 Rebecca L. Holberton1*
Rebecca L. Holberton1* Philip D. Taylor2,3Lindsay M. Tudor4Kathleen M. O'Brien5Glen H. Mittelhauser6Ana Breit7
Philip D. Taylor2,3Lindsay M. Tudor4Kathleen M. O'Brien5Glen H. Mittelhauser6Ana Breit7- 1Laboratory of Avian Biology, School of Biology and Ecology, University of Maine, Orono, ME, United States
- 2Department of Biology, Acadia University, Wolfville, NS, Canada
- 3Bird Studies Canada, Port Rowan, ON, Canada
- 4Bird Group, Maine Department of Inland Fisheries and Wildlife, Bangor, ME, United States
- 5United States Fish and Wildlife Service–Rachel Carson National Wildlife Refuge, Wells, ME, United States
- 6Maine Natural History Observatory, Gouldsboro, ME, United States
- 7School of Biology and Ecology, University of Maine, Orono, ME, United States
The Gulf of Maine has long been recognized as a major stopover area for shorebirds in fall. Knowing how birds move within and beyond the region will be paramount to protecting threatened shorebird habitat. To determine stopover behavior during fall migration (2013–2017) in Maine, 180 (104 AHY, 76 HY) Semipalmated Sandpipers, Calidris pusilla, were tracked using VHF radiotelemetry and an extensive array of automated receivers (Motus Wildlife Tracking System). Birds tagged at three locations along the Maine coastline showed no effect of age class or stopover site on body condition (body mass, estimated fat mass) or stopover length (post-capture detection period). However, movement after departure varied greatly among sites. Few birds captured at the northern-most site (“Downeast,” n = 71), which had the greatest amount of mudflats and offshore roost sites and the least amount of human disturbance, were detected beyond the initial tagging location, suggesting that they, like birds in the Bay of Fundy just to the north, initiated trans-oceanic flights from that location. At the Downeast site, leaner birds remained significantly longer than fatter birds, suggesting that time of departure there depended on energy reserves, which would be critical for making extensive flights. In contrast, over half of the birds tagged further south (Popham Beach, n = 59; Rachel Carson NWR, n = 50) were later detected at coastal locations to the north (few) or to the south (most). Stopover period at these sites was independent of fat, suggesting that other factors (e.g., feeding/roosting site availability, human activity) influenced departure decisions. In Maine, Semipalmated Sandpipers, regardless of age, may move north (Downeast) or south (e.g., Cape Cod, Rhode Island, Long Island Sound) where the local topography, habitat characteristics (feeding/roosting sites), and/or lower human activity, may best enable them to initiate trans-oceanic flights to the wintering grounds. Future study should determine if variation in stopover behavior is population-specific and if population-segregation occurs in Maine. Use of automated VHF radiotelemetry has led to a greater understanding of stopover behavior and the degree of connectivity among stopover sites, which should be taken into account for conserving migratory bird habitat across broad spatial scales.
Introduction
Many Arctic-breeding shorebird populations have declined rapidly over the past few decades, with some populations decreasing by as much as 60% (for review, see 2012 North American Bird Conservation Initiative Report; U.S. Shorebird Conservation Plan Partnership, U.S. Shorebirds of Conservation Concern, 2015). Hunting on the wintering grounds and habitat loss experienced throughout the annual cycle are believed to be the primary factors underlying overall species declines (Brown et al., 2017). Within a species, trends for individual breeding populations may differ due to disparate factors that individual breeding populations experience on their respective breeding or wintering areas as well as along population-specific migratory routes. For example, although Semipalmated Sandpipers, Calidris pusilla, breed across the North American Arctic, different breeding populations show a high degree of geographic segregation in their respective wintering areas and migration routes (Andres et al., 2012; Gratto-Trevor et al., 2012a; Brown et al., 2017). Western and central breeding populations appear to be stable or slightly increasing over the past few decades, but eastern Semipalmated Sandpiper populations have shown little to no increase, with some indications that they continue to decline (Andres et al., 2012; Gratto-Trevor et al., 2012b; Morrison et al., 2012; Smith et al., 2012; Brown et al., 2017). This pattern prompted Andres et al. (2012) to propose that eastern breeding populations of this species be considered “of high conservation concern.” Understanding the behavior and ecology of different Semipalmated Sandpiper populations throughout the annual cycle will be key to determining how each breeding population is ultimately regulated and, consequently, how to manage resources to support them.
Recently, using light-level geolocators, Brown et al. (2017) confirmed the degree of geographic segregation of different Semipalmated Sandpiper breeding populations during migration as well as during the stationary periods of breeding and wintering. In spring, individuals from western populations of this species retrace much of their southward journey through the interior of North America as they move north to return to their respective breeding areas. In contrast, eastern-breeding Semipalmated Sandpipers exhibit an “elliptical migration” pattern between the wintering and breeding areas: in spring, these birds depart the wintering grounds in the Caribbean and along the east coast of South America and move northward along the U.S. Atlantic coastline. When most birds reach New Jersey's Delaware Bay, they stage there for as much as several weeks to acquire sufficient energy reserves needed to initiate extended flights that bypass New England on the way to their respective breeding grounds (Gratto-Trevor and Dickson, 1994; Gratto-Trevor et al., 2012a; Figures 2, 3 in Brown et al., 2017). In autumn, while up to 500,000 Semipalmated Sandpipers stage each year in the Bay of Fundy region, as many as 100,000 more arrive along the Gulf of Maine coastline, from the Bay of Fundy to Cape Cod (L. Tudor, unpubl. data, McNeil and Burton, 1977; Fefer and Schettig, 1980; Lank, 1983; Hicklin, 1987; Dunn et al., 1988; Tudor, 2002; Maine Department of Inland Fisheries Wildlife, 2015). Most of these birds rest and refuel in coastal estuaries before they initiate non-stop flights over the North Atlantic to reach the wintering grounds (Gratto-Trevor et al., 2012a; Brown et al., 2017).
Birds preparing for extended non-stop flight require stopover sites that offer abundant high quality food, low predation pressure, and roost sites where they can efficiently rest as they refuel before departing. The Gulf of Maine region (primarily the upper Bay of Fundy and coastal Maine) offers a wide diversity of features that support feeding and roosting, including numerous tidal mudflats, marshes, and beaches, as well as many islands and rocky ledge outcroppings offshore. Unfortunately, suitable shorebird habitat along the entire Atlantic coast continues to be threatened by increased human activities and rapid sea level rise. Understanding shorebird movement within and between stopover areas along the coast, therefore, is key to identifying and managing resources needed by birds during this critical stage of the annual cycle.
Much of what is known about stopover behavior and ecology of Semipalmated Sandpipers during fall migration in eastern North America comes from studies in the Bay of Fundy (McNeil and Burton, 1977; Hicklin, 1987; Mawhinney et al., 1993; Hicklin and Chardine, 2012; Mann et al., 2017); less is known about their movements in the Gulf of Maine region to the south (Dunn et al., 1988). Maine's coastal habitats, in particular, are under increasing pressure from residential and commercial development, coastal engineering, aquaculture, rockweed harvest, threats of sea level rise due to climate change, as well as disturbance issues associated with recreationalists and pet owners, which have recently escalated in Maine (Tyrrell, 2005; Maine Department of Inland Fisheries Wildlife, 2015). Semipalmated Sandpipers are currently listed as a Species of Special Concern and a Priority Species for conservation in Maine (Maine Department of Inland Fisheries Wildlife, 2015). Documenting shorebird behavior during migration in Maine has become a priority for effective conservation of the region's coastal habitats.
To document stopover behavior of individual Semipalmated Sandpipers in Maine, we capitalized on the international network of automated VHF radiotelemetry stations recently established as the Motus Wildlife Tracking System (described in Taylor et al., 2017). We first initiated a study at one location that, through an extensive collaboration among federal and state resource management agencies, several NGOs, and private landowners, subsequently led to an opportunity to add two more sites that varied in local topography as well as in the intensity of human activities (e.g., shellfish harvesting, beach recreation) during fall (July–October) migration (see Figure 1). The locations included a relatively undisturbed site in northern Maine (Pleasant Bay in Downeast Maine), a state-owned beach with extensive recreational use in mid-coast Maine (Popham Beach State Park), and a site situated within the National Wildlife Refuge system in southern Maine (Rachel Carson National Wildlife Refuge) that has limited human recreational use of shorebird habitat during fall migration (Figure 1).
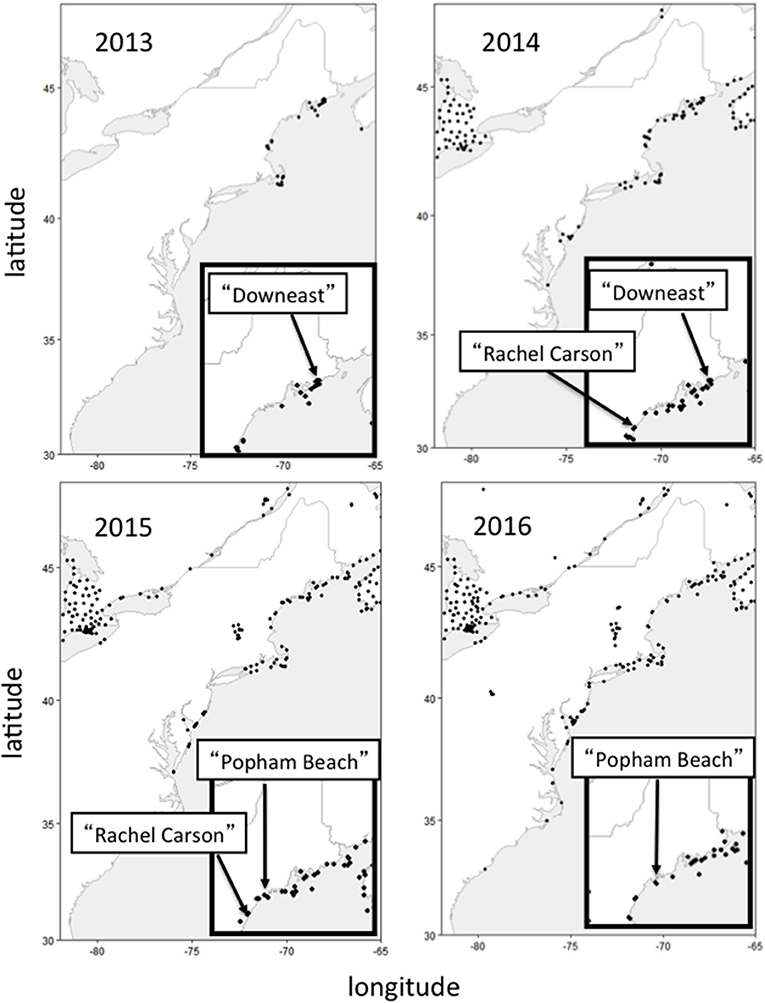
Figure 1. Location (solid circles) of individual automated VHF telemetry towers, comprising the Motus Wildlife Tracking Array, that were active during each year in which tagging activities occurred in Maine. Insert maps show the specific locations (see text for detailed description) where Semipalmated Sandpipers were captured and tagged each year (see text for detailed description of each site's topography, levels of human activity, and the towers designated as “local”).
Our objectives were to determine if groups of individuals using these sites differed in morphological characteristics, which might indicate spatial segregation by different breeding populations, as this could be used to understand changes in bird abundance in Maine possibly reflecting the disparate trends reported for different breeding populations (Andres et al., 2012; Gratto-Trevor et al., 2012b; Morrison et al., 2012; Smith et al., 2012; Brown et al., 2017). During the entire study, multiple automated receiver towers within the Motus array extended along the Atlantic coastline from Atlantic Canada to as far south as the Carolinas, a flyway that includes major shorebird staging areas during fall migration (Figure 1). The extensive array of automated towers allowed us to examine stopover length (as estimated by the post-capture length of stay at the tagging location) and subsequent regional-scale movements (beyond the initial capture locations) of individuals on stopover at these different sites.
We tested several hypotheses made before and after data were collected from all three locations. We first examined age-related differences in body condition and stopover length. As young birds on their first migration may not be as efficient in foraging or in predator detection (Cresswell, 1994; Fernandez and Lank, 2006; Stillman et al., 2007; van den Hout et al., 2017), we predicted, a priori, that young birds would have lower energy reserves at the time of capture and/or longer stopover periods to rest and refuel, compared to adults. We also predicted, a priori, that, regardless of age, birds with greater energy reserves at the time of capture would be more ready to depart (have shorter post-capture detection period) than birds with less fat (c.f. Williams et al., 2007). Finally, the ability to opportunistically compare bird movements at three sites in Maine allowed us to examine, a posteriori, potential differences in stopover behavior related to site-based differences in key topographical features (geographic location with respect to a goal, tidal mudflats vs. sandy beach, human disturbance level). We assumed throughout the analyses that age, body condition, and stage of stopover did not influence the likelihood of a bird being captured.
Materials and Methods
Tagging Locations, Automated Radiotelemetry System, and Tower Deployment
Bird capture and tagging activities, and automated receiver tower deployments were undertaken at three locations along the Maine coast designated as: (1) “Downeast” (2013–14, Figure 1), a relatively undisturbed site in Pleasant Bay, Downeast Maine that offers extensive feeding areas during low tide and many roost sites on rocky outcroppings offshore, (2) “Popham” (2015–16, Figure 1), a public recreation area at Popham Beach State Park in mid-coast Maine that is used extensively by recreational beach-goers and hikers throughout the tidal cycle, and (3) “Rachel Carson” (2014–15, Figure 1), situated within the Rachel Carson National Wildlife Refuge in southern Maine that offers limited recreational use near where shorebirds feed on exposed mudflats during low tide but experiences extensive recreational use along sandy beaches where shorebirds are seen throughout the tidal cycle.
At each tagging location, we deployed two or more automated receiver stations that contributed to the Motus array. Motus towers designated as “local” for each of the three tagging locations, as well as additional towers that were active along the coastline from the Bay of Fundy to South Carolina during the study periods for each year of the studies, are shown in Figure 1. All components of the telemetry system, including descriptions of the VHF radiotags and automated receivers (“Sensorgnomes”) are described by Taylor et al. (2017), and detailed maps of all tower locations for each year are available on the Motus web page (www.motus.org). The towers and associated antennas at each tagging location were positioned with respect to local topography to maximize detection of tagged birds moving within and beyond the sites. Each VHF tag was confirmed to be operating when it was deployed. In addition to the tag deployment days, towers were checked every 2–4 weeks during and for several months after the region's fall migration period (approx. early July through early November) to confirm operation and to collect detection data for processing (described below). Data from our tagged birds detected beyond our local tagging sites were processed by Motus (www.motus.org), downloaded using the “tagme” function in the Motus R package, and post-processed as described in Crewe et al. (2018).
“Downeast,” Maine (Washington County, 44.55° N, 67.80° W)—From July through November, 2013 and 2014, we deployed two automated radiotelemetry receiver towers in the Pleasant Bay region within 5 km of the capture locations (Figure 1). This area includes the estuaries of the Mill, Harrington, and Pleasant Rivers and contains important shorebird habitat during fall migration. Extensive tidal mudflats associated with the numerous creeks and river outflows provide valuable feeding areas during low tide. In addition to vegetated saltpans, numerous rocky ledges offshore provide roosting sites during high tide. The area surrounding the study site is rural and not heavily developed. Human disturbance is low as there is limited sandy beach and, during the fall migration period, the exposed tidal mudflats are used for limited shellfish harvesting by hand. In both years, one telemetry station (Pineo Point, Washington County: “PINEO”: 44.5645o N, 67.8066o W) was set up immediately adjacent to extensive mudflats exposed during low tides at the southern tip of a peninsula located between the Mill and Harrington Rivers. Only one antenna (174o) was deployed on the PINEO tower in 2013. Three antennas (195, 151, 243o) were deployed on the same tower in 2014. In both years, a second tower was set up at Seal Cove (“SECO”: 44.5431o N, 67.7528o W) south and east from Pineo Point, on the eastern side of Pleasant Bay. Only one antenna was deployed on the SECO tower in 2013 (239o) and three were deployed in 2014 (225, 263, 333o). Several additional towers deployed and operated by USFWS-Maine Coastal Islands National Wildlife Refuge and other Motus participants in 2013 and 2014 were included as part of the array designated for this study as “local”. These were located at the southern opening of Pleasant Bay: Nash Island (“NASH” 2013–14: 44.4648o N, 67.746o W; 2013–14) and Jordan's Delight (“JORDL” 2013: 44.44269o N, 67.8241o W), on the nearby Petit Manan Peninsula just to the east (PMP 2013 = PMP1 2014: 44.4131o N, 67.9058o W; PMP_WL 2013: 44.4015o N, 67.8965o W; PMP2 2014: 44.4085o N, 67.9050o W), and on Petit Manan Island about 10 km to the southeast of Pleasant Bay (PMI 2013: 44.5385o N, 67.8805o W). Additional towers, considered outside the “local” tagging location, were deployed in 2013–14 (by multiple Motus participants) along the coast to the north and east and at locations on the mainland and on offshore islands to the southwest (Figure 1).
“Popham,” Popham Beach State Park (Sagadahoc County, 43.4417o N, 69.4759o W)—This public beach area is bordered by the mouth of the Kennebec River to the north and the Morse River to the south. Only one telemetry tower was deployed in 2015 (“POPH”: 43.736o N, 69.7997o W; 2015 antenna directions = 128, 164, 224o). This tower was redeployed, at the same location, in 2016 (2016 antenna directions = 84, 159, 244o) along with a new tower (“SEGUIN”: 43.7099 o N, 69.7596o W; antenna directions = 224, 278, 344o) set up on Seguin Island, one of several small rocky offshore islands within 1–2 kilometers east of the beach. These two towers, designated as “local” for this study, collectively provided extensive detection coverage in the immediate area, including the sandy beach and the small tidal lagoon behind it at the mouth of the Morse River, Seawall Beach just to the south, and the few rocky ledge outcroppings just offshore from these areas. Coverage also extended to the local area's tidal marsh inlets. The Park provides activities for the public such as swimming, kayaking, fishing, picnicking, and hiking and is considered the most visited public state park in Maine during the summer and fall. Dogs are not allowed on the beach 1 April−30 September to protect nesting seabirds and shorebirds as well as birds on migration. Additional towers outside the “local” tagging location were deployed in 2015–16 (by multiple Motus participants) along the coast to the north and east and along the coast to the south (Figure 1).
“Rachel Carson,” Rachel Carson National Wildlife Refuge (RCNWR, 43.2100° N, 70.3228° W, York and Cumberland Counties)–This study area includes the Webhannet, Little, and Mousam Rivers, with estuarine areas dominated by tidal mudflats and saltmarsh. The sandy beaches are heavily visited by people, particularly during the late summer months that overlap with shorebird migration (Aug.–Sept.). Pedestrian access to the estuarine areas (where birds were captured during low tide) is limited and human disturbance in those areas is generally low. Unlike the Downeast and Popham Beach locations, the Refuge area lacks offshore roosting sites.
Two telemetry towers, considered “local” for this study, were each deployed in 2014 and 2015: “FURBISH” (43.2819° N, 70.5817° W, 2014 antenna directions = 17, 197, 85, 260°; 2015 antenna directions = 18, 85, 197°, and “WNERR” (43.3351° N, 70.5491° W, 2014 antenna directions = 223°; 2015 antenna directions = 145, 240, 281°). Additional towers, considered outside the “local” tagging location, were deployed during the 2014–15 season (by multiple Motus participants) along the coast to the north and east and along the coast to the south (Figure 1).
Capture and Handling
All birds were captured during daylight hours and most birds were captured by mist net while feeding on exposed mudflats. The 2–3 h period of rising tide leading to peak high tide provided the greatest capture rates as birds concentrated in large numbers on exposed feeding areas as these were being gradually submerged by the rising water. During the 6 h surrounding the period of peak high tide, such feeding areas were entirely unavailable and birds either moved to feeding areas further upstream in tidal marshes or moved to available roost sites on exposed rocky ledges offshore or on sandy beaches along the mainland (as confirmed with hand-held telemetry receivers). Roosting birds were captured by mist nets on the beach during the high tide period at Popham Beach and at Rachel Carson NWR and by a net gun (rifle cartridge or CO2 powered) deployed from the bow of a small boat as birds roosted on exposed rocky ledges offshore at the Downeast site.
Birds were handled by crews that worked at more than one site and an attempt was made to standardize measuring techniques among different crew members. Regardless of capture method, birds were immediately placed in small cloth bags until processed for body mass (with a hanging Pesola spring scale or table balance, to nearest 0.5 g), wing length (flattened wing to nearest 0.5 mm), and culmen length (to nearest 0.5 mm) and to an age class (adult: After Hatching Year = AHY; juvenile: Hatching Year = HY; Unknown = U) based on plumage characteristics (Pyle, 1997). We also estimated the amount of lipid energy stores independent of body size, using a formula developed by Dunn et al. (1988) based on Semipalmated Sandpipers collected in Maine: estimated fat mass = total body mass – fat free mass; fat free mass = (−9.0513 + [0.3134 X wing length]). Estimated fat mass was not available for three birds because either wing length or body mass was not recorded. After processing, each bird was banded with a uniquely numbered aluminum USGS band, a pair of color bands that indicated they were banded in Maine, and a green 3-character plastic flag band for individual identification that could be read from afar.
We attached coded VHF radiotags (Lotek model NTQB-2), using a small drop of quick drying “super glue” to a few clipped body feathers along the back just above the “rump.” Tags were applied only to birds weighing at least 20 g but no more than 34 g so as to increase the opportunity to monitor bird movements during the period leading up to departure from the immediate area. Although this may have produced a sample population not truly representative of the groups of birds on stopover at these areas, birds weighing <20 g may not have been doing well (and thus may have been influenced by tagging), and those above 34 g may have been more likely to depart the site too soon after tagging to reveal much information about movement within the stopover area. As with many shorebird species, adult Semipalmated Sandpipers pass through the region in fall earlier than young of the year and capture efforts were targeted to yield an equal number of each age class at each site each year if weather conditions allowed.
All activities related to bird capture and handling were reviewed and approved by UMaine IACUC (#A2013-01-02 to RLH) and were performed under federal and state permits to U.S. Fish & Wildlife Service (to KMO) and Maine Department of Inland Fisheries and Wildlife (to LMT).
Data Handling and Analyses
Detection data collection, storage, and extraction are described in Taylor et al. (2017) and Crewe et al. (2018). Detection data (initial and final timestamp of individual VHF coded signals) for each bird were compiled into a spreadsheet containing information as to age class, body mass, and body size measurements. As our proxy for length of stopover at each of the three study locations, we determined the period of time between the release of the bird at the time of tag attachment and its final detection by any one of the antennas mounted on towers designated as “local” at each of the three sites. For the model of the nanotags used throughout the study, each tower had an estimated detection radius of up to 10 km under ideal conditions.
Statistical analyses were done using R, version 3.3.0 (2016-03-10), Copyright © 2016, The R Foundation for Statistical Computing, on a Windows 10 PC. For all analyses, we fit generalized linear models (using the glm function in base R) with “site” and “age” as categorical predictors. For culmen, wing length, mass, and estimated fat mass (EFM), we fit models with an identity link and Gaussian errors. For the amount of time spent at the local site after being tagged and released (post-capture detection period, PCDP) we fit a model with an identity link and Gaussian errors, but we transformed the response using a square root transformation (to improve fit by reducing the influence of long-staying individuals on the parameter estimates). For the model assessing whether a bird remained at the initial tagging area, or disappeared completely from the initial tagging area (suggesting a likely direct departure over water to the wintering grounds), we fit a binomial model (logit link and binomial errors).
To assess the validity of assumptions regarding detection at local sites, and thus our ability to infer different patterns of departure at the three sites, we fit simple multi-state mark-release-recaptures models using the RMark interface in R to program Mark (White and Burnham, 1999; www.phidot.org). We fit a single model with two states (local and non-local; defined as above) with time-varying survivorship for each state and location and with detection probability allowed to vary between the two states (but common among locations). Capture periods were defined as the 24 h day beginning at midnight. For all models, we assessed overall model fit by examining standard errors of coefficients and using residual plots. We report the results of an analysis of deviance table (F-tests) and interpret interactions using plots.
Results
Summary information for each site and age class, including morphometrics and body condition at the time of capture, stopover length (PCDP), and the proportion of birds detected beyond the initial tagging location can be found in Table 1.
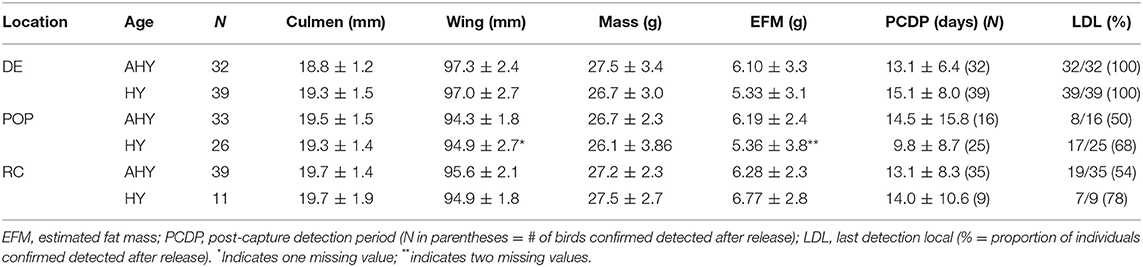
Table 1. Summary (mean ± standard deviation) of morphometric and detection data from Semipalmated Sandpipers tagged at three locations in the Gulf of Maine (DE, Downeast 2013–14; POP, Popham Beach 2015–16; RC, Rachel Carson NWR 2014–15; years pooled for each location).
Across Site Summaries
We found a significant effect of site on wing length [F(2, 173) = 20.6, p < 0.001] and on culmen length [F(2, 174) = 3.3, p = 0.04], but no evidence for differences in morphological features with age [culmen: F(1, 174) = 0.4, p = 0.5; wing: F(2, 173) = 0.04, p = 0.8]. Birds captured at the Downeast site had the smallest culmens (~19 vs. ~19.5 mm; Table 1), but larger wings than those captured at Popham Beach and Rachel Carson (~97 vs. ~95 mm; Table 1; Figure 2). We found no evidence for an effect of age [F(1, 172) = 1.34, p = 0.25] or site [F(2, 172) = 0.85, p = 0.43] [or their interaction: F(2, 172) = 0.49, p = 0.62)] on mass or on size-corrected estimated fat mass (EFM) [age: F(1, 173) = 1.2, p = 0.30; site: F(2, 173) = 0.87, p = 0.40], suggesting that all individuals, regardless of location or age, were in similar energetic condition at the time of capture (Table 1).
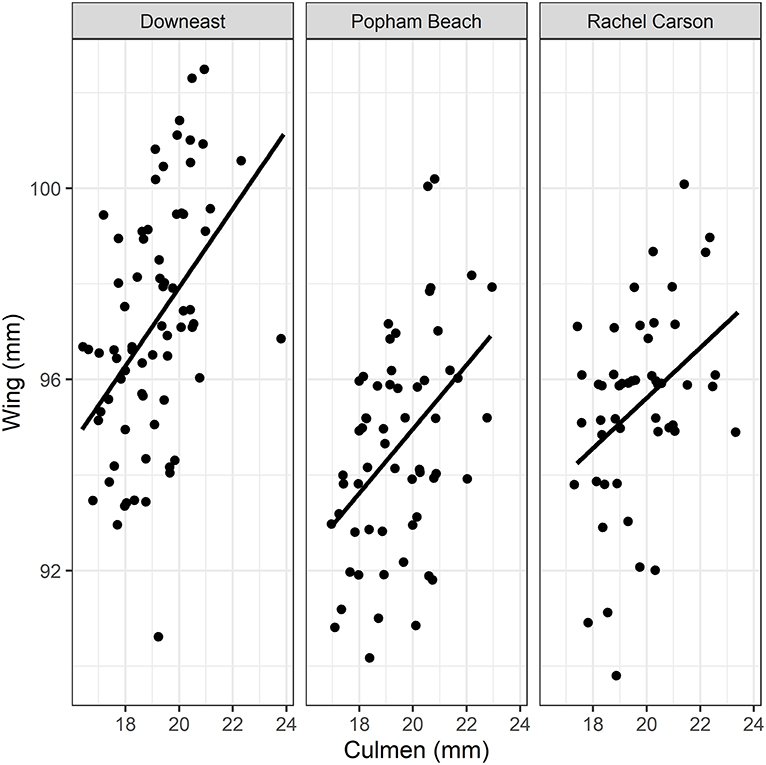
Figure 2. Relationships between flattened wing length (to nearest 0.5 mm) and bill culmen (to nearest 0.5 mm) for Semipalmated Sandpipers (ages, years pooled) captured at each of the three study locations. While there was no difference across each site, in either metric, birds captured at the Downeast site had smaller culmens, but larger wings than those captured at Popham Beach and Rachel Carson.
We found no effect of age [F(1, 143) = 0.04, p = 0.8] on PCDP, but evidence for a significant interaction between site and EFM [F(2, 143) = 5.1, p = 0.007) on PCDP. Individuals from Popham and Rachel Carson remained in the local area irrespective of their original energy stores (Popham Beach 2015–16: R2 = 0.0407, P = 0.1325, df = 56; Rachel Carson NWR 2014–15: R2 = 0.0114, P = 0.4899, df = 43, Figure 3). In contrast, the amount of time that birds on stopover remained at the Downeast site declined linearly with increasing fat mass (Downeast 2013–14: R2 = 0.2664, P < 0.0001, df = 63, Figure 3).
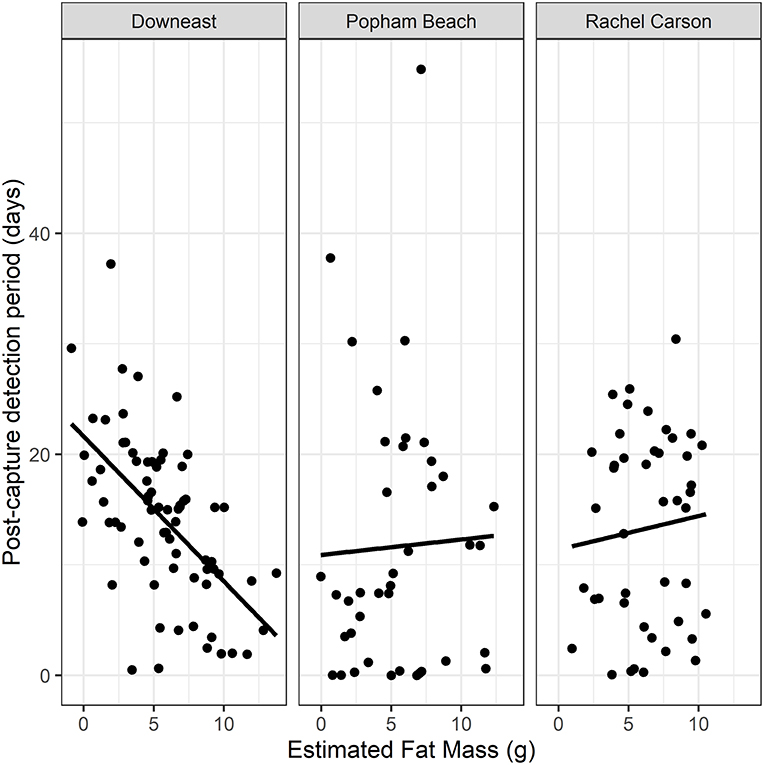
Figure 3. Relationship between energetic condition (size-corrected Estimated Fat Mass, EFM) at the time of capture and the number of days individuals remain detected in the local area after being captured and tagged (Post-Capture Detection Period, PCDP) at each of the three tagging locations (years and ages pooled). Only the group of birds captured at the Downeast site showed a significant effect of EFM on PCDP (Downeast 2013–14: R2 = 0.2664, P < 0.0001, df = 63; Popham Beach 2015–16: R2 = 0.0407, P = 0.1325, df = 56; Rachel Carson NWR 2014–15: R2 = 0.0114, P = 0.4899, df = 43).
Daily probability of detection (Pd) was high when birds were in the designated local tagging area (Pd = 0.96; 0.950–0.974; estimate plus lower and upper CL) but was considerably lower when birds moved out of the local area (the non-local state: Pd = 0.22; 0.18–0.25). After accounting for detection probability, daily “survivorship” (remaining in either state, Ps) was initially high and similar for birds at all three sites (Downeast: Ps = 0.99; 0.976–0.994; Popham: Ps = 0.99; 0.972–0.995; Rachael Carson: Ps = 0.98; 0.96–0.96) but declined much more sharply at the Downeast site than the other two (final daily survivorship, Pds: Downeast: Pds = 0.45; 0.25–0.68; Popham: Pds = 0.72; 0.50–0.87; Rachael Carson: Pds = 0.78; 0.61–0.89).
The probability of a bird being detected at a foreign (non-local) tower differed significantly among sites (Chi = 46.7, df = 2, p < 0.001); no Downeast birds had their final detections at foreign towers, whereas more than 50% of the birds tagged at Rachel Carson (59%, ages pooled) and Popham Beach (61%, ages pooled) were detected beyond the local area (Figure 4). That Downeast birds were not likely to be detected at any other location to the north or south, and that their daily survivorship in the simple multi-state model declined much more quickly than the other sites, suggests that, similar to birds staging in the Bay of Fundy, they departed directly from the Pleasant Bay area on their way to the wintering grounds in eastern South America.
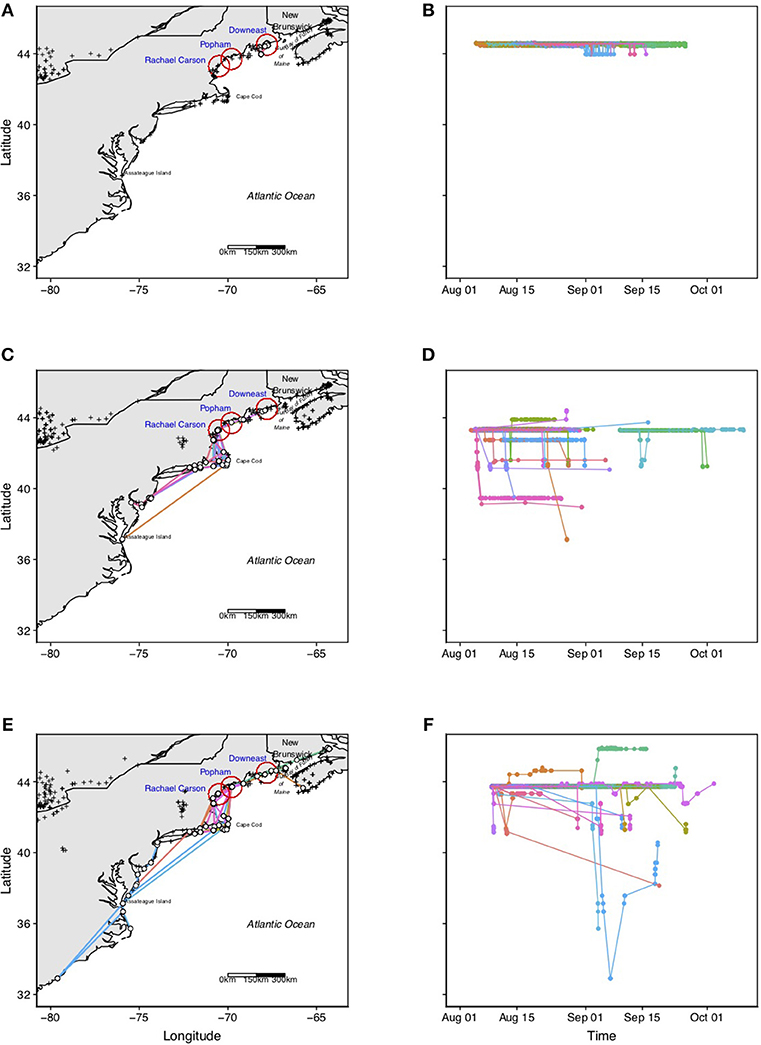
Figure 4. Plots showing spatial as well as temporal post capture movements of individual Semipalmated Sandpipers (ages, years pooled) tagged at each of the three tagging locations: “Downeast” (A,B), “Popham” (C,D), and “Rachel Carson” (E,F). Crosses represent locations of Motus towers present during each of the deployment periods. White dots represent towers where Semipalmated Sandpipers were detected. Colored lines represent tracks of different individual birds, and are consistent within a given site. Sample sizes and detailed information about each site's activities are provided in the text. Note that no birds tagged at the Downeast stopover site in Pleasant Bay were detected beyond the local tagging area, suggesting that these birds initiated trans-oceanic flights from this area. In contrast, more than 50% of the birds tagged at Popham Beach and Rachel Carson NWR were detected to the north (few) and to the south (many), with birds concentrating in coastal southern New England (Cape Cod and the islands, Rhode Island, Long Island Sound) before their final detection, suggesting that this site is also a major departure area for birds making trans-oceanic flights to wintering grounds in eastern South America. A few birds were detected as far south as Delaware Bay, the Carolinas and Virginia.
Individual Sites
Downeast (2013–2014)
The five towers deployed in the Pleasant Bay area provided an approximate local detection space of 30 × 50 km = 1,500 km2 during the 2013–14 migration periods (Figure 1). Numerous towers beyond the area to the north (Bay of Fundy, NS) and to the south, in particular, the array of four towers, oriented NW-SE (perpendicular to the coastline) ~40 km beyond the Downeast tagging area, provided opportunities to detect birds moving along the coast in either direction beyond the denoted local stopover area (Figure 1).
A total of 71 birds (32 AHY, 39 HY, years pooled) were tagged and released at the Downeast site in Pleasant Bay; all of these were later detected at towers within or beyond the tagging location. The post-capture detection period (PCDP) for Semipalmated Sandpipers tagged at this site (and meeting the criterion for inclusion in PCDP) ranged from 0.62 to 27.7 days for adults and from 0.50 to 37.2 days for juveniles (Table 1).
Most post-capture movements of birds captured and tagged in Pleasant Bay remained within the immediate area, with daily movements commonly occurring between exposed mudflats at the mouth of the rivers during low tide and offshore ledge and islands to roost during peak high tide (not shown). All of the 71 birds detected after release had final detections at towers within the local tagging area (Figure 4). The majority of these final detections (39/71 = 55%) were made by towers along the southern boundary of Pleasant Bay, with most of these (26/39 = 67%) made by the tower on Nash Island (Figures 1, 4).
Popham (2015–2016)
The location of the two towers (2015 POP only, 2015 and 2016 POP and SEGUIN, Figure 1) designated as “local” at the Popham Beach tagging site resulted in a local approximate detection space of 20 × 20 km = 400 km2 for 2015 and 20 × 40 km = 800 km2 for 2016 (Figure 1). Numerous towers were active along the coast to the northeast and to the southwest beyond the tagging area during June through November in both years (Figure 1).
A total of 59 birds (33 AHY, 26 HY) were tagged and released at the Popham Beach site but only 41 birds (16 AHY, 25 HY) were confirmed to have been subsequently detected at towers within or beyond the local tagging location (for unknown reasons, 18 tags confirmed to be active at the time of deployment were not later detected by any tower; these birds are excluded from tracking data analyses).
More than half (25/41 = 61%) of the birds tagged at the Popham Beach were last detected at the initial tagging location (Table 1, Figure 4). The post-capture detection period (PCDP) for Semipalmated Sandpipers tagged at this site (and meeting the criterion for inclusion in PCDP) ranged from 0.0 to 54.8 days for adults and from 0.02 to 30.3 days for juveniles (Table 1).
Three individuals tagged at Popham were later detected at sites to the NE, including one HY bird (Motus #20769, Figures 5A,B) that remained at the local site from 21 August through 4 September, after which it moved directly to the upper Bay of Fundy where it remained for at least another 12 days. Another bird (HY, Motus #15515, Figures 5A,B) tagged at Popham Beach was later detected to the north at Cutler, Maine, then flying by the western coast of Nova Scotia 1.6 h later. Another (HY; Motus #20771, Figures 5A,B) was detected at Grand Manan Island at the mouth of the Bay of Fundy.
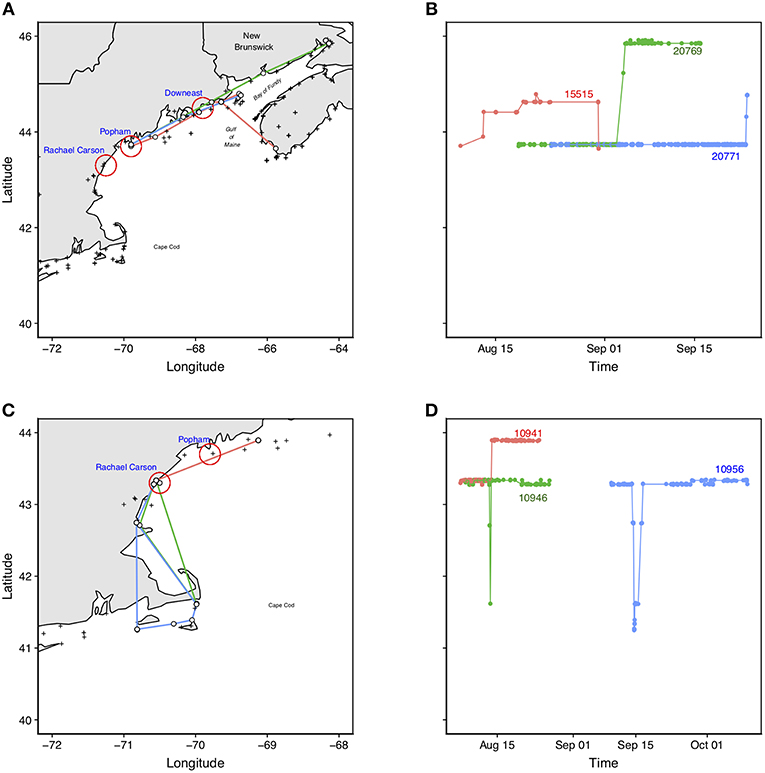
Figure 5. Examples of movements of individual Semipalmated Sandpipers (noted by Motus ID#) tagged at Popham Beach (A,B) and Rachel Carson NWR (C,D). The panels on the left hand side (A,C) show the pathways of the birds within the region; panels on the right hand side (B,D) show the latitudinal movements of the same birds through time. Birds tagged at Popham: bird Motus #20769 (green) moved northeast to the upper part of the Bay of Fundy; bird Motus #15515 (red) moved northward and was last detected at the southwest coast of Nova Scotia; bird Motus #20771 (blue) moved northward and was last detected at Grand Manan Island. Birds tagged at Rachael Carson: Two birds, Motus #10956 (blue) and #10946 (green) made forays south to the Cape Cod area before returning to Rachel Carson NWR where they were both last detected; bird Motus#10941 (red) moved northward and was last detected at the mouth of the Bay of Fundy. These patterns collectively reveal the connectivity among areas in the Bay of Fundy, the Gulf of Maine, and southern New England.
Most birds (8 AHY, 5 HY) whose last detections were at towers beyond the immediate area, however, were detected south of it, with final detections distributed around the southern New England coast (Southern Maine, Cape Cod and the islands of Martha's Vineyard and Nantucket, Block Island, RI, and Long Island Sound) with some individuals being last detected as far away as Cape May, NJ, Delaware, and Chesapeake Bays, Virginia, and North and South Carolinas (Figure 4).
Rachel Carson NWR (2014–2015)
The estimated local detection area, based on the two local towers was 20 × 40 km = 800 km2, oriented northeast-southwest parallel to the coastline (Figure 1). Multiple towers further north and south were active in both years along the coast (Figure 1).
Due to weather constraints, the 2015 capture period was limited to the early half of the migration season for southern Maine, which biased captures to only adults. In 2014–15, 50 birds were captured and tagged: 44 (35 AHY, 9 HY) of these birds had confirmed detections after being released. The remaining six birds were excluded from tracking analyses.
Slightly more than half (59%) of the birds (ages pooled) tagged at Rachel Carson were never detected beyond the local tagging site (Table 1). Of the 18 individuals (16 AHY, 2 HY) that were last detected beyond the local area, 14 went as far south as Cape Cod (or further) with four of those individuals moving these distances within 24 h of capture (Figure 4). The rest of the birds remained at the local tagging site for between 7 and 12 days (Figure 4). Two individuals (Motus #10956, #10946, Figures 5C,D) traveled south to Cape Cod for a short period (<48 h) and returned to the local site, and two others traveled <30 km to the SW. One individual (Motus #10941, Figures 5C,D) traveled ~100 km to the NE.
Discussion
Stopover behavior of individual Semipalmated Sandpipers varied not only in the amount of time they spent at stopover sites along the coast of Maine, but also in subsequent movements within and beyond the region. This variation did not appear to be age-related, but may be in response to ecological, topographical, and environmental factors that varied among the sites, providing support for some of our predictions but not all. Counter to our predictions, we failed to find any support for age-related differences in stopover behavior: during their first migration, young birds on stopover in the Gulf of Maine do not appear to remain longer at stopover sites than adults even though they may be less efficient at foraging and detecting predators (Cresswell, 1994; Fernandez and Lank, 2006; Stillman et al., 2007; van den Hout et al., 2017).
We found partial support for our prediction that the amount of energy reserves birds had at the time of capture would influence the amount of time remained after tagging, but only for birds captured at the Downeast site. And, in contrast to birds tagged further south, few Semipalmated Sandpipers tagged in Pleasant Bay at the Downeast site were detected making forays out of the local tagging area before departing from it altogether. Finally, unlike birds tagged at the other two locations further south, none of the Downeast birds had final detections beyond the initial tagging location.
Site-based difference in final detection patterns suggests that birds arriving on stopover in the Downeast area have a different migratory strategy or route compared to most birds stopping over at sites to the south, which could indicate some level of population or sex class segregation occurring along the Maine coast. Our data provide little support, however, for sex-related segregation. While birds at the Downeast site had smaller bills compared to birds on stopover further south, potentially representing a sex-based bias with males moving to the north and females arriving further south along the Maine coast, this is counter-intuitive. Within breeding populations, females of this species have longer bills than males (Gratto-Trevor et al., 2012a; Hicklin and Chardine, 2012;), and birds with longer bills (i.e., females) may be better at foraging in deeper mud substrates (Harrington, 1982). If the availability of optimal feeding substrates (mudflats), which is greater in the northern Gulf of Maine compared to the southern areas, influences sex-biased movements, we would have expected to see the opposite pattern, with larger billed birds (females) more robustly represented at the Downeast site compared to Popham Beach and Rachel Carson.
We cannot rule out the possibility, however, that birds arriving at the Downeast site are from the same population as those staging in the Bay of Fundy, and that birds from other populations stopover further south. Flattened wing and bill (culmen) lengths for both age classes of birds captured at the Downeast site in this study are consistent with those observed in Semipalmated Sandpipers on stopover in the Bay of Fundy during the same time period (wing: ~97 mm, bill: ~19 mm; see Figure 2, Anderson et al., 2019). As both of these characters are significantly different from birds captured at Popham Beach and Rachel Carson, the data suggest that more than one breeding population is moving through the Gulf of Maine region during fall migration and these populations may show some spatial segregation. However, it is unclear as to which populations birds arriving further south represent. The fact that birds captured in southern Maine had larger bill size than those on stopover at the Downeast site suggests a bias toward central rather than eastern populations arriving in northern Maine, but this is counter to previous population-level morphometric analyses and recent tracking data: bill size increases across an east to west cline, birds staging in the Bay of Fundy are predominantly from eastern breeding populations (Gratto-Trevor et al., 2012a; Hicklin and Chardine, 2012; Miller et al., 2013), and central and western populations do not move as far east as the Gulf of Maine/Bay of Fundy region during fall migration (Brown et al., 2017). Finally, we cannot rule out potential site-based differences in measurement error: in spite of attempts to standardize measurements across the three studies, only one of the researchers took all of the morphological measurements at the first of the three sites (Downeast) while several others collected the data at the other two sites. It would be useful, however, to confirm, in future studies, which Semipalmated Sandpiper populations move through the Gulf of Maine in order to better link events occurring on the breeding grounds with changes in bird numbers at different stopover locations in the region.
Although the underlying mechanisms influencing variation in stopover behavior are unknown, our results clearly show site-based differences in Semipalmated Sandpiper stopover behavior in Maine. The fact that few birds tagged in at the Downeast site (Pleasant Bay) made forays out of the area before resuming migration, and the fact that none were detected anywhere else along the Atlantic coastline strongly suggest that the Pleasant Bay area, like the Bay of Fundy, provides an opportunity to rest and refuel before initiating an extensive trans-oceanic flight to South America. The fact that the timing of departure from the site after tagging was significantly influenced by the amount of fat reserves at the time of capture further reinforces this idea.
Unlike the Downeast site, departure decisions, which were not influenced by energetic condition, at the two southern sites were likely influenced by factors affecting the birds' ability to efficiently rest and refuel. Both Popham Beach State Park and Rachel Carson Wildlife Refuge areas have much greater human activity compared to the Downeast region, and such activity has been shown to directly and negatively influence the amount of time shorebirds spend feeding and resting (Burger, 1993; Thomas et al., 2003; Schlacher et al., 2013; Mayo et al., 2015). In particular, Popham Beach has extremely high public use during peak shorebird migration period (L. Tudor, unpublished data). While the tidal pools at Popham Beach offer areas for shorebirds to feed, these pools are also heavily visited by the public for recreation during both high and low tides. Shorebirds feeding on mudflats located outside the park are exposed to additional human and pet-related activities associated with the surrounding residential development areas (such as free-roaming dogs observed chasing shorebirds on the mudflats, M. Fahay, pers. obs., L. Tudor, pers. obs.). Roosting sites away from human activity are limited at Popham Beach. Similarly, although birds at Rachel Carson NWR were captured while feeding on tidal mudflats, which were within the marsh away from the beach and public hiking areas, these areas are much smaller in area than those in Pleasant Bay and are entirely unavailable for roosting during high tide. Undisturbed roosting areas are limited at Rachel Carson NWR and birds may actively seek them out: many Semipalmated Sandpipers were observed roosting during high tide in the more remote beach areas fenced off earlier in the season to protect nesting shorebirds from public disturbance (K. O'Brien, pers. obs.).
We failed to find age-related patterns in movement behavior at all three stopover sites in spite of the fact that earlier studies found that young birds may not be as efficient in foraging or predator detection as adults (Stillman et al., 2007; van den Hout et al., 2017). Although predation pressure can influence migration strategies in shorebirds (Lank et al., 2003; Sprague et al., 2008), we were not able to systematically collect data on predators. We did, however, note aerial predators when detected, and frequently observed Peregrine falcons (Falco peregrinus) and merlins (F. columbarius) actively attacking shorebird flocks during high and low tides at all three sites.
The Pleasant Bay area (Downeast site) is located along the northeast-southwest oriented coastline contiguous with the Bay of Fundy (Figure 1). Not only is it strategically situated as a jumping off site to fly non-stop over the North Atlantic, it offers expansive tidal mudflats for feeding and numerous offshore rocky ledge outcroppings for roosting, critical resources for shorebirds on migration. These offshore roosting sites are not only safer, away from terrestrial predators and human activity, they are only a few kilometers from the feeding areas. Similar to those in the Bay of Fundy, these tidal mudflats contain greater densities of macroinvertebrates than sand flats and sandy beaches that dominate southern Maine coast (Napolitano and Ackman, 1990; Napolitano et al., 1992). In the northern Gulf of Maine, such mudflats offer higher densities of the nutritionally rich amphipod, Corophium volutator, considered a high quality food for shorebirds, making up 86% of the diet of Semipalmated Sandpipers in the Bay of Fundy and influencing the spatial distribution of the birds feeding there (Napolitano and Ackman, 1990; Napolitano et al., 1992; Hamilton et al., 2003). Corophium volutator is found in greater abundance in the northern Gulf of Maine than in the south (Commito, 1982; Larsen and Doggett, 1991), making stopover sites there the ideal combination of lower human activity and greater abundance of high quality food in close proximity to relatively isolated roosting sites away from terrestrial predators.
In contrast, birds arriving at sites further south in the Gulf of Maine may experience not only greater human activity, but also fewer mudflats with lower quality and less abundant food, as well as fewer safe roost sites nearby. Thus, these birds may opt to do a “hop”/“skip” strategy (c.f. Warnock, 2010) to move either north (to the Downeast area or Bay of Fundy) or south along the coastline to reach a more strategic and more suitable site to prepare for and initiate a trans-oceanic flight. Indeed, we detected birds making such northward movements, but most birds moving beyond their initial tagging area moved southward, with many birds concentrating in and apparently departing from the southern New England coast (e.g., Cape Cod, Rhode Island, Long Island Sound, Figures 4, 5). The fact that at least two individuals returned to the Cape Cod area after making forays further south suggests that Cape Cod and the islands, which, like the Downeast/Bay of Fundy region, serve as a strategic “jumping off” point for birds initiating a trans-oceanic flight to the east coast of South America. It is not known if the birds detected in Virginia or the Carolinas (Figures 4, 5) arrived there by remaining along the coastline or if they were unable to continue an overwater flight initiated further north due to storms or other reasons.
Ocean acidity, which has been shown to severely affect calcium metabolism and thus, productivity of shelled invertebrates (Jacobson et al., 2009), is increasing. For Semipalmated Sandpipers stopping over in the northern Gulf of Maine/Bay of Fundy region, a significant decline in C. volutator, with its high concentration of energy-rich long chain fatty acids, could not only affect the birds' preparation for successful trans-oceanic flight (Maillet and Weber, 2006), but could also have long-term consequences on individual health through reduced immune function and increased tissue damage (Buehler et al., 2010; Eikenaar et al., 2019). Because more than 75% of the world's population of Semipalmated Sandpipers are believed to collectively stage in the Downeast and Bay of Fundy regions in fall, declines in available habitats and energy-rich food sources in this area could seriously impact the species stability altogether (Maillet and Weber, 2006).
Not only are areas along the Atlantic coastline experiencing increased pressure from development, the current rate of sea level rise in the Gulf of Maine is apparently accelerating, with some models predicting more than a half meter rise in sea level by 2100 (Jacobson et al., 2009). Such a rise will significantly and permanently reduce currently available feeding and roosting habitats, perhaps more rapidly than new ones can develop. This realization should prompt resource managers to identify potential feeding and roosting areas now in order to sustain shorebird populations in the future.
In summary, the use of automated VHF radiotelemetry to track individual movements of Semipalmated Sandpipers revealed important site-based, and not age-based, differences in migratory behavior and movements within and beyond the Gulf of Maine region. Impending sea level rise, ocean acidification, and increased human activity will have profound impacts on future shorebird populations worldwide (Galbraith et al., 2005; Iwamura et al., 2013; Fraser et al., 2018). Future research and resource management should be directed toward factors that affect individual behavior, including local as well as regional scale topography, habitat characteristics, human activity, and the degree of connectivity among different sites at the regional and continental scale.
Data Availability
The telemetry data for this study can be found under Projects #8 (Downeast), #25 (Rachel Carson), and #110 (Popham Beach) on the Motus Wildlife Tracking System Motus (www.motus.org).
Ethics Statement
All activities related to bird capture and handling were reviewed and approved by UMaine IACUC (#A2013-01-02 to RH) and were performed under federal and state permits to U.S. Fish and Wildlife Service (to KO'B) and Maine Department of Inland Fisheries and Wildlife (to LT).
Author Contributions
RH, LT, and KO'B developed the project and secured funding for it. RH, LT, KO'B, and GM conducted and supervised all aspects of the fieldwork, including tower assembly and deployment, and capturing and tagging birds. Data analyses were conducted by PT, AB, and RH were reviewed and approved by GM, KO'B, and LT. All authors contributed to the writing and editing of the manuscript.
Funding
Funding was awarded, collaboratively or as individuals, by the Maine Outdoor Heritage Fund (to RH and LT), US Fish & Wildlife Service Region 5 Division of Natural Resources - National Wildlife Refuge System (to KO'B), the federally funded State Wildlife Grant Program (to LT), and the Eastern Maine Conservation Initiative (to RH). This project was supported by the USDA National Institute of Food and Agriculture, Hatch Project #ME0-21609 through the Maine Agricultural and Forestry Experimental Station (to RH); Maine Agricultural and Forest Experiment Station Publication Number 3684.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
As per U.S. Fish and Wildlife policy (http://www.fws.gov/policy/117fw1.html), “The findings and conclusions in this article are those of the author(s) and do not necessarily represent the views of the U.S. Fish and Wildlife Service.”
Acknowledgments
We thank the numerous personnel and volunteers who helped capture and tag birds, set up and operate towers within and beyond our three focal sites, and/or provided valuable assistance with data handling: University of Maine (Sean Rune, Wesley Wright), Maine Department of Inland Fisheries and Wildlife (Brad Allen, Jason Czapiga, MaryEllen Wickett, Lisa Bates, Matthew O'Neal), Biodiversity Research Institute (Patrick Keenan, Kevin Reagan), Maine Bird Observatory (Donna Kausen), US Fish and Wildlife Service National Wildlife Refuges (Maine Coastal Islands National Wildlife Refuge, Rachel Carson National Wildlife Refuge), Popham Beach State Park managers Meagan Hennessey and Brian Murray, Friends of Seguin Island Lighthouse, and Seguin Island Ferry Service (Capt. Ethan DeBery), and Motus Wildlife Tracking Network (John Brzustowski, Stu MacKenzie). We are also grateful for the support of private landowners (the Mudge and Marshall families) who allowed towers to be deployed on their properties at the Downeast location. Michael Fahey and Ken Janes provided numerous observations of sandpipers, including resightings of tagged birds. All activities related to bird capture and handling were reviewed and approved by UMaine IACUC (#A2013-01-02 to RH) and were performed under federal and state permits to US Fish and Wildlife Service and Maine Department of Inland Fisheries and Wildlife (KO'B and LT).
References
Anderson, A. M., Friis, C., Gratto-Trevo, C. L., Morrison, R. I. G., Smith, P. A., and Nol, E. (2019). Consistent declines in wing lengths of Calidridine sandpipers suggest a rapid morphometric response to environmental change. PLoS ONE 14:e0213930. doi: 10.1371/journal.pone.0213930
Andres, B. A., Gratto-Trevor, C., Hicklin, P., Mizrahi, D., Morrison, R. I. G., and Smith, P. A. (2012). Status of the semipalmated sandpiper. Waterbirds 35, 146–148. doi: 10.1675/063.035.0114
Brown, S., Gratto-Trevor, C., Porter, R., Weiser, E. L., Mizrahi, D., Bentzen, R., et al. (2017). Migratory connectivity of Semipalmated Sandpipers and implications for conservation. Condor 119, 207–224. doi: 10.1650/CONDOR-16-55.1
Buehler, D. M., Tieleman, B. I., and Piersma, T. (2010). Indices of immune function are lower in red knots, Calidris canutus, recovering protein than in those storing fat during stopover in Delaware Bay. Auk 127, 394–401. doi: 10.1525/auk.2010.09017
Commito, J. A. (1982). Importance of predation by infaunal polychaetes in controlling the structure of a soft-bottom community in Maine, USA. Mar. Biol. 68, 77–81. doi: 10.1007/BF00393144
Cresswell, W. (1994). Age-dependent choice of Redshank (Tringa totanus) feeding location: profitability or risk? J. Anim. Ecol. 63, 589–600. doi: 10.2307/5225
Crewe, T. L., Crysler, Z., and Taylor, P. (2018). Motus R Book: A Walk Through the Use of R for Motus Automated Radio-Telemetry Data. Port Rowan, ON: Bird Studies Canada. Available online at: https://motus.org/MotusRBook/ (accessed August, 2019).
Dunn, P. O., May, T. A., McCollough, M. A., and Howe, M. A. (1988). Length of stay and fat content of migrant Semipalmated Sandpipers in eastern Maine. Condor 90, 824–835. doi: 10.2307/1368839
Eikenaar, C., Hegemann, A., Packmore, F., Kleudgen, I., and Isaksson, C. (2019). Not just fuel: energy stores are correlated with immune function and oxidative damage in a long-distance migrant. Curr. Zool. 65, 1–8. doi: 10.1093/cz/zoz009
Fefer, S. I., and Schettig, P. A. (1980). An Ecological Characterization of Coastal Maine. U.S. Fish and Wildlife Service Report, FWS/OBS-80/S-29.
Fernandez, G., and Lank, D. B. (2006). Sex, age, and body size distributions of Western Sandpipers during the nonbreeding season with respect to local habitat. Condor 108, 547–557. doi: 10.1650/0010-5422(2006)108[547:SAABSD]2.0.CO;2
Fraser, K. C., Davies, K. T. A., Davy, C. M., Ford, A. T., Tyler Flockhart, D. T., and Martins, E. G. (2018). Tracking the conservation promise of movement ecology. Front. Ecol. Evol. 6:150. doi: 10.3389/fevo.2018.00150
Galbraith, H., Jones, R., Park, R., Clough, J., Herrod-Julius, S., Harrington, B., et al. (2005). Global Climate Change and Sea Level Rise: Potential Losses of Intertidal Habitat for Shorebirds. USDA Forest Service General Technical Report PSW-GTR-191.
Gratto-Trevor, C., Morrison, R. I. G., Mizrahi, D., Lank, D. B., Hicklin, P., and Spaans, A. L. (2012a). Migratory connectivity of Semipalmated Sandpipers: winter distribution and migration routes of breeding populations. Waterbirds 35, 83–95. doi: 10.1675/063.035.0109
Gratto-Trevor, C., Smith, P. A., Morrison, R. I. G., Aubry, Y., and Cotter, R. (2012b). Population trends in Semipalmated Sandpipers from migration counts. Waterbirds 35, 96–105. doi: 10.1675/063.035.0110
Gratto-Trevor, C. L., and Dickson, H. L. (1994). Confirmation of elliptical migration in a population of Semipalmated Sandpipers. Wilson Bull. 106, 78–90.
Hamilton, D. J., Barbeau, M. A., and Diamond, A. W. (2003). Shorebirds, mud snails, and Corophium volutator in the upper Bay of Fundy, Canada: predicting bird activity on intertidal mudflats. Canad. J. Zool. 81, 1358–1366. doi: 10.1139/z03-130
Harrington, B. A. (1982). Morphometric variation and habitat use of Semipalmated Sandpipers during a migratory stopover. J. Field Ornithol. 53, 258–262.
Hicklin, P. W., and Chardine, J. W. (2012). The morphometrics of migrant Semipalmated Sandpipers in the Bay of Fundy: evidence for declines in the eastern breeding population. Waterbirds 35, 74–82. doi: 10.1675/063.035.0108
Iwamura, T., Possingham, H. P., Chades, I., Minton, C., Murray, N. J., Rogers, D. I., et al. (2013). Migratory connectivity magnifies the consequences of habitat loss from sea-level rise for shorebird populations. Proc. R. Soc. B 280:20130325. doi: 10.1098/rspb.2013.0325
Jacobson, G. L., Fernandez, I. J., Mayewski, P. A., and Schmitt, C. V. (2009). Maine's Climate Future: An Initial Assessment. Earth Science Faculty Scholarship, 177. Available online at: https://digitalcommons.library.umaine.edu/ers_facpub/177
Lank, D. B. (1983). Migratory behavior of Semipalmated Sandpipers (Doctoral dissertation). Cornell University, Ithaca, NY, United States.
Lank, D. B., Butler, R. W., Ireland, J., and Ydenberg, R. C. (2003). Effects of predation danger on migration strategies of sandpipers. Oikos 103, 303–319. doi: 10.1034/j.1600-0706.2003.12314.x
Larsen, P. F., and Doggett, L. F. (1991). The macroinvertebrate fauna associated with the mud flats of the Gulf of Maine. J. Coast. Res. 7, 365–375.
Maillet, D., and Weber, J.-M. (2006). Performance-enhancing role of dietary fatty acids in a long-distance migrant shorebird: the Semipalmated sandpiper. J. Exp. Biol. 209, 2686–2695. doi: 10.1242/jeb.02299
Maine Department of Inland Fisheries and Wildlife (2015). Maine's Wildlife Action Plan. Augusta, ME: Maine Department of Inland Fisheries and Wildlife.
Mann, H. A. R., Hamilton, D. J., Paquet, J. M., Gratto-Trevor, C. L., and Neima, S. G. (2017). Effects of extreme tidal events on Semipalmated Sandpiper (Calidris pusilla) migratory stopover in the Bay of Fundy, Canada. Waterbirds 40, 41–49. doi: 10.1675/063.040.0106
Mawhinney, K., Hicklin, P. W., and Boates, J. S. (1993). A re-evaluation of the numbers of migrant Semipalmated Sandpipers, Calidris pusilla, in the Bay of Fundy during fall migration. Canad. Field-Nat. 107, 19–23.
Mayo, T. W., Paton, P. W. C., and Peter, V. (2015). Responses of birds to humans at a coastal barrier beach: Napatree Point, Rhode Island. Northeast. Nat, 22, 501–512. doi: 10.1656/045.022.0302
McNeil, R., and Burton, J. (1977). Southbound migration of shorebirds from the Gulf of St. Lawrence. Wilson Bull. 89, 167–171.
Miller, M. P., Gratto-Trevor, C., Haig, S. M., Mizrahi, D. S., Mitchell, M. M., and Mullins, T. D. (2013). Population genetics and evaluation of genetic evidence for subspecies in the Semipalmated Sandpiper (Calidris pusilla). Waterbirds 36, 166–178. doi: 10.1675/063.036.0206
Morrison, R. I. G., Mizrahi, D. S., Ross, R. K., Ottema, O. H., de Pracontal, N., and Narine, A. (2012). Dramatic declines of Semipalmated Sandpipers on their major wintering areas in the Guianas, northern South America. Waterbirds 35, 120–134. doi: 10.1675/063.035.0112
Napolitano, A., and Ackman, R. G. (1990). Anatomical distribution of lipids and their fatty acids in the Semipalmated sandpiper, Calidris pusilla L. from Shepody Bay, New Brunswick, Canada. J. Exp. Mar. Biol.Ecol. 144, 113–124. doi: 10.1016/0022-0981(90)90023-6
Napolitano, G. E., Ackman, R. G., and Parrish, C. C. (1992). Lipids and lipohilic pollutants in in three species of migratory shorebirds and their food in Shepody Bay (Bay of Fundy, New Brunswick). Lipids 27, 785–790. doi: 10.1007/BF02535850
Pyle, P. (1997). Identification Guide to North American Birds, Part I. Bolinas, CA: Slate Creek Press.
Schlacher, T. A., Nielsen, T., and Weston, M. A. (2013). Human recreation alters behaviour profiles of non-breeding birds on open-coast sandy shores. Estuar. Coast. Shelf Sci. 118, 31–42. doi: 10.1016/j.ecss.2012.12.016
Smith, P. A., Gratto-Trevor, C. L., Collins, B. T., Fellows, S. D., Lanctot, R. B., Liebezeit, J., et al. (2012). Trends in abundance of Semipalmated Sandpipers: evidence from the Arctic. Waterbirds 35, 106–118. doi: 10.1675/063.035.0111
Sprague, A. J., Hamilton, D. J., and Diamond, A. W. (2008). Site safety and food affect movements of Semipalmated Sandpipers (Calidris pusilla) migrating through the upper Bay of Fundy. Avian Conserv. Ecol. 3:4. doi: 10.5751/ACE-00252-030204
Stillman, R. A., West, A. D., Caldow, R. W. G., Le, V., and Dit Durell, S. E. A. (2007). Predicting the effect of disturbance on coastal birds. Ibis 149, 73–81. doi: 10.1111/j.1474-919X.2007.00649.x
Taylor, P. D., Crewe, T. L., Mackenzie, S. A., Lepage, D., Aubry, Y., Crysler, Z., et al. (2017). The Motus Wildlife Tracking System: a collaborative research network to enhance the understanding of wildlife movement. Avian Conserv. Ecol. 12:8. doi: 10.5751/ACE-00953-120108
Thomas, K., Kvitek, R. G., and Bretz, C. (2003). Effects of human activity on the foraging behavior of sanderlings, Calidris alba. Biol. Conserv. 109, 67–71. doi: 10.1016/S0006-3207(02)00137-4
Tudor, L. (2002). Coastal Migratory Shorebird Management System and Data Base. Report to Maine Department of Inland Fisheries and Wildlife, July, 2002.
Tyrrell, M. C. (2005). “Gulf of Maine marine habitat primer,” in Gulf of Maine Council on the Marine Environment. P. 54. Available online at: http://www.gulfofmaine.org/habitatprimer (accessed 2019).
U.S. Shorebird Conservation Plan Partnership, U.S. Shorebirds of Conservation Concern. (2015). Available online at: https://www.shorebirdplan.org/science/assessment (accessed 2017).
van den Hout, P. J., Piersma, T., ten Horn, J., Spaans, B., and Lok, T. (2017). Individual shifts toward safety explain age-related foraging distribution in a gregarious shorebird. Behav. Ecol. 28, 419–428. doi: 10.1093/beheco/arw173
Warnock, N. (2010). Stopping vs. staging: the difference between a hop and a jump. J. Avian Biol. 41, 621–626. doi: 10.1111/j.1600-048X.2010.05155.x
White, G. C., and Burnham, K. P. (1999). Program MARK: survival estimation from populations of marked animals. Bird Study 46(Suppl.), 120–138. doi: 10.1080/00063659909477239
Keywords: Semipalmated Sandpipers, Calidris pusilla, migration, stopover, radiotelemetry, shorebirds, habitat
Citation: Holberton RL, Taylor PD, Tudor LM, O'Brien KM, Mittelhauser GH and Breit A (2019) Automated VHF Radiotelemetry Revealed Site-Specific Differences in Fall Migration Strategies of Semipalmated Sandpipers on Stopover in the Gulf of Maine. Front. Ecol. Evol. 7:327. doi: 10.3389/fevo.2019.00327
Received: 29 April 2019; Accepted: 14 August 2019;
Published: 13 September 2019.
Edited by:
Brett K. Sandercock, Norwegian Institute for Nature Research (NINA), NorwayReviewed by:
Jonathan B. Cohen, State University of New York College of Environmental Science and Forestry, United StatesJaime A. Collazo, North Carolina State University, United States
Copyright © 2019 Holberton, Taylor, Tudor, O'Brien, Mittelhauser and Breit. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Rebecca L. Holberton, cmViZWNjYS5ob2xiZXJ0b25AbWFpbmUuZWR1