 Camilo Carneiro
Camilo Carneiro Tómas G. Gunnarsson2
Tómas G. Gunnarsson2 José A. Alves
José A. Alves- 1Department Biology and CESAM, University of Aveiro, Aveiro, Portugal
- 2South Iceland Research Centre, University of Iceland, Laugarvatn, Iceland
The timing of annual events is key for organisms that exploit seasonal resources, as deviations from optimal timing might result in considerable fitness costs. Under strong time selection, individuals likely have fewer suitable strategies available than when selection is more relaxed, hence both consistency and flexibility might be advantageous depending on the life history or annual cycle stage. For migrants using both the arctic and the tropics during their annual cycle, the faster warming at higher latitudes than elsewhere in the range may lead to mismatches with local environmental conditions. Additionally, while individuals might already be limited in responding to changes at each stage, the potential degree of a given response will likely also be limited by responses at previous stages of the annual cycle. Contrary to other migratory waders breeding in Iceland, Icelandic whimbrels Numenius phaeopus islandicus have not changed arrival dates during the past 30 years, suggesting high individual consistency in spring arrival timing and a potential limitation in responding to a changing environment. After repeatedly tracking 12 individual Icelandic whimbrels at least twice throughout their annual cycle between 2012 and 2018, we investigated individual consistency of spring arrival date and other annual stages and migration strategy, and explored differences between sexes and seasons. Individuals were more consistent on timing of spring than autumn migration, and the most consistent stage was departure from the wintering sites. Timing of laying was the stage that varied the most, and no overall significant difference between sexes was observed, except on spring stopover duration. While lower consistency in laying dates might allow individuals to track the advancement of spring, consistency at departure from the wintering sites, stopover duration, and arrival into Iceland might limit the degree of advancement. Transgenerational changes in the migratory behavior of other wader species allows population level responses to a changing phenology, but seems unlikely for Icelandic whimbrels, given the stable dates of spring arrival in this population. Under continuing advancement of spring onset, it is thus important to acquire information on the timing of spring arrival of recruits and on the ontogeny of migration to understand how migratory schedules are defined and might influence responses of long-distance migrants to environmental change.
Introduction
The annual cycle of many animals comprises migratory periods during which individuals travel to exploit seasonally available resources (Newton, 2007). Timing of specific events, such as breeding, is therefore fundamental (Alerstam and Lindström, 1990), but selection for optimal timing of events might not be equally strong between different stages of the annual cycle. Due to the influence that timing of breeding can have on breeding success and thus individual fitness (Perrins, 1970; Drent, 2006), selection on timing during the preceding spring migration is expected to be stronger than in autumn (McNamara et al., 1998). Under strong time selection, the available strategies (e.g., schedules) for individuals are likely to be fewer than when selection is more relaxed and individuals can perform a given task (e.g., migration) over a wider time window (Madsen, 2001; Warnock et al., 2004). Therefore, both consistency and flexibility can be advantageous depending on the life history of a given species (Vardanis et al., 2016).
For migrants breeding in arctic and sub-arctic environments, selection on timing of breeding is expected to be strong, given the relatively short time window with favorable environmental conditions for reproduction. However, climate is changing at a faster pace than some species are able to adjust to (Both and Visser, 2001), and the situation can become more complicated in the arctic, where temperatures are increasing more rapidly than at lower latitudes (Høye et al., 2007; Serreze et al., 2009; Cohen et al., 2014). Whereas, individual consistency can differ among distinct annual events (e.g., Conklin et al., 2013, Verhoeven et al., 2019), the performance of individuals at a given stage may also depend on the conditions experienced in previous stages (Harrison et al., 2011; O'Connor et al., 2014). Hence, consistent behavior at one stage might thus limit the degree of change in the subsequent one. Detailed data throughout the annual cycle (e.g., Senner et al., 2014) is therefore required to understand how individuals and populations might be limited in their capacity to respond to a changing environment (Marra et al., 2015).
Previous research has indicated that most species of waders (Charadrii) breeding in Iceland have been advancing their spring arrival dates (e.g., golden plover Pluvialis apricaria, common snipe Gallinago gallinago, black-tailed godwit Limosa limosa), accompanying the trend of temperature change, but Icelandic whimbrels Numenius phaeopus islandicus were an exception (Gunnarsson and Tómasson, 2011, Gill et al., 2014). We update this information by adding 9 years of data of first arrival for birds recorded in South Iceland (see Gunnarsson and Tómasson, 2011 for details) and confirm that arrival dates show no significant trends since 1988 for this population (Figure 1). Whimbrels are long-lived birds (typical lifespan: 11 years; Robinson, 2018) that are long-distance migrants. The Icelandic subspecies breeds predominantly in Iceland, with an estimated population of 256,000 pairs (Skarphéðinsson et al., 2016), and winters in West Africa (Gunnarsson and Guðmundsson, 2016; Carneiro et al., 2019). In spring, their departure from the wintering sites occurs on the second fortnight of April and arrival into Iceland from late April to early May, with individuals performing one of two migratory strategies: a non-stop flight or two flights divided by a relatively short stopover; in autumn, migratory movements occur from late June to late August and only the direct flight strategy has been recorded in this season (Alves et al., 2016, Carneiro et al., 2019). Icelandic whimbrels are site faithful to the breeding territory, monogamous, and most reproduce with the same partner from year to year (BWPi, 2006), with males arriving earlier and departing later from breeding sites than females (Carneiro et al., 2019).
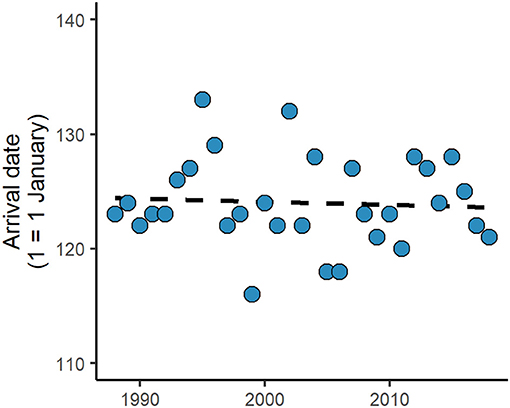
Figure 1. Arrival dates of whimbrels into South Iceland in the 31-year period from 1988 to 2018, updated from Gunnarsson and Tómasson (2011), showing no significant trend (day of the year = −0.027 * year + 178.11, n = 31, R2 = 0.004, p = 0.734).
The low variation of Icelandic whimbrels arrival dates into Iceland and the lack of a population advancement (Figure 1) in response to increasing temperatures may suggest consistency of individual spring arrival dates. Individual consistency in this population is currently unknown, but, at the same time, important to unravel in order to understand potential limitations to population-level responses. Hence, we quantified (1) individual timing consistency in arrival dates to Iceland, and at five other annual stages (departure and arrival during autumn migration, departure of spring migration, stopover duration in spring, and laying date) in order to investigate possible constraints throughout the year; (2) individual consistency in migratory strategy; and tested for (3) seasonal and sex differences in timing consistency. Along with a lack of population variation in spring arrival date, and a higher pressure for breeding timing than for arrival into the wintering sites, we expected higher consistency during spring than autumn migration, but lower consistency in laying dates than during other stages, as this also depends on the schedule of the partner and on their return. For spring migration, consistency might be higher at departure because conditions are likely to be more stable at the wintering sites than at breeding areas, where weather conditions are more variable (as stochastic weather events can reduce food availability and lead to mortality; Vepsäläinen, 1969, Marcström et al., 1979) and the occurrence of a stopover during migration might also influence arrival. However, during autumn, consistency should not vary from departure to arrival due to the direct strategy. Because males arrive earlier than females to defend territories and also attend broods longer (BWPi, 2006), we anticipated that males should show higher consistency on spring migration and less so at autumn departure than females.
Methods
Fieldwork was carried out on whimbrels at breeding grounds in the southern lowlands of Iceland (63.8°N; 20.2°W), between 2012 and 2018. Nests were searched for and upon finding, the incubation stage estimated through egg floatation (Liebezeit et al., 2007), and the laying date was back calculated from stage of incubation. Nests were monitored until hatching and for those that hatched, laying date was back calculated from the hatching date considering an incubation period of 25 days after the last egg laid (mean ± se: 24.8 ± 0.2 days, n = 24 nests found when laying, and hatch recorded). Because whimbrels were individually marked (see below), we were able to identify replacement clutches which were not included in the analyses.
Two hundred and twenty-six adult birds were caught on the nest, using a nest trap (Moudry TR60; www.moudry.cz). Birds were individually marked with a unique metal ring, issued by the Icelandic ringing scheme, and a combination of color rings. Geolocators were fitted on a leg flag to a subgroup of 86 individuals (number of geolocators deployed per year-2012: 10; 2013: 3; 2014: 10; 2015: 30; 2016: 40; 2017: 40). The device was replaced whenever possible each breeding season. Sixty-two devices were retrieved one or more years later (number of geolocators retrieved per year-2013: 5; 2014: 4; 2015: 5; 2016: 14; 2017: 20; 2018: 14). For tags retrieved two or more years later, data on two autumn migrations were recorded. We used the Intigeo-W65A9RJ model from 2012 to 2014 and Intigeo-C65 in the following years (Migrate Technology Ltd.). One device stopped logging in mid-winter, another shortly after departure from Iceland, and a third one was damaged and contained no data. Sixty five individuals were sexed using biometrics following Katrínardóttir et al. (2013), 22 molecularly (as in Katrínardóttir et al., 2013), eight through behavioral observation (copulating position, assuming males on top) and two remained undetermined. Geolocator data analysis and determination of individual departure and arrival timings were performed using light, and temperature, conductivity and wet contacts as described in Carneiro et al. (2019). Due to the accuracy of geolocators (Phillips et al., 2004), we considered arrival and departures to/from the general area. For example, spring arrival was arrival into Iceland instead of arrival into the breeding territory (although some individuals have been observed on the breeding territory on the day of arrival into Iceland). We consider stopover any stop during travel between breeding and wintering locations, irrespective of length of stay, site quality or previous or future flight distance and duration (i.e., we do not discriminate from staging; Warnock, 2010). Although stops of few hours may be undetected with geolocator data, stopovers of Icelandic whimbrels are usually of several days (Carneiro et al., 2019), with the minimum stopover duration recorded during this study being 6 days.
Icelandic whimbrels show two migratory strategies in spring: a direct non-stop flight or two flights with a stopover in between (henceforward: “direct” and “stopover”; Carneiro et al., 2019). However, and adding to previous information, during this study one individual was recorded undertaking a stopover during autumn migration. To understand individual consistency in migratory strategy we calculated the percentage of individuals that changed strategy during the tracking period and the direction of change (i.e., from direct to stopover, from stopover to direct, or both).
Repeatability (R) was estimated in a mixed effects model framework, using 1,000 bootstrap iterations to estimate the confidence intervals, with R package rptR (Stoffel et al., 2017). Given that R takes into account both within- and between-individual variances, it does not translate into absolute consistency (see Conklin et al., 2013), and therefore we also calculated the mean individual range (difference between the latest and earliest record for each individual for each stage, in days) and the absolute interannual difference (absolute difference between consecutive years for each individual within stage, in days) in order to better evaluate individual consistency. To test for differences in consistency between stages and sexes, we fit a generalized linear model with absolute interannual difference as the dependent variable and stage, sex, and their interaction as explanatory variables (with family = Poisson due to the positively skewed dependent variable). In this analysis we did not account for the dependency of member of the same pair, because we could have only used those individuals that nested with the same partner in consecutive years, which would result in a reduced sample size. Data were analyzed in R Core Team (2018) and results are shown as mean ± se.
Results
We recorded individual level data spanning 2–7 years, with a median of 3 years for autumn migration (n = 16 individuals), and 2 years for spring migration (n = 12 individuals), and for laying date (n = 70 individuals). Hence, autumn migration, spring migration, and laying date were recorded during a median of 27, 18, and 18%, and up to 54, 46 and 64%, of whimbrels typical lifespan (Robinson, 2018), respectively. During the present study, one individual was recorded making a stopover during autumn for the first time. In that season, one individual (out of 16) switched strategy between years, from a stopover to a direct one (Table 1). A change in the opposite direction, from direct to stopover in the following year, was observed in a higher proportion during spring migration (three out of 12 individuals; Table 1). No individual was observed changing strategy in both directions (Table 1).

Table 1. Direction and proportion of individuals that changed migratory strategy on each season (Autumn and Spring), from a stopover to direct, direct to stopover or on both directions; n = number of individuals.
Along the annual cycle, Icelandic whimbrels showed the highest consistency of timing at spring departure (Figures 2, 3 and Table 2). Despite low R values for spring arrival, individuals showed relatively smaller mean individual ranges (Table 2) and small absolute interannual differences (Figure 3 and Table 3) than at autumn migration stages. Such low R values result from a relatively low variance among individuals compared to the variance within (Figure 2), which arises from the change in migratory strategies. Spring stopover duration was relatively consistent, with low mean individual range and low absolute interannual differences (Table 1 and Figure 3). During autumn migration, both departure, and arrival timings showed similar consistency (Figures 2, 3 and Table 2), as in this season individuals seldom change strategy and almost always perform a direct flight (Table 1). However, contrary to autumn departure, autumn arrival absolute interannual difference was not statistically different from laying dates (Table 3). Laying date was the least consistent stage of the annual cycle (Figures 2, 3 and Table 2). When considering the absolute interannual difference, we found no overall difference between sexes (Table 3), but males showed a lower absolute interannual difference of stopover duration (Figure 3 and Table 2), and a mean individual range in autumn ca. 2.5 days longer than females (Table 2).
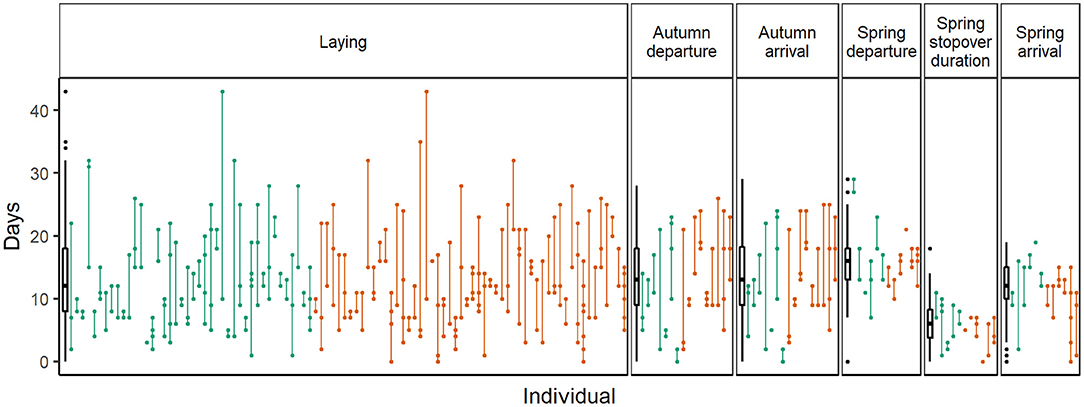
Figure 2. Population and individual interannual variation at each stage of the annual cycle for female (green) and male (orange) Icelandic whimbrels. Population data are summarized by the boxplot on the left of each panel (showing the median and 25 and 75% quartiles, whiskers extending up to 1.5 times the inter quartile range from the median and points beyond that are individually marked) and aims to serve as a reference to visually compare individual variation. Each data point shows the timing or duration of the event in a given year and all observations of the same individual are linked with a vertical line; dates were standardized by subtracting the minimum value observed for each stage at the population level. Note that at spring departure one individual left the wintering area considerably early (shown as an outlier), creating and apparent large variation at this stage if only the boxplot (population) is considered; the same individual then performed the longest stopover recorded (upper outlier). Unfortunately, we do not have repeated tracks of this individual to explore consistency in these stages, as it did not return to the breeding sites after the geolocator was replaced.
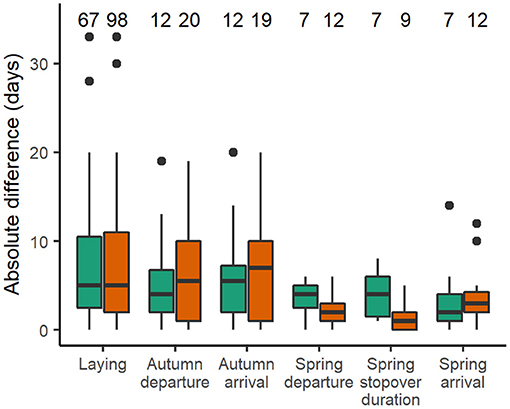
Figure 3. Absolute interannual difference in timing for each annual stage, grouped by sex (females in green, males in orange); boxes show the median and 25 and 75% quartiles, whiskers extend up to 1.5 times the inter quartile range from the median and points beyond that are individually marked; the number above each boxplot shows the sample size.
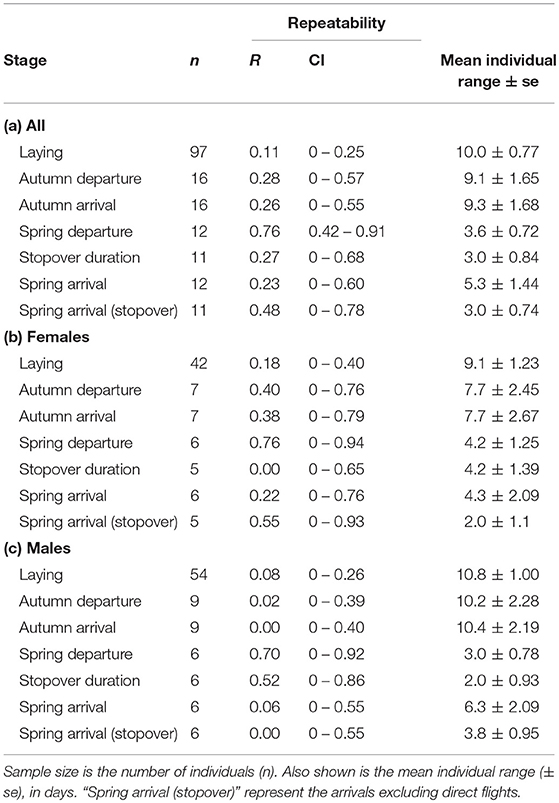
Table 2. Repeatability (R) with 95% confidence intervals (CI) of timing of annual events and spring stopover duration, for all individuals (a) and by sex (b and c).
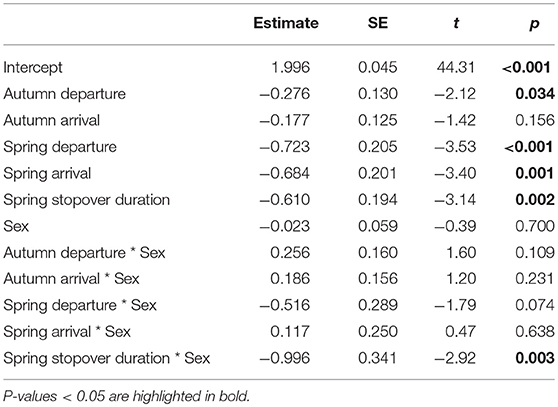
Table 3. Results of the general linear model testing the effects of stage and sex on absolute interannual differences; estimates for stages are in relation to laying and for sex it is of male in relation to female.
Discussion
By tracking individuals over multiple years and over a considerable part of their lifespan, it is possible to quantify relevant levels of individual consistency or flexibility regarding the phenology of important events during the annual cycle. Both consistency and flexibility can be advantageous depending on the life history of each species (Vardanis et al., 2016), and can be essential to understand the capacity and rate of population responses to changing environments (Gill et al., 2014). The arrival dates of Icelandic whimbrels in spring have been stable over the past 30 years (Figure 1; Gunnarsson and Tómasson, 2011), despite a spring advancement of temperatures that drives resources for waders locally (Alves et al., 2019). At the individual level, we show that Icelandic whimbrels were more consistent in timing of spring than autumn migration, and most consistent at departure from the wintering sites. Timing of laying was the stage of the annual cycle that varied the most and no overall significant difference between sexes was observed, except for males lower absolute interannual difference of stopover duration.
During autumn migration, a lower consistency in timing was observed compared to spring (Tables 2, 3 and Figures 2, 3) and the values at arrival mirror the ones at departure because in nearly all occasions individuals flew directly from Iceland to the wintering sites (Table 1; Carneiro et al., 2019). On the other side of the Atlantic, Hudsonian whimbrels showed the same general pattern, as inter-individual variation was greater at autumn departure and arrival dates, than at spring departure and arrival dates (Johnson et al., 2016). Given the expected stable conditions in the wintering area, one could anticipate repeatability to be high during autumn migration (Nussey et al., 2005). In fact, despite a lower consistency in relation to spring, Icelandic whimbrels are still reasonably consistent in autumn migration timings, with a median individual departure range of 9 days and median absolute interannual difference of 5 days, which are in line, or even lower than those observed on other long distance migratory birds (median range of departure of ca. 15 days in individual continental black-tailed godwits L. l. limosa, Verhoeven et al., 2019; median absolute interannual difference of ca. 4, 5, and 6 days in bar-tailed godwits L. lapponica, marbled godwits L. fedoa and red-backed shrikes Lanius collurio, respectively, Conklin et al., 2013, Pedersen et al., 2018, Ruthrauff et al., 2019; mean absolute interannual difference of 12.9 in great reed warblers Acrocephalus arundinaceus, Hasselquist et al., 2017). The observed variation in departure dates from Iceland is likely explained by the prior variation in laying dates and breeding success, since successful breeders tend to depart later (pers. obs.). The mean individual range recorded for males (ca. 2.5 days larger than females; Table 2) is likely due to their longer attendance of broods.
The relative low repeatability of laying date (Table 2 and Figures 2, 3) may be partially explained by the variation in arrival dates into the breeding sites and partner arrival timing and return. Nevertheless, under a scenario of arctic amplification and spring advancement (Høye et al., 2007; Serreze et al., 2009; Gill et al., 2014; Alves et al., 2019), flexibility on laying dates might be beneficial, allowing individuals to track the local conditions and breed successfully. But the potential advancement of laying might be constrained by previous annual events. While spring arrival dates showed some variability (mean individual range at arrival: 5.3 ± 1.4 days), it was mostly due to variation in migratory strategy (Table 1), with the occurrence of a stopover augmenting variation on arrival date after a consistent departure from the wintering sites (mean individual range at departure: 3.6 ± 0.7 days). When considering only the spring arrival of individuals that had a stopover (which is the common strategy), we find higher consistency of arrival dates (mean individual range at arrival: 3.0 ± 0.7 days; Table 2). Furthermore, stopover duration also shows considerable consistency. Hence, individuals tend to be consistent throughout spring migration, starting at departure, which might limit how much laying dates can vary after arrival. If individuals would advance the departure date from the wintering sites, the capacity of tracking the advancement of resource availability in the breeding sites would be higher. However, climatic conditions in the wintering areas seem to be more stable than at the breeding sites (Høye et al., 2007; Serreze et al., 2009; Cohen et al., 2014), and thus unlikely to trigger individual responses at a sufficient rate that allows individuals to track the changes at the breeding sites. In fact, over the last 30 years the population of Icelandic whimbrels showed a stable spring arrival date (Figure 1), despite the increasing temperatures in the breeding grounds (Gunnarsson and Tómasson, 2011; Alves et al., 2019).
Populations can change migration timing through transgenerational variation in phenology (Gill et al., 2014). Such a mechanism was identified in Icelandic black-tailed godwits (L. l. islandica), that have advanced their arrival date into Iceland and tracked the advancement of spring onset (Gill et al., 2014). Similar to whimbrels, individual godwits are consistent in their timing of spring arrival, but recruits tend to migrate earlier and drive the population timing (Gill et al., 2014). The lack of an advancement in whimbrels population arrival dates into Iceland, together with individual consistency, suggests none, or little, transgenerational changes of migration timing. While black-tailed godwits spend the winter in the temperate region, at a maximum of ca. 3,000 km from Iceland (Alves et al., 2012), Icelandic whimbrels migrate longer distances to winter in the tropical or subtropical region, ca. 6,000 km from the breeding sites (Gunnarsson and Guðmundsson, 2016; Carneiro et al., 2019). By wintering closer to Iceland (Alves et al., 2013), godwits might adjust arrival dates to the local environment (Alves et al., 2012), lay as soon as conditions are adequate and produce young early in the season, that are more likely to recruit and ultimately drive population changes (Alves et al., 2019). Whimbrels, on the other hand, by wintering further and likely with no cue of the environmental conditions in Iceland, might have narrowed the variation in timing of departure to a later date than godwits and other waders breeding in Iceland, which in turn reduces the variation at spring arrival, the time between arrival and laying and, consequently, laying dates, limiting the possibilities for transgenerational changes. The arrival date into the breeding sites can vary considerably with spring migratory strategy, with individuals arriving earlier after a direct strategy consequently having a longer period in Iceland before breeding and be more likely to track the advancement of spring onset. However, a direct strategy in spring is uncommon, and although our data on its variation is limited, no individual changed from a stopover to a direct strategy, suggesting that individuals might not track the advancement of spring onset through a change in migratory strategy.
Whimbrels show no advancement of arrival dates into Iceland while spring onset is advancing (Figure 1; Gunnarsson and Tómasson, 2011; Alves et al., 2019), but there is no indication of a population decline (Skarphéðinsson et al., 2016). In Iceland, whimbrels are one of the latest waders to arrive and have a relatively short breeding period when compared to other species breeding in the same area (Gunnarsson, unpublished data). The period length of available minimum resources for breeding is unknown, but if it is wider than that required for successful breeding by whimbrels, it could be that they have been reproducing within that window even while the environmental conditions are advancing. However, under continuous advancement of spring conditions, consistency might prevent individuals from responding to changes when breeding and resource time windows mismatch. At such a hypothetical point one might observe the recruits performing with a different phenology and allow the population to respond to environmental changes, similarly to Icelandic black-tailed godwits. While monitoring population size, it is thus important to acquire knowledge on resource dynamics at the breeding areas (e.g., does the peak abundance of a given resource influence chick growth and survival?), and on the ontogeny of migration and associated timing (e.g., does hatching date affect the migration timing of recruits?). Links between resources and ontogeny will allow understanding how migratory schedules are defined and forecast population-level responses of long-distance migrants to environmental change.
Data Availability
The datasets generated for this study are available on request to the corresponding author.
Ethics Statement
This study was carried out in accordance with the recommendations of Icelandic Institute of Natural History. The protocol was approved by the Icelandic Institute of Natural History.
Author Contributions
All authors designed and carried the study. CC performed the analysis and lead the writing with substantial discussion and inputs from TG and JA. All authors read and approved the final version.
Funding
This work was funded by RANNIS (Grants: 130412-052 and 152470-052), the University of Iceland Research Fund, and by FCT/MCTES to CESAM (UID/AMB/50017/2019), and individual grants (PD/BD/113534/2015 and SFRH/BPD/91527/2012), through National Funds, and ProPolar.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We are thankful for the logistic support of the Icelandic Soil Conservation Service, particularly to Anne Bau and Jóna Maria; Snæbjörn Pálsson for support with genetic sex determination of birds; Verónica Méndez, Borgný Katrínardóttir, and Edna Correia for fieldwork support, our group members in Iceland and Portugal for fruitful discussions, Kristinn Jónsson for kindly allowing us to work on his land and Gunnar Tómasson for sharing arrival dates for whimbrels. Last, we are thankful to Daniel Ruthrauff, Brett Sandercock, and a reviewer for their suggestions, which helped to improve our manuscript.
References
Alerstam, T., and Lindström, Å. (1990). “Optimal bird migration: the relative importance of time, energy, and safety,” in Bird Migration: Physiology and Ecophysiology, ed E. Gwinner (Berlin, Heidelberg: Springer Berlin Heidelberg), 331–351. doi: 10.1007/978-3-642-74542-3_22
Alves, J. A., Dias, M. P., Méndez, V., Katrínardóttir, B., and Gunnarsson, T. G. (2016). Very rapid long-distance sea crossing by a migratory bird. Sci. Rep. 6:38154. doi: 10.1038/srep38154
Alves, J. A., Gunnarsson, T. G., Hayhow, D. B., Appleton, G. F., Potts, P. M., Sutherland, W. J., et al. (2013). Costs, benefits, and fitness consequences of different migratory strategies. Ecology 94, 11–17. doi: 10.1890/12-0737.1
Alves, J. A., Gunnarsson, T. G., Potts, P. M., Gélinaud, G., Sutherland, W. J., and Gill, J. A. (2012). Overtaking on migration: does longer distance migration always incur a penalty? Oikos 121, 464–470. doi: 10.1111/j.1600-0706.2011.19678.x
Alves, J. A., Gunnarsson, T. G., Sutherland, W. J., Potts, P. M., and Gill, J. A. (2019). Linking warming effects on phenology, demography, and range expansion in a migratory bird population. Ecol. Evol. 9, 2365–2375. doi: 10.1002/ece3.4746
Both, C., and Visser, M. E. (2001). Adjustment to climate change is constrained by arrival date in a long-distance migrant bird. Nature 411, 296–298. doi: 10.1038/35077063
Carneiro, C., Gunnarsson, T. G., and Alves, J. A. (2019). Faster migration in autumn than in spring: seasonal migration patterns and non-breeding distribution of Icelandic whimbrels Numenius phaeopus islandicus. J. Avian Biol. 50, 1–8. doi: 10.1111/jav.01938
Cohen, J., Screen, J. A., Furtado, J. C., Barlow, M., Whittleston, D., Coumou, D., et al. (2014). Recent Arctic amplification and extreme mid-latitude weather. Nat. Geosci. 7, 627–637. doi: 10.1038/ngeo2234
Conklin, J. R., Battley, P. F., and Potter, M. A. (2013). Absolute consistency: individual versus population variation in annual-cycle schedules of a long-distance migrant bird. PLoS ONE 8:e54535. doi: 10.1371/journal.pone.0054535
Drent, R. H. (2006). The timing of birds' breeding seasons: the Perrins hypothesis revisited especially for migrants. Ardea 94, 305–322.
Gill, J. A., Alves, J. A., Sutherland, W. J., Appleton, G. F., Potts, P. M., and Gunnarsson, T. G. (2014). Why is timing of bird migration advancing when individuals are not? Proc. R. Soc. B Biol. Sci. 281, 20132161–20132161. doi: 10.1098/rspb.2013.2161
Gunnarsson, T. G., and Guðmundsson, G. A. (2016). Migration and non-breeding distribution of Icelandic Whimbrels Numenius phaeopus islandicus as revealed by ringing recoveries. Wader Study 123, 44–48. doi: 10.18194/ws.00031
Gunnarsson, T. G., and Tómasson, G. (2011). Flexibility in spring arrival of migratory birds at northern latitudes under rapid temperature changes. Bird Study 58, 1–12. doi: 10.1080/00063657.2010.526999
Harrison, X. A., Blount, J. D., Inger, R., Norris, D. R., and Bearhop, S. (2011). Carry-over effects as drivers of fitness differences in animals. J. Anim. Ecol. 80, 4–18. doi: 10.1111/j.1365-2656.2010.01740.x
Hasselquist, D., Montràs-Janer, T., Tarka, M., and Hansson, B. (2017). Individual consistency of long-distance migration in a songbird: significant repeatability of autumn route, stopovers and wintering sites but not in timing of migration. J. Avian Biol. 48, 91–102. doi: 10.1111/jav.01292
Høye, T. T., Post, E., Meltofte, H., Schmidt, N. M., and Forchhammer, M. C. (2007). Rapid advancement of spring in the High Arctic. Curr. Biol. 17, 449–451. doi: 10.1016/j.cub.2007.04.047
Johnson, A. S., Perz, J., Nol, E., and Senner, N. R. (2016). Dichotomous strategies? The migration of Whimbrels breeding in the eastern Canadian sub-Arctic. J. Field Ornithol. 87, 371–383. doi: 10.1111/jofo.12173
Katrínardóttir, B., Pálsson, S., Gunnarsson, T. G., and Sigurjónsdóttir, H. (2013). Sexing Icelandic Whimbrels Numenius phaeopus islandicus with DNA and biometrics. Ringing Migr. 28, 43–46. doi: 10.1080/03078698.2013.811160
Liebezeit, J. R., Smith, P. A., Lanctot, R. B., Schekkerman, H., Tulp, I., Kendall, S. J., et al. (2007). Assessing the development of shorebird eggs using the flotation method: species—specific and generalized regression models. Condor 109, 32–47. doi: 10.1650/0010-5422(2007)109%5B32:ATDOSE%5D2.0.CO;2
Madsen, J. (2001). Spring migration strategies in pink-footed geese Anser brachyrhynchus and consequences for spring fattening and fecundity. Ardea 89, 43–55.
Marcström, V., Mascher, J. W., and Marcstrom, V. (1979). Weights and fat in Lapwings Vanellus vanellus and Oystercatchers Haematopus ostralegus starved to death during a cold spell in spring. Ornis Scand. 10, 235–240. doi: 10.2307/3676067
Marra, P., Cohen, E. B., Loss, S. R., Rutter, J. E., and Tonra, C. M. (2015). A call for full annual cycle research in animal ecology. Biol. Lett. 11:20150552. doi: 10.1098/rsbl.2015.0552
McNamara, J. M., Welham, R. K., and Houston, A. I. (1998). The timing of migration within the context of an annual routine. J. Avian Biol. 29, 416–423. doi: 10.2307/3677160
Nussey, D. H., Postma, E., Gienapp, P., and Visser, M. E. (2005). Selection on heritable phenotypic plasticity in a wild bird population. Science 310, 304–306. doi: 10.1126/science.1117004
O'Connor, C. M., Norris, D. R., Crossin, G. T., and Cooke, S. J. (2014). Biological carryover effects: linking common concepts and mechanisms in ecology and evolution. Ecosphere 5, 1–11. doi: 10.1890/ES13-00388.1
Pedersen, L., Jackson, K., Thorup, K., and Tøttrup, A. P. (2018). Full-year tracking suggests endogenous control of migration timing in a long-distance migratory songbird. Behav. Ecol. Sociobiol. 72:139. doi: 10.1007/s00265-018-2553-z
Perrins, C. M. (1970). The timing of birds breeding seasons. Ibis 112, 242–255. doi: 10.1111/j.1474-919X.1970.tb00096.x
Phillips, R. A., Silk, J. R. D., Croxall, J. P., Afanasyev, V., and Briggs, D. R. (2004). Accuracy of geolocation estimates for flying seabirds. Mar. Ecol. Progr. Series 266, 265–272. doi: 10.3354/meps,266265
R Core Team (2018). R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing. Available online at: https://www.r-project.org/ (accessed March 1, 2019).
Robinson, R. A. (2018). Birdfacts: Profiles of Birds Occurring in Britain & Ireland (BTO Research Report 407) Thetford. Available online at: http://www.bto.org/birdfacts (accessed February 15, 2019).
Ruthrauff, D. R., Tibbitts, T. L., and Gill, R. E. (2019). Flexible timing of annual movements across consistently used sites by Marbled Godwits breeding in Alaska. Auk 136, 1–11. doi: 10.1093/auk/uky007
Senner, N. R., Hochachka, W. M., Fox, J. W., and Afanasyev, V. (2014). An exception to the rule: Carry-over effects do not accumulate in a long-distance migratory bird. PLoS ONE 9:e86588. doi: 10.1371/journal.pone.0086588
Serreze, M. C., Barrett, A. P., Stroeve, J. C., Kindig, D. N., and Holland, M. M. (2009). The emergence of surface-based Arctic amplification. Cryosphere 3, 11–19. doi: 10.5194/tc-3-11-2009
Skarphéðinsson, K. H., Katrínardóttir, B., Guðmundsson, G. A., and Auhage, S. N. V. (2016). Mikilvæg Fuglasvæði á Íslandi. Garðabæ: Fjölrit Náttúrufræðistofnunar.
Stoffel, M. A., Nakagawa, S., and Schielzeth, H. (2017). rptR: repeatability estimation and variance decomposition by generalized linear mixed-effects models. Methods Ecol. Evol. 8, 1639–1644. doi: 10.1111/2041-210X.12797
Vardanis, Y., Nilsson, J. Å., Klaassen, R. H. G., Strandberg, R., and Alerstam, T. (2016). Consistency in long-distance bird migration: contrasting patterns in time and space for two raptors. Anim. Behav. 113, 177–187. doi: 10.1016/j.anbehav.2015.12.014
Vepsäläinen, K. (1969). The effect of the cold spring 1966 upon the Lapwing (Vanellus Vanellus) in Finland. Ornis Fenn. 45, 33–47.
Verhoeven, M. A., Loonstra, A. H. J., Senner, N. R., McBride, A. D., Both, C., and Piersma, T. (2019). Variation from an unknown source: large inter-individual differences in migrating black-tailed godwits. Front. Ecol. Evol. 7, 1–9. doi: 10.3389/fevo.2019.00031
Warnock, N. (2010). Stopping vs. staging: the difference between a hop and a jump. J. Avian Biol. 41, 621–626. doi: 10.1111/j.1600-048X.2010.05155.x
Keywords: flexibility, consistency, annual cycle, whimbrel, individual, environmental change, timing, phenology
Citation: Carneiro C, Gunnarsson TG and Alves JA (2019) Why Are Whimbrels Not Advancing Their Arrival Dates Into Iceland? Exploring Seasonal and Sex-Specific Variation in Consistency of Individual Timing During the Annual Cycle. Front. Ecol. Evol. 7:248. doi: 10.3389/fevo.2019.00248
Received: 26 April 2019; Accepted: 14 June 2019;
Published: 02 July 2019.
Edited by:
Brett K. Sandercock, Norwegian Institute for Nature Research (NINA), NorwayReviewed by:
Daniel Robert Ruthrauff, U.S. Geological Survey, Alaska, United StatesErica Nol, Trent University, Canada
Copyright © 2019 Carneiro, Gunnarsson and Alves. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Camilo Carneiro, Y2FtaWxvZmNhcm5laXJvQGdtYWlsLmNvbQ==