 Cecile Richard-Hansen1*
Cecile Richard-Hansen1* Damien Davy2Guillaume Longin3Laurent Gaillard1
Damien Davy2Guillaume Longin3Laurent Gaillard1 François Renoux4Pierre Grenand5
François Renoux4Pierre Grenand5 Raphaëlle Rinaldo3
Raphaëlle Rinaldo3- 1Office National de la Chasse et de la Faune Sauvage - UMR EcoFoG, Kourou, France
- 2CNRS - OHM Oyapock, USR LEEISA 3456, Cayenne, France
- 3Parc Amazonien de Guyane, Rémire-Montjoly, France
- 4Independent Researcher, Paris, France
- 5IRD-OHM Oyapock USR 3456, Cayenne, France
Hunting sustainability in Amazonian ecosystems is a key challenge for modern stakeholders. Predictive models have evolved from first mostly biological data-based to more recent modeling including human behavior. We analyze here the hunting data collected in French Guiana through a panel of indices aiming at drawing the puzzle of parameters influencing hunting activity and impact in various socio ecological conditions across the country. Data were collected from five different study sites differing in cultural origins and remoteness from market economy, and over a 10 years period. Most indices show an impact on wildlife populations, and using a full set of indicators allowed us to better understand some underlying mechanisms that lead to a community's hunting profile. The results showed that there are noticeable differences between the study sites in the practices and the ways hunters face the changes in environment and resources availability.
Introduction
Hunting sustainability in Amazonian ecosystems is a key challenge for modern stakeholders (Weinbaum et al., 2013; de Oliveira et al., 2018; Van Vliet, 2018). In temperate ecosystems, managing hunting deals most of the time with recreational activities, severe habitat loss and land use changes in highly industrialized/agricultural lands, and sometimes regulation of overpopulation issues, induced by ecosystem changes. At the opposite, neotropical wildlife managers often face the critical issue of human subsistence (Sarti et al., 2015; Van Vliet et al., 2017), with very different and poorly understood sociocultural drivers and human-animal relationships (Alves and Van Vliet, 2018; Shaffer et al., 2018a), but also occurring in a changing world.
In conservation and ecological literature, this topic peaked following the first Robinson and Redford (1991) sustainability model (Robinson and Redford, 1994; Alvard et al., 1997; Bodmer et al., 1997; Peres, 1997; Slade et al., 1998), then slowed down (Weinbaum et al., 2013) until recent researches bring new modeling approaches and insights in the late 2010s, including more and more human related dimensions. The first sustainability models were mainly based on ecological and biological data and provided a basic useful tool to detect clear overharvesting situations. They were widely used and sometimes misused when concluding to sustainability for harvest levels below the maximum potential yield, without assessing local ecologic and demographic parameters (Levi et al., 2011b). On the other hand, many studies reported that hunting considered unsustainable through the use of these indices has continued for decades with little to no evidence for prey depletion (Shaffer et al., 2018b). One of the main criticism is that they are static on-off indices, extrapolating, and inferring sustainability from spatial and temporal punctual situations (Levi et al., 2009, 2011b). They suppose closed systems, not taking into account possible animal movements in and out the catchment area. This is a major issue, as source-sink processes are thought to play a major role in animal densities balance on a larger scale (Novaro et al., 2000; Takashina and Mougi, 2015; Shaffer et al., 2018b; da Silva Chaves et al., 2019). From an operational point of view, another limitation comes also from the large amount of data needed if one wants to go further a first diagnose derived from maximum production conditions. In the absence of reliable data, uncertainty accumulates and predictions often failed (Van Vliet and Nasi, 2008). Finally, very few is known about the hunters' behaviors, choices and constraints that lead to the measured hunting profile.
The further step in modeling sustainability included spatial parameters, taking into account the variability in the distribution of habitats, of species and of hunters (Siren et al., 2004; Ohl-schacherer et al., 2007; Levi et al., 2009, 2011b; Van Vliet et al., 2010; Takashina and Mougi, 2015). Such biodemographic models allowed to include gradual depletion of vulnerable species from a central foraging place, multiple or growing number of settlements (Shaffer et al., 2018b), and probability of animals moving within the area through a diffusivity parameter (Levi et al., 2009). They were successfully used to predict patterns of game densities and depletion in Guyana (Shaffer et al., 2017, 2018b).
Finally, last generation models include human behavior in the predictive sustainability. Hunter behavior is derived from predators' one, and based on optimal foraging and diet-breath theories (Rowcliffe et al., 2003; Levi et al., 2011a). Underlying hypothesis are that hunters tend to maximize their offtake considering opportunities and constraints, generally as central-place foragers. The more complex models consider a multi-preys system with various prey value (mainly protein intake), probability of encounter (declining with the distance from settlement according to species rarity, detectability and vulnerability) and kill rates, as well as the hunter's investment (increasing with distance to the settlement), and various limitations as cartridge availability and the need to come back home. These models aim at explaining the proportional representation of different species (prey profile) (Levi et al., 2011a).
Long term monitoring allows detecting the changes in game populations. Ecological studies directly compare density, abundance or biomass over places with various hunting pressure, and control “undisturbed” plots, sometimes accounting for habitat variability (Hill et al., 1997; Peres, 1997, 2000; Siren et al., 2004; Haugaasen and Peres, 2005). Other indicators of ecological changes can be based on animal performance (group size, reproductive success, and body mass), habitat impact and habitat use, providing relevant information on the population–habitat system and status, and quantitative basis for flexible management decisions (Morellet et al., 2007). In tropical areas, various indicators of species or ecosystem conservation status based on hunting data have been used (Milner-Gulland and Akçakaya, 2001; Jerozolimski and Peres, 2003; Ohl-schacherer et al., 2007; Parry et al., 2009b; Weinbaum et al., 2013; Constantino, 2016). Hunting data are theoretically easier to collect than ecological ones, and are used to build indices of impact on game population based on various models and hypothesis as reported above. Without concluding with a yes/no answer on sustainability, continuous trends of validated indicators reveal potential issues for the future, and monitoring allows to confirm or to correct the trends. Moreover, we argue here that a large set of hunting-based indicators informs not only on the impact of hunting, but helps to understand the modalities of the practice and of its changes over times and/or places (Parry et al., 2009a).
In French Guiana, hunting and wildlife management are still parsimonious and need modernization and deployment. As a piece of a “western” country dropped in tropical environment, the area hosts at the same time several indigenous populations still relying on bush meat for their subsistence, modern cities with full market access and a gradient of rural areas keeping strong cultural and alimentary relationships with the forest. For the last 10 years, the government has expressed a growing interest for developing adapted management rules, based on local and scientific knowledge (Richard-Hansen and Hansen, 2004).
This study intended in first objective to document the changes in hunting-based indicators of game-species conservation status over 10 years across several villages of French Guiana. Moreover, our panel of indicators informs not only on the wildlife status, but also on the different strategies deployed by hunters according to various contexts, or to face the variations in their natural resources, helping to better understand their underlying behaviors and incentives.
Study Area
French Guiana (2°7′-5°44″ North, and 51°38′-54°35′ West), is a French overseas department, situated between Suriname and the Brazilian state of Amapá, covering ~85,000 km2 in the eastern Guiana Shield. Altitude generally ranges between 0 and 200 m above sea level (mean 140 m) with some peaks ranging from 500 to 800 m. Annual rainfall ranges from 3,600 mm (north-east) to 2,000 mm (south and west). Mean annual temperature is of 25.7°C. Evergreen rainforest covers more than 90% of the country (FAO, 2010). In 2014, 88% of the population (250,400 people) lived in the coastal strip in human-modified areas (artificial, agricultural, and disturbed areas) covering ~1,000 km2 (ONF, 2016). Outside this area, the average population density is 0.04 people km2 (INSEE, 2012). Forest logging is restricted to less than a third of the territory, under National Forest Office control. The area used to be considered as well-preserved (Taber et al., 2008), but presently suffers from persistent and growing impacts of illegal goldmining (Hammond et al., 2007; Dezécache et al., 2017), inducing habitat destruction, rivers asphyxia, and uncontrolled hunting.
Mainland French hunting laws were not suitable to the tropical ecological and sociological contexts, and consequently do not apply in French Guiana. However, conservation laws protect the most vulnerable species, and several protected areas regulate access to hunting. Since the last 10 years, things are on the move. Most recent regulations established hunting bags for some species, based on a number of pieces allowed to harvest by day and hunter. In 2018, a new law created a specific French-guianan hunting license, freely and systematically attributed to all residents asking for it until 2020.
Many different cultural and ethnic groups are present in French Guiana: Creole, Bushinengue (Aluku, Paamaka, Saamaka, Ndjuka), Hmong, Chinese, Brazilian, Surinamese, people coming from mainland France, and six different Amerindian groups (Wayãpi, Wayana, Teko, Kali'na, Palikur, Arawak-Lokono) coexist in the country. Owing to the first article of the French 1958 Constitution, stating equality of rights to all French citizens, a specific status cannot be recognized for native populations, and there are no indigenous reserve nor specific rights given to these populations. However, specific uses and livelihoods are recognized. In 2007 a national park was created (Parc amazonien de Guyane) in the southern part of the region, an area encompassing the historical territories of several indigenous populations of various ethnical origins, with the willingness to protect and promote both natural and cultural richness of the area. To fit with both the constitutional law and the spirit of the project, “communities of inhabitants traditionally subsisting from the forest” are allowed to hunt in the Park. These communities also benefit from exception of recent regulation law on hunting bags and seasons. This exception rule also applies in “Zones of Common Use Rights” outside the Park, as several native groups also live in the most northern area (Kali'na, Palikur, Arawak, Saamaka, Ndjuka) (Filoche et al., 2017). However, protected species remain strictly forbidden to harvest for anybody. The National Park authorities are willing to set up co-constructed management rules, taking into account traditional and cultural needs. A particularity to underline is that, as French citizens, local communities benefit from national social incomes and/or may work and have salaries, allowing the purchase of motorboats, gas, firearms and cartridges, which influences hunting patterns (Tritsch et al., 2015).
Methods
Hunting Data Collection
The first hunting surveys were initiated between 2000 and 2002, to document hunting practices, quantify and describe the harvest, initiate a monitoring of its impact, and try to estimate its sustainability (Grenand, 2002). In selected study sites, hunters voluntarily shared the results of their hunting activity, recorded by local field assistants. Hunters' self-monitoring was attempted but did not succeed, because most hunters rejected or rapidly gave up, with exception of very few motivated participants. Data collected for each hunt included the number of pieces of all species harvested, the duration of the hunt (from home to home, including traveling to the hunting area), the number of hunters, and information about the practice (weapon, means of transport used). We distinguished the harvests occurring during other activities (agriculture, transport), considered as opportunistic catches. The hunt location was mapped according to the hunter's indications on a 5 × 5 km grid. A form was filled even in case of unsuccessful hunts. The harvested biomass was estimated afterwards, from a database of mean weights of animals issued from local data when possible (Richard-Hansen et al., 1999; Richard-Hansen pers. data) or from literature when not available.
Around 10 years later, from 2010 to 2015, new surveys were conducted, some of them on the same sites. Surveys conducted between 2000 and 2002 are grouped as survey 1, and those conducted in the 2010s as survey 2 (Table 1). Among all the study sites, we selected five for which data were the more complete in both surveys, and lasted at least several months. These study sites represent a gradient from higher subsistence and isolated conditions (Trois Sauts, the most southern site) to more rural context, with growing access to market economy (Saint-Georges de l'Oyapock). Trois Sauts, Camopi, and Elahe are located in the southern part, within the National park, are mostly inhabited by Amerindian communities (Wayãpi, Teko, and Wayana), and cannot be reached by road (Figure 1). Régina and Saint-Georges are mostly Créole villages, with a mixed population including Palikur (Amerindian community) and Brazilian people, in the northern part of French Guiana. Conducting hunting surveys in larger towns proved to be more challenging, and could not be addressed here.
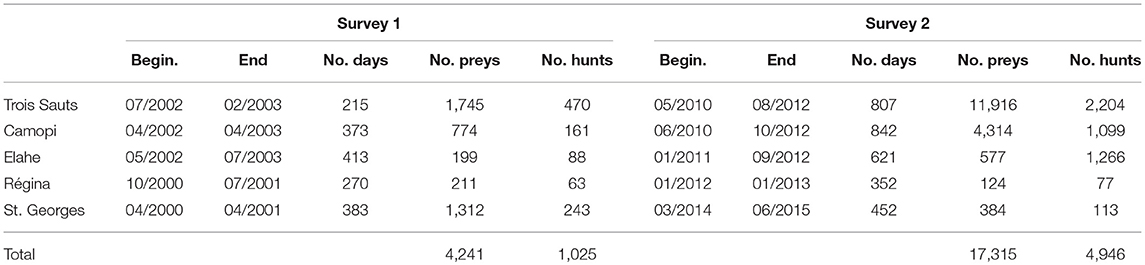
Table 1. Characteristics of the two hunting surveys conducted in French Guiana.
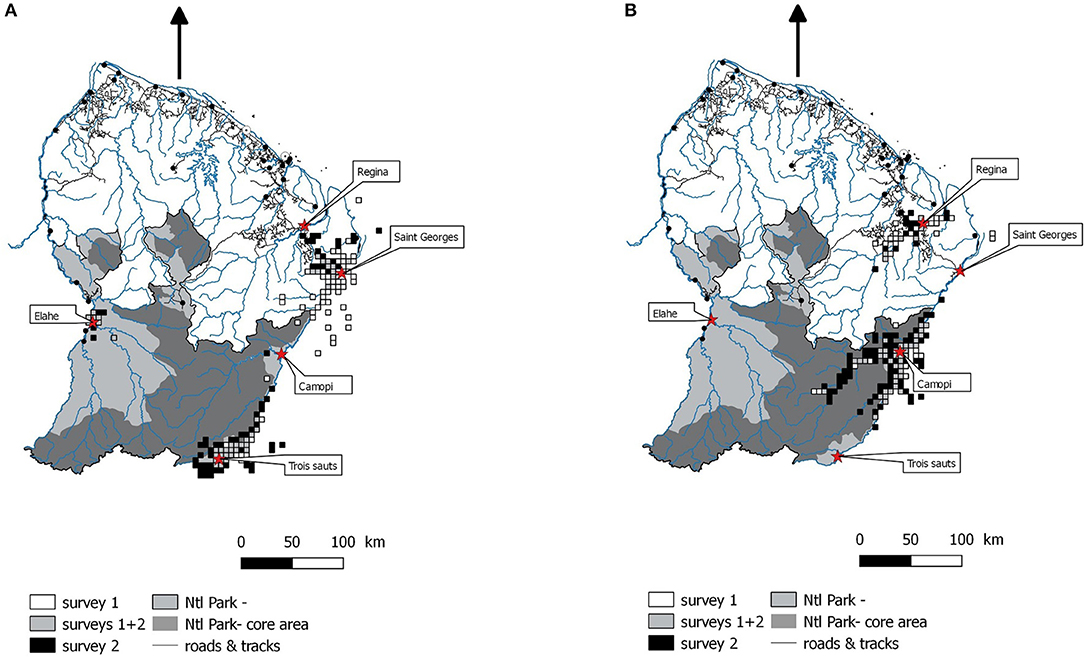
Figure 1. Map of the five study sites, and the catchments areas during surveys 1 and 2. Catchment areas were represented on different maps to avoid overlapping between sites. (A) catchment areas of Trois Sauts, Saint-Georges, and Elahe villages. (B) catchment areas of Camopi and Régina. Each square is a 5 × 5 unit with at least one kill recorded during the considered survey. White cells: catchment during survey 1 only; black cells: catchments during survey 2 only; gray cells: catchments during both surveys.
Data Analysis
Variables describing the hunting strategies included the mean duration of the hunting trips, the proportion of daily vs. overnight trips; the proportion of hunts using a motorized vehicle (may be car or motorboat), and the number of hunters. Variables describing the hunting results included the total number of preys and the total biomass returned in each hunt, the mean prey biomass, the proportion of species or species groups (prey profile), and in particular the Rodents/ungulates ratio. The Catch per unit effort (CPUE = biomass/hunter/hour) was calculated as an integrated indicator of returns over effort. The mean number of preys and the mean biomass collected by each hunter in a single trip were also computed. Duration and time related indicators were calculated excluding opportunistic catches, for which the spent time was not related to hunting effort. Some large species known to be the most vulnerable to hunting pressure (Robinson and Redford, 1986, 1991) were analyzed separately, or grouped together as sensitive species: the tapir (Tapirus terrestris), the black spider monkey (Ateles paniscus), the Artiodactyla (peccaries and deers: Tayassu pecari, Pecari tajacu, Mazama nemorivaga, and Mazama americana), and the large Cracid black curassow (Crax alector).
The hunting spatial pattern was estimated by the total catchment area, calculated by the number of grid cells with at least one catch. The shape of the catchment area was described by the perimeter/area ratio. For each prey, the straight-distance from the catch to the village was calculated from the centers of 5 × 5 grid cells.
All indicators reflect an underlying hypothesis regarding impact of harvest on the game population and/or an associated hunting practice, according to the different theoretical frameworks presented above (Table 2).
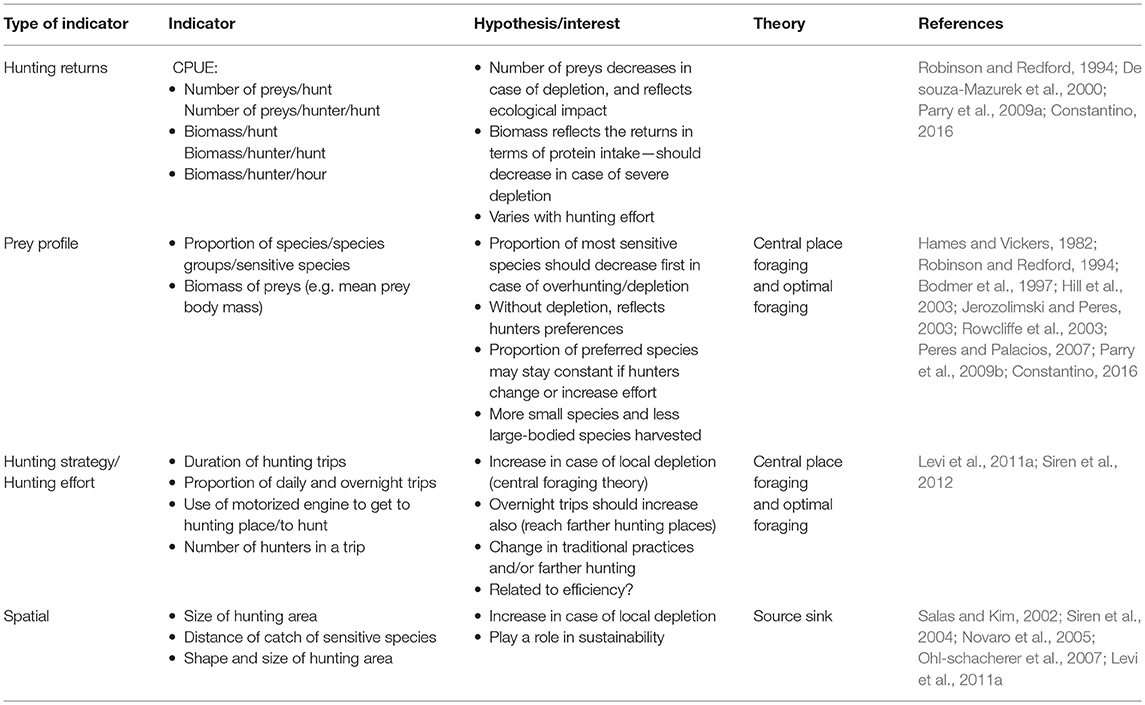
Table 2. Variables, indicators and related hypothesis used to explore the hunting impact and practices in French Guiana.
More data were collected during the second survey because a much larger effort has been involved in the program. However, we checked that sampling enclosed both rainy and dry season in each place and survey, to avoid bias related to seasonal activities, and that no indicators were correlated with monitoring effort (as the number of days of study in each village and survey) (Spearman correlation r < 0.5, p > 0.2 for all comparisons). Only one weak correlation was found, with the use of motor transportation (r = 0.6, p = 0.05), but we assumed no causality. To explore the relationships between hunting strategies and hunting returns, together with time-(two surveys) and site-scales effects, we first ran stepwise regressions to fit generalized linear models with backward selection of candidate variables, and selection criteria based on AIC. Response variables were log transformed, and Poisson regression was used on count data (number of preys), with a goodness-of-fit chi-squared test on the residual deviance. To closer examine the time effect on our indicators, mean values from surveys 1 and 2 were compared by student t-test or Wilcoxon tests, and proportions by khi square tests. Spearman rank correlation test were conducted on paired variables.
Results
General Patterns
Five thousand nine hundred and seventy one hunts and Twenty one thousand five hundred and fifty six kills were recorded during the whole study, in the five study sites.
We first explored the effects of some possible explanatory parameters on selected indicators as response variables. Do the number of hunters participating in a hunt, the duration of the trip, the use of motor to reach the hunting place, the date of study (survey) or the study site influence hunting results, in terms of total number of preys, total biomass, or mean prey biomass? Spearman correlation matrix between variables showed first that the number of hunters was positively correlated with the duration of the hunt (r = 0.82, p < 0.01). The total biomass harvested by hunt increased with the number of hunters (r = 0.64, p < 0.05) but not with the duration of the hunt (r = 0.2, p > 0.0.5), while the biomass per hunter is not significantly correlated. The duration of motorized hunts is significantly higher (7.5 vs. 5.7 h; p < 0.01 Student t-test), and these motorized hunting trips provided more biomass (19.3 vs. 13.1 kg; p < 0.01 Student t-test) and more preys (4.3 vs. 2.5 preys; p < 0.01 Student t-test), with preys being a little bigger (4.7 vs. 3.9 kg; p < 0.01 Student t-test). However, the biomass collected for each hunter is not different (12.5 and 12 kg/hunter of biomass, t-test p > 0.5).
Overnight trips globally provided less preys (24%) but more biomass (38%) that expected on the basis of days allocated (30% of hunting days), both proportions being significantly different from null hypothesis (χ2 tests, p < 0.001).
General linear models including all explanatory variables (number of hunters, duration, use of motor engine, survey, and site) returned poor fitting and predictive power. According to the selected model (quasi poisson regression, full model), the most important significant positive parameters on the total number of preys was the number of hunters and the use of motor to go hunting (Table 3). The goodness-of-fit chi-square test on residual deviance was not significant (p > 0.5), indicating the model fits the data. Negative parameters corresponded to sites effects, mainly in Régina and Saint-Georges. The survey variable had no significant effect. The full model was selected to partly explain the total biomass returns (R2 = 0.18, p < 0.001, Gaussian regression). The number of hunters and the same sites as previous analysis (Régina and Saint-Georges) positively influenced the returned biomass, while Trois Sauts site and the survey negatively did.
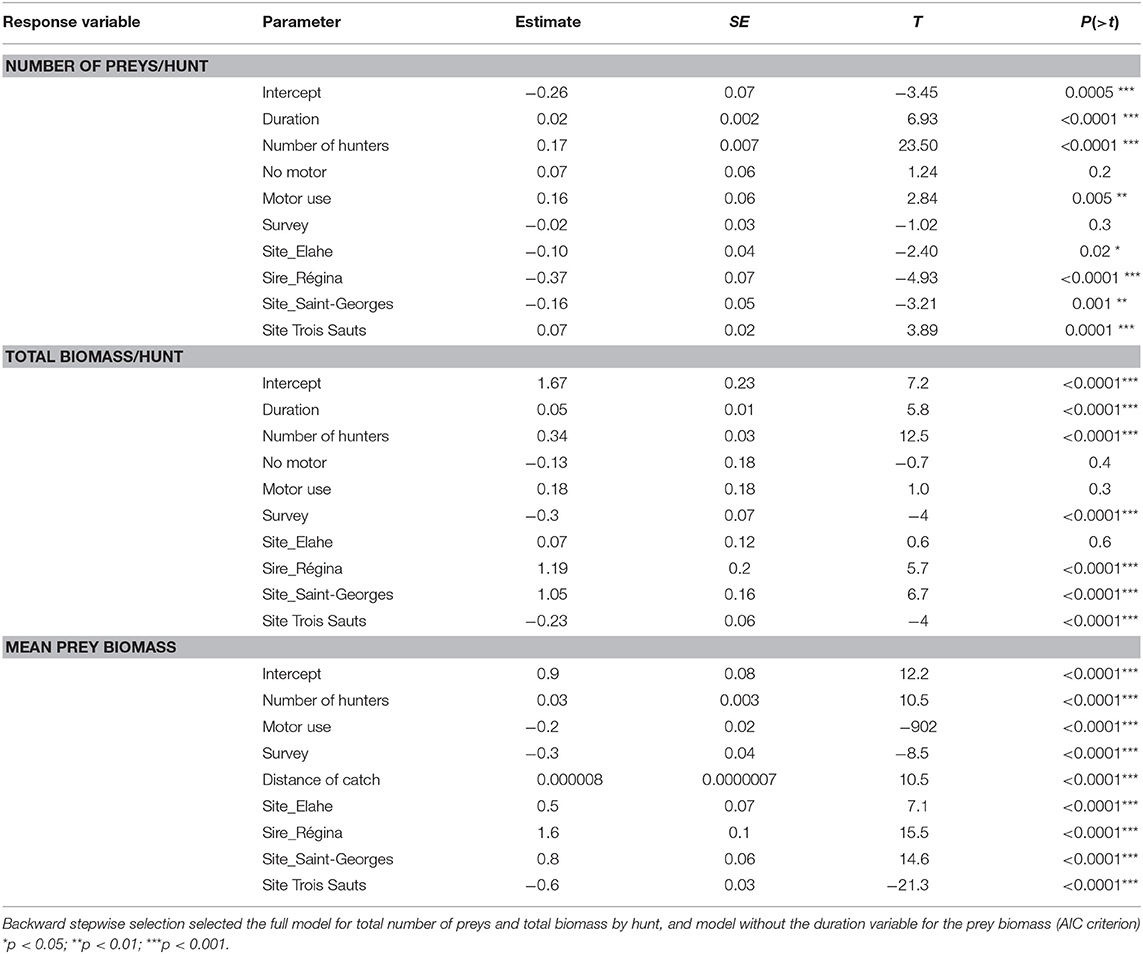
Table 3. Variables explaining the variation of the hunting returns, in terms of (i) number of preys by hunt (quasipoisson regression, N = 3,192, Dispersion parameter taken to be 0.2899811, GOF test on residuals >0.5) (ii) total biomass by hunt (gaussian regression, N = 2,889, adjusted R-squared: 0.18, p-value: <2.2e-16) (iii) mean biomass of individual preys, N = 16,328, adjusted R-squared = 0.21, p < 0.0001).
Mean prey biomass (logged) was negatively influenced by the survey and motor use, positively in Régina and Saint George and Elahe, and negatively in and Trois Sauts (R2 = 0.21, p < 0.001). The duration of hunt has no significant influence, and was not selected in the best model after backwards selection, based on AIC values.
These first results show that general models on the whole dataset explain only a small part of the variance, and that the site effect is strong. Trois Sauts site differs the most from others, with higher number of preys but lower total and prey biomass. On the other hand, Régina and Saint-Georges sites generally have patterns that are more similar.
Spatial Patterns
The size of the hunting areas vary from 200 to 2,775 km2, as estimated by the number of 25 km2 grid squares with at least one catch recorded (Table 4), according to the survey and the study site. The shape of the main hunting area (excluding isolated square grids cf. map Figures 1A,B) was characterized by the perimeter/area ratio. For all study sites, it was around 0.3 (up to 0.44 for Régina), indicating an elongated shape. Rounded shapes corresponding to the same areas would have a ratio inferior to 0.1, except for Elahe.
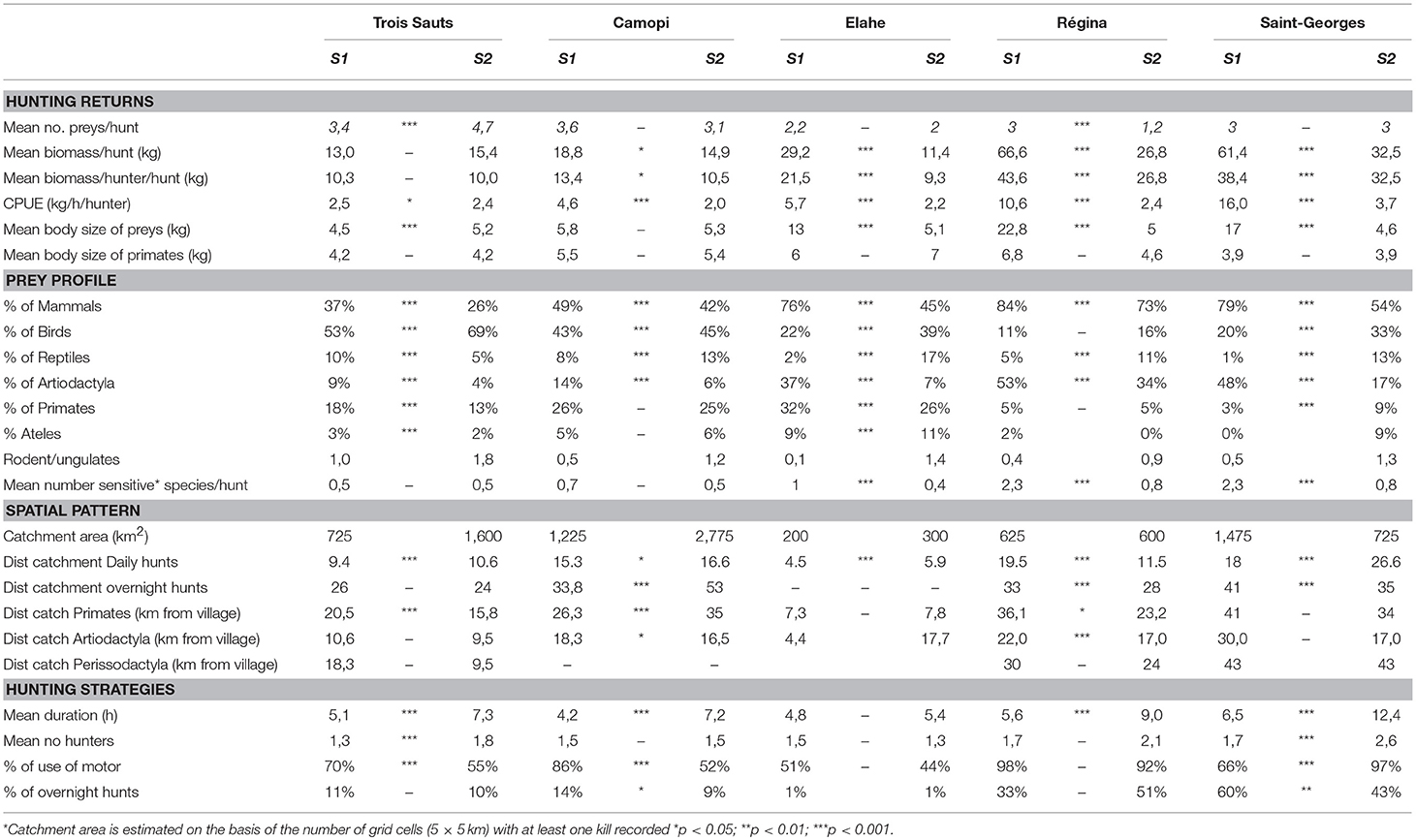
Table 4. Changes in various indices between two hunting surveys (S1 and S2) conducted 10 years apart in five study sites of French Guiana.
Mean catchment distance during daily hunts is lower for birds (13.8 km) than mammals (21.7 km), and Reptiles (27.3 km) (p < 0.001 Student t-tests for paired comparisons) (Figure 2). Among mammals, it is the highest for Tapirus terrestris (28 km) and large primates (26 and 29 km from settlements for Ateles and Alouatta, 25 km for Cebus apella vs. 8 km for Saguinus, paired t-tests < 0.001). Rodents and Artiodactyla are both harvested at around 20 km from settlements. For Rodents, the distance is higher for Agouti paca (26 km) than Dasysprocta leporina (10 km, p < 0.0001 t-test), and among Artiodactyla, it is higher for peccaries (respectively, 21 and 22 km for Pecari tajacu and Tayassu pecari) than for brocket deers (15 and 14 km for Mazama americana and Mazama nemorivaga, respectively, p < 0.0001). Among birds, largest distances of hunting are recorded for the largest species as black curassow Crax alector (24 km), trumpet birds (Psophia crepitans, 22 km), or Penelope guan (Penelope marail, 17 km), while the smaller species from Columbiform, Passeriform, Psittaciforms orders are collected within a ten km radius (6, 8, and 9 km, respectively). Among Reptiles, the crocodilian are harvested nearest villages (28 km) than Squamates (mainly Iguana iguana, 35 km).
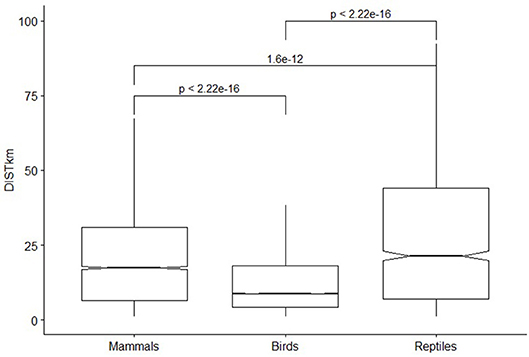
Figure 2. Mean distances and quartiles of catch of mammals, birds and reptiles groups during the whole study. Probability of Student paired tests is indicated for each comparison. Box plots are figured without outliers.
Trends Over 10 Years
Trends of most indices were compared between the two surveys period. Most indicators showed clear changes between the two surveys, but also varying according to the study site.
Hunting Returns
The total biomass per hunt and the various hunting yields (CPUE, biomass harvested by hunter) tend to decrease everywhere between the two surveys, but less markedly in Trois Sauts (Table 4, Mann Whitney test). As suggested by the results of general linear models, the number of preys harvested by hunting trip does not change a lot between the two surveys, although it tend to increase in Trois Sauts and decrease in Régina. The mean biomass of preys (e.g., body size) significantly decreased in three sites, whereas no changes were observed in Camopi, and it slightly increased in Trois Sauts. The mean biomass of primates did not change, indicating no shift to smaller primate species.
Prey Profiles
The proportion of mammals significantly decreased (Chi square test, p < 0.001 for all sites, comparing survey 1 and 2), and the proportions of birds increased (Chi square test, p < 0.001 for all sites except Elahe) in all prey profiles of survey 2 compared to survey 1 (Table 4). Regarding Reptiles, their proportions increased in all sites but Trois Sauts, where their proportion decreased. Among mammals, the proportion of Artiodactyla clearly decreased everywhere, while the primates' one decreased in Trois sauts and Elahe but remained stable in Camopi and Régina, and increased in Saint-Georges. In particular, the largest, most vulnerable and preferred primate species Ateles paniscus presented the same trends as the primates group considered as a whole. The mean number of sensitive species (see Methods) harvested per hunt tend to decrease, except in Trois Sauts and Camopi. The Rodents/Ungulates ratio increased in all sites.
Hunting Strategies
Considering the hunting strategies, the mean duration of daily hunts significantly increased in all sites (Table 3, Mann Whitney test all p < 0.01, except for Elahé, Table 4). The mean number of hunters participating in a hunting trip tended to increase in Trois Sauts and Saint-Georges. The proportion of motorized access to hunting decreased in Trois Sauts and Camopi, and increased in Saint-Georges, and the proportions of overnight trips show no clear tendency.
Spatial Pattern
The size of total catchment area increased in the three southern villages (Trois Sauts, Camopi, and Elahé) but not in the two other ones (Régina and Saint-Georges), as estimated by the number of quadrats with at least one catch recorded (Table 4). The map shows that the most clear spread of hunting area occurred in Trois Sauts, from which hunters clearly went farther for hunting during survey 2 (black squares indicate areas used only during survey 2, Figures 1A,B). In Camopi, we observe a more intensive use of middle distance areas, and some new rivers explored (Figure 1B). One consequence is a higher overlap of hunting areas between villages (Camopi/Trois Sauts, and Camopi/Saint-Georges de l'Oyapock) during survey 2.
Mean catchment distances tend to increase for daily hunts (but decreased in Régina), but for overnights hunts only in Camopi. The straight distance of catch of sensitive species as primates, Artiodactyla or Perissodactyla did not change markedly (Table 4).
Inter-sites Differences
Many previous results pointed how indicators' changes over year differed according to study sites. A full analysis is currently under process to characterize a hunting typology in French Guiana, but we can point out here some major differences between our five study sites.
Hunting Strategies
Overnight trips were much more common in Saint-Georges de l'Oyapock and Régina, representing around half of the records (54 and 43% of occurrences, Table 5). Most overnight hunts lasted 3 days in average (87% between 2 and 4 days), so this hunting practice represents up to 78% of hunting days recorded in Saint-Georges de l'Oyapock. Regarding the number of preys and the harvested biomass, overnight trips bring back a lower amount of game or biomass than expected on the basis of proportion of days allocated χ2 tests p < 0.001), except in Elahe in which 23% of catches and 21% of biomass occurred during the 2% of time spent in overnight hunting trips. In Camopi, less preys but more biomass than expected was harvested during overnight trips (χ2 test p < 0.001)
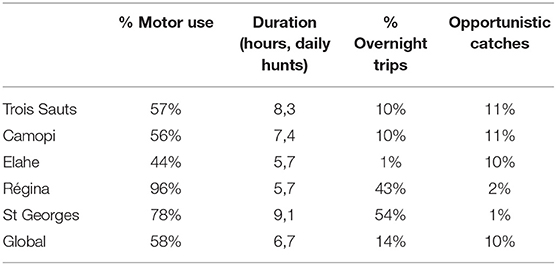
Table 5. Hunting strategies in five study sites of French Guiana.
Voluntary daily hunts were also longer in Saint-Georges de l'Oyapock (9 h in average, Table 5) than in other sites (6.7 h in average, t-test, p < 0.001). Using a motor vehicle (car, boat or motorcycle) to reach the hunting area is much more common in Saint-Georges and Régina (78 and 96%) than in the southern sites, with a minimum of 44% in Elahé.
Biomass harvested per hour and per hunter (CPUE) is of same magnitude for the three southern villages (Trois Sauts, Camopi, and Elahe: respectively, 2.5, 2.4, and 3 kg/hunter/hour), and clearly lower than hunting yields from Régina and Saint-Georges de l'Oyapock (6.8 and 10.2 kg/h/h, respectively).
In southern sites (Elahe, Trois Sauts, and Camopi), opportunistic harvests (i.e., performed during another activity as travel or agriculture) provide a 10–14% proportion of catches, while it is negligible in Régina and Saint-Georges (Table 5).
Prey Profile
Over the whole study, mammals represent 36% of the number of animals harvested, birds 56% and Reptiles 8%. However, 86% of the biomass is mammals, and 8% birds. There are large differences between the five study sites regarding the prey profile (Figure 3). Mammals, and in particular ungulates clearly dominate the harvest for Saint-Georges de l'Oyapock and Régina, while birds (67% of preys in Trois Sauts) and particularly toucans (36% of preys in Trois Sauts) are the most hunted species in southern Amerindian villages. For these villages, primates are an important part of the harvest (14–27%), representing 19–26% of the biomass. Regarding protein intake, the large amount of harvested toucans brings <3% of the biomass consumed. Tapir hunting provides 12% (Camopi) to 39% (Régina) of the biomass harvested.
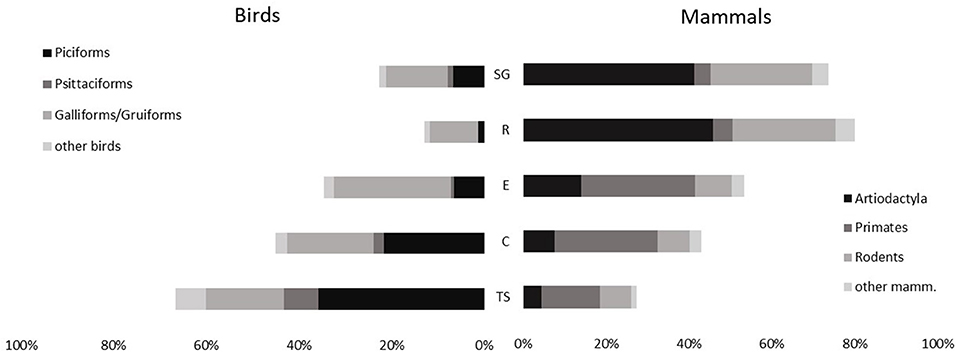
Figure 3. Comparative proportions of main taxa harvested in the five study sites. SG, Saint-Georges de l'Oyapock; R, Régina; E, Elahe; C, Camopi, TS, Trois Sauts.
The Rodent/Ungulate ratio is higher in the three southern sites (1.28, 0.96, and 0.63 in Trois Sauts, Camopi, and Elahe, respectively), than the two other ones (0.33 and 0.21 in Saint-Georges de l'Oyapock and Régina, respectively).
In all sites, overnight trips bring back a larger proportion of large primates (Ateles, Alouatta, and Cebus), large birds (Galliform), Crocodilians and Iguanas than expected on the basis of number of days spent.
Spatial Patterns
Among study sites, Elahe has the most spatially concentrated hunting pattern, the majority of hunts taking place within 4 km, except for primates (Figure 4). Hunters from Trois Sauts mostly hunt within a 10 km radius, except for primates, and rodents. For Camopi, the distance is the highest for primates. The two northern sites, Régina and Saint-Georges, have the wider spatial patterns, catching all preys over 20 km away from the village. The total catchment area, estimated through the number of grid cells (5 × 5 km) with at least one catch, was the largest for Camopi with 121 grid cells (3,025 km2), and the minimum area value is found in Elahe (Figure 1A).
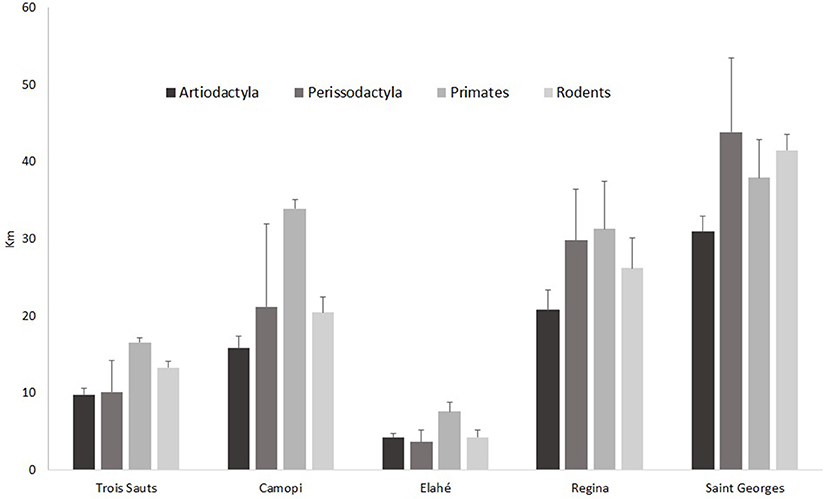
Figure 4. Mean distances of catch of mammals groups of species in the five study sites, and 95% confident interval.
Discussion
The main objectives of the study was to assess the hunting impacts on the animal community and to scan its evolution over a 10-year period, using hunting-based indicators according to various theoretical frames presented in the introduction. We also wanted to explore hunters' strategies in relation with hunting results, and how they changed or adapted over the years.
Concerning impact on game populations, we can underline that hunting areas and distances recorded here are generally higher in our study comparing to literature on Amazonian traditional hunting patterns. Traditional central foragers generally spread over a 10 km radius (Constantino, 2015; Shaffer et al., 2018b), while it is the smallest mean distance recorded in this study. We showed that catch distances of species known to be vulnerable as tapirs and larges primates are significantly higher than other species, which is coherent with central foraging theory predictions, and denote their progressive depletion around villages. Among larger bodied species, artiodactyla (peccaries and brocket deers) are encountered at comparatively smaller distances from central places, whereas they are considered as preferred items and vulnerable species. Despite their body mass, brocket deers are considered as to be less vulnerable and preferred species than peccaries, because they are difficult to hunt (Levi et al., 2011a), or taboos (Shepard et al., 2012), and their harvest is often considered as sustainable (Hurtado-Gonzales and Bodmer, 2004). In this study, they appear to persist closer from villages than other large game species. Peccaries are generally used as indicator species, strongly impacted by overhunting, but we previously found that their abundances in French Guiana are highly unpredictable and driven by large temporal and geographical scale process not only directly related to local hunting pressure (Richard-Hansen et al., 2018, 2019). The Rodent/Ungulate ratio is a commonly used indicator, useful because synthetic, but it is however strongly influenced by the presence of white lipped peccaries, in the environment and consequently in the harvest. It therefore has to be carefully interpreted, and considering a larger context. Birds follow the same predictable pattern, with large species being probably extirpated around villages, and smaller species harvested around them. Rodents are generally considered as second choice, but we found that the Agouti paca seems to be depleted in the vicinity of villages also. Note that Reptiles (mainly green iguanas and crocodilians) are also harvested at very large distances, which would indicate that these games are among the most depleted around villages.
Hunters' practices influence only slightly the quantitative outcomes of the hunts. The total hunting returns seem to increase with some allocated efforts, as the number of hunters and the use of motorized engines, but not the real benefits for each hunter. The overnight trips allowed harvesting smaller number but higher biomass of preys compared to the expected quantities based on the number of allocated days, which proved they are efficient for large game harvesting. Overnight and longer hunting trips and as well as motor use allow farther hunting from the village (as they do for slash and burn agriculture, Tritsch et al., 2015), giving access to less disturbed areas with larger-sized and more abundant game, but requires more hunters, probably for cost sharing and/or security (Siren et al., 2012). The “benefit” is counterbalanced by the necessity of sharing the total amount harvested.
Trends Over Time
Comparing over 10 years, many indices point out a probable diminution of game populations around all the studied sites, with clear differences between them. The most general tendency observed indicating the decreasing resources is the growing time allocated to hunting, while the total biomass brought back home generally decreased despite this increasing effort, with the noticeable exception of Trois Sauts. Although we found that using motorized vehicles or boat may increase the yields, as well as going to overnight hunting trips, hunters did not systematically increase this behavior to face the decreasing yields. At the opposite, although increasing the number of hunters does not increase the amount of biomass, we found a general tendency to go hunting in larger groups. In the northern part, this can be related to a need of sharing costs and increase security. In the southern villages, younger people tend to loose traditional knowledge because of time spent in school, and need to be accompanied by eldest for cultural transmission (P. Grenand, obs. pers.). These examples also show that the hunters' behavior is not always optimal, or that this optimization has to deal with many complex constraints. Concomitantly with longer trips, we found that hunting areas expanded, and/or partly shifted. In Trois Sauts, for example, hunters used more intensively the upriver area, which is not farther from the village, but harder to reach because of strong rapids and rocky bars on the river, increasing the effort for hunters. This probably explain that proportion and hunting distances of sensitive species of primates for example did not increase substantially. This area used to be prospected for long time by a small part of the population, leaving upriver from the falls, but increasing the population size also probably leading to a social reorganization of hunting territories (Tritsch et al., 2015).
The other expected changes when facing decreasing resources is shifting to more resilient and/or less preferred species, generally smaller ones (Table 2). This is illustrated in our results by the general increase of the Rodent/Ungulates ratio, and decrease of the size of preys and the proportion of mammals. Hunting rodents and smaller species is generally a second choice (Suárez et al., 1995; Levi et al., 2011a), but these species with higher reproductive rates (Robinson and Redford, 1986) are less impacted by hunting pressure and more resilient. However, when hunting turns to more recreational or commercial activity, hunters tend to focus on a few attractive species, and are not systematically shifting to smaller preys (Redford and Robinson, 1987). In Régina and Saint-Georges, ungulates are undoubtedly preferred preys, but their proportion nonetheless decreased clearly between the two surveys, while the proportion of rodents increased in Régina and not Saint-Georges.
Diet breath theory also predicts that much preferred species will be pursued with increasing effort until intense depletion (Levi et al., 2011a). Primates are a highly preferred game for Amerindian populations but are very sensitive to harvest owing to their very low reproductive rates (Bodmer, 1995; Robinson, 2000). The proportion of primates in the prey profile of Trois Sauts previously increased from 11.3 to 19.3% between 1977/78 and 95/96 (Ouhoud-Renoux, 1998b), when the hunting technology shifted from bow to firearms (Grenand, 1995), increasing their hunting efficiency. However, we observed that this proportion tend to stabilize until our first survey in 2002 (18%), and began to decrease after 10 additional years of harvesting (13% in 2012). Increasing effort can be measured by the increasing of time and/or distances allocated to reach favorite game species. The mean distance of catch of primates (and in particular for Ateles) effectively increased in Camopi, but not in other sites, and even decreased in Trois Sauts, probably in relation with a change in hunting area (see above). The mean biomass of primates did not change, indicating no shift to smaller species. These results indicate that the depletion of large primates is not achieved around Trois Sauts or Camopi, as hunters still manage to find these favorite games, but that the decline is initiated.
Our results also highlight the differences between our five study sites, and particularly between the three most traditional villages of the National park in one hand (southern sites) and the two rural ones (northern sites) in the other hand. The three autochthonous villages living for great part from subsistence hunting (Trois Sauts, Camopi, and Elahe) present some similarities in their practices and in the evolution of their practices over time. They used to hunt more birds than mammals, and larger proportion of primates and smaller birds as Toucans. The larger bird consumption is generally related to subsistence contexts (Benítez-López et al., 2017), as the larger width of the diet (Grenand, 1980, 2002). Comparing to rural sites (Régina and Saint-Georges), these hunters go hunting more often alone, with less motorized transport mean, for daily hunts on smaller distances. Harvesting animals is an everyday reality, regularly implemented during other activities as transportation, fishing, or slash-and-burn agriculture (“opportunistic catches”) (Grenand, 1980). In this place, there is really no market, subsistence and proteins need are absolute for everyday meals. In this situation, hunters manage to come back with a similar amount of biomass, increasing efforts, distances, or shifting preys.
In Régina and Saint-Georges de l'Oyapock, most hunters use motor vehicles and boats to go farther and for longer hunts farther from the village, more often with especially dedicated overnight trips. The depletion in the 21st km away from the villages is probably strong, as proved by the long hunting distances recorded. However, the small proportion of primates harvested is not easy to interpret: they could have been severely depleted, but as they are not pursued as preferred game, this indicator could be non-representative in this case. Large mammals and particularly ungulates are the main preys, providing hunters higher biomass yields although they hunt in larger groups. These high yields do not reflect healthy populations, but increased efforts over very large areas, allowed by modern means and money. In these sites, hunting is still a strong tradition, and provides protein complement to low income rural populations. However, bushmeat is not a survival need anymore, and access to market modifies the relationships with natural resources in contradictory ways, providing both alternative protein source, and bushmeat trade opportunities. Hunters tend to reduce they prey choice, seeking mainly ungulates, tending to less traditional prey profiles (Redford and Robinson, 1987), but wild meat remains on the menu (Alves and Van Vliet, 2018). According to their incentives, hunters will not face the changes in their environment on the same way.
Spatial Patterns
As often underlined, the use of space is the major key (Levi et al., 2011b; McNamara et al., 2015; Takashina and Mougi, 2015; Constantino et al., 2018; Shaffer et al., 2018b; Van Vliet et al., 2018). Most models rely on central foraging models, with a homogeneous access to the forest from the settlement. However, the size and shape of a hunting area depend both on geographical (penetrability of the environment) and economic parameters (De souza-Mazurek et al., 2000; Siren et al., 2012; Siren and Wilkie, 2015). Infrastructure expansion has been widely related to the increase of wild meat harvest and trade in the tropics (Laurance et al., 2015; Benítez-López et al., 2017). As long as there is no new access paths, hunting will be concentrated in reduced areas, constrained by physical limits. This explains in particular why the hunting areas expanded few across the years, despite decreasing available resources. In our study, most hunting areas are already strongly enlarged, and widely elongated along major access ways. In Camopi, the social incomes together with the presence of two major rivers allowed the inhabitants, by equipping themselves by canoe and motor to increase their predation and agricultural zone (Tritsch et al., 2015). Other parameters can limit the expansion of the area. In Elahe, although the village is also located along two main rivers, hunters remain on a small territory, apparently taking few advantage of this opportunity to expand their catchment area. This is partly due to competition with other populations for access to the resources (Davy, pers. data), as the territory of this village is surrounded both by other villages (Wayana and Businenge) and illegal gold-miners. In Trois Sauts, hunting territories spread from 770 to 1,180 km2 between the years 1976/1977 and 1994/1995, to face the population increase (Ouhoud-Renoux, 1998a). Methodologies used to estimate the superficies are not directly comparable, but it seems that 20 years later, their territory increased again a little. Indeed, the main changes consist in the creation of new settlements related to population increase, rather than increasing size of the hunting territories of initial villages we sampled (Davy et al., 2012). In this case, there is no competition with other groups, but topography do not allow further expansion.
We think that the hunting impact on wildlife populations around our study sites is manifest, but still concentrated around settlements and access paths. The larger, linear and ramified shapes of the catchment areas in our study sites enhance the sustainability of the harvest (Constantino, 2015; Tritsch et al., 2015), diluting the harvest on larger superficies, and providing more sources-sink exchanging systems along elongated frontiers than a circular central foraging traditional catchment area (Salas and Kim, 2002). Although the area are larger than estimated in more traditional contexts (Constantino et al., 2018), this can be viewed as positive changes through modernized practices. Allowing spatial expansion of hunting would spread the impacted area, but may increase the sustainability, as long as the total offtake does not increase (Stearman and Redford, 1995; Ouhoud-Renoux, 1998a; Grenand, 2002). Improving sustainability of hunting in isolated villages may therefore go through favoring their movements, as it was implemented in some ethnodevelopment projects (Stearman and Redford, 1995). Constantino et al. (2018) provide a full analysis of the integration of hunting and source areas in the delimitation of indigenous lands in Brazil, taking into account population growth. They propose useful scenarii to be implemented, as redistributing villages respecting distances between them to ensure functional source-sink systems. This may be discussed with communities, as it may echo a behavior currently spontaneously emerging in some places (Tritsch et al., 2015), but in French Guiana may be difficult owing to the very large hunting territories recorded.
The sustainability diagnosis of a harvest depends not only on the scale of the catchment area (Robinson and Redford, 1994), but also on the scale of the analysis (Hill and Padwe, 2000; Shaffer et al., 2017). Shaffer et al. (2018b) concluded that the WaiWai hunting may be considered as sustainable within the Indigenous reserve as a whole, because sensitive species will be extirpated from <13% of the total area of their Reserve, and persist in the catchment area. Novaro et al. (2000) estimated the size of refugia (area with unharvested population) needed to prevent overharvesting, in relation to reproductive parameter of species and proportionally of the size of catchment area. A buffer area of 15 km around the main catchment area of Camopi represent a total area of more than 11,000 km2. Proportionally to the hunted area of <3,000 km2, 73% are non-hunted and act as refugia and adjacent source for game species, which fulfill the requirements for sustainable use of most vulnerable species as spider monkey and tapir (Novaro et al., 2000). The situation is the same for Trois Sauts, insuring sustainability on large scale, but probably hardly in Elahe, because of neighboring populations, as well as Régina and Saint-Georges for which only the southern part of the catchment area is surrounded by non-hunted areas. However, even in most favorable cases, the sustainability issue has not to deal with the species conservation but more with the survival of these human communities. Finding proteins every day may however become very hard for them, who probably reached their maximal capacity of spatial extension.
Sustainability Models and Indicators
In the literature, most models are elaborated from data from one study site or community, and assuming hunters acting with optimal behaviors. We saw here that hunters' behaviors are complex, not always optimal, and driven by many sociological, cultural, economic and geographical constraints. Comparing with Levi et al. (2011a) results and predictions, some of our results do not fit their hypothesis, and some do, depending on the socio-economic and cultural context. As predicted by Levi's model, for example, we found hunters' return rates remaining consistent even when vulnerable species were depleted around the village of Trois Sauts, but they decreased clearly in the other sites. However, they predicted that lower-value game do not expand into the diet in a depletion scenario, but we found a general increase of the rodent/ungulate ratio, of birds and smaller preys. The game value is a strong cultural parameter, not only dealing with protein amount or probability of killing. The proportion of large primates, being considered to be the most vulnerable species, is generally thought as an indicator of game depletion and hunting sustainability. However, in some places, their proportion in prey profile decreased less than ungulates ‘one, probably because they are very actively researched in those communities. Many reasons may lead to kill or not kill an apparently valuable prey, as complex dietary taboos, taste of the meat, cultural or religious bans that are likely to change along the time. In French Guiana, the Busi nenge community (Maroon's descendants) has a strong demand for Ateles killing for ritual mourning ceremonies. Wayãpi had a strong relationship to living environment, and their harvest used to be controlled by the precept “No doing too much” (Grenand and Grenand, 1996). Those populations coexist in French Guiana, aside urban and rural ones, having very different constraints and incentives.
Conclusion
Our results alert on the risk of relying on too few indicators to assess hunting impact, sustainability or game depletion in different contexts. Indicators sometimes give apparently contradictory conclusions, but this is without accounting hunters' strategies. For example, a stable CPUE value can mask an increasing harvest together with a proportional increasing effort, therefore not going toward sustainability but overharvest, until a tipping point. Using a full set of indicators allowed us not only to look for hunting impact and sustainability, but also on some of the underlying mechanisms that lead to a community's hunting profile. We saw that each community has a different answer to the changing conditions, according to its own values and constraints. Integrative indices are useful, but may reflect different realities. Modeling detect general trends, but human behavior are complex to model and management needs to adapt to local constraints (Renoux and de Thoisy, 2016).
Long term monitoring with hunters' participation remains the best way to understand and accompany the changes in behaviors, practices and impacts of hunting. It provides basic elements for a flexible management, which can be improved by the integration or recovering of traditional knowledge (Berkes et al., 2000). Modern tools of adaptive management include the concept of learning processes to improve the knowledge of the system functioning, in particular in situations of high uncertainty levels (Keith et al., 2011).
However, our results suggest also that hunting management in French Guiana should be considered differently in southern and northern part of the territory, owing to different hunting practices, accessibility level to the wildlife resource, gradient of modernity in the livelihoods, and former and cumulated human impacts. Moreover, we highlighted that the sustainability of high-level harvests depends mainly of the presence and persistence of large non-impacted areas surrounding the catchment areas. Southern French Guiana remains quite preserved by its remoteness, but this relative quietness is presently severely threatened by the diffuse sprawl of illegal goldmining.
Author Contributions
PG coordinated the Silvolab Program in 2000, and co-supervised the hunting program of the Parc amazonien de Guyane from 2010 to 2012 with DD, CR-H, and RR. GL conducted most field work of this same program. FR and LG conducted hunting surveys in northern villages in surveys 1 and 2, respectively. CR-H participated to fieldwork and/or supervision of all programs, made the analysis and wrote the initial draft of the manuscript. All authors revised the manuscript text.
Funding
Ministère de l'Environnement. French Ministry of Environment (Programme Ecoystemes Tropicaux, Mission pour la Création du Parc de la Guyane, DIREN Guyane, DEAL Guyane, Parc amazonien de Guyane, ONCFS), Labex CEBA, and Labex DRIIHM (OHM Oyapock, CNRS).
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We are very grateful to all hunters that accepted to participate to the study, on a voluntary basis, and shared their hunting results, as well as Amerindian leaders Jacky Pawey, Thomas Palassissi, Raymond Lassouka, René Maïpouri, Laurent Pilauku, Albert Mifsud (†), Guy Barcarel, and Roger Labonté who allowed this study. Many thanks to all people that contributed to collect the data across the years, Pierre Alounawalé, Lucienne Taloekaïdoe, Henry Civette, Gérard Jean-Baptiste, Thierry Yawalou, Luc Lassouka, Yves Kouyouli, Jean-Michel Miso, René Maïpouri, Loïc Zidock, Laurent Pilauku, Jean-François Maillard, and Manfred Ulitzka. We are grateful to referees who helped enhancing the first draft of this work.
References
Alvard, M. S., Robinson, J. G., Redford, K. H., and Kaplan, H. (1997). The sustainability of subsistence hunting in the neotropics. Conserv. Biol. 11, 977–982. doi: 10.1046/j.1523-1739.1997.96047.x
Alves, R. R. N., and Van Vliet, N. (2018). “Wild Fauna on the menu,” in Ethnozoology: Animals in Our Lives, eds R. N. N. Aves and U. P. Albuquerque (Oxford, UK: Academic Press), 167–194. doi: 10.1016/B978-0-12-809913-1.00010-7
Benítez-López, A., Alkemade, R., Schipper, A. M., Ingram, D. J., Verweij, P. A., Eikelboom, J. A. J., et al. (2017). The impact of hunting on tropical mammal and bird populations. Science 356:6334. doi: 10.1126/science.aaj1891
Berkes, F., Colding, J., and Folke, C. (2000). Rediscovery of traditional ecological knowledge as adaptive management. Ecol. Appl. 10, 1251–1262. doi: 10.1890/1051-0761(2000)010[1251:ROTEKA]2.0.CO;2
Bodmer, R. E. (1995). “Susceptibility of mammals to overhunting in amazonia,” in Integrating People and Wildlife for a Sustainable Future, eds J. Bissonnette and P. R. Krausman (Bethesda, MD: The Wildlife Society, 292–295.
Bodmer, R. E., Eisenberg, J. F., and Redford, K. H. (1997). Hunting and the likelihood of extinction of Amazonian mammals. Conserv. Biol. 11, 460–466. doi: 10.1046/j.1523-1739.1997.96022.x
Constantino, P. A. L. (2015). Dynamics of hunting territories and prey distribution in Amazonian Indigenous Lands. Appl. Geogr. 56, 222–231. doi: 10.1016/j.apgeog.2014.11.015
Constantino, P. A. L. (2016). Deforestation and hunting effects on wildlife across Amazonian indigenous lands. Ecol. Soc. 21:3. doi: 10.5751/ES-08323-210203
Constantino, P. A. L., Benchimol, M., and Antunes, A. P (2018). Designing Indigenous Lands in Amazonia: securing indigenous rights and wildlife conservation through hunting management. Land Use Policy 77, 652–660. doi: 10.1016/j.landusepol.2018.06.016
da Silva Chaves, L., Caselli, C. B., Nascimiento, A., and Alves, R. R. N. (2019). “Techniques to evaluate hunting sustainability,” in Methods and Techniques in Ethnobiology and Ethnoecology, eds U. P. Albuquerque, L. V. F. C. da Cunha, R. F. P. De Lucena, and R. R. N. Alves (New York, NY: Humana Press), 323–333. doi: 10.1007/978-1-4939-8919-5_21
Davy, D., Tritsch, I., and Grenand, P. (2012). Construction et restructuration territoriale chez les Wayãpi et Teko de la commune de Camopi, Guyane française. Revista Franco-Brasilera de Geografia 16. doi: 10.4000/confins.7964
de Oliveira, J. V., de faria Lopes, S., Barboza, R. R. D., and Alves, R. R. N. (2018). To preserve, or not to preserve, that is the question: urban and rural student attitudes towards wild vertebrates. Environ. Dev. Sustain. 21, 1271–1289. doi: 10.1007/s10668-018-0083-5
De souza-Mazurek, R. R., Pedrhino, T., FEliciano, X., Hilario, W., Geroncio, S., and Marcelo, E. (2000). Subsistence hunting among the Waimiri Atroari Indians in central Amazonia, Brazil. Biodivers. Conserv. 9, 579–596. doi: 10.1023/A:1008999201747
Dezécache, C., Faure, E., Gond, V., DSalles, J. M., Vieilledent, G., and Hérault, B. (2017). Gold-Rush in a forested El Dorado: deforestation leakages and the need for regional cooperation. Environ. Res. Lett. 12:034013. doi: 10.1088/1748-9326/aa6082
FAO (2010). FAO, Global Forest Resources Assessment. Available online at: www.fao.org
Filoche, G., Davy, D., Guigner, A., and Armanville, F. (2017). La construction de l'État français en Guyane à l'épreuve de la mobilité des peuples amérindiens. Crit. Int. 75, 71–88. doi: 10.3917/crii.075.0071
Grenand, P. (1980). Introduction à l'univers Wayãpi. Ethnoécologie des indiens du Haut Oyapock. (Guyane Française). Paris: Ste d'Etude Linguistiques et Anthropologiques de France SELAF.
Grenand, P. (1995). “De l'arc au fusil. Un changement technologique chez les Wayãpi de Guyane,” in Transitions Plurielles. Exemples dans Quelques Sociétés des Amériques, eds F. Grenand and V. Randa (Paris: Peeters, 23–53.
Grenand, P. (2002). La Chasse en Guyane: vers une Gestion Durable. SILVOLAB- Programme Ecosystèmes Tropicaux.
Grenand, P., and Grenand, F. (1996). ≪ II ne faut pas trop en faire ≫ Connaissance du vivant et gestion de l'environnement chez les Wayapi (Amérindiens de Guyane). Cahier Sci. Hum. 32, 51–63.
Hames, R. B., and Vickers, W. T. (1982). Optimal diet theory as a model to explain variability in Amazonian hunting. Am. Ethnol. 9, 358–378. doi: 10.1525/ae.1982.9.2.02a00090
Hammond, D. S., Gond, V., de Thoisy, B., Forget, P.-M., and DeDijn, B. P. E. (2007). Causes and consequences of a tropical forest gold rush in the Guiana Shield, South America. Ambio 36, 661–670. doi: 10.1579/0044-7447(2007)36[661:CACOAT]2.0.CO;2
Haugaasen, T., and Peres, C. A. (2005). Mammal assemblage structure in Amazonian flooded and unflooded forests. J. Trop. Ecol. 21, 133–145. doi: 10.1017/S026646740400207X
Hill, K., McMillan, G., and Farina, R. (2003). Hunting-related changes in game encouter rates from 1994 to 2001 in the Mbaracayu Reserve, Paraguay. Conserv. Biol. 17, 1312–1323. doi: 10.1046/j.1523-1739.2003.01135.x
Hill, K., and Padwe, J. (2000). “Sustainability of Aché hunting in the Mbaracayu Reserve, Paraguay,” in Hunting for Sustainability in Tropical Forests, eds J. G. Robinson and E. L. Bennett (New York, NY: Columbia University Press, 79–105.
Hill, K., Padwe, J., Bejyvagi, C., Bepurangi, A., Jakugi, F., Tykuarangi, R., et al. (1997). Impact of hunting on large vertebrates in the Mbaracayu reserve, Paraguay. Conserv. Biol. 11, 1339–1353. doi: 10.1046/j.1523-1739.1997.96048.x
Hurtado-Gonzales, J. L., and Bodmer, R. (2004). Assessing the sustainability of brockett deer hunting in the Tamshiyacu-Tahuayo Communal Reserve, northeastern Peru. Biol. Conserv. 116, 1–7. doi: 10.1016/S0006-3207(03)00167-8
INSEE (2012). Recensements de la Population. Populations Légales en Vigueur à Compter du 1er Janvier 2013. www.insee.fr. Available online at: www.insee.fr
Jerozolimski, A., and Peres, C. A. (2003). Bringing home the biggest bacon: a cross-site analysis of the structure of hunter-kill profiles in Neotropical forests. Biol. Conserv. 111 415–425. doi: 10.1016/S0006-3207(02)00310-5
Keith, D. A., Martin, T. G., McDonald-Madden, E., and Walters, C. (2011). Uncertainty and adaptive management for biodiversity conservation. Biol. Conserv. 144, 1175–1178. doi: 10.1016/j.biocon.2010.11.022
Laurance, W. F., Peletier-Jellema, A., Geenen, B., Harko Koster, H., Verweij, P., Van Dijck, P., et al. (2015). Reducing the global environmental impacts of rapid infrastructure expansion. Curr. Biol. 25, R255–R268. doi: 10.1016/j.cub.2015.02.050
Levi, T., Lu, F., Yo, D. W., and Mangel, M. (2011a). The behaviour and diet breadth of central-place foragers: an application to human hunters and Neotropical game management. Evol. Ecol. Res. 13, 171–185. Available oline: http://www.evolutionary-ecology.com/abstracts/v13/2663.html
Levi, T., Shepard, G. H., Ohl-Schacherer, J., Wilmers, C. C., Peres, C. A., and Yu, D. W. (2011b). Spatial tools for modeling the sustainability of subsistence hunting in tropical forests. Ecol. Appl. 21, 1802–1818. doi: 10.1890/10-0375.1
Levi, T., Shepard, G. H. J., Ohl-Schacherer, J., Peres, C. A., and Yu, D. W. (2009). Modelling the long-term sustainability of indigenous hunting in Manu National Park, Peru: landscape-scale management implications for Amazonia. J. Appl. Ecol. 46, 804–814. doi: 10.1111/j.1365-2664.2009.01661.x
McNamara, J., Kusimi, J. M., Rowcliffe, J. M., Cowlishaw, G., Brenyah, A., and Milner-Gulland, E. J. (2015). Long-term spatio-temporal changes in a West African bushmeat trade system. Conserv. Biol. 29, 1446–1457. doi: 10.1111/cobi.12545
Milner-Gulland, E. J., and Akçakaya, H. R. (2001). Sustainability indices for exploited populations. Trends Ecol. Evol. 16, 686–692. doi: 10.1016/S0169-5347(01)02278-9
Morellet, N., Gaillard, J.-M., Hewison, A. J. M., Ballon, P., Boscardin, Y., Duncan, P., et al. (2007). Indicators of ecological change: new tools for managing populations of large herbivores. J. Appl. Ecol. 44, 634–643. doi: 10.1111/j.1365-2664.2007.01307.x
Novaro, A. J., Funes, M. C., and Walker, S. (2005). An empirical test of source–sink dynamics induced by hunting. J. Appl. Ecol. 42, 910–920. doi: 10.1111/j.1365-2664.2005.01067.x
Novaro, A. J., Redford, K. H., and Bodmer, R. E. (2000). Effect of hunting in source-sink systems in the neotropics. Conserv. Biol. 14, 713–721. doi: 10.1046/j.1523-1739.2000.98452.x
Ohl-schacherer, J., Shepard, G. H. J., Kaplan, H., Peres, C. A., Levi, T., and Yu, D. W. (2007). The sustainability of subsistence hunting by matsigenka native communities in Manu National Park, Peru. Conserv. Biol. 21, 1174–1185. doi: 10.1111/j.1523-1739.2007.00759.x
ONF (2016). Carte D'occupation des sol sur la Bande Littorale Guyanaise en 2015. GeoGuyane. Available online at: https://www.geoguyane.fr/geonetwork/srv/0906df56-3bf2-4df3-8ad8-645df6f03308
Ouhoud-Renoux, F. (1998a). De l'outil à la prédation. Technologie culturelle et ethno-écologie chez les Wayãpi du haut Oyapock (Guyane Française). Ph.D, Paris X.
Ouhoud-Renoux, F. (1998b). Se nourrir à Trois sauts. Analyse diachronique de la prédation chez les wayapis du haut-Oyapock(Guyane Française). JATBA 40, 181–206. doi: 10.3406/jatba.1998.3668
Parry, L., Barlow, J., and Peres, C. A. (2009a). Allocation of hunting effort by Amazonian smallholders: implications for conserving wildlife in mixed-use landscapes. Biol. Conserv. 142, 1777–1789. doi: 10.1016/j.biocon.2009.03.018
Parry, L., Barlow, J., and Peres, C. A. (2009b). Hunting for sustainability in tropical secondary forests. Conserv. Biol. 23, 1270–1280. doi: 10.1111/j.1523-1739.2009.01224.x
Peres, C. A. (1997). “Evaluating the sustainability of subsistence hunting in tropical forests,” in CSERGE Working Papers GEC 97-22 (London), 1–39.
Peres, C. A. (2000). Effects of subsistence hunting on vertebrate community structure in Amazonian forests. Biol. Conserv. 14, 240–253. doi: 10.1046/j.1523-1739.2000.98485.x
Peres, C. A., and Palacios, E. (2007). Basin-wide effects of game harvest on vertebrate population densities in Amazonian forests: implications for animal-mediated seed dispersal. Biotropica 39, 304–315. doi: 10.1111/j.1744-7429.2007.00272.x
Redford, K. H., and Robinson, J. G. (1987). The game of choice: patterns of indian and colonist hunting in the neotropics. Am. Anthropol. 89, 650–667. doi: 10.1525/aa.1987.89.3.02a00070
Renoux, F., and de Thoisy, B. (2016). Hunting management: the need to adjust predictive models to field observations. Ethnobiol. Conserv. 5:1. doi: 10.15451/ec2016-6-5.1-1-13
Richard-Hansen, C., Ackermann, L., Berzins, R., Devillechabrolle, J., Foch, T., Goguillon, B., et al. (2018). The return of White-lipped peccaries in two main protected areas of French Guiana: update and insights into an uncommon population dynamics. Suiform Soundings 17, 37–40.
Richard-Hansen, C., Berzins, R., Petit, M., Rux, O., Goguillon, B., and Clément, L. (2019). “Movements of white-lipped Peccary in French Guiana,” in Movement Ecology of Neotropical Forest Mammals, eds R. Reyna-Hurtado and C. A. Chapman (Cham: Springer Nature), 57–75. doi: 10.1007/978-3-030-03463-4_5
Richard-Hansen, C., and Hansen, E. (2004). “Hunting and wildlife management in French Guiana : current aspects and future prospects,” in People in Nature: Wildlife Conservation in south and central America, eds K. M. Silvius, R. E. Bodmer and J. M. V. Fragoso (New York, NY: Columbia University Press), 400–410. doi: 10.7312/silv12782-025
Richard-Hansen, C., Vié, J. C., Vidal, N., and Keravec, J. (1999). Biometrical data on 40 species of mammals from French Guiana. J. Zool. 247, 419–428. doi: 10.1111/j.1469-7998.1999.tb01005.x
Robinson, J. G. (2000). “Calculating Maximum Sustainable Harvests and Percentage offtakes.,” in Hunting for Sustainability in Tropical Forests, eds J. G. Robinson and E. L. Bennett (New York, NY: Columbia University Press, 521–524.
Robinson, J. G., and Redford, K. H. (1986). Intrinsic rate of natural increase in neotropical forest : relationships to phylogeny and diet in mammals. Oecologia 68, 516–520. doi: 10.1007/BF00378765
Robinson, J. G., and Redford, K. H. (1994). Measuring the sustainability of hunting in neotropical forests. Oryx 28, 249–256. doi: 10.1017/S0030605300028647
Robinson, J. G., and Redford, K. H. (eds.). (1991). “Sustainable harvest of neotropical forest mammals,” in Neotropical Wildlife Use and Conservation, (Chicago: University of Chicago Press, 415–429.
Rowcliffe, J. M., Cowlishaw, G., and Long, A. J. (2003). A model of human hunting impacts in multi-prey communities. J. Appl. Ecol. 40, 872–889. doi: 10.1046/j.1365-2664.2003.00841.x
Salas, E., and Kim, J. B. (2002). Spatial factors and stochasticity in the evaluation of sustainable hunting of tapirs. Conserv. Biol. 16, 86–96. doi: 10.1046/j.1523-1739.2002.00225.x
Sarti, F. M., Adams, C., Morsello, C., Van Vliet, N., Schor, T., Yagüe, B., et al. (2015). Beyond protein intake: bushmeat as source of micronutrients in the Amazon. Ecol. Soc. 20:22. doi: 10.5751/ES-07934-200422
Shaffer, C. A., Milstein, M. S., Suse, P., Marawanaru, E., Yukuma, C., Wolf, T. M., et al. (2018a). Integrating ethnography and hunting sustainability modeling for primate conservation in an Indigenous Reserve in Guyana. Int. J. Primatol. 39, 945–968. doi: 10.1007/s10764-018-0066-2
Shaffer, C. A., Milstein, M. S., Yukuma, C., Marawanaru, E., and Suse, P. (2017). Sustainability and comanagement of subsistence hunting in an indigenous reserve in Guyana. Conserv. Biol. 31, 1119–1131. doi: 10.1111/cobi.12891
Shaffer, C. A., Yukuma, C., Marawanaru, E., and Suse, P. (2018b). Assessing the sustainability of Waiwai subsistence hunting in Guyana by comparison of static indices and spatially explicit, biodemographic models. Anim. Conserv. 21, 148–158. doi: 10.1111/acv.12366
Shepard, G. H. Jr., Levi, T., Goes Neves, E., Peres, C. A., and Yu, D. W. (2012). Hunting in ancient and modern Amazonia: rethinking sustainability. Am. Anthropol. 114, 652–667. doi: 10.1111/j.1548-1433.2012.01514.x
Siren, A., Cardenas, J.-C., Hambäck, P., and Parvinen, K. (2012). Distance Friction and the Cost of Hunting in Tropical Forests. Bogota: Ediciones Uniandes.
Siren, A., Hamback, P., and Machoa, J. (2004). Including spatial heterogeneity and animal dispersal when evaluating hunting: a model analysis and an empiracal assessment in an amazonian community. Conserv. Biol. 18, 1315–1329. doi: 10.1111/j.1523-1739.2004.00024.x
Siren, A., and Wilkie, D. S. (2015). The effects of ammunition price on subsistence hunting in an Amazonian village. Oryx 50, 47–55. doi: 10.1017/S003060531400026X
Slade, N. A., Gomulkiewicz, R., and Alexander, H. M. (1998). Alternatives to Robinson and redford's method of assessing overharvest from incomplete demographic data. Conserv. Biol. 12, 148–155. doi: 10.1046/j.1523-1739.1998.96273.x
Stearman, A. M., and Redford, K. H. (1995). Game management and cultural survival: the Yuquí ethnodevelopment Project in lowland Bolivia. Oryx 29, 29–33. doi: 10.1017/S0030605300020846
Suárez, E., Stallings, J., and Suárez, L. (1995). Small-mammal hunting by two ethnic groups in north-western Ecuador. Oryx 29, 35–41. doi: 10.1017/S0030605300020858
Taber, A., Chalukian, S. C., Altrichter, M., Minkowski, K., Lizárraga, L., Sanderson, E., et al. (2008). El Destino de los Arquitectos de los Bosques Neotropicales: Evaluacion de la Distribucion y el Estado de Conservation de los Pecaries Labiados y los Tapires de Tierras Bajas Wildlife Conservation Society/Tapir Specialist Group/Wildlife Trust/Grupo Especialista de la CSE/UICN en Cerdos, Pecaries e Hipopotamos.
Takashina, N., and Mougi, A. (2015). Maximum sustainable yields from a spatially-explicit harvest model. J. Theor. Biol. 383, 87–92. doi: 10.1016/j.jtbi.2015.07.028
Tritsch, I., Marmoex, C., Davy, D., Thibaut, B., and Gond, V. (2015). Toward a revival of indigenous mobility in French Guiana? Contemporary transformations of the Wayãpi and Teko territories. Bull. Latin Am. Res. 34, 19–34. doi: 10.1111/blar.12204
Van Vliet, N. (2018). “Bushmeat crisis” and “cultural imperialism” in wildlife management? Taking value orientations into account for a more sustainable and culturally acceptable wildmeat sector. Front. Ecol. Evol. 6:112. doi: 10.3389/fevo.2018.00112
Van Vliet, N., Milner-Gulland, E. J., Bousquet, F., Saqalli, M., and Nasi, R. (2010). Effect of small-scale heterogeneity of prey and hunter distributions on the sustainability of bushmeat hunting. Conserv. Biol. 24, 1327–1337. doi: 10.1111/j.1523-1739.2010.01484.x
Van Vliet, N., Moreno, J., Gómez, J., Zhou, W., Fa, J. E., Golden, C., et al. (2017). Bushmeat and human health: assessing the evidence in tropical and sub-tropical forests. Ethnobiol. Conserv. 6:3. doi: 10.15451/ec2017-04-6.3-1-45
Van Vliet, N., Muhindo, J., Nyumu, J. K., Mushagalusa, O., and Nasi, R. (2018). Mammal depletion processes as evidenced from spatially explicit and temporal local ecological knowledge. Trop. Conserv. Sci. 11, 1–15. doi: 10.1177/1940082918799494
Van Vliet, N., and Nasi, R. (2008). Why do models fail to assess properly the sustainability of duiker (Cephalophus spp.) hunting in Central Africa? Oryx 42, 392–399. doi: 10.1017/S0030605308000288
Keywords: sustainable hunting, French Guiana, indices, livelihoods, diachronic
Citation: Richard-Hansen C, Davy D, Longin G, Gaillard L, Renoux F, Grenand P and Rinaldo R (2019) Hunting in French Guiana Across Time, Space and Livelihoods. Front. Ecol. Evol. 7:289. doi: 10.3389/fevo.2019.00289
Received: 31 January 2019; Accepted: 18 July 2019;
Published: 08 August 2019.
Edited by:
Nathalie Van Vliet, Center for International Forestry Research, IndonesiaReviewed by:
Pratheesh C. Mammen, Institute for Climate Change Studies, IndiaPedro De Araujo Lima Constantino, Independent Researcher, Brazil
Copyright © 2019 Richard-Hansen, Davy, Longin, Gaillard, Renoux, Grenand and Rinaldo. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Cecile Richard-Hansen, Y2VjaWxlLnJpY2hhcmQtaGFuc2VuQGVjb2ZvZy5nZg==