Abstract
Recent declines in the migratory North American populations of monarchs (Danaus plexippus) have necessitated efforts to evaluate the current status of the species, including worldwide populations. While monarchs originate from North America and may be ancestrally migratory, they have expanded throughout many parts of the world over the past 200 years. Most of these newer populations no longer migrate and face a variety of threats across a wide range of habitats, but we lack a comprehensive review of locations and characteristics of these worldwide populations. We thus delineated the current range of monarchs and their status throughout the world, recording over 90 countries, islands, and island groups where monarchs occur (74 with recent documented sightings) and known features of these populations. We discuss the major differences between these populations, focusing on morphology, migration, overwintering, natural enemies, larval diet, and genetics. The differences documented here provide the species with adaptive capacity, thus better allowing the species to adapt to novel changes in its environment. We end with a discussion of current gaps in our understanding of monarchs worldwide and directions for future research.
Introduction
Monarchs (Danaus plexippus) are well-known within North America for their long distance migration to overwintering sites along the western coast and in central Mexico. These colorful orange and black butterflies have also expanded to occupy areas throughout the world, from Australia to Spain. With the recent declines in both the eastern and western North American populations, the U.S. Fish and Wildlife Service (USFWS) was petitioned to list monarchs under the Endangered Species Act (ESA) of 1973 (Center for Biological Diversity, 2014). As part of this process, the USFWS is conducting a Species Status Assessment (SSA) to evaluate the status and viability of the species. This SSA requires the species be evaluated as a whole, including in locations outside of the eastern and western North American populations. This process thus necessitated a better understanding of worldwide monarch populations, including where monarchs currently exist, threats faced, and how these populations contribute to the adaptive capacity of the species. Adaptive capacity, or the ability of a species to adjust to novel changes in its physical and biological environment, is important to evaluate to understand the continued success and viability of the species (Nicotra et al., 2015). This mini-review summarizes the findings on monarchs throughout the world, primarily focusing on areas outside of the eastern and western North American populations.
Background and Classification of Monarchs Worldwide
Monarchs were not recorded outside of North America until the mid-nineteenth century, when they colonized areas across both the Pacific and Atlantic Oceans (Zalucki and Clarke, 2004; Fernández-Haeger et al., 2015; although see also Zhan et al., 2014 regarding genetic evidence on potential earlier timing of dispersal events). They established new populations, using available milkweed host plants (primarily Asclepias spp.) and often became non-migratory in the face of year-round suitable temperatures. Monarchs continue to reside in many of these areas worldwide, although North American individuals vastly outnumber the combined numbers of individuals in all the other regions.
We initially limited this review to the migratory subspecies of monarch, D. plexippus plexippus, as this was the subspecies that was petitioned to be listed under the ESA. However, the subspecies categorization is not well-defined for monarchs (e.g., there are non-migratory monarchs that live year-round in areas where others migrate), and most of the literature only refers to monarchs at the species level. Thus, we examined the entire worldwide range of the species, D. plexippus.
To determine where monarchs are located worldwide, we first built a database of all observations of the species published in scientific papers (including Ackery and Vane-Wright, 1984; Zalucki and Clarke, 2004; Patrick and Patrick, 2012; Fernández-Haeger et al., 2015). Observations were removed from locations where we now know it was likely another species [e.g., butterflies classified as monarchs that occur in southern South America are likely Danaus erippus (the southern monarch; Malcolm and Slager, 2015)]. We then looked for current evidence of monarch occupation, which we defined as a sighting in the twenty-first century, by first conducting an extensive literature search to locate countries and islands where monarchs have been recently observed. We also searched iNaturalist, a citizen science platform, and the photo sharing site Flickr for posted monarch sightings with photographic documentation in locations throughout the world. All photos were vetted by the authors, and records were not used if the species could not be verified or if the photo was taken in a butterfly exhibit (as monarchs present might have been imported from other areas). Observations were also excluded that were likely monarchs passing through (e.g., there are occasional sightings of monarchs in England, but no observed breeding). Based on the differences discussed below, we then grouped these countries and islands into eight different geographic regions (Table 1). In total, 90 countries, islands, or island groups were identified as having been historically occupied by monarchs (Figure 1). Of those, 74 have verified sightings since 2000 (Table 1). While the monarch now resides in many worldwide locations, they do not reside in all climatically suitable locations (Zalucki and Rochester, 1999).
Figure 1
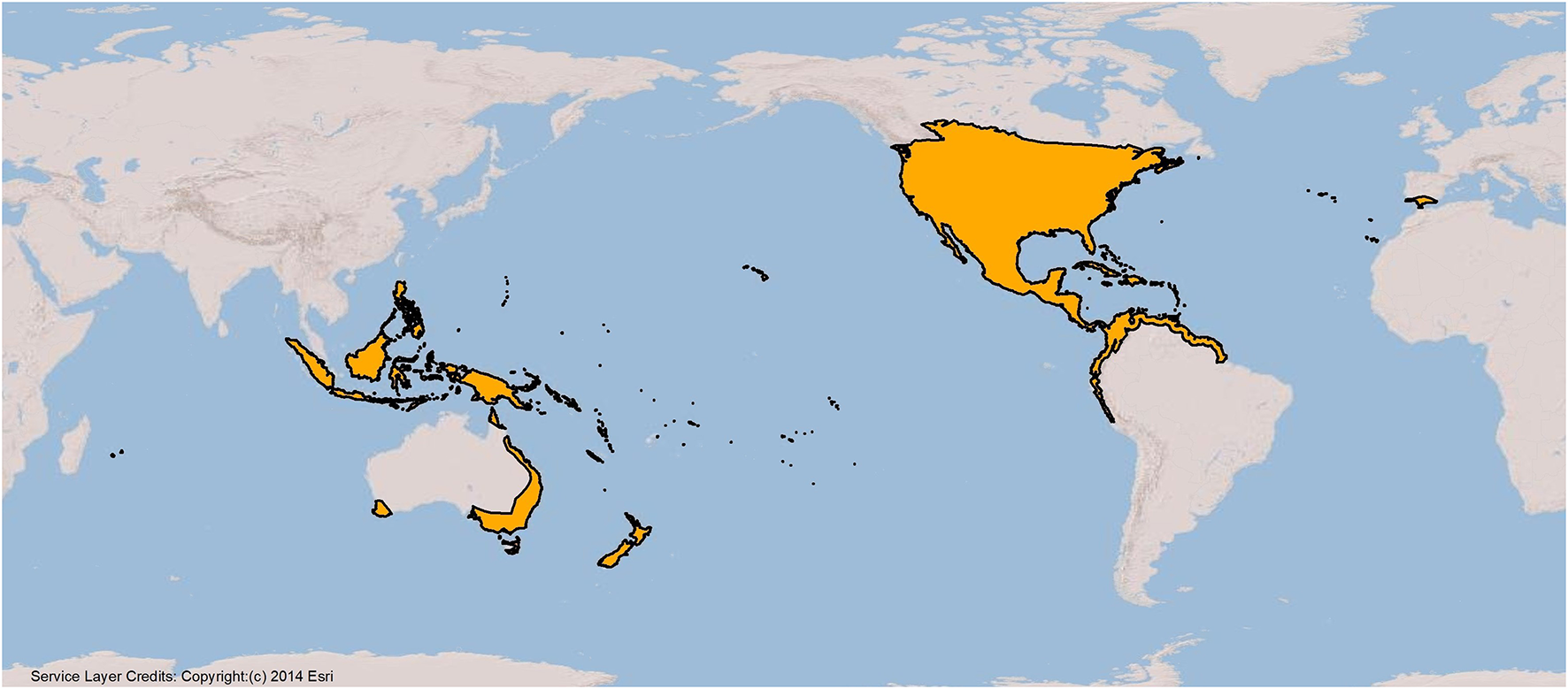
Map showing global range of monarchs (orange shows known range).
Table 1
| Region | Country/Island/Island Group |
|---|---|
| Australia, New Zealand, and Indo-Pacific Islands | American Samoa*, Australia, Brunei*, Commonwealth of the Northern Mariana Islands, Cook Islands, Federated States of Micronesia, Fiji, French Polynesia, Guam, Indonesia*, Johnston Atoll, Kiribati, Malaysia*, Marquesas Islands, Marshall Islands, Mauritius*, Nauru, New Caledonia, New Zealand, Norfolk Island, Palau, Papua New Guinea, Philippines*, Réunion, Samoa, Society Islands, Solomon Islands, Timor-Leste*, Tokelau*, Tonga, Tuvalu, Vanuatu, Wallis and Futuna* |
| Central America and Caribbean | Anguilla, Antigua and Barbuda, Bahamas, Barbados, Belize, Bermuda, Bonaire, British Virgin Islands*, Cayman Islands*, Costa Rica, Cuba, Dominica, Dominican Republic, El Salvador, Grenada, Guadeloupe, Guatemala, Haiti, Honduras, Jamaica, Martinique, Montserrat*, Nicaragua, Panama, Puerto Rico, Saba, Saint Barthélemy*, Saint Kitts and Nevis, Saint Lucia*, Saint Martin, Saint Vincent and the Grenadines*, Sint Eustatius, Sint Maarten, Turks and Caicos Islands, US Virgin Islands |
| Eastern North America | Canada (Eastern)†, Mexico†, Saint Pierre and Miquelon, United States (Eastern)† |
| Hawaii | United States (Hawaii)† |
| Iberian Peninsula | Azores, Canary Islands, Gibraltar, Madeira, Morocco, Portugal, Spain |
| South America and Aruba | Aruba, Colombia, Curaçao, Ecuador, French Guiana*, Guyana, Peru, Suriname, Trinidad and Tobago, Venezuela |
| Southern Florida | United States (Florida)† |
| Western North America | Canada (Western)†, Mexico (Western)†, United States (Western)† |
Locations with occurrences of monarchs.
Indicates that the country/island has evidence of historical occupation, but no evidence has been found of monarch occupation since 2000;
Indicates that the country is listed in multiple regions.
Monarchs are known to have occupied 90 countries, islands, or island groups, which we grouped into eight worldwide regions, with 74 recent documented sightings.
We next examined the literature for differences between monarchs throughout the world in morphology, larval diet, natural enemies, migration, overwintering, genetics, and population sizes and trends, to help us better understand the potential worldwide sources of adaptive capacity for the species.
Worldwide Differences
Morphology
Morphological differences that contribute to the adaptive capacity of the species include wing structure and coloration differences. Wing length was examined in non-migratory monarchs throughout the western hemisphere (from Costa Rica, Puerto Rico, Hawaii, and southern Florida), as well as eastern and western migratory monarchs (Altizer and Davis, 2010). There were differences in shape and size between non-migratory and migratory populations, and non-migratory populations had relatively smaller wings. Additionally, within the eastern population, long-distance migrants tend to have redder coloration (Davis, 2009). Redder coloration is associated with the ability to fly for longer periods of time, although the mechanism for this correlation is unknown (Davis et al., 2012). Recent research also suggests that long-distance migration is a selective force within populations, with longer migration distances positively correlated with longer and larger wings (Flockhart et al., 2017).
Larval Diet
Monarchs rely on milkweed as their host plant, but larval diet still contributes adaptive capacity through variation in species of milkweed consumed. Within North America, there are 108 milkweed species in the genus Asclepias, of which at least 33 are known to be used as larval host plants, as well as at least three species of milkweed vines in the genera Cynanchum and Funastrum (Woodson, 1954; Lynch and Martin, 1993; Yeargan and Allard, 2005). Outside of the eastern and western North American populations, monarchs use Asclepias spp. and closely related species in the subfamily Asclepiadoideae as host plants. In most cases outside of the Americas, these host plants are introduced. Some areas with resident monarchs (e.g., Micronesia and Hawaii) are associated with the common ornamental milkweed, Calotropis gigantea (Buden and Miller, 2003; Buden and Tennent, 2017). Monarchs in Morocco are associated with both A. curassavica and Gomphocarpus fruticosus (also known as A. fruticosa; Fernández-Haeger et al., 2015). Larvae in Australia use C. procera, A. curassavica, and G. fruticosus, of which the latter two have a restricted range due to their inability to tolerate frost and dry conditions (James, 1993; Zalucki, 1993). Monarch larvae in New Zealand and other islands use introduced species including G. fruticosus, Araujia sericifera, and Oxypetalum caeruleum. Larvae in the Azores have been observed consuming Gossypium arboreum and some species of the genus Euphorbia, although these species may not be suitable as host plants (Ramsay, 1964; Neves et al., 2001).
Natural Enemies
Predation, parasitism, and disease impact monarchs throughout their range and thus contribute to the species' adaptive capacity. One natural enemy is the tachinid fly, which impacts monarchs in Australia (Gibbs, 1994), Hawaii (Etchegaray and Nishida, 1975), and throughout Central America and into South America (Arnaud, 1978; Toma, 2010). In Hawaii, parasitism rates from tachinid flies ranged from 0 to 42% (Etchegaray and Nishida, 1975), and in Australia, rates fluctuate throughout the year, going from very low to 100% of sampled monarchs in February (Smithers, 1973). For comparison, the largest North American study estimated tachinid fly parasitism at 13% (Oberhauser et al., 2007). Another parasitoid, a wasp in the Pteromalus genus, is also known to attack monarch pupae in other locations (Ramsay, 1964).
The protozoan parasite, Ophryocystis elektroscirrha (OE), infects monarchs throughout Australia, Central and South America (Altizer et al., 2000), and Hawaii (Pierce et al., 2014b). Infection rates averaged 35% in Hawaii (range: 4–85%; Pierce et al., 2014b), with Australian infection rates averaging between under 10 and almost 66% (Altizer et al., 2000; Barriga et al., 2016). These average rates of OE infection are lower than those observed in the non-migratory population in southern Florida (75–100%), but higher than average rates in the eastern (<10%) and western (5–30%) North American monarch populations (Altizer and de Roode, 2015). Sternberg et al. (2013) further determined that in lab settings, monarchs from South Florida had lower OE spore loads (relative to eastern migratory monarchs) and were less likely to become infected, potentially indicating that non-migratory southern Florida monarchs have increased resistance to OE (however, see also Altizer, 2001). Furthermore, the OE parasites from Florida have been shown to cause higher parasite loads than those from the eastern population (Altizer, 2001). Outside of North America, these high rates of OE infection may not be as detrimental to monarchs. Although the Hawaiian strand of OE is particularly virulent, Hawaiian monarchs are both more resistant to and tolerant of OE (Sternberg et al., 2013).
Monarchs have a number of vertebrate and invertebrate predators that have been studied in North America, and are likely to have many predators outside of North America as well (Oberhauser et al., 2015). There are documented bird predators of monarchs in Australia and Oahu, Hawaii (Smithers, 1973; Stimson and Berman, 1990). Additionally, Australia also has a number of recorded spider and insect predators of monarchs, including mantids, ants, and wasps (Smithers, 1973).
Migration and Overwintering
Monarchs worldwide exhibit varying overwintering and migratory behaviors (with migration potentially being an ancestral trait; Zhan et al., 2014). This variation creates a range of behavioral adaptive capacity. Eastern North American monarchs migrate upwards of 4,000 km every fall (Solensky, 2004), to overwinter in mountainous forests, which provide a unique, protective microclimate (Williams and Brower, 2015). Western North American monarchs also migrate in the fall, flying up to hundreds of kilometers to primarily coastal overwintering groves, which provide a slightly different specific microclimate (Jepsen and Black, 2015; Pyle, 2015). There are fewer monarchs in the western population, spread out among hundreds of overwintering sites (compared to fewer than 20 sites in Mexico; Vidal and Rendón-Salinas, 2014; Jepsen and Black, 2015). Western North American overwintering monarchs may also have a shorter diapause compared to those in eastern North America (Herman et al., 1989), and there may be differences in mating behavior at the different overwintering grounds (Brower et al., 1995).
While these long-distance migrations are well-studied, many locations worldwide have non-migratory monarchs and year-round or winter breeding, including Central America (Ackery and Vane-Wright, 1984), southern Florida (Brower, 1961), along the Gulf Coast (Howard et al., 2010), and southern California (Satterfield et al., 2016), as well as throughout many Pacific Islands. Monarchs in Australia employ both migratory and non-migratory strategies concurrently (James, 1993), with monarchs breeding year round in a northeastern coastal area, but overwintering without breeding at two other sites (Smithers, 1977). This strategy of partial migration (where some individuals migrate and others do not) thus seems common throughout the monarchs' worldwide range, although the proportion of migrants to non-migrants varies greatly.
Australian monarchs have been recorded flying as far as 380 km northeast, forming transient autumn roosts and eventually overwintering roosts. These autumn and winter roost sites are similar in configuration and are often adjacent to major breeding grounds (James, 1993). These sites tend to be inland (20–80 km), have some protection from southerly and westerly winds, and have trees and bushes for roosting [primarily Melaleuca styphelioides (a native tree) and Lantana camara (a naturalized, invasive plant that is also a nectar source for migrating monarchs)]. These sites have high numbers of males during autumn and at the end of overwintering, likely reflecting differences in male and female behavior (James, 1993). Relative to North American monarchs, Australian monarchs spend a shorter time in overwintering aggregations (about 2–4 months, compared to 4–5 months in North America; James, 1993).
Monarchs in New Zealand are non-migratory (one study showed that <3% of recaptured butterflies had flown more than 20 km; Wise, 1980). However, these non-migratory monarchs form overwintering clusters, using Quercus spp., Eucalyptus spp., Cedrus libani, and other species of trees in locations that are both sunny and sheltered (Ramsay, 1964). These sites have nectar sources used by adults, and the colonies vary in size from tens of monarchs to thousands (Ramsay, 1964).
Genetics
The genetics of monarch populations worldwide reflect the widespread variation in dispersal ability, gene flow, and both genetic and allelic diversity that contribute to the adaptive capacity of the species. Genetic results from genome-wide analyses (Zhan et al., 2014) and microsatellite data (Pierce et al., 2014a, 2015) suggest multiple dispersal events from an ancestral North American population, reflecting the capacity for the species to repeatedly expand its range to take advantage of new geographies and resources. Information on gene flow also reflects the wide adaptive capacity of monarchs, as they differentiate and persist at both low and high levels of gene flow. For example, low genetic differentiation among North American monarchs suggest high levels of gene flow between eastern and western populations, even though they display differing migratory behavior (Pierce et al., 2015). Similarly, monarchs from Pacific and Hawaiian Islands or Spain, Portugal, and Aruba also have the capacity to persist and differentiate in their behaviors despite low levels of gene flow that result in more highly genetically differentiated populations (Pierce et al., 2014a). In addition, monarch populations have been established and have persisted across a wide range of genetic and allelic diversity. North American monarchs have higher allelic and genetic diversity and allelic richness than monarch populations in the Pacific and this disparity increases the further the populations exist away from North America (Pierce et al., 2014a).
The adaptive capacity of monarchs worldwide is reflected through the genetically-based estimates of numbers of distinct populations. Pierce et al. (2015) estimate three genetic populations: North America, island populations, and Ecuador, based on genetic clustering analysis of microsatellite markers. Population structure analyses in Pierce et al. (2014a) support a total of seven worldwide populations: North America (including USA, Mexico, Costa Rica, Belize, Puerto Rico, and Bermuda), South America (Ecuador), Aruba, Spain, Portugal/Morocco, the Hawaiian Islands, and a series of Pacific islands (including Australia and New Zealand). Furthermore, Zhan et al. (2014) found genetic distinction among all geographically sampled locations (17), but also reported structure analyses that resulted in 2–11 populations. Additionally, recent research indicates that there may be genetic differentiation between migratory and non-migratory Mexican monarchs (Pfeiler et al., 2017). Microsatellite analyses of monarchs in several locations in the Pacific (Australia, New Zealand, New Caledonia, Fiji, and Samoa) indicate that these monarchs are genetically distinct from other areas and have lower allelic diversity than North American monarchs (Shephard et al., 2002; Pierce et al., 2014a). While specific findings vary, these results show that monarchs worldwide reflect complex population structuring based on extensive dispersal and varying levels of gene flow, which create a wide capacity for the species to adapt to novel environmental and ecological changes in the future.
Population Size and Trends
Despite limited information on population estimates and population size trends outside of North America, there is potential adaptive capacity for the species resulting from monarchs persisting at a range of population sizes worldwide. James (1993) notes that Australian overwintering colony cluster sizes from 1978 to the 1990s were much smaller than those reported in the 1960s (with maximum numbers of 3,500 between 1978 and the 1990s, and ~40,000 as a maximum number in the 1960s). There are also reports of Pacific Islands having boom and bust cycles when first colonized, with monarchs quickly becoming very common before defoliating available host plants (and perhaps moving on to colonize nearby islands; Zalucki and Clarke, 2004).
Discussion
As summarized here, the range of characteristics of worldwide monarchs contribute to the species' adaptive capacity. We know that variation occurs in monarch morphology, with butterflies having varying wing shape, color, and length throughout their range. This variation may be associated with differing migration behavior, which also varies throughout occupied habitat, from the long-distance migrations of North America, to populations exhibiting partial migration, and island monarchs that are now non-migratory. As monarchs have expanded their range, larvae still need suitable milkweed host plants, but the milkweed species (and sometimes genera) can vary from what is common in North America. Monarchs have a suite of natural enemies that they encounter throughout the world. OE is one of the more well-studied natural enemies, with infection and virulence rates varying greatly between populations. The genetics of monarchs throughout their range provide insight into how monarchs expanded and now differ from region to region. By having healthy populations with a variety of these characteristics, the species has more capacity to adapt to future changes in its physical and biological environment.
While there is much that we know about monarchs living outside of the migratory North American populations, many areas of study would benefit from further research. One of the biggest information gaps is the lack of basic information on monarch locations, numbers, and trends throughout much of their range. This information is particularly important with the documented declines of the North American populations. Additionally, without more surveys, it is difficult to delineate range boundaries for many countries. For example, monarchs are presumably not located on every one of Indonesia's over 17,000 islands, but they are probably on multiple islands. Therefore, island nations where we shaded in the entire country likely overrepresent the monarch range (Figure 1).
It is also important for us to better understand threats and their impacts for monarchs worldwide. While some threats are known and discussed above, many are not. Threats that impact monarchs in North America (including habitat loss, insecticides, and climate change, among others; Belsky and Joshi, 2018) may also impact non-North American monarchs in similar or different ways. For example, climate change may impact the suitable breeding range of monarchs in ways similar to North America (Lemoine, 2015), but it may also impact monarch-occupied habitat on Pacific Islands through sea-level rise ([IPCC] Intergovernmental Panel on Climate Change, 2014). These future research efforts, both on novel threats and on monarch locations and population trends, can provide a better understanding of monarchs worldwide, including their contribution to the adaptive capacity of the species, and their likelihood to persist into the future.
Statements
Author’s note
The findings and conclusions in this article are those of the authors and do not necessarily represent the views of the U.S. Fish and Wildlife Service.
Author contributions
KN, LD, and KV contributed to the initial conception and design of this review. LD and KN worked on worldwide analyses. KV and KN completed literature reviews of the worldwide differences. KN wrote the manuscript. All authors contributed to manuscript revisions and approved the submitted version.
Acknowledgments
We thank Erik Olson for his work on the species distribution map. We greatly appreciate the feedback we received from experts on an earlier draft of this paper, including Karen Oberhauser, Stephen Malcolm, Sarina Jepsen, Pablo Jaramillo López, and Chip Taylor, as well as the many reviewers of the SSA draft. We are also extremely grateful for the valuable insight of our SSA team, who helped us define and refine what was needed for a review of monarchs worldwide. We additionally thank two reviewers Jeffrey Marcus and Ayse Tenger-Trolander for their useful feedback, and to citizen scientists for uploading information on monarchs worldwide.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
1
AckeryP. R.Vane-WrightR. I. (1984). Milkweed Butterflies and Their Cladistics and Biology. British Museum (Natural History). Ithaca, NY: Cornell University Press. 425pp.
2
AltizerS.DavisA. K. (2010). Populations of monarch butterflies with different migratory behaviors show divergence in wing morphology. Evolution64, 1018–1028. 10.1111/j.1558-5646.2010.00946.x
3
AltizerS.de RoodeJ. C. (2015). Monarchs and their debilitating parasites: immunity, migration, and medicinal plant use, in Monarchs in a Changing World: Biology and Conservation of an Iconic Insect, eds OberhauserK. S.NailK. R.AltizerS. M. (Ithaca, NY: Cornell University Press), 83–93.
4
AltizerS. M. (2001). Migratory behaviour and host-parasite co-evolution in natural populations of monarch butterflies infected with a protozoan parasite. Evol. Ecol. Res.3, 611–632. Available online at: http://www.evolutionary-ecology.com/issues/v03n05/kkar1286.pdf
5
AltizerS. M.OberhauserK. S.BrowerL. P. (2000). Associations between host migration and the prevalence of a protozoan parasite in natural populations of adult monarch butterflies. Ecol. Entomol.25, 125–139. 10.1046/j.1365-2311.2000.00246.x
6
ArnaudP. H.Jr. (1978). A host-parasite catalog of North American Tachinidae (Diptera). USDA Science and Education Administration Miscellaneous Publications No. 1319. 860.
7
BarrigaP. A.SternbergE. D.LefèvreT.de RoodeJ. C.AltizerS. (2016). Occurrence and host specificity of a neogregarine protozoan in four milkweed butterfly hosts (Danaus spp.)J. Inverteb. Pathol.140, 75–82. 10.1016/j.jip.2016.09.003
8
BelskyJ.JoshiN. K. (2018). Assessing role of major drivers in recent decline of monarch butterfly population in North America. Front. Environ. Sci.6:86. 10.3389/fenvs.2018.00086
9
BrowerL. P. (1961). Studies on migration of monarch butterfly. 1. Breeding populations of Danaus plexippus and D. gilippus Berenice in South Central Florida. Ecology42, 76–83. 10.2307/1933269
10
BrowerL. P.FinkL. S.LeongK.OberhauserK.AltizerS.TaylorO.et al. (1995). On the dangers of interpopulational transfers of monarch butterflies–discussion. Bioscience45, 540–544. 10.2307/1312699
11
BudenD. W.MillerJ. Y. (2003). Butterflies of Pohnpei, Eastern Caroline Islands, Micronesia. Pac. Sci.57, 1–8. 10.1353/psc.2003.0003
12
BudenD. W.TennentW. J. (2017). New records of butterflies (Lepidoptera) from the Federated States of Micronesia with remarks on geographic variation in Hypolimnas bolina (Linnaeus). Pac. Sci.71, 83–90. 10.2984/71.1.8
13
Center for Biological Diversity Center for Food Safety, Xerces SocietyBrowerL. (2014). Petition to protect the monarch butterfly (Danaus plexippus plexippus) under the Endangered Species Act. 159pp.
14
DavisA. K. (2009). Wing color of monarch butterflies (Danaus plexippus) in eastern North America across life stages: migrants are redder than breeding and overwintering stages. Psyche J. Entomol.2009, 1–5. 10.1155/2009/705780
15
DavisA. K.ChiJ.BradleyC.AltizerS. (2012). The Redder the Better: Wing color predicts flight performance in monarch butterflies. PLoS ONE7:e41323. 10.1371/journal.pone.0041323
16
EtchegarayJ. B.NishidaT. (1975). Reproductive activity, seasonal abundance and parasitism of monarch butterfly, Danaus plexippus (Lepidoptera, Danaidae) in Hawaii. Proc. Hawai. Entomol. Soc.22, 33–39.
17
Fernández-HaegerJ.JordanoD.ZaluckiM. P. (2015). Monarchs across the Atlantic Ocean: what's happening on the other shore? in Monarchs in a Changing World: Conservation of an Iconic Butterfly, eds OberhauserK. S.NailK. R.AltizerS. (Ithaca, NY: Cornell University Press), 247–256.
18
FlockhartD. T. TFitz-geraldB.BrowerL. P.DerbyshireR.AltizerS.HobsonK. A.et al. (2017). Migration distance as a selective episode for wing morphology in a migratory insect. Mov. Ecol.5, 1–9. 10.1186/s40462-017-0098-9
19
GibbsG. (1994). The Monarch Butterfly (Mobil New Zealand Nature Series).Auckland: Reed Books Publishing.
20
HermanW. S.BrowerL. P.CalvertW. H. (1989). Reproductive tract development in monarch butterflies overwintering in California and Mexico. J. Lepidopt. Soc.43, 50–58.
21
HowardE.AschenH.DavisA. K. (2010). Citizen science observations of monarch butterfly overwintering in the southern United States. Psyche J. Entomol.2010, 1–6. 10.1155/2010/689301
22
[IPCC] Intergovernmental Panel on Climate Change (2014). AR5 Climate Change 2014: Impacts, Adaptation, and Vulnerability. Contribution of Working Group II to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change. Cambridge; New York, NY: Cambridge University Press.
23
JamesD. G. (1993). Migration biology of the monarch butterfly in Australia, in Biology and Conservation of the Monarch Butterfly. eds MalcolmS. B.ZaluckiM. P. (Los Angeles, CA: Natural History Museum), 189–200.
24
JepsenS.BlackH. S. (2015). Understanding and conserving the western North American monarch population, in Monarchs in a Changing World: Biology and Conservation of an Iconic Butterfly, eds OberhauserK. S.NailK. R.AltizerS. (Ithaca, NY: Cornell University Press), 147–156.
25
LemoineNP. (2015). Climate change may alter breeding ground distributions of eastern migratory monarchs (Danaus plexippus) via range expansion of Asclepias host plants. PLoS ONE10:e0118614. 10.1371/journal.pone.0118614
26
LynchS. P.MartinR. A. (1993). Milkweed host plant utilization and cardenolide sequestration by monarch butterflies in Louisiana and Texas, in Biology and Conservation of the Monarch Butterfly, eds MalcolmS. B.ZaluckiM. P. (Los Angeles, CA: Natural History Museum), 107–123.
27
MalcolmS. B.SlagerB. H. (2015). Migration and host plant use by the southern monarch, Danaus erippus, in Monarchs in a Changing World: Conservation of an Iconic Butterfly, eds OberhauserK. S.NailK. R.AltizerS. (Ithaca, NY: Cornell University Press), 225–235.
28
NevesV. C.FragaJ. C.SchaeferH.VieiraV.Bivar de SousaA.BorgesP. V. (2001). The occurrence of the monarch butterfly, Danaus plexippus L. in the Azores, with a brief review of its biology. Life Marine Sci. 18A, 17–24. Available online at: https://repositorio.uac.pt/bitstream/10400.3/150/1/pp17_24_Neves_et_al_18A.pdf
29
NicotraA. B.BeeverE. A.RobertsonA. L.HofmannG. E.O'LearyJ. (2015). Assessing the components of adaptive capacity to improve conservation and management efforts under global change. Cons. Biol.29, 1268–1278. 10.1111/cobi.12522
30
OberhauserK.GebhardI.CameronC.OberhauserS. (2007). Parasitism of monarch butterflies (Danaus plexippus) by Lespesia archippivora (Diptera : Tachinidae). Am. Midland Natur.157, 312–328. 10.1674/0003-0031(2007)157[312:POMBDP]2.0.CO;2
31
OberhauserK. S.AndersonM.AndersonS.CaldwellW.De AndaA.HunterM.et al. (2015). Lacewings, wasps, and flies—oh my: insect enemies take a bite out of monarchs, in Monarchs in a Changing World: Biology and Conservation of an Iconic Insect, eds OberhauserK. S.NailK. R.AltizerS. M. (Ithaca, NY: Cornell University Press), 71–82.
32
PatrickB.PatrickH. (2012). Butterflies of the South Pacific. Dunedin: Otago University Press.
33
PfeilerE.Nazario-YepizN. O.Pérez-GalvezF.Chávez-MoraC. A.Loustalot LacletteM. R.Rendón-SalinasE.et al. (2017). Population genetics of overwintering monarch butterflies, Danaus plexippus (Linnaeus), from central Mexico inferred from mitochondrial DNA and microsatellite markers. J. Heredity108, 163–17510.1093/jhered/esw071
34
PierceA. A.AltizerS.ChamberlainN. L.KronforstM. R.de RoodeJ. C. (2015). Unraveling the mysteries of monarch migration and global dispersal through molecular genetic techniques, in Monarchs in a Changing World: Biology and Conservation of an Iconic Butterfly, eds OberhauserK. S.NailK. N.AltizerS. (Ithaca, NY: Cornell University Press),257–267.
35
PierceA. A.de RoodeJ. C.AltizerS.BartelR. A. (2014b). Extreme heterogeneity in parasitism despite low population genetic structure among monarch butterflies inhabiting the Hawaiian Islands. PLoS ONE9:e100061. 10.1371/journal.pone.0100061
36
PierceA. A.ZaluckiM. P.BanguraM.UdawattaM.KronforstM. R.AltizerS.et al. (2014a). Serial founder effects and genetic differentiation during worldwide range expansion of monarch butterflies. Proc R Soc B281, 1–6. 10.1098/rspb.2014.2230
37
PyleR. M. (2015). Monarchs in the mist: new perspectives on monarch distribution in the Pacific Northwest, in Monarchs in a Changing World: Biology and Conservation of an Iconic Insect, OberhauserK. S.NailK. R.AltizerS. M. (Ithaca, NY: Cornell University Press), 236–246.
38
RamsayG. W. (1964). Overwintering swarms of the monarch butterfly (Danaus plexippus (L.)) in New Zealand. N. Zeal. Entomol.3, 10–16. 10.1080/00779962.1964.9722821
39
SatterfieldD. A.VillablancaF. X.MaerzJ. C.AltizerS. (2016). Migratory monarchs wintering in California experience low infection risk compared to monarchs breeding year-round on non-native milkweed. Integr. Comp. Biol.56, 343–352. 10.1093/icb/icw030
40
ShephardJ. M.HughesJ. M.ZaluckiM. P. (2002). Genetic differentiation between Australian and North American populations of the monarch butterfly Danaus plexippus (L.) (Lepidoptera: Nymphalidae): an exploration using allozyme electrophoresis. Biol. J. Lin. Soc.75, 437–452. 10.1046/j.1095-8312.2002.00034.x
41
SmithersC. N. (1973). Note on natural enemies of Danaus plexippus (L.) (Lepidoptera: Nymphalidae) in Australia. Austr. Entomol. Mag. 1, 37–40.
42
SmithersC. N. (1977). Seasonal distribution and breeding status of Danaus plexippus (L.) (Lepidoptera: Nymphalidae) in Australia. Austr. J. Entomol.16, 175–184. 10.1111/j.1440-6055.1977.tb00082.x
43
SolenskyM. J. (2004). Overview of monarch migration, in The Monarch Butterfly: Biology and Conservation, eds OberhauserK. S.SolenskyM. J. (Ithaca, NY: Cornell University Press), 79–83.
44
SternbergE. D.LiH.WangR.GowlerC.de RoodeJ. C. (2013). Patterns of host-parasite adaptation in three populations of monarch butterflies infected with a naturally occurring protozoan disease: virulence, resistance, and tolerance. Am. Nat.182:E235–E248. 10.1086/673442
45
StimsonJ.BermanM. (1990). Predator induced color polymorphism in Danaus-plexippus L. (Lepidoptera, Nymphalidae) in Hawaii. Heredity65, 401–406. 10.1038/hdy.1990.110
46
TomaR. (2010). Contribuição ao conhecimento de espécies venezuelanas de Lespesia Robineau-Desvoidy (Diptera, Tachinidae, Exoristinae), com descrições de novas espécies. Rev. Bras. Entomol.54, 165–172. 10.1590/S0085-56262010000200002
47
VidalO.Rendón-SalinasE. (2014). Dynamics and trends of overwintering colonies of the monarch butterfly in Mexico. Biol. Conserv.180, 165–17510.1016/j.biocon.2014.09.041
48
WilliamsE. H.BrowerL. P. (2015). Microclimatic protection of monarch butterflies provided by Mexico's high elevation Oyamel fir forests: a review, in Monarchs in a Changing World: Biology and Conservation of an Iconic Butterfly, eds OberhauserK. S.NailK. R.AltizerS. (Ithaca, NY: Cornell University Press), 109–116.
49
WiseK. A. J. (1980). Monarch butterfly dispersal in New Zealand. Rec. Auckl. Inst. Museum. 17, 157–173.
50
WoodsonR. E.Jr. (1954). The North American species of Asclepias L. Ann. Missouri Bot. Garden. 41, 1–211. 10.2307/2394652
51
YearganK. V.AllardC. M. (2005). Comparison of common milkweed and honeyvine milkweed (Asclepiadaceae) as host plants for monarch larvae (Lepidoptera: Nymphalidae). J. Kansas Entomol. Soc.78, 247–251. 10.2317/0407.40.1
52
ZaluckiM. P. (1993). Sex around the milkweed patch–the significance of patches of host plants in monarch reproduction, in Biology and Conservation of the Monarch Butterfly, eds MalcolmS. B.ZaluckiM. P. (Los Angeles, CA: Natural History Museum), 69–76.
53
ZaluckiM. P.ClarkeA. R. (2004). Monarchs across the Pacific: the Columbus hypothesis revisited. Biol. J. Lin. Soc.82, 111–121. 10.1111/j.1095-8312.2004.00322.x
54
ZaluckiM. P.RochesterW. A. (1999). Estimating the effect of climate on the distribution and abundance of Danaus plexippus: a tale of two continents, in “The 1997 North American Conference on the Monarch Butterfly, eds HothJ.MerinoL.OberhauserK.PisantyI.PriceS.WilkinsonT. (Montreal: Commission for Environmental Cooperation), 151–163.
55
ZhanS.ZhangW.NiitepoldK.HsuJ.HaegerJ. F.ZaluckiM. P.et al. (2014). The genetics of monarch butterfly migration and warning colouration. Nature514, 317–321. 10.1038/nature13812
Summary
Keywords
monarch, Danaus plexippus, migration, worldwide range, natural enemies, morphology, genetics, larval diet
Citation
Nail KR, Drizd L and Voorhies KJ (2019) Butterflies Across the Globe: A Synthesis of the Current Status and Characteristics of Monarch (Danaus plexippus) Populations Worldwide. Front. Ecol. Evol. 7:362. doi: 10.3389/fevo.2019.00362
Received
21 May 2019
Accepted
11 September 2019
Published
27 September 2019
Volume
7 - 2019
Edited by
Wayne E. Thogmartin, United States Geological Survey (USGS), United States
Reviewed by
Jeffrey M. Marcus, University of Manitoba, Canada; Ayse Tenger-Trolander, University of Chicago, United States
Updates
Copyright
© 2019 Nail, Drizd and Voorhies.
This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Kelly R. Nail kelly_nail@fws.gov
This article was submitted to Conservation, a section of the journal Frontiers in Ecology and Evolution
Disclaimer
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.