E. (1985). The parasitic cowbirds and their hosts. Proc. West. Found. Vertebr. Zool.2, 225–302.
33
GarciaA. C. L.ValiatiV. H.GottschalkM. S.RohdeC.ValenteV. L. S. (2008). Two decades of colonization of the urban environment of Porto Alegre, southern Brazil, by Drosophila paulistorum (Diptera, Drosophilidae). Iheringia Sér. Zool.98, 329–338. 10.1590/S0073-47212008000300007
34
GhalamborC. K.HueyR. B.MartinP. R.TewksburyJ. J.WangG. (2006). Are mountain passes higher in the tropics? Janzen's hypothesis revisited. Integr. Comp. Biol.46, 5–17. 10.1093/icb/icj003
35
GotelliN. JEllisonA. M. (2004). A Primer of Ecological Statistics. Sunderland, MA: Sinauer Associates, Inc.
36
GrimmN. B.ChapinF. S.III.BierwagenB.GonzalezP.GroffmanP. M.LuoY.et al. (2013). The impacts of climate change on ecosystem structure and function. Front. Ecol. Environ.11, 474–482. 10.1890/120282
37
HeY.ZhangY. (2005). Climate Change from 1960 to 2000 in the Lancang River Valley, China. Mt. Res. Dev.25, 341–348. 10.1659/0276-4741(2005)025[0341:CCFTIT]2.0.CO;2
38
HerzogS. K.MartínezR.JørgensenP. M.TiessenH. (eds.). (2011). Climate change and biodiversity in the tropical andes, in Inter-American Institute for Global Change Research (IAI) and Scientific Committee on Problems of the Environment (SCOPE) (Sao Jose dos Campos).
39
HiltyS. L.BrownW. L. (1986). A Guide to the Birds of Colombia. Princeton, NJ: Princeton University Press.
40
HodkinsonI. D. (2005). Terrestrial insects along elevational gradients: species and community responses to altitude. Biol. Rev.80, 479–513. 10.1017/S1464793105006767
41
HolmesR. T. (2011). Avian population and community processes in forest ecosystems: long-term research in the Hubbard Brook Experimental Forest. For. Ecol. Manage.262, 20–32. 10.1016/j.foreco.2010.06.021
42
IUCN (2017). The IUCN Red List of Threatened Species. Version 2016-3. Available online at: www.iucnredlist.org
43
JanzenD. H. (1967). Why mountain passes are higher in the tropics” and revisit of the topic. Am. Nat.101, 233–249. 10.1086/282487
44
JohnsgardP. A. (1967). Sympatry changes and hybridization incidence in mallards and black ducks. Am. Midl. Nat.77, 51–63. 10.2307/2423425
45
JohnsgardP. A (2015). Global Warming and Population Responses among Great Plains Birds. Zea E-Books. Book 26. Availabe online at: http://digitalcommons.unl.edu/zeabook/26
46
LarsenT. H.BrehmG.NavarreteH.FrancoP.GomezH.MenaJ. L. (2011). Range shifts and extinctions driven by climate change in the tropical andes: synthesis and directions, in Climate Change and Biodiversity in the Tropical Andes, eds HerzogS. K.MartínezR.JørgensenP. M.TiessenH. (Sao Jose dos Campos: Inter-American Institute for Global Change Research and Scientific Committee on Problems of the Environment), 47–67.
47
La SorteF. A.LeeT. M.WilmanH.JetzW. (2009). Disparities between observed and predicted impacts of climate change on winter bird assemblages. Proc. R. Soc. B276, 3167–3174. 10.1098/rspb.2009.0162
48
LessiB. F.Rodrigues-PiresJ. S.BatisteliA. F.MacGregor-ForsI. (2016). Vegetation, urbanization, and bird richness in a Brazilian periurban area. Ornitol. Neotrop.27, 2013–2010.
49
LinaresE.AvendañoJ.MartínezA.RojasA. (2013). El Caracol Gigante Africano Achatina Fulica, un Visitante Indeseado. Bogotá: Miniguía, Instituto de Ciencias Naturales, Universidad Nacional de Colombia.
50
Martínez-HerreraÓ. J. (2015). El proceso de urbanización en los municipios de la Sabana de Bogotá. Ánfora22, 85–111.
51
MarzluffJ. M. (2016). A decadal review of urban ornithology and a prospectus for the future. Ibis159, 1–13. 10.1111/ibi.12430
52
MarzluffJ. M.RodewaldA. D. (2008). Conserving biodiversity in urbanizing areas: nontraditional views from a bird's perspective. Cities Environ.1, 1–27. 10.15365/cate.1262008
53
MedinaF. M.BonnaudE.TershyB. R.ZavaletaE. S.DonlanC. J.BradfordS. K.et al. (2001). A global review of the impacts of invasive cats on island endangered vertebrates. Glob. Change Biol.17, 3503–3510. 10.1111/j.1365-2486.2011.02464.x
54
MonahanW. B.KoenigW. D. (2006). Estimating the potential effects of sudden oak death on oak-dependent birds. Biol. Conserv.127, 146–157. 10.1016/j.biocon.2005.08.005
55
MontañezG.ArcilaO.PachecoJ. C.HernándezY.GraciaJ.LancherosH. (1994). Hacia Dónde va la Sabana de Bogotá: Modernización, Conflicto, Ambiente y Sociedad. Bogotá: Universidad Nacional de Colombia. Centro de Estudios Sociales: SENA.
56
MuñozM. C.Fierro-CalderónK.Rivera-GutierrezH. F. (2007). Las aves del campus de la Universidad del Valle, una isla verde urbana en Cali, Colombia. Ornitol. Colomb.5, 5–20.
57
NaranjoL. G. (1995). Patrones de reproducción en dos poblaciones aisladas de Agelaius icterocephalus (Aves:Icteridae). Caldasia18, 89–100.
58
National Audubon Society (2017). Christmas Bird Counts. Available online at: http://www.audubon.org/conservation/science/christmas-bird-count
59
NavaD. E.NascimentoA. M.SteinC. P.HaddadM. L.BentoJ. M. S.ParraJ. (2007). Biology, thermal requirements, and estimation of the number of generations of Zaprionus indianus (Diptera: Drosophilidae) for the main fig producing regions of Brazil. Flor. Entomol.90, 495–501. 10.1653/0015-4040(2007)90[495:BTRAEO]2.0.CO;2
60
NogalesM.VidalE.MedinaF. E.BonnaudE.TershyB. R.CampbellK. J.et al. (2013). Feral cats and biodiversity conservation: the urgent prioritization of island management. Bioscience63, 804–810. 10.1525/bio.2013.63.10.7
61
OlivaresA. (1969). Aves de Cundinamarca.Bogotá: Universidad Nacional de Colombia.
62
Ortega-ÁlvarezR.MacGregor-ForsI. (2011). Dusting-off the file: a review of knowledge on urban ornithology in Latin America. Landsc. Urban Plan.101, 1–10. 10.1016/j.landurbplan.2010.12.020
63
Pérez-ArbeláezE. (1935). Plantas útiles de Colombia. Medellín: Editorial Victor Hugo.
64
PerrinsC. M.GoslerA. G. (2011). Birds, in Wytham Woods: Oxford's Ecological Laboratory, eds SavillP.PerrinsC.KirbyK.FisherN. (New York, NY: Oxford University Press), 145–172.
65
PoundsJ. A.FogdenM. P. L.CampbellJ. H. (1999). Biological response to climate change on a tropical mountain. Nature398, 611–615. 10.1038/19297
66
RaynorG. S. (1975). Techniques for evaluating and analyzing Christmas Bird Count data. Am. Birds29, 626–633.
67
RenjifoL. M.Amaya-VillarrealA. M.Burbano-GirónJ.Velásquez-TibatáJ. (2016). Libro Rojo de Aves de Colombia. Volumen II: Ecosistemas Abiertos, Secos, Insulares, Acuáticos Continentales, Marinos, Tierras Altas del Darién y Sierra Nevada de Santa Marta y Bosques Húmedos del Centro, Norte Y Oriente del País. Bogotá: Editorial Pontificia Universidad Javeriana y Editorial Instituto Alexander von Humboldt.
68
RenjifoL. M.GómezM. F.Velásquez-TibatáJ.Amaya-VillarrealA. M.KattanG. H.Amaya-EspinelJ. D.et al. (2014). Libro rojo de aves de Colombia. Volumen I: Bosques Húmedos de los Andes y la Costa Pacífica. Bogotá: Editorial Pontificia Universidad Javeriana e Instituto Alexander von Humboldt.
69
RojasE.ArceB.PeñaA.BoshellF.AyarzaM. (2010). Cuantificación e interpolación de tendencias locales de temperatura y precipitación en zonas alto andinas de Cundinamarca y Boyacá (Colombia). Corpoica. Cienc. Tecnol. Agrop.11, 173–182.
70
RomeroJ. A. (2010). Transformación urbana de la ciudad de Bogotá, 1990–2010: efecto espacial de la liberalización del comercio. Perspec. Geogr.15, 85–112.
71
RootT. L.PriceJ. T.HallK. R.SchneiderS. H.RosenzweigC.PoundsJ. A. (2003). Fingerprints of global warming on wild animals and plants. Nature421, 57–60. 10.1038/nature01333
72
RosselliL.StilesF. G. (2012a). Wetland habitats of the Sabana de Bogotá Andean Highland Plateau and their birds. Aquat. Conserv. Mar. Freshw. Ecosyst.22, 303–317. 10.1002/aqc.2234
73
RosselliL.StilesF. G. (2012b). Local and landscape environmental factors are important for the conservation of endangered wetland birds in a high andean plateau. Waterbirds35, 453–469. 10.1675/063.035.0310
74
RosselliL.StilesF. G.CamargoP. A. (in press). Changes in the avifauna in a high Andean cloud forest in Colombia over a 24-year period. J. Field Ornithol.
75
SalmiT.AnttilaA. M. P.Ruoho-AirolaT.AmnellT. (2002). Detecting Trends of Annual Values of Atmospheric Pollutants by the Mann-Kendall Test and Sen's Slope Estimates - The Excel Template Application MAKESENS. Publications on air quality N°31. Helsinki: Finnish Meteorological Institute.
76
Secretaría Distrital de Planeación (2016). Región Metropolitana de Bogotá: Una visión de Ocupación del suelo. Colección Integración Regional 11. Alcaldía Mayor de Bogotá.
77
SekerciogluC. H.SchneiderS. H.FayJ. P.LoarieS. R. (2008). Climate change, elevational range shifts, and bird extinctions. Conserv. Biol.22, 140–150. 10.1111/j.1523-1739.2007.00852.x
78
SeressG.LikerA. (2015). Habitat urbanization and its effects on birds. Acta Zool. Acad. Sci. Hungaricae61, 373–408. 10.17109/AZH.61.4.373.2015
79
SoykanC. U.SauerJ.SchuetzJ. G.LeBaronG. S.DaleK.LanghamG. M. (2016). Population trends for North American winter birds based on hierarchical models. Ecosphere7, 1–16. 10.1002/ecs2.1351
80
StattersfieldA. J.CrosbyM. J.LongA. J.WegeD. C. (1998). Endemic Bird Areas of the World. Priorities for Biodiversity Conservation.BirdLife Conservation Series 7. Cambridge: BirdLife International.
81
StilesF. G. (1990). La avifauna de la Universidad de Costa Rica y sus alrededores a través de veinte anaos (1968–1989). Rev. Biol. Trop.38, 361–381.
82
StilesF. G.RosselliL. (1998). Inventario de las aves de un bosque altoandino: comparación de dos métodos. Caldasia20, 29–43.
83
van der HammenT.StilesF. G.RosselliL.ChisacáM. L.CamargoG.GuillotG.et al. (2008). Protocolo de Recuperación y Rehabilitación Ecológica de Humedales en Centros Urbanos. Bogotá: Secretaría Distrital de Ambiente –SDA; Alcaldía Mayor de Bogotá.
84
Vásquez-MuñozJ. L.Castaño-VillaG. J. (2008). Identificación de áreas prioritarias para la conservación de la avifauna en la zona urbana del municipio de Medellín, Colombia. Bol. Cient. Mus. Hist. Nat.12, 51–61.
85
Velásquez-TibatáJ.GutiérrezA.CarrilloE. (2000). Primer registro de parasitismo reproductivo en el Cucarachero de Pantano Cistothorus apolinari por el Chamón Maicero Molothrus bonariensis. Cotinga14, 102–103.
86
Velásquez-TibatáJ.SalamanP.GrahamC. H. (2013). Effects of climate change on species distribution, community structure, and conservation of birds in protected areas in Colombia. Region. Environ. Change13, 235–248. 10.1007/s10113-012-0329-y
87
VillanedaM.RosselliL. (2011). Abundancia de Molothrus bonariensis (Icteridae) en los humedales de la Sabana de Bogotá (Colombia), su asociación con la vegetación y su relación con la presencia de otras especies de interés para conservación. Ornitol. Colomb.11, 37–48.
88
VisserM. E.BothC. (2005). Shifts in phenology due to global climate change: the need for a yardstick. Proc. R. Soc. B272, 2561–2569. 10.1098/rspb.2005.3356
89
WilbyR. L.PerryG. L. W. (2006). Climate change, biodiversity and the urban environment: a critical review based on London, UK. Prog. Phys. Geogr.30, 73–98. 10.1191/0309133306pp470ra
Summary
Keywords
Andes, conservation, Audubon's Christmas Bird Counts, neotropical mountains, global warming, land use change, urban ornithology
Citation
Stiles FG, Rosselli L and De La Zerda S (2017) Changes over 26 Years in the Avifauna of the Bogotá Region, Colombia: Has Climate Change Become Important?. Front. Ecol. Evol. 5:58. doi: 10.3389/fevo.2017.00058
Received
10 March 2017
Accepted
19 May 2017
Published
19 June 2017
Volume
5 - 2017
Edited by
Amanda D. Rodewald, Cornell University, United States
Reviewed by
Nicole Michel, National Audubon Society, United States; Francisco J. Escobedo, Del Rosario University, Colombia
Updates
Copyright
© 2017 Stiles, Rosselli and De La Zerda.
This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Loreta Rosselli lrosselli@udca.edu.co
This article was submitted to Urban Ecology, a section of the journal Frontiers in Ecology and Evolution
Disclaimer
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.