 Yussuf A. Wato1,2†
Yussuf A. Wato1,2† Herbert H. T. Prins1*
Herbert H. T. Prins1* Ignas M. A. Heitkönig1
Ignas M. A. Heitkönig1 Geoffrey M. Wahungu3Shadrack M. Ngene2Steve Njumbi4
Geoffrey M. Wahungu3Shadrack M. Ngene2Steve Njumbi4 Frank van Langevelde1,5
Frank van Langevelde1,5- 1Resource Ecology Group, Wageningen University, Wageningen, Netherlands
- 2Kenya Wildlife Service, Nairobi, Kenya
- 3National Environment Management Authority, Nairobi, Kenya
- 4International Fund for Animal Welfare, East Africa, Nairobi, Kenya
- 5School of Life Sciences, Westville Campus, University of KwaZulu-Natal, Durban, South Africa
Water is a scarce resource in semi-arid savannas where over half of the African elephants (Loxodonta africana) populations occur and may therefore influence their movement pattern. A random search is expected for an animal with no information on the location of the target resource, else, a direction-oriented walk is expected. We hypothesized that elephants movement patterns show a stronger directional orientation toward water sources in the dry season compared to the wet season. We investigated the movement paths of four male and four female elephants with hourly GPS fixes in Tsavo National Park, Kenya in 2012–2013. Consistent with our predictions, the movement paths of elephants had longer step lengths, longer squared net displacements, and were directed toward water sources in the dry season as compared to the wet season. We argue that African elephants know the location of dispersed water resources, enabling them to survive with scarce resources in dry savannas. These results can be used in conservation and management of wildlife, through for instance, protection of preferred water sources.
Introduction
The movement paths of animals represent behavioral and ecological processes, such as navigation, migration, dispersal, and food searching (Benhamou, 2004) and the distribution of the resources (de Jager et al., 2014). For instance, the movement strategies used by an animal when foraging in a landscape with dispersed resources would be different from those of animals foraging in an area with clustered resources (Bartumeus, 2009). It is generally hypothesized that animals increase tortuosity of their movement paths in areas with high resource density (Bartumeus et al., 2005; Hengeveld, 2007; Bartumeus, 2009). Consequently, the squared net displacement of the animal decreases and the time spent in utilizing these resources increases, leading to efficient resource use (Turchin, 1991). On the other hand, straight and less tortuous movement paths with high net displacement are more efficient in landscapes with dispersed resources (Turchin, 1998; De Knegt et al., 2007; Roshier et al., 2008). Therefore, analysis of the movement paths of animals could give an understanding of the relationship between the resource distribution and foraging efficiency.
Previous studies on movement path analyses were mostly carried out on insects, birds, and small mammals (Turchin, 1991; Viswanathan et al., 1996; Atkinson et al., 2002). However, recent advances in radio-telemetry have made it possible to collect vast quantities of movement data in space and time for both large terrestrial and marine mammals (Austin et al., 2004; Boyce et al., 2010; Sims et al., 2012). Although the movement parameters to be measured vary with the objectives of the study (Marsh and Jones, 1988), generally, parameters such as the distance covered between successive relocations, the turn angles, the directionality of the track and the relationship of the track with properties of the environment that the animal passes through, form the basis of movement path analysis (Root and Kareiva, 1984; Marsh and Jones, 1988; Hengeveld, 2007; Dray et al., 2010; de Knegt et al., 2011; Calenge, 2015; Kölzsch et al., 2015). These movement patterns may in turn determine the frequency with which the animal will encounter the object of interest, which may be food, water, mates, or escape from predation (Marsh and Jones, 1988). To increase resource use efficiency, a random search is expected for a forager with no information on the location of the target resource, whereas a more direction-oriented ballistic walk is expected for a forager with information on the target resource (Valeix et al., 2010). Knowledge on how animals move in their environment can give critical insight on animal's behavior that may be used in the effective management and conservation of species under study.
Water is a scarce resource in semi-arid savanna, where over half of the African elephant (Loxodonta africana) populations occur, and may therefore influence the movement strategies used by elephants. Elephants are water dependent and they usually have to drink water every two to three days (Stokke and Du Toit, 2002; Redfern et al., 2005; Smit et al., 2007). To survive in dry savannas, it is therefore critical for elephants to be able to efficiently find the sparsely distributed water sources, especially during the dry season. Based on the elephant's water requirements and the scarcity of water during the dry season, we expect that the movement pattern of the elephant will reflect these seasonal contrasts in water distribution. Although it is not in doubt that the distance to water is a primary environmental factor influencing habitat use by elephants (Verlinden and Gavor, 1998; Chamaillé-Jammes et al., 2007; Smit et al., 2007; Hilbers et al., 2015; Wato et al., 2016; Sianga et al., 2017), it remains unclear how the behavioral responses of elephants change as a result of water scarcity (Polansky et al., 2015). Here, we analyse the movement paths of four male and four female elephants to address the hypothesis that elephant movement patterns show a stronger directional orientation toward water sources in the dry season compared to the wet season. We predict that the movement path for male and female elephants are less tortuous, have longer step lengths, longer net displacements and smaller turning angles and will show stronger directionality toward water sources in the dry season than in the wet season. Past reports indicate that elephants remember and re-visit previously visited sites (De Beer and Van Aarde, 2008; Prins and Van Langevelde, 2008; de Knegt et al., 2011; Polansky et al., 2015) and pass on the information of their historical migration routes through generations (McComb et al., 2001; Moss et al., 2011). Thus, longer step lengths and higher directionality of elephant movement paths toward water sources in the dry season is an indication that elephants use information to travel to these water sources.
Materials and Methods
Study Area
We conducted this study in the Tsavo Conservation Area in Kenya, a semi-arid ecosystem spanning an area of ~48,300 km2, located at 2°-4° S and 37.5°-39.5° E in the southern part of Kenya (Omondi et al., 2008; Ngene et al., 2012). The area is characterized by a bi-modal rainfall with long rains in mid-March to May, short rains in November to December (Tyrrell and Coe, 1974) and a mean annual rainfall of 250–500 mm (Tyrrell and Coe, 1974; Prins and Loth, 1988). The two rainfall seasons are separated by a 5 months long dry season typically ranging from June through October (Tyrrell and Coe, 1974; Leuthold and Leuthold, 1978; Omondi et al., 2008). There are two permanent rivers in Tsavo (Galana river and Tsavo river) and several seasonal rivers, with Voi and Tiva rivers flowing for a short time in the rainy season (Ayeni, 1975). Other sources of water are the numerous natural waterholes which fill up with water during the rainy season. Some of these waterholes can hold water throughout the short dry season (January-March) but all the natural waterholes dry up around July-August during the long dry season (June to October) (Ayeni, 1975; Mukeka, 2010). In addition, there are three wind-pumped boreholes and a few water reservoirs located around tourist facilities and community owned ranches with constant water supply for animals in the peak of the dry months.
Elephants GPS Data
We used radio-telemetry data from GPS-collared elephants in the Tsavo Conservation Area to investigate the differences in elephants movement patterns between the wet and dry season. During the wet season, there is abundant water for wildlife in the Tsavo ecosystem (Omondi et al., 2008; Mukeka, 2010). However, this area has sparsely distributed permanent water sources in the dry season when the only available water sources for wildlife are reduced to two perennial rivers, three boreholes, and a few water pools constantly refilled by the hoteliers and neighboring community ranchers (Ayeni, 1975). Water has been identified as key resource that affects elephants distribution and their spatial habitat use (Chamaillé-Jammes et al., 2007; Harris et al., 2008; Smit and Grant, 2009; Polansky et al., 2015). For instance, in drier environments, elephants take an average interval of 3 days to drink water and the duration of re-visiting water points differ between sexes (Stokke and Du Toit, 2002), with bull elephants drinking every 3–5 days while breeding herds every 2–4 days (Viljoen, 1989; Leggett, 2006). Furthermore, the breeding herds have been reported to forage close to water sources in the dry season compared to the male elephants (Harris et al., 2008; Sianga et al., 2017).
We monitored four female and four male elephants fitted with satellite-linked GPS collars between March 2012 and June 2013 in the Tsavo Conservation Area in Kenya (Figure 1). The individuals that were collared were randomly selected from five sectors in the Tsavo Conservation Area to represent elephant movement patterns across the entire park. The procedure for fitting GPS collars are described in Ngene et al. (2012). The GPS collars transmitted hourly fixes and the data were automatically transmitted to a web-linked database at the Tsavo East Research Station in Kenya. The GPS had an error of ~10 meters for relocation fixes and some hours had missing values caused by obstruction of signals by, for instance, heavy cloud cover or dense tree canopies (Hebblewhite et al., 2007). In our analysis we considered only the successive time steps with GPS fixes.
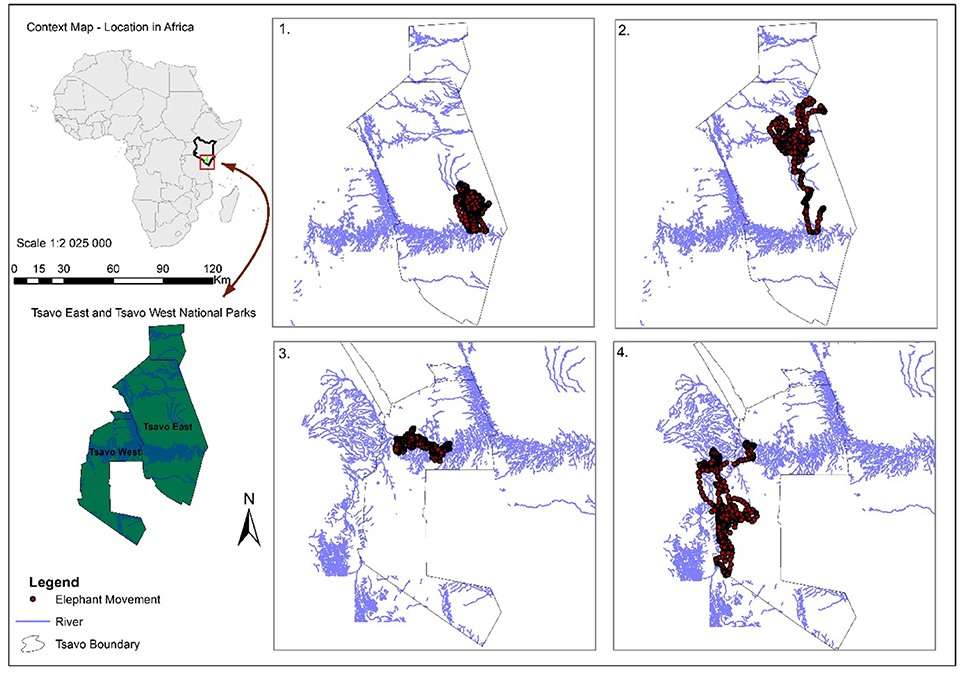
Figure 1. Study areas and the hourly GPS fixes (black dots) for a male elephants in the dry (1) and the wet season (2); and for a female elephants in the dry (3) and the wet season (4). In both sexes the individuals' movement paths seem to be tightly anchored on a river in the dry season.
We analyzed elephant movement patterns for the males and the females in two seasons: the long dry season (June to October 2012) and the long wet season (March to May 2013). These two seasons are distinctly different in the amount of rainfall and would therefore show the relationship between the change in movement pattern related to water availability.
Data Analysis
We calculated the distance covered by each elephant per hour based on the hourly GPS-fixes. We recorded the distance between successive hours to represent a single movement path (i.e., step length) based on the methods described by Root and Kareiva (1984), Marsh and Jones (1988), Turchin (1998), and Hengeveld (2007). We calculated the turn angle as a measure of the change of direction between successive steps with a zero degrees turn corresponding to locomotion on a straight line without change of direction, a negative angle representing a turn to the left and a positive angle representing a turn to the right (Calenge, 2015). We then analyzed the distribution of step lengths, turning angles, and squared net displacement distances (NDD) for both sexes and seasons with AdehabitatLT animal movement analysis package in R (Calenge, 2015). We calculated the parameters of turn angle distributions such as the mean resultant length and the mean direction using CircStats package Version 0.2-4 in R (Lund and Agostinelli, 2015). The mean direction vector represents the mean orientation of the turn angles while the mean resultant length shows the strength of directionality and the concentration of the angles distribution around the mean (R = 0 represents a dispersed turn angles distribution and R = 1 shows that all angles are equal to the mean direction vector) (Lund and Agostinelli, 2015). We only analyzed the movement paths that were directed toward the nearest water source to focus on the effects of water on movement path. Thus, all the movement paths for the elephants that were further than 15 km from the nearest water sources and those for the individuals returning from drinking water were excluded from the analysis on the assumption that they were foraging and not seeking for water. Since the range of water re-visitation frequency for the Tsavo elephants were 1–4 days for the females and 2–5 days for the males in the dry season, we also excluded all the movement paths of the day following an elephant's visitation to the water sources in the dry season. The rest of the movement paths were included in the analysis. In order to establish whether the directionality changed with distance from the water source, or whether proximity had no effect, we also stratified our analysis to 5, 10 and 15 km from the water source. We analyzed the effect of the fixed variables; season, sex and distance from the nearest water source, on elephant's movement pattern using linear mixed effects models (LMMs). We used the ID of the elephant as a random effect variable to account for variation due to individual differences. We also checked for the interaction effects between sex and season in our analyses. We performed these analyses using R package lme4 (Bates et al., 2013).
Results
The step lengths per hour for the elephants were significantly longer in the dry season compared to the wet season (Table 1A). The step lengths changed with distance from the nearest water point, with the step lengths further from the water (15 km) being significantly shorter than those closer to water points (5 and 10 km) (Table 1A). Also, step lengths were longer in the 10 km group than in the 5 km group. Even though both male and female elephants have longer step lengths in the dry season as compared to the wet season, the results showed a significant interaction effect of sex and season (Figure 2A). The male elephants have shorter step lengths than the females in the wet (Table 1A) and a longer step length in the dry season (Figure 2A). Similarly, the squared net displacement for the elephants were significantly longer in the dry season compared to the wet season (Table 1B). The squared net displacements were significantly longer further away from water (15 and 10 km) as compared to distance closer to the water (5 km) (Table 1B). Furthermore, the squared net displacement was also significantly affected by the interaction between sex and season with squared net displacement for the males being longer than the females in the wet season (Table 1B). Both sexes had similarly longer squared net displacements in the dry season (Figure 2B). However, the turn angles for both sexes were large in both the wet and the dry season and did not show any significant difference between the seasons.
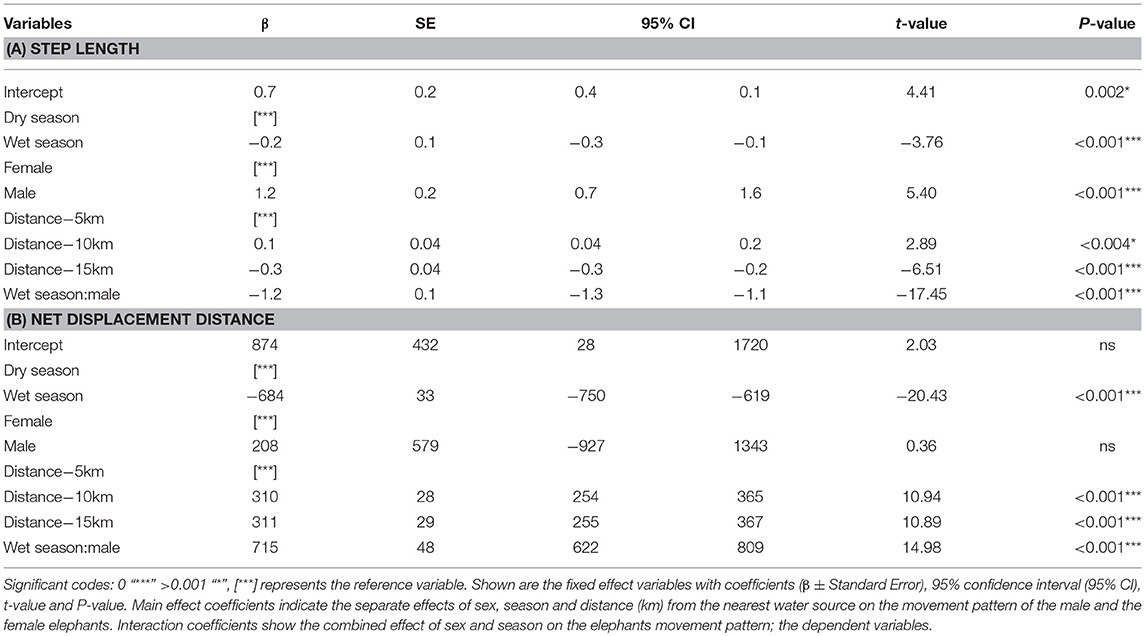
Table 1. Results of the LMM models for step length (A) and net displacement distances (B).
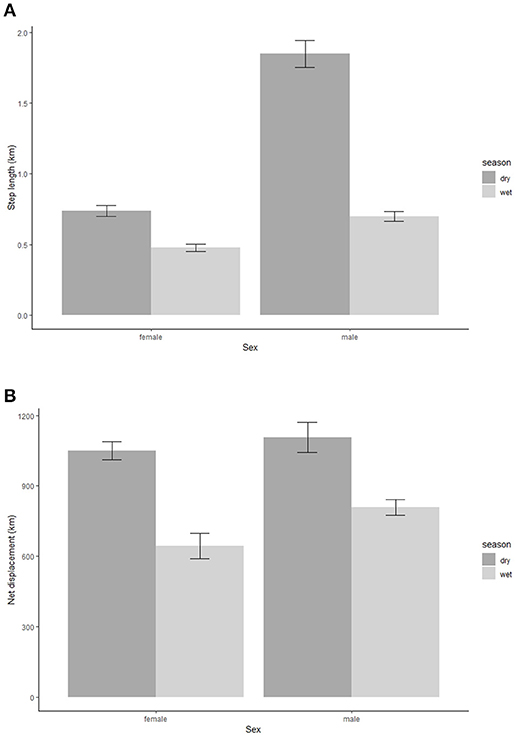
Figure 2. Step length (A) and squared net displacement distances (B) for the female and the male elephants movement paths in the wet (black bars) and the dry (gray bars) season.
The mean resultant length of the turning angles for both sexes showed strong directionality in the dry season compared to the wet season (Figure 3). The resultant mean length of the turning angles for females were in the same range with males in wet season but much lower than the males in the dry season, although not significantly different (Figure 3).
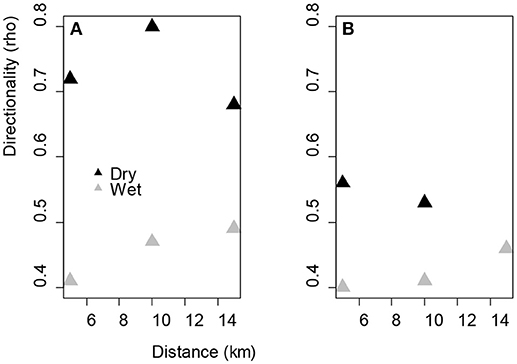
Figure 3. Directionality of the movement patterns at different distances to water sources for (A) male elephants in the wet (Rho at 5 km = 0.41, 10 km = 0.47, 15 km = 0.49) and dry season (Rho at 5 km = 0.72, 10 km = 0.8, 15 km = 0.68) and (B) females in the wet (Rho at 5 km = 0.40, 10 km = 0.41, 15 km = 0.46) and dry season (Rho at 5 km = 0.56, 10 km = 0.53). Rho = 0 represents a dispersed turn angles distribution, hence indicates weak directionality, and R = 1 shows that all angles are equal to the mean direction vector, hence indicates strong directionality. Females were not recorded to be at 15 km from the nearest water point in the dry season.
Discussion
The study of animal movement patterns in relation to resource distribution is one of the novel ways to link behavior of individuals to the spatial distribution of resources (Schick et al., 2008; Giuggioli and Bartumeus, 2010). Resource distribution varies in space and time, and can occur in a spectrum ranging from over-dispersed, random, in patches to highly aggregated clusters (Prins and Van Langevelde, 2008; de Knegt et al., 2011). We examined the role of water distribution on the movement pattern of elephants. In this study, we showed how elephant movement patterns change as a result of seasonal variation in water distribution. The results support our predictions that the movement paths of both male and female elephants are less tortuous, resulting in longer step lengths, and have longer squared net displacements in the dry season compared to the wet season. Furthermore, the mean length of the turning angle showed strong directionality toward water sources for both the sexes in the dry season. The movement paths that were removed were for individuals returning from the water source, those that were beyond 15 km from the nearest water source and the paths for all individuals for the day following their return from drinking water during the dry season, which would not contribute to our conclusion.
The Tsavo Conservation Area is an ecosystem undergoing pronounced scarcity of water and in the long dry season, two perennial rivers and three boreholes serve as the primary water source for wildlife. Most of the area is far away from water for much of the year. Most wildlife species, and particularly elephants, require regular water intake (Stokke and Du Toit, 2002; Redfern et al., 2005) and have to travel between the foraging sites and watering points to meet their energy and water requirements. For instance, in Kruger National Park, elephants drink water every 2 days during the dry season (Young, 1970), and other studies have shown that the duration of water re-visitation is sex-dependent (Viljoen, 1989; Leggett, 2006). The movement paths of elephants are thus expected to be influenced by the water distribution, and the Tsavo elephants appear to have information about the water locations which supports findings in previous studies (Polansky et al., 2015). Therefore, regular re-visitation of watering points may explain the long step lengths and squared net displacements and the strong directionality toward water sources in the dry season (Chamaillé-Jammes et al., 2013; Polansky et al., 2015). The use of information about the location of the water sources is especially apparent when they show this behavior at long distances from the water sources. A few studies have found a relationship between resource distribution and the movement patterns of other wildlife species (Prins, 1996; Loureiro et al., 2007; Valeix et al., 2010). For instance, in a study of lions in arid savannas, their step lengths and squared net displacements were longer as they headed toward waterholes with high aggregation of prey species (Valeix et al., 2010). Similarly, the study of Eurasian badgers showed that their movement paths were less tortuous as they headed toward their dens and latrine sites (Loureiro et al., 2007).
During the wet season, there is abundant food and water for wildlife in the Tsavo ecosystem (Omondi et al., 2008; Mukeka, 2010). In addition to the perennial rivers that flow throughout the year, most of the natural waterholes across Tsavo ecosystem fill up with water during rainy seasons (Tyrrell and Coe, 1974; Ayeni, 1975). This may explain the short step lengths and squared net displacement distances during the wet season. Some of the natural waterholes have water throughout the rainy season and a few of them extend to the short dry season (January-March) (Ayeni, 1975), hence elephants are not water limited then. Similar movement patterns have been reported for foragers in sites of abundant resources. For instance, the movement paths of lions hunting close to a waterhole where there were high prey species congregations, had shorter step lengths and squared net displacement distances and their movement paths were more tortuous than when they were further away from a waterhole (Valeix et al., 2010).
Although water is limiting for both sexes in the dry season, female elephants rarely moved further than 10 km from the nearest water source to forage in the dry season while male elephants accessed forage sites beyond 15 km (Sianga et al., 2017). Furthermore, in the dry season, the directionality of movement path for male elephants was much stronger than the female elephants. This is in line with past studies that reported that breeding herds rarely roam far away from drinking water in drier environments (Viljoen, 1989; Leggett, 2006; Young and Van Aarde, 2010). In these mixed herds, the increased costs associated with moving long distances to far foraging sites may be especially stressful for infants and juveniles (Lee and Moss, 1986; Loveridge et al., 2006) and could lead to increased calf mortality (Loveridge et al., 2006; Foley et al., 2008; Young and Van Aarde, 2010). Our results support the hypothesis that elephants initially seek habitats closer to water in the dry season, regardless of the distribution of food (De Beer et al., 2006; Illius, 2006; Chamaillé-Jammes et al., 2007; Evans and Harris, 2008; Wato et al., 2016). However, if there are many water sources, elephants choose those water sources with more vegetation and avoid those that are not associated with suitable vegetation (Harris et al., 2008).
The difference between male and female elephants movement patterns may be also be explained by their social organization (Archie et al., 2011; Moss et al., 2011) and the difference in foraging strategy between the sexes (Lee et al., 2011). The foraging range of male elephants is larger than that of females as they take more risks and disperse to unfamiliar habitats to seek for food and mates (Lee et al., 2011; Skarpe et al., 2014). This foraging behavior may have advantages such as accessing far foraging grounds and water points in dry seasons (Lee et al., 2011; Lindsay, 2011). Moreover, the mixed herd comprises of individuals of different ages and the group's movement is affected by, for instance, calves that may not be able to move fast and far from water sources like the adult elephants (Ngene et al., 2010). The large herds also spread widely while foraging and, probably, while heading to the water sources to drink. Conversely, the bulls move and forage alone or in a bachelor herd without calves to retard their speed (Ngene et al., 2010). Thus, bulls may travel far to forage but also walk in a less spread formation toward the watering point. The difference in foraging strategies among different sexes are common in other sex-dimorphic species like the red deer (Gordon et al., 1989), moose (Miquelle et al., 1992), and many others (Moss et al., 2011). Generally, the differences in foraging strategies in many species appears to be driven by factors such as energy need requirements, reproductive status of an individual, body sizes and the social context, all of which differ between sexes (Miquelle et al., 1992; Lindsay, 2011).
This study shows that water distribution determines the movement paths of elephants. It supports other studies and models that indicated that animals often adjust their movement pattern in relation to critical and scarce resources (Chamaillé-Jammes et al., 2007; Hengeveld, 2007). Our findings reveal that movement paths of African elephants show strong directionality in dry seasons driven by water distribution. We demonstrate that environmental variables can be used to predict general movement patterns of large herbivores and that the findings can be used in conservation and management of wildlife, through for instance, protection of preferred, or critically needed water sources. The method in this study can be replicated to examine directed movement of other species, of which their cognitive abilities are less well-known.
Ethics Statement
Approval for animal collaring was not required as per the local legislation. Collaring was done for management purposes by the relevant authority (the Kenya Wildlife Service) executed and supervised by the veterinarians of the mentioned authority. Thus, we adhered to all ethical requirements to ensure animal safety.
Author Contributions
YW helped in research design, data collection, analysis and writing. HP, FvL, IH, GW, and SMN in research design, data analysis and writing. SN assisted in data collection and editing of the paper.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
A special thanks to the Kenya Wildlife Service (KWS), International Fund for Animal Welfare (IFAW), Wageningen University and NUFFIC for the financial and technical support for this project. Our gratitude goes to Tsavo research and park management team for assistance in data collection and field logistics. This is a part of the publicly defended Ph.D. thesis of the first author (see Wato et al., 2016).
References
Archie, E. A., Moss, C. J., and Alberts, S. C. (2011). “Friends and relations: kinship and the nature of female elephant social relationships,” in The Amboseli Elephants: A Long-Term Perspective on a Long-Lived Mammal, eds C. J. Moss and H. Croze (Chicago, IL: University of Chicago Press), 238–245.
Atkinson, R. P. D., Rhodes, C. J., Macdonald, D. W., and Anderson, R. M. (2002). Scale-free dynamics in the movement patterns of jackals. Oikos 98, 134–140. doi: 10.1034/j.1600-0706.2002.980114.x
Austin, D., Bowen, W. D., and McMillan Austin, J. I. (2004). Intraspecific variation in movement patterns: modeling individual behaviour in a large marine predator. Oikos 105, 15–30. doi: 10.1111/j.0030-1299.1999.12730.x
Ayeni, J. S. O. (1975). Utilization of waterholes in Tsavo National Park (East). Afr. J. Ecol. 13, 305–323. doi: 10.1111/j.1365-2028.1975.tb00142.x
Bartumeus, F. (2009). Behavioral intermittence, lévy patterns, and randomness in animal movement. Oikos 118, 488–494. doi: 10.1111/j.1600-0706.2009.17313.x
Bartumeus, F., Da Luz, M. G. E., Viswanathan, G. M., and Catalán, J. (2005). Animal search strategies: a quantitative random-walk analysis. Ecology 86, 3078–3087. doi: 10.1890/04-1806
Bates, D., Maechler, M., Bolker, B., Walker, S., Christensen, R. H. B., Singmann, H., et al. (2013). lme4: Linear Mixed-Effects Models Using Eigen and S4. R package version 10–4. Available online at: https://cran.r-project.org/package=lme4
Benhamou, S. (2004). How to reliably estimate the tortuosity of an animal's path: straightness, sinuosity, or fractal dimension? J. Theor. Biol. 229, 209–220. doi: 10.1016/j.jtbi.2004.03.016
Boyce, M. S., Pitt, J., Northrup, J. M., Morehouse, A. T., Knopff, K. H., Cristescu, B., et al. (2010). Temporal autocorrelation functions for movement rates from global positioning system radiotelemetry data. Philos. Trans. R. Soc. B 365, 2213–2219. doi: 10.1098/rstb.2010.0080
Calenge, C. (2015). Analysis of Animal Movements in R: the adehabitatLT Package. Office national de la chasse et de la faune sauvage.
Chamaillé-Jammes, S., Mtare, G., Makuwe, E., and Fritz, H. (2013). African elephants adjust speed in response to surface-water constraint on foraging during the dry-season. PLoS ONE 8:e59164. doi: 10.1371/journal.pone.0059164
Chamaillé-Jammes, S., Valeix, M., and Fritz, H. (2007). Managing heterogeneity in elephant distribution: interactions between elephant population density and surface-water availability. J. Applied Ecol. 44, 625–633. doi: 10.1111/j.1365-2664.2007.01300.x
De Beer, Y., Kilian, W., Versfeld, W., and van Aardea, R. J. (2006). Elephants and low rainfall alter woody vegetation in Etosha National Park, Namibia. J. Arid Environ. 64, 412–421. doi: 10.1016/j.jaridenv.2005.06.015
De Beer, Y., and Van Aarde, R. J. (2008). Do landscape heterogeneity and water distribution explain aspects of elephant home range in southern Africa's arid savannas? Journal of Arid Environ. 72, 2017–2025. doi: 10.1016/j.jaridenv.2008.07.002
de Jager, M., Bartumeus, F., Kölzsch, A., Weissing, F. J., Hengeveld, G. M., Nolet, B. A., et al. (2014). How superdiffusion gets arrested: ecological encounters explain shift from Lévy to Brownian movement. Proc. R. Soc. B. 281:20132605. doi: 10.1098/rspb.2013.2605
De Knegt, H. J., Hengeveld, G. M., van Langevelde, F., de Boer, W. F., and Kirkman, K. P. (2007). Patch density determines movement patterns and foraging efficiency of large herbivores. Behav. Ecol. 18, 1065–1072. doi: 10.1093/beheco/arm080
de Knegt, H. J., van Langevelde, F., Skidmore, A. K., Delsink, A., Slotow, R., Henley, S., et al. (2011). The spatial scaling of habitat selection by African elephants. J. Anim. Ecol. 80, 270–281. doi: 10.1111/j.1365-2656.2010.01764.x
Dray, S., Royer-Carenzi, M., and Calenge, C. (2010). The exploratory analysis of autocorrelation in animal-movement studies. Ecol. Res. 25, 673–681. doi: 10.1007/s11284-010-0701-7
Evans, K. E., and Harris, S. (2008). Adolescence in male African elephants, Loxodonta africana, and the importance of sociality. Anim. Behav. 76, 779–787. doi: 10.1016/j.anbehav.2008.03.019
Foley, C., Pettorelli, N., and Foley, L. (2008). Severe drought and calf survival in elephants. Biol. Lett. 4, 541–544. doi: 10.1098/rsbl.2008.0370
Giuggioli, L., and Bartumeus, F. (2010). Animal movement, search strategies and behavioural ecology: a cross-disciplinary way forward. J. Anim. Ecol. 79, 906–909. doi: 10.1111/j.1365-2656.2010.01682.x
Gordon, I. J., Journal, T., and Apr, N. (1989). Vegetation community selection by ungulates on the Isle of Rhum. J. Appl. Ecol. 26, 65–79. doi: 10.2307/2403650
Harris, G. M., Russell, G. J., van Aarde, R. I., and Pimm, S. L. (2008). Rules of habitat use by elephants Loxodonta africana in southern Africa: insights for regional management. Oryx 42, 66–75. doi: 10.1017/S0030605308000483
Hebblewhite, M., Percy, M., and Merrill, E. H. (2007). Are all global positioning system collars created equal? Correcting habitat-induced bias using three brands in the central canadian rockies. J. Wildl. Manage. 71, 2026–2033. doi: 10.2193/2006-238
Hengeveld, G. M. (2007). Moving to Eat: Animal Foraging Movements in a Heterogeneous Environment. Ph.D. dissertation, Wageningen University, Wageningen, The Netherlands.
Hilbers, J. P., Van Langevelde, F., Prins, H. H., Grant, C. C., Peel, M. J., Coughenour, M. B., et al. (2015). Modeling elephant-mediated cascading effects of water point closure. Ecol. Appl. 25, 402–415. doi: 10.1890/14-0322.1.sm
Illius, A.W. (2006). “Linking functional responses and foraging behaviour to population dynamics,” in Large Herbivore Ecology, Ecosystem Dynamics and Conservation, eds K. Danell, R. Bergström, P. Duncan, and J. Pastor (Cambridge: Cambridge University Press), 71–96.
Kölzsch, A., Alzate, A., Bartumeus, F., de Jager, M., Weerman, E. J., Hengeveld, G. M., et al. (2015). Experimental evidence for inherent Lévy search behaviour in foraging animals. Proc. R. Soc. B. 282:20150424. doi: 10.1098/rspb.2015.0424
Lee, P., Lindsay, W., and Moss, C. (2011). “Ecological patterns of variability in demographic rates” in The Amboseli Elephants, eds C. J. Moss, H. Croze, and C. L. Phyllis (Chicago, IL: University of Chicago Press), 74–88.
Lee, P. C., and Moss, C. J. (1986). Early maternal investment in male and female African elephant calves. Behav. Ecol. Sociobiol. 18, 353–361. doi: 10.1007/BF00299666
Leggett, K. E. A. (2006). Home range and seasonal movement of elephants in the Kunene Region, northwestern Namibia. African Zool. 41, 17–36. doi: 10.1080/15627020.2006.11407332
Leuthold, B. M., and Leuthold, W. (1978). Ecology of the giraffe in Tsavo East National Park, Kenya. Afric. J. Ecol. 16, 1–20. doi: 10.1111/j.1365-2028.1978.tb00419.x
Lindsay, K. (2011). “Habitat use, diet choice and nutritional status in female and male Amboseli elephants,” in The Amboseli Elephants: A Long-Term Perspective on a Long-Lived Mammal, eds C. J. Moss, H. Crozem and P. C. Lee (Chicago, IL; London: University of Chicago Press), 51–73.
Loureiro, F., Rosalino, L. M., Macdonald, D. W., and Santos-Reis, M. (2007). Path tortuosity of Eurasian badgers (Meles meles) in a heterogeneous Mediterranean landscape. Ecol. Res. 22, 837–844. doi: 10.1007/s11284-006-0325-0
Loveridge, A. J., Hunt, J., Murindagomo, F., and Macdonald, D. (2006). Influence of drought on predation of elephant (Loxodonta africana) calves by lions (Panthera leo) in an African wooded savannah. J. Zool. 270, 523–530. doi: 10.1111/j.1469-7998.2006.00181.x
Lund, U., and Agostinelli, C. (2015). Circstats: Circular Statistics, From “Topics in Circular Statistics”(2001). S-plus original by Lund, U. R port by Agostinelli, C. R package version 0.2–4. Available online at: http://CRAN.R-project.org/package=CircStats
Marsh, L. M., and Jones, R. E. (1988). The form and consequences of random walk movement models. J. Theor. Biol. 133, 113–131. doi: 10.1016/S0022-5193(88)80028-6
McComb, K., Moss, C., Durant, S. M., Baker, L., and Sayialel, S. (2001). Matriarchs as repositories of social knowledge in African elephants. Science 292, 491–494. doi: 10.1126/science.1057895
Miquelle, D. G., Peek, J. M., and Ballenberghe, V. (1992). Sexual segregation in Alaskan moose. Wildlife Monogr. 122, 3–57. doi: 10.1017/CBO9781107415324.004
Moss, C. J., Croze, H., and Lee, P. C. (2011). The Amboseli Elephants: A Long-Term Perspective on a Long-Lived Mammal. Chicago, IL: Oxford University Press.
Mukeka, J. M. (2010). Analyzing the Distribution of the African Elephant (Loxodonta africana) in Tsavo, Kenya. Masters thesis, Miami University.
Ngene, S. M., Njumbi, S., Ngoru, B., Nzisa, M., Bitok, E., Poghon, J., et al. (2012). “Satellite-linked GPS collars deployed on elephants in Tsavo Ecosystem, Kenya,” in Recent Advances in Satellite Research and Development, eds S. Gardiner, and K. P. Olsen (New York, NY: Nova Publishers).
Ngene, S. M., van Gils, H. A. M. J., van Wieren, S. E., Rasmussen, H., Skidmore, A. K., Prins, H. H. T., et al. (2010). The ranging patterns of elephants in Marsabit protected area, Kenya: the use of satellite-linked GPS collars. Afric. J. Ecol. 48, 386–400. doi: 10.1111/j.1365-2028.2009.01125.x
Omondi, P., Bitok, E., Mukeka, J., Mayienda, R., and Litoroh, M. (2008). Total Aerial Count of Elephants and Other Large Mammal Species of Tsavo–Mkomazi Ecosystem'. Kenya Wildlife Service Report.
Polansky, L., Kilian, W., Wittemyer, G., Polansky, L., Kilian, W., and Wittemyer, G. (2015). Elucidating the significance of spatial memory on movement decisions by African savannah elephants using state–space models. Proc. R. Soc. B. 282:20143042. doi: 10.1098/rspb.2014.3042
Prins, H. H. T. (1996). Ecology and Behaviour of the African Buffalo: Social Inequality and Decision Making. London: Chapman & Hall.
Prins, H. H. T., and Loth, P. E. (1988). Rainfall patterns as background to plant phenology in northern Tanzania. J. Biogeogr. 15, 451–463. doi: 10.2307/2845275
Prins, H. H. T., and Van Langevelde, F. (2008). Resource Ecology: Spatial and Temporal Dynamics of Foraging. Berlin: Springer.
Redfern, J. V., Grant, C. C., Gaylard, A., and Getza, W. M. (2005). Surface water availability and the management of herbivore distributions in an African savanna ecosystem. J. Arid Environ. 63, 406–424. doi: 10.1016/j.jaridenv.2005.03.016
Root, R. B., and Kareiva, P. M. (1984). (Pieris Rapae): ecological consequences and adaptive significance of markovian movements in a patchy environment. Ecology 65, 147–165.
Roshier, D. A., Doerr, V. A., and Doerr, E. D. (2008). Animal movement in dynamic landscapes: Interaction between behavioural strategies and resource distributions. Oecologia 156, 465–477. doi: 10.1007/s00442-008-0987-0
Schick, R. S., Loarie, S. R., Colchero, F., Best, B. D., Boustany, A., Conde, D. A., et al. (2008). Understanding movement data and movement processes: current and emerging directions. Ecol. Lett. 11, 1338–1350. doi: 10.1111/j.1461-0248.2008.01249.x
Sianga, K., van Telgen, M., Vrooman, J., Fynn, R. W. S., and van Langevelde, F. (2017). Spatial refuges buffer landscapes against homogenization and degradation by large herbivore populations and facilitate vegetation heterogeneity. Koedoe 59:a1434. doi: 10.4102/koedoe.v59i2.1434
Sims, D. W., Humphries, N. E., Bradford, R. W., and Bruce, B. D. (2012). Lévy flight and Brownian search patterns of a free-ranging predator reflect different prey field characteristics. J. Ecol. 81, 432–442. doi: 10.1111/j.1365-2656.2011.01914.x
Skarpe, C., Du Toit, J. T., and Moe, S. R. (2014). Elephants and Savanna Woodland Ecosystems: A Study From Chobe National Park, Botswana. West Sussex, UK: John Wiley & Sons.
Smit, I. P. J., and Grant, C. C. (2009). Managing surface-water in a large semi-arid savanna park: effects on grazer distribution patterns. J. Nature Conserv. 17, 61–71. doi: 10.1016/j.jnc.2009.01.001
Smit, I. P. J., Grant, C. C., and Devereux, B. J. (2007). Do artificial waterholes influence the way herbivores use the landscape? Herbivore distribution patterns around rivers and artificial surface water sources in a large African savanna park. Biol. Conserv. 136, 85–99. doi: 10.1016/j.biocon.2006.11.009
Stokke, S., and Du Toit, J. T. (2002). Sexual segregation in habitat use by elephants in Chobe National Park, Botswana. Afric. J. Ecol. 40, 360–371. doi: 10.1046/j.1365-2028.2002.00395.x
Turchin, P. (1991). Translating foraging movements in heterogeneous environments into the spatial distribution of foragers. Ecology 72, 1253–1266. doi: 10.2307/1941099
Turchin, P. (1998). Quantitative Analysis of Movement: Measuring and Modeling Population Redistribution in Animals and Plants. Sunderland: Sinauer Associates.
Tyrrell, J., and Coe, M. (1974). The rainfall regime of Tsavo National Park, Kenya and its potential phenological significance. J. Biogeogr. 1, 187–192.
Valeix, M., Loveridge, A. J., Davidson, Z., Madzikanda, H., Fritz, H., and Macdonald, D. W. (2010). How key habitat features influence large terrestrial carnivore movements: waterholes and African lions in a semi-arid savanna of north-western Zimbabwe. Landscape Ecol. 25, 337–351. doi: 10.1007/s10980-009-9425-x
Verlinden, A., and Gavor, I. K. (1998). Satellite tracking of elephants in northern Botswana. Afric. J. Ecol. 36, 105–116. doi: 10.1046/j.1365-2028.1998.00118.x
Viljoen, P. (1989). Habitat selection and preferred food plants of a desert-dwelling elephant population in the northern Namib Desert, South West Africa/Namibia. Afric. J. Ecol. 27, 227–240.
Viswanathan, G. M., Afanasyev, V., Buldyrev, S. V., Murphy, E. J., Prince, P. A., and Stanley, H. E. (1996). Lévy flight search patterns of wandering albatrosses. Nature 381, 413–441. doi: 10.1038/381413a0
Wato, Y. A., Heitkönig, I. M. A., van Wieren, S. E., Wahungu, G., Prins, H. H. T., and van Langeveldea, F. (2016). Prolonged drought results in starvation of African elephant (Loxodonta africana). Biol. Conserv. 203, 89–96. doi: 10.1016/j.biocon.2016.09.007
Young, E. (1970). Water as a Factor in the Ecology of Game in the Kruger National Park. PhD Thesis. University of Pretoria.
Keywords: Tsavo, wildlife, movement, step length, savanna, directionality, conservation
Citation: Wato YA, Prins HHT, Heitkönig IMA, Wahungu GM, Ngene SM, Njumbi S and van Langevelde F (2018) Movement Patterns of African Elephants (Loxodonta africana) in a Semi-arid Savanna Suggest That They Have Information on the Location of Dispersed Water Sources. Front. Ecol. Evol. 6:167. doi: 10.3389/fevo.2018.00167
Received: 12 August 2018; Accepted: 03 October 2018;
Published: 25 October 2018.
Edited by:
Thomas Wassmer, Siena Heights University, United StatesReviewed by:
Sarah-Anne Jeanetta Selier, South African National Biodiversity Institute, South AfricaMonique De Jager, Utrecht University, Netherlands
Copyright © 2018 Wato, Prins, Heitkönig, Wahungu, Ngene, Njumbi and van Langevelde. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Herbert H. T. Prins, aGVyYmVydC5wcmluc0B3dXIubmw=
†Present Address: Yussuf A. Wato, World Wide Fund for Nature, Nairobi, Kenya