 Kester Bull-Hereñu
Kester Bull-Hereñu Louis P. Ronse De Craene
Louis P. Ronse De Craene- 1Museo Nacional de Historia Natural, Santiago, Chile
- 2Fundación Flores, Santiago, Chile
- 3Royal Botanic Garden Edinburgh, Edinburgh, United Kingdom
The flower of Malesherbia Ruiz & Pav. (Passifloraceae) is a suitable model to study how far growth constraints throughout ontogeny are causal for the variation in the proportions of reproductive structures. The Malesherbia flower is characterized by a marked hypanthium subtending five alternating sepal and petal lobes plus a coronal rim. In Malesherbia, the size relation between hypanthial tube and perianth lobes conditions the general aspect that the flower of a given species may display. For instance, flowers of taxa belonging to the section Malesherbia are characterized by a predominant hypanthium much similar to tubular flowers with reduced erect perianth lobes and a protruding paracorollar cylinder, while the opposite is true for the remaining species of the genus resembling a radiate ten-parted open flower with a reduced corona. Further morphological variation in the genus includes the bimodal distribution of absolute size of the mature flower, with some species showing much smaller dimensions (e.g., M. humilis) and also the variability in the level of aggregation of the inflorescences ranging from uniflorous (M. lactea), through racemose (M. densiflora, M. lirana) up to very condensed and globular in shape (M. fasciculata). In this work we studied under SEM the flower morphogenesis of 14 Malesherbia species collected in the Andean and desertic region of septentrional Chile. Against expectations, our data showed that the growth of petal lobe primordia is relatively faster in tubular flowers than in radiate ones, despite the presence of a much showier perianth at maturity in the latter. Absolute flower size could also be related to absolute meristem size and a relative developmental arrest was detected in the flower buds of very condensed inflorescences. Our results support the idea of a common constraint throughout ontogeny in which earlier inception and faster organ growth leads to relatively smaller dimensions coupled with earlier maturation and arrest of elongation.
Introduction
Floral phenotypic variation observed in nature is possible due to the existence of unique developmental pathways that lead from floral meristems to mature flowers. How these developmental processes might have diverted through time is doubtless an intriguing question for biologists. One way of dealing with this issue is by trying to understand the drivers that push the development in response to environmental signals and selective forces. A complementary way of understanding these changes is through the lenses of developmental constraints that may explain why certain changes in the morphogenetic process itself lead to particular phenotypic changes or variation (Ronse de Craene, 2018). Understanding principles behind the shifts of developmental processes, if there are any at all, enables us to understand floral morphology and its diversity from an organism’s point of view.
Malesherbia is a South American genus originally considered as the sole representative of the family Malesherbiaceae, but recently placed within the family Passifloraceae (Angiosperm Phylogeny Group, 2016). The genus consists of 27 species that inhabit arid and Andean regions of Perú, Chile and Argentina since the Pliocene colonization of the territory (Gengler-Nowak, 2002; Guerrero et al., 2013). Although the basic bauplan of a Malesherbia flower is constant (five alternating sepal and petal lobes subtended by a manifest hypanthium, five stamens attached to a trilocular ovary elevated by an androgynophore and three protruding styles: Harms, 1894; Ricardi, 1967), the floral morphology looks pretty variable in nature (Figure 1). Clearly, the size relation between hypanthial tube and perianth lobes conditions the general aspect that a given species may display. For example, flowers of taxa belonging to the section Malesherbia (Figures 1A–D) are characterized by having a predominant hypanthium with reduced erect perianth lobes and a prominent corona protruding as a paracorollar cylinder. This construction looks pretty much like a tubular flower of the Asterids (Figures 1A–D). In the remaining species though, the perianth itself is much showier and remains open evoking rather a typical radiate flower (Figures 1E–N). Here, the sepals share a similar coloration as the petals, which creates the aspect of a ten-parted open flower with a corona limited to a discrete toothed rim (Figures 1E–N).

Figure 1. Chilean Malesherbia species studied in this work. (A–D) Species whose flowers show a prominent reddish tubular shape, all of them belonging to the section Malesherbia. (E–N) Species with flowers of radiate shape including minute flowers and globular inflorescence and corresponding to the remaining four sections of the genus. (A) M. tocopillana Ricardi, (B) M. corallina Muñoz-Schick & R.Pinto. Note the prominent corona encircling stamens. (C) M. tenuifolia D.Don. Note the spatial separation between the five anthers and three pink stigmas. (D) M. auristipulata Ricardi, (E) M. rugosa Gay, (F) M. obtusa Phil., note the pilose ovary at the center. (G) M. densiflora Phil. (H) M. linearifolia (Cav.) Pers. (I) M. deserticola Phil. (J) M. fasciculata D.Don, note its condensed inflorescence similar to a head or large glomerule, (K) M. paniculata D.Don seen from the back. Note the hypanthium that sustains the perianth, (L) M. lirana Gay. Picture courtesy of Ludovica Santilli. (M) Annual M. humilis Poepp, note the tiny flowers. (N) M. lanceolata Ricardi. (O) Phylogenetic relationship among the studied species after Guerrero et al. (2013). Differences in branch length are for diagrammatic purposes only.
Further morphological variation in the genus includes the absolute size of the mature flower, with some species showing much smaller flowers as the ones of the annual species M. humilis (below 9 mm in flower width, Ricardi, 1967; Figure 1M); and also variability at the level of aggregation of the inflorescences ranging from uniflorous (M. lactea), through racemose (M. densiflora, M. lirana) up to very condensed and globular in shape as in M. fasciculata (Ricardi, 1967; Figure 1J).
The reason for this variation is linked to changes in the sizes of the meristem and timing of their development (Ronse de Craene, 2018). For example, meristem size has been shown to be correlated with the absolute size of Eucryphia flowers, as the small Eucryphia milliganii develops from smaller flower meristems (Bull-Hereñu et al., 2018). Comparable reduction in size of flower organs in relation to their meristematic size have been reported for the reduction in carpel size in Oxalis (Bull-Hereñu et al., 2016), the reduced sixth staminode of Globba (Iwamoto et al., 2020 this volume), the transformation of stamens into staminodes in Byttneria (Malvaceae) or Sideroxylon (Sapotaceae, Ronse de Craene and Bull-Hereñu, 2016), and even the abortion of the “fifth” petal of Koelreuteria (Sapindaceae, Cao et al., 2018).
On the other hand, the particular globose aspect of M. fasciculata could be developmentally explained simply by the inhibition of growth of the inflorescence internodes after flower production, which would account for an open inflorescence type I sensu Bull-Hereñu and Classen-Bockhoff (2011a). However, as the sister species to M. fasciculata is the uniflorous M. lactea (Gengler-Nowak, 2003), an alternative plausible scenario to explain the globose condensed inflorescence could be the transformation of the ancestor’s flower meristem into a flower-like inflorescence meristem = Floral unit meristem (FUM) capable of producing many flowers (Claßen-Bockhoff and Bull-Hereñu, 2013; Claßen-Bockhoff, 2016). FUMs are related to condensed inflorescences of e.g., Asteraceae, Apiaceae, and further pseudanthia in general. They originate from a meristem that initially enlarges to a great extent, giving rise to a considerable spherical or concave meristem, followed by rapid fractionation of flowers covering its surface. Those meristems, also treated as “MC” meristems (Bull-Hereñu and Classen-Bockhoff, 2011b), normally produce floral buds with high phyllotactic numbers (high number of parastichies) resulting in a low arc-angle of the floral buds on it (see “Open II” meristems in Bull-Hereñu and Classen-Bockhoff, 2011a). As the two alternatives show different generative patterns through ontogeny, screening of inflorescence development would help discriminating between them.
In this study we aim to document the morphogenesis of Malesherbia flowers with focus on perianth, flower size and inflorescence structure, to ascertain whether developmental patterns are influenced by particular constraints that might explain differences in their later mature appearance.
We hypothesize that tubular flowers with reduced and erect petals and sepals would have perianth primordia arising from relatively smaller or inhibited meristematic tissues. Similarly, we expect that minute flowers of M. humilis develop from comparatively smaller floral buds and that the glomerulate inflorescence of M. fasciculata responds to either a inhibition of inflorescence internode growth or a unique appearance of a FUM in the genus.
Materials and Methods
Material Sampling and Microscopy
This study comprises the observation of fourteen of the sixteen accepted Malesherbia species for Chile (Gengler-Nowak, 2003; Figure 1). Flower buds were collected from wild populations in the field (Table 1) and conserved in ethanol 70% before preparation for microscopy. The two Chilean Malesherbia species not considered here are M. lactea for which no reproductive material could be obtained, and M. campanulata (Ricardi, 1967), whose taxonomic identity is somewhat dubious and probably represents a variety nested within M. lanceolata. According to their floral shape, species were designated under one of the four categories: (1) “Tubular,” corresponding to the four species of the Malesherbia section, characterized by tiny almost erect perianth lobes and a prominent floral tube; (2) “Radiate” corresponding to species with open and showy sepal and petal lobes; (3) “Minute,” corresponding to the markedly smaller flowers of the annual M. humilis; and (4) “Glomerulate,” corresponding to the odd M. fasciculata that presents a unique condensed inflorescence resembling more or less a capitulum of Asteraceae or Apiaceae (Figure 1).
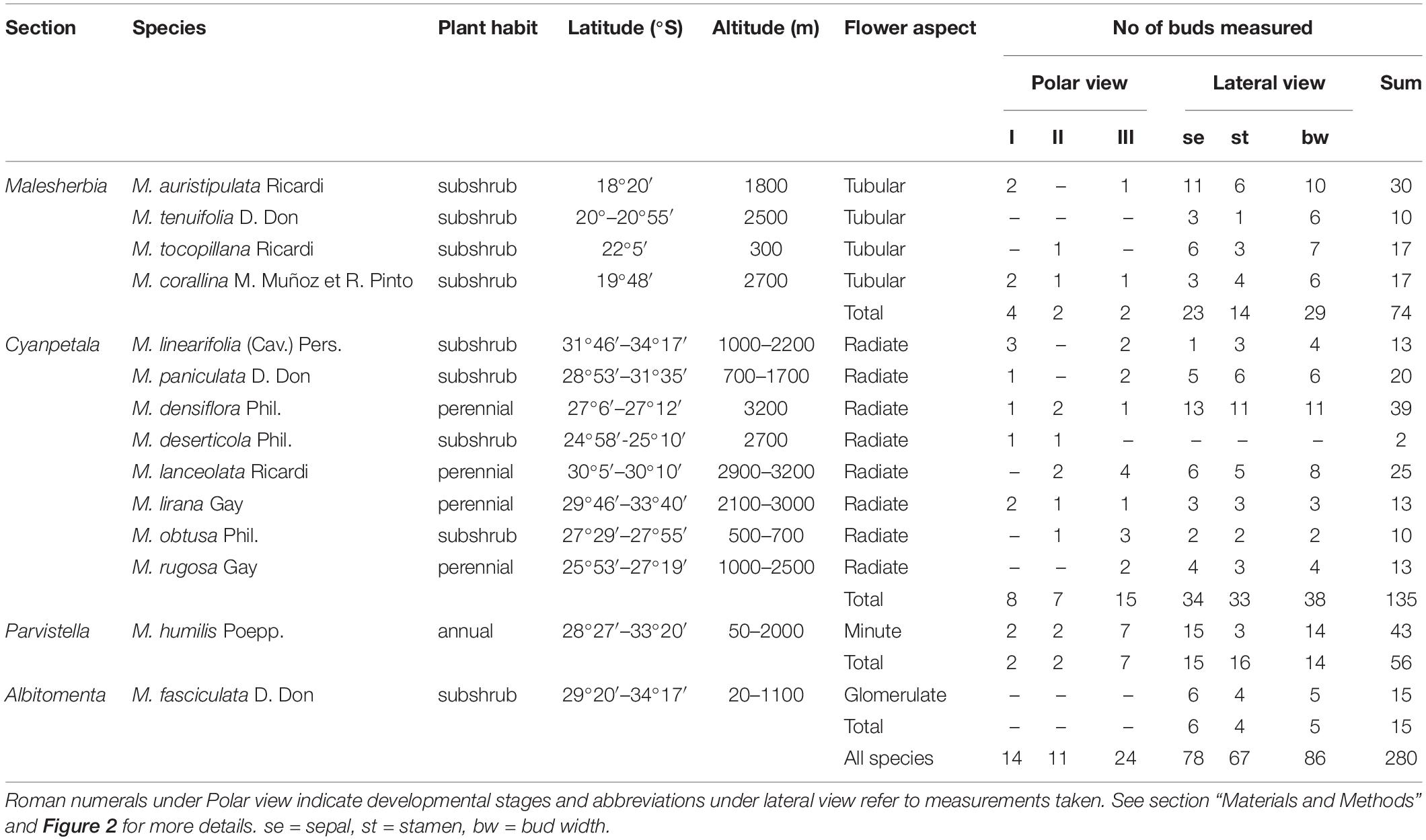
Table 1. Material studied in this work including their taxonomic and morphological identity, geographical provenance and N observed.
Flowers from different stages were dissected under a light microscope (Zeiss Stemi SV6), dehydrated in an alcohol gradient and acetone, dried in a K850 critical-point dryer (Quorum Technologies), further dissected, mounted on aluminum stubs, coated with platinum (Emitech K575X) and observed with a scanning electron microscope (SEM) (Leo Supra 55-VP).
Observations, Measurements and Analysis
From the material visualized from polar views, three stages of flower development were defined (Figure 2): (I) Sepal inception, characterized by the existence of a bare floral meristem (fm) surrounded by the sepal lobes; (II) Hypanthial groove, characterized by the appearance of a ridge surrounding the central floral meristem; and (III) Stamen initiation, characterized by the recognition of anther primordia on the flower. The images obtained from stages I–III were used to measure bud width (bw) and monitor the time of inception of petal primordia. In these stages a total of 49 floral buds were screened (Table 1).

Figure 2. Developmental stages and measurements taken. (A–C) definition of stages I–III on polar views; the key organs or attributes whose presence defines the respective stage marked in white. (D–F) Examples of measurements obtained from lateral views. (A) M. linearifolia (Radiate) showing stage I defined by the presence of a bare floral meristem (fm) surrounded by five just initiated sepal primordia (se). The dashed line represents the measurement of the bud width (bw) from the tip of one of the sepal primordia (on the right) toward the opposite side of the flower bud at the merging point of two other sepal primordia. (B) M. lanceolata (Radiate) showing stage II defined by the appearance of a groove separating the perianth from the future androecium, marking the position of the future hypanthial tube. A white line illustrates a section of this groove. Dashed line represents the measurement of bud width as explained in panel (A). (C) M. obtusa (Radiate) showing stage III defined by the recognition of stamen primordia (st). Dashed line same as in panels (A) and (B). (D) Example of measurements taken from the lateral view of a young flower bud of M. auristipulata (Tubular) represented by dashed lines comprising petal (pe), anther (an) and sepal length (se), and bud width (bw). (E) Example of measurements taken from the lateral view of a relatively older flower bud of M. humilis (Minute) showing petal and sepal length plus bud width. (F) Similar as in panels (D) and (E) but at a younger stage of M. humilis. fm = floral meristem, st = stamen, se = sepal, se = sepal length, pe = petal length, an = anther length, bw = bud width.
Buds in stage III and older were also recorded from lateral views (Figures 2D–F) to measure the length of organs such as sepal lobes (se), petal (pe) and anthers (an) primordia, and flower bud width (bw) as well (Table 1). From lateral views, 231 measurements were taken, which added to the polar views led to a total of 280 measurements taken from SEM micrographs (Table 1). Data obtained were then assigned to one of the four groups defined above, according to the species of provenance (Table 1).
Bud width was screened in stages I–III in order to inspect for differences in the size of the floral bud among the Tubular, Radiate and Minute groups.
Data regarding organ length were analyzed by plotting petal primordium length (pe) against bud width, and anther and sepal length, in order to understand whether there were differences in the relative rates of elongation of the petals between Tubular and Radiate groups.
Finally, the Glomerulate group was qualitatively inspected in terms of inflorescence development relative to the other groups.
Results
Early Developmental Stages I–III
Floral development in Malesherbia starts with the quincuncial inception of sepal lobes around a bare floral apex (Stage I, Figures 3A,D,G,J). Radiate species seem to possess a somewhat larger floral meristem (Figures 3G,J), but this is not numerically corroborated for all species of both groups (Figure 5A). Later on, and after the sepal lobes have grown slightly, a groove surrounding the floral meristem (fm) can be recognized, separating the perianth from the central zone where the stamens will arise (Stage II, Figures 3B,E,H,K). This groove represents a circumferential border separating an external side that will expand to form the hypanthium, and an inner side that will rise to build the androgynophore (see Figure 6N). At this point Tubular species present the appearance of petal primordia in between the sepal lobes (white arrowheads, Figures 3B,E), while these are completely lacking in the Radiate ones (open arrowheads, Figures 3H,K). Interestingly, sepal lobes in species of the Radiate group are markedly more advanced (Figures 3H,K). Later on, as the floral bud enlarges a bit more in width, stamen primordia appear simultaneously on the central floral meristem (stage III, Figures 3C,F,I,L). At this stage, petal primordia are markedly visible in the Tubular group (white arrowheads, Figures 3C,F), while these are either absent or very difficult to see in Radiate species (open arrowheads, Figures 3I,L). Radiate and Tubular species do not differ in the width of their floral buds at any stage (Figure 5A), as the range of the values obtained in both groups fully overlaps. A slight tendency in the increase of the floral width from Stages I through III can be seen in both groups, qualitatively (Figure 3) and numerically (Figure 5A).
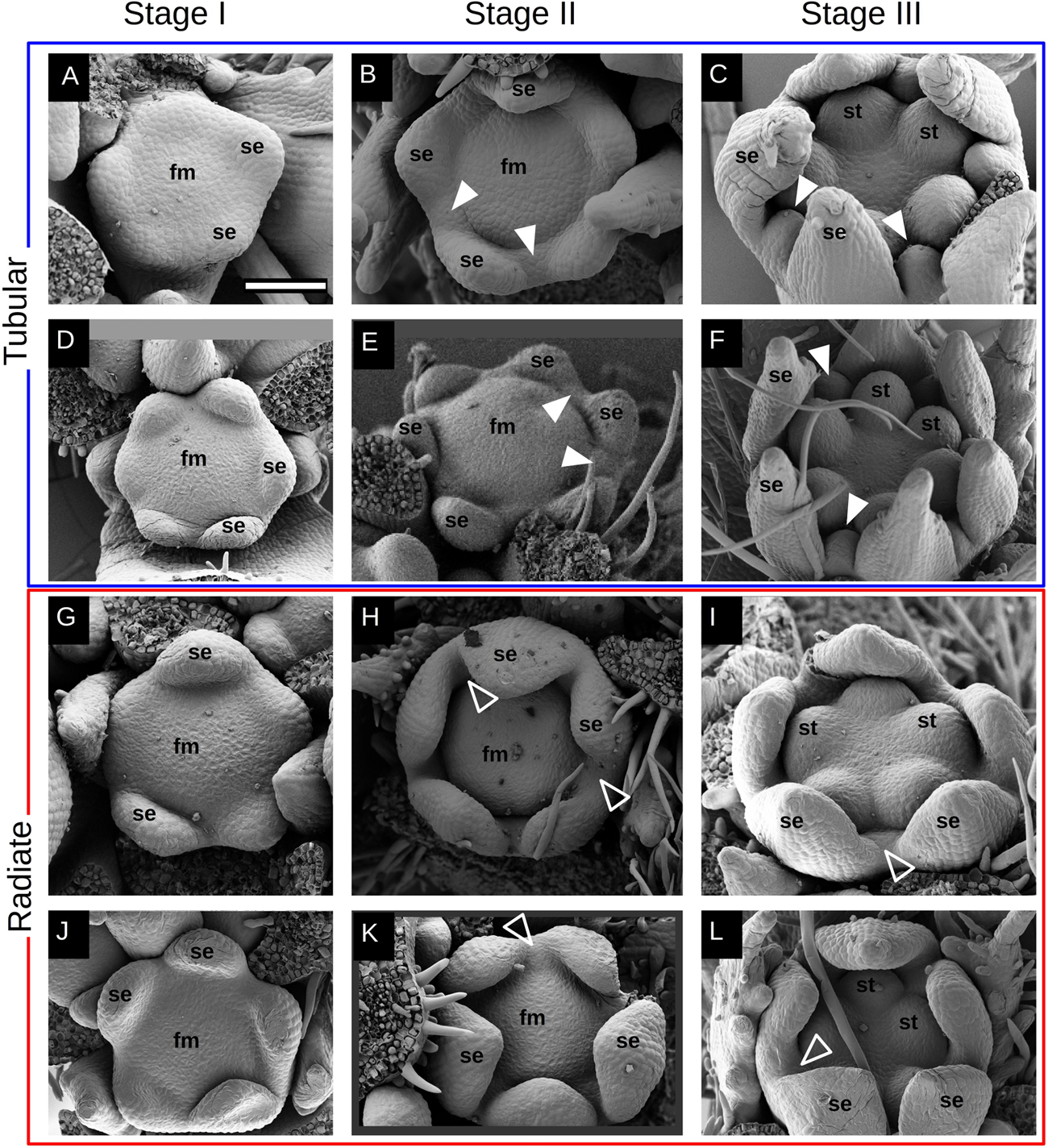
Figure 3. Early development of Malesherbia flower. Images in a same row belong to the same species: Tubular group (blue frame): (A–C) M. tocopillana; and (D–F) M. auristipulata; Radiate group (red frame): (G–I) M. lanceolata; (J–L) M. lirana. Images in the same column correspond to a common developmental stage: (A,D,G,J) stage I: sepal inception; (B,E,H,K) stage II: hypanthium groove initiation; (C,F,I,L) stage III: stamen initiation. (A) Sepal primordia (se) arising around a young flower meristem (fm) of Tubular M. tocopillana. (B) Same species showing a clear hypanthial rim and emergence of petal primordia between sepal lobes (white arrowheads). (C) M. tocopillana showing five round stamen primordia (st), well defined sepal lobes and in between already developing petal primordia (arrowheads). (D) Tubular M. auristipulata flower initiating sepal primordia. (E) Slightly older flower showing petal initiation (arrowheads) between sepal primordia (se). (F) Petal primordia expanding (arrowheads) in between sepal primordia and surrounding the emerging stamen primordia in the center. (G) Radiate M. lanceolata showing sepal (se) inception and a bare flower meristem (fm) in the center. Note the slightly larger size of the young flower in comparison to the Tubular species shown in panels (A) and (D). (H) A clear hypanthial rim is established in the young flower while sepal lobes expand toward the center. At this point, no petal primordium can be seen in the space between sepal primordia (open arrowheads) in contrast with the Tubular species shown in panels (B) and (E). (I) Same species with marked stamen primordia in the center still lacking petal primordia between sepal lobes (open arrowhead). (J) Radiate M. lirana developing flower with a bare flower meristem in the center (fm) and emerging sepal lobes (se). (K) Flower primordium showing hypanthial rim and a sustained growth of sepal lobes. Note the larger sized sepal lobes at this point in Radiate species (H,K), compared to Tubular species (B,E). (L) Stamen primordia clearly shown in the flower center while petal primordia are still lacking (open arrowhead). Bar in panel (A) for all Figures = 100μm. fm = flower meristem, se = sepal, st = stamen. White arrowhead = petal primordium, open arrowhead = petal primordium still not initiated.
Qualitative Differences in Petal, Sepal and Anther Development
After their inception, floral organs begin to elongate (Figures 4, 6). The difference in growth rate of petal primordia becomes evident when comparing species of both groups with either similar anther or petal sizes. Taking a similar anther size as a reference, it is noticeable that while petal primordia begin to acquire a characteristic shape in Tubular species (Figures 4A,G), petals are just visible as small protrusions in Radiate species (Figures 4C,J). For a similar petal size in Tubular (Figures 4B, 6D) and Radiate (Figures 4D, 6E) species, differences in stamen size are evident, being larger in the latter. Comparative observations of the perianth also reveal a slight difference in the inception point of petal lobes: Petal primordia are clearly inserted in between the sepal lobes and at a similar distance from the center in tubular species (Figures 4E, 6C), while in Radiate ones, petal primordia are initiated later and more to the inside of the hypanthium (Figures 4F, 6A,B). This difference in position of petal inception is also visible at later stages (Figures 6D,E).
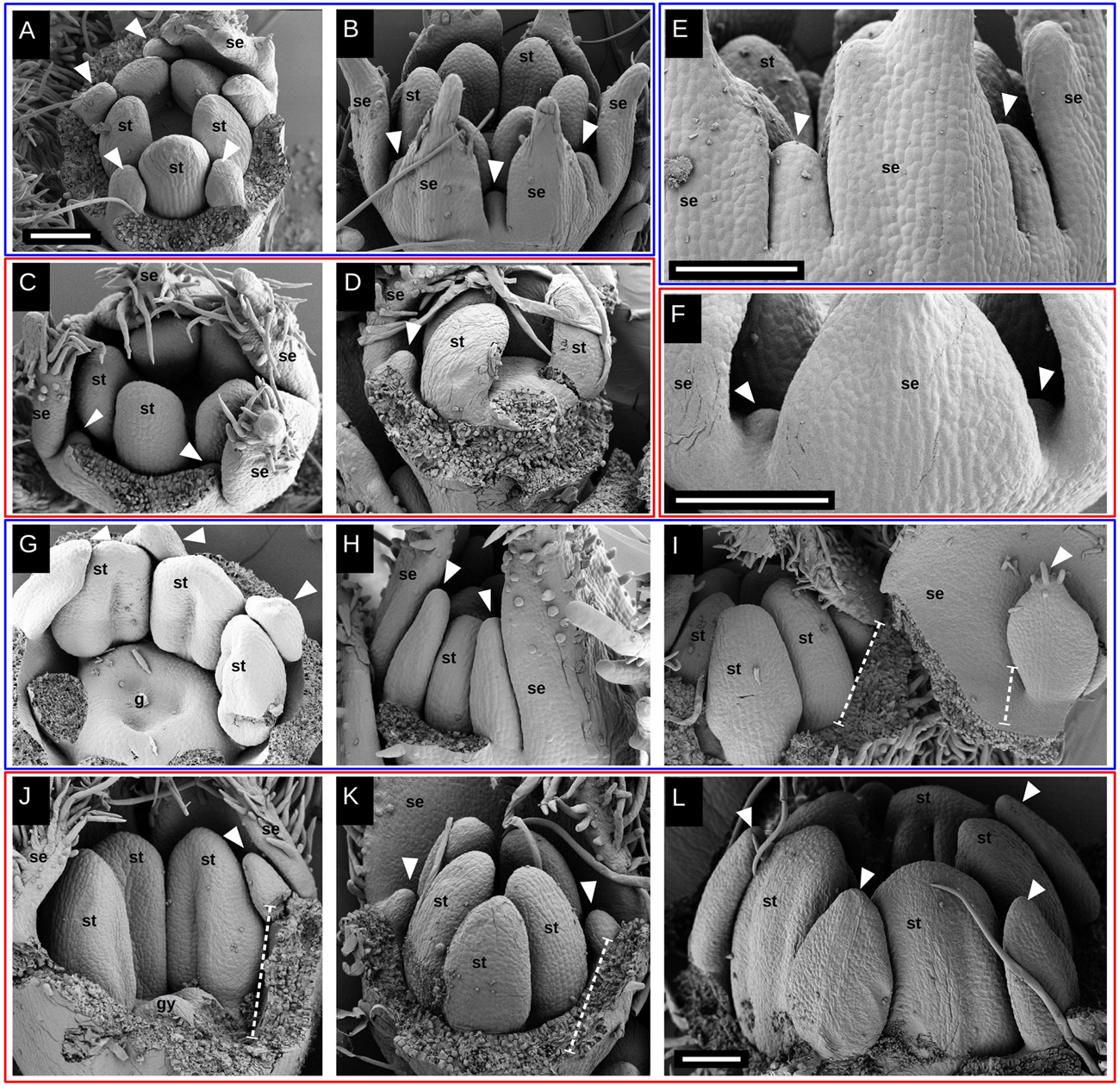
Figure 4. Petal elongation during flower development in Malesherbia. Tubular (blue frame) and Radiate (red frame) species at similar stages for comparison: (A) and (B) v/s (C) and (D); (E) v/s (F); (G,H) and (I) v/s (J,K) and (L). (A) Tubular M. corallina showing petals (arrowheads) elongating and alternating with stamens (st). Sepal lobes removed. (B) Tubular M. auristipulata at a comparable stage showing erect growing young petals. (C) M. lanceolata showing similar stamen (st) primordium size as in Tubular species (A) and (B), but with markedly smaller petal primordia shown by arrowheads. (D) M. densiflora at similar stage regarding stamen primordium size and also showing a small protruding petal primordium (arrowhead). (E) View of perianth of Tubular M. auristipulata showing alternating sepal (se) and petal (arrowheads) lobes all inserted on the external perimeter. (F) Comparable view of Radiate M. obtusa showing much more retarded petal primordia situated more internally than in panel (E). (G) M. auristipulata; young flower showing stamen (st) with thecal ridge differentiation and well-formed petals outgrowing stamens (arrowheads). A trilocular gynoecium (g) covers the center of the flower. Sepals removed. (H) Tubular M. tocopillana at a similar stage as G and equivalent large young petals (arrowheads) outgrowing stamens (st). Sepals (se) enclose flower bud, therefore one sepal is removed. (I) M. tenuifolia showing ventral face of petal (arrowhead) and sepal (se) plus a fraction of the hypanthium (dashed line). This part of the perianth has been torn open from the flower bud that shows young stamens (st). Note the growing trichomes on the petal tip indicating tissue maturation. (J) Radiate M. lanceolata young flower showing gynoecium and stamen differentiation at a similar stage as (G–I). Note the markedly smaller petal primordium (arrowhead) than in previous species. (K) M. densiflora at a similar stage as (J), also showing quite retarded and small petal primordia. (L) M. densiflora showing petals (arrowheads) differentiated to a similar extent as shown for Tubular species in panels (G–I), but with a much larger overall size. Note that even at this size, the petals still do not overgrow the stamens. All bars = 100 μm. fm = flower meristem, se = sepal, st = stamen, gy = gynoecium, white arrowhead = petal primordium, dashed line: hypanthium outline.
While petal lobes have reached and surpassed anthers in Tubular species (Figure 4H), they still are lagging far behind the anther tips in Radiate species (Figure 4K). A more mature aspect of petal primordia in a Tubular species can be seen in Figure 4I showing trichome development on its tip. A similar petal lobe aspect in a Radiate species can be seen in a far more advanced bud stage with larger anthers (Figure 4L).
When taking a similar petal size for both Radiate and Tubular species (Figures 6D,E), it is evident that the anthers of Radiate species appear to be more developed (Figure 6D). This is also true when observing older flower buds as the pair of Tubular and Radiate species shown in Figures 6F,G.
Similar observations can be made when taking sepals as reference organs. Figures 6H,K present a comparable petal size and shape but with much more reduced sepal lobes in the Tubular species (Figure 6H) than in the Radiate ones (Figure 6K). A same tendency is encountered when comparing a general view of the perianth in Figures 6I,J,L. The first two figures represent Tubular species with sepals approximately double the petal size, while Radiate species at this stage present sepals three to four times larger than the petals. Conversely, Radiate species achieve a relative petal size closer to the 50% of the sepal only when they are much more older and larger, as can be seen in Figure 6M.
The petal size pattern observed is reverted when the young flower has grown to a certain extent: petals of Radiate species are capable of overgrowing the floral width as seen in Figure 6N (see also Figure 5B), while Tubular species attain a relative petal size kept below the flower width as seen in Figure 6P. Young flowers of Tubular species show a prominent hypanthium already at this young age and tightly arranged petal lobes at its top (Figures 6O,P), while Radiate species seem to have their corolla more loosely arranged within the flower bud (Figure 6N).
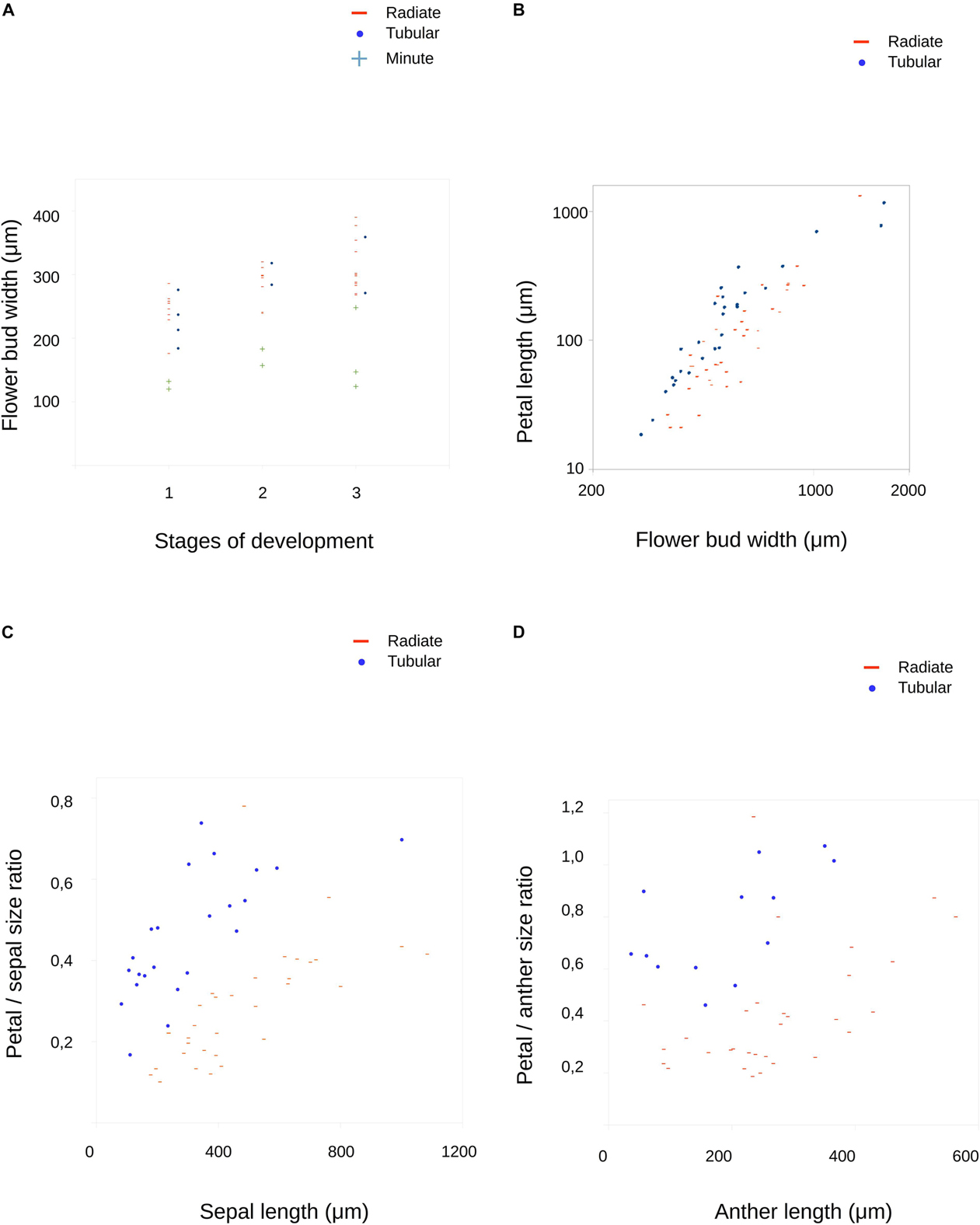
Figure 5. Results of measurements on developing flowers of Malesherbia. (A) Flower bud width along three developmental stages. Three groups are plotted: Radiate (red stripes), Tubular (blue dots) and Minute (green plus symbol, see Table 1 and main text for more explanations). Note that data ranges of Radiate and Tubular species fully overlap, while values for Minute are the smallest overall and well outside the ranges of the other two. (B) Petal primordium length plotted in relation to bud width. Note that Radiate flowers present smaller values in general. Note the logarithmic scale in both axes. (C) Sepal length plotted against relative petal size. (D) Anther length plotted against relative petal size. Blue dots = Tubular group, red stripes = Radiate group, green plus symbol = Minute group.
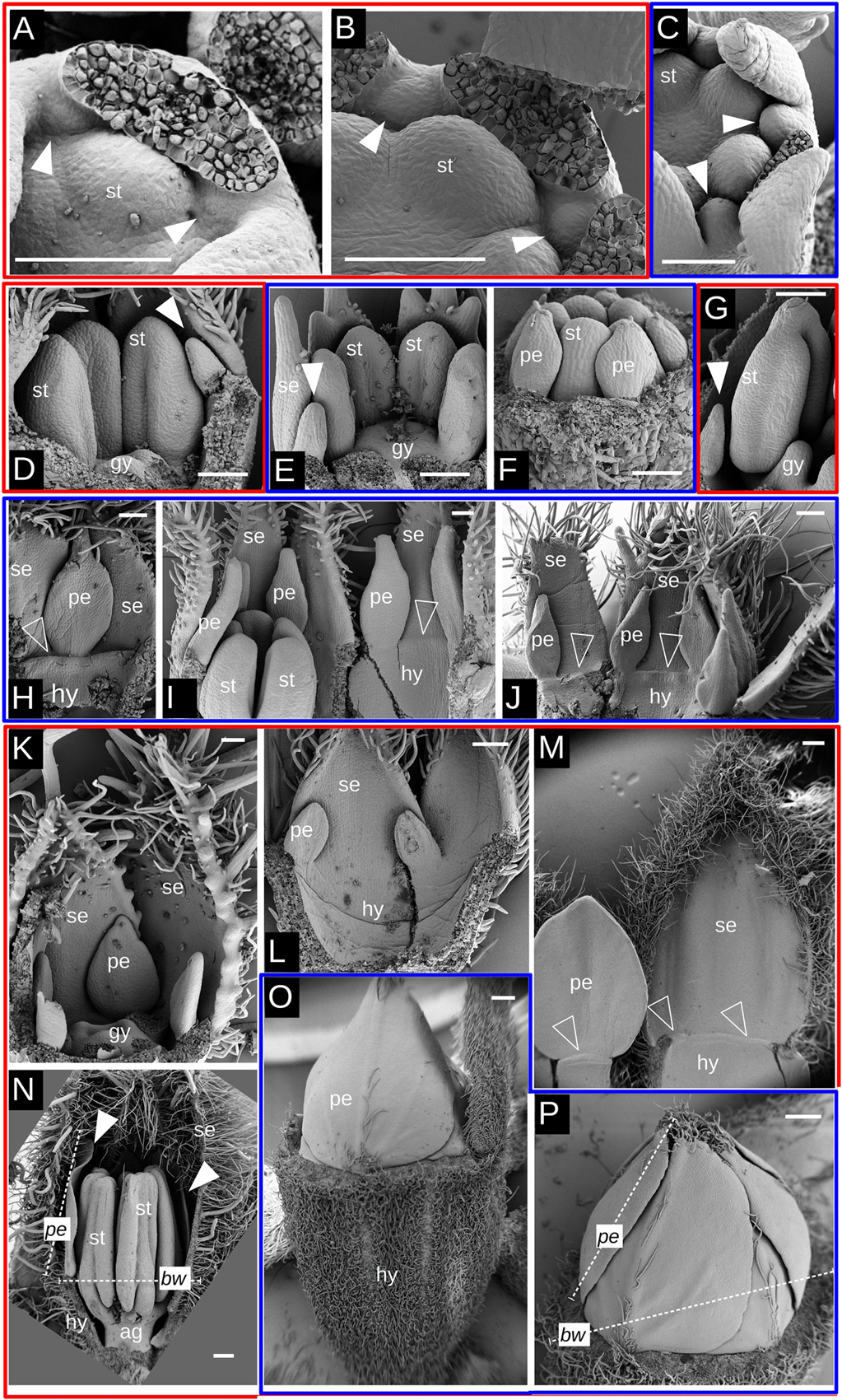
Figure 6. SEM images of petal initiation and elongation in Malesherbia. Red frames surround Radiate species and blue frames Tubular species. (A) M. rugosa (Radiate) showing petal initiation (arrowheads) between stamen primordium (st) and sepals. (B) M. paniculata (Radiate) at a slightly older stage than in panel (A) showing slightly larger petal primordia. Note the position of the petal between stamens (st) and sepals. (C) M. tenuifolia (Tubular) with petal primordia placed well between sepals and being part of the dorsal margin of the flower. (D) M. lanceolata showing a small young petal (arrowhead) (E) M. auristipulata with a similar sized young petal (arrowhead) as in panel (D), but with stamens of clearly smaller size. Note the more external position of the petal than in panel (D). Same magnification in panels (D) and (E). (F) M. tenuifolia showing petal primordia (pe) attached at the tip of a developing hypanthium. Also part of stamens (st) visible. Same magnification in panels (D) and (E) (G) M. linearifolia showing young petal of comparable size as in panel (F), but with a much larger stamen (st). (H) Young petal (pe) and sepal (se) of M. auristipulata of a fairly similar size. Open arrowhead signals an incipient corona along the top of the hypanthium (hy) (I) M. tocopillana flower torn open to show part of the hypanthium, stamens, petals of about half the size of the sepals. (J) Perianth and hypanthium of M. auristipulata flower showing petals roughly half the size of the sepals. (K) M. rugosa showing petals of a comparable size as in panel (H), but with much larger sepals (se). Same magnification in panels (H) and (K). (L) Fraction of perianth of a M. lanceolata flower (Radiate). Note the difference in size between petal and sepal primordia in contrast to the situation in Tubular flowers shown in panels (I) and (J). Same magnification in panels (I,J) and (L). (M) M. lanceolata, showing petal of around half the size of the sepal. Note that this proportion is achieved in a much later ontogenetic stage in this Radiate flowers than for Tubular ones as shown in panels (I) and (J). (N) M. rugosa young flower with well developed organs. At this advanced stage, the petal length (pe) is close to or already larger than the bud width (wd). See panel (P) for the opposite case. (O) M. tenuifolia young flower showing petals folded in the bud and an important hypanthium formed. (P) M. tenuifolia (Tubular) showing petals folded with cochlear aestivation. At this advanced stage, the petal length (pe) does not surpass the flower bud width (bw), contrary to a Radiate flower as shown in N. Bars (A–L) = 100 μm, (M–P) = 200 μm. Same magnification among (D–G); (H,K); (I,J) and (L) se = sepal, pe = petal, st = stamen, gy = gynoecium, hy = hypanthium, ag = androgynophore, white arrowhead = petal primordium, open arrowhead = incipient corona, pe = petal length, bw = bud width, dashed white line = example of measurement.
Quantitative Elongation of Floral Organs
We quantitatively monitored the behavior of petal growth by plotting floral bud width values against petal length (Figure 5B). Tubular species present measurable petal primordia at smallest flower widths (blue dots, Figure 5B) and the overall tendency to be larger than the ones of Radiate species (red stripes) at similar bud width (Figure 5B).
The growth of petal primordium size relative to sepal or anther primordium size is shown as both increase during bud growth (Figures 5C,D). Here, the results found are similar as before: Tubular species present in general relatively larger petal primordia for a given referential organ size. For example, in Tubular species, petals equal anther size when anther primordia reach a length of around 400 μm (blue dots, Figure 5D), while in Radiate species petals do not reach anther size at the stages here screened, but attain a size of around 80% of the respective anther (red stripes, Figure 5D). A similar observation is made for the size relationship petal/sepal (Figure 5C), as the petal relative size in Tubular species is larger than in Radiate species, although not achieving the complete sepal size, but around 75%, while Radiate species achieve around 50% when sepals achieve a size of around 1,000 μm.
Corona Growth
As in other Passifloraceae, Malesherbia flowers present a corona rim that surrounds the stamens. This is more prominent in Tubular species (Figures 1A–D) where the corona forms a small tube, even reaching the level of the anthers as in M. auristipulata (Figure 1D). In the remaining species, the corona is limited to a whitish or yellowish toothed rim (Figures 1H–N). A corona is already well differentiated in Tubular young flower buds (Figures 7A–C). It protrudes ventrally from the hypanthial tissue just below petal and sepal attachment (Figures 7B,D–H). The distal margin of the corona is always somewhat toothed but to different degrees, varying from more or less even (Figures 7A,D,I), to markedly irregular (Figures 7C,H,L). Corona inception occurs when floral organs and hypanthium are already well developed and there seems to be no difference in the petal size of Tubular and Radiate species when it is initiated (Figures 7E–G). The first evidence of the corona appears as a swelling of the hypanthial tissue below the petal insertion (Figures 7E–G) that later expands and establishes its own growing margin (Figures 7H–K). This tissue may proliferate and elongate markedly in Tubular species (Figures 7D,I), remain more or less arrested in Radiate ones (Figures 7J,K), or also generate irregular crests (Figure 7L).
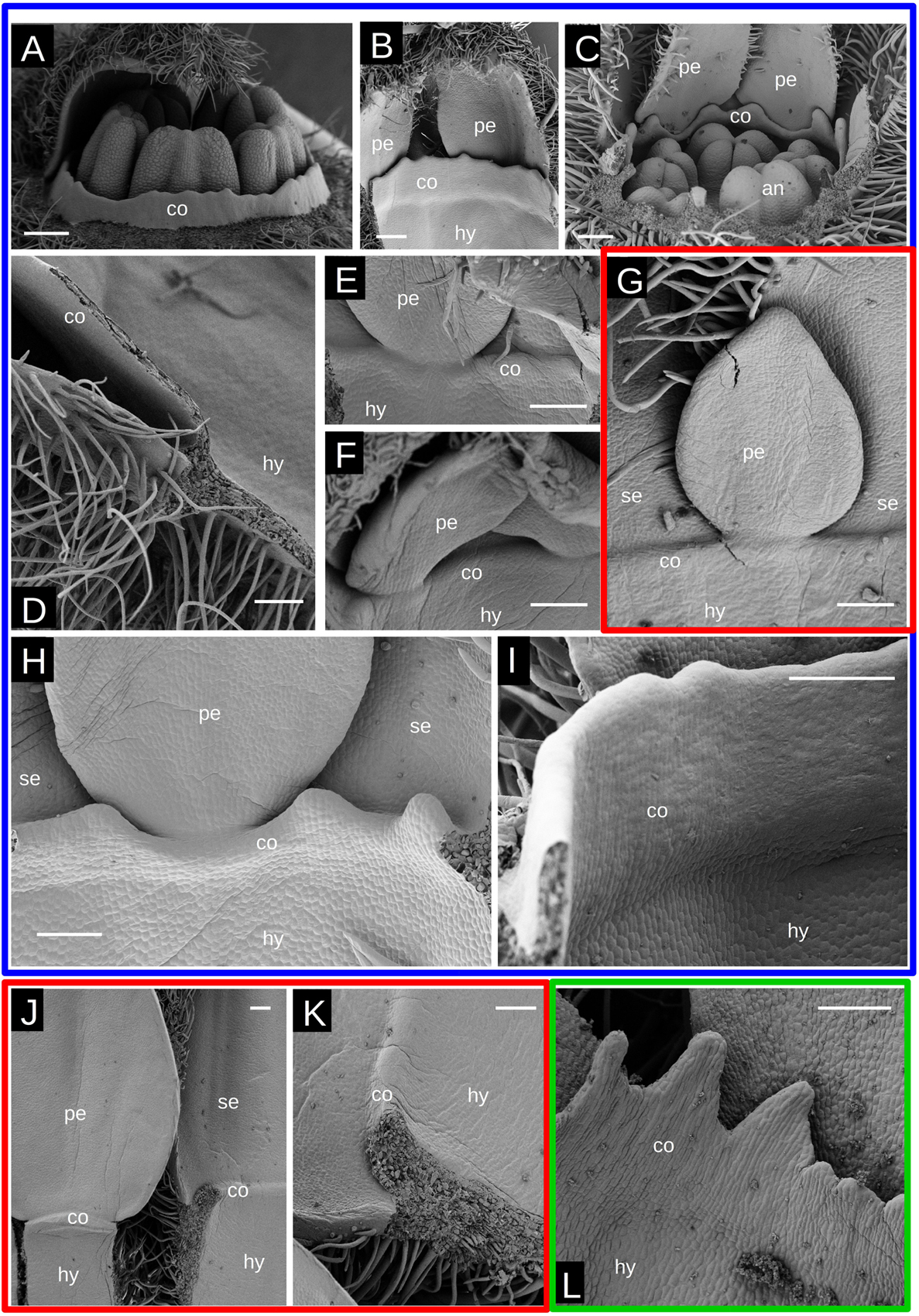
Figure 7. Corona growth in Malesherbia. Red frame surrounds a Radiate species, green frame the minute species and blue frames surround Tubular species. (A) M. corallina showing a corona rim surrounding stamens. (B) Same species showing the perianth and corona rim from its ventral side. (C) M. tocopillana showing its more irregular corona surrounding androecium. (D) Detail of longitudinally sectioned well developed corona in M. auristipulata. (E) Corona being initiated in M. corallina. (F) Corona initiation in M. tenuifolia. (G) Corona initiation in M. densiflora. Note similar petal size in panels (E–G) (same magnification). (H) Corona growth in M. tocopillana. Note the irregular toothed shape. (I) Corona growth in M. auristipulata resembling a solid wall. (J) Delayed corona growth in M. lanceolata. (K) Same as in panel (J) showing detail of the slow growing corona. (L) Corona growth in M. humilis showing prominent teeth. co = corona, se = sepal, pe = petal, an = anther, hy = hypanthium. Same magnification panels (E–G). Bars (A–C) = 200 μm, (D–L) = 100 μm.
Glomerulate Inflorescence and Minute Flowers
The development of the condensed glomerulate inflorescence of M. fasciculata is shown in Figure 8A. Many undifferentiated flower primordia subtended by bracts surround an inflorescence meristem (Figure 8A). In contrast, inflorescences of Tubular (Figure 8B) and Radiate (Figures 8C,D) species show floral primordia growing and maturing soon after being produced by the inflorescence apex. Bracteoles and sepals can be seen arising on flower primordia (Figures 8B–D). The size of the flower buds is quite different among species, but in particular in the Glomerulate inflorescence (Figure 8E) they show the smallest dimensions (compare with Figures 8D,F, same magnification).

Figure 8. Scanning electron micrographs of flower and inflorescence buds in Malesherbia. Red frame surround Radiate species, green frame the minute species and blue frames surround Tubular species. Glomerulate species without a frame. (A–F) Inflorescences, (G–I) Malesherbia humilis (Minute), (J–K) Radiate species. (A) Developing inflorescence of M. fasciculata (Glomerulate). Here, the inflorescence meristem is not particularly more expanded than the other species, nor any particularly higher phyllotaxis is manifest at floral bud initiation, thus there is no evidence for a FUM. (B) M. tocopillana, (C) M. deserticola, (D) M. linearifolia. (E) M. fasciculata; note the smaller size than (D) (same magnification D,E). (F) M. tocopillana; note larger bud size than M. fasciculata in panel (E) (same magnification E,F). (G) Stage I of M. humilis. (H) Same species in stage II. (I) Stage III of M. humilis. (J) Stage I of Radiate M. rugosa. (K) Stage II in Radiate M. paniculata. (L) Stage III of M. rugosa. Note the overall smaller bud size of M. humilis (G–I) compared to the equivalent stage of Radiate species (J–L), same magnification (G–L). All bars = 100 μm. im = inflorescence meristem, fm = flower meristem, se = sepal, st = stamen, br = subtending bract, bl = bracteole.
When observing the stages I–III of flower development of the Minute group (Figures 8G–I), they all appear clearly smaller than any other Radiate species (Figures 8J–L, same magnification), which is numerically corroborated by the measurements (Figure 5A). The size of M. humilis (Minute) floral bud width at these stages corresponds to nearly half of the rest.
Discussion
Floral morphogenesis in Malesherbia shows that Tubular flowers initiate their petals earlier than Radiate flowers, and also with a faster growth rate. The petal inception in Tubular species occurs in stage II, while Radiate flowers initiate their petals in stage III or even later. The faster growth rate of Tubular petals is recognized both by their higher absolute dimension for a given bud width, and also by their higher relative size in relation to the sepal or anther within the same bud.
This fact may be somehow counterintuitive as Tubular species present the smallest petals at maturity, hence the expectation would be that these would grow slower from the beginning on, but the opposite is true.
Can this difference in growth rate be explained in terms of meristematic tissues? No evident difference in flower meristem size was found between Radiate and Tubular groups, but the position of the petal primordium was different: in Tubular flowers it is situated well between sepal lobes and at the same level, while in Radiate flowers, the position of the petal primordium is shifted more to the center. This positional shift in Radiate species is probably a consequence of the later initiation of the petal primordium in stage III or later. At this time, sepal lobes are considerably more expanded in Radiate flowers and the space between them is probably no longer available; therefore the site of initiation of the petal is pushed toward the next available space more to the center of the bud. This difference may be linked to the size difference of petals at maturity, as in Tubular flowers the petals remain confined between the erect sepal lobes and corona, and have limited space for expansion, while in Radiate flowers the more inward position of the petals allows for a greater expansion.
Interestingly enough, an initial slower organ growth rate of the petal primordia seems here to be related with a larger mature organ size. This is widely recognized for the petal development in several core eudicots (see Ronse De Craene, 2008). For example, in Caryophyllaceae petals lag very much behind the stamens, even arising from common stamen-petal primordia, but they overtake the stamens in size prior to anthesis (Wei and Ronse De Craene, 2019). Some evidence in the literature has shown this pattern to exist in other flower types affecting other organs. For example, the size of the style in heterostylous Oxalis (Oxalidaceae) species has been shown to be inversely related to its initial growth rate, implying that initially faster growing carpels attain a smaller mature size (Bull-Hereñu et al., 2016). In analogy, in a comparative study of the flower development of Eucryphia (Cunoniaceae) it has been shown that relatively slow-growing androecia attain larger dimensions at maturity (Bull-Hereñu et al., 2018). At the organismic level, the same phenomenon has been described in the slow-growing regional varieties of the composite Chaetanthera moenchioides that attain larger sizes in the adult state (Bull-Hereñu and Arroyo, 2009). Also in controlled experiments, a temperature induced slower plant growth can lead to larger phenotypes as demonstrated for the larger capitula in Microseris pygmaea (Battjes and Bachmann, 1994) and Senecio (Abbott and Schmitt, 1985) and larger petals in Capsella (Neuffer and Paetsch, 2013). This principle allows for hypothesis testing in natural occurring patterns, as e.g., the increase of floral size with elevation: overall lower temperatures in mountain environments that slow growth rates in plants living there, could lead to larger floral aspects at maturity (see Bull-Hereñu et al., 2018).
The corona in Malesherbia arises when the organs are fully formed, representing a late morphogenetic phenomenon, similar to the documented observations for the corona of the sister genus Passiflora (Prenner, 2014; Claßen-Bockhoff and Meyer, 2016). In Malesherbia, the corona seems to initiate when petals have attained a certain size both in Radiate and Tubular flowers, which implies a comparatively earlier initiation in Tubular species, as Radiate petal lobes have been shown to develop slower. The earlier initiation of the corona in Tubular species correlates with its larger dimension and a small perianth at maturity. In radiate species the opposite occurs: a late initiation of the corona is linked to its reduced dimension and a large perianth at maturity. As for petal size, no relation could be found between initial meristematic size and corona size. Such changes in shape of the flower and morphometric adaptations are realized here as a matter of space occupation. Building a long tube is correlated with a longer corona and prevents petals from expanding, while a shorter tube leads to a reduced corona and an expanded perianth.
The Minute flowers of Malesherbia humilis proved to be formed by comparatively smaller flower meristems. This phenomenon has been also observed in Eucryphia milliganii, a considerably smaller flower within a genus with larger flowers, that also originates from relatively smaller flower meristems (Bull-Hereñu et al., 2018). Similar corroboration has been found for smaller cleistogamous flowers of Viola (Violaceae, Mayers and Lord, 1984), Collomia (Polemoniaceae, Minter and Lord, 1983) or Pseudostellaria (Caryophyllaceae, Luo et al., 2012) that develop from comparatively smaller meristematic sizes.
Smaller flower meristems were also observed in M. fasciculata, a species that also presents small flower size at maturity, comparable to that of M. humilis. Unfortunately, these flower meristems could only be observed qualitatively at early inception (Figures 8A,E), as no measurements of stages I-III could be performed due to a lack of material. The globular inflorescence of M. fasciculata does not arise from a FUM as it shows an active inflorescence meristem and no meristematic expansion followed by flower fractionation typical of a FUM (Claßen-Bockhoff and Bull-Hereñu, 2013; Claßen-Bockhoff, 2016). This means that the globose aspect of the inflorescence of M. fasciculata is due mainly to inhibition of internode elongation. Here the floral segregation seems to occur very rapidly, as flower primordia appear homogeneous all around the inflorescence meristem. A parallel between inhibited flower meristem size and inhibited inflorescence internodes is here manifest and probably speaks for a common tendency for changing growth rates at various levels of the development. This faster development here is again related to smaller flower and inflorescence sizes, similar to the perianth of Tubular species.
Conclusion
We found a general tendency in Malesherbia flower ontogeny in showing that timing of development of organs leads to manifest phenotypic variation. As already seen elsewhere for other species, an initial rapid development of organs correlates here with smaller dimensions of petals in Tubular species and in the compact globose inflorescence. Earlier inception of the corona in Tubular species could also account for petal size reduction and a larger corona tube. The absolute size of the meristem was also found to correlate with smaller flower size at maturity. The high diversity in flower morphology among species of Malesherbia demonstrates that subtle changes in growth rates are responsible for floral diversification and evolution.
Data Availability Statement
The datasets generated for this study are available on request to the corresponding author.
Author Contributions
KB-H collected and prepared the material, made observations with the SEM, performed imaging and measurements, as main writing. LR together with KB-H analyzed and interpreted the data and contextualized it in the present day disciplinary discussion. All authors contributed to the article and approved the submitted version.
Funding
This work has been funded by Conicyt-Fondecyt project nr. 11150847.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We appreciate the help of following persons in the field in collecting floral materials: Alan Bull, Diego Penneckamp, Camila Gómez, Stefany Navarrete, Jimena Arriagada, Jaime Martínez, Catalina Rivera, Markus Jerominek, Maria Will, Isidora Sepúlveda, Augusto Cornejo, Ludovica Santilli, and Vanezza Morales. We also appreciate the field access facilities and permissions granted by CONAF, GASCO. We also thank Frieda Christie for technical assistance with the SEM. The Royal Botanic Garden Edinburgh (RBGE) was supported by the Scottish Government’s Rural and Environmental Science and Analytical Services Division.
References
Abbott, R. J., and Schmitt, J. (1985). Effect of environment on percentage female ray florets per capitulum and outcrossing potential in a self-compatible composite (Senecio vulgaris L. var hibernicus Syme). New Phytol. 101, 219–229. doi: 10.1111/j.1469-8137.1985.tb02828.x
Angiosperm Phylogeny Group (2016). An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG IV. Bot. J. Linn. Soc. 181, 1–20. doi: 10.1111/boj.12385
Battjes, J., and Bachmann, K. (1994). Phenotypic plasticity of capitulum morphogenesis in Microseris pygmaea (Asteracea: Lactuceae). Ann. Bot. 73, 299–305. doi: 10.1006/anbo.1994.1035
Bull-Hereñu, K., and Arroyo, M. T. K. (2009). Phenological and morphological differentiation in annual Chaetanthera moenchioides (Asteraceae) over an aridity gradient. Plant. Syst. Evol. 278, 159–167. doi: 10.1007/s00606-008-0126-8
Bull-Hereñu, K., and Classen-Bockhoff, R. (2011a). Ontogenetic course ans spatial constraints in the appearance and disappearance of the terminal flower in inflorescences. Int. J. Plant. Sci. 172, 471–498. doi: 10.1086/658922
Bull-Hereñu, K., and Classen-Bockhoff, R. (2011b). Open and closed inflorescences: more than simple opposites. J. Exp. Bot. 62, 79–88. doi: 10.1093/jxb/erq262
Bull-Hereñu, K., Ronse De Craene, L., and Pérez, F. (2016). Flower meristematic size correlates with heterostylous morphs in two Chilean Oxalis (Oxalidaceae) species. Flora 221, 14–21. doi: 10.1016/j.flora.2016.02.009
Bull-Hereñu, K., Ronse De Craene, L., and Pérez, F. (2018). Floral meristem size and organ number correlation in Eucryphia Cav. (Cunoniaceae). J. Plant Res. 131, 429–441. doi: 10.1007/s10265-018-1030-0
Cao, L., Liu, J., Lin, Q., and Ronse de Craene, L. (2018). The floral organogenesis of Koelreuteria bipinnata and its variety K. bipinnata var integrifolia (Sapindaceae): evidence of floral constraints on the evolution of monosymmetry. Plant Syst. Evol. 304, 923–935. doi: 10.1007/s00606-018-1519-y
Claßen-Bockhoff, R. (2016). The shoot concept: still up to date? Flora 221, 46–53. doi: 10.1016/j.flora.2015.11.012
Claßen-Bockhoff, R., and Bull-Hereñu, K. (2013). Towards an ontogenetic understanding of inflorescence diversity. Ann. Bot. 112, 1523–1542. doi: 10.1093/aob/mct009
Claßen-Bockhoff, R., and Meyer, C. (2016). Space matters: meristem expansion triggers corona formation in Passiflora. Ann. Bot. 117, 277–290.
Gengler-Nowak, K. (2002). Reconstruction of the biogeographical history of malesherbiaceae. Bot. Rev. 68, 171–188. doi: 10.1663/0006-8101(2002)068[0171:rotbho]2.0.co;2
Gengler-Nowak, K. (2003). Molecular phylogeny and taxonomy of Malesherbiaceae. Syst. Bot. 28, 333–344.
Guerrero, P. C., Rosas, M., Arroyo, M. T. K., and Wiens, J. J. (2013). Evolutionary lag times and recent origin of the biota of an ancient desert (Atacama–Sechura). Proc. Natl. Acad. Soc. U.S.A. 110, 11469–11474. doi: 10.1073/pnas.1308721110
Harms, H. (1894). “Malesherbiaceae,” in Die Natürlichen Pflanzenfamilien. III Teil Abt 6a, ed. A. Engler (Leipzig: Verlag von Wilhelm Engelmann), 65–68.
Iwamoto, A., Ichigooka, S., Cao, L., and Ronse De Craene, L. P. (2020). Floral development reveals the existence of a fifth staminode on the labellum of basal Globbeae. Front. Ecol. Evol. 8:133. doi: 10.3389/fevo.2020.00133
Luo, Y., Bian, F., and Luo, Y. (2012). Different patterns of floral ontogeny in dimorphic flowers of Pseudostellaria heterophylla (Caryophyllaceae). Int. J. Plant Sci. 173, 150–160. doi: 10.1086/663166
Mayers, A., and Lord, E. (1984). Comparative flower development in the cleistogamous species Viola odorata. III. A histological study. Bot. Gaz. 145, 83–91. doi: 10.1086/337430
Minter, T., and Lord, E. (1983). A comparison of cleistogamous and chasmogamous floral development in Collomia grandiflora Dougl. Ex Lindl. (Polemoniaceae). Am. J. Bot. 70, 1499–1508. doi: 10.1002/j.1537-2197.1983.tb10853.x
Neuffer, B., and Paetsch, M. (2013). Flower morphology and pollen germination in the genus Capsella (Brassicaceae). Flora 208, 626–640. doi: 10.1016/j.flora.2013.09.007
Prenner, G. (2014). Floral ontogeny in Passiflora lobata (Malpighiales, Passifloraceae) reveals a rare pattern in petal formation and provides new evidence for interpretation of the tendril and corona. Plant. Syst. Evol. 300, 1285–1297. doi: 10.1007/s00606-013-0961-0
Ronse de Craene, L., and Bull-Hereñu, K. (2016). Obdiplostemony: the occurrence of a transitional stage linking robust flower configurations. Ann. Bot. 117, 709–724. doi: 10.1093/aob/mcw017
Ronse De Craene, L. P. (2008). Homology and evolution of petals in the core eudicots. Syst. Bot. 33, 301–325. doi: 10.1600/036364408784571680
Ronse de Craene, L. P. (2018). Understanding the role of floral development in the evolution of angiosperm flowers: clarifications from a historical and physicodynamic perspective. J. Plant Res. 1313, 367–393. doi: 10.1007/s10265-018-1021-1
Keywords: perianth, petals, sepals, hypanthium, corona, flower development, morphogenesis
Citation: Bull-Hereñu K and Ronse De Craene LP (2020) Ontogenetic Base for the Shape Variation of Flowers in Malesherbia Ruiz & Pav. (Passifloraceae). Front. Ecol. Evol. 8:202. doi: 10.3389/fevo.2020.00202
Received: 30 January 2020; Accepted: 02 June 2020;
Published: 08 July 2020.
Edited by:
Alistair P. McGregor, Oxford Brookes University, United KingdomReviewed by:
Simone Pádua Teixeira, University of São Paulo, BrazilJoseph Francis McKenna, Oxford Brookes University, United Kingdom
Copyright © 2020 Bull-Hereñu and Ronse De Craene. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Kester Bull-Hereñu, a2VzdGVyQGxhYm9yYXRvcmlvZmxvcmVzLmNs