 Virginia Iglesias
Virginia Iglesias Simon G. Haberle
Simon G. Haberle Andrés Holz
Andrés Holz Cathy Whitlock
Cathy Whitlock- 1Montana Institute on Ecosystems, Montana State University, Bozeman, MT, United States
- 2School of Culture, History and Language, Australian National University, Canberra, ACT, Australia
- 3Department of Geography, Portland State University, Portland, OR, United States
- 4Department of Earth Sciences, Montana State University, Bozeman, MT, United States
Analyses of long-term ecosystem dynamics offer insights into the conditions that have led to stability vs. rapid change in the past and the importance of disturbance in regulating community composition. In this study, we (1) used lithology, pollen, and charcoal data from Mallín Casanova (47°S) to reconstruct the wetland, vegetation, and fire history of west-central Patagonia; and (2) compared the records with independent paleoenvironmental and archeological information to assess the effects of past climate and human activity on ecosystem dynamics. Pollen data indicate that Nothofagus-Pilgerodendron forests were established by 9,000 cal yr BP. Although the biodiversity of the understory increased between 8,480 and 5,630 cal yr BP, forests remained relatively unchanged from 9,000 to 2,000 cal yr BP. The charcoal record registers high fire-episode frequency in the early Holocene followed by low biomass burning between 6,500 and 2,000 cal yr BP. Covarying trends in charcoal, bog development, and Neoglacial advances suggest that climate was the primary driver of these changes. After 2,000 cal yr BP, the proxy data indicate (a) increased fire-episode frequency; (b) centennial-scale shifts in bog and forest composition; (c) the emergence of vegetation-fire linkages not recorded in previous times; and (d) paludification in the last 500 years possibly associated with forest loss. Our results therefore suggest that Nothofagus-Pilgerodendron dominance was maintained through much of the Holocene despite long-term changes in climate and fire. Unparalleled fluctuations in local ecosystems during the last two millennia were governed by disturbance-vegetation-hydrology feedbacks likely triggered by greater climate variability and deforestation.
Introduction
Instrumental and tree-ring records from the southern Andes show a strong warming trend during the last 50 years coupled with a decline in precipitation (Villalba et al., 2003). With rising temperatures, glaciers in the North and South Patagonian icefields are receding up to 100 times faster than at any time since the Little Ice Age (Glasser et al., 2011). These trends are projected to continue in the coming decades and lead to increased moisture deficits, further ice recession, tree mortality, and greater risk of fires (Cabré et al., 2016).
Paleoenvironmental data from Patagonia suggest that past changes in climate and land use have profoundly influenced not only the water balances and disturbance regimes, but also the composition and distribution of vegetation (e.g., Huber et al., 2004; Markgraf et al., 2013). Prior to ca. 18,000 cal yr BP, cold, dry and windy conditions were associated with extensive glaciation in western Patagonia and led to the extirpation of several plant species over much of their former range (Barreda et al., 2007). Open steppe/heath prevailed in unglaciated areas, and trees either migrated to northern latitudes or survived in local refugia (Pastorino and Gallo, 2002). Higher temperatures and a shift in dominance from year-round Antarctic air during glacial times to more humid Pacific winds in the Holocene (Heusser, 2003) promoted the reorganization of vegetation (e.g., Iglesias et al., 2014). In particular, Nothofagus-dominated forests expanded throughout the western Andes. This increase in woody fuel allowed a progressive rise in regional fire activity that peaked between 10,000 and 7,000 cal yr BP (Whitlock et al., 2006). The Late Holocene was characterized by fluctuations in moisture availability that resulted in spatial and temporal variability in vegetation composition and probability of fire (Fletcher and Moreno, 2012). In recent centuries, deforestation has affected forest diversity and soil structure, and these changes threaten ecosystem services, including carbon storage, forage production, and water supply (Veblen et al., 2011). In this study, we reconstruct the Holocene vegetation and fire history in west-central Patagonia (Mallín Casanova 47°38′36.86″S; 72°58′30.81″W; 126 m elev), and assess the impact of climate, fire, and volcanic eruptions on vegetation and wetland dynamics. By analyzing the long-term history of this site, we offer insights into the conditions that have driven change in terrestrial and wetland communities in the past.
Study Area
Lake Casanova and the surrounding ombrotrophic Sphagnum bog (Mallín Casanova) are situated in the Baker River watershed, between the North and South Patagonian icefields (Figure 1). The rugged topography of the area results from Pleistocene glaciation and postglacial alluvial and slope activity (Niemeyer et al., 1984; Sernageomin, 2002). Precipitation is associated with frontal storm systems that migrate eastwards along the path of the Southern Westerlies (Garreaud et al., 2013). Uplift of low-level winds over the Andes produces year-round orographic precipitation on the west side of the mountains (e.g., 4,300 mm yr−1 on San Pedro island; Dirección Meteorológica de Chile, 2017). Conversely, forced subsidence on the leeward slopes causes adiabatic warming of the air masses, resulting in increasingly drier conditions toward the east (e.g., 800 mm yr−1 in Cochrane; Dirección Meteorológica de Chile, 2017).
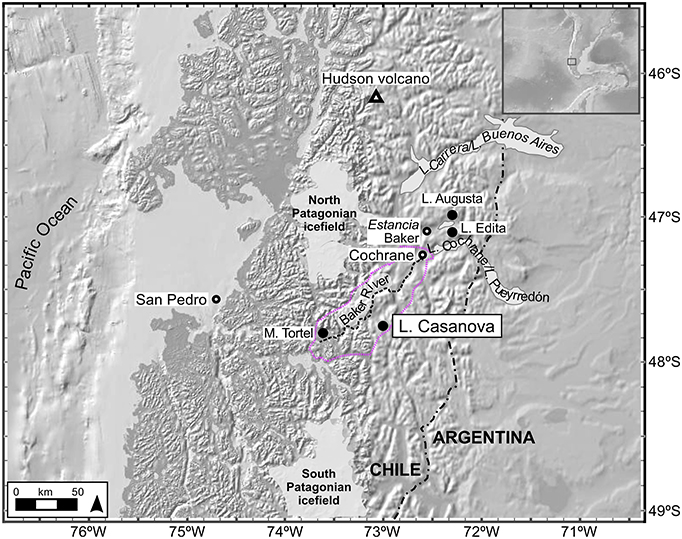
Figure 1. Study area and location of L. Casanova and other sites mentioned in the manuscript. M. Casanova is the ombrotrophic Sphagnum bog surrounding the lake. The coring site is located southeast of L. Casanova. The approximate boundaries of the Baker River watershed are shown in color.
The present-day west-to-east precipitation gradient leads to a transition from evergreen cool rainforests of Nothofagus nitida-N. betuloides-Pilgerodendron in the lowland (precipitation = > 1,500 mm yr−1) to deciduous forests of Nothofagus pumilio (precipitation = 1,500–400 mm yr−1) at higher elevations to steppe characterized by a matrix of tussock grasses, herbs, and cushion shrubs (precipitation <500 mm yr−1). M. Casanova is surrounded by N. betuloides-Pilgerodendron forest with open shrub/grassland and Sphagnum peatland (precipitation = 3,000 mm yr−1). The presence of burned trees on the bog and homogeneous stand structure attest to the effects of recent fires (Holz et al., 2012), which burned both tree crowns and surface fuels. These mixed-severity fires are associated with interannual moisture variability (Holz and Veblen, 2009, 2012). Since ca. 1930 AD, the wetland has been surrounded by Estancia Casanova, a small family farm (Holz and Veblen, 2011).
Methods
A 1-m-long, 80-mm-diameter piston sampler was used to collect a 350-cm-long sediment core from Mallín Casanova at the southeastern margin of the lake. The record provides evidence of environmental change from the beginning of sediment deposition at the wetland to present day. Core segments were transported to the Australian National University, Canberra, where they were split into working and archival halves and subsampled. Description of the sedimentary structures and biological components was performed visually following Schnurrenberger et al. (2001). Magnetic susceptibility (SI) was measured at 1-cm contiguous intervals to assess changes in inorganic sediment input to the wetland (Gedye et al., 2000).
Eight charcoal samples were submitted for AMS radiocarbon dating to the W. M. Keck Carbon Cycle AMS facility (University of California, Irvine) and calibrated with the ShCal13 calibration curve (Hogg et al., 2013; Table 1). Chronologies were developed by modeling depth as a function of the explicitly combined probability distribution of each calibrated date and prior expectations about temporal variability in sediment accumulation rates (Blaauw and Christen, 2011). Specifically, the prior distribution for sediment accumulation rates had a mean of 0.05 cm yr−1 and a shape equal to 1.5. Depths were adjusted by removing volcanic ashes >1 cm in thickness on the assumption that these layers were deposited in a negligible span of time.
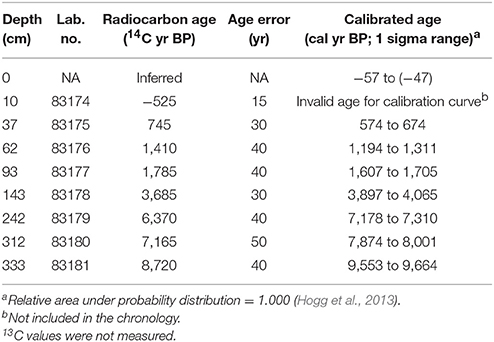
Table 1. Radiocarbon and calibrated dates from M. Casanova.
Past vegetation dynamics were inferred from changes in pollen abundance through time. Standard techniques were employed to extract pollen residues from 0.5 cm−3 sediment samples at 2–5 cm intervals (Faegri and Iversen, 1989). Pollen identification was based on published atlases (Heusser, 1971; Markgraf and D'Antoni, 1978) and a modern reference collection, and performed at 250 and 400x magnification. Pollen percentages (both terrestrial and aquatic) were calculated as a function of the sum of terrestrial pollen types, which in all cases exceeded 250 grains per sample to ensure replicability (Iglesias et al., 2016b).
Empetrum and Caryophyllaceae were excluded from the terrestrial pollen sum. Modern pollen-vegetation studies suggest that, due to the very limited pollen dispersal capabilities of these taxa, they are depicted in the pollen rain only when locally present in the landscape (Fletcher and Thomas, 2007; Iglesias et al., 2016b). This high fidelity is likely to be reinforced in peat deposits, where local vegetation is overrepresented in the pollen spectrum (Janssen, 1973). For these reasons, we assumed that variations in the pollen abundance of Empetrum and Caryophyllaceae probably reflected changes in wetland composition rather than changes in terrestrial vegetation at the watershed-scale (Huber et al., 2004), although we recognize that both taxa have wider ecological distributions.
For this study, pollen types exceeding 3% of the terrestrial pollen sum are discussed in detail, while poorly represented taxa were grouped according to their ecological affinities. Nothofagus dombeyi-type includes N. pumilio, N. dombeyi, N. nitida, N. betuloides, and N. antarctica. Currently, only N. betuloides and N. antarctica grow in the study area. Cupressaceae pollen is attributed to Pilgerodendron uviferum, although Fitzroya cupressoides and, less likely, Austrocedrus chilensis may have been long-distance contributors. Pollen from Hydrangea, Eucryphia/Caldcluvia-type, Myrtaceae, Weinmannia, Coriaria, Drimys, Pseudopanax, and Saxegothaea are grouped into “Other rainforest taxa.” “Other shrubs” includes Schinus, Embothrium, Escallonia, Verbenaceae, Rhamnaceae, and Berberis. “Other grassland taxa” corresponds to pollen from xerophytic shrubs and herbs, such as Acaena, Phacelia, and Mulinum. Selected pollen percentages were plotted as a function of time. The resulting pollen diagram was zoned by visual inspection to aid in the description of the stratigraphic changes.
In order to obtain a local high-resolution fire reconstruction, macroscopic charcoal (i.e., particles > 125 μm in diameter) was extracted from contiguous 1-cm thick samples and quantified under a binocular dissecting microscope (Whitlock and Larsen, 2001). Changes in sediment accumulation rates and sampling variability were accounted for by converting charcoal concentrations to charcoal accumulation rates (CHAR; particles cm−2 yr−1) and interpolating the data to the median time resolution.
A locally weighted scatterplot smoother was used to isolate the high-frequency component of the charcoal time series (i.e., positive residuals of the model) from the long-term trends in CHAR (i.e., “background charcoal”). Comparison of the charcoal data and fire scars in tree rings of P. uviferum shows high correlation coefficients and suggests that the CHAR times series constitutes a record of biomass burning within 10 km of the bog (Holz et al., 2012). Large CHAR values (i.e., positive residuals of the model that exceeded the 95th percentile of a locally fit Gaussian distribution; “charcoal peaks”) were tested for significance with a Poisson distribution and interpreted as local fire episodes (i.e., fire events occurring within the time span of the charcoal peak; Higuera et al., 2009). Local fire episodes were summarized as fire-episode frequency (i.e., fire episodes 1,000 years−1).
To explore the relationship between variations in fire and climate, we calculated the probability of fire episodes occurring at times of regional glacier advance or retreat as:
with Pr(firegl): probability of fire at times of glacier advances; Sum(Fire episodesgl): total number of fire episodes at times of glacier advances; Total samplesgl: number of sample at times of glacier advances; Pr(firenon-gl): probability of fire at times of glacier retreat; Sum(Fire episodesgl): total number of fire episodes at times of glacier retreat and Total samplesnon-gl: number of sample at times of glacier retreat. Times of glacier advances and retreat were obtained from Aniya (2013). A Chi-squared test for proportions was used to assess if the probabilities of fire at times of glacier advances and retreat were statistically different at the 0.05 alpha-level (Table 2).
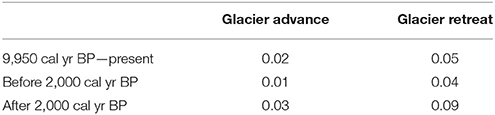
Table 2. Probability of fire episodes at times of glacier advances or glacial retreat.
Long-term changes in community composition were inferred from a dissimilarity matrix produced for the terrestrial taxa tallied in the pollen samples. Specifically, we calculated the squared chord distance (SCD) between every sample and the first sample of the record (i.e., 344 cm depth; 9,840 cal yr BP) as an assessment of their dissimilarity from the oldest pollen assemblages. We interpreted this time series as a measure of long-term trends in the stability of the vegetation. Under the premise that stable ecosystems remain mostly unchanged over time or fluctuate around an equilibrium point, our null hypothesis was that SCDs would be equal to the median Holocene SCD. SCDs larger than the median would provide evidence of statistically significant change in vegetation composition. In order to account for change in sampling effort, rates of change between pairs of consecutive samples were also estimated. Rates of change were defined as:
Rates of change of zero would result from the comparison of identical samples (i.e., no temporal changes in composition = stable plant communities), while non-zero values would be indicative of changes in vegetation composition or unstable communities between times t and t–1. Because we acknowledge the possibility of sampling errors, we bootstrapped confidence intervals for the median Holocene SCD and for rates of change equal to zero. SCDs and rates of change were visually compared with charcoal data, climate variability inferred from independent proxies, and the archeological and historical record. The same procedure was applied to aquatic taxa and interpreted as a proxy of wetland dynamics.
Spectral analysis was conducted on the terrestrial SCD time series to detect possible changes in the periodicity of oscillations in vegetation composition. Specifically, a dynamic Lomb-Scargle periodogram was used to identify periodic signals in the unevenly spaced time series (Press and Rybicki, 1989). All analyses and figures were performed with R (R Core Team, 2015).
Results
Chronology
Seven AMS radiocarbon dates and the inferred age of the top sample were used in the construction of a calibrated age-depth model (Table 1). The radiocarbon age corresponding to the sample extracted at 10-cm depth (−525 ± 15 14C yr BP) was judged to be anomalously young and not included in age model development. The estimated age-depth relationship was relatively linear, suggesting quite stable sediment accumulation rates (0.01–0.07 cm yr−1; Figure 2).
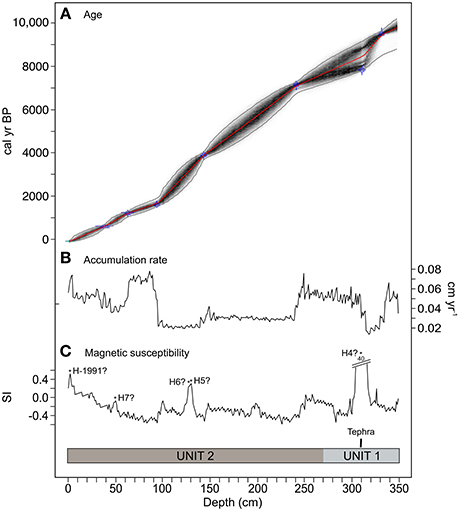
Figure 2. (A) Age-depth model (The red curve shows the weighted mean age for each depth. Dark and light gray lines depict the more likely age-depth models and 95% confidence intervals, respectively. Blue areas represent the probability distributions of the calibrated dates), (B) sediment accumulation rates and (C) magnetic susceptibility for M. Casanova. Lithology and tephra –probably derived from eruptions of the Hudson volcano (H)- are shown. Depths are given in true core measurements, not the adjusted depths used in the development of the chronology.
Lithology
Two lithological units were visually identified in the core from M. Casanova (Figure 2). The basal unit (Unit 1: 350–275 cm depth; 9,950–7,840 cal yr BP) was composed of fine clay. With the exception of two peaks at 326 and 310 cm depth (9,220 and 8,520 cal yr BP; 0.19 and 39.62 SI, respectively), magnetic susceptibility was always negative (median = −0.18 SI), suggesting that the sediment was mainly composed of diamagnetic minerals, such as quartz, calcite, and/or organic matter. A large peak in magnetic susceptibility at 310 cm depth corresponds with a 4-cm-thick tephra, possibly derived from the H4 eruption of the Hudson volcano. H4 tephra deposits are widespread in Patagonia. AMS radiocarbon dates place the mean age of this tephra at 8,440 cal yr BP in lake cores and 7,891 cal yr BP in outcrops, bog, and soil samples (Stern et al., 2016). Due to this systematic difference between radiocarbon dates, which has been attributed to the percolation of humic acids in soils and bogs (Bertrand et al., 2012) as well as to 14C reservoir effects in lakes (Stern et al., 2016), the age of this tephra was not employed in the development of the core chronology.
The uppermost unit (Unit 2: 275–0 cm depth; 7,840 cal yr BP-present) was characterized by brown fibrous peat, low magnetic susceptibility (median = 0.20 SI), and overall low sedimentation rates (median = 0.03 cm yr−1). Gray-brown fine material was observed at 258 and 251 cm depth (7,510 and 7,320 cal yr BP, respectively). Weakly positive magnetic susceptibility values at those depths, as well as in the 136–131 and 27–0 cm depth segments (3,630–3,400 and 450 cal yr BP-present, respectively) indicate the presence of paramagnetic minerals (e.g., iron oxides, iron carbonates, and/or iron silicates). Although no lithological changes were visually identified in this unit, it is possible that magnetic enrichment of these three segments resulted from the deposition of basaltic to andesitic tephra produced during the H5 (pooled mean age = 3,840 cal yr BP), H6 (pooled mean age = 2,740 cal yr BP) and 1991 AD eruptions of the Hudson volcano (Haberle and Lumley, 1998).
Pollen and Charcoal Records
The pollen record from M. Casanova was visually divided in five pollen zones to aid in the description of vegetation and fire history reconstructions (Figures 3, 4). Pollen data are publicly available at Neotoma Paleoecology Database (www.neotomadb.org), and charcoal data have been deposited in the Global Charcoal Database (www.gpwg.org).
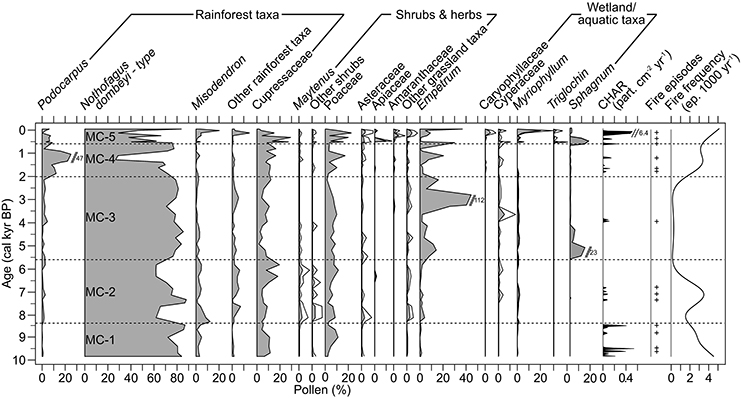
Figure 3. Pollen percentages of selected taxa and macroscopic charcoal data from M. Casanova. Exaggeration curves depict 3*taxa percentages.
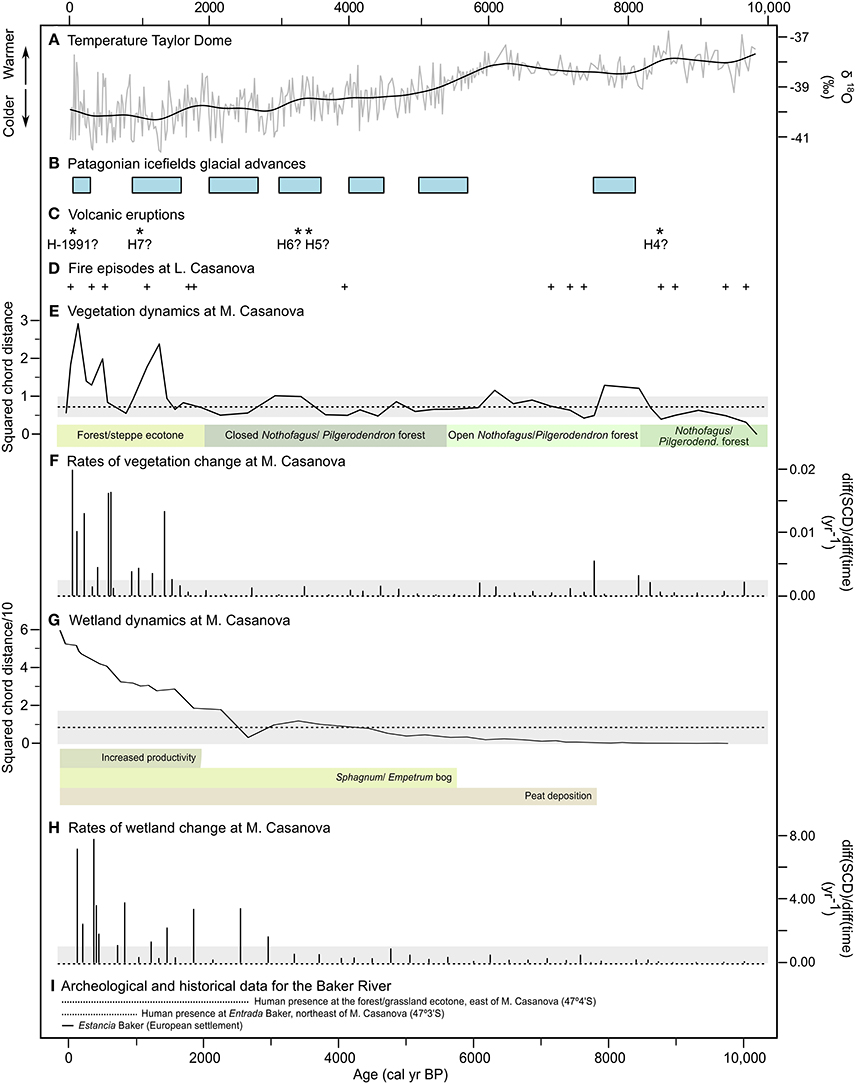
Figure 4. (A) δ18O record from Taylor Dome (Grootes et al., 1993). (B) Holocene glacial advances of the Patagonian icefields (Aniya, 2013). (C) Possible events of tephra deposition associated with known eruptions of the Hudson volcano. (D) Reconstructed fire episodes from M. Casanova charcoal data. Long-term dynamics in (E) vegetation and (G) wetland composition as inferred from the squared chord distance (SCD) between every pollen sample and the first sample of the record. The median Holocene SCDs (dashed line) and their confidence intervals (gray shading) are shown. Pollen zones and wetland characteristics (as interpreted from changes in lithology, sedimentation rates, magnetic susceptibility, and the presence/absence of Sphagnum spores) are indicated. Estimated rates of change in (F) vegetation and (H) wetland composition inferred from pollen data. Confidence intervals for 0 yr−1 (i.e., no change) are depicted in gray. (I) Archeological and historical information for the study area (Martinic, 1977; Mena and Jackson, 1991; Fuentes-Mucherl et al., 2012).
Zone MC-1 (350–308 cm depth; 9,950–8,480 cal yr BP) was dominated by N. dombeyi-type (72–86%), Poaceae (<11%) and Cupressaceae (<11%). Other rainforest taxa, Other shrubs, and Other grassland taxa, including Asteraceae and Amaranthaceae, were poorly represented in the pollen assemblage (1<% in all cases). The vegetation probably resembled present-day Nothofagus/Pilgerodendron forest, and was relative stable (rates of change <0.003% yr−1; Haberle and Bennett, 2001). Aquatic and wetland pollen and spores were virtually absent (<0.7 %; rates of change <0.005% yr−1), suggesting that the wetland was poorly developed at the time, or that the modern lake extended to the coring site. CHAR levels oscillated between 0 and 0.49 particles cm−2 yr−1, and local fire-episode frequency steadily declined from 4.8 episodes 1,000 yr−1 at 9,950 cal yr BP to 1.7 episodes 1,000 yr−1 at the top of the zone.
Zone MC-2 (308–195 cm depth; 8,480–5,630 cal yr BP) continued to be dominated by N. dombeyi-type (65–88%) and Cupressaceae (15–19%), but pollen from Misodendron (<12.3%), Maytenus (<3%), Other shrubs (<3%), and Other grassland taxa (<2%) was higher than before. This increase in palynological richness implies diversification of the understory and/or opening of the Nothofagus/Pilgerodendron forest. High rates of change in terrestrial SCDs point to fast changes in vegetation composition at the 8,320–8,150 and 7,660–7,510 cal yr BP periods. These fluctuations are evidenced by pronounced variability in N. dombeyi-type pollen percentages. Empetrum, Myriophyllum, and Cyperaceae rose up to 4%, but the aquatic rates of change remained low (<0.01% yr−1). CHAR was lower than before (<0.15 particles cm−2 yr−1), and fire-episode frequency declined to 0.1 after reaching a local maximum of 3.5 fire episodes 1,000 yr−1 at 7,100 cal yr BP.
Zone MC-3 (195–101 cm depth; 5,630–2,000 cal yr BP). N. dombeyi-type increased to 85.4% at expense of Misodendron (<3%), Cupressaceae (<12%), Other rainforest taxa (<2.5%), Maytenus (<0.6%), and Other shrubs (<1.5%). Vegetation-pollen calibrations indicate that the vegetation was dominated by closed Nothofagus/Pilgerodendron forest (rates of change <0.003% yr−1; Haberle and Bennett, 2001). Empetrum pollen and Sphagnum spores rose and reached peaks of 112 and 23% of the total terrestrial sum, respectively, suggesting the establishment of the modern Sphagnum bog at the beginning of this zone. Although Sphagnum values were erratic, modern studies suggest that the presence of spores in the sediment is indicative of moss accumulation (Halsey et al., 2000). Terrestrial rates of change remained stable throughout the zone while aquatic SCDs began to slowly rise at ca. 5,000 cal yr BP. CHAR declined to <0.08 particles cm2 yr−1, and so did local fire-episode frequency, which ranged between 0 and 2 fire episodes 1,000 yr−1.
Zone MC-4 (101–31 cm depth; 2,000–530 cal yr BP). N. dombeyi-type (<77%), Misodendron (>1.6%), and Other rainforest taxa (<2%) declined, and Podocarpus and Cupressaceae pollen percentages were higher than before (<47.4 and <27.3%, respectively). CHAR and fire-episode frequency increased to 0.19 particles cm2 yr−1 and 4.8 fire episodes 1000 yr−1, respectively, suggesting that the forest supported more frequent and possibly more severe fires than before. Terrestrial and aquatic SCDs greater than the Holocene median and high rates of change imply large and rapid variations in terrestrial and bog vegetation throughout the period. Dynamic spectral analysis of terrestrial SCDs shows high power at 300- and 700-year periods during the last 2000 years (Figure 5).
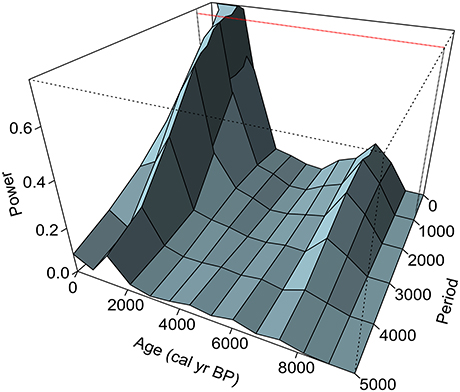
Figure 5. Dynamic Lomb-Scargle periodogram for the SCD time series. Note the increase in power at 700- and 300-year periods during the last 2,000 years. Power values larger than 0.78 are statistically significant (p = 0.05; depicted with a red line).
Zone MC-5 (31–0 cm depth; 530 cal yr BP-present) was characterized by rapid fluctuations in pollen percentages of most taxa (terrestrial rates of change >0.003% yr−1). Overall, Podocarpus and N. dombeyi-type declined to >3.4% and >26.8%, respectively, while Cupressaceae (<29.3%), Poaceae (<22.6%), Apiaceae (<9.8%), and Other grassland taxa (<7%) increased. This pollen assemblage compares well with modern samples from the forest/grassland ecotone found in the wet Chonos archipelago in westernmost Patagonia (44–47°S; Haberle and Bennett, 2001). CHAR (0–6.4 particles cm2 yr−2), pollen of Caryophyllaceae (<5%), Cyperaceae (<3%), Myriophyllum (<21%), and Triglochin (<3%), terrestrial rates of change, and aquatic SCDs reached their highest Holocene values. Fire-episode frequency increased to Zone 1 levels (4.8 fire episodes 1,000 yr−1).
Discussion and Conclusions
Vegetation and Fire History of West-Central Patagonia
During the Last Glacial Maximum (LGM; 23,000–19,000 cal yr BP), sea-surface temperatures off the coast of Chile were ~6°C lower than at present (Lamy et al., 2004; Kaiser et al., 2007). Reconstructions of glacier extent suggest that the North and South Patagonian icefields along with their outlet glaciers coalesced to form a >1200-m-thick ice sheet (Caldenius, 1932; Glasser et al., 2008; ~38–56°). Rapid thinning of the ice sheet at 17,500 cal yr BP was associated with changes in atmospheric circulation and regional warming, and resulted in ice-free conditions in the study area by 15,000 cal yr BP (Hulton et al., 2002; Boex et al., 2013).
As the glaciers retreated, deep glacially-carved valleys filled with a system of lakes. Present-day Lake General Carrera/Lake Buenos Aires and Lake Cochrane/Lake Pueyrredón (Figure 1) are the remnants of a glacial lake that extended between 46 and 48°S. Dammed in the west by the icefields and mountains, the glacial lakes drained into the Atlantic Ocean (Clapperton, 1993). Between 12,800 and 8,000 cal yr BP, the water breached the ice dam resulting in a regional drainage reversal toward the west. The watershed of the Baker River, where M. Casanova is located, adopted its modern configuration at this time (Turner et al., 2005; Bell, 2008).
In late-glacial times (17,500–10,000 cal yr BP), rising winter and annual insolation coupled with changes in atmospheric circulation led to increasingly higher temperatures in Antarctica (Figure 4A) and southern South America, forcing a poleward shift of the Southern Westerlies to a position south of the study region (Rojas et al., 2009). This trend toward warmer conditions and longer growing seasons peaked in the early Holocene and favored the expansion of tree populations throughout western Patagonia (Moreno, 2004; Iglesias et al., 2016a). Initial tree expansion was time transgressive, occurring as early as 16,500 cal yr BP at 41°S (Whitlock et al., 2006), at 15,000 at 44–45°S (Haberle and Bennett, 2004; Massaferro et al., 2005), at 12,500 at 46°S (Lumley and Switsur, 1993), and not until 7,000 cal yr BP at 52°S (Markgraf and Huber, 2010). This north-to-south pattern represents a response to changes in effective moisture associated with warming and a southward shift of the westerly storm tracks (Iglesias et al., 2016a). It is possible that poorly developed soils and the presence of ice in high-elevation valleys (Aniya, 2013) were contributing factors to the late colonization of areas adjacent to the Patagonian icefields.
Pollen data from Lake Augusta (Villa-Martínez et al., 2012) and Lake Edita (Henríquez et al., 2016) located on the eastern flanks of the Andes at 47°S (Figure 1) indicate that expansion of Nothofagus began with ice recession and continued until 9,000 cal yr BP. At M. Casanova, Nothofagus (possibly N. betuloides) and Cupressaceae (likely Pilgerodendron) dominated mountain slopes before 8,480 cal yr BP (Figure 3). Deposition of fine clay and low percentages of aquatic pollen and spore percentages (<0.7%) suggest the presence of open water.
Fire frequencies declined from 4.8 episodes 1,000 yr−1 at 9,840 cal yr BP to 1.7 episodes 1,000 yr−1 at 8,500 cal yr BP (Figure 3). Frequent fires prior to 9,000 cal yr BP are consistent with charcoal records throughout western Patagonia. Frequent burning at the time is attributed to increasing fuel loads coupled with an early onset of the fire season and higher probability of lightning (Haberle and Bennett, 2004; Moreno, 2004; Whitlock et al., 2007; de Porras et al., 2012; Villa-Martínez et al., 2012; Markgraf et al., 2013; Iglesias and Whitlock, 2014). Although it is possible that anthropogenic ignitions were an additional cause of fire, these early-Holocene fire episodes pre-date archeological evidence of humans in the study area.
Between 8,480 and 5,630 cal yr BP, vegetation continued to be dominated by Nothofagus forest. Higher-than-before pollen percentages of Maytenus and other shrubs and herbs, however, point to a more diverse understory and/or forest openings (Figure 3). CHAR was lower than before (<0.15 particles cm−2 yr−1), and fire-episode frequency declined to 0.1 fire episodes 1,000 yr−1 after reaching a maximum of 3.5 fire episodes 1,000 yr−1 at 7,100 cal yr BP. Forest expansion and reduced fire activity have been inferred at sites on both sides of the Andes (e.g., Haberle and Bennett, 2004; Moreno, 2004; Markgraf et al., 2007; Abarzúa and Moreno, 2008; Wille and Schäbitz, 2009; de Porras et al., 2012; Iglesias et al., 2016a) in association with the onset of effectively wetter conditions during the mid-Holocene (Berger and Loutre, 1991; Liu et al., 2003).
Increasingly cooler/wetter conditions during the late Holocene (5,800 cal yr BP-present) supported advances of outlet glaciers from the Patagonian icefields (Aniya, 2013; Figure 4) and aquifer recharge in southern Patagonia (51°S; Larson et al., 2008). At 5,630 cal yr BP, closed forest prevailed in the M. Casanova watershed, and fire activity declined sharply and remained very low until 2,000 cal yr BP (CHAR <0.08 particles cm2 yr−1). Similar trends toward closed forest and reduced fires are observed south of the study site (e.g., Villa-Martínez and Moreno, 2007; Mancini, 2009; Moreno et al., 2009; Wille and Schäbitz, 2009; Echeverría et al., 2014). These conditions contrast with steppe expansion at 44°S (de Porras et al., 2012, 2014) and may be explained by a southward shift and/or weakening of the northern margin of the Southern Westerlies, which would have prevented storm tracks—and therefore precipitation and lightning—from reaching northern Patagonia (Lamy et al., 2010).
Peat accumulation started at the coring site at 7,840 cal yr BP (Figure 2). A rapid increase of Empetrum pollen and Sphagnum spores at 5,800 cal yr BP, nonetheless, suggests that maximum development of the ombrotrophic bog did not occur until the onset of effectively more humid conditions between 5,800 and 2,000 cal yr BP (Figure 3). The rise of Empetrum pollen to its Holocene maximum at 3,500–2,700 cal yr BP is associated with peaks in magnetic susceptibility possibly derived from the H5 and H6 eruptions. This association suggests that Empetrum populations resisted the effects of tephra deposition on soil permeability and geochemistry. Low CHAR at times of Empetrum-Sphagnum bog expansion has also been reported in Fuego-Patagonia (50°S) during the Holocene (Huber and Markgraf, 2003). Huber et al. (2004) interpret the relationship between low fire occurrence and Sphagnum growth as evidence of infrequent burning of forest and bog during periods of higher effective moisture conducive to peat accumulation.
Between 2,000 and 530 cal yr BP, rising lake levels and forest expansion south of 44°S have been recorded east and west of the Andes and are attributed to higher effective moisture (Mancini, 2009; de Porras et al., 2014; Echeverría et al., 2014; Iglesias et al., 2016a). At M. Casanova, faster peat accumulation and lower-than-before magnetic susceptibility (Figure 2) suggest increased bog productivity (Figure 4). Higher effective moisture may also explain the expansion of Podocarpus on poorly drained fluvio-glacial deposits west of the lake or at higher elevations in the study area between 2,000 and 1,000 cal yr BP (Pérez et al., 2016; Figure 3). Rising CHAR at M. Casanova (Figure 3) is consistent with fire-history reconstructions throughout Patagonia that suggest increased biomass burning during this period (Szeicz et al., 2003; Haberle and Bennett, 2004; Villa-Martínez and Moreno, 2007; de Porras et al., 2014; Iglesias and Whitlock, 2014).
The last 530 years are characterized by changes in bog hydrology at M. Casanova, including the establishment of Caryophyllaceae, Triglochin, and other wetland plants, and rapid deposition of peaty sediments (Figures 2–4). The presence of submersed aquatic taxa, such as Myriophyllum, indicates that areas of the bog were flooded at times. High percentages of Asteraceae, Apiaceae, Amaranthaceae, and other herbaceous taxa and a decline in arboreal pollen suggest a loss of forest cover in recent centuries (Figure 3). Frequent fires (up to 5 fire episodes 1,000 yr−1; Figure 3) would have favored the expansion of disturbance-adapted taxa (e.g., Amaranthaceae and herbs) at expense of less-tolerant trees. A decline of arboreal taxa in the last 500 years has also been reported north (44°S; Haberle and Bennett, 2004; de Porras et al., 2014) and south of the study area (50–55°S; Huber et al., 2004; Mancini, 2009). Deforestation through burning and, more recently, logging probably reduced evapotranspiration and facilitated paludification (Holz and Veblen, 2011).
Effects of Climate, Disturbance, and Human Impact on Ecosystem Stability
Paleoenvironmental data from L. Augusta (47°S) suggest that Nothofagus forests prevailed relatively unchanged in the watershed from the time of their establishment at ca. 9,800 cal yr BP to European arrival in the area in the twentieth century. Villa-Martínez et al. (2012) attribute this stability to the high tolerance of N. pumilio, the dominant tree, to changes in precipitation. The pollen record from M. Casanova shows that Nothofagus-Pilgerodendron forests have existed in the study area for about 9,000 years despite long-term changes in climate and fire (Figure 3). Rates of change statistically equal to zero indicate that terrestrial pollen percentages did not vary significantly over the period from 9,000 to 1,400 cal yr BP (Figure 4). The temporal and taxonomic resolution of the pollen time series, however, does not allow examining whether this stability resulted from Nothofagus' resistance to environmental change, its rapid recovery after disturbance (i.e., resilience), or trade-offs in the Nothofagus species [i.e., deciduous, fire-adapted N. antarctica vs. deciduous, obligate-seeder, fire-sensitive N. pumilio vs. evergreen, obligate-seeder, fire-sensitive N. betuloides and N. nitida (Correa, 1984), all of which grow at this latitude and whose pollen cannot be differentiated].
Two periods of rapid change before 1,400 cal yr BP were identified in the terrestrial SCD series. One occurred between 8,320 and 8,150 cal yr BP, when shrub taxa, Asteroideae, herbs, and grasses expanded at the expense of Nothofagus. The other one took place from 7,660 to 7,510 cal yr BP as a result of a pronounced increase in Nothofagus, and was coeval with the start of peat accumulation in the wetland (Figure 4). A reorganization of the ecosystem toward more open vegetation occurred between these two periods of rapid change (8,150–7,660 cal yr BP) and coincided with renewed glaciation in the Patagonian icefields (Aniya, 2013; Bourgois et al., 2016), suggesting that low temperatures probably limited seedling survival. Additionally, tephra from the H4 eruption of the Hudson volcano (Figures 2, 4) may have promoted the acidification of the soils and precluded tree establishment in the study area. It is possible, however, that the change in pollen composition registered from 7,660 to 7,510 cal yr BP did not reflect changes in terrestrial vegetation but rather a shift in depositional environments (i.e., shift from fine clay to peat; Figure 2).
Charcoal times series from the Baker River watershed (i.e., Mallín Tortel [47°48′S, 73°28′W] and M. Casanova; Holz et al., 2012) show synchronous changes in fire activity in the last 7,000 years, including a pronounced rise in biomass burning at 2,000 cal yr BP. Comparison of reconstructed fire episodes at M. Casanova and the glacial record from the Patagonian icefields indicates that the probability of fire was significantly lower during times of ice advance (Table 2; Figure 4). Increased effective moisture and/or lower temperatures are likely to have favored ice expansion and bog productivity and suppressed fire activity (Figure 4).
During the last 2,000 years, fire-episode frequency was as high as that of the early Holocene (>3 fire episodes 1,000 yr−1), despite wetter and/or cooler conditions (Table 2; Figure 4). The large SCDs indicate that, in contrast with the early-and mid-Holocene wetland and vegetation history, the bog has been highly variable since ca. 1,800 cal yr BP, while Nothofagus-Pilgerodendron forests have experienced pronounced and rapid departures from the long-term median composition after ca. 1,400 cal yr BP (Figure 4). The frequency spectrum of terrestrial squared chord distances reveals statistically significant power at centennial-scales during the last 2,000 years (300-year cycles; Figure 5), indicating that changes in vegetation occurred in a quasi-periodic manner. This periodicity matches that reconstructed for late-Holocene fire episodes (Figure 3), pointing to fire-induced forest loss followed by forest recovery/fuel accumulation and renewed fire activity in ca. 300-year cycles. These shifts in local ecosystem dynamics could have been an abrupt response to human impact and/or resulted from nonlinear climate-vegetation-fire linkages.
Humans arrived in Patagonia ca. 18,500 years ago (Dillehay et al., 2015) and lived throughout the region as small, dispersed groups until 7,000 cal yr BP, when population densities increased (Pérez et al., 2016). Resource availability in highly variable ecosystems would have been unpredictable, and when combined with the greater demands from growing human populations (Pérez et al., 2016), would have motivated the use of the Baker River watershed. The increasing presence of people in the area in the last two millennia may have therefore been an adaptation strategy in response to reduced resources per capita. Higher human population densities, in turn, possibly reinforced climate-disturbance-vegetation feedbacks operating at the local scale.
Comparison of reconstructed patterns of human occupation and environmental records from the central Patagonian Andes in Chile (44°S) suggests that hunter-gatherers likely increased the probability of fire, particularly in the last 3,000 years (Méndez et al., 2016). For example, Holz and Veblen (2011) reported burning of Pilgerodendron stands primarily by the Kawésqar canoeing group to secure dry firewood and canoe-building material. Similarly, fires set by aboriginal peoples to hunt guanacos (Lama guanicoe) and rheas (Rhea spp.) in northern Patagonia (~41–43°S) were described by early European explorers and missionaries (Musters, 1871; Cox, 1963).
The shift in vegetation, fire activity and hydrology observed at M. Casanova after ca. 2,000 cal yr BP may therefore be partly anthropogenic. Faunal and lithic analyses from a small rock shelter in the Chacabuco Valley to the east (47°4′48″S, 72°16′20″; ca. 40 km from the study site) suggest use of the forest/grassland ecotone as early as 2,800 cal yr BP (Fuentes-Mucherl et al., 2012). Conversely, archeological evidence from the Baker River watershed shows human presence only after 1,000 cal yr BP (Mena and Jackson, 1991), and permanent settlement is not recorded until the establishment of the Estancia Baker by European ranchers in 1902 AD (ca. 35 km from the study site; Martinic, 1977). The late settlement record in this watershed, however, may reflect a lack of older archeological data, rather than an absence of people before Europeans.
The possibility of human ignitions at M. Casanova does not rule out climate as a driving force of change in local ecosystem dynamics reconstructed for the last 2,000 years that include (a) high fire activity; (b) unprecedented variability in bog composition; (c) emergence of regular fluctuations in the relative abundance of trees and shrubs that seem related to post-fire regeneration; and (d) paludification in association with forest loss. Decadal- and centennial-scale climate variability in the mid- and high latitudes of Patagonia has been ascribed to the Southern Annular Mode (Moreno et al., 2014), El Niño-Southern Oscillation (Whitlock et al., 2006), and to changes in solar irradiance associated with the Vries/Suess solar cycle (Lüdecke et al., 2015). Heightened moisture variability probably favored fires, and well-drained, acidic, tephra-rich soils may have slowed the post-fire regeneration of sensitive taxa. Fire, whether natural or anthropogenic in origin, led to forest loss in the last 500 years, which, in turn, contributed to paludification. Thus, the long-term trend toward cooler, effectively wetter conditions, combined with extreme wet and dry events and more frequent disturbance created a unique set of conditions that led to unprecedented change at M. Casanova starting at ca. 2,000 cal yr BP.
Author Contributions
VI and SH: Designed research; SH and CW: Contributed reagents; VI: Counted pollen samples and analyzed data; VI, SH, AH, and CW: Interpreted results; VI: Wrote the paper; SH, AH, and CW: Edited the manuscript.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
This work was supported by grants from the National Science Foundation (0966472, 0956552, 0602166), the National Geographic Society (7988-06), and the Department of Archaeology and Natural History at the Australian National University. We thank Philip Roberts for counting charcoal samples.
References
Abarzúa, A., and Moreno, P. I. (2008). Changing fire regimes in the temperate rainforest region of southern Chile over the last 16,000 yr. Quat. Res. 69, 62–71. doi: 10.1016/j.yqres.2007.09.004
Aniya, M. (2013). Holocene glaciations of Hielo Patagónico (Patagonia Icefield), South America: a brief review. Geochem. J. 47, 97–105. doi: 10.2343/geochemj.1.0171
Barreda, V., Anzótegui, L. M., Prieto, A. R., Aceñolaza, P., Bianchi, M. M., Borromei, A. M., et al. (2007). Diversificación y cambios de las angiospermas durante el Neógeno en Argentina. Ameghiniana 11, 173–191.
Bell, C. M. (2008). Punctuated drainage of an ice-dammed quaternary lake in Southern South America. Geogr. Ann. 90, 1–17. doi: 10.1111/j.1468-0459.2008.00330.x
Berger, A., and Loutre, M. F. (1991). Insolation values for the climate of the last 10 million years. Quat. Sci. Rev. 10, 297–317. doi: 10.1016/0277-3791(91)90033-Q
Bertrand, S., Araneda, A., Vargas, P., Jana, P., Fagel, N., and Urrutia, R. (2012). Using the N/C ratio to correct bulk Chilean Patagonia. Quat. Geochronol. 12, 23–29. doi: 10.1016/j.quageo.2012.06.003
Blaauw, M., and Christen, J. A. (2011). Flexible paleoclimate age-depth models using an autoregressive gamma process. Bayesian Anal. 6, 457–474. doi: 10.1214/11-BA618
Boex, J., Fogwill, C., Harrison, S., Glasser, N. F., Hein, A., Schnable, C., et al. (2013). Rapid thinning of the late Pleistocene Patagonian Ice Sheet followed migration of the Southern Westerlies. Nat. Sci. Rep. 3:2118. doi: 10.1038/srep02118
Bourgois, J., Cisternas, M. E., Braucher, R., Bourles, D., and Frutos, J. (2016). Goemorphic records along the General Carrera (Chile)-Buenos Aires (Argentina glacial lake (46-48S), climate inferences, and glacial rebound for the past 7-9 ka. J. Geol. 124, 27–53. doi: 10.1086/684252
Cabré, M. F., Solman, S., and Núñez, M. (2016). Regional climate change scenarios over southern South America for future climate (2080-2099) using the MM5 Model. Mean, interannual variability and uncertainties. Atmósfera 29, 35–60. doi: 10.20937/ATM.2016.29.01.04
Caldenius, C. C. (1932). Las glaciaciones cuaternarios en la Patagonia y Tierra del Fuego. Geografiska Annaler, 14, 1–164 (English summary 144–157).
Clapperton, C. M. (1993). Quaternary Geology and Geomorphology of South America. Amsterdam: Elsevier.
Correa, M. E. (1984). Flora Patagónica. Buenos Aires: Instituto Nacional de Tecnología Agropecuaria.
Cox, G. (1963). Viajes a las regiones septentrionales de Patagonia 1862-1863. Anal. Univ. Chile 23, 3–239.
de Porras, M. E., Maldonado, A., Abarzúa, A. M., Cárdenas, M. L., Francois, J. P., Martel-Cea, A., et al. (2012). Postglacial vegetation, fire and climate dynamics at Central Chilean Patagonia (Lake Shaman, 44°S). Quat. Sci. Rev. 50, 71–85. doi: 10.1016/j.quascirev.2012.06.015
de Porras, M. E., Maldonado, A., Quintana, F., Martel-Cea, A., Reyes, O., and Méndez, C. (2014). Environmental and climatic changes at central Chilean Patagonia since the Late Glacial (Mallín El Embudo, 44°S). Clim. Past 10, 1063–1078. doi: 10.5194/cp-10-1063-2014
Dillehay, T. D., Ocampo, C., Saavedra, J., Sawakuchi, A. O., Vega, R. M., Pino, M., et al. (2015). New archaeological evidence for an early human presence at Monte Verde, Chile. PLoS ONE 10:e0141923. doi: 10.1371/journal.pone.0141923
Dirección Meteorológica de Chile (2017). Climatología. Available online at: http://www.meteochile.cl/PortalDMC-web/index.xhtml (Accessed May 31, 2017).
Echeverría, M. E., Sottile, G. D., Mancini, M. V., and Fontana, S. L. (2014). Nothofagus forest dynamics and palaeoenvironmental variations during the mid and late Holocene, in southwest Patagonia. Holocene 24, 957–969. doi: 10.1177/0959683614534742
Fletcher, M., and Moreno, P. I. (2012). Have the Southern Westerlies changed in a zonally symmetric manner over the last 14,000 years? A hemisphere-wide take on a controversial problem. Quat. Int. 253, 32–46. doi: 10.1016/j.quaint.2011.04.042
Fletcher, M.-S., and Thomas, I. (2007). Modern pollen-vegetation calibration relationships in western Tasmania, Australia. Rev. Palaeobot. Palynol. 146, 146–168. doi: 10.1016/j.revpalbo.2007.03.002
Fuentes-Mucherl, F., Mena, F., Blanco, J., and Contreras, C. (2012). Excavaciones en Alero Gianella, curso medio del valle de Chacabuco (Andes centro patagónicos). Magallania 40, 259–265. doi: 10.4067/S0718-22442012000200013
Garreaud, R. D., Lopez, P., Minvielle, M., and Rojas, M. (2013). Large scale control on the Patagonian climate. J. Clim. 26, 215–230. doi: 10.1175/JCLI-D-12-00001.1
Gedye, S. J., Jones, R. T., Tinner, W., Ammann, B., and Oldfield, F. (2000). The use of mineral magnetism in the reconstruction of fire history: a case study from Lago di Origlio, Swiss Alps. Palaeogeogr. Palaeoclimatol. Palaeoecol. 164, 101–110. doi: 10.1016/S0031-0182(00)00178-4
Glasser, N. F., Harrison, S., Jansson, K. N., Anderson, K., and Cowley, A. (2011). Global sea-level contribution from the Patagonian Icefields since the Little Ice Age maximum. Nat. Geosci. 4, 303–307. doi: 10.1038/ngeo1122
Glasser, N. F., Jansson, K. N., Harrison, S., and Kleman, J. (2008). The glacial geomorphology and Pleistocene history of South America between 38 degrees S and 56 degrees S. Quat. Sci. Rev. 27, 365–390. doi: 10.1016/j.quascirev.2007.11.011
Grootes, P. M., Stuiver, M., White, J. W. C., Johnsen, S. J., and Jouzel, J. (1993). Comparison of oxygen isotope records from the GISP2 and GRIP Greenland ice cores. Nature 366, 552–554. doi: 10.1038/366552a0
Haberle, S. G., and Bennett, K. D. (2001). Modern pollen rain and lake mud-water interface geochemistry along environmental gradients in southern Chile. Rev. Palaeobot. Palynol. 117, 93–107. doi: 10.1016/S0034-6667(01)00079-3
Haberle, S. G., and Bennett, K. D. (2004). Postglacial formation and dynamics of North Patagonian Rainforest in the Chonos Archipelago, Southern Chile. Quat. Sci. Rev. 23, 2433–2452. doi: 10.1016/j.quascirev.2004.03.001
Haberle, S. G., and Lumley, S. H. (1998). Age and origin of tephras recorded in postglacial lake sediments to the west of the southern Andes, 44°S to 47°S. J. Volcanol. Geotherm. Res. 84, 239–256. doi: 10.1016/S0377-0273(98)00037-7
Halsey, L. A., Vitt, D. H., and Gignac, L. D. (2000). Sphagum-dominated peatlands in North America since the last glacial maximum: their occurrence and extent. Bryologist 103, 334–352. doi: 10.1639/0007-2745(2000)103[0334:SDPINA]2.0.CO;2
Henríquez, W. I., Villa-Martínez, R., Vilanova, I., de Pol-Holz, R., and Moreno, P. I. (2016). The last glacial termination on the eastern flank of the Patagonian Andes (47°S). Clim. Past 13, 879–895. doi: 10.5194/cp-13-879-2017
Heusser, C. J. (1971). Pollen and Spores of Chile: Modern types of the Pteridophyta, Gymnospermae, and Angiospermae. Tucson, AZ: University of Arizona Press.
Heusser, C. J. (2003). Ice Age in the Southern Andes: A Chronicle of Paleoenvironmental Events. Amsterdam: Elsevier.
Higuera, P. E., Brubaker, L. B., Anderson, P. M., Hu, F. S., and Brown, T. A. (2009). Vegetation mediated the impacts of postglacial climatic change on fire regimes in the south central Brooks Range, Alaska. Ecol. Monogr. 79, 201–219. doi: 10.1890/07-2019.1
Hogg, A. G., Hua, Q., Blackwell, P. G., Buck, C. E., Guilderson, T. P., Heaton, T. J., et al. (2013). ShCal13 Southern Hemisphere calibration, 0-50,000 cal yr BP. Radiocarbon 55, 1889–1903. doi: 10.2458/azu_js_rc.55.16783
Holz, A., Haberle, S., Veblen, T. T., De Pol-Holz, R., and Southon, J. (2012). Fire history in western Patagonia from paired tree-ring fire-scar and charcoal records. Clim. Past 8, 451–466. doi: 10.5194/cp-8-451-2012
Holz, A., and Veblen, T. T. (2009). Pilgerodendron uviferum: the southernmost tree-ring fire recorder species. Ecoscience 16, 322–329. doi: 10.2980/16-3-3262
Holz, A., and Veblen, T. T. (2011). The amplifying effects of humans on fire regimes in temperate rainforests in western Patagonia. Palaeogeogr. Palaeoclimatol. Palaeoecol. 311, 82–92. doi: 10.1016/j.palaeo.2011.08.008
Holz, A., and Veblen, T. T. (2012). Wildfire activity in rainforests in western Patagonia linked to the Southern Annular Mode. Int. J. Wildland Fire 21, 114–126. doi: 10.1071/WF10121
Huber, U. M., and Markgraf, V. (2003). European impact on fire regimes and vegetation dynamics at the steppe–forest ecotone of southern Patagonia. Holocene 13, 567–579. doi: 10.1191/0959683603hl647rp
Huber, U. M., Markgraf, V., and Schäbitz, F. (2004). Geographical and temporal trends in Late Quaternary fire history in Fuego-Patagonia, South America. Quat. Sci. Rev. 23, 1079–1097. doi: 10.1016/j.quascirev.2003.11.002
Hulton, N. R. J., Purves, R. S., McCulloch, R. D., Sugden, D. E., and Bentley, M. J. (2002). The Last Glacial Maximum and deglaciation in southern South America. Quat. Sci. Rev. 21, 233–241. doi: 10.1016/S0277-3791(01)00103-2
Iglesias, V., Markgraf, V., and Whitlock, C. (2016a). 17,000 years of vegetation, fire and climate change in the eastern foothills of the Andes (lat. 44°S). Palaeogeogr. Palaeoclimatol. Palaeoecol. 457, 195–208. doi: 10.1016/j.palaeo.2016.06.008
Iglesias, V., Quintana, F., Nanavati, W., and Whitlock, C. (2016b). Interpreting modern and fossil pollen data along a steep environmental gradient in northern Patagonia. Sage 27, 1008–1018. doi: 10.1177/0959683616678467
Iglesias, V., and Whitlock, C. (2014). Fire responses to postglacial climate change and human impact in northern Patagonia (41-43°S). Proc. Natl. Acad. Sci. U.S.A. 111, E5545–E5554. doi: 10.1073/pnas.1410443111
Iglesias, V., Whitlock, C., Markgraf, V., and Bianchi, M. M. (2014). Postglacial history of the Patagonian forest/steppe ecotone (41-43°S). Quat. Sci. Rev. 94, 120–135. doi: 10.1016/j.quascirev.2014.04.014
Janssen, C. R. (1973). “Local and regional pollen deposition,” in Quaternary Plant Ecology, eds H. J. P. Birks and R.G. West (New York, NY: Wiley & Sons), 31–42.
Kaiser, J., Lamy, F., Arz, H. W., and Hebbeln, D. (2007). Dynamics of the millennial-scale sea surface temperature and Patagonian Ice Sheet fluctuations in southern Chile during the last 70 kyr (ODP Site 1233). Quat. Int. 161, 77–89. doi: 10.1016/j.quaint.2006.10.024
Lamy, F., Kaiser, J., Ninnemann, U., Hebbeln, D., Arz, H. W., and Stoner, J. (2004). Antarctic Timing of Surface Water Changes off Chile and Patagonian Ice Sheet Response. Science 304, 1959–1962. doi: 10.1126/science.1097863
Lamy, F., Kilian, R., Arz, H., Francois, J. P., Kaiser, J., Prangem, M., et al. (2010). Holocene changes in the positions and intensity of the southern westerly wind belt. Nat. Geosci. 3, 695–699. doi: 10.1038/ngeo959
Larson, S. A., Moy, C. M., Dunbar, R. B., and Moreno, P. I. (2008). Lacustrine carbonate records of climate variability in SW Patagonia. AGU Fall Meeting, PP41C-1466. San Francisco.
Liu, Z., Brady, E., and Lynch-Stieglitz, J. (2003). Global ocean response to orbital forcing in the Holocene. Paleoceanography 18, 1041–1060. doi: 10.1029/2002PA000819
Lüdecke, H.-J., Weiss, C. O., and Hempelmamm, A. (2015). Paleoclimate forcing by the solar DeVries/Suess cycle. Clim. Past Discuss. 11, 279–305. doi: 10.5194/cpd-11-279-2015
Lumley, S. H., and Switsur, R. (1993). Late Quaternary chronology of the Taitao Peninsula, southern Chile. J. Quat. Sci. 8, 161–165. doi: 10.1002/jqs.3390080208
Mancini, M. V. (2009). Holocene vegetation and climate changes from a peat pollen record of the forest-steppe ecotone, southwest of Patagonia (Argentina). Quat. Sci. Rev. 28, 1490–1497. doi: 10.1016/j.quascirev.2009.01.017
Markgraf, V., and D'Antoni, H. L. (1978). Pollen Flora of Argentina. Tucson, AZ: University of Arizona Press.
Markgraf, V., and Huber, U. (2010). Late and postglacial vegetation and fire history in Southern Patagonia and Tierra del Fuego. Palaeogeogr. Palaeoclimatol. Palaeoecol. 297, 351–366. doi: 10.1016/j.palaeo.2010.08.013
Markgraf, V., Iglesias, V., and Whitlock, C. (2013). Late and postglacial vegetation and fire history from Cordón Serrucho Norte, northern Patagonia. Palaeogeogr. Palaeoclimatol. Palaeoecol. 371, 109–118. doi: 10.1016/j.palaeo.2012.12.023
Markgraf, V., Whitlock, C., and Haberle, S. (2007). Vegetation and fire history during the last 18,000 cal yr B.P. in Southern Patagonia: Mallin Pollux, Coyhaique, Province Aisen (45°41′30″ S, 71°50′30″ W, 640 m elevation). Palaeogeogr. Palaeoclimatol. Palaeoecol. 254, 492–507. doi: 10.1016/j.palaeo.2007.07.008
Martinic, M. B. (1977). Ocupación y colonización de la Región Septentrional del Antiguo Territorio de Magallanes, entre los paralelos 47° y 49 Sur. Anales del Instituto Patagónico 8, 5–57.
Massaferro, J., Brooks, S. J., and Haberle, S. G. (2005). The dynamics of chironomid assemblages and vegetation during the Late Quaternary at Laguna Facil, Chonos Archipelago, southern Chile. Quat. Sci. Rev. 24, 2510–2522. doi: 10.1016/j.quascirev.2005.03.010
Mena, F., and Jackson, D. (1991). Tecnología y subsistencia en el alero entrada Baker, región de Aisén, Chile. Anales del Instituto Patagónico Serrano de Ciencias Sociales 20, 169–204.
Méndez, C., de Porras, M. E., Maldonado, A., Reyes, O., Nuevo Delaunay, A., and García, J.-L. (2016). Human effects in Holocene fire dynamics of central western Patagonia (~44°S, Chile). Front. Ecol. Evol. 4:100. doi: 10.3389/fevo.2016.00100
Moreno, P. I. (2004). Millennial-scale climate variability in northwest Patagonia over the last 15,000 yr. J. Quat. Sci. 19, 35–47. doi: 10.1002/jqs.813
Moreno, P. I., François, J. P., Villa-Martínez, R. P., and Moy, C. M. (2009). Millennial-scale variability in Southern Hemisphere westerly wind activity over the last 5000 years in SW Patagonia. Quat. Sci. Rev. 28, 25–38. doi: 10.1016/j.quascirev.2008.10.009
Moreno, P. I., Vilanova, I., Villa-Martínez, R., Garreaud, R. D., Rojas, M., and De Pol-Holz, R. (2014). Southern Annular Mode-like changes in southwestern Patagonia at centennial timescales over the last three millennia. Nat. Commun. 5:4375. doi: 10.1038/ncomms5375
Musters, G. C. (1871). At Home with the Patagonians: A Year's Wanderings over Untrodden Ground from the Straits of Magellan to the Río Negro. London: Murray.
Niemeyer, H. R., Skarmeta, J. M., Fuenzalida, R. P., and Espinosa, W. N. (1984). Hojas Península de Taitao y Puerto Aisén, Región Aisén del General Carlos Ibañez del Campo. Santiago: Servicio Nacional de Minería.
Pastorino, M., and Gallo, L. (2002). Quaternary evolutionary history of Austrocedrus chilensis, a cypress native to the Andean Patagonian forest. J. Biogeogr. 29, 1167-1178. doi: 10.1046/j.1365-2699.2002.00731.x
Pérez, S. I., Postillone, M. B., Rindel, D., Gobbo, D., Gonzalez, P. N., and Bernal, V. (2016). Peopling time, spatial occupation and demography of the Late Pleistocene-Holocene human population from Patagonia. Quat. Int. 425, 214–223. doi: 10.1016/j.quaint.2016.05.004
Press, W. H., and Rybicki, G. B. (1989). Fast algorithm for spectral analysis of unevenly sampled data. Astrophys. J. 388, 277–280. doi: 10.1086/167197
R Core Team (2015). R: A Language and Environment for Statistical Computing. Vienna: R Foundation for Statistical Computing.
Rojas, M., Pi, M., Kageyama, M., Crucifix, M., Hewitt, C., Abe-Ouchi, A., et al. (2009). The southern westerlies during the last glacial maximum in PMIP2 simulations. Clim. Dyn. 32, 525–548. doi: 10.1007/s00382-008-0421-7
Schnurrenberger, D. B., Kelts, K. R., Johnson, T. C., Shane, L. C. K., and Ito, E. (2001). National lacustrine core repository (LacCore). J. Paleolimnol. 25, 123–127. doi: 10.1023/A:1008171027125
Sernageomin (2002). Carta Geológica de Chile, N°75, Escala: 1,000,000, Hoja 3. Servicio Nacional de Geología y Minería, Subdireccion Nacional de Geologia, Chile.
Stern, C. R., Moreno, P. I., Henríquez, W. I., Villa-Martínez, R., Sagredo, E., Aravena, J. C., et al. (2016). Holocene tephrochronology around Cochrane (~47°S), southern Chile. Andean Geol. 43, 1–19. doi: 10.5027/andgeoV43n1-a01
Szeicz, J. M., Haberle, S. G., and Bennett, K. D. (2003). Dynamics of north Patagonian rainforests from fine-resolution pollen, charcoal and tree-ring analysis, Chonos Archipelago, southern Chile. Austral Ecol. 28, 413–422. doi: 10.1046/j.1442-9993.2003.01299.x
Turner, K. J., Fogwill, C. J., McCulloch, R. D., and Sugden, D. E. (2005). Deglaciation of the eastern flank of the North Patagonian Icefield and associated continental-scale lake diversions. Geogr. Ann. 87, 363–374. doi: 10.1111/j.0435-3676.2005.00263.x
Veblen, T. T., Holz, A., Paritsis, J., Raffaele, E., Kitzberger, T., and Blackhall, M. (2011). Adapting to global environmental change in Patagonia: what role for disturbance ecology? Austral. Ecol. 36, 891–903. doi: 10.1111/j.1442-9993.2010.02236.x
Villalba, R., Lara, A., Boninsegna, J. A., Masiokas, M., Delgado, S., Aravena, J. C., et al. (2003). Large-scale temperature changes across the southern Andes: 20th century variations in the context of the past 400 years. Clim. Change 59, 177–232. doi: 10.1023/A:1024452701153
Villa-Martínez, R., Moreno, P. I., and Valenzuela, M. A. (2012). Deglacial and postglacial vegetation changes on the eastern slopes of the central Patagonian Andes (47 degrees S). Quat. Sci. Rev. 32, 86–99. doi: 10.1016/j.quascirev.2011.11.008
Villa-Martínez, R. P., and Moreno, P. I. (2007). Pollen evidence for variations in the southern margin of the westerly winds in SW Patagonia over the last 12,600 years. Quat. Res. 68, 400–409. doi: 10.1016/j.yqres.2007.07.003
Whitlock, C., Bianchi, M. M., Bartlein, P. J., Markgraf, V., Marlon, J., Walsh, M., et al. (2006). Postglacial vegetation, climate, and fire history along the east side of the Andes (lat. 41-42.5° S), Argentina. Quat. Res. 66, 187–201. doi: 10.1016/j.yqres.2006.04.004
Whitlock, C., and Larsen, C. P. S. (2001). “Charcoal as a fire proxy,” in Tracking Environmental Change Using Lake Sediments: Terrestrial, Algal, and Siliceous indicators, Vol. 3, eds J. P. Smol, H. J. P. Birks and W. M. Last (Dordrecht: Kluwer Academic Publishers), 75–97.
Whitlock, C., Moreno, P. I., and Bartlein, P. J. (2007). Climatic controls of Holocene fire patterns in southern South America. Quat. Res. 68, 28–36. doi: 10.1016/j.yqres.2007.01.012
Keywords: anthropogenic impact, charcoal, climate, pollen, stability, vegetation, Holocene, forest
Citation: Iglesias V, Haberle SG, Holz A and Whitlock C (2018) Holocene Dynamics of Temperate Rainforests in West-Central Patagonia. Front. Ecol. Evol. 5:177. doi: 10.3389/fevo.2017.00177
Received: 23 October 2017; Accepted: 28 December 2017;
Published: 26 January 2018.
Edited by:
Valentí Rull, Instituto de Ciencias de la Tierra Jaume Almera (CSIC), SpainReviewed by:
Antonio Maldonado, Centro de Estudios Avanzados en Zonas Aridas, ChileColin Long, University of Wisconsin–Oshkosh, United States
Copyright © 2018 Iglesias, Haberle, Holz and Whitlock. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Virginia Iglesias, dmlyZ2luaWEuaWdsZXNpYXNAbXN1Lm1vbnRhbmEuZWR1