Abstract
Environmental change associated with urbanization is considered one of the major threats to biodiversity. Some species nevertheless seem to thrive in the urban areas, probably associated with selection for phenotypes that match urban habitats. Previous research defined different “copying styles” in distress behavior during the handling of birds. These behaviors vary along a continuum from “proactive” to “reactive” copers. By studying avian distress behaviors we aimed to broaden our understanding of the relationship between coping styles and urbanization. Using a large-scale comparative study of seven paired rural and urban sites across Europe, we assayed distress behaviors during handling of urban and rural-dwelling populations of the great tit Parus major. We detected no consistent pairwise differences in breath rate between urban and rural habitats. However, urban great tits displayed more distress calling (fear screams) and higher pecking rate (handling aggression) than rural birds. These findings suggest that urban great tits have a more proactive coping strategy when dealing with stressful conditions. This finding is in line with previous studies implying that urban great tits are more explorative, less neophobic, and display shorter flight distances than their rural counterparts, representing further aspects of the same “proactive,” coping strategy. Future research should investigate whether reported differences in distress behavior are due to local adaption caused by natural selection or due to phenotypic plasticity.
Introduction
Urban environments are expanding worldwide with an unprecedented speed (United Nations, 2014); environmental change associated with urbanization is being considered as one of the main current threats to biodiversity (Turner et al., 2004). However, urbanization also represents a potential source of selection and a new avenue to study evolutionary processes (Donihue and Lambert, 2014; Isaksson, 2015; Alberti et al., 2017; Hendry et al., 2017).
For wildlife, urban habitats differ in many respects from rural habitats, including microclimate, food abundance, pollution, abundance of exotic species, and predation risk (Luniak and Pisarski, 1982; Marzluff et al., 2001; Gaston, 2010; Gil and Brumm, 2014). Many of these factors induce stress, defined in a broad sense as changes away from an organisms' physiological homeostasis that emerge in response to a change in the environment (sensu Badyaev, 2005; Tuomainen and Candolin, 2012). This is especially the case for factors that represent, or are associated with increased perceived predation risk, such as elevated disturbance due to the presence of humans, pet mammals, and cars (Buchanan and Partecke, 2012). The obvious hypothesis based on this known disturbance or perceived threat is that urban animals would have an enhanced stress response than rural-dwelling animals. However, the results are mixed. For example, some studies reveal higher corticosterone levels (avian stress hormones) in urban compared to rural bird populations (Schoech et al., 2007; Fokidis et al., 2009; Zhang et al., 2011), whereas other studies found the opposite pattern or no significant differences (Partecke et al., 2006; French et al., 2008; Abolins-Abols et al., 2016).
Nevertheless, whatever the direction of the effects, they are likely to influence aspects of behavior, physiology, and life history of urban-dwellers in their coping response to the urban environment (Tuomainen and Candolin, 2012; Sprau and Dingemanse, 2017). Consequently, stress-responses, i.e., the behavioral and physiological means by which animals cope with environmental stress, play an important role in local adaptation (Badyaev, 2005). Hence, comparing stress-responses between urban- and rural-dwelling populations can shed light on the mechanisms underpinning adaptation to urban habitats. For example, predatory attack is most likely one of the highest stresses that a prey animal can experience, and this is irrespective of whether the animal is habituated to the urban environment. Behavioral responses to such threats are commonly referred to as distress behaviors (Møller and Ibáñez-Álamo, 2012). Previous research has shown that distress behaviors displayed during handling are parts of the same phenotypic syndrome that encapsulates an entire suite of behavioral and physiological traits that individual display in order to adaptively respond to stressful situations (Koolhaas et al., 1999, 2010; Carere and van Oers, 2004; Coppens et al., 2010; Carvalho et al., 2013; Class et al., 2014; Kluen et al., 2014). Responses can be categorized into different “coping styles,” varying along a continuum from “proactive” to “reactive” copers, and they relate to the way organisms deal with stressors. Summarizing, proactive individuals are more aggressive, explorative, neophilic, and risk-prone than reactive individuals (Carere et al., 2010).
Accordingly, birds displaying a high frequency of distress behaviors represent proactive individuals. We focus in this paper on three distress behaviors easily recorded during handling: distress calling, handling aggression, and breath rate. Distress calling rate has previously been related to proactivity: in black-capped chickadees Poecile atricapillus, distress calling rate was positively related to exploration rate, which is one of the main characteristics of the proactive coping style (Guillette and Sturdy, 2011). In siskins Carduelis spinus, individuals uttering more often distress calls, displayed also bolder behaviors in front of a novel object (Mateos-González and Senar, 2012; Pascual and Senar, 2014). In the Japanese quail Coturnix japonica, distress calling was positively related to the excretion of corticosterone metabolites, implying that high levels of distress calling are related to proactivity (Niall Daisley et al., 2005). Distress calling has therefore been used as a proxy of proactivity in several studies (Andersen, 2012; Pascual and Senar, 2014; Thorsteinsen, 2015; Richardson et al., 2016). Aggression has also been regarded as a typical response of proactive copers to stressful situations (Koolhaas et al., 1999; Carere et al., 2010), and pecking rate during handling is classically used to compare level of proactivity among individuals (Brommer and Kluen, 2012; van den Brink et al., 2012a,b; Class et al., 2014; Kluen et al., 2014; Dubuc-Messier et al., 2017). Breath rate has also been suggested as an indicator of coping style, with breath rates being higher in proactive individuals than in reactive ones (Carere and van Oers, 2004; van Oers and Carere, 2007; Torné-Noguera et al., 2014; Charmantier et al., 2017). Additionally, breath rate has been found to correlate positively to exploration rate, which as previously stated, is one of the main characteristics of the proactive coping style (Charmantier et al., 2017). The three behaviors have also been found to be heritable (Koenig et al., 1991; Brommer and Kluen, 2012). Therefore, these three distress behaviors can be used as an accurate proxy of coping style and their study may allow a broader understanding of the role of coping style mediating processes of urbanization in birds.
Recent work has found that distress behaviors during handling differ between urban and rural population of birds. For example, an interspecific comparison revealed that urban birds showed higher frequencies of distress calling when handled than their rural counterparts (Møller and Ibáñez-Álamo, 2012). Aggression, another typical response to stressful situations (Koolhaas et al., 1999), has also been found to differ in inter-specific comparisons between urban and rural individuals during handling (Møller and Ibáñez-Álamo, 2012). At the intra-specific level, urban birds have also been shown to display different breath rates compared to their rural counterparts during handling (Torné-Noguera et al., 2014; Abolins-Abols et al., 2016; Charmantier et al., 2017). Yet, this relationship may differ between species. To date, studies comparing distress behaviors between urban and rural bird populations (Møller and Ibáñez-Álamo, 2012; Torné-Noguera et al., 2014) have lacked sufficient replication to draw firm conclusions regarding the relationship between urbanization and avian distress behavior (Evans et al., 2009).
The main aim of the present paper was to investigate whether urban and rural populations of the great tits (Parus major) differ consistently in their behavioral response to an acute perceived threat, assayed during capturing and handling (Møller and Nielsen, 2010; Markó et al., 2013; Class et al., 2014; Kluen et al., 2014; Dubuc-Messier et al., 2017). To acquire a robust insight into the relationship between urbanization and distress behavior, we assayed three different distress behaviors (distress calling rate, pecking rate, and breath rate) in seven replicate pairs of urban and rural/forest populations across Europe. Previous work on this species has revealed many differences between urban and rural great tits in terms of morphology (Horak et al., 1995; Senar et al., 2014; Biard et al., 2017), physiology (Andersson et al., 2015; Salmón et al., 2016; Toledo et al., 2016), genetics (Björklund et al., 2010; Riyahi et al., 2015), life-history (Berressem et al., 1983; Schmidt and Einloft-Achenbach, 1984; Isaksson and Andersson, 2007; Hedblom and Soderstrom, 2012; Bailly et al., 2015; Vaugoyeau et al., 2016; Charmantier et al., 2017), population dynamics (Horak and Lebreton, 1998), and behavior (Slabbekoorn and Peet, 2003; Salaberria and Gil, 2010; Riyahi et al., 2017). Urban great tits have been found to be more explorative and less neophobic than rural tits (Tryjanowski et al., 2016; Charmantier et al., 2017; Riyahi et al., 2017), to display shorter flight initiation distances (Møller et al., 2015), and to show a higher problem solving performance than rural birds (Preiszner et al., 2017). Based on this growing body of literature, which suggests that a proactive coping strategy fits urban conditions, we predict that urban great tit populations show a higher distress calling rate, pecking rate and breath rate compared to their rural conspecifics.
Materials and methods
Study areas and sampling
Great tits were caught in seven distinct cities paired with a close rural site: Granada (Spain), Barcelona (Spain), Montpellier (France), Munich (Germany), Paris (France), Malmö (Sweden), and Tartu (Estonia; Table 1). Paired sites within localities were separated with distances ranging from 5 to 106 km (Table 1). Natural habitats were either forest or rural areas, but we refer to them as rural habitats for simplicity. To quantify the degree of urbanization in each locality we used aerial images from Google Maps following the methods previously described to assess the effect of the degree of urbanization on wild bird populations (Seress et al., 2014; Vincze et al., 2017). Briefly, each locality was represented by a 1 × 1 km2 rectangular area around the birds' capture site. The content in each rectangle was evaluated by dividing the image in 100 × 100 m2 cells and considering three land-cover characteristics in each: proportion of buildings, vegetation (including cultivated fields), and paved surfaces. The different land-cover measures obtained per site fitted into a principal component analysis (PCA) to estimate an urbanization score (PC1) per site (Table 1). The PC1 values were multiplied with −1 to obtain more positive values in the more urbanized localities.
Table 1
| Rural | Urban | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Locality | Period sampled | Latitude | Longitude | N | PC1 | Latitude | Longitude | N | PC1 | Distance (km) |
| Granada | 2015–2016 | 36°56′N | 3°30'W | 19 | −0.99 | 37°12′N | 3°36′W | 10 | 1.29 | 27 |
| Barcelona | 2015–2016 | 41°23′N | 2°10′E | 77 | −2.40 | 41°23′N | 2°09′E | 49 | 2.76 | 5 |
| Montpellier | 2014–2016 | 43°40′N | 3°40′E | 97 | −1.60 | 43°36′N | 3°52′E | 273 | 1.99 | 20 |
| Munich | 2014–2015 | 47° 58′N | 11° 14′E | 442 | −1.55 | 48° 8′N | 11° 34′E | 208 | 3.05 | 31 |
| Paris | 2015–2016 | 48°16′N | 2°41′E | 55 | −2.58 | 48°51′N | 2°20′E | 48 | 2.27 | 80 |
| Malmö | 2015 | 55°40′N | 13°31′E | 37 | −2.56 | 55°36′N | 13°02′E | 41 | 2.03 | 35 |
| Tartu | 2015–2016 | 58°7′N | 25°5′E | 134 | −2.17 | 58°23′N | 26°43′E | 49 | 0.46 | 106 |
Sampling localities of urban and rural great tits (Parus major) across Europe.
We print the name of each site, year of sampling, locations, sample size (n), urbanization index (PC1), and the distance between the rural and the urban site. The lower the urbanization index, the more urbanized the population's habitat; Munich being the most and Tartu the least urbanized habitats among the cities.
Great tits were captured using spring traps, mist nets or funnel traps (Senar et al., 1997). Though trapping methods differed among localities, they were identical in six out of seven urban/rural pair sites (only Granada differed). Sampling was conducted during the main breeding season at each site (April–June), except for the Barcelona population, which was sampled from January to April. Overall, we quantified distress behaviors of 1,539 individual great tits. All individuals represent independent data points as re-captures were not included in our analyses comparing distress behavior between habitats (see below). However, behavioral information of individuals recorded in multiple years was used to estimate repeatability (68 birds for breath rate and 22 for pecking and distress calling rates, see below). All birds were sexed and aged (yearling vs. adult) according to plumage characteristics (Svensson, 1992).
Behavioral tests
We performed measurements of distress behavior within 5 min after capture. We recorded three behaviors known to reflect coping style and stress responsiveness and/or fighting propensity during handling (Carere and van Oers, 2004; Fucikova et al., 2009; Laiolo et al., 2009; Markó et al., 2013; Torné-Noguera et al., 2014). Breath rate, distress call rate, and pecking rate were recorded in the same consecutive order. The target traits were not recorded in all localities: in Montpellier and Munich only breath rate was recorded.
Breath rate was recorded by counting the number of breast respiratory movements within 30 s, while holding the wings fixed and also ensuring reduced disturbance by visual and sound cues (Markó et al., 2013; Torné-Noguera et al., 2014). Pecking rate and distress calling rate were subsequently quantified by counting the number of pecks and distress calls that were emitted during the next 15 s of handling (Fucikova et al., 2009; Laiolo et al., 2009). Pecking rate refers to the number of pecks against a straight finger positioned at 1–2 cm from the beak of the focal bird while holding its legs (Markó et al., 2013). Distress calling rate was defined as the number of vocalized distress calls recorded during the same pecking rate trial (Markó et al., 2013). Breath rate was measured by multiple observers in Montpellier (n = 4) and Munich (n = 14). In the other localities, a single observer recorded all data in both the rural and urban site.
Statistical analyses
To test for general differences in distress behavior between the urban and rural habitats across the seven localities, data were analyzed using univariate linear mixed-effects models (LMM), in which breath rate, pecking rate, or distress calling rate were sequentially included as the response variable. As fixed effects, we included habitat (rural vs. urban), sex and age (yearling vs. adult). Locality (n = 7 levels) and population (n = 2 populations* 7 localities = 14 populations) were included as random effects. LRT refers to a likelihood ratio test. As breath rate was measured by multiple observers in two of the localities (see above), we also included observer as random effect (23 levels, nested within locality) in models explaining variation in breath rate. Since the seven localities represented a wide range of different habitats, a habitat-specific residual variance was fitted to account for unequal residual variances between the two types (rural vs. urban). Breath rate was analyzed assuming a Gaussian error distribution, while pecking rate and distress call were analyzed assuming a Poisson error distribution.
Bivariate LMM were used to estimate among-population and among-individual-within-population covariances (and correlations) between each unique combination of two behaviors, where the two focal behaviors were entered as response variables (following procedures detailed by Dingemanse and Dochtermann, 2013).
Only a priori considered combinations of predictor variables were entered into the statistical models, which was based on biological thinking. This approach to the explicit test of a priori formulated hypotheses has been suggested to allow more general inferences than more traditional exploratory analyses in which various possible interactions are tested sequentially or based on statistical algorithm (Dochtermann, 2010). Accordingly, we were interested in the interaction between habitat and locality, because the difference between urban and rural habitats might differ among localities (see also below). We also included the interaction between habitat and sex, because previous analyses revealed sex differences in the expression of some of the focal behaviors (Markó et al., 2013). We also included an interaction term between habitat and age to test for age-specific differences between urban and rural populations (Markó et al., 2013). In cases where the interaction between sex and habitat was significant, we additionally modeled the sexes separately, in order to understand for which sex the differences between habitats were statistically supported.
We assumed that previous “capture history” did not affect distress behavior, an assumption that was confirmed for the Barcelona site, where the number of preceding captures had no effect on distress behaviors measured during the present study [distress calling rate: F(1, 119) = 1.4, p = 0.32; pecking rate: F(1, 119) = 0.6, p = 0.52; breath rate: F(1, 119) = 0.2, p = 0.73]. Furthermore, we can infer that “capture history” should not be an important confounder, as the data used in our analyses were largely based on first captures.
Adjusted repeatability was calculated following Nakagawa and Schielzeth (2010) using series of univariate LMMs where individual identity and population identity were fitted as a random effects, and where sex, age, and habitat as fixed effects. We repeated these analyses for rural and urban birds separately to calculate repeatability within each habitat type (urban vs. rural).
All statistical analyses were carried out using the package lme4, version 1.1-12 (Bates et al., 2011) and MCMCglmm version 2.23 (Hadfield, 2010) within the R (version 3.3.1) computing environment (R Development Core Team, 2017). For the modeling based on MCMCglmm, we ran Markov chains up to 230,000 iterations with 30,000 iterations of burn in and with 500 iterations of thinning interval (longer runs, such as 1,300,000 iterations, did not improve the results). For prior definition, we followed a technique called parameter expansion as was suggested by Hadfield (2010), and set V = 1, nu = 0.02, and alpha.V = 1,000. We repeated each run for each model 3–4 times to check the stability of results. After each run, the trace and distribution of all estimated parameters were checked visually, as well as autocorrelation between iterations. Furthermore, mixing and convergence were checked with Gelman-Rubin statistics (Gelman and Rubin, 1992).
Results
The selected urban areas had significantly higher urbanization scores than the selected rural areas [F(1, 13) = 96.08, p < 0.0001] based on the satellite data, confirming that our assignments were appropriate.
Distress calling rate showed a low and non-significant between-year repeatability within individuals (Table 2). However, repeatabilities of pecking and breath rates were of higher magnitude, and were similar between rural and urban populations (Table 2). Distress calling rate and pecking rate were positively correlated within populations (Table 3) but distress calling and pecking rates did not correlate significantly with breath rate (Table 3), suggesting that breath rate was independent from the two other traits. None of the between-population correlations were statistically significant (Table 3).
Table 2
| Individual repeatability | |||||
|---|---|---|---|---|---|
| r | SE | −95%CI | +95%CI | p | |
| DISTRESS CALLING RATE | |||||
| Unadjusted repeatability | |||||
| All birds | 0.02 | 0.08 | 0 | 0.28 | 0.27 |
| Adjusted repeatability | |||||
| All birds | 0.02 | 0.09 | 0 | 0.32 | 0.26 |
| PECKING RATE | |||||
| Unadjusted repeatability | |||||
| All birds | 0.59 | 0.18 | 0.13 | 0.84 | <0.001 |
| Rural birds | 0.59 | 0.3 | 0 | 0.97 | 0.09 |
| Urban birds | 0.54 | 0.23 | 0 | 83 | 0.04 |
| Adjusted repeatability | |||||
| All birds | 0.61 | 0.19 | 0.15 | 0.87 | <0.001 |
| Rural birds | 0.38 | 0.32 | 0 | 0.91 | 0.04 |
| Urban birds | 0.55 | 0.22 | 0 | 0.84 | 0.04 |
| BREATH RATE | |||||
| Unadjusted repeatability | |||||
| All birds | 0.48 | 0.10 | 0.27 | 0.65 | <0.001 |
| Rural birds | 0.43 | 0.2 | 0 | 0.76 | 0.01 |
| Urban birds | 0.49 | 0.11 | 0.25 | 0.68 | <0.001 |
| Adjusted repeatability | |||||
| All birds | 0.35 | 0.10 | 0.14 | 0.54 | <0.001 |
| Rural birds | 0.39 | 0.21 | 0 | 0.74 | 0.08 |
| Urban birds | 0.37 | 0.12 | 0.13 | 0.59 | <0.001 |
Within-individual and between-year repeatability of distress calling, pecking, and breath rates.
Univariate linear mixed-effects models were performed fitting bird identity as a random effect. First, we present models where effects of sex, age, habitat, locality, and population, were not included (i.e., estimates represent the unadjusted repeatability). Second, we present models where the mentioned effects were controlled for (i.e., estimates represent adjusted repeatability; sensu Nakagawa and Schielzeth, 2010). We also present repeatability estimates separately for the urban and rural habitat (though for distress calling rate, variance and the sample size was too small to analyze the two habitats separately). Repeatability (r), standard error (SE), lower and upper confidence interval (CI), and significance (p-value) is provided. Sample size for breath rate: nrural = 32, nurban = 104 individuals measured twice in different years, for pecking and distress calling rates: nrural = 12, nurban = 32 individuals measured twice in different years.
Table 3
| Correlations | ||||||
|---|---|---|---|---|---|---|
| Among-populations | Within-populations | |||||
| r | −95%CI | +95%CI | r | −95%CI | +95%CI | |
| Distress calling vs. Pecking rate | 0.54 | −0.07 | 0.85 | 0.58 | 0.49 | 0.68 |
| Distress calling vs. Breath rate | 0.14 | −0.41 | 0.69 | −0.01 | −0.13 | 0.03 |
| Pecking rate vs. Breath rate | 0.01 | −0.63 | 0.65 | 0.04 | −0.06 | 0.16 |
Correlations coefficients at different levels among distress calling, pecking, and breath rates.
We print the correlation coefficient (r) and the credible intervals (CI) between-population (the correlation of population means of the individual traits across different populations) and within-population (the correlation of individual-specific measurements within populations). When the upper and lower CI's does not cross over zero the correlation can be considered as significant (high-lighted in bold face).
Distress calling rate was higher in birds from urban compared to rural habitats (Figure 1, Table 4). There was a non-significant trend toward an interaction between sex and habitat on distress calling rate (Table 4). Though the difference in distress calling rate between urban and rural areas tended to be stronger in females compared to males, urban great tits of both sexes had higher distress calling rates compared to their rural conspecifics. The interaction between locality and habitat could not be tested because models incorporating this fixed effect did not converge.
Figure 1
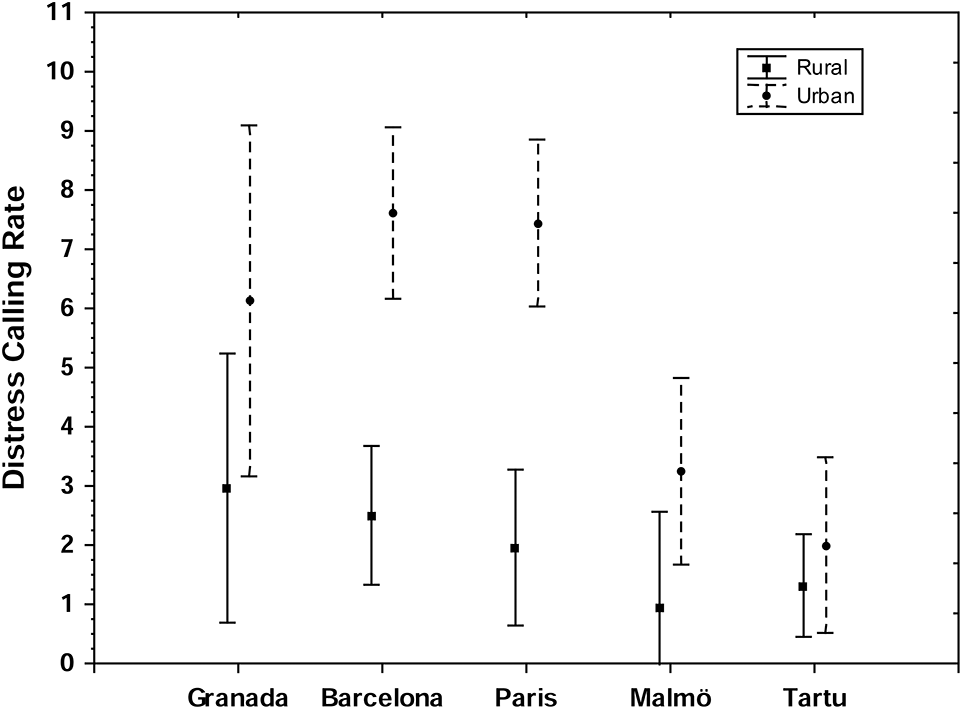
Between-individual variation (mean with standard error) in distress calling rate along habitats (urban vs. rural) and localities (see Table 4; both sexes combined). Distress calling rate is number of calls during 15 s.
Table 4
| LRT | p | ß | −95% CI | +95% CI | |
|---|---|---|---|---|---|
| FIXED EFFECTS | |||||
| (Intercept) | −1.94 | −3.13 | 0.80 | ||
| Sex [male] | 3.46 | 0.06 | 0.92 | 0.18 | 1.67 |
| Age [yearling] | 2.54 | 0.11 | −0.44 | −1.20 | 0.33 |
| Habitat [urban] | 15.44 | <0.001 | 2.72 | 1.25 | 4.04 |
| Habitat*Sex [urban, male] | 3.44 | 0.06 | −1.08 | −2.05 | 0.10 |
| Habitat*Age [urban, yearling] | 0.56 | 0.46 | −0.06 | −1.05 | 0.95 |
| RANDOM EFFECTS | |||||
| Variance | |||||
| Locality | <0.001 | 0.99 | <0.001 | 0.00 | 2.01 |
| Population | 3.19 | 0.07 | 0.01 | 0.00 | 1.69 |
| Error | 4.40 | 2.09 | 8.60 | ||
Statistical summary of the analysis of distress calling rate between rural and urban habitats.
The table shows the results of LMMs, in which the significance of terms was evaluated by likelihood-ratio test (LRT). Means of the parameter estimates (ß), variance components and their respective 95%CI intervals were obtained from the posterior distribution of the corresponding MCMCglmm. In square bracket we indicate to which factor level the parameter estimate corresponds.
In the full model for pecking rate, there was no significant effect of habitat, age or their interaction (habitat: LRT = 2.75, p = 0.10; age LRT = 0.00, p = 0.98; habitat × age: LRT = 0.29, p = 0.59). Both locality and population were significantly related with great tit pecking rate (locality: LRT = 7.24, p = 0.01; population: LRT = 2.41, p < 0.01). In addition, males displayed higher pecking rates than females (sex: LRT = 4.63, p = 0.03). However, we also found a non-significant trend for an interaction between sex and habitat on pecking rate (sex × habitat: LRT = 3.34, p = 0.07). A post-hoc analysis separating the sexes, revealed that there was an effect of habitat on pecking rate in males (LRT = 4.79, p = 0.03, Figure 2, Table 5) but not in females (LRT = 0.01, p = 0.95). The interaction between locality and habitat was non-significant (LRT: 0.34, p = 0.95) in the model for males indicating that the higher pecking rate of urban than rural males was consistent across localities (Figure 2).
Figure 2
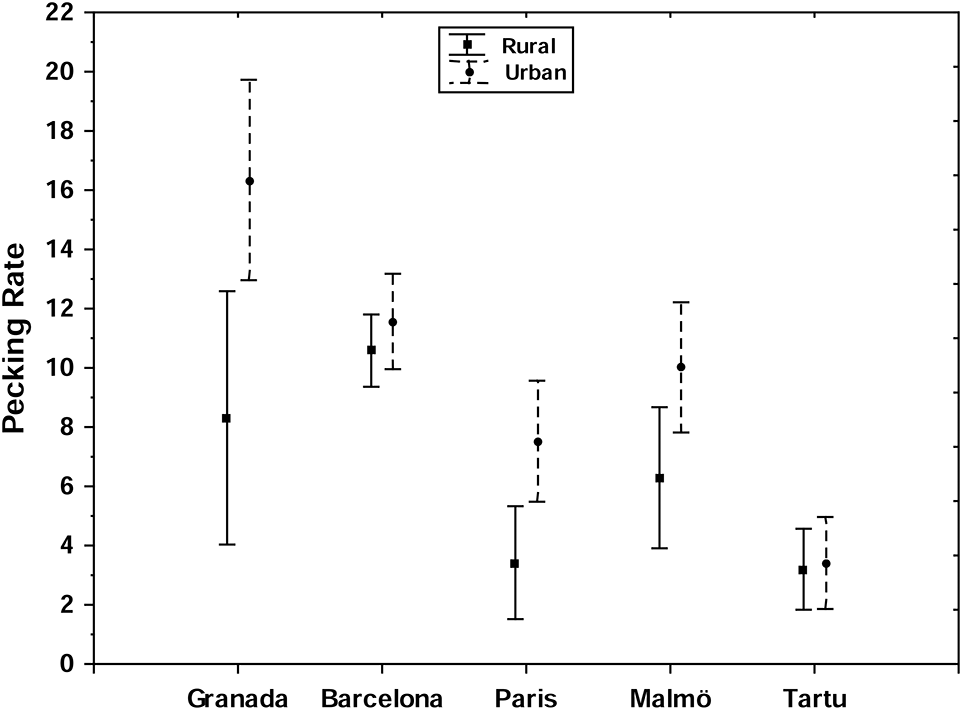
Between-individual variation (mean with standard error) in pecking rate along habitats (urban vs. rural) and localities (see Table 5). Pecking rate is number of peckings during 15 s.
Table 5
| LRT | p | ß | −95% CI | +95% CI | |
|---|---|---|---|---|---|
| FIXED EFFECTS | |||||
| (Intercept) | 1.55 | 0.64 | 2.61 | ||
| Age [yearling] | 0.76 | 0.38 | −0.07 | 0.21 | 0.32 |
| Habitat [urban] | 4.79 | 0.03 | −0.46 | 0.02 | 1.09 |
| Habitat*Age [urban, yearling] | 0.11 | 0.75 | −0.02 | −0.38 | 0.35 |
| RANDOM EFFECTS | |||||
| Variance | |||||
| Locality | 4.14 | 0.04 | 0.04 | 0.00 | 3.59 |
| Population | 4.78 | 0.03 | 0.01 | 0.00 | 0.52 |
| Error | 0.32 | 0.13 | 0.57 | ||
Statistical summary of the analysis of pecking rate for male great tits between rural and urban habitats.
The table shows the results of LMMs, in which the significance of terms was evaluated by likelihood-ratio test (LRT). Means of the parameter estimates (ß), variance components, and their respective 95%CI intervals were obtained from the posterior distribution of the corresponding MCMCglmm. In square bracket we indicate to which factor level the parameter estimate corresponds.
Breath rate did not differ between urban and rural habitats when controlling for sex, age, locality, population, and observer effects (Table 6). The interaction between locality and habitat was also non-significant (LRT < 0.001, p = 0.99), indicating that this lack of difference was consistent across localities.
Table 6
| LRT | p | ß | −95% CI | +95% CI | |
|---|---|---|---|---|---|
| FIXED EFFECTS | |||||
| (Intercept) | 62.64 | 60.35 | 65.89 | ||
| Sex [male] | 0.649 | 0.42 | −0.16 | −1.51 | 1.38 |
| Age [yearling] | 1.186 | 0.28 | 0.83 | −0.76 | 2.49 |
| Habitat [urban] | 0.432 | 0.51 | 1.25 | −2.44 | 4.10 |
| Habitat*Sex [urban, male] | 0.696 | 0.40 | −0.71 | −2.62 | 1.37 |
| Habitat*Age [urban, yearling] | 0.353 | 0.55 | −0.53 | −2.31 | 2.22 |
| RANDOM EFFECTS | |||||
| Variance | |||||
| Locality | 0.001 | 1.00 | <0.001 | 0.00 | 19.10 |
| Observer | 0.321 | 0.57 | 2.954 | 0.00 | 10.92 |
| Population | 16.500 | <0.001 | 7.632 | 2.35 | 15.68 |
| Error | 84.28 | 76.63 | 95.40 | ||
Statistical summary of the analysis of breath rate between rural and urban habitats.
The table shows the results of LMMs, in which the significance of terms was evaluated by likelihood-ratio test (LRT). Means of the parameter estimates (ß), variance components and their respective 95%CI intervals were obtained from the posterior distribution of the corresponding MCMCglmm. In square bracket we indicate to which factor level the parameter estimate corresponds.
Discussion
The present study reveals consistent differences in distress calling rate between urban and rural populations of great tits, across multiple European localities, with urban birds having, in general, a higher distress calling rate than their rural conspecifics. In line with this finding, pecking rate showed similar tendencies, but only for males. By contrast, we did not find a general difference in breath rate between urban and rural birds.
Distress behaviors displayed during handling have been hypothesized to reflect the coping style of the different individuals in the presence of an acute stressful situation (Brommer and Kluen, 2012; Class et al., 2014; Kluen et al., 2014; Class and Brommer, 2016). Among the three measured distress behaviors, only distress calling rate showed a clear habitat effect. The higher distress calling rate of urban great tits compared to their rural counterparts is consistent with findings of previous studies that compared bird species (Møller and Ibáñez-Álamo, 2012). Given that high distress calling rates have been related to a proactive coping style (Guillette and Sturdy, 2011; Pascual and Senar, 2014), our results support the view that urban birds are more proactive than rural birds.
Because distress calling and pecking rates were correlated, we expected similar habitat differences for the two traits. Indeed, pecking rate was related to urbanization, but only for males. This sexual difference has previously been observed (Markó et al., 2013), and is congruent with the general observation that males are more aggressive than females. Under this general view, the higher aggression of urban male great tits could be boosted by intense intra-specific competition for a limited number of territories and a potentially higher breeding density in urban areas (Chamberlain et al., 2009). In line with this finding, elevated levels of male territorial aggression have been found in urban song sparrows Melospiza melodia, compared to rural male conspecifics. However, this pattern is not necessarily supported in other species (Newman et al., 2006; Bókony et al., 2010; Atwell et al., 2014; Hasegawa et al., 2014). Furthermore, bird species living in urban landscapes have been shown to peck in lower frequencies in response to handling than those inhabiting rural habitats (Møller and Ibáñez-Álamo, 2012). Yet, Møller and Ibáñez-Álamo (2012) did not discriminate between males and females, which hampers a straightforward comparison with our findings. Taken together, these inconsistent results do not allow assessing whether pecking rate is more related to aggression and the latter trait that differ between habitat types, or if it is per se a distress behavior that is subjected to selection. In any case, both aggression and behavior upon handling have been regarded as typical indicators of the same proactive coping style (Koolhaas et al., 1999; Carere et al., 2010; Class et al., 2014; Kluen et al., 2014; Dubuc-Messier et al., 2017). Accordingly, our data on great tit pecking rate suggests that urban males are more proactive than rural males. The interaction between habitat and locality was not significant, indicating that the pecking rate differences among habitats was consistent across localities.
In songbirds, breath rate has been proposed as an indicator of acute stress resulting from a predatory attack, and more generally, as an indicator of stress sensitivity and coping style (Carere and van Oers, 2004; van Oers and Carere, 2007; Torné-Noguera et al., 2014). Indeed, some studies found higher breath rates in urban great tits compared to their rural counterparts (Torné-Noguera et al., 2014; Charmantier et al., 2017). Our study suggests that this finding does not represent a general pattern. In other species, such as dark-eyed juncos Junco hyemalis, urban birds even had a lower breath rate than their rural conspecifics (Abolins-Abols et al., 2016). Our study further reveals that breath rate did not correlate with any of the other behaviors in great tits. This is surprising, since a positive association between pecking rate and breath rate has previously been documented in the closely related blue tit Cyanistes caeruleus (Brommer and Kluen, 2012). However, breath rate has been shown to be affected by seasonal, annual and other ecological factors, hence it may be especially prone to micro-climatic and habitat variation which may mask links to both general urbanization and other behaviors (Torné-Noguera et al., 2014; Charmantier et al., 2017).
The relatively low repeatability of distress calling, potentially due to the low number of repeats per individual (Dingemanse and Dochtermann, 2013; Markó et al., 2013), points to a conservative measure of differences among habitats. Despite the low repeatability, there existed strong differences in distress calling among rural and urban individuals, what points to important differences among habitats, suggesting that distress calling may have evolved in response to urban-induced selection pressures. The between-year repeatability estimates for pecking and breath rate that we recorded, were generally higher than repeatabilities for other behaviors reported in literature (Bell et al., 2009). Given that male pecking rate was different across the habitats and that the repeatability was high, this particular trait seems part of the “urban behavioral phenotype.” This finding is also supported by a recent study indicating a non-random distribution of behavioral types along urban gradients, but little behavioral plasticity in response to within-individual variation to urbanization (Sprau and Dingemanse, 2017).
The combined datasets from different European projects across the distribution of great tits enabled us to conduct a cross-population comparison over a large geographic scale. It entailed study-specific differences in capture time, trapping methods, and observer effects. Nevertheless, we were able to detect consistent differences between habitats, implying that effects reported should be conservative. Altogether, our data suggest that urban great tits, especially males, have a more proactive coping strategy when dealing with stressful conditions than rural birds. This finding is in line with studies showing that urban great tits are more explorative and less neophobic than their rural counterparts (Tryjanowski et al., 2016; Charmantier et al., 2017; Riyahi et al., 2017), and display shorter flight initiation distances (Møller, 2008, 2012), which represent other aspects of the proactive coping strategy. Findings on the great tit are also in line with data from other bird species, for which urban populations have also been found to display proactive behaviors (Evans et al., 2010; Carrete and Tella, 2017), but these studies compared few populations (Miranda et al., 2013). Our study included 14 populations from a large geographic area and revealed consistent results across localities, allowing to generalize the impact of urbanization on great tit coping styles.
This proactive strategy of urban great tits could be advantageous in the urban environment. Proactive individuals, for instance, more rapidly discover and use new food resources (van Overveld and Matthysen, 2010), and are less afraid of novel objects or environments than reactive birds (Tryjanowski et al., 2016; Charmantier et al., 2017; Riyahi et al., 2017), which could help urban dwellers to take profit of the new feeding opportunities that a city can provide (e.g., Fisher and Hinde, 1949). Our study therefore implies that urban birds may adapt to human disturbance by a process of local adaptation, as suggested previously (Partecke et al., 2006; Sol et al., 2013). Future research should investigate whether the differences in distress behaviors reported here have an underlying genetic base, hence are due to local adaption caused by divergent natural selection, or originate from phenotypic plasticity.
Statements
Ethics statement
Behavioral data was recorded during ringing operations. Capture and ringing was performed under individual ringing permits delivered by the ringing office within each country.
Author contributions
The study was conceived by JCS, LG, VT, CB, GM-R, and CI. The data were collected by VT, CB, GM-R, PaS, JR, PhS, ND, AC, VD, and HN. The statistical analyses were conducted by JCS and LG. The manuscript was written by JCS and LG with input from all other co-authors. Contributed funding and materials: JCS, VT, CB, GM-R, PhS, ND, AC, CI.
Funding
This work was supported by funds from the Ministry of Economy and Competitivity, Spanish Research Council (CGL-2016-79568-C3-3-P; to JCS. CGL2015-70639-P; to LG. CGL2014-55969-P; to GM-R), from the National Research, Development and Innovation Office (Hungary) (K-115970; to LG), from the Estonian Ministry of Education and Research (institutional research funding IUT no. 34-8; to VT), from the European Research Council (ERC-2013-StG-337365-SHE; to AC), from the OSU-OREME (to AC), from the Deutsche Forschungsgemeinschaft (DFG) (SP 1450/3-1; to PhS), from the European Union a Marie Curie Re-Integration grant (CIG 322217; to CI) and from the Swedish Research council (C0361301; to CI).
Acknowledgments
We thank the Institute of Parks and Gardens of Barcelona (Parcs i Jardins), the mairie de Montpellier and the cities of Paris and Rueil-Malmaison, for allowing us to sample birds in their city parks. We also thank the National Forest Office (ONF) for allowing us to survey tit populations breeding in state-owned forest, the late Leopoldo Gil, who always supported our work, for allowing us to sample birds in Can Catà forest area, and Hermanitas de la Asunción for allowing us to sample birds in the Sarria area. We also thank Lluïsa Arroyo, Arnaud Grégoire, Marcel Lambrechts, Mar Comas, Nuria Mora, Abelardo Requena, Alexia Mouchet, Maria Moiron, Robin Abbey-Lee, Natalia Perez Ruiz, Mattia Bessone, Rui Machado, Lucila Fernandez, Alexander Hutfluss, Diogo Barros, Gülperi Stenhouse, Josefa Bleu, Simon Agostini, Beatriz Decencière, Erika Beaugeard, Jeanne Dupuy, Eva Du Tien Hat, Laura Grosvalet, Lucie Mathieu, Juliette Rabdeau, Marine Ramirez, Baptiste Vancostenoble, Antonin Waterschoot, Audrey Fournier, Estelle Garrouste, Marilou Keck, Alice Le Priol, Cécilia Mondet, Amélie Pinel, and Quentin Pionneaud for help during field work. Anders P. Møller also provided useful advice during the early stages of the work.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
1
Abolins-AbolsM.HopeS. F.KettersonE. D. (2016). Effect of acute stressor on reproductive behavior differs between urban and rural birds. Ecol. Evol.6, 6546–6555. 10.1002/ece3.2347
2
AlbertiM.MarzluffJ.HuntV. M. (2017). Urban driven phenotypic changes: empirical observations and theoretical implications for eco-evolutionary feedback. Philos. Trans. R. Soc. B Biol. Sci.372:20160029. 10.1098/rstb.2016.0029
3
AndersenM. (2012). What is Maintaining Variation in Personalities in Wild Populations of Great Tits and Blue Tits? Master thesis, University of Oslo, Oslo.
4
AnderssonM. N.WangH. L.NordA.SalmonP.IsakssonC. (2015). Composition of physiologically important fatty acids in great tits differs between urban and rural populations on a seasonal basis. Front. Ecol. Evol.3:93. 10.3389/fevo.2015.00093
5
AtwellJ. W.CardosoG. C.WhittakerD. J.PriceT. D.KettersonE. D. (2014). Hormonal, behavioral, and life-history traits exhibit correlated shifts in relation to population establishment in a novel environment. Am. Nat. 184, E147–E160. 10.1086/678398
6
BadyaevA. V. (2005). Stress-induced variation in evolution: from behavioural plasticity to genetic assimilation. Proc. Biol. Sci.272, 877–886. 10.1098/rspb.2004.3045
7
BaillyJ.ScheiflerR.BertheS.Clément-DemangeV. A.LeblondM.PasteurB.et al. (2015). From eggs to fledging: negative impact of urban habitat on reproduction in two tit species. J. Ornithol.157, 377–392. 10.1007/s10336-015-1293-3
8
BatesD.MaechlerM.BolkerB. (2011). lme4: Linear Mixed-Effects Models Using S4 Classes. Available online at: http://CRAN.R-project.org/package=lme4
9
BellA. M.HankisonS. J.LaskowskiK. L. (2009). The repeatability of behaviour. A meta-analysis. Anim. Behav.77, 771–783. 10.1016/j.anbehav.2008.12.022
10
BerressemK. G.BerressemH.SchmidtK. H. (1983). Vergleich der Brutbiologie von Höhlenbrütern in innerstädtischen und stadtfernen Biotopen. J. Ornithol.124, 431–445. 10.1007/BF01640362
11
BiardC.BrischouxF.MeillèreA.MichaudB.NivièreM.RuaultS.et al. (2017). Growing in cities: an urban penalty for wild birds? A study of phenotypic differences between urban and rural great tit chicks (Parus major). Front. Ecol. Evol.5:79. 10.3389/fevo.2017.00079
12
BjörklundM.RuizI.SenarJ. C. (2010). Genetic differentiation in the urban habitat: the great tits (Parus major) of the parks of Barcelona city. Biol. J. Linn. Soc. 99, 9–19. 10.1111/j.1095-8312.2009.01335.x
13
BókonyV.KulcsárA.LikerA. (2010). Does urbanization select for weak competitors in house sparrows?Oikos119, 437–444. 10.1111/j.1600-0706.2009.17848.x
14
BrommerJ. E.KluenE. (2012). Exploring the genetics of nestling personality traits in a wild passerine bird: testing the phenotypic gambit. Ecol. Evol.2, 3032–3044. 10.1002/ece3.412
15
BuchananK. L.ParteckeJ. (2012). The endocrine system: can homeostasis be maintained in a changing world?, in Behavioural Responses to a Changing World: Mechanisms and Consequences, eds CandolinU.WongB. B. M. (Oxford: Oxford University Press), 32–45.
16
CarereC.CaramaschiD.FawcettT. W. (2010). Covariation between personalities and individual differences in coping with stress. Converging evidence and hypotheses. Curr. Zool.56, 728–740.
17
CarereC.van OersK. (2004). Shy and bold great tits (Parus major): body temperature and breath rate in response to handling stress. Physiol. Behav.82, 905–912. 10.1016/S0031-9384(04)00312-9
18
CarreteM.TellaJ. L. (2017). Behavioral correlations associated with fear of humans differ between rural and urban burrowing owls. Front. Ecol. Evol. 5:54. 10.3389/fevo.2017.00054
19
CarvalhoC. F.LeitaoA. V.FunghiC.BatalhaH. R.ReisS.MotaP. G.et al. (2013). Personality traits are related to ecology across a biological invasion. Behav. Ecol. 24, 1081–1091. 10.1093/beheco/art034
20
ChamberlainD. E.CannonA. R.TomsM. P.LeechD. I.HatchwellB. J.GastonK. J. (2009). Avian productivity in urban landscapes: a review and meta-analysis. Ibis151, 1–18. 10.1111/j.1474-919X.2008.00899.x
21
CharmantierA.DemeyrierV.LambrechtsM.PerretS.GrégoireA. (2017). Urbanization is associated with divergence in pace-of-life in great tits. Front. Ecol. Evol.5:53. 10.3389/fevo.2017.00053
22
ClassB.BrommerJ. E. (2016). Senescence of personality in a wild bird. Behav. Ecol. Sociobiol.70, 733–744. 10.1007/s00265-016-2096-0
23
ClassB.KluenE.BrommerJ. E. (2014). Evolutionary quantitative genetics of behavioral responses to handling in a wild passerine. Ecol. Evol.4, 427–440. 10.1002/ece3.945
24
CoppensC. M.BoerS. F.de KoolhaasJ. M. (2010). Coping styles and behavioural flexibility. Towards underlying mechanisms. Philos. Trans. R. Soc. B Biol. Sci.365, 4021–4028. 10.1098/rstb.2010.0217
25
DingemanseN. J.DochtermannN. A. (2013). Quantifying individual variation in behaviour: mixed-effect modelling approaches. J. Anim. Ecol. 82, 39–54. 10.1111/1365-2656.12013
26
DochtermannN. A. (2010). Behavioral syndromes: carryover effects, false discovery rates, and a priori hypotheses. Behav. Ecol.21, 437–439. 10.1093/beheco/arq021
27
DonihueC. M.LambertM. R. (2014). Adaptive evolution in urban ecosystems. Ambio44, 194–203. 10.1007/s13280-014-0547-2
28
Dubuc-MessierG.RéaleD.PerretP.CharmantierA. (2017). Environmental heterogeneity and population differences in blue tits personality traits. Behav. Ecol. 28, 448–445. 10.1093/beheco/arw148
29
EvansJ.BoudreauK.HymanJ. (2010). Behavioural syndromes in urban and rural populations of song sparrows. Ethology116, 588–595. 10.1111/j.1439-0310.2010.01771.x
30
EvansK. L.GastonK. J.SharpS. P.McGowanA.HatchwellB. J. (2009). The effect of urbanisation on avian morphology and latitudinal gradients in body size. Oikos118, 251–259. 10.1111/j.1600-0706.2008.17092.x
31
FisherJ.HindeR. A. (1949). The opening of milk bottles by birds. Br. Birds42, 347–357.
32
FokidisH. B.OrchinikM.DevicheP. (2009). Corticosterone and corticosteroid binding globulin in birds: relation to urbanization in a desert city. Gen. Comp. Endocrinol.160, 259–270. 10.1016/j.ygcen.2008.12.005
33
FrenchS. S.FokidisH. B.MooreM. C. (2008). Variation in stress and innate immunity in the tree lizard (Urosaurus ornatus) across an urban-rural gradient. J. Comp. Physiol. B Biochem. Syst. Environ. Physiol.178, 997–1005. 10.1007/s00360-008-0290-8
34
FucikovaE.DrentP. J.SmitsN.van OersK. (2009). Handling stress as a measurement of personality in great tit nestlings (Parus major). Ethology115, 366–374. 10.1111/j.1439-0310.2009.01618.x
35
GastonK. J. (2010). Ecological Reviews: Urban Ecology. Cambridge: Cambridge University Press.
36
GelmanA.RubinD. B. (1992). Inference from iterative simulation using multiple sequences. Stat. Sci.7, 457–472. 10.1214/ss/1177011136
37
GilD.BrummH. (eds.). (2014). Avian Urban Ecology. Behavioural and Physiological Adaptations, 1st Edn. Oxford: Oxford University Press.
38
GuilletteL. M.SturdyC. B. (2011). Individual differences and repeatability in vocal production: stress-induced calling exposes a songbird's personality. Naturwissenschaften98, 977–981. 10.1007/s00114-011-0842-8
39
HadfieldJ. D. (2010). MCMC methods for multi-response generalized linear mixed models: the MCMCglmm R package. J. Stat. Softw.33, 1–20. 10.18637/jss.v033.i02
40
HasegawaM.LigonR. A.GiraudeauM.WatanabeM.McGrawK. J. (2014). Urban and colorful male house finches are less aggressive. Behav. Ecol.25, 641–649. 10.1093/beheco/aru034
41
HedblomM.SoderstromB. (2012). Effects of urban matrix on reproductive performance of Great Tit (Parus major) in urban woodlands. Urban Ecosyst.15, 167–180. 10.1007/s11252-011-0204-5
42
HendryA. P.GotandaK. M.SvenssonE. I. (2017). Human influences on evolution, and the ecological and societal consequences. Philos. Trans. R. Soc. B Biol. Sci.372, 1712. 10.1098/rstb.2016.0028
43
HorakP.LebretonJ. D. (1998). Survival of adult Great tits Parus major in relation to sex and habitat; a comparision of urban and rural populations. Ibis140, 205–209. 10.1111/j.1474-919X.1998.tb04380.x
44
HorakP.MandR.OtsI.LeivitsA. (1995). Egg size in the great tit Parus major: individual, habitat and geographic differences. Ornis Fennica72, 97–114.
45
IsakssonC. (2015). Urbanization, oxidative stress and inflammation: a question of evolving, acclimatizing or coping with urban environmental stress. Funct. Ecol. 29, 913–923. 10.1111/1365-2435.12477
46
IsakssonC.AnderssonS. (2007). Carotenoid diet and nestling provisioning in urban and rural great tits Parus major. J. Avian Biol. 38, 564–572. 10.1111/j.0908-8857.2007.04030.x
47
KluenE.SiitariH.BrommerJ. E. (2014). Testing for between individual correlations of personality and physiological traits in a wild bird. Behav. Ecol. Sociobiol.68, 205–213. 10.1007/s00265-013-1635-1
48
KoenigW. D.StanbackM. T.HoogeP. N.MummeR. L. (1991). Distress calls in the acorn woodpecker. Condor93, 637–643. 10.2307/1368195
49
KoolhaasJ. M.BoerS. F.de CoppensC. M.BuwaldaB. (2010). Neuroendocrinology of coping styles. Towards understanding the biology of individual variation. Front. Neuroendocrinol.31, 307–321. 10.1016/j.yfrne.2010.04.001
50
KoolhaasJ. M.KorteS. M.De BoerS. F.Van Der VegtB. J.Van ReenenC. G.HopsterH.et al. (1999). Coping styles in animals: current status in behavior and stress-physiology. Neurosci. Biobehav. Rev.23, 925–935. 10.1016/S0149-7634(99)00026-3
51
LaioloP.BandaE.LemusJ. A.AguirreJ. I.BlancoG. (2009). Behaviour and stress response during capture and handling of the red-billed chough Pyrrhocorax pyrrhocorax (Aves: Corvidae). Biol. J. Linn. Soc.96, 846–855. 10.1111/j.1095-8312.2008.01174.x
52
LuniakM.PisarskiB. (1982). Animals in Urban Environment. Wroclaw: Polish Academy Sciences, Inst. Zoology.
53
MarkóG.AzcárateM.HegyiG.HercegG.LacziM.NagyG.et al. (2013). Behavioural responses to handling stress in the Great Tit: within-individual consistency and the effect of age, sex and body condition. Ornis Hungarica21, 12–25. 10.2478/orhu-2013-0012
54
MarzluffJ. M.DonnellyR. E.BowmanR. (eds.). (2001). Avian Ecology and Conservation in An Urbanizing World.New York, NY: Kluwer Academic Publishers.
55
Mateos-GonzálezF.SenarJ. C. (2012). Melanin-based trait predicts individual exploratory behaviour in siskins, Carduelis spinus. Anim. Behav. 83, 229–232. 10.1016/j.anbehav.2011.10.030
56
MirandaA. C.SchielzethH.SonntagT.ParteckeJ. (2013). Urbanization and its effects on personality traits: a result of microevolution or phenotypic plasticity?Glob. Change Biol.19, 2634–2644. 10.1111/gcb.12258
57
MøllerA. P. (2008). Flight distance of urban birds, predation, and selection for urban life. Behav. Ecol. Sociobiol. 63, 63–75. 10.1007/s00265-008-0636-y
58
MøllerA. P. (2012). Reproductive behaviour, in Behavioural Responses to a Changing World: Mechanisms and Consequences, eds CandolinU.WongB. B. M. (Oxford: Oxford University Press), 106–118.
59
MøllerA. P.Ibáñez-ÁlamoJ. D. (2012). Escape behaviour of birds provides evidence of predation being involved in urbanization. Anim. Behav. 84, 341–348. 10.1016/j.anbehav.2012.04.030
60
MøllerA. P.NielsenJ. T. (2010). Fear screams and adaptation to avoid imminent death: effects of genetic variation and predation. Ethol. Ecol. Evol. 22, 183–202. 10.1080/03949371003707968
61
MøllerA. P.TryjanowskiP.DíazM.KwiecinskiZ.IndykiewiczP.MitrusC.et al. (2015). Urban habitats and feeders both contribute to flight initiation distance reduction in birds. Behav. Ecol. 26, 861–865. 10.1093/beheco/arv024
62
NakagawaS.SchielzethH. (2010). Repeatability for Gaussian and non-Gaussian data: a practical guide for biologists. Biol. Rev. Camb. Philos. Soc.85, 935–956. 10.1111/j.1469-185X.2010.00141.x
63
NewmanM. M.YehP. J.PriceT. D. (2006). Reduced territorial responses in dark-eyed juncos following population establishment in a climatically mild environment. Anim. Behav. 71, 893–899. 10.1016/j.anbehav.2005.08.007
64
Niall DaisleyJ.BromundtV.MöstlE.KotrschalK. (2005). Enhanced yolk testosterone influences behavioral phenotype independent of sex in Japanese quail chicks Coturnix japonica. Horm. Behav.47, 185–194. 10.1016/j.yhbeh.2004.09.006
65
ParteckeJ.SchwablI.GwinnerE. (2006). Stress and the city: urbanization and its effects on the stress physiology in European Blackbirds. Ecology87, 1945–1952. 10.1890/0012-9658(2006)87[1945:SATCUA]2.0.CO;2
66
PascualJ.SenarJ. C. (2014). Antipredator behavioural compensation of proactive personality trait in male Eurasian siskins. Anim. Behav. 90, 297–303. 10.1016/j.anbehav.2014.02.002
67
PreisznerB.PappS.PipolyI.SeressG.VinczeE.LikerA.et al. (2017). Problem-solving performance and reproductive success of great tits in urban and forest habitats. Anim. Cogn.20, 53–63. 10.1007/s10071-016-1008-z
68
R Development Core Team (2017). R: A Language and Environment for Statistical Computing. Vienna: R Foundation for Statistical Computing.
69
RichardsonK. M.EwenJ. G.BrekkeP.DoerrL. R.ParkerK. A.ArmstrongD. P. (2016). Behaviour during handling predicts male natal dispersal distances in an establishing reintroduced hihi (Notiomystis cincta) population. Anim. Conserv.20, 135–143. 10.1111/acv.12296
70
RiyahiS.BjörklundM.Mateos-GonzalezF.SenarJ. C. (2017). Personality and urbanization: behavioural traits and DRD4 SNP830 polymorphisms in great tits in Barcelona city. J. Ethol.35, 101–108. 10.1007/s10164-016-0496-2
71
RiyahiS.Sánchez-DelgadoM.CalafellF.MonkD.SenarJ. C. (2015). Combined epigenetic and intraspecific variation of the DRD4 and SERT genes influence novelty seeking behavior in great tit Parus major. Epigenetics10, 516–525. 10.1080/15592294.2015.1046027
72
SalaberriaC.GilD. (2010). Increase in song frequency in response to urban noise in the great tit Parus major as shown by data from the Madrid (Spain) city noise map. Ardeola57, 3–11.
73
SalmónP.NilssonJ. F.NordA.BenschS.IsakssonC. (2016). Urban environment shortens telomere length in nestling great tits, Parus major. Biol. Lett.12:20160155. 10.1098/rsbl.2016.0155
74
SchmidtK. H.Einloft-AchenbachH. (1984). Könnenisolierte meisenpopulationen in städtenihrenbestanderhalten?Vogelwelt105, 97–105.
75
SchoechS. J.BowmanR.BridgeE. S.BoughtonR. K. (2007). Baseline and acute levels of corticosterone in Florida Scrub-Jays (Aphelocoma coerulescens): effects of food supplementation, suburban habitat, and year. Gen. Comp. Endocrinol.154, 150–160. 10.1016/j.ygcen.2007.05.027
76
SenarJ. C.ConroyM. J.QuesadaJ.Mateos-GonzalezF. (2014). Selection based on the size of the black tie of the great tit may be reversed in urban habitats. Ecol. Evol.4, 2625–2632. 10.1002/ece3.999
77
SenarJ. C.DomènechJ.CarrascalL. M.MorenoE. (1997). A funnel trap for the capture of tits. Butll. GCA14, 17–24.
78
SeressG.LipovitsÁ.BókonyV.CzúniL. (2014). Quantifying the urban gradient: a practical method for broad measurements. Landsc. Urban Plan.131, 42–50. 10.1016/j.landurbplan.2014.07.010
79
SlabbekoornH.PeetM. (2003). Birds sing at a higher pitch in urban noise. Nature424, 267–268. 10.1038/424267a
80
SolD.LapiedraO.González-LagosC. (2013). Behavioural adjustments for a life in the city. Anim. Behav. 85, 1101–1112. 10.1016/j.anbehav.2013.01.023
81
SprauP.DingemanseN. J. (2017). An approach to distinguish between plasticity and non-random distributions of behavioral types along urban gradients in a wild passerine bird. Front. Ecol. Evol. 5:92. 10.3389/fevo.2017.00092
82
SvenssonL. (1992). Identification Guide to European Passerines. Stockholm: L. Svensson.
83
ThorsteinsenC. (2015). Reproductive Success and Survival during the Breeding Season in Relation to Individual Behaviour in the Great Tit, Parus Major. Oslo: University of Oslo.
84
ToledoA.AnderssonM. N.WangH. L.SalmónP.WatsonH.BurdgeG. C.et al. (2016). Fatty acid profiles of great tit (Parus major) eggs differ between urban and rural habitats, but not between coniferous and deciduous forests. Sci. Nat.103, 1–11. 10.1007/s00114-016-1381-0
85
Torné-NogueraA.Pagani-Nú-ezE.SenarJ. C. (2014). Great Tit (Parus major) breath rate in response to handling stress: urban and forest birds differ. J. Orn. 155, 315–318. 10.1007/s10336-013-1025-5
86
TryjanowskiP.MøllerA. P.MorelliF.BiadunW.BrauzeT.CiachM.et al. (2016). Urbanization affects neophilia and risk-taking at bird-feeders. Sci. Rep. 6:28575. 10.1038/srep28575
87
TuomainenU.CandolinU. (2012). Behavioural responses to human-induced environmental change. Biol. Rev. 86, 640–657. 10.1111/j.1469-185X.2010.00164.x
88
TurnerW. R.NakamuraT.DinettiM. (2004). Global urbanization and the separation of humans from nature. Bioscience54, 585–590. 10.1641/0006-3568(2004)054[0585:GUATSO]2.0.CO;2
89
United Nations (2014). World Urbanization Prospects: The 2014 Revision, Highlights (ST/ESA/SER.A/352).New York, NY: Department of Economic and Social Affairs/Population Division 3.
90
van den BrinkV.DolivoV.FalourdX.DreissA. N.RoulinA. (2012a). Melanic color-dependent antipredator behavior strategies in barn owl nestlings. Behav. Ecol. 23, 473–480. 10.1093/beheco/arr213
91
van den BrinkV.HenryI.WakamatsuK.RoulinA. (2012b). Melanin-based coloration in juvenile kestrels (Falco tinnunculus) covaries with anti-predatory personality traits. Ethology118, 673–682. 10.1111/j.1439-0310.2012.02057.x
92
van OersK.CarereC. (2007). Long-term effects of repeated handling and bleeding in wild caught Great Tits Parus major. J. Ornithol.148, 185–190. 10.1007/s10336-007-0200-y
93
van OverveldT.MatthysenE. (2010). Personality predicts spatial responses to food manipulations in free-ranging great tits (Parus major). Biol. Lett.6, 187–190. 10.1098/rsbl.2009.0764
94
VaugoyeauM.AdriaensenF.ArtemyevA.BanburaJ.BarbaE.BiardC.et al. (2016). Interspecific variation in the relationship between clutch size, laying date and intensity of urbanization in four species of hole-nesting birds. Ecol. Evol.6, 5907–5920. 10.1002/ece3.2335
95
VinczeE.SeressG.LagiszM.NakagawaS.DingemanseN. J.SprauP. (2017). Does urbanization affect predation of bird nests? A meta-analysis. Front. Ecol. Evol.5:29. 10.3389/fevo.2017.00029
96
ZhangS.LeiF.LiuS.LiD.ChenC.WangP. (2011). Variation in baseline corticosterone levels of Tree Sparrow (Passer montanus) populations along an urban gradient in Beijing, China. J. Orn. 152, 801–806. 10.1007/s10336-011-0663-8
Summary
Keywords
urbanization, distress behavior, handling, coping strategy, distress calling, pecking rate, local adaptation, great tit
Citation
Senar JC, Garamszegi LZ, Tilgar V, Biard C, Moreno-Rueda G, Salmón P, Rivas JM, Sprau P, Dingemanse NJ, Charmantier A, Demeyrier V, Navalpotro H and Isaksson C (2017) Urban Great Tits (Parus major) Show Higher Distress Calling and Pecking Rates than Rural Birds across Europe. Front. Ecol. Evol. 5:163. doi: 10.3389/fevo.2017.00163
Received
05 June 2017
Accepted
29 November 2017
Published
18 December 2017
Volume
5 - 2017
Edited by
Patrick S. Fitze, Museo Nacional de Ciencias Naturales (CSIC), Spain
Reviewed by
Deseada Parejo, Universidad de Extremadura, Spain; Gonçalo C. Cardoso, Universidade do Porto, Portugal
Updates
Copyright
© 2017 Senar, Garamszegi, Tilgar, Biard, Moreno-Rueda, Salmón, Rivas, Sprau, Dingemanse, Charmantier, Demeyrier, Navalpotro and Isaksson.
This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Juan Carlos Senar jcsenar@bcn.cat
This article was submitted to Behavioral and Evolutionary Ecology, a section of the journal Frontiers in Ecology and Evolution
Disclaimer
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.