 Fredrik Dalerum
Fredrik Dalerum Liam O. K. Selby4
Liam O. K. Selby4 Christian W. W. Pirk
Christian W. W. Pirk- 1Research Unit of Biodiversity (UMIB, UO-PA-CSIC), University of Oviedo, Oviedo, Spain
- 2Department of Zoology, Stockholm University, Stockholm, Sweden
- 3Mammal Research Institute, Department of Zoology and Entomology, University of Pretoria, Pretoria, South Africa
- 4Department of Zoology and Entomology, University of Pretoria, Pretoria, South Africa
Promoting co-existence between humans and their physical and ecological environment, including wildlife, has been given an increased importance due to a recent shift of society to become environmentally sustainable. However, humans and large carnivores have been in conflict throughout history. One of the most prominent reasons for this conflict is damages to livestock and domestic animals. Population reduction or even local eradication has often been used as a damage mitigation strategy. However, number of carnivore damages need to be positively related to carnivore densities for population reduction to be an effective damage limitation tool. Sweden is a country in northern Europe with frequent human-carnivore conflicts, spurred by an intense and polarized public debate. We use a 20-year data set on brown bear (Ursus arctos), Eurasian lynx (Lynx lynx) and wolf (Canis lupus) and their damages in Sweden to evaluate if temporal variation in carnivore densities has caused an equivalent variation in the number of damages to cattle, sheep and domestic dogs, if such relationships differed between the carnivore species and damage types, and if there were geographic scale dependencies in these relationships. We observed contradictory effects of large carnivore densities on damages, which included both positive and negative effects. Differences occurred between carnivore species, damage types, geographic areas, and spatial scales. However, wolf densities appeared to have been positively related to the number of damages more often than bear and lynx densities. Our results highlight that large carnivore damages can be highly context dependent, and that other factors than the size of local or regional carnivore populations may be more important damage determinants. Such an interpretation implies that population reduction may not necessarily be an effective method for limiting large carnivore damages, and highlight that damage mitigation strategies need to be flexible over time and space. We recommend further studies identifying the contexts in which large carnivore densities influence damages to livestock and domestic animals, as well as studies aimed at identifying other factors that may be related to the number of damages.
Introduction
Conservation biology has seen a major paradigm shift in the past 50 years, with the current focus being on sustainable incorporation of human societies within their geophysical and biological environment (Mace, 2014). Within this focus, the promotion of coexistence between humans and wildlife is of obvious importance (Frank et al., 2019). However, such coexistence is not without issues, and diverging interests can cause intense conflict between human activities and wildlife populations (Woodroffe et al., 2005; Leader-Williams et al., 2010; Redpath et al., 2013). Conflict can arise, for example, when wildlife damage property, crops or livestock or threaten to kill people. Human-wildlife conflicts are also influenced by the social, cultural, and political background of the people involved, and therefore often extend beyond direct physical conflict between humans and wildlife (Madden, 2004).
Large carnivores are particularly conflict-prone, since their predatory nature often put them at odds with animal owners or hunters (Linnell et al., 2001; Kruuk, 2002; Treves and Karanth, 2003; Woodroffe et al., 2007; Inskip and Zimmermann, 2009). Large carnivores also have slow reproductive rates, low non-human related mortality rates and large area requirements, which results in low population densities (Ewer, 1973). These biological characteristics make large carnivores particularly sensitive to persecution (Purvis et al., 2000), which has lead to large historical losses of carnivore diversity (Dalerum et al., 2009; Dalerum, 2013). Identifying sustainable approaches to human-carnivore coexistence is therefore a central component of current large carnivore management and conservation worldwide (Clarke et al., 2005; Clark and Rutherford, 2014; Hovardas, 2018).
The complex issues causing carnivore-human conflicts call for conflict resolution strategies that target both the direct damages as well as the psychological and social dimensions of occurring conflicts (Dickman, 2010). The relative effectiveness of different strategies depends of the nature of each specific conflict. It is therefore of vital importance to characterize any underlying causes for occurring conflicts (Redpath et al., 2013). Since damages are reported as one of the most prominent causes for conflict (van Eeden et al., 2017), several damage mitigation strategies have been suggested and implemented with varying success, including improved protective fencing and deployment of guard dogs for preventing carnivore attacks (Shivik, 2006; van Eeden et al., 2017, 2018). However, such damage mitigation strategies will be largely ineffective if the conflict is mostly caused by psychological or social issues (Naughton-Treves et al., 2003, but see Karlsson and Sjöström, 2011 for an alternative view). In such cases, strategies aimed at changing public attitudes, for instance by financial compensation or long-term information campaigns, are likely more effective (Nyhus et al., 2003; Kunkel et al., 2016). Such programs can also be implemented in cases were damages are difficult or too costly to prevent (Swenson and Andrén, 2005).
Sweden is a forested and relatively sparsely populated country in northern Europe. It hosts four large carnivore species: brown bear (Ursus arctos, hereafter referred to as “bear”), Eurasian lynx (Lynx lynx, hereafter referred to as “lynx”), gray wolf (Canis lupus, hereafter referred to as “wolf”), and wolverine (Gulo gulo). Human-carnivore conflicts are widespread and large carnivore populations are of increasing concern in rural areas (Sandström et al., 2015). Conflict is complicated by an often intense and polarized public debate, where conservationists and nature-enthusiasts stand in opposition to hunters, livestock farmers and rural residents (Ericsson and Heberlein, 2003; Sandström et al., 2009). Much of the conflict is centered on perceived fear, where hunters and farmers fear for the safety of their pets and livestock and the rural residents fear mostly for the lives of humans, especially that of children (Frank et al., 2015). Despite several mitigation efforts to reduce the conflict (Karlsson and Sjöström, 2011; Lundmark and Matti, 2015; Sjölander-Lindqvist et al., 2015), there are strong pressures to adapt a stricter management with heavy population reduction or even local, regional or national eradication (Sjölander-Lindqvist, 2015). Conflicts between wolves and Sámi reindeer herders, for instance, have previously been regarded as unsolvable. Wolves have therefore actively been prevented from establishing in the northern parts of the country where reindeer herding is practiced (Eriksson and Dalerum, 2018).
In this study we evaluate if temporal variation in the densities of bear, lynx and wolves have led to corresponding variation in the number of damages to cattle, sheep, and domestic dogs in Sweden during a 20-year time period, from 1999 to 2018. Such relationships are critical to establish the effectiveness of various conflict resolution strategies in this country. We also evaluate if damages are associated with temporal variation in livestock or domestic dog density, as well as contrast our analyses across different spatial scales and geographic areas of Sweden. Despite their potential social costs (Boström and Grahn, 2008), we have omitted damages by wolverines since they up until recently primarily have been causing damages to semi-domesticated reindeer, for which no reliable damage records exists. Our specific aims are: (i) to determine if the number of damages to cattle, sheep and domestic dogs are related to temporal variations in large carnivore density, (ii) to determine if any such relationships differ between the three types of damages and between the three species of large carnivores, (iii) to determine if there is a scale dependence in the relationships between carnivore densities and the number of damages, with an expected stronger relationship at smaller spatial scales. Our study covers a period of rapid expansion of the Swedish bear and wolf populations (Swenson et al., 2017; Eriksson and Dalerum, 2018), but a decline of the Swedish lynx population (Widman and Elofsson, 2018). We focus on damages to cattle and sheep because of the economical importance of these types of livestock damages (Widman and Elofsson, 2018), and also on domestic dogs since damages to them are associated with conflict in rural areas (Frank et al., 2015).
Methods
Study Region
Sweden ranges from 55° 20 N to 69° 03 N and covers a land area of 438,600 km2. Approximately 70% of Sweden is covered by forest, most of which is commercial. Only 3% is regarded as built up areas and 8% consists of agricultural land (Statistics Sweden, 2013). Human population is approximately 9 million, with an average density of 24.2 people/km2 (Statistics Sweden, http://www.scb.se). However, humans are unevenly distributed with the most densely populated areas being concentrated to the southern part of the country and at urban centers along the east coast. Climatic and environmental conditions are varied, with mean annual temperatures of 10°C in the south and 8°C in the north of Sweden. The mean temperature averages 18°C in July and 2°C in January in the southernmost parts of the country, but there are large annual fluctuations in temperature.
Sweden is divided into 21 counties, each of which has its own county board. For large carnivore management purposes, the counties are grouped into three regions, the north (consisting of the counties Norrbotten, Västerbotten, Västernorrland, and Jämtland), the central (consisting of Dalarna, Gävleborg, Örebro, Stockholm, Uppsala, Värmland, Västmanland, and Västra Götaland) and the southern (consisting of Blekinge, Halland, Jönköping, Kalmar, Kronoberg, Östergotland, Skåne, and Södermanland) management region. The county of Gotland is an island in the Baltic Sea with no large carnivore presence and is hence excluded from these management regions.
Our study focused on 20 of Sweden's 21 counties. Similarly to Gotland, the island of Öland has also lacked large carnivore presence in recent history and was excluded from area estimates for the county of Kalmar. About half of Sweden's land area, from the central parts and northwards, are defined as a reindeer grazing zone, and can be utilized for semi domesticated reindeer husbandry by the native Sámi people (Swedish Reindeer Husbandry Act, 1971). Unfortunately, there are no public data on reindeer damages, which prevented them from being included in our analyses. Moreover, although the reindeer grazing zone has large implications for Sweden's large carnivore management policies (e.g., Eriksson and Dalerum, 2018), we have not evaluated if the relationships between carnivore densities and damages to sheep, cattle, and domestic dogs differed between areas inside and outside the reindeer grazing zone, since we argue that reindeer husbandry does not directly influence husbandry practices of other domestic species.
Estimation of Carnivore Densities
Population monitoring of all carnivores is managed by Swedish governmental agencies (the Swedish Environmental Protection Agency and regional county boards). We used the annual results from these public surveys as a base for our estimates of temporal variation in carnivore densities across counties, management regions and for the whole of Sweden. For each carnivore species, we converted the estimated number of bears or reproductive units of lynx and wolf per county, management area and for the whole Sweden into densities expressed as number of animals or reproductive units per 1,000 km2. While we appreciate that a smaller spatial resolution may yield additional insights, we have restricted our analyses to these relatively coarse scales for two reasons. First, reliable data on carnivore densities at higher spatial resolution are not readily available from the national surveys. Secondly, we regard these coarse spatial scale as the most germane for national and regional management.
Bear Population Data
Bears are monitored by a combination of volunteer observation during the moose hunt and regional mark-recapture studies based on genetic analyses of collected feces (Kindberg et al., 2011). The observations are routinely recorded annually by hunters during the first week of the moose hunt. Each hunting team records the number of bears they see in an area during their hunt, along with the number of man-hours spent in the field (Kindberg et al., 2011). Fecal collection occurs during targeted surveys. Genetic mark-recapture surveys have been conducted in the counties of Norrbotten (2016), Västerbotten (2004, 2009, 2014), Västernorrland (2004, 2015), Jämtland (2006, 2015), Dalarna (2001, 2012, 2017), Gävleborg (2001, 2012, 2017), and Värmland (2012). We compiled the estimated population sizes from all of these surveys and extracted the number of observed adult bears as well as the number of observation hours from 1999 to 2018. For each of the mark-recapture surveys, we calculated a conversion factor between number of bears observed per man hour and the estimated population size based on genetic mark-recapture, and used the average of the conversion factors for each county to estimate the brown bear population for years with only observations. We omitted the surveys in Dalarna (2001) and Värmland (2012), due to a very low number of observations on which to base conversion factors. We only included counties in the northern management region as well as Dalarna, Gävleborg, and Värmland, as this is regarded as the distribution of the Swedish bear population (Swenson et al., 2017). Estimated annual bear numbers are given in Table S1.
Lynx and Wolf Population Data
The lynx and wolf populations are monitored using a combination of snow tracking, radio telemetry and DNA analyses. The monitoring targets the quantification of number of reproductive units, which for lynx represents a family group consisting of a breeding female with kittens (Andrén et al., 2002) and for wolves represents wolf packs or scent-marking pairs (Liberg et al., 2012). Results are published in annual reports. We collated data from these reports on estimated number of reproductive units of lynx and wolves in each county during the period of 1999 to 2018. In cases where the same reproductive unit was observed in more than one county, we divided it with the number of counties in which it was observed before using it in the annual summary (Eriksson and Dalerum, 2018). For instance, a reproductive unit observed in two counties was counted as 0.5 in each of these counties, and a reproductive unit observed in three counties was counted as 0.33 in each county. Annually reported numbers of reproductive units are given in Table S1.
Estimation of Damages
To receive compensation for carnivore damages on livestock or domestic dogs, each damage has to be reported to the regional county boards, after which it is inspected by certified field personnel. Data from these reports are recorded and stored in a database coordinated and financed by the Swedish Environmental Protection Agency. We extracted all confirmed damages to cattle, sheep and domestic dogs by bear, lynx and wolf during the period 1999 to 2018. We used the raw number of killed, injured, or missing animals, which were pooled for each county, management region and for the whole Sweden. Years with reported damages are given in Table S2.
Estimation of Livestock and Dog Numbers
The Swedish Board of Agriculture maintains registries of all cattle, sheep and domestic dogs. We extracted data on adult cattle and sheep for each county for the period 1999 to 2018 and data on dogs for the period 2011 to 2018. We had to restrict the time period for dogs since they have only been recorded consistently by the board since 2011. All cattle and sheep are required by Swedish law to be housed outside during the summer, and we regard temporal variation in animal husbandry condition to have been minimal. The number of livestock and dogs were converted to densities and are expressed as number of animals or dogs per 100 km2.
Data Analysis
We used generalized linear mixed models to evaluate the effect of carnivore population densities on the number of livestock and dog damages. We ran one separate model for each carnivore species (i.e., brown bear, Eurasian lynx, and wolf) and damage type (i.e., cattle, sheep, and domestic dogs). We also ran models for each carnivore species and damage type on three spatial scales using data from the whole country pooled, data from each management region and data for each county. In each model, we used the raw number of damages as response variable, large carnivore densities, livestock or dog owner densities and their two-way interaction as predictors. For the regional and county scale models, we also added management region and the three-way interaction between region, carnivore density and livestock or dog density as additional predictors. For all models, we added year as a random covariate, and for the county scale we added county size as an additional fixed predictor and county as a random grouping variable. All models were fitted using a log link and a Poisson error distribution. We have provided alpha errors of fixed effects based on Satterthwaite's approximation of residual degrees of freedom (Satterthwaite, 1946). We restricted all analyses on bear damages to the parts of Sweden comprising the central and northern management region, since the population has not yet expanded south of this area (although bears may occasionally occur). We also restricted our analyses to data sets (e.g., for each carnivore species, damage type, and geographic area) that contained at least 3 years with recorded damages. With this restriction, we ran analyses on all carnivores and damage types at a national scale and at all management regions except the southern region for bears. We omitted damages to sheep by lynx and wolves for 1999–2000 since these years consisted of substantial outliers. Counties included in the county scale models are reported in Table S2. All statistical analyses were conducted using the statistical environment R, version 3.5.3 compiled for Linux (http://www.r-project.org) and the packages lme4 (Bates et al., 2015) and lmerTests (Kuznetsova et al., 2017).
Results
Carnivore Populations
During the study period, bears had the highest densities in the central and northern parts of Sweden (Figure 1A), whereas both lynx (Figure 1B) and particularly wolves (Figure 1C) had the highest densities in the central parts. Densities of bears (Figure 1A) and wolves (Figure 1C) increased during the study period, particularly in the central management region, whereas lynx exhibited a stable trend in densities from 2007 (Figure 1B).
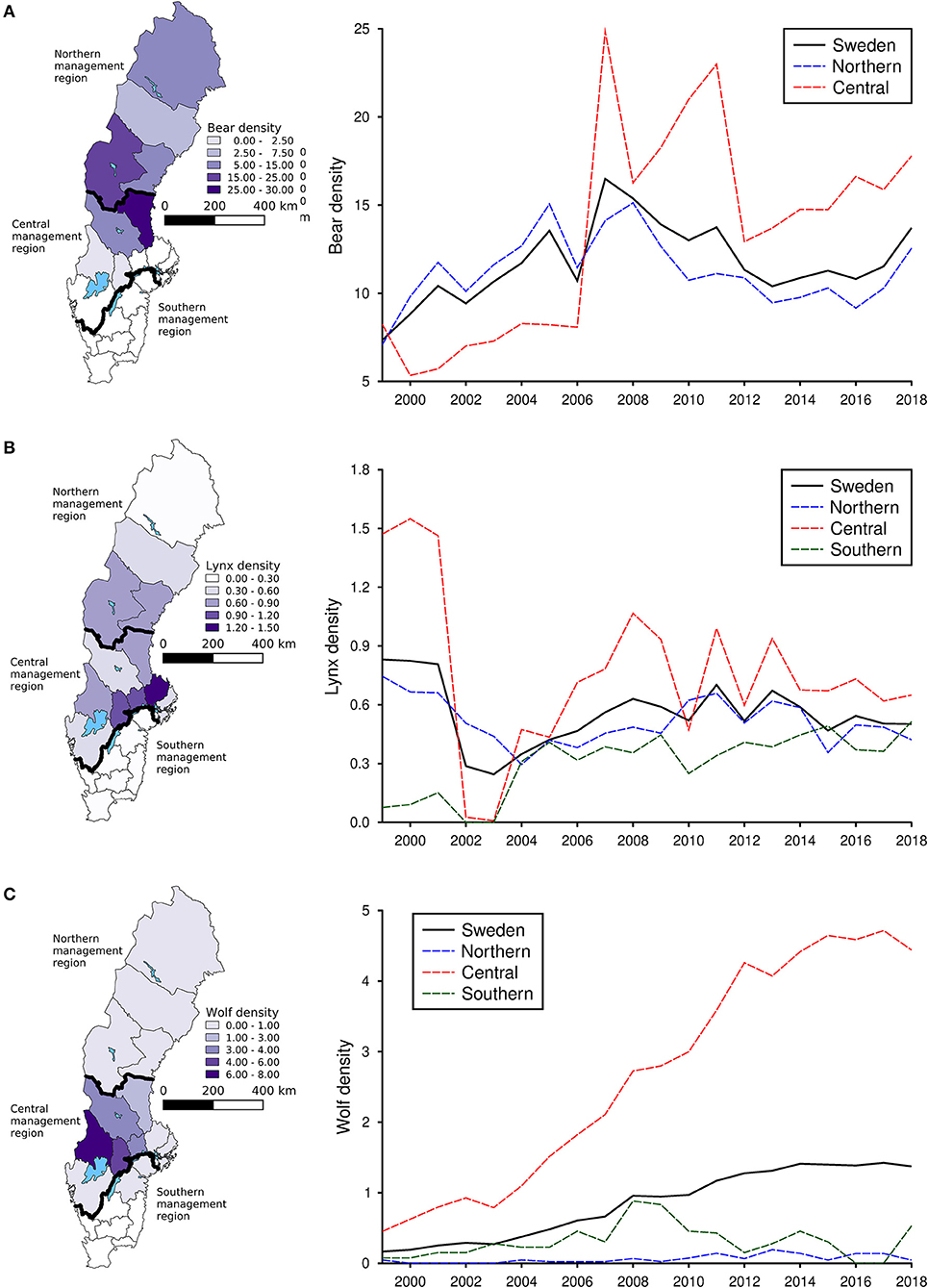
Figure 1. Average densities of brown bear (A), Eurasian lynx (B), and gray wolf (C) in each county of Sweden from 1999 to 2018 as well as the national and regional trends for the same time period. Densities of bears represent number of bears per 1,000 km2, densities for lynx represents number of family units (i.e., mothers with offspring) per 1,000 km2 and densities for wolves represents number of packs or territorial pairs per 1,000 km2. Densities were calculated for each spatial scale as the number of animals divided by the total area of each specific spatial unit (i.e., Sweden, each management region, or, in the maps, each county).
Damages to Cattle
During the study period, an average of 3.2 damages to cattle occurred by bears annually, 1.9 occurred by lynx and 6.3 by wolves. Bear damages to cattle occurred primarily in the northern parts of Sweden (Figure 2A), whereas damages to cattle by lynx (Figure 2B) and wolves (Figure 2C) occurred in central and south western Sweden. For all regions and carnivore species, number of damaged cattle showed substantial annual variation.
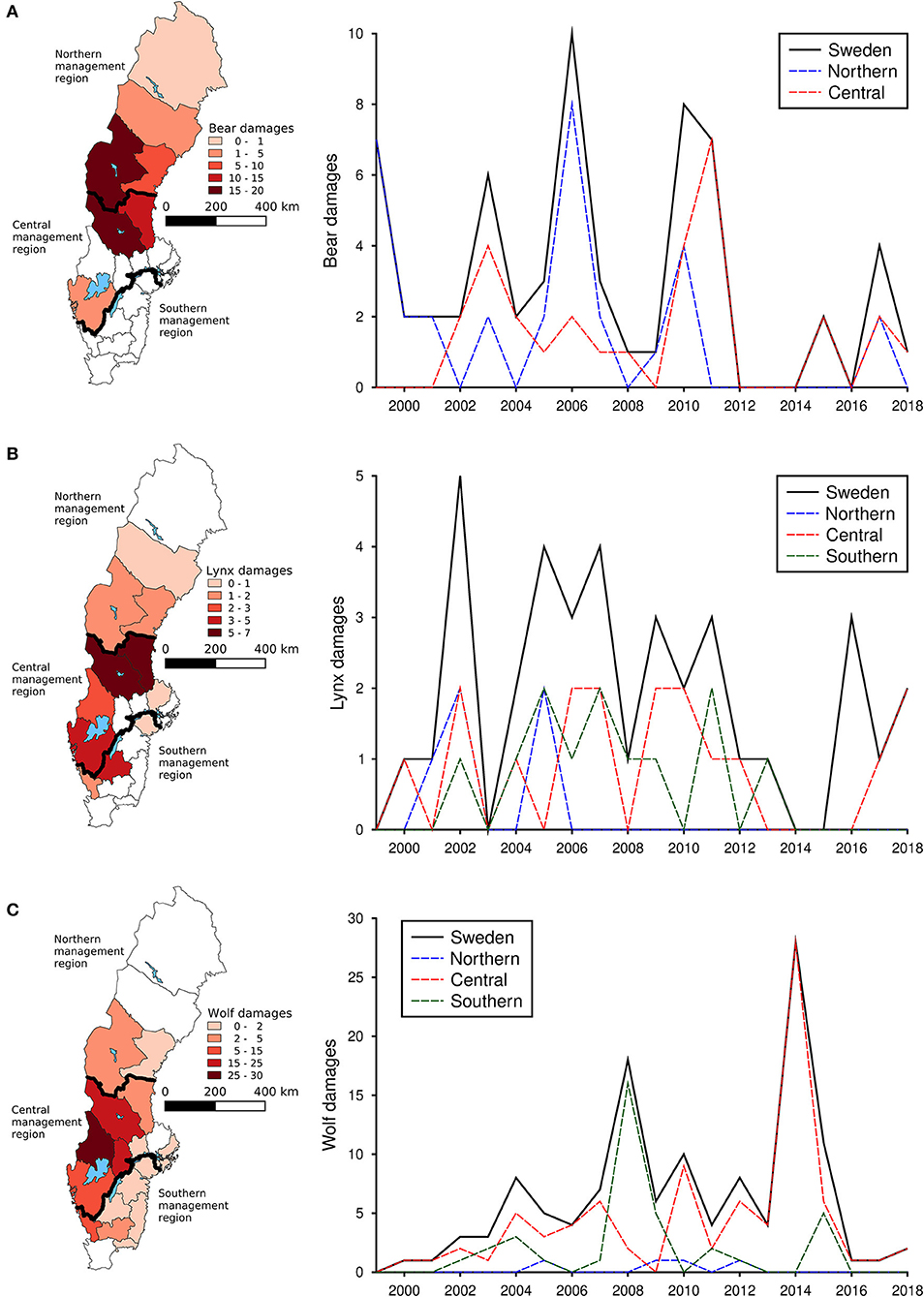
Figure 2. Total number of damaged cattle by brown bear (A), Eurasian lynx (B), and gray wolf (C) in each county of Sweden from 1999 to 2018 as well as the national and regional trends for the same time period.
There were no significant effects of bear density on number of damaged cattle for either geographic scale or region, nor was there any significant interaction effects suggesting that the effects of bear density was influenced by cattle density (Table 1). However, there was a trend for a positive relationship between cattle density and number of damages in the northern region (β = 0.77, SEβ = 0.44, p = 0.078).
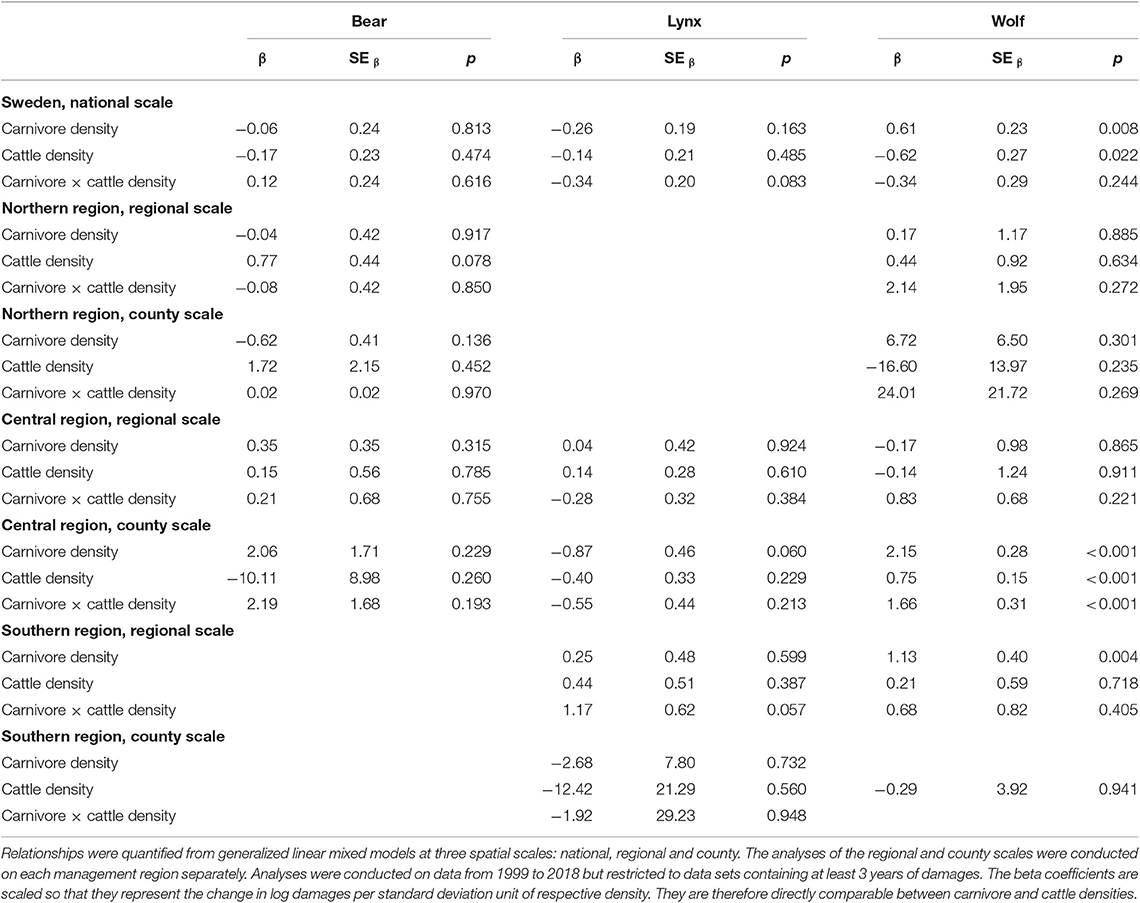
Table 1. Beta coefficients describing the relationships between number of damaged cattle by brown bears, Eurasian lynx and gray wolves and densities of respective carnivore and cattle, as well interaction coefficients describing if the density of cattle influences the relationship between number of damaged cattle and densities of carnivores.
There was a trend for a negative relationship between lynx densities and cattle damages in the central region, but only at the county scale (β = −0.87, SEβ = 0.46, p = 0.060), and lynx density had a positive relationship to cattle densities in the southern region, but also only for the county scale (β = 1.17, SEβ = 0.62, p = 0.057). Lynx densities did not influence the number of damaged cattle either at the national or the regional scale, nor did cattle densities influence such relationships (Table 1). We note that the number of reported lynx damages to cattle remained low, with a maximum of 5 reported damages for a given year in the whole country.
Wolf densities were positively related to number of cattle damages at the national scale (β = 0.61, SEβ = 0.23, p = 0.008), in the southern region at the regional scale (β = 1.13, SEβ = 0.40, p = 0.022) and in the central region at the county scale (β = 2.15, SEβ = 0.48, p < 0.001). There was also a significant positive interaction between wolf and cattle densities in the central region (β = 1.66, SEβ = 0.31, p < 0.001), suggesting that the effects of wolf densities were higher in years of high cattle density. However, wolf densities were not significantly related to cattle damages at neither the regional nor the county scale in the northern region or at the regional scale in the central region (Table 1).
Damages to Sheep
During the study period, an average of 56.3 damages to sheep occurred by bears annually, 123.2 occurred by lynx and 225.9 by wolves. Bear damages to sheep occurred in the western and central parts of Sweden (Figure 3A), lynx damages were distributed across the whole Sweden except for the most northern county of Norrbotten (Figure 3B), whereas damages by wolves were distributed across the western counties of Västra Götaland, Värmland, and Dalarna (Figure 3C). Bear damages to sheep declined during the study period (Figure 3A), whereas damages by both lynx (Figure 3B) and wolves (Figure 3C) increased, particularly in the southern and central management regions.
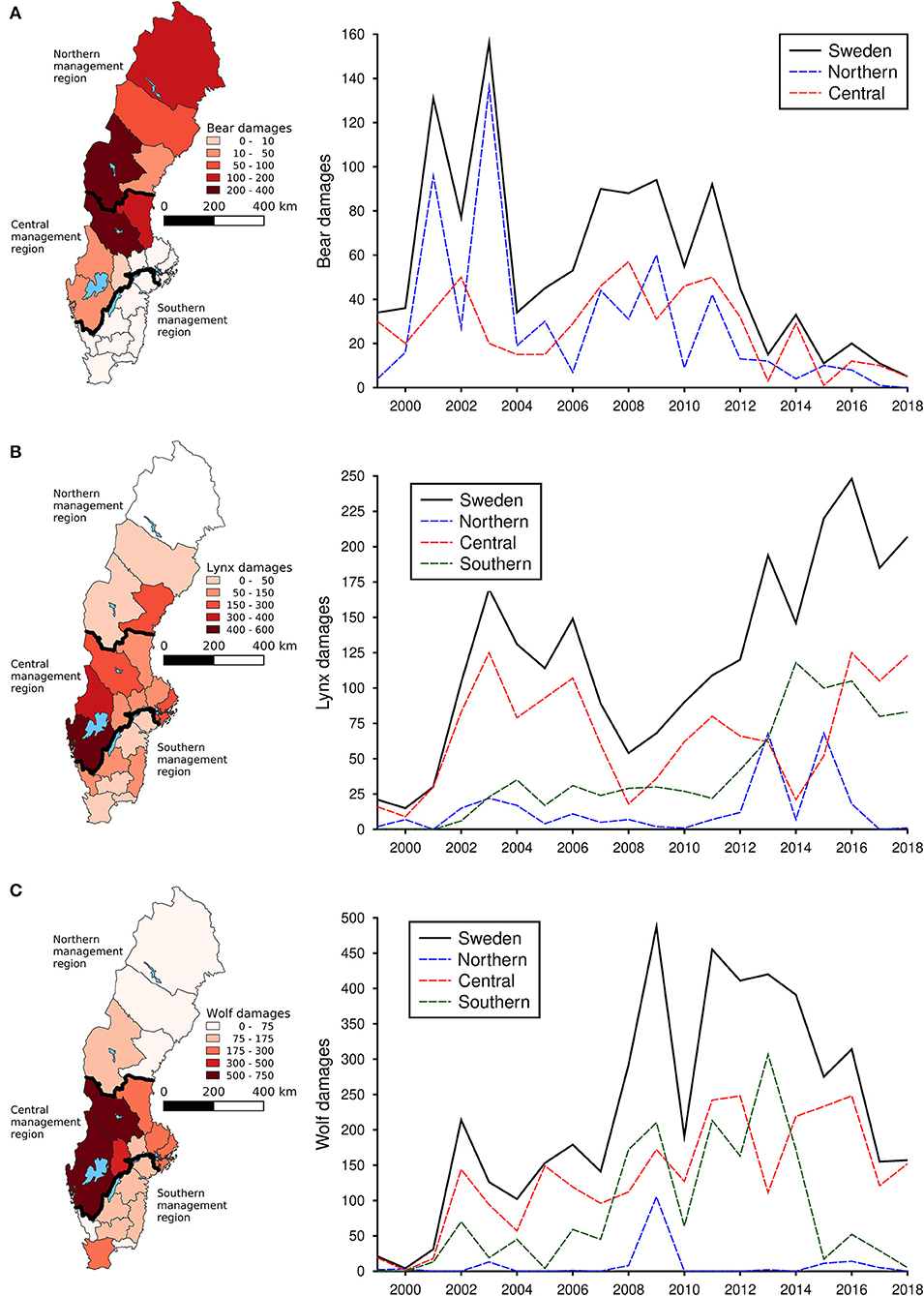
Figure 3. Total number of damaged sheep by brown bear (A), Eurasian lynx (B), and gray wolf (C) in each county of Sweden from 1999 to 2018 as well as the national and regional trends for the same time period.
Bear densities were positively related to the number of bear damaged sheep at the national scale (β = 0.37, SEβ = 0.18, p = 0.043), at the regional scale in the central region (β = 0.54, SEβ = 0.23, p = 0.021) and at the county scale in the northern region (β = 0.52, SEβ = 0.06, p < 0.001). There was also a significant negative interaction effect of bear and sheep densities at the county scale in the northern region, with effects of bear density being weaker during high sheep densities (β = −0.43, SEβ = 0.15, p = 0.005), and a trend for the opposite interaction effect in the central region (β = 0.66, SEβ = 0.35, p = 0.061) (Table 2).
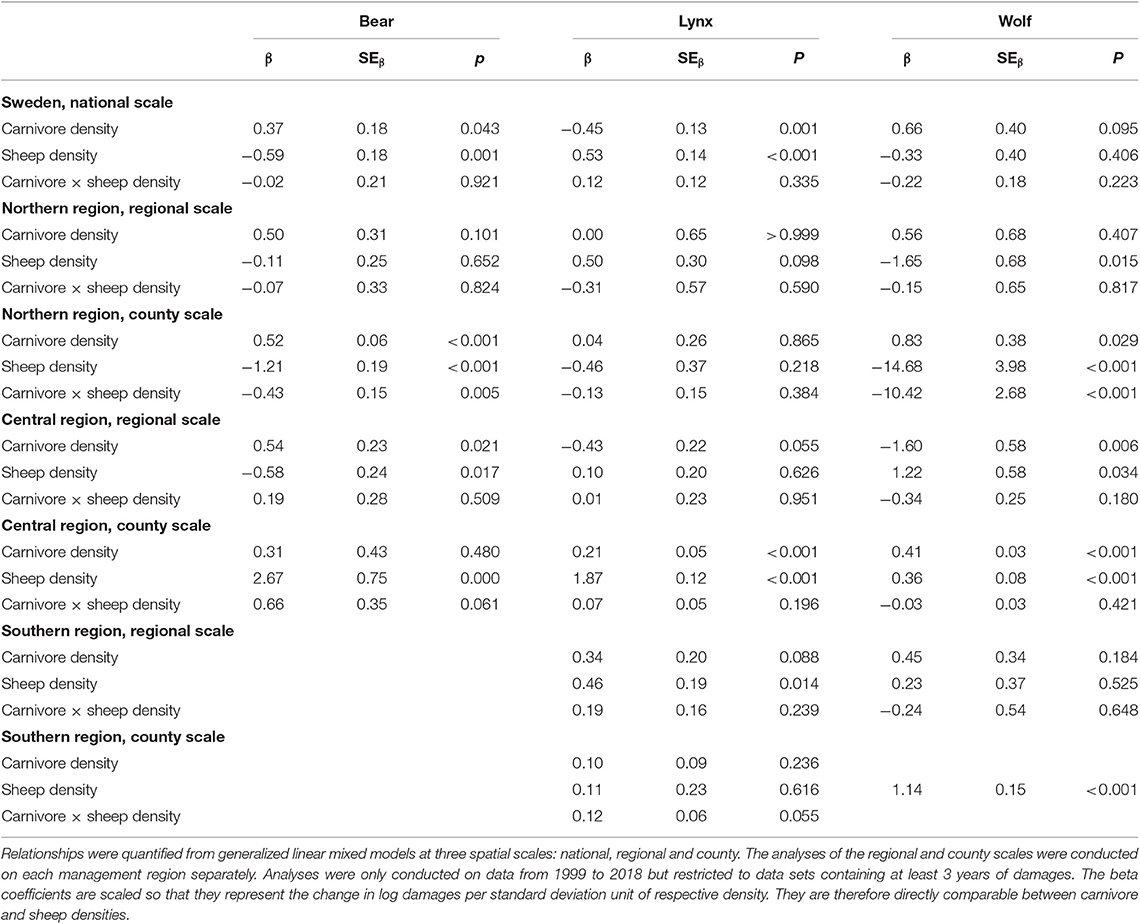
Table 2. Beta coefficients describing the relationships between number of damaged sheep by brown bears, Eurasian lynx and gray wolves and densities of respective carnivore and sheep, as well interaction coefficients describing if the density of sheep influences the relationship between number of damaged sheep and densities of carnivores.
Lynx densities were positively related to lynx damages to sheep in the central region at the county scale (β = 0.21, SEβ = 0.05, p < 0.001), and there was a trend for lynx densities to also be positively related to damages in the southern region at the regional scale (β = 0.34, SEβ = 0.20, p = 0.088). However, lynx densities were negatively related to sheep damages at the national scale (β = −0.45, SEβ = 0.13, p = 0.001) and at the regional scale in the central region (β = −0.43, SEβ = 0.22, p = 0.055). At the national scale (β = 0.53, SEβ = 0.14, p < 0.001), as well as in the central (county scale: β = 1.87, SEβ = 0.12, p < 0.001) and southern region (regional scale: β = 0.46, SEβ = 0.19, p = 0.014), there were positive relationships between sheep density and damages to sheep. There were no significant interaction effects between lynx and sheep densities on sheep damages for any scale or region, nor any effects of lynx or sheep densities on damages in the northern region (Table 2).
At the county scale in the northern (β = 0.83, SEβ = 0.38, p = 0.029) and central regions (β = 0.43, SEβ = 0.03, p < 0.001), wolf densities were positively related to wolf damages to sheep, and there was a trend for a positive relationship at the national scale (β = 0.66, SEβ = 0.40, p = 0.095). However, at the regional scale, wolf densities in the central region were instead negatively related to damages (β = −1.60, SEβ = 0.58, p = 0.006), and there was a significant negative interaction between wolf- and sheep densities at the county scale for the northern region (β = −10.42, SEβ = 2.68, p < 0.029), suggesting that the effect of wolf density declined with increasing sheep densities. Sheep densities were negatively related to wolf damages to sheep in the northern region (regional scale: β = −1.65, SEβ = 0.68, p = 0.015; county scale: β = −14.68, SEβ = 3.98, p < 0.001) but positively related to damages in the central region (regional scale: β = 1.22, SEβ = 0.58, p = 0.034; county scale: β = 0.36, SEβ = 0.08, p < 0.001). Apart from the northern region, there were no significant interaction effects between wolf and sheep densities on sheep damages (Table 2).
Damages to Dogs
During the study period, an average of 2.7 damages to dogs occurred by bears annually, 12.6 occurred by lynx and 29.2 by wolves. Bear damages to dogs occurred primarily in the northern parts of Sweden (Figure 4A), whereas damages by both lynx (Figure 4B) and wolves (Figure 4C) were distributed across almost the whole Sweden. Damages to domestic dogs did not show any distinct trends over time for either bears, lynx or wolves (Figure 4).
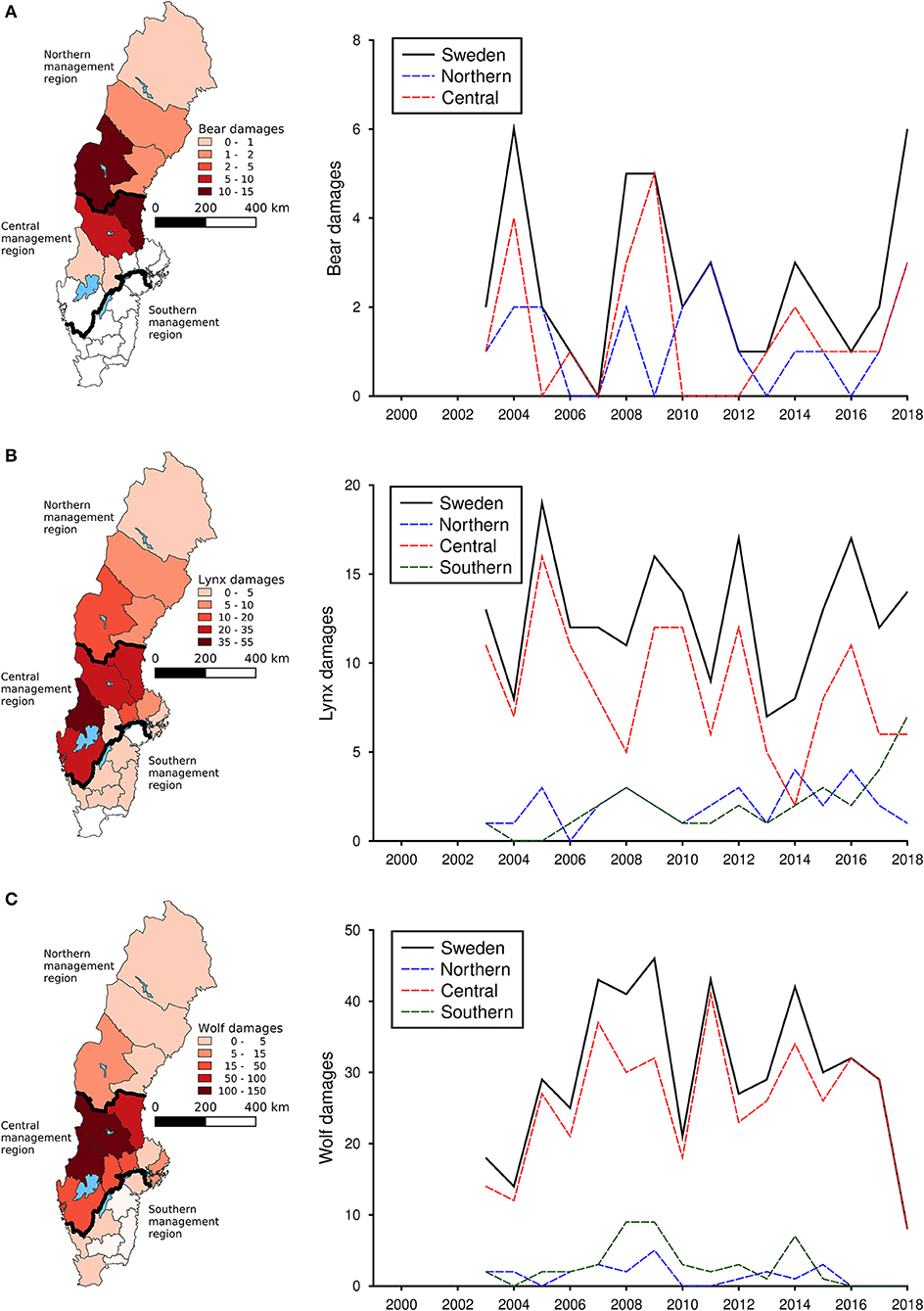
Figure 4. Total number of damaged domestic dogs by brown bear (A), Eurasian lynx (B), and gray wolf (C) in each county of Sweden from 1999 to 2018 as well as the national and regional trends for the same time period.
There was a trend for a positive effect of bear density on the number of dogs attacked by bears, but only in the northern region at the regional scale (β = 1.58, SEβ = 0.88, p = 0.072). For either the national scale or the central region, bear density was not related to bear attacks on dogs (Table 3). Similarly, lynx densities were not related to lynx attacks on dogs for any scale or region (Table 3). At the national scale, there was a trend for the number of attacked dogs to be positively related to wolf density (β = 2.79, SEβ = 1.49, p = 0.061), and there was also a trend for an interaction between wolf and dog densities (β = 3.19, SEβ = 1.77, p = 0.071), suggesting that the effect of wolf density may have been stronger during years with more dogs. However, wolf densities were not related to wolf attacks on dogs at any of the smaller geographic scales (Table 3).
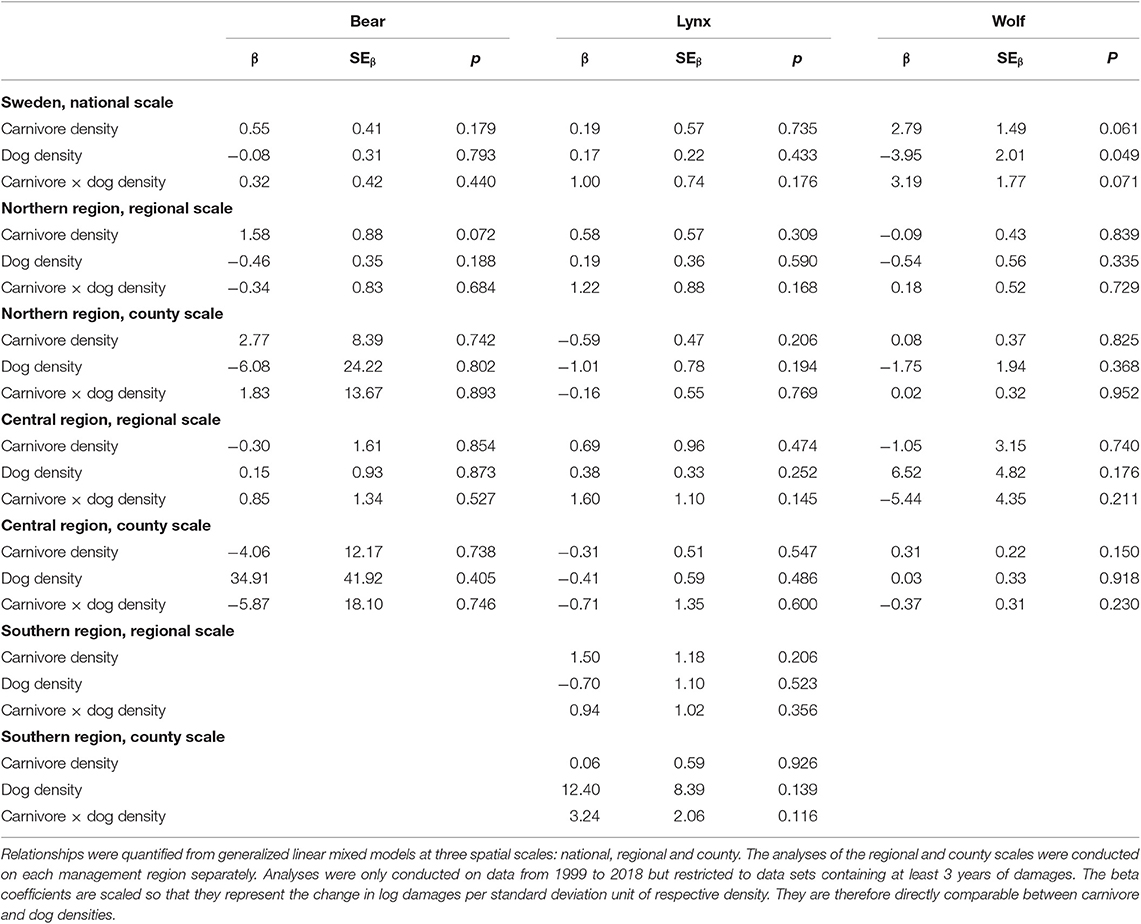
Table 3. Beta coefficients describing the relationships between number of damaged domestic dogs by brown bears, Eurasian lynx and gray wolves and densities of respective carnivore and dogs (proxied by the density of dog owners), as well interaction coefficients describing if the density of dogs influences the relationship between number of damaged dogs and densities of carnivores.
Discussion
We found inconsistencies regarding the effect of temporal variation in large carnivore densities on large carnivore damages to cattle, sheep, and dogs, which suggest that the relationships between large carnivore densities and carnivore damages may have been highly context dependent. These observations also highlight that increased large carnivore densities may not necessarily lead to an increased number of carnivore attacks, but that the number of attacks could also be regulated by other factors. Such context dependence would agree with recent suggestions of context dependencies also in the secondary ecological effects of predation risk, which has been suggested to depend on various factors such as resource availability and the immediate exposure to predation risk exhibited by individual prey at any given time (Middleton et al., 2013; Périquet et al., 2017; Chizzola et al., 2018). The observed inconsistencies included both positive and negative relationships between carnivore densities and number of damages, as well as interaction effects between carnivore and livestock or dog densities. Inconsistencies also occurred across carnivore species, damage types and geographic scales and regions. As with context dependencies in ecological systems (Chamberlain et al., 2014; Haswell et al., 2017), we argue that improving our understanding of context dependencies in the interface between ecological and socio-ecological systems, where environmental managers operate, ought to be a prioritized area of environmental management research in general, and for large carnivore management and conservation in particular.
Because individual carnivore attacks on livestock and domestic animals are highly localized events (Treves et al., 2004), we predicted stronger relationships between temporal variation in carnivore population densities and the number of attacks at small spatial scales. Our results only partly supported this prediction. For instance, in the central management region we did find stronger effects of lynx and wolf densities on damages to cattle at the county than the regional scale. However, these relationships were positive for wolves and negative for lynx. We also found indications of a national scale effect of wolf density on domestic dog attacks, although not statistically significant, but no indications of effects at smaller spatial scales. We suggest several explanations for these observed discrepancies. First, our smallest spatial scale, the administrative scale of county, may have been too large to capture localized effects of carnivore densities for all damage types (e.g., Treves et al., 2004). More localized effects could, for instance, have been caused by variation in territory size and in movement patterns within territories. Contrarily, the spatial scale may have been too small to enable meaningful temporal resolution of damages due to limited sample sizes of attacks (Signorini, 1991). Additionally, the wide-ranging nature of these species could have complicated scale dependencies, with individuals not necessarily captured by the national surveys (e.g., dispersing males or other individuals that were not part of lynx family groups or wolf packs) causing attacks. Attacks could also partly have been carried out by specific individuals or groups (e.g., specific wolf packs, Olson et al., 2015) or categories of individuals (e.g. males, Johansson et al., 2015), which may have caused higher incidences of attacks and damages than what would have been predicted based on average densities.
The relationships between densities and damages differed between the three carnivore species as well as between damage types. Despite contradictory results, including both positive and negative effects of densities, as well as interactions between live stock or dog and carnivore densities, we note three overall patterns in our results. First, damages appear to have been more related to temporal variation in densities of wolves than to variation in bear and lynx densities. Second, damages to sheep appear to have been more related to variation in carnivore densities than damages to either cattle or dogs. Third, damages to dogs appeared to have had no relationships to temporal variation in carnivore densities, except possibly for relationships with wolf densities on a national scale. From a conflict resolution perspective, these general patterns provide important insights for policy development (Treves et al., 2016). For instance, since all detected relationships between wolf densities and damages were positive, reduced wolf populations at spatial scales from national to county may reduce the number of damages by wolves. However, our results do not necessarily support such a conclusion for lynx or bears. Particularly for damages to domestic dogs, restricting carnivore populations may not have noticeable effects on the number of attacks. In cases where relationships between carnivore densities and number of damages are weak, other methods, more directly focused on either avoiding individual attacks or limiting the potential consequences of an attack, may be more successful (van Eeden et al., 2017). In addition, we reiterate previous arguments for the importance of considering both damage related and other causes for conflict when developing conflict mitigation strategies between large carnivores and people (Conover, 2002; Ericsson and Heberlein, 2003; Suryawanshi et al., 2014; van Heel et al., 2017).
Damages of all types and by all three carnivore species occurred mainly in the western and central parts of Sweden. Although this spatial pattern may partly be caused by geographic variation in animal holding practices or native prey densities, this part of the country has seen the most rapid increase in bear and wolf densities during the past few decades (Swenson et al., 2017; Eriksson and Dalerum, 2018). Particularly for wolves, it is clear that the national policy restricting wolves to south of the reindeer grazing zone may have caused an increased number of wolf damages especially in the central region. Providing that the national population size remains constant, an expansion of the wolf population into the reindeer grazing zone would therefore likely decrease the number of damages in central and southern Sweden, particularly the number of sheep damages. Such a decline is expected since, under the scenario of an expanded distribution range but a constant population size, the densities in the current distribution range would by necessity decline. However, allowing such a range expansion would need to carefully consider economic, social and cultural issues related to an established wolf population, although it seems ecologically feasible for wolves to exist in northern Sweden (Eriksson and Dalerum, 2018). We did observe a strong concentration of damages also by bears and lynx in the central region, but the relationships between damages and bear and lynx densities were not consistent. We suggest that focusing national economic and policy incentives for non-lethal damage control to this region is likely to be highly effective in reducing the overall number of damages caused by large carnivores nationally.
To conclude, using a 20-year data set from Sweden, we found contradictory results with regards to the relationships between bear, lynx and wolf densities and damages to cattle, sheep and livestock. Instead, our results suggest that the effects of large carnivore densities on the number of damages may have either been context dependent, or that damages were regulated by other factors than national or regional carnivore densities. Although we did observe positive relationships between densities and damages for some carnivores, damage types and geographic regions and scales, we also observed a lack of relationships, negative relationships, as well as dependencies of livestock densities on the effects of large carnivore densities. In addition, we observed differences in the effects of carnivore densities on damages between the three carnivore species as well as between damage types. Despite the observed variation, wolf densities appeared to have been positively related to the number of damages more often than bear and lynx densities, and damages to sheep appeared to have been more sensitive to increased carnivore densities than damages to cattle and domestic dogs. We urge for further studies aimed at identifying in what contexts that variation in large carnivore densities influences damages to livestock and domestic animals, at what spatial scales such density dependencies in damages occur, and also what other factors than carnivore densities that may regulate number of damages. Such information will be paramount to develop effective human-carnivore conflict mitigation strategies both in Sweden and elsewhere.
Data Availability Statement
The dataset for this study is available in figshare (https://doi.org/10.6084/m9.figshare.11423013).
Ethics Statement
Ethical review and approval was not required because the analyses were based on public data that were not collected specifically for this study.
Author Contributions
FD conceptualized the study, coordinated and conducted data collation, designed and conducted data analyses, and wrote parts of the manuscript. LS assisted with data collation, data analyses, and manuscript writing. CP assisted with writing the manuscript.
Funding
Funding was provided by the Ministry of Economy and Competitiveness in Spain (grant number RYC2013-14662) and by the National Research Foundation in South Africa (grant numbers SFP2008072900003, IFR2011032400087, CPRR13081929244 and IFR150119112493).
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The reviewer, AO, declared past co-authorships with one of the authors, FD, to the handling Editor.
Acknowledgments
We are grateful to administrators at the Swedish Hunting Association for providing brown bear observation data and at the Swedish Board of Agriculture for providing data on number of registered domestic dogs. We are also grateful to the editors of this special topic for the invitation to participate in it.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fevo.2019.00507/full#supplementary-material
References
Andrén, H., Linnell, J. D. C., Liberg, O., Ahlqvist, P., Andersen, R., Danell, A., et al. (2002). Estimating total Lynx lynx lynx population size from censuses of family groups. Wildl. Biol. 8, 299–306. doi: 10.2981/wlb.2002.027
Bates, D. M., Mäechler, M., Bolker, B., and Walker, S. (2015). Fitting linear mixed-effects models using lme4. J. Stat. Softw. 67, 1–48. doi: 10.18637/jss.v067.i01
Boström, G., and Grahn, P. (2008). Estimating cost functions for the four large carnivores in Sweden. Ecol. Econ. 68, 517–524. doi: 10.1016/j.ecolecon.2008.05.008
Chamberlain, S. A., Bronstein, J. L., and Rudgers, J. A. (2014). How context dependent are species interactions? Ecol. Lett. 17, 881–890. doi: 10.1111/ele.12279
Chizzola, M., Belton, L., Ganswindt, A., Greco, I., Hall, G., Swanepoel, L., et al. (2018). Landscape level effects of lion presence (Panthera leo) on two contrasting prey species. Front. Ecol. Evol. 6:191. doi: 10.3389/fevo.2018.00191
Clark, S. B., and Rutherford, M. B. (eds.). (2014). Large Carnivore Conservation: Integrating Science and Policy in the North American West. Chicago, IL: University Chicago Press. doi: 10.7208/chicago/9780226107547.001.0001
Clarke, T. W., Rutherford, M. B., and Casey, D. (eds.). (2005). Coexisting with Large Carnivores – Lessons from Greater Yellowstone. New York, NY: Island Press.
Conover, M. R. (2002). Resolving Human-Wildlife Conflicts. The Science of Wildlife Damage Management. Boca Raton, FL: CRC Press. doi: 10.1201/9781420032581
Dalerum, F. (2013). Phylogenetic and functional diversity in large carnivore assemblages. Proc. R. Soc. B 280:20130049. doi: 10.1098/rspb.2013.0049
Dalerum, F., Cameron, E. Z., Kunkel, K., and Somers, M. J. (2009). Diversity and depletions in continental carnivore guilds: implications for prioritizing global carnivore conservation. Biol. Lett. 5, 35–38. doi: 10.1098/rsbl.2008.0520
Dickman, A. (2010). Complexities of conflict: the importance of considering social factors for effectively resolving human-wildlife conflict. Anim. Cons. 13, 458–466. doi: 10.1111/j.1469-1795.2010.00368.x
Ericsson, G., and Heberlein, T. A. (2003). Attitudes of hunters, locals, and the general public in Sweden now that the wolves are back. Biol. Cons. 111, 149–159. doi: 10.1016/S0006-3207(02)00258-6
Eriksson, T., and Dalerum, F. (2018). Identifying potential areas for an expanding wolf population in Sweden. Biol. Cons. 220, 170–181. doi: 10.1016/j.biocon.2018.02.019
Frank, B., Glikman, J. A., and Marchini, S. (eds.). (2019). Human-Wildlife Interactions: Turning Conflict into Coexistence. Cambridge: Cambridge University Press. doi: 10.1017/9781108235730
Frank, J., Johansson, M., and Flykt, A. (2015). Public attitude towards the implementation of management actions aimed at reducing human fear of brown bears and wolves. Wildl. Biol. 121, 122–130. doi: 10.2981/wlb.13116
Haswell, P. M., Kusak, J., and Hayward, M. W. (2017). Large carnivore impacts are context-dependent. Food Webs 12, 3–13. doi: 10.1016/j.fooweb.2016.02.005
Hovardas, T. (ed.). (2018). Large Carnivore Conservation and Management. London: Routeledge. doi: 10.4324/9781315175454
Inskip, C., and Zimmermann, A. (2009). Human-felid conflict: a review of patterns and priorities worldwide. Oryx 43, 18–34. doi: 10.1017/S003060530899030X
Johansson, Ö., McCarthy, T., Samelius, G., Andrén, H., Tumursukh, L., and Mishra, C. (2015). Snow leopard predation in a livestock dominated landscape in Mongolia. Biol. Cons. 184, 251–258. doi: 10.1016/j.biocon.2015.02.003
Karlsson, J., and Sjöström, M. (2011). Subsidized fencing of livestock as a means of increasing tolerance for wolves. Ecol. Soc. 16:16. doi: 10.5751/ES-03878-160116
Kindberg, J., Swenson, J. E., Ericsson, G., Bellemain, E., Miquel, C., and Taberlet, P. (2011). Estimating population size and trends of the Swedish brown bear Ursus arctos population. Wildl. Biol. 17, 114–123. doi: 10.2981/10-100
Kruuk, H. (2002). Hunter and Hunted: Relationships Between Carnivores and People. Cambridge: Cambridge University Press. doi: 10.1017/CBO9780511614996
Kunkel, K., Shafqat, H., and Khatiwada, A. (2016). “A review of lessons, successes and pitfalls of livestock insurance schemes,” in Snow Leopards of the World: Conservation from Genes to Landscape, eds T. McCarthy, and D. Mallon (London: Academic Press), 173–178.
Kuznetsova, A., Brockhoff, P. B., and Christensen, R. H. B. (2017). lmerTest package: tests in linear mixed effects models. J. Stat. Softw. 82, 1–26. doi: 10.18637/jss.v082.i13
Leader-Williams, N., Adams, W. M., and Smith, R. J. (eds.). (2010). Trade-Offs in Conservation. Chichester: Wiley-Blackwell. doi: 10.1002/9781444324907
Liberg, O., Aronson, Å., Sand, H., Wabakken, P., Maartmann, E., Svensson, L., et al. (2012). Monitoring of wolves in Scandinavia. Hystrix 23, 29–34. doi: 10.4404/hystrix-23.1-4670
Linnell, J. D., Swenson, J. E., and Andersen, R. (2001). Predators and people: conservation of large carnivores is possible at high human densities if management policy is favourable. Anim. Cons. 4, 345–349. doi: 10.1017/S1367943001001408
Lundmark, C., and Matti, S. (2015). Exploring the prospects for deliberative practices as a conflict- reducing and legitimacy enhancing tool: the case of Swedish carnivore management. Wildl. Biol. 21, 147–156. doi: 10.2981/wlb.00009
Madden, F. (2004). Creating coexistence between humans and wildlife: global perspectives on local efforts to address human-wildlife conflict. Hum. Dimens. Wildl. 9, 247–257. doi: 10.1080/10871200490505675
Middleton, A. D., Kauffman, M. J., McWhirther, E. J., Jimenez, M. J., Cook, R. C., Cook, J. G., et al. (2013). Linking anti-predator behavior to prey demography reveals limited risk effects of an actively hunting large carnivore. Ecol. Lett. 16, 1023–1030. doi: 10.1111/ele.12133
Naughton-Treves, L., Grossberg, R., and Treves, A. (2003). Paying for tolerance: rural citizens' attitudes toward wolf depredation and compensation. Cons. Biol. 17, 1500–1511. doi: 10.1111/j.1523-1739.2003.00060.x
Nyhus, P. J., Fisher, H., Madden, F., and Osofsky, S. (2003). Taking the bite out of wildlife damage: the challenges of wildlife compensation schemes. Cons. Pract. 4, 39–41. doi: 10.1111/j.1526-4629.2003.tb00061.x
Olson, E. R., Van Deelen, T. R., Wydeven, A. P., Ventura, S. J., and MacFarland, D. M. (2015). Characterizing wolf-human conflicts in Wisconsin, USA. Wildl. Soc. Bull. 39, 676–688. doi: 10.1002/wsb.606
Périquet, S., Richardson, P., Cameron, E. Z., Ganswindt, A., Belton, L., Loubser, E., et al. (2017). Effects of lions on behavior and endocrine stress in plains zebras. Ethology 123, 667–674. doi: 10.1111/eth.12638
Purvis, A., Gittleman, J. L., Cowlishaw, G., and Mace, G. M. (2000). Predicting extinction risk in declining species. Proc. R. Soc. B 267:20001234. doi: 10.1098/rspb.2000.1234
Redpath, S. M., Young, J., Evely, A., Adams, W. M., Sutherland, W. J., Whitehouse, A., et al. (2013). Understanding and managing conservation conflicts. Trends Ecol. Evol. 28, 100–109. doi: 10.1016/j.tree.2012.08.021
Sandström, C., Johansson, M., and Sjölander-Lindqvist, A. (2015). The management of large carnivores in Sweden — challenges and opportunities. Wildl. Biol. 21, 120–121. doi: 10.2981/wlb.00143
Sandström, C., Pellikka, J., Ratamaki, O., and Sande, A. (2009). Management of large carnivores in Fennoscandia: new patterns of regional participation. Hum. Dim. Wildl. 14, 37–50. doi: 10.1080/10871200802304726
Satterthwaite, F. E. (1946). An approximate distribution of estimates of variance components. Biometrics Bull. 2, 110–114. doi: 10.2307/3002019
Shivik, J. A. (2006). Tools for the edge: whats new for conserving carnivores? Bioscience 56, 253–259. doi: 10.1641/0006-3568(2006)056[0253:TFTEWN]2.0.CO;2
Signorini, D. (1991). Sample size for Poisson regression. Biometrika 78, 446–450. doi: 10.1093/biomet/78.2.446
Sjölander-Lindqvist, A. (2015). Targeted removal of wolves: analysis of the motives for controlled hunting. – Wildl. Biol. 21, 138–146. doi: 10.2981/wlb.00011
Sjölander-Lindqvist, A., Johansson, M., and Sandström, C. (2015). Individual and collective responses to large carnivore management: the roles of trust, representation, knowledge spheres, communication and leadership. Wildl. Biol. 21, 175–185. doi: 10.2981/wlb.00065
Suryawanshi, K. R., Bhatia, S., Bhatnagar, Y. V., Redpath, S., and Mishra, C. (2014). Multiscale factors affecting human attitudes toward snow leopards and wolves. Cons. Biol. 28, 1657–1666. doi: 10.1111/cobi.12320
Swedish Reindeer Husbandry Act (1971). Rennäringslagen 1971:437. Swedish Ministry of Rural Affairs, Stockholm [In Swedish].
Swenson, J., and Andrén, H. (2005). “A tale of two countries: large carnivore depredation and compensation schemes in Sweden and Norway,” in People and Wildlife: Conflict or Co-existence? eds R. Woodroffe, S. Thirgood, and A. Rabinowitz (Cambridge: Cambridge University), 323–339. doi: 10.1017/CBO9780511614774.021
Swenson, J. E., Schneider, M., Zedrosser, A., Söderberg, A., Franzén, R., and Kindberg, J. (2017). Challenges of managing a European brown bear population; lessons from Sweden, 1943–2013. Wildl. Biol. 2017:wlb.00251. doi: 10.2981/wlb.00251
Treves, A., and Karanth, K. U. (2003). Human-carnivore conflict and perspectives on carnivore management worldwide. Conserv. Biol. 17, 1491–1499. doi: 10.1111/j.1523-1739.2003.00059.x
Treves, A., Krofel, M., and McManus, J. (2016). Predator control should not be a shot in the dark. Front. Ecol. Environ. 14:7. doi: 10.1002/fee.1312
Treves, A., Naughton-Treves, L., Harper, E., Mladenoff, D. J., Rose, R. A., Sickley, T. A., et al. (2004). Predicting human-carnivore conflict: a spatial model derived from 25 years of data on wolf predation on livestock. Cons. Biol. 18, 114–125. doi: 10.1111/j.1523-1739.2004.00189.x
van Eeden, L. M., Crowther, M. S., Dickman, C. R., Macdonald, D. W., Ripple, W. J., Ritchie, E. G., et al. (2017). Managing conflict between large carnivores and livestock. Cons. Biol. 32, 26–34. doi: 10.1111/cobi.12959
van Eeden, L. M., Eklund, A., Miller, J. R. B., López-Bao, J. V., Chapron, G., Cejtin, M. R., et al. (2018). Carnivore conservation needs evidence-based livestock protection. PLoS Biol. 16:e2005577. doi: 10.1371/journal.pbio.2005577
van Heel, B. F., Boerboom, A. M., Fliervoet, J. M., Lenders, H. J. R., and van den Born, R. J. G. (2017). Analysing stakeholders' perceptions of wolf, lynx and fox in a Dutch riverine area. Biodiv. Cons. 26, 1723–1743. doi: 10.1007/s10531-017-1329-5
Widman, M., and Elofsson, K. (2018). Costs of livestock depredation by large carnivores in Sweden 2001 to 2013. Ecol. Econ. 143, 188–198. doi: 10.1016/j.ecolecon.2017.07.008
Woodroffe, R., Frank, L. G., Lindsey, P. A., Ranah, S., and Romanach, S. (2007). Livestock husbandry as a tool for carnivore conservation in Africa's community rangelands: a case-control study. Biodiv. Cons. 16, 1245–1260. doi: 10.1007/s10531-006-9124-8
Keywords: human-wildlife conflict, predation, livestock, brown bear, Eurasian lynx, wolf, sheep, cattle
Citation: Dalerum F, Selby LOK and Pirk CWW (2020) Relationships Between Livestock Damages and Large Carnivore Densities in Sweden. Front. Ecol. Evol. 7:507. doi: 10.3389/fevo.2019.00507
Received: 26 July 2019; Accepted: 16 December 2019;
Published: 10 January 2020.
Edited by:
Laurentiu Rozylowicz, University of Bucharest, RomaniaReviewed by:
Juan Carlos Blanco, Fundación Oso Pardo (Brown Bear Foundation), SpainAndres Ordiz, Norwegian University of Life Sciences, Norway
Copyright © 2020 Dalerum, Selby and Pirk. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Fredrik Dalerum, ZGFsZXJ1bWpvaGFuQHVuaW92aS5lcw==