 Benjamin Luke Payne
Benjamin Luke Payne Jakob Bro-Jørgensen
Jakob Bro-Jørgensen- Mammalian Behaviour and Evolution Group, Department of Evolution, Ecology and Behaviour, Institute of Integrative Biology, University of Liverpool, Neston, United Kingdom
Throughout Africa, conservancies under community and private management are becoming increasingly important as a complement to the protection offered by the established core network of protected areas (PAs), which are largely under national management. However, opportunities for creating new conservation areas are restricted by increasing demand on land use by growing human populations, and it is unclear how effectively the current protected area network captures spatial priorities for conservation. Taking into account climate-induced range-shifts, we first identified spatial priorities for antelope conservation in Africa by gap analysis of the network of PAs listed with an IUCN category in the World Database for Protected Areas. For three countries from which information were available, we then assessed to what extent the gaps identified were covered by PAs not listed with an IUCN category, for the latter making a distinction between whether management was referred to as community-based or not. The results showed limited overlap, suggesting that the success of community-based and privately managed PAs in covering spatial priorities from a continent-wide perspective could be increased by more strategic land use planning at the national level.
Introduction
Protected areas (PAs) constitute a cornerstone in conservation. Currently around 15% of the Earth’s land surface is under protection (UNEP-WCMC et al., 2019), not far off the 17% Aichi target set for 2020 set by the UN Convention on Biological Diversity (UNEP, 2010). However, whether this coverage is sufficient to effectively preserve biodiversity is the subject of debate, especially following E.O. Wilson’s call for half the Earth be set aside for wildlife conservation (Larsen et al., 2015; Wilson, 2016). A major factor influencing the proportion of land surface requiring protection is whether PAs are placed strategically.
Historically, the core networks of national parks and reserves were not designed with the sole aim of covering the areas of highest conservation priority systematically; rather, conservation often had to contend with marginal lands that were of limited use for other human activities (Joppa and Pfaff, 2009). Ideally, the more recently established PAs supplementing the core networks would fill their gaps, but because creation of PAs is often opportunistic, and the underlying motivation varies, the extent to which they do so is unclear. Here we focus on three countries in sub-Saharan Africa for which data were available, i.e., Kenya, Tanzania and Namibia, to explore whether communal and private conservancies as a whole are strategically located within the PA network.
Regardless of the answer to this question, the important contribution of communal and private conservancies to conservation in recent years is indisputable (Dudley et al., 2018). In Kenya, a boom in communal and private conservancies over the past two decades involves the Northern Rangeland Trust and the Maasai Mara Wildlife Conservancies Association as key players (Nelson and Cooney, 2018). Over the same timeframe, Tanzania has promoted the establishment of Wildlife Management Areas to engage local communities in conservation (Bluwstein et al., 2018). This policy transfers wildlife user rights to committees of village representatives that negotiate contracts with private investors, with the national government and conservation organizations as advisors. Positive conservation impacts of Wildlife Management Areas are evident despite some challenges in their implementation, notably relating to the role of the government and the level of taxation (Lee, 2018; Nelson and Cooney, 2018). In Namibia, the legislative framework has since the mid-1990s allowed conditional rights to manage and benefit from natural resources to be transferred by creation of communal conservancies where local communities enter into joint ventures or lease arrangements with tourism or trophy hunting enterprises (Naidoo et al., 2016; Nelson and Cooney, 2018).
But has the proliferation of communal and private conservancies covered spatial priorities from a continental perspective effectively? Pinpointing optimal locations for PAs is complicated by the threat posed by climate change. Even PA networks that protect biodiversity well at present may not necessarily do so in the future because of the climate-induced range-shifts expected for many species (Payne and Bro-Jørgensen, 2016b; Keeley et al., 2018). This is especially the case if habitat fragmentation prevents wildlife from moving between PAs, whether because of natural barriers or human land-uses, infrastructural development and fencing (Payne and Bro-Jørgensen, 2016a; Wilson et al., 2016). Identifying the localities that preserve biodiversity most effectively long-term thus requires consideration not only of how changes in climate are likely to affect habitat suitability for wildlife, but also whether connectivity in the landscape will allow animals to track habitat changes. Here we use species distribution modeling to accommodate the expected consequences of climate-change (Guisan et al., 2017).
Ungulates are well suited as indicator taxa to assess how effectively PA networks cover areas of high biodiversity value because of their species richness and ubiquity (Bro-Jørgensen, 2016). Ungulates are often keystone species integral to ecosystem functioning, be it as prey for carnivores (Hopcraft et al., 2010), dispersers of seeds (Feer, 1995), architects of habitats (Prins and van der Jeugd, 1993; Augustine and McNaughton, 2004; Bond, 2008) or contributors to nutrient cycling (McNaughton and Georgiadis, 1986). In Africa, the extraordinary radiation of antelopes makes bovids particularly useful as a barometer of ecosystem health and its response to environmental change (Veldhuis et al., 2019), and in this study, we use bovids as our indicator taxon. Antelope conservation also demands increased attention in its own right as shown by around a third of the species being listed as threatened, and two-thirds with declining population sizes, on the global IUCN Red List (IUCN, 2020).
Taking into account predicted range-shifts because of climate change, we first identified the key priority areas for antelope conservation outside the core PA network in Africa. We then assessed how well community-based and privately managed PAs in the three focal countries captured these spatial priorities by testing their locations relative to locations selected at random. Our results indicate that the success is mixed and that several species are of urgent concern. The findings suggest that that PA network design can be improved by (i) strategic support for locally managed PAs in priority areas, which in turn highlights the need to mainstream conservation priorities into land-use planning at national level, and (ii) enhanced collaboration across national borders.
Methods
Species Distribution Models
We rasterized ESRI shape files of the species distributions maps for 72 African antelope species from the IUCN Red List1 to a 10’ grid scale. Using data on climatic conditions between 1950 and 2000 from WorldClim (Hijmans et al., 2005), we then modeled “presence or absence” as a function of annual precipitation (log), and hottest and coldest monthly temperature using quadratic generalized linear models (GLMs) in the R package BIOMOD (Thuiller et al., 2009); selection of the three predictive variables was based on a principal component analysis and variable importance assessment (Thuiller et al., 2010) of 34 environmental variables describing climate, soil, elevation, evapotranspiration and land cover. We evaluated AIC-selected species distribution models derived from 70% of the data against the remaining 30% by quantifying the area-under-the-curve (AUC), sensitivity, and specificity (Swets, 1988) and noted model accuracy to range from “high” (AUC > 0.9; 69 species) to “useful” (AUC > 0.7; 3 species). Subsequently, we predicted future ranges by informing the species distribution models by climate projections according to three Atmosphere-Ocean Global Circulation Models (AOGCMs), i.e., UKMO HADCM3, NCAR CCSM3, and BCCR BCM2. We then produced multi-climate-model ensemble forecasts of species distributions by requiring that predictions agreed under at least two of the three AOGCMs climate models, thereby minimizing the effect of inconsistencies in the climate models. Since the IUCN distribution maps are alpha shapes indicating the Extent of Occurrence (EOO) rather than the Area of Occupancy (AOO), we followed Thuiller et al. (2006) in applying a weighted “Human Footprint” filter in the simulations to decrease the probability of species populating human-affected areas. The “Human Footprint” incorporates human population density, land transformation, human access, and power infrastructure (Sanderson et al., 2002). Note that the human footprint was modeled as a static filter since projections were not available. The forecasts were based on the balanced A1B greenhouse gas emission storyline (Akçakaya et al., 2014).
We used two alternative approaches to predict the effect of climate change on species distributions by 2080. The first, conservative approach aims to reflect a future where species are prevented from dispersing outside their current ranges because of widespread human land-use; here we only included projected distributions that fell within the current distribution. The second approach aims to reflect what the species distribution might be without human interference; here we modeled the future bioclimatic envelope, i.e., the area projected to be climatically suitable and which is connected spatiotemporally to the current bioclimatic envelope, defined as the area of climatically suitable habitat connected to the current range. Intermediate time steps for assessing connectivity were 2030 and 2050.
Gap Analysis
We used the Marxan software (Ball et al., 2009) to conduct gap analyses of the PA network in Africa based on the forecasts for 2080 produced by the species distribution models; separate analyses were conducted for the conservative and envelope approaches. Marxan selects the set of planning units that best represents specified biodiversity features and allows users to define the total area of the set, the desired emphasis on spatial clustering of units, and the presence of any PAs that are mandatory in the final solution. Data on the PA network came from the UNEP-WCMC/IUCN World Database on Protected Areas (WDPA) (UNEP-WCMC, 2012). We set all PAs listed with an IUCN PA category as mandatory in the final solution; assuming these are generally more consolidated, we henceforth refer to these as “core PAs” and PAs listed without an IUCN PA category as “supplementary PAs.” For species with a range below 20,000 km2, we set protection of the entire range as mandatory because this limit defines the threshold for the extent of occurrence (EOO) below which a species qualifies as “vulnerable” on the IUCN Red List. For other species, we followed Ball et al. (2009) in setting the proportion of the range of species x requiring protection by relating it to a theoretical species y, which requires either 20% (low protection) or 30% (high protection) protection of its 1,000 cell range (∼34,400 km2) using the formula: (xp/yp) ≈ (xt/yt)0.5, where p is the area protected, and t is the total range-size (Ardron et al., 2010). We set the boundary length modifier to reflect a high cost (10,000) to prioritize fewer, larger PAs and thereby promote the connectivity required under climate change. Each species was assigned a penalty factor whereby threatened species were considered more important to the solution (critically endangered 5,000, endangered 4,000, vulnerable 3,000, near threatened 2,000, least concern 1,000; the silver dik-dik [Madoqua piacentinii], which has no threat status, was set as 3,000 because of a similar size range to other vulnerable species). Because of computational restrictions associated with the large dataset, we conducted 1,000 repetitions using simulated annealing and the final solution identified priority areas where at least 750 repetitions agreed (Ardron et al., 2010).
Overlap Between Marxan Priority Areas and Supplementary PAs
For the analysis, we focused on the only three African countries for which the PA descriptions in WDPA included extensive references to community management, i.e., Kenya, Tanzania and Namibia. For these countries, we considered PAs as “community-managed” if either the designation, government type or management authority referred to them as such; none of these community-managed PAs were listed with an IUCN PA category. The PAs which were neither listed with an IUCN PA category nor qualified as “community-managed” are referred to as “supplementary PAs under other management.”
To test whether the overlap between supplementary PAs and Marxan solutions differed from random, we generated randomized values by (i) creating a set of points to represent all cells in a country except the core PA cells; (ii) taking a random selection of those points to represent the number of cells in the Marxan solution for the country; (iii) taking another random selection of cells equal to the number of supplementary PA cells in the country; (iv) determining the number of cells that overlap with the selections created under (ii) and (iii); (v) repeating steps (iii) and (iv) 10,000 times; and (vi) comparing the actual values to the average of the randomizations generated under (v) using X2-tests. Using a similar approach, we also compared the overlap between the Marxan solution and the supplementary PAs according to their management type separately; in this instance, fewer points were included in the sets created under step (i) to exclude cells of the alternative management type. All statistical analyses were conducted in R (R Development Core Team, 2019).
We found only minor differences between the results relating to the high and low protection scenarios, and for simplicity, we therefore display only the former and refer to discrepancies in the text.
Results
Gap Analysis for Africa
The priority areas for antelope conservation identified by the gap analyses of the core PA network in Africa are shown together with the extent of the core PA network and supplementary PAs in the WDPA in Figure 1; separate solutions are illustrated for the bioclimatic envelope approach (Figure 1A), and the conservative approach where species are unable to disperse (Figure 1B).
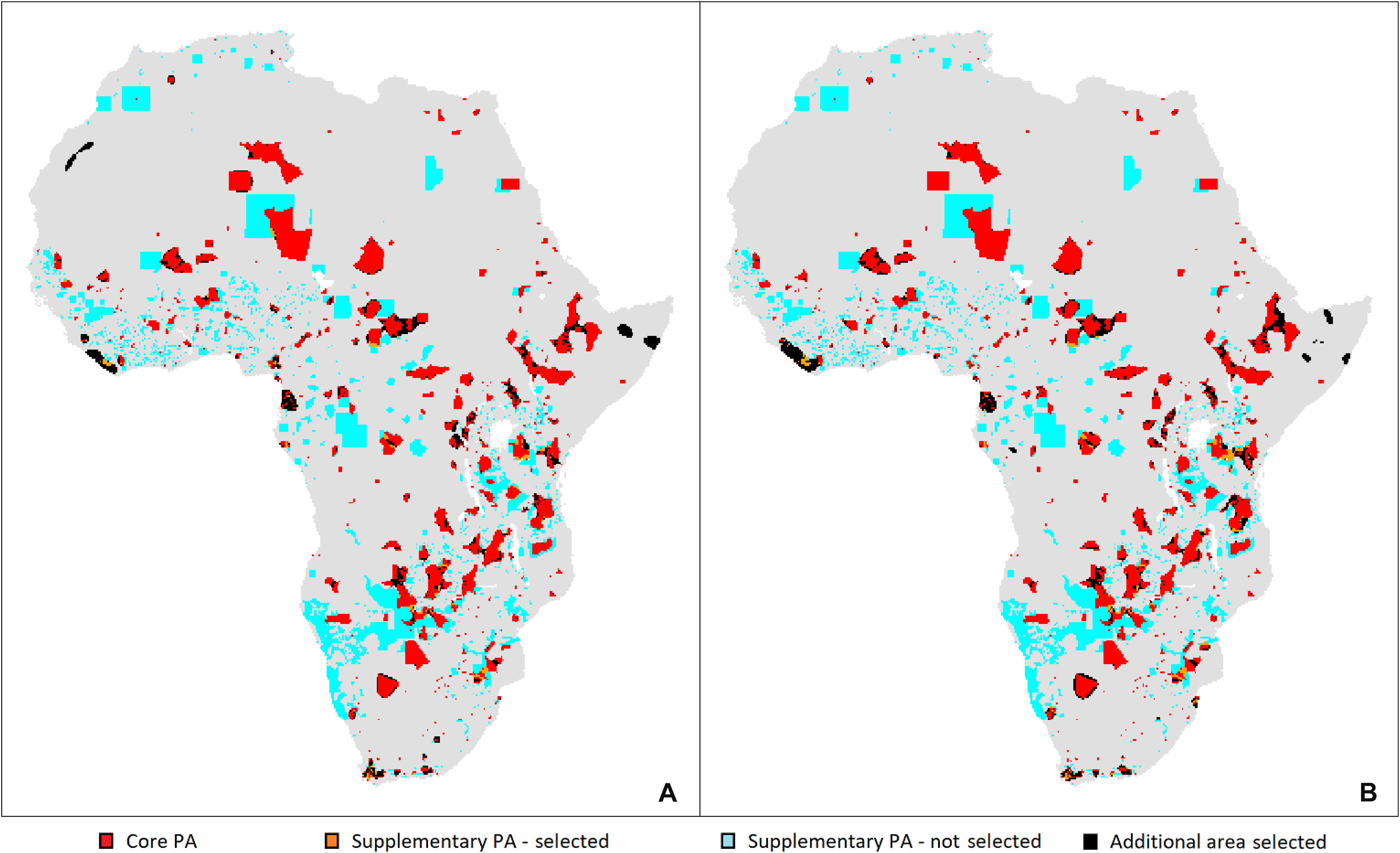
Figure 1. Priority areas for antelope conservation in Africa under climate change. Gaps in the core protected area network identified by Marxan are shown in relation to current protection, where “selected” refers to Marxan priority areas. (A) Solution under the bioclimatic envelope approach. (B) Solution under the conservative approach where species are unable to disperse.
Kenya
Predicted Local Extinctions and Colonizations
Projected patterns in antelope biodiversity in Kenya by 2080 are shown in Figure 2, which indicates both the predicted overlap in bioclimatic envelopes and species richness predicted under the conservative approach. Of the 35 bovid species currently in the country, the hirola (Beatragus hunteri) is forecast to have no suitable area remaining by 2080 when modeling the spatiotemporally connected bioclimatic envelope. When adopting the conservative approach, assuming that dispersal from their current distribution is not possible, this forecast is shared by three additional species, i.e., sable antelope (Hippotragus niger), Ader’s duiker (Cephalophus adersi) and bongo (Tragelaphus eurycerus). The bioclimatic envelope of seven antelope species not currently recorded as extant in the country are forecast to extend into Kenya by 2080, i.e., Soemmering’s gazelle (Nanger soemmerringii), which has been recorded as a vagrant species in the north until recently (Kingdon, 1982), kob (Kobus kob), formerly present in the west of the country (Kingdon, 1982), southern reedbuck (Redunca arundinum), red-flanked duiker (Cephalophus rufilatus), bay duiker (Cephalophus dorsalis), natal red duiker (Cephalophus natalensis), and Sharpe’s grysbok (Raphicerus sharpei).
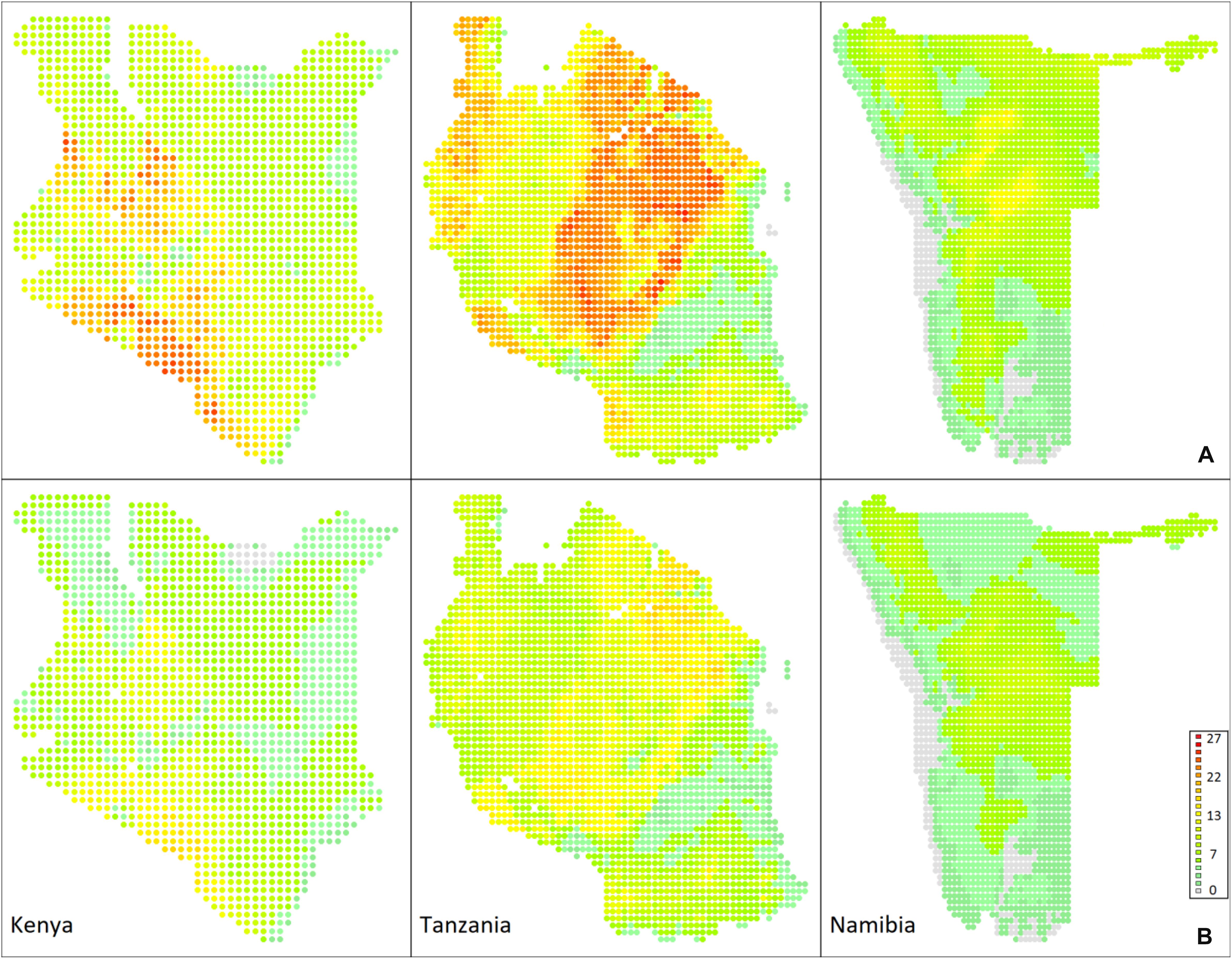
Figure 2. Antelope biodiversity forecasts for 2080 in three sub-Saharan countries. (A) Heat map of the number of species for which bioclimatic envelopes overlap at a site. (B) Heat map of the species richness predicted under the conservative approach where species are unable to disperse.
Gap Analysis for the Core PA Network
Of the priority areas identified by Marxan to supplement the core network of PAs in Africa, those in Kenya include an area improving the connectivity between Tsavo East and West national parks (NPs) in the south as well as an area connecting Tsavo and Amboseli NPs, resulting in a large transfrontier park between Kenya and Tanzania (Figure 3). In the north of the country, a priority area expands Sibiloi NP on the east bank of Lake Turkana to the east and north to join Murle NP and Chelbi Wildlife Sanctuary across the Ethiopian border. Under the high protection scenario only, a priority area in the west of the country connects to the Amudat Community Wildlife Area across the border in Uganda. Finally, priority areas expand the existing core PAs in the center of the country.
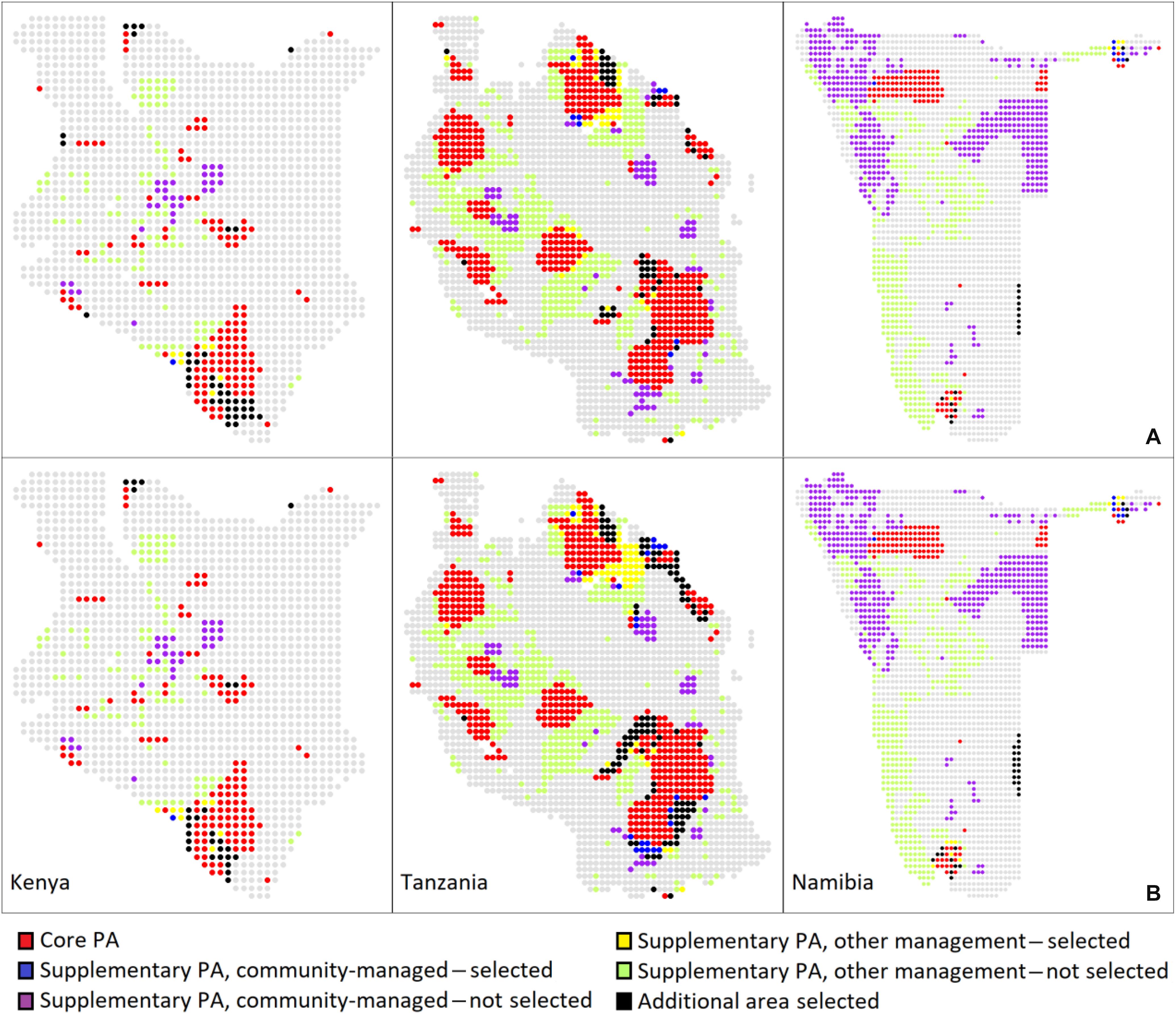
Figure 3. PA networks and priority areas in three sub-Saharan countries. Areas selected for protection to complement the core PA network in a continent-wide gap analysis are shown in relation to the current PA network from WDPA. (A) Solutions under the bioclimatic envelope approach. (B) Solutions under the conservative approach where species are unable to disperse.
Do Supplementary PAs Fill Gaps Effectively?
The supplementary PAs in Kenya were not more likely to cover the African-wide priority areas for antelope conservation located in the country than if they had been situated at random; this was the case whether supplementary PAs was considered as a whole or divided by their type of management (Table 1).
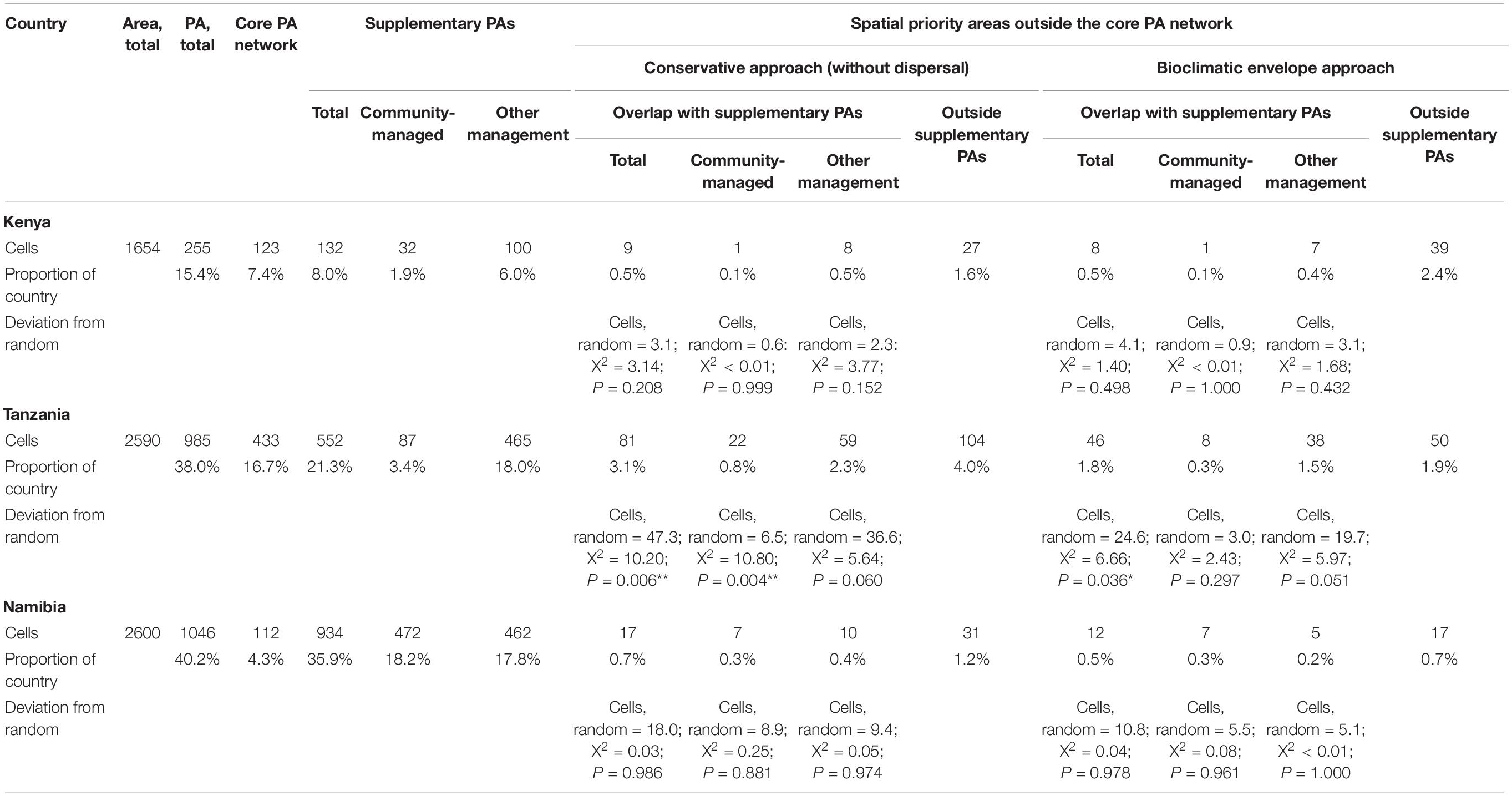
Table 1. Protected area coverage according to WDPA (UNEP-WCMC 2012) and spatial priority areas for antelope conservation in three African countries (*P < 0.05; **P < 0.01).
Tanzania
Predicted Local Extinctions and Colonizations
Projected patterns in antelope biodiversity by 2080 in Tanzania, the country with the highest species richness, are shown in Figure 2, which indicates both the predicted overlap in bioclimatic envelopes and species richness predicted under the conservative approach. Of 37 bovid species currently in the country, the Ader’s duiker is forecast to have no suitable area remaining by 2080, both when modeling the spatiotemporally connected bioclimatic envelope and when assuming that dispersal from their current distribution is not possible. The bioclimatic envelope of five antelope species not currently present in Tanzania are forecast to extend into the country by 2080, i.e., the black-fronted duiker (Cephalophus nigrifrons), yellow-backed duiker (Cephalophus silvicultor), bay duiker, Salt’s dikdik (Madoqua saltiana), and Günther’s dikdik (Madoqua guentheri).
Gap Analysis for the Core PA Network
Priority areas identified by Marxan expand Kilimanjaro and Mikomazi NPs in the northeast to form the large transfrontier park with Kenya mentioned previously. Priority areas also extend Ngorongoro Conservation Area and Serengeti NP and in the high protection scenario using the conservative approach (without dispersal), the transfrontier park covers a contiguous area from Tsavo NPs in Kenya to Lake Victoria in Tanzania (Figure 3). In the southeast of the country, priority areas expand the Selous Game Reserve to the northwest, connecting to the Mufindi Scarp and Kigogo Forest Reserve in the high protection scenario without dispersal; in the scenario without dispersal, Selous Game Reserve also is expanded to the south to connect with community PAs such as Tunduru.
Do Supplementary PAs Fill Gaps Effectively?
As a whole, the supplementary PAs in Tanzania were significantly more likely to cover the African-wide priority areas for antelope conservation located in the country than if they had been situated at random (Table 1). Under the conservative approach only, this also held for community-managed supplementary PAs taken separately; for supplementary PAs under other management, there was a tendency to capture antelope priority areas both under the conservative and envelope approaches (Table 1).
Namibia
Predicted Local Extinctions and Colonizations
Projected patterns in antelope biodiversity in Namibia by 2080 are shown in Figure 2, which indicates both the predicted overlap in bioclimatic envelopes and species richness predicted under the conservative approach. All the 20 bovid species currently in the country retain a spatiotemporally connected bioclimatic envelope within the country by 2080, however, six species are forecast to have no suitable area remaining if dispersal from their current distribution is not possible, i.e., the African buffalo (Syncerus caffer), sitatunga (Tragelaphus spekii), sable antelope, southern lechwe (Kobus leche), southern reedbuck and Sharpe’s grysbok. The bioclimatic envelope of four antelope species not currently present in the country are forecast to extend into Namibia by 2080, i.e., Cape grysbok (Raphicerus melanotis), gray rhebok (Pelea capreolus), bohor reedbuck (Redunca redunca), and lowland nyala (Tragelaphus angasi).
Gap Analysis for the Core PA Network
Priority areas identified by Marxan expand the Ai-Ais Hot Springs NP in the south of the country, and on the eastern border, priority areas connect to the Kgalagadi Transfrontier Park in neighboring Botswana and South Africa (except in the low protection envelope scenario) (Figure 3). In the northeast of the country, priority areas are identified adjacent to Mudumu and Nkasa Rupara NPs in the Zambezi Region, which would strengthen the Kavango-Zambezi Transfrontier Conservation Area linking PAs in Angola, Botswana, Zambia, and Zimbabwe.
Do Supplementary PAs Fill Gaps Effectively?
The supplementary PAs in Namibia were not more likely to cover the African-wide priority areas for antelope conservation located in the country than if they had been situated at random; this held whether supplementary PAs were considered as a whole or divided by their type of management (Table 1).
Discussion
According to our analysis, the PAs supplementing the core PA network in Tanzania are well-placed to capture priorities in antelope conservation at a continental scale, whereas in Kenya and Namibia no significant association appeared. These findings suggest that whereas the supplementary PAs make a valuable contribution to antelope conservation in all three countries, strategic support for conservation initiatives in priority areas may improve the design of PA networks. Our gap analysis recommends expansion of current PAs and increased interconnectivity, which will benefit a wide range of antelopes that qualify as landscape-species in that their requirements reflect more general conservation priorities at the ecosystem level. These taxa include migrating species such as the blue wildebeest (Connochaetes taurinus) and the gazelles, nomadic species such as the common eland (Tragelaphus oryx) and oryxes, and the many species that move seasonally between dry season concentration areas and wet season dispersal areas, e.g., hartebeest (Alcelaphus buselaphus) and the African buffalo (Lamprey, 1963). Several of the priority areas connect PAs across national borders, which underlines the importance of international collaborative networks to establish and manage transfrontier parks.
Our study also identifies particular species which should be afforded special attention in PA design and management because either global or local extinction is predicted. The bioclimatic envelope for the critically endangered hirola, which is extant in Kenya only, is projected to have disappeared by 2080, calling for careful ecological monitoring as well as the establishment of an ex situ population. Recent research supports habitat degradation as a major cause underlying the species’ decline (Ali et al., 2017). If confined to its current distribution, global extinction is also projected for the critically endangered Ader’s duiker, which is extant in Kenya and Tanzania only, however, a spatiotemporally connected bioclimatic envelope remains in Kenya, indicating PAs adjacent to its current distribution as a management priority. In addition, two subspecies of conservation concern are forecast to disappear from Kenya by 2080 if restricted to their current range, i.e., the critically endangered mountain bongo (T. e. isaaci), which occurs in Kenya only, and Roosevelt’s sable antelope (H. n. roosevelti), which is considered critically endangered at the national level. Spatiotemporally connected bioclimatic envelopes remain for both species, again highlighting that PAs adjoining their current distribution may be essential to allow dispersal. The range of the sable antelope, which is now restricted to Shimba Hills National Reserve, previously extended to Tsavo and Malindi (Kock and Goss, 1995), pointing to dispersal corridors and possibly translocation as priorities. For the mountain bongo, the island characteristics of its montane forest habitat present a challenge for protecting a sufficiently interconnected network of reserves to allow dispersal in what is a densely populated part of the country. In Namibia, local extinction is projected for several humid-adapted species, all classified as “least concern” on the IUCN Red List and limited to the extreme northeast of the country where their distributions constitute only a small fraction of their global ranges.
The partial overlap between locally managed PAs and gaps in the core network of primarily nationally managed PAs demonstrates the value of local conservation initiatives to achieve strategic goals. Nonetheless, our study also indicates that many priority areas remain without any protection, bringing the importance of strengthening strategic land-use planning at the national and multinational levels to the fore. Mainstreaming of the conservation agenda into policy-making is an Aichi strategic goal (UNEP, 2010), and at national levels, we recommend a wider application of multi-sector zoning approaches to land-use mapping to promote the allocation of land according to its underlying potential. A priority in this context is also the formulation of explicit fencing policies (Durant et al., 2015). For NGOs providing support for community conservation initiatives, our findings likewise underscore the importance of taking wider spatial priorities into account to maximize beneficial effects.
In the study, we used WDPA to explore gaps in PA networks. WDPA is a valuable resource as the most comprehensive database for PAs available, however, as it relies on data entered by a diverse set of users, ensuring that the same standards are uniformly applied is a challenge which WDPA are making commendable efforts to address. Still, we came across several inconsistencies and omissions in the database. In particular, a few countries have yet to categorize important national parks by IUCN management type, and not all communal and private conservancies are in the database, in spite of the best practice guidelines from the IUCN (Dudley, 2008; Dudley et al., 2014). In line with the advice in the WDPA guidelines, we therefore stress that the present results are to be taken as indicative of general issues, and suggestive of particular concerns worth further investigation, rather than as the basis for firm conclusions.
In closing, we note that although the success of PAs in averting threats to wildlife varies (Leverington et al., 2010; Geldmann et al., 2019), rates of declines in biodiversity are typically far lower inside than outside PAs (Gray et al., 2016), and strengthening both the design and management of PAs remains of paramount importance for conservation. For this purpose, our analysis illustrates how mapping of continent-wide conservation priorities can inform land-use planning and guide policies at national level. A pressing need is now to ensure that data on PA networks are reported in a more consistent manner across the globe to improve the quality of such analyses.
Data Availability Statement
All datasets generated for this study are included in the article/supplementary material.
Author Contributions
BP and JB-J conceived and designed the study. BP performed the analyses. JB-J wrote the manuscript with input from BP.
Funding
Funding was provided by the Duncan Norman Trust.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We thank M. Baylis, J. Hurst, N. Pettorelli, and I. Saccheri, W. Thuiller and two reviewers for comments.
Footnotes
References
Akçakaya, H. R., Butchart, S. H. M., Watson, J. E. M., and Pearson, R. G. (2014). Preventing species extinctions resulting from climate change. Nat. Clim. Change 4, 1048–1049. doi: 10.1038/nclimate2455
Ali, A. H., Ford, A. T., Evans, J. S., Mallon, D. P., Hayes, M. M., King, J., et al. (2017). Resource selection and landscape change reveal mechanisms suppressing population recovery for the world’s most endangered antelope. J. Appl. Ecol. 54, 1720–1729. doi: 10.1111/1365-2664.12856
Ardron, J. A., Possingham, H. P., and Klein, C. J. (2010). Marxan Good Practices, 2nd Edn, Victoria: Pacific Marine Analysis and Research Association.
Augustine, D. J., and McNaughton, S. J. (2004). Regulation of shrub dynamics by native browsing ungulates on East African rangeland. J. Appl. Ecol. 41, 45–58. doi: 10.1111/j.1365-2664.2004.00864.x
Ball, I. R., Possingham, H. P., and Watts, M. (2009). “Marxan and relatives: software for spatial conservation prioritisation,” in Spatial Conservation Prioritisation: Quantitative Methods and Computational Tools, eds A. Moilanen, K. A. Wilson, and H. P. Possingham (Oxford: Oxford University Press), 185–195.
Bluwstein, J., Homewood, K., Lund, J. F., Nielsen, M. R., Burgess, N., Msuha, M., et al. (2018). A quasi-experimental study of impacts of Tanzania’s wildlife management areas on rural livelihoods and wealth. Sci. Data 5:180087. doi: 10.1038/sdata.2018.87
Bond, W. J. (2008). What limits trees in C4 grasslands and savannas? Annu. Rev. Ecol. Evol. Syst. 39, 641–659. doi: 10.1146/annurev.ecolsys.39.110707.173411
Bro-Jørgensen, J. (2016). “Our antelope heritage – why the fuss?,” in Antelope Conservation: from Diagnosis to Action. Conservation Science and Practice Series, eds J. Bro-Jørgensen and D. Mallon (Oxford: Wiley-Blackwell), 1–10. doi: 10.1002/9781118409572.ch1
Dudley, N., Groves, C., Redford, K. H., and Stolton, S. (2014) Where now for protected areas? Setting the stage for the 2014 World Parks Congress. Oryx 48, 496–503.
Dudley, N., Jonas, H., Nelson, F., Parrish, J., Pyhala, A., Stolton, S., et al. (2018). The essential role of other effective area-based conservation measures in achieving big bold conservation targets. Glob. Ecol. Conserv. 15:e00424. doi: 10.1016/j.gecco.2018.e00424
Durant, S. M., Becker, M. S., Creel, S., Bashir, S., Dickman, A. J., Beudels-Jamar, R. C., et al. (2015). Developing fencing policies for dryland ecosystems. J. Appl. Ecol. 52, 544–551.
Feer, F. (1995). Seed dispersal in African forest ruminants. J. Trop. Ecol. 11, 683–689. doi: 10.1017/s0266467400009238
Geldmann, J., Manica, A., Burgess, N. D., Coad, L., and Balmford, A. (2019). A global-level assessment of the effectiveness of protected areas at resisting anthropogenic pressures. Proc. Natl. Acad. Sci. U.S.A. 116, 23209–23215. doi: 10.1073/pnas.1908221116
Gray, C. L., Hill, S. L. L., Newbold, T., Hudson, L. N., Börger, L., Contu, S., et al. (2016). Local biodiversity is higher inside than outside terrestrial protected areas worldwide. Nat. Commun. 7:12306. doi: 10.1038/ncomms12306
Guisan, A., Thuiller, W., and Zimmermann, N. E. (2017). Habitat Suitability And Distribution Models: With Applications in R. Cambridge: Cambridge University Press.
Hijmans, R. J., Cameron, S. E., Parra, J. L., Jones, P. G., and Jarvis, A. (2005). Very high resolution interpolated climate surfaces for global land areas. Intern. J. Climatol. 25, 1965–1978. doi: 10.1002/joc.1276
Hopcraft, J. G. C., Olff, H., and Sinclair, A. R. E. (2010). Herbivores, resources and risks: alternating regulation along primary environmental gradients in savannas. Trends Ecol. Evol. 25, 119–128. doi: 10.1016/j.tree.2009.08.001
IUCN (2020). The IUCN Red List of Threatened Species. Available at: http://www.iucnredlist.org (accessed April 15, 2020).
Joppa, L. N., and Pfaff, A. (2009). High and far: biases in the location of protected areas. PLoS One 14:e8273. doi: 10.1371/journal.pone.008273
Keeley, A. T. H., Ackerly, D. D., Cameron, D. R., Heller, N. E., Huber, P. R., Schloss, C. A., et al. (2018). New concepts, models, and assessments of climate-wise connectivity. Environ. Res. Lett. 13:073002. doi: 10.1088/1748-9326/aacb85
Kingdon, J. (1982). East African Mammals, An Atlas of Evolution in Africa. Vol. IIIC (Bovids). New York, NY: Academic Press.
Kock, R. A., and Goss, T. (1995). Sable sighting in Tsavo East National Park. J. East Afr. Nat. Hist. 84, 19–23. doi: 10.2982/0012-8317(1995)84[19:ssiten]2.0.co;2
Lamprey, H. F. (1963). Ecological separation of the large mammal species in the tarangire game reserve, Tanganyika. Afr. J. Ecol. 1, 63–92. doi: 10.1111/j.1365-2028.1963.tb00179.x
Larsen, F. W., Turner, W. R., and Mittermeier, R. A. (2015). Will protection of 17% of land by 2020 be enough to safeguard biodiversity and critical ecosystem services? Oryx 49, 74–79. doi: 10.1017/s0030605313001348
Lee, D. E. (2018). Evaluating conservation effectiveness in a Tanzanian community wildlife management area. J. Wildlife Manag. 82, 1767–1774. doi: 10.1002/jwmg.21549
Leverington, F., Costa, K. L., Pavese, H., Lisle, A., and Hockings, M. (2010). A global analysis of protected area management effectiveness. Environ. Manag. 46, 685–698. doi: 10.1007/s00267-010-9564-5
McNaughton, S. J., and Georgiadis, N. J. (1986). Ecology of African grazing and browsing animals. Annu. Rev. Ecol. Syst. 17, 39–65.
Naidoo, R., Weaver, L. C., Diggle, R. W., Matongo, G., Stuart-Hill, G., and Thouless, C. (2016). Complementary benefits of tourism and hunting to communal conservancies in Namibia. Conserv. Biol. 30, 628–638. doi: 10.1111/cobi.12643
Nelson, F., and Cooney, R. (2018). Communities Hold The Key To Expanding Conservation Impact In Africa. Available at: https://www.iucn.org/crossroads-blog/201810/communities-hold-key-expanding-conservation-impact-africa (accessed April 18, 2020).
Payne, B., and Bro-Jørgensen, J. (2016a). A framework for prioritizing conservation translocations to mimic natural ecological processes under climate change: a case study with African antelopes. Biol. Conserv. 201, 230–236. doi: 10.1016/j.biocon.2016.07.018
Payne, B., and Bro-Jørgensen, J. (2016b). Disproportionate climate-induced range loss forecast for the most threatened antelopes. Curr. Biol. 26, 1200–1205. doi: 10.1016/j.cub.2016.02.067
Prins, H. H. T., and van der Jeugd, H. P. (1993). Herbivore population crashes and woodland structure in East Africa. J. Ecol. 81, 305–314.
R Development Core Team (2019). R: A Language And Environment for Statistical Computing. Vienna: R Foundation for Statistical Computing. Available at: http://www.R-project.org (accessed December 10, 2019).
Sanderson, E. W., Jaiteh, M., Levy, M. A., Redford, K. H., Wannebo, A. V., and Woolmer, G. (2002). The human footprint and the last of the wild. Bioscience 52, 891–904.
Swets, J. A. (1988). Measuring the accuracy of diagnostic systems. Science 240, 1285–1293. doi: 10.1126/science.3287615
Thuiller, W., Broennimann, O., Hughes, G., Alkemade, J. R. M., Midgley, G. F., and Corsi, F. (2006). Vulnerability of African mammals to anthropogenic climate change under conservative land transformation assumptions. Glob. Change Biol. 12, 424–440. doi: 10.1111/j.1365-2486.2006.01115.x
Thuiller, W., Lafourcade, B., and Araújo, M. (2010). Presentation Manual for BIOMOD. Grenoble: Université Joseph Fourier.
Thuiller, W., Lafourcade, B., Engler, R., and Araújo, M. B. (2009). BIOMOD - a platform for ensemble forecasting of species distributions. Ecography 32, 369–373. doi: 10.1111/j.1600-0587.2008.05742.x
UNEP (2010). Convention on Biological Diversity, Decision X/2: Strategic Plan for Biodiversity 2011-2020. Available at: https://www.cbd.int/doc/decisions/cop-10/cop-10-dec-02-en.pdf (accessed December 20, 2019).
UNEP-WCMC, IUCN, and NGS, (2019). Protected Planet Live Report 2019. Cambridge: UNEP-WCMC, IUCN & NGS.
Veldhuis, M. P., Ritchie, M. E., Ogutu, J. O., Morrison, T. A., Beale, C. M., Estes, A. B., et al. (2019). Cross-boundary human impacts compromise the Serengeti-Mara ecosystem. Science 363, 1424–1428. doi: 10.1126/science.aav0564
Keywords: conservation management, species distribution modeling, national parks, community-based conservation, Bovidae
Citation: Payne BL and Bro-Jørgensen J (2020) Conserving African Ungulates Under Climate Change: Do Communal and Private Conservancies Fill Gaps in the Protected Area Network Effectively? Front. Ecol. Evol. 8:160. doi: 10.3389/fevo.2020.00160
Received: 20 December 2019; Accepted: 07 May 2020;
Published: 16 June 2020.
Edited by:
R. Terry Bowyer, University of Alaska Fairbanks, United StatesReviewed by:
Jacob Goheen, University of Wyoming, United StatesRobin Naidoo, World Wildlife Fund, United States
Copyright © 2020 Payne and Bro-Jørgensen. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Jakob Bro-Jørgensen, YnJvQGxpdi5hYy51aw==
†Present address: Benjamin Luke Payne, Natural England, York, United Kingdom