Abstract
Climate change is forecasted to generate a range of evolutionary changes and plastic responses. One important aspect of avian responses to climate change is how weather conditions may change nestling growth and development. Early life growth is sensitive to environmental effects and can potentially have long-lasting effects on adult phenotypes and fitness. A detailed understanding of both how and when weather conditions affect the entire growth trajectory of a nestling may help predict population changes in phenotypes and demography under climate change. This review covers three main topics on the impacts of weather variation (air temperature, rainfall, wind speed, solar radiation) on nestling growth. Firstly, we highlight why understanding the effects of weather on nestling growth might be important in understanding adaptation to, and population persistence in, environments altered by climate change. Secondly, we review the documented effects of weather variation on nestling growth curves. We investigate both altricial and precocial species, but we find a limited number of studies on precocial species in the wild. Increasing temperatures and rainfall have mixed effects on nestling growth, while increasing windspeeds tend to have negative impacts on the growth rate of open cup nesting species. Thirdly, we discuss how weather variation might affect the evolution of nestling growth traits and suggest that more estimates of the inheritance of and selection acting on growth traits in natural settings are needed to make evolutionary predictions. We suggest that predictions will be improved by considering concurrently changing selection pressures like urbanization. The importance of adaptive plastic or evolutionary changes in growth may depend on where a species or population is located geographically and the species’ life-history. Detailed characterization of the effects of weather on growth patterns will help answer whether variation in avian growth frequently plays a role in adaption to climate change.
Introduction
Avian growth can affect fledgling survival and recruitment, and early life experiences are known to impact adult phenotypes (Haywood and Perrins, 1992; Lindström, 1999; Maness and Anderson, 2013; Merrill and Grindstaff, 2018). Given gradual warming and increasingly stochastic weather conditions under climate change, some growth strategies might result in higher fitness than others. Therefore, predicting future changes in fitness and demography under climate change could depend on understanding the effects of current weather variation on offspring growth. The consequences of weather variation on offspring development will likely be complex and will, to some degree, depend on a species’ life-history and ecological niche.
Growth patterns in birds vary, and differences among populations may be adaptations to their local environments (Ricklefs, 1968; Emlen et al., 1991; Starck and Ricklefs, 1998). Global patterns of variation in avian growth within species can generate hypotheses for how the changing climate will influence growth-related traits. For example, given the observation that individuals tend to be smaller in warmer habitats both within and across species (Bergmann’s Rule; Bergmann, 1847 as cited in Salewski and Watt, 2017) we might predict that for a given species, natural selection will favor nestlings that fledge at a smaller size in a warmer climate (Millien et al., 2006). Introduced House Sparrows Passer domesticus in North America and Australia both exemplify predictions from Bergmann’s rule: birds in the colder regions of both continents are on average larger (Johnston and Selander, 1964, 1973; Andrew et al., 2018). Clines in body size could be driven by natural selection developmental constraints, or both (Fleischer and Johnston, 1984; Andrew et al., 2018). Effects on size during development are important because many bird species approach their final structural size before fledging, meaning there may be little opportunity for compensation later in life (Gill and Prum, 2019).
Plastic adjustment of growth could allow rapid adaptation to changing conditions, but evolution of growth traits might be required if current environmental cues no longer predict future environments. To predict possible phenotypic changes in nestling growth we need to understand how weather variation affects nestling growth, how new weather conditions under climate change are changing the selection on avian growth traits, and the additive genetic and environmental variance of growth traits. Some studies have identified carry-over effects of growth on adult fitness with negative and positive consequences for body size (Gardner et al., 2011; Teplitsky and Millien, 2014; van Gils et al., 2016), but few studies examine whether patterns in growth are adaptive or maladaptive. Examining growth-environment relationships jointly with patterns of genetic change is important because phenotypic plasticity can mask evolutionary change (Merilä et al., 2001; Bonnet et al., 2017; Dobson et al., 2017), and plastic responses can contribute to an adaptive or maladaptive response to changing weather conditions (Ghalambor et al., 2007; Snell-Rood et al., 2018). An essential aspect of studying growth is to quantify it in a way that allows among population, individual and genetic variation to be estimated (Box 1).
Box 1. Methods for quantifying growth.
Avian growth can be investigated using a broad range of traits and different statistical modeling approaches. Often, measurements of size or shape near fledging are used because these are more comparable across studies and can predict post-fledging survival (Maness and Anderson, 2013). However, single time point measurements can hide variation in growth among individuals if individuals follow different trajectories to the same final size (Figure 1).
Repeated measurements made throughout nestling development can be used to fit a curve to the data as a function of age. For growth data, logistic models are often used, and the model’s asymptote, maximum relative growth rate, and timing of maximum growth may be estimated as biological parameters of interest describing growth (Figures 1, 2A; Tjørve and Tjørve, 2010; Aldredge, 2016). The unified family of growth models proposed by Tjørve and Tjørve (2010, 2017) allows flexible, comparable, and biologically interpretable estimates from various growth model families. Biologically, the asymptote should provide an estimate of the maximum size reached by a nestling. However, the asymptote in many models may not be a good indication of a nestling’s fledging mass, as a nestling could fledge before the maximum mass is reached, or in some avian species nestlings reach an asymptote and then decline in mass before fledging.
Such function-valued trait approaches summarize the growth trajectory and enable researchers to explore whether different environments affect the growth rate, timing, or size of a nestling. Linear and polynomial functions could also be used, with the terms of the polynomial being treated as the parameters of interest. Instead of fitting functions to a growth trajectory, sometimes the average difference between consecutive daily measurements (e.g., Peck et al., 2004; Harter, 2007; Divoky et al., 2015) or the daily deviation from growth in ideal conditions are used as an indication of daily growth (Keller and Noordwijk, 1993).
Character-state models provide a flexible method for modeling growth, where one can estimate age-specific measurements as separate, yet correlated traits (Roff and Wilson, 2014; Figure 2B). Note that character-state and polynomial functions are equivalent in many situations. In a character-state approach covariances among age-specific masses can be fitted across all ages, or be restricted to consecutive ages only, in what is called an “ante-dependence” model (see the supplementary material of Hadfield et al., 2013; Thomson et al., 2017; Hadfield, 2019). Both function-valued trait and character state approaches to modeling growth are identical to models that measure trait plasticity across environments, but replace environmental variation with age (Falconer, 1952; Via and Lande, 1985; Jong, 1990; Gavrilets and Scheiner, 1993; Chevin et al., 2013). Function-valued trait approaches can perform poorly if individuals or genotypes have different curve shapes and can be harder to interpret biologically (Roff and Wilson, 2014). Generally, function-valued trait approaches use fewer parameters. Therefore, when sufficient data are available, comparison of function-valued and character-state approaches might be valuable in determining when during ontogeny function-valued trait approaches are not capturing variation during growth (Morrissey and Liefting, 2016). If a linear model is used, environmental effects on the slope or intercept of the line could be detected, yet a gain or loss of mass at any specific time during growth, which could be biologically important, would be difficult or impossible to detect. A character-state might be better able to determine when during ontogeny the environment affects a trait. Houslay (2017) provides accessible tutorials implemented in the R package MCMCglmm for both character state and function-valued trait approaches that could be used to model avian growth (Hadfield, 2010).
Function-valued trait and character-state approaches can be analyzed in a mixed model framework, so that hierarchical (co)variances are estimated (function-valued trait approach, Aldredge, 2016; character-state approach, Thomson et al., 2017). Further, Bayesian models now allow the fitting of flexible models that can estimate the fixed effects of weather and environmental variation on specific parameters or age-specific traits in a single model (Hadfield, 2010; Bürkner, 2017).
FIGURE 1
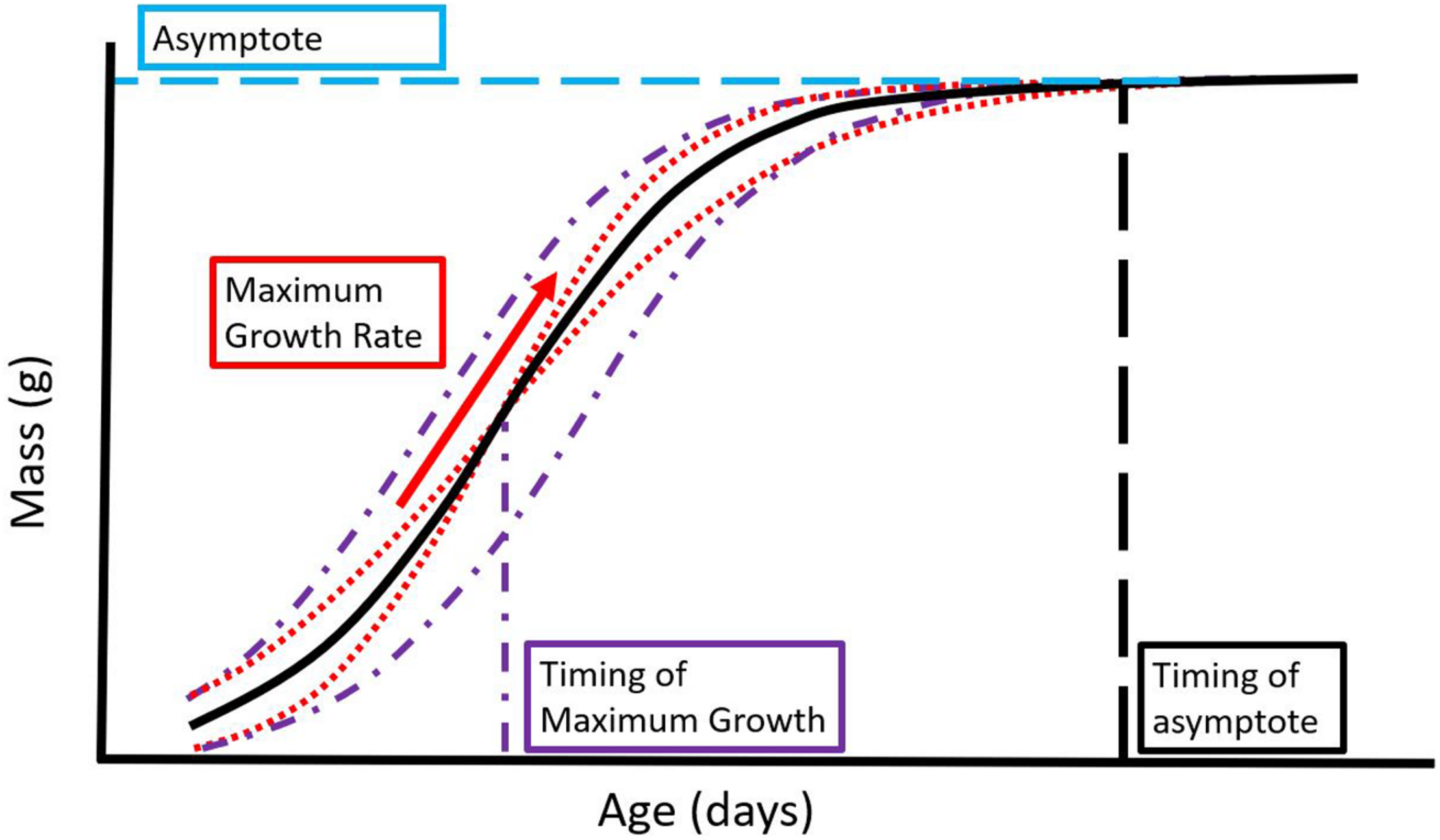
Parameters estimated in a standard logistic curve. The maximum growth rate (red arrow) gives an indication of the speed of growth, the asymptote gives an indication of the nestlings’ maximum size achieved during the period of study (blue horizontal dash), the timing of maximum growth gives an indication of when during ontogeny maximum growth occurs (purple/vertical dash + dotted line), and the timing of the asymptote might indicate developmental timing (black vertical dashed line). Five growth curves are shown, with the same asymptote but varying in the timing of maximum growth (purple dash and dotted lines) or in maximum growth rate (red dotted lines). All curves have the same asymptote as the reference curve (thick black curve) but vary in their trajectory. If a study of nestling growth only measures mass at the asymptote as in this example, variation throughout the trajectory of the growth curve is hidden. Whether this variation impacts fitness is currently unknown.
FIGURE 2
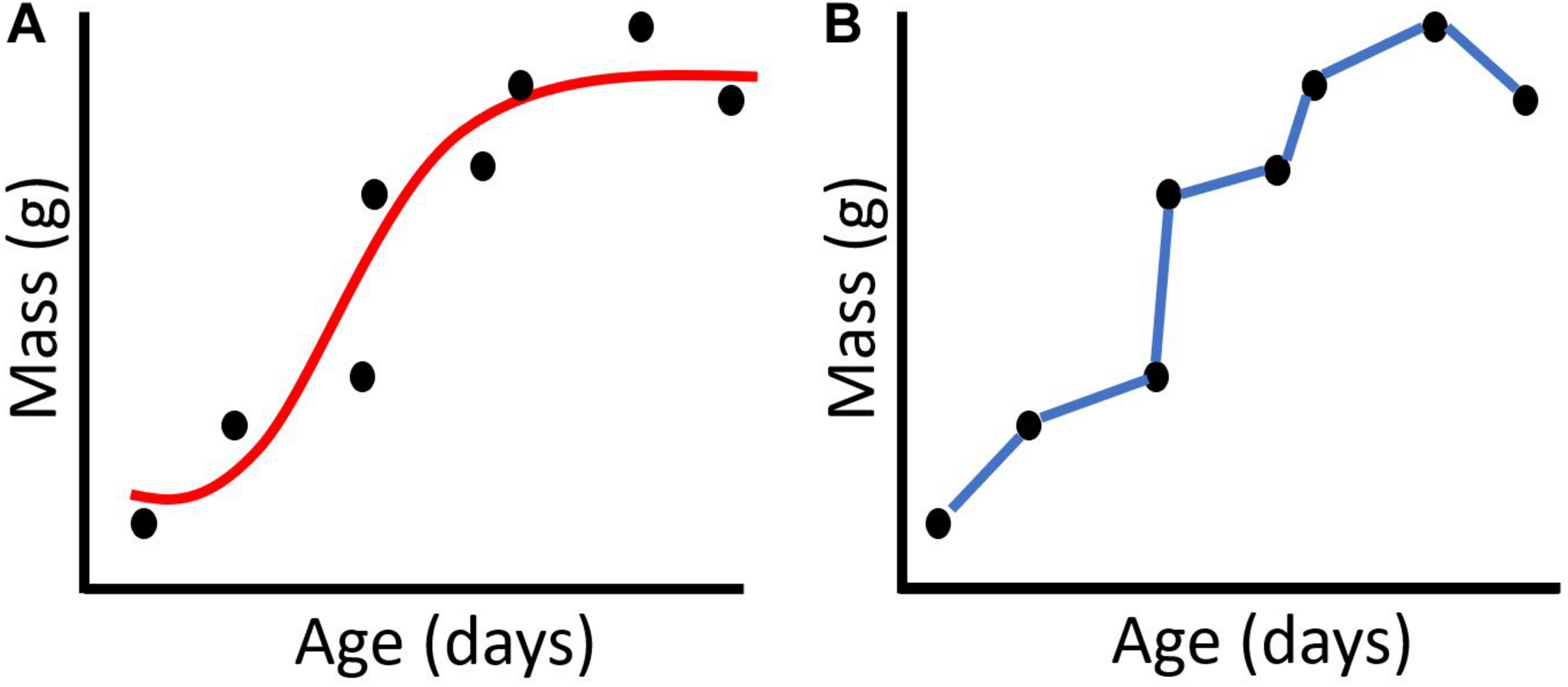
Hypothetical example of a (A) function-valued trait approach and a (B) character-state approach for modeling a chick growth curve.
In this review, we (1) highlight why studying weather-related changes in avian growth might be important to understand avian responses to climate change, (2) review the currently documented effects of weather on avian growth in wild populations, and (3) highlight that understanding the selection operating on growth traits and the inheritance of growth traits is needed to predict future phenotypic responses. We define weather as local within-year variation in temperature (of air and sea surface), solar radiation, precipitation and wind, and climate as the average weather across multiple years for a given region. We explore the effects of minimum, maximum, average, and duration (heatwaves, rainfall) of weather variables.
Importance of Studying Weather-Related Changes in Avian Growth
Understanding weather-related changes in avian growth is important for two reasons: some individuals or genotypes might survive better than others in stressful weather conditions, and weather variation may change a population’s demography through influences on nestling traits.
Plasticity of Avian Growth and Development
Because of genetic, environment, or parental effects individuals will differ in their growth. Understanding the causes of these growth differences is important because some individuals may have higher fitness under changing conditions (Kruuk, 2004; Wilson et al., 2010). In theory, the same individual or genotype could also produce several different phenotypes given different weather conditions (phenotypic plasticity), and the plasticity of an individual’s phenotype could also vary among individuals because of genetic, permanent environment, or parental effects. Plastic changes have gained recognition as common responses to changing environmental conditions (Ghalambor et al., 2007; Charmantier and Gienapp, 2014; Snell-Rood et al., 2018). Central questions in evolutionary ecology are (1) whether plasticity will allow adaptation to new weather conditions, and (2) how plasticity might affect the rate of evolutionary change.
Studying the plasticity of avian growth requires the estimation of how growth patterns can change for a given individual (or a given genotype) across different environments. Such measures are impossible at the individual-level because a bird only grows once. However, in long-lived and philopatric species, associations between growth and weather variation may be likely to result from plastic rather than evolutionary responses because the population genetic background may change little from 1 year to another. In a wild setting, a cross-fostering experiment across a weather gradient (experimental or natural) may reveal if different families have on average different or similar growth patterns across environments (for a discussion on artificial breeding designs, see Roff and Wilson, 2014). To our knowledge, no studies have analyzed cross-fostering in wild birds, with the goal of measuring differences among families in changes in growth patterns across weather gradients.
Because a proportion of the variation in growth traits are determined by parental behavior we might be able to measure the plasticity of nestling growth by considering nestling growth traits as parental traits in species where faithful pairs repeatedly reproduce (Lloyd and Martin, 2004; Wilson et al., 2005). Nestling growth traits that are measured across years for parents can then be considered repeated measures of a parental trait. We can then try to examine the plasticity of the parental care contribution to nestling growth by examining how nestling growth traits vary across environments for a given parent or pair of parents. Importantly, because reproductive investment is a complex function of age and parental condition, such a model will require careful consideration of parental age or breeding experience (McCleery et al., 2008) and the weather conditions parents experience. Consequently, informing such a model will likely require large sample sizes but will provide valuable information on the suite of growth responses individual parents or parental genotypes could generate.
One of the challenges in studying plastic responses in wild populations is determining the environmental variable to which an organism responds (Scheiner, 1993; Gienapp and Brommer, 2014). This problem is further complicated because a trait could be affected by multiple weather variables, interacting weather variables, or even the same weather variables in different ways at different times throughout ontogeny (Figure 3). Recent statistical techniques try to address the difficulty of identifying time windows of weather variation that affect a trait of interest (van de Pol et al., 2016). The van de Pol et al. (2016) method is an exploratory approach that compares multiple windows within a period of time (relative to the biological variable or an absolute time frame) to determine those that best predict (relative to all windows examined) the trait of interest. To understand when during the breeding season weather affects nestling growth, it may be useful to explore weather variation within the prelaying, prenatal, and growth stages (Figure 3). Importantly, we mainly discuss physical growth in this review, but many of the impacts of weather also affect the development (e.g., cognition, thermoregulation, motor function) of nestlings. Understanding how and when during the breeding season development is affected by weather is an additional complexity that will be important to study for a more complete understanding of the impacts of weather.
FIGURE 3
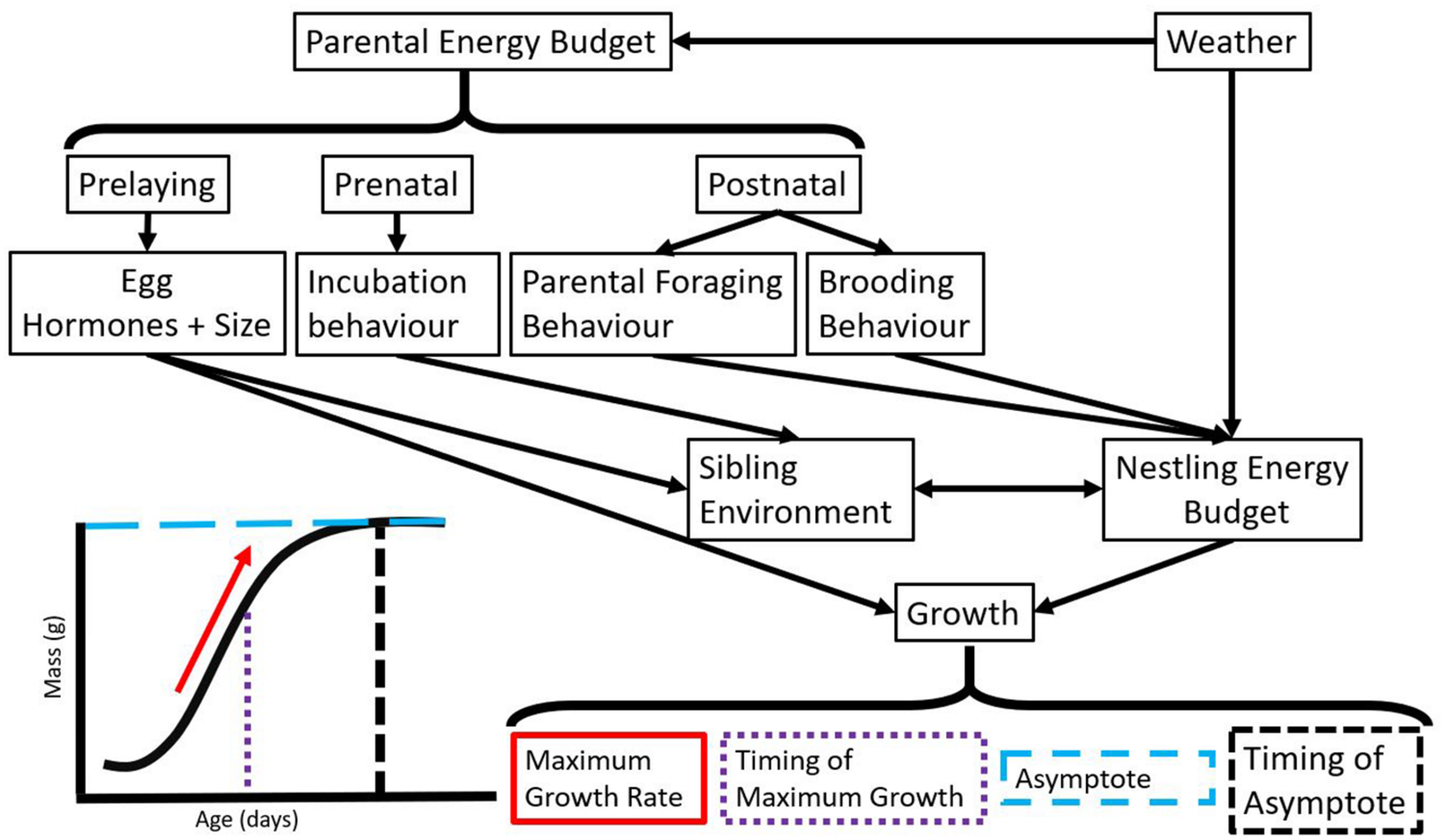
Paths by which weather can impact variation in growth as described by parameters estimated from a general logistic curve. While we display physical growth in this figure, it will be useful to map the development of tissue functions (e.g., cognition, thermoregulation, motor function) onto this growth curve to compare and contrast development and physical growth over the nestling period. Many of the factors that impact growth are also likely to impact development.
In the context of growth and development, identifying environmental correlates may still be challenging because weather variation among prelaying, prenatal, and postnatal periods might be tightly correlated – making it difficult to disentangle during which period weather variation affects a trait. For more easily manipulated weather effects like temperature, experimental work may allow researchers to measure the effects of different time windows by manipulating temperature during specific time periods (Andrew et al., 2017; Andreasson et al., 2018, 2020a; Figure 3). Exploring effects of weather variation outside the breeding season will be valuable if weather patterns are expected to have long-term effects on the resources available or the condition of parents during the breeding season. Understanding the environmental cues that affect the expression of a character is essential because plasticity might allow persistence in the face of changing weather conditions (Vedder et al., 2013), but could also increase the speed of extinction if cues are no longer informative (Reed et al., 2010). Further, in seabirds nestling growth is routinely used as an indicator of environmental conditions (Cairns, 1988). Inferences from such traits require an intimate understanding of the relationship between environmental variation and the measured biological trait (Grémillet and Charmantier, 2010; Brisson-Curadeau et al., 2017).
Nestling Growth and Demography
Weather effects on nestling traits can also affect the demographics of bird populations (Bryant, 1978; Salaberria et al., 2014). In many avian species, growth is likely to affect survival to fledging (nestling survival) and post-fledging survival (juvenile survival). In a review of predictors of juvenile survival, Maness and Anderson (2013) found (in a subset of quality-controlled studies) that nine out of ten studies identify an effect of nestling mass on juvenile survival, but noted that few studies assess whether growth rates affected juvenile survival. Given evidence that growth traits can influence nestling and juvenile survival (Maness and Anderson, 2013), the effects of variation in growth caused by weather patterns might most strongly affect demography in short-lived bird species (Sæther and Bakke, 2000). However, if nestling growth traits determine adult survival or fecundity, weather-induced nestling variation might change the demography of a broad range of avian species. During early life, environmental conditions, like weather, are expected to play a role in shaping adult phenotypes and fitness (Lindström, 1999). A meta-analysis of the effects of early developmental conditions in birds and mammals found that poor early developmental conditions increases the rate of decline in fecundity with age, but developmental conditions do not influence the age-specific decline in survival in adults (Cooper and Kruuk, 2018). However, the latter result relies heavily on mammal studies, since only one avian study in this meta-analysis investigates the effects of the developmental environment on survival senescence (Hammers et al., 2013). Future studies on birds should investigate the effects of early life conditions (including weather) on traits expressed in adults.
Documented Weather Effects on Avian Growth
Understanding current responses to weather enable us to extrapolate responses to predict future changes. Here, we review the effects of weather on growth and discuss the different ontogenic pathways through which weather could impact growth (Figure 3). To cover studies investigating the effect of weather on nestling growth, we searched in the ISI Web of Science on November 2nd, 2020. We used the keywords ‘avian’ OR ‘bird’ + ‘growth’ OR ‘development’ + ‘climate change’ OR ‘global warming’ OR ‘weather’ OR ‘temperature’ OR ‘wind’ OR ‘rainfall’ OR ‘precipitation’ OR ‘sunshine’ OR ‘solar radiation’ OR ‘solar’ OR ‘insolation’ OR ‘experiment’ + ‘ offspring’ OR ‘juvenile’ OR ‘nestling,’ which brought up 989 references. We restricted papers to experimental and observational studies that examined the effect of weather variation on nestling growth, leaving 36 studies. Specifically, we retained studies that examined the effect of weather variables (wind, precipitation, solar radiation, temperature, ice- cover, sea-surface temperature) on nestling phenotypes during the growth period. Weather effects identified from our literature search are included in Tables 1–4.
TABLE 1
| Species | Temperature variable | Association with growth | Study |
| Zebra Finch Taeniopygia guttata | Mean daily maximum during growth | Mean mass (–) and tarsus length (0) | Andrew et al., 2017 |
| European bee-eater Merops apiaster | Mean daily maximum between hatching and measurements | Residual mass (0) | Arbeiter et al., 2016 |
| Lesser kestrel Falco naumanni | Maximum during 2-day period before measurement | Mass gain in adobe cavity nests (–) Mass gain in wooden nest boxes (–) | Catry et al., 2015 |
| European roller Coracias garrulus | Maximum during 2-day period before measurement | Mass gain in adobe cavity nests (0) Mass gain in wooden nest boxes (–) | Catry et al., 2015 |
| Common fiscal Lanius collaris | Daily maximum | Diurnal change in mass at age 6 days (–) | Cunningham et al., 2013 |
| Cliff Swallow Petrochelidon pyrrhonota | Mean daily maximum during growth | Nestling mass at age 10 days (–) | Imlay et al., 2018 |
| Lance-tailed manakin Chiroxiphia lanceolata | Mean daily maximum during breeding season | Linear growth rate (–) | Jones and DuVal, 2019 |
| Superb Fairy wren Malurus cyaneus | Mean maximum before growth | Mass at age 14 days (+, – quadratic) | Kruuk et al., 2015 |
| Superb Fairy wren Malurus cyaneus | Mean maximum during period the end of the previous breeding season | Mass at age 14 days (–) | Kruuk et al., 2015 |
| Lapland longspur Calcarius lapponicus | Mean maximum during growth | Daily growth rate (+) | Pérez et al., 2016 |
| Pied flycatcher Ficedula hypoleuca | Mean and maximum between mass measurements | Age-specific mass at age 7, 10, and 13 days (+) | Siikamäki, 1996 |
| Great tit Parus major | Mean daily during growth | Residual wing length (+) | Eeva et al., 2020 |
| Pied flycatcher Ficedula hypoleuca | Mean daily during growth | Residual wing length (+, – quadratic) | Eeva et al., 2020 |
| Pied flycatcher Ficedula hypoleuca | Mean during growth | Subarctic population residual mass (+) Temperate population residual mass (+) | Eeva et al., 2002 |
| Wryneck Jynx torquilla | Mean hourly during the daytime between measurements | Daily growth of mass and 8th primary from age 14 to 16 days (–) | Geiser et al., 2008 |
| White stork Ciconia ciconia | Mean daily during 7 days of growth | Relative growth rate (+) | Kosicki and Indykiewicz, 2011 |
| Adelaide Rosella Platycercus elegans adelaidae | Mean day and night during growth Night-time standard deviation | Nestling linear growth in mass and bill surface area (+) Nestling linear growth in mass (–) | Larson et al., 2018 |
| Blue tit Cyanistes caeruleus | Mean during 2-day period before measurement | Average 2-day increase in mass (–), head-bill length (–), tarsus length (–), fourth primary length (–) | Mainwaring and Hartley, 2016 |
| Spotless starling Sturnus unicolor | Mean during growth of nestling from first brood Mean during growth of nestling from second brood | Wing length (+) and bill length (+) at age 14 days Mass (–), tarsus length (–), wing length (–), and bill length (–) at age 14 days | Salaberria et al., 2014 |
| Curlew sandpiper Calidris ferruginea* | Mean between growth measurements | Observed/expected growth of mass (+) Observed/expected growth of bill length (+) | Schekkerman et al., 1998 |
| Chaffinch Fingilla coelebs | Mean daily minimum during linear growth | Linear mass (0) and tarsus (0) growth rate | Bradbury et al., 2003 |
| Linnet Carduelis cannabina | Mean daily minimum during linear growth | Linear mass (0) and tarsus (0) growth rate | Bradbury et al., 2003 |
| Skylark Alauda arvensis | Mean daily minimum during linear growth | Linear mass (+, – quadratic) and tarsus (+, – quadratic) growth rate | Bradbury et al., 2003 |
| Yellowhammer Emberiza citrinella | Mean daily minimum during linear growth | Linear mass (0) and tarsus (0) growth rate | Bradbury et al., 2003 |
| Tengmalm’s owl Aegolius fenereus | Mean daily during the breeding season | Nestling duration (0) | Kouba et al., 2015 |
| Golden plover* Pluvialis apricaria | Mean daily minimum 2 days prior to measurement Mean daily minimum over age interval | Residual mass (0) Residual change in mass (+) | Machín et al., 2018 |
Associations between aspects of temperature and nestling growth traits identified from our literature search.
Temperature variables are sorted in the order maximum, mean, and minimum temperature and alphabetically by first author. Columns indicate the species, the weather variable found to be associated with a growth trait, the growth trait and whether the association was positive (+), negative (–) or not significant (0), and the study reference. * Precocial species.
TABLE 2
| Species | Precipitation variable | Association with growth | Study |
| Canada goose Branta canadensis | Cumulative before and after hatch (June/July) | Principle Component 1 of Gosling Size (+) | Brook et al., 2015 |
| Grasshopper Buzzard Butastur rufipennis | Cumulative post-hatch (June/July) | Days to grow from 10 to 90% of final mass (–) Grams per day mass gain from 0 to 10 days (+) | Buij et al., 2013 |
| Tree Swallow Tachycineta bicolor | Cumulative 3 days prior to measurement | Residual mass (–) | Cox et al., 2019 |
| Eurasian bittern Botaurus stellaris | Cumulative between measurements | Daily increase in mass (–) and tarsus length (–) | Kaspryzkowski et al., 2014 |
| Pied flycatcher Ficedula hypoleuca | Cumulative between mass measurements | Age-specific masses at age 7, 10, and 13 days (–) | Siikamäki, 1996 |
| European bee-eater Merops apiaster | Mean daily between hatching and measurement | Residual mass (0) | Arbeiter et al., 2016 |
| Chaffinch Fingilla coelebs | Mean daily during linear growth | Linear mass (0) and tarsus (0) growth rate | Bradbury et al., 2003 |
| Linnet Carduelis cannabina | Mean daily during linear growth | Linear mass (0) and tarsus (0) growth rate | Bradbury et al., 2003 |
| Skylark Alauda arvensis | Mean daily during linear growth | Linear mass (0) and tarsus (0) growth rate | Bradbury et al., 2003 |
| Yellowhammer Emberiza citrinella | Mean daily during linear growth | Linear mass (–) and tarsus growth rate (–) | Bradbury et al., 2003 |
| Wryneck Jynx torquilla | Mean hourly during the day between measurements | Daily growth of mass from age 3–5 and 5–7 days (–) | Geiser et al., 2008 |
| Blue tit Cyanistes caeruleus | Mean daily 5 days prior to measurement | Nestling mass at age 11 days (+) | Grzędzicka, 2019 |
| White stork Ciconia ciconia | Mean daily during 7 days of growth | Relative growth rate (–) | Kosicki and Indykiewicz, 2011 |
| Tengmalm’s owl Aegolius fenereus | Mean daily during the breeding season | Nestling duration (0) | Kouba et al., 2015 |
| Fairy wren Malurus cyaneus | Mean before nestling growth | Mass at age 14 days (+) | Kruuk et al., 2015 |
| Blue tit Cyanistes caeruleus | Mean during 2-day period before measurement | Average 2 days increase in mass (+), head-bill length (0), tarsus length (+), and fourth primary length (+) | Mainwaring and Hartley, 2016 |
| Lapland longspur Calcarius lapponicus | Mean daily during growth | Daily growth rate (–) | Pérez et al., 2016 |
| Gambel’s white crowned sparrow Zonotrichia leucophrys gambelii | Mean daily during growth | Daily growth rate (–) | Pérez et al., 2016 |
| Pied flycatcher Ficedula hypoleuca | Mean daily during growth | Subarctic population residual mass (0) Temperate population residual mass (0) | Eeva et al., 2002 |
| Great tit Parus major | Maximum number of consecutive rainy days during growth | Residual wing length (+) | Eeva et al., 2020 |
| Pied flycatcher Ficedula hypoleuca | Maximum number of consecutive rainy days during growth | Residual wing length (+) | Eeva et al., 2020 |
Associations between aspects of precipitation and nestling growth traits identified from our literature search.
Precipitation variables are sorted in the order of cumulative, mean, and number of days with, precipitation and alphabetically by first author. Columns indicate the species, the weather variable found to be associated with a growth trait, the growth trait and whether the association was positive (+), or negative (–) or not significant (0), and the study reference.
TABLE 3
| Species | Weather Variable | Association with growth | Study |
| Blue tit Cyanistes caeruleus | Mean wind speed during 2-day period before measurement | Average 2 day increase in mass (0), head-bill length (0), tarsus length (–), and fourth primary length (+) | Mainwaring and Hartley, 2016 |
| Eurasian bittern Botaurus stellaris | Mean wind speed between measurements | Daily increase in mass (–) and tarsus length (–) | Kaspryzkowski et al., 2014 |
| Tengmalm’s owl Aegolius fenereus | Mean daily wind speed during breeding season | Nestling duration (0) | Kouba et al., 2015 |
| European bee-eater Merops apiaster | Mean daily windspeed between hatching and measurement | Residual mass (0) | Arbeiter et al., 2016 |
| Mean daily sunshine between hatching and measurement | Residual mass (+) | Arbeiter et al., 2016 | |
| Chaffinch Fingilla coelebs | Mean daily sunshine hours during linear | Linear mass (0) and tarsus (0) growth rate | Bradbury et al., 2003 |
| Linnet Carduelis cannabina | Mean daily sunshine hours during linear | Linear mass (0) and tarsus (0) growth rate | Bradbury et al., 2003 |
| Skylark Alauda arvensis | Mean daily sunshine hours during linear growth | Linear mass (0) and tarsus (–) growth rate | Bradbury et al., 2003 |
| Yellowhammer Emberiza citrinella | Mean daily sunshine hours during linear | Linear mass (0) and tarsus growth rate (0) | Bradbury et al., 2003 |
| Barn Swallow Hirundo rustica | Temperature on day of nestling measurement Mean temperature between hatching and measurement | Nestling mass at age 8–12 days (–) – High wind speed dampened effect – High rainfall increased effect Nestling mass at age 8–12 days (–) – High wind speed dampened effect | Facey et al., 2020 |
| American kestrel Falco sparverius | Principle Component 1 of weather during nestling growth period (associated with increased wind, rainfall, and cold air temperature) | Nestling mass (–) and length of tenth primary (–) at age 20.5–24 days | Dawson and Bortolotti, 2000 |
Associations between combined weather effects, wind speed, sunshine, and nestling growth traits identified from our literature search.
Columns indicate the species, the weather variable found to be associated with a growth trait, the growth trait and whether the association was positive (+), negative (–), or not significant (0), and the study reference.
TABLE 4
| Species | Experimental manipulation | Association with growth | Study |
| Blue Tit Cyanistes caeruleus | Heating of nest during growth | Increase in mass with age (–) Tarsus and wing length at age 14 days (0) | Andreasson et al., 2018 |
| Zebra Finch Taeniopygia guttata | Mass and tarsus length at age 28 days (–) | Andrew et al., 2017 | |
| Tree Swallow Tachycineta bicolor | Growth rate constant of mass (+), length of ninth primary feather (+), and length of tarsus (0) | Dawson et al., 2005 | |
| Great Tit Parus Major | Mass at age 15 days (–) | Rodríguez and Barba, 2016b | |
| Great Tit Parus Major | Cooling of nest during growth | Mass at age 15 days (0) Tarsus length at age 15 days (–) | Rodríguez and Barba, 2016a |
| Great Tit Parus Major | Heating of nest during incubation | Age-specific tarsus length, and mass (0) Mass and tarsus length growth rate (0) | Álvarez and Barba, 2014 |
| Carolina Wrens Thryothorus ludovicianus | Nestling period (–) Mass at age 9 days (–) | Mueller et al., 2019 | |
| European starlings | Heating of nest during incubation | Nestling period (0) Mass at age 3 days (0) Ratio of mass to tarsus cubed at age 10 days (0) | Reid et al., 2000 |
| Prothonotary warbler Protonotaria citrea | Nestling period (–) Mass at age 8 days (+) | Mueller et al., 2019 | |
| Tree Swallows Tachycineta bicolor | Variation in mass at 1 day of age (–) | Ardia et al., 2009 | |
| Heating at origin nest during incubation Heating at nest of rearing during incubation | Residual Mass at age 4 and 7 days (+) Residual Mass at age 10 and 13 days (+) | Pérez et al., 2008 | |
| Cooling at origin nest during incubation Cooling at nest of rearing during incubation Cooling nest temperatures during incubation | Residual (–) and absolute mass (–) at age 4 and 7 days Absolute mass at age 10 days (–) | Ardia et al., 2010 | |
| Blue tits Cyanistes caeruleus | Growth rate (–) Mass at age 14 days (–) | Nilsson et al., 2008 | |
| Mountain blackeye Chlorocharis emiliae | Warming of nest and rain protection during incubation and growth | Nestling period (–), mass growth rate constant (+), wing length growth rate constant (+), tarsus length growth rate constant (+) | Mitchell et al., 2020 |
| Tree Swallow Tachycineta bicolor | Experimental trimming of female ventral feathers | Nestling asymptotic mass (+) | Tapper et al., 2020 |
| Blue tits Cyanistes caeruleus | Experimental trimming of parent’s ventral feathers (Control and experimental broods were both enlarged) | Mass of nestlings at age 14 days (+) – Only for first time breeding females Wing length (+) and tarsus length (0) at age 14 days | Nord and Nilsson, 2019 |
| Experimental trimming of parent’s ventral feathers (Control and experimental broods were both enlarged) | Mass (0), tarsus length (0), and wing length (0) at age 14 days | Andreasson et al., 2020a |
Associations between experimental manipulations of environmental conditions and nestling growth traits.
Columns indicate the species, the weather variable found to be associated with a growth trait, the growth trait and whether the association was positive (+), negative (–), or not significant (0), and the study reference.
Weather Impact of Growth Through Prenatal Effects
In addition to any effects of weather conditions on nestling growth in birds, conditions during embryonic development could affect post-hatching growth. A mother may influence the size of an egg and the hormones in an egg, potentially in response to environmental cues (Love and Williams, 2008; Bentz et al., 2013). The importance of egg size on nestling growth traits is somewhat uncertain. Effects of short-term weather variation on egg size tend to be limited (Nager and Noordwijk, 1992; Christians, 2002; Thomson and Hadfield, 2017; Griffith et al., 2020), and some studies note that egg size affects hatching traits, but this effect deteriorates through ontogeny (Krist, 2011; Williams, 2012).
Weather variation can, but does not always, influence maternal hormones transported into the egg, and these components can affect nestling growth (Schwabl, 1996; Groothuis et al., 2005; Addison et al., 2008; Groothuis and Schwabl, 2008; Ruuskanen et al., 2016). In particular, weather impacts on food may have a strong effect on maternal hormones deposited in the egg and could aid the matching of brood size to environmental conditions (Verboven et al., 2003; Gasparini et al., 2007; Vergauwen et al., 2012; Benowitz-Fredericks et al., 2013; Müller and Groothuis, 2013; Merkling et al., 2016). Other than hormonal influences, weather could also indirectly impact nestling growth through antioxidants, immunoglobins and antimicrobial agents (Williams, 2012), but these effects might be more indirect, as these compounds are less likely to affect growth directly and instead likely help nestlings survive in the face of environmental challenges (e.g., parasites).
If weather patterns during the prenatal period can predict the environment that a nestling will develop and grow in, a mother may be able to provide cues or influence the phenotype of her offspring to better match its future environment. However, the extent to which mothers provide such signals, and whether these signals are adaptive, is mostly untested in the wild. Whether a species has such cues may depend on the predictability of the environment it evolved in, and whether such cues remain adaptive will depend on how the predictability of the environment changes. Parents may even provide auditory cues to their prenatal offspring to signal warm environments resulting, for instance, in lighter 13-day old nestlings in zebra finches Taeniopygia guttata (Mariette and Buchanan, 2016).
Predicting changes in growth caused by climate change may depend on knowing the effects of an embryo’s thermal environment on the nestling phenotype. Weather can directly affect the thermal environment that avian embryos experience and influence parental incubation behavior (DuRant et al., 2010; Griffith et al., 2016). Embryo thermal tolerance varies among species, with species from colder climates tending to have broader tolerances and lower optimums (Webb, 1987). However, even temperature variation within the range tolerated by an embryo can impact a nestling phenotype (Hepp et al., 2006; DuRant et al., 2010; Nord and Nilsson, 2011).
More experiments warming nest microclimates during incubation on a wide range of species might help clarify the impacts of a warmer incubation environment on nestling growth. Nest microclimate experiments are valuable because they reveal the combined effect of indirect thermal effects on the parents and direct effects on embryos (Table 4). Current results from experiments that warm the nest microclimate are variable and different species from the same study location can have contrasting effects on nestling growth (e.g., Mueller et al., 2019; Table 4). Few studies have experimentally cooled nests, but those that do find that it decreases nestling size and growth (Table 4). A cross-fostering experiment in tree swallows provides an interesting insight because nest warming during incubation increases the early age mass of nestlings incubated in a warm nest (raised by parents in a control nest) and the late-age mass of nestlings incubated in a control nest but raised by parents from a warm nest (Pérez et al., 2008). The cross-fostering experiment suggests carry-over effects of warming on parental care during growth and direct benefits of warming on young nestlings.
Post-hatching Weather Effects on Avian Growth
Variation in weather during the growth period is likely to affect chicks both directly and indirectly. Growth can be influenced by changes in thermal environments (e.g., Cunningham et al., 2013; McKinnon et al., 2013; Tables 1, 4), and changes in food availability and parental care (e.g., Keller and Noordwijk, 1994). Scientists have long been interested in the effects of weather on growth and both historical and recent studies provide a baseline for making qualitative and quantitative predictions for changes in growth under different weather conditions (Lack and Lack, 1951; Hawksley, 1957; Tables 1–4). Predicting how climate change will impact avian growth depends on understanding how nestlings are affected by different weather components and how these weather components are expected to change under climate change.
A Warmer Growing Environment Can Positively or Negatively Impact Nestling Growth
Average global temperatures are expected to continue to increase with climate change, and for many regions, the frequency of heatwaves and variability of thermal environments are expected to increase (Intergovernmental Panel on Climate Change, 2018). Increasing temperatures can result in either challenging or improved growing conditions for nestlings and can operate either directly through thermal changes experienced by nestlings or indirectly by affecting parents’ foraging or brooding behavior (Andreasson et al., 2020b; Table 1 and Figure 3). The effects of warming may vary among species, populations, and environments because some populations may be closer to their thermal limits or may be more susceptible to dehydration. For example, species living in environments below their thermal optimum may benefit from increased natural air temperatures during development (e.g., McKinnon et al., 2013), while increased air temperatures or heatwaves may have strong negative impacts on species living close to their thermal limits (Conradie et al., 2019). The effect of thermal changes depends on the ability of nestlings to thermoregulate. If temperatures are beyond a nestling’s thermal neutral zone (the temperature range that body temperature is maintained with little energy expenditure), nestlings will have inefficient physiological processes unless they allocate energy to thermoregulation. Further, nestlings might use limited energy resources on thermoregulation, instead of growth, to prevent body temperatures outside their thermal maximum (e.g., Andreasson et al., 2018).
In many bird species, warmer temperatures are associated with faster growth, heavier asymptotes, heavier age-specific masses, or have no effects at all (Dyrcz, 1974; Hiraldo et al., 1990; McCarty and Winkler, 1999; see Table 1). But, warming temperatures can also correlate with slower growth and smaller nestlings. In a population of blue tits in Lancashire in the United Kingdom, warmer air temperatures correlate with slower increases of tarsus length, head size, feather length, and body mass (Mainwaring and Hartley, 2016), and in nestling Eastern kingbirds Tyrannus tyrannus high temperatures covary with smaller age-specific masses (Murphy, 1985). Similarly, in arid environments, warmer temperatures correlate with smaller nestlings (Cunningham et al., 2013; Wiley and Ridley, 2016; Andrew et al., 2017; Van de Ven, 2017). In arid environments, warming may frequently approach the thermal maximum of many bird species, resulting in smaller nestlings and increases in nestling mortality either from decreases in parental foraging or direct challenges for nestlings (Cunningham et al., 2013; Wiley and Ridley, 2016; Van de Ven, 2017; Conradie et al., 2019). Studies that experimentally heat passerine nestlings tend to find that warmer conditions slow growth and result in smaller nestlings in warm climates (Rodríguez and Barba, 2016b; Andrew et al., 2017; Table 4), but that warmer conditions have a positive or mixed effect on nestling mass and size in cool temperate environments (Dawson et al., 2005; Andreasson et al., 2018; Table 4). In many instances, it will be important to explore specific aspects of temperature (e.g., maximum, minimum, mean) because they may affect growth differently (Table 1). For example, crimson rosella Platycercus elegans nestlings are heavier when the minimum nest temperature is high, but nestlings are lighter when the mean nest temperature is high (Larson et al., 2015). Finally, birds that produce several broods could face different weather conditions for each brood. For example, because temperatures are warmer when spotless starlings Sturnus unicolor produce their second brood, increasing temperatures result in smaller 14-day old nestlings, while the opposite effect is found for first broods (Salaberria et al., 2014). Different temperature regimes during early and late broods suggest nestlings in late broods might be more strongly impacted by warming conditions unless parents can adjust nesting locations (presumably to cooler locations) for their second broods.
Sensitivity to Thermal Variation Could Vary Throughout Nestling Ontogeny
To understand the impacts of temperature variation, we also need to investigate the potentially varying impacts of a thermal challenge during different time points in ontogeny. To predict warming effects on growth patterns it may be important to identify periods of weather within a season that might impact growth and determine how these time periods are predicted to change.
Development of thermoregulation varies among species, but endothermy develops after hatching in birds and earlier during ontogeny in precocial species than in altricial species (Dunn, 1975; Whittow and Tazawa, 1991; Nichelmann and Tzschentke, 2002; Price and Dzialowski, 2018). Because of better thermoregulatory abilities, young precocial nestlings may be better able to survive temperature challenges than young altricial species (Hohtola and Visser, 1998). Within a species, mass is an important determinant of thermoregulatory ability (Visser, 1998), but most studies investigate relationships between size and thermoregulation among species and it is unknown how this relates to variation among nestlings within a species. In the context of a warming environment, smaller birds could be better able to cope with heat stress through faster evaporative cooling (McKechnie and Wolf, 2010), but evaporative cooling causes water loss, leaving small birds vulnerable to dehydration (Whitfield et al., 2015). Consequently, small nestlings may be more susceptible to high temperatures in water-limited environments than large nestlings because of potential dehydration (Whitfield et al., 2015). However, many small birds that live in hot and arid environments will have adaptive strategies, like facultative hyperthermia, that allow tolerance of challenging high temperatures (Gerson et al., 2019; Freeman et al., 2020).
To predict the effects of a warming environment on avian growth, it will be important to know whether hot temperatures during early life can improve tolerance of warmer environments later in life. Evidence from poultry suggests temperature conditioning during incubation and early ages can improve growth performance in warm and cold environments (Nichelmann and Tzschentke, 2002; Shinder et al., 2002; Loyau et al., 2015; Oke et al., 2020; reviewed in Nord and Giroud, 2020). No study has investigated the effects of short-term prenatal exposure to hot or cold temperatures in wild birds, but continuous exposure to low incubation temperatures generally reduces a nestling’s thermal regulatory ability (reviewed in Nord and Giroud, 2020).
How Does Adult Thermoregulation Relate to Offspring Thermoregulation?
According to the ‘climate variability hypothesis,’ high latitude birds may be better able to tolerate temperature variation (Stevens, 1989). This hypothesis suggests that organisms that experience more variable climates should have wider thermal tolerances, and climate variation tends to increase with latitude (Stevens, 1989). A meta-analysis of non-migratory birds supports the climate variability hypothesis – adults of bird species living in variable environments can tolerate a broader range of temperatures (Khaliq et al., 2014). Importantly, migratory bird species do not demonstrate a trend toward increased or decreased tolerance to environmental conditions with latitude. However, when trying to predict the impacts of weather changes on bird species, the variability of the environments where the species evolved may be important. Given this empirical result on adults, we might predict that the offspring of species adapted to more variable environments can tolerate more warming. Note, however, that the relevance of tolerance in adults depends on how tolerance of offspring is related to adult tolerance. If offspring have broader or narrower thermal niches than adults, they may have better or worse resilient than adults to direct effects of temperature challenges. No studies of wild birds have measured thermal tolerance development and compared it to adult birds’ thermal tolerance (Nord and Giroud, 2020).
Weather Impacts Growth Through Changes in Food Availability
Weather variation affects the timing of food abundance, which, depending on when reproduction occurs, can affect the food available for nestling growth (Naef-Daenzer and Keller, 1999; Both et al., 2006; Daunt et al., 2006; Visser et al., 2006). Major components of climate change are shifts in the mean and variability of weather conditions. A consequence of shifting weather conditions is that cues used by birds to time their breeding may become unreliable predictors of food abundance (Bonamour et al., 2019). The mismatch hypothesis suggests that species at lower trophic levels are better able to track shifts in climatic variation than are those at upper trophic levels, resulting in top consumers being less able to time their reproductive events to match food abundance (Stenseth et al., 2002; Thackeray et al., 2016). Because of changing or variable climatic conditions, many birds may have fewer available food resources while offspring develop. For example, both rainfall changes and an inability to temporarily track shifts in prey species can decrease food availability during nestling growth (e.g., black kites Milvus migransHiraldo et al., 1990; Lapland longspurs and white crowned sparrows Pérez et al., 2016). In Baird’s sandpiper Calidris bairdii, increases in asynchrony of hatching date with peak biomass of crane flies (Tipulidae) correlates with a slower maximum growth rate (McKinnon et al., 2012). In thick-billed murres Uria lomvia, earlier ice break-up covaries with decreases in Arctic cod Boreogadus saida and increases in capelin Mallotus villosus in the murre diet. Further, hatching later relative to the ice breakup is associated with smaller 14-day old nestlings (Gaston and Elliott, 2014).
Some avian nestlings demonstrate a remarkable ability to persist under periods of scarce food. Laboratory research and avian husbandry suggest that, in some bird species, nestling growth is incredibly flexible in response to food restriction. Chickens Gallus gallus domesticus, Japanese quail Coturnix japonica, and mallards Anas platyrhynchos raised on restricted diets pause their growth (sometimes for months) then resume typical growth trajectories when food restriction stops, with no evident effect on adult morphology (Jordan, 1953; Singsen et al., 1964; reviewed in Schew and Ricklefs, 1998). Similarly, some seabird species can slow chick growth and delay fledging in years with low food abundance or poor weather conditions (Barrett and Rikardsen, 1992; Weimerskirch et al., 2001; Chiaradia and Nisbet, 2006; Kuepper et al., 2018), and white-fronted bee-eaters Merops bullockoides can pause growth during periods of low food availability (Emlen et al., 1991). Some species can maintain growth early in the nestling stage because of spare yolk (Romanoff, 1944), but nestlings may be more vulnerable to food shortages when the yolk is depleted. For many species extending growth will not be possible because the time window for breeding is restricted (e.g., many Arctic breeding birds). If the timing of the breeding season is constrained, it may be optimal to mature at a smaller size rather than arrest growth until suitable food sources become available.
Life-History Strategy May Determine How Parental Care Changes Under More Difficult Weather Conditions
Weather can also have indirect effects on growth by influencing the foraging or brooding behavior of parents (Taylor, 1983; Machmer and Ydenberg, 1990). The degree to which birds can adjust their parental effort is likely to depend on their life-history, possibly resulting in differing magnitudes of impacts of weather variation in species with high versus low adult survival (Ghalambor and Martin, 2001). For instance, under unpredictable or stressful conditions, long-lived species may invest less in reproductive effort, including parental care, with a stronger impact on nestling growth. All species face trade-offs in their allocation of resources into reproduction and survival (Stearns, 1989). These trade-offs may be particularly apparent in altricial species and in species with nestlings dependent on parents for food or thermoregulation. Parents can cope with fewer resources by increasing foraging effort at the expense of their body condition – helping chicks to survive challenging weather conditions. For example, in tree swallows, increases in rainfall correlate with increases in parental provisioning at the expense of declines in adult body condition (Cox et al., 2019). Alternatively, parents may invest less in reproduction during stressful situations to preserve or maintain their body condition. Yellow-nosed albatrosses Diomedea chlororhynchos appear to increase provisioning in response to poor nestling condition only if food is plentiful, limiting the costs of reproduction for themselves in unfavorable years (Weimerskirch et al., 2001). Future work could determine which strategy is adaptive for different life-histories under unpredictable or extreme food conditions.
Other Components of Weather Can Also Affect Nestling Growth
Climate change is increasing the frequency of heavy precipitation events (Intergovernmental Panel on Climate Change, 2018), and rainfall can have variable effects on avian growth (Table 2). The effects of rainfall on nestling growth may depend on when rainfall occurs relative to a species’ breeding season (Kruuk et al., 2015). During nestling growth, rainfall can decrease growth rates and age-specific morphometrics (Siikamäki, 1996; Kosicki and Indykiewicz, 2011; Kaspryzkowski et al., 2014; Pérez et al., 2016; Cox et al., 2019, but see Mainwaring and Hartley, 2016; Grzędzicka, 2019; Table 2). Decreases in growth can result from nestlings having difficulty thermoregulating because of wet downy feathers or difficult foraging conditions for parents (Nye, 1964; Keller and Noordwijk, 1994). In great tits, days with some rainfall greater than 1 mm result in decreases in daily mass gain by nestlings. The negative association between rainfall and daily mass gain in great tit nestlings is suspected to be caused by decreases in foraging of parents because experimental trapping of adults has negative effects on growth comparable to daily rainfall (Keller and Noordwijk, 1994). In hot and dry regions, prey abundance may increase in rainy conditions and will improve foraging conditions for parents and nestling growth rates (Sicurella et al., 2014).
In some studies rainfall has mixed effects on growth or only impacts nestlings at specific ages. Robinson et al. (2017) categorized 3 years of their study on Arctic peregrine falcons Falco peregrinus tundrius as cool and wet or as warm and dry. In cool and wet years, nestlings grew the fastest, grew to a lower-than-average asymptote, and attain their maximum growth rate at a younger age (Robinson et al., 2017). In little auks Alle alle, rainfall during the nestling period correlates with decreases in the masses of young nestlings, while in red-capped larks Calandrella cinerea monthly rainfall has a positive effect on the mass of older nestlings, but no effect on young nestlings (Konarzewski and Taylor, 1989; Ndithia et al., 2017).
In contrast to rainfall during nestling growth, rainfall outside the breeding season might increase avian growth rates and masses because of the generally beneficial effects of rainfall on vegetation and insect abundance. In a population of superb fairy wrens Malurus cyaneus, increases in rainfall prior to nestling growth result in heavier nestling masses (Kruuk et al., 2015). Higher rainfall during the nestling stage is associated with increases in gosling size in the Canada goose Branta canadensis, but the authors suggest that high rainfall during growth is likely associated with high rainfall prior to the nestling stage (Brook et al., 2015). The authors suggest that disentangling whether rainfall during growth had a negative effect on Canada goose nestling growth might be hard because the observed effect was a combination of earlier rainfall effects on vegetation and direct effects of rainfall on nestling size.
In many regions, global climate change is expected to increase ocean wind speeds but decrease land wind speeds (Torralba et al., 2017). Increases in wind speeds often correlate with lower age-specific nestling masses in seabirds. Common terns Sterna hirundo, for example, display reductions in growth when wind speeds are high (Langham, 1968), maybe because of a reduction in the ability of parents to capture prey (Taylor, 1983). Interestingly, high wind speeds do not appear to affect nestling growth of a relative of common terns, sandwich terns Thalasseus sandvicensis, perhaps because of differences between these two species in foraging behavior or morphology (Langham, 1968; Taylor, 1983). In the little auk, wind speed and decreases in visibility correlate with lower masses in nestlings older than 5 days (Konarzewski and Taylor, 1989). Environmental differences at breeding locations might determine whether a weather variable impacts growth in black-legged kittiwakes Rissa tridactyla, wind speeds do not affect nestlings on a colony in the Gulf of Alaska, but high wind speeds do reduce the growth of nestlings on a colony in the Norwegian Sea (Elliott et al., 2014; Christensen-Dalsgaard et al., 2018). Marine birds may need to change or increase their foraging behavior with changing weather conditions, or offspring will need to adjust to lower or variable food provisioning.
Components of foraging behavior in birds can vary among individuals, and individuals may change their behavior differently in response to weather conditions (Woo et al., 2008; Patrick et al., 2014). Differences among individuals in foraging and how foraging behavior develops may be essential to determine which strategies are adaptive in an altered environment. Beyond effects on foraging, wind may stress (e.g., via noise) nestlings directly, resulting in altered growth phenotypes (Crino et al., 2020). Evidence from blue tits suggests that increases in wind speeds negatively affect the growth of fourth primary feathers, mass and tarsus, so a decline in average wind speeds on land may result in more favorable growing conditions for some land birds (Table 3; but see Sicurella et al., 2014).
Variation in sea-surface temperature is also likely to impact food availability for many seabirds. Average sea-surface temperatures are expected to increase globally, with some regions warming more rapidly than others (Intergovernmental Panel on Climate Change, 2013). Sea-surface temperature changes are likely to affect marine bird species by changing their prey species’ distribution, abundance, and phenology, generally resulting in lower food availability and, in extreme cases, in mass seabird die-offs (Piatt et al., 2020). Associations between warm sea-surface temperatures and slower daily mass gain are found in numerous seabird species (Bertram et al., 1991; Hedd et al., 2002; Gjerdrum et al., 2003; Smithers et al., 2003; Peck et al., 2004; Ancona et al., 2011; but see Pinaud et al., 2005). Additionally, sea-ice coverage, age, and distribution will be important for provisioning in ice-associated seabirds (Gaston and Elliott, 2014; Divoky et al., 2015). Generally, increases in sea-surface temperatures correlate with decreases in nestling masses in seabirds, likely because of low prey availability during breeding.
Lastly, the interaction of different weather effects may change our predictions of future change because interactions among weather variables or other changing variables could enhance or ameliorate negative effects. For example, in a study of barn swallows Hirundo rustica, nestling mass between age 8–12 days is negatively associated with increasing temperatures, and this effect is stronger during heavier rainfall, but weaker during high wind speeds (Facey et al., 2020).
Variation in Weather Conditions May More Strongly Impact Later-Hatching Nestlings
The hatching order of a nestling within a brood might play an important role in how weather interacts with growth. In species with asynchronous hatching, older nestlings can outcompete younger nestlings for food, and younger nestlings will frequently grow more slowly, fledge at a smaller size, or die from starvation or aggression from older nestlings (Mock and Parker, 1998). Siblicide can be facultative or obligate. When siblicide is facultative, aggression and siblicide within a brood may be less frequent when food availability and weather conditions are favorable (e.g., Bortolotti et al., 1991; Reynolds, 1996). Egg-hormones, parental incubation, and parental feeding behavior might all modulate competition among nestlings (Mock and Parker, 1997; Müller and Groothuis, 2013). Weather conditions could act as cues for changes in physiology or behavior that promote or impede competition among nestlings. If climate change generally results in weather conditions that are unfavorable for growth (less food, increased thermal stress), the strongest impacts of changing weather conditions will likely be on later hatching nestlings.
Weather and the Evolution of Avian Growth
Because weather can influence avian growth in many ways, a change in weather will likely result in altered selection on nestling growth traits and influence the evolution of nestling growth traits that are heritable. To predict the evolution of avian growth traits, we need to understand the evolvability of these traits as well as natural selection acting on them. Both these components of evolution, i.e., growth evolvability and natural selection on growth, can change according to environmental variations.
Estimation of the Evolutionary Potential of Growth Is Challenging
Little is known about the heritability of nestling growth traits under different weather conditions, making it difficult to predict how and whether nestling traits will evolve in response to changing climate. Similarly, little is known about how variation in weather affects natural selection on growth traits.
Fewer studies have investigated the heritability of growth parameters such as the asymptote, growth rate, and timing of maximum growth (Figure 1) in the wild. Tarsus length at 13–15 days of age in European starlings is heritable when analyzed using either offspring-midparent regressions (h2 = 0.43 [SE = 0.12]) or full-sibling analyses (h2 = 0.30 [SE = 0.22]) in a partial cross fostering experiment (Smith and Wettermark, 1995). Such high heritability for tarsus length is similar to findings of many other avian quantitative genetic studies (Postma and Brommer, 2014). However, when Smith and Wettermark (1995) used an analysis of variance to estimate the heritability of logistic growth curve components fit to the starling’s masses, they found the asymptote to be heritable (h2 = 0.25 [SE = 0.21]) but not the inflection point or growth constant. Age-specific size and mass measurements also have been found to be heritable in medium ground finches, and great tits (h2 of fledging mass = 0.24 [SE = 0.02]; Garant et al., 2005).
Advances in statistical techniques used in quantitative genetics now allow a more accurate measurement of the evolutionary parameters of nestling growth in wild populations. Mixed models called ‘animal models’ that use between-individual relatedness within a population and can handle uneven sampling designs offer an improvement over offspring-midparent regressions (Kruuk, 2004; Wilson et al., 2010). Use of an animal model in house sparrows shows the nestling age-specific masses and tarsus lengths to be heritable (h2 for mass at 5 days = 0.57 [SE = 0.19], mass at 10 days = 0.86 [SE = 0.13]; tarsus at 5 days = 0.81 [SE = 0.22], tarsus at 10 days = 0.63 [SE = 0.11]; Bonneaud et al., 2009). In a more recent large cross-fostering experiment of blue tits, age-specific masses throughout a nestling’s growth period are found to have low heritabilities using an animal model (h2 ranged from 0.07 [SE = 0.04] at 0 days to 0.09 (SE = 0.03] at 6 days (Hadfield et al., 2013). The authors attributed the low additive genetic variances and heritabilities of nestling age-specific masses to their multivariate statistics and experimental design, where they controlled for the effects of viability selection when estimating (co)variances among nestlings (Hadfield et al., 2013). Analyses accounting for missing data from individuals that do not survive until nestling mass measurement can help avoid biases in estimates of selection and additive genetic variances for nestling traits (see Hadfield, 2008 for a thorough discussion)
In poultry, the heritability of growth parameters (asymptote, the timing of maximum growth, growth rate, age-specific mass/size) can be moderately heritable (e.g., h2 range = 0.15 – 0.66; Grossman and Bohren, 1985; Mignon-Grasteau, 1999; N’Dri et al., 2006; Dana et al., 2011; Haunshi et al., 2012) and selection on domestic species provides evidence that growth traits can evolve under these controlled settings (Marks, 1990 as cited in Steigner et al., 1992; Noordwijk and Marks, 1998; Zuidhof et al., 2014). Heritability of avian growth might be higher in husbandry settings compared to wild populations because of controlled rearing conditions, which reduce the amount of environmental variation. Quantitative genetic studies on avian growth in wild conditions are presently too scarce to make more robust conclusions on the expected levels of heritability, so we encourage more studies to estimate the additive genetic variance of nestling growth parameters in wild avian populations.
Heritability Is Environmentally Dependent
An important note relevant to weather conditions is that heritability measures are only applicable to the population and the environment where they are measured. The denominator in the calculation of heritability, phenotypic variance, is equal to the sum of genetic variance and environmental variance. Hence, an increase in the environmental variance of a trait results in a decrease in the heritability estimate. Additionally, environmental variation can impact the level of additive genetic variation estimated for a given trait (Gebhardt-Henrich and Noordwijk, 1994; Charmantier and Garant, 2005; Wood and Brodie, 2016). Early papers measuring the heritability of growth traits noted that poor environmental conditions during growth might restrict the expression of additive genetic variance, resulting in lower heritability of the fledgling or adult phenotypic traits in unfavorable environments (Gebhardt-Henrich and Noordwijk, 1991, 1994; Gebhardt-Henrich, 1992). The original hypothesis of environmental influence on the heritability of growth by Noordwijk (1982 as cited in Noordwijk and Marks, 1998) suggests that under unfavorable conditions, a nestling might not reach its genetically determined size, but instead would be constrained because of maturation at a set age. However, under favorable conditions a nestling will reach a final size before age-induced maturation. Therefore, the hypothesis suggests that variation in the asymptote under unfavorable conditions might be more likely to reflect environmental differences among nestlings and the variation under favorable conditions might reflect genetic differences among individuals (Noordwijk and Marks, 1998). A change in the genes that underlie the phenotypic expression of a trait could also cause a change in additive genetic variation between environments (Wood and Brodie, 2016). For example, in a hot environment, genes that play a role in heat tolerance might largely determine the additive genetic variance in a growth trait, while in a thermoneutral environment, genes that play a role in metabolism and growth might explain most of the differences among individual growth trajectories. Therefore, weather variation could affect rates of evolution through an increase or decrease in heritability by increasing or decreasing the environmental and/or the additive genetic contribution to variation in nestling growth traits. Lastly, because heritability is dependent on environmental variance, directly reporting the additive genetic variance of a trait scaled by the mean will allow accurate comparisons of evolvability across traits and species (Houle, 1992; Hansen et al., 2011). It is important for researchers to report both metrics when trying to evaluate the general evolvability of any trait.
While we might not know how heritability will change with different weather patterns, we do know that annual changes in the heritability of nestling size traits in response to changes in environmental conditions have been observed. Following a brood size manipulation experiment in great tits, Gebhardt-Henrich and Noordwijk (1991) found that heritability of mass at 15 days of age (near asymptotic mass) might be lower in large broods (average h2 = 0.40 [SE = 0.54]) than in small broods (average h2 = 0.75 [SE = 0.36]), but only in years when environmental conditions are unfavorable. Notably, standard errors are large, and results are not statistically significant, but this study is one of the earliest to postulate an effect of environmental conditions on evolutionary parameters in nestling traits. In a comparison of age-specific size measurements following a brood size manipulation in blue tits, Kunz and Ekman (2000) detected high heritability estimates for age-specific tarsus (age 8 and 10 days), wing (age 6 days), and mass (age 6 and 8 days) measurements in smaller compared to larger broods. While not focused on weather conditions, other more recent studies have noted higher heritability in more favorable environments (Charmantier et al., 2004; Garant et al., 2005).
Experimental studies on poultry provide insight into how weather impacts might influence the evolutionary response of chick growth traits. Marks (1996) examined the response to artificial selection for increased 4-week body mass in restricted versus full nutritional diet lines of Japanese quail (Coturnix japonica) and found that the restricted diet line had much lower phenotypic increases (no changes in phenotype for many generations) in 4-week body mass. The quail chick’s growth conditions are artificial but could indicate that evolutionary changes in nestling size will be slower in environments where weather conditions reduce food availability, as is expected for climate change in many ecosystems.
Generally, these studies indicate that the heritability of fledging mass increases under favorable conditions, but we should cautiously extrapolate from the current literature because the taxonomic range of current results is restricted, and many studies do not investigate the effects of weather. Further, no studies to our knowledge evaluated the changing heritability of growth rates or the timing of maximum growth in wild populations.
Perspectives on Adding Complexity and Interactive Effects
Beyond the genetic variance of a trait, the evolution of a trait also depends on its genetic covariance with other traits (Willham, 1972; Arnold et al., 2008; Walsh and Lynch, 2018). The evolution of growth traits may be constrained because of genetic correlations among traits. For example, genes that increase asymptotic mass may decrease the maximum growth rate. Therefore, weather variation could affect the evolution of growth traits by influencing the genetic covariance among traits. To date, no study has investigated how weather variation shapes genetic covariances among chick growth traits. Hence it is difficult to determine if genetic constraints or the effect of weather on genetic constraints will have an important role in the evolutionary response of avian growth traits to climate change.
Finally, in addition to a given growth trait’s evolutionary trajectory being dependent on genetic correlations, selection on growth traits induced by climate change can interact with other selective forces. For instance, comparative studies have shown that adult and nestling birds are repeatedly smaller in cities than in surrounding rural areas (Bailly et al., 2016; Caizergues et al., 2018). However, as with weather effects, such trends have not yet been attributed to either plastic or evolutionary processes. Several factors that directly influence growth and are affected by climate change (e.g., food availability) are also affected by growing urbanization. Predictive models will hence need at some point to include the complexity of these different and interacting selective forces. Some authors (e.g., Grimm et al., 2008) have hypothesized that since urban areas are ‘heat islands,’ cities provide an interesting opportunity to study how global warming will impact specific traits (Rivkin et al., 2019). Describing growth curves in urban birds could, hence, be informative in understanding how warmer temperatures and changes in resources influence avian growth. Gene flow from populations adapted to urban habitats into populations in more natural habitats could also provide individuals pre-adapted to warmer conditions (e.g., adaptation with gene flow; Tigano and Friesen, 2016).
Conclusion
Human-induced warming has resulted in average global temperatures increasing by 1°C since the preindustrial period, and temperatures are likely to increase by 1.5°C or more in the next 2–3 decades (Intergovernmental Panel on Climate Change, 2018). If the impacts of weather on a species are known, qualitative or quantitative predictions for growth trait changes from weather-growth models can be used to predict future growth trait changes. If the additive genetic variance and fitness associated with differences in growth traits can be estimated, these models could be improved with evolutionary and demographic information (Jenouvrier and Visser, 2011; Vedder et al., 2013).
We have outlined multiple weather components that affect growth traits of avian species. The future of this research will involve trying to predict adaptive responses to these changes. We think the main questions to address next are: (1) what growth trajectories are adaptive in an environment altered by climate change? (2) what is the potential for a given population of birds to adapt to climate change through either plastic or evolutionary adjustments in growth and development? (3) how will environmental changes alter parental effects on growth? and (4) is the potential for adaptation of growth or development altered by the predictability of weather or environmental conditions? To answer these questions, we need more information on the relationship between individual fitness and growth curves and the genetic (co)variance of chick growth traits.
Statements
Author contributions
DS conducted the literature review. DS, VLF, and AC conceived the project idea and wrote the manuscript. All the authors contributed to the article and approved the submitted version.
Funding
We acknowledge grants from the Natural Sciences and Engineering Council of Canada (Grant # 493789-16), a TD Fellowship in Arctic Environmental Issues, and Queen’s University Graduate Awards for funding to conduct this research.
Acknowledgments
We thank Arnaud Grégoire, Céline Teplitsky, the Friesen lab at Queen’s University and the Ecologie Evolutive Empirique, Communication & Coopération team at the CEFE University for helpful discussion. We are also thankful for comments from the two reviewers and editor that helped improve this manuscript.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
1
AddisonB.Benowitz-FredericksZ. M.HipfnerJ. M.KitayskyA. S. (2008). Are yolk androgens adjusted to environmental conditions? A test in two seabirds that lay single-egg clutches.Gen. Comp. Endocrinol.1585–9.
2
AldredgeR. A. (2016). Using non-linear mixed effects models to identify patterns of chick growth in House Sparrows Passer domesticus.Ibis15816–27.
3
ÁlvarezE.BarbaE. (2014). Behavioural responses of great tits to experimental manipulation of nest temperature during incubation.Ornis Fenn.91220–230.
4
AnconaS.Sánchez-ColónS.RodríguezC.DrummondH. (2011). El Niño in the Warm Tropics: local sea temperature predicts breeding parameters and growth of blue-footed boobies.J. Anim. Ecol.80799–808. 10.1111/j.1365-2656.2011.01821.x
5
AndreassonF.HegemannA.NordA.NilssonJ. -Å (2020a). Experimental facilitation of heat loss affects work rate and innate immune function in a breeding passerine bird.J. Exp. Biol.223jeb219790. 10.1242/jeb219790
6
AndreassonF.NilssonJ. -ÅNordA. (2020b). Avian reproduction in a warming world.Front. Ecol. Evol.8:576331. 10.3389/fevo.2020.576331
7
AndreassonF.NordA.NilssonJ. -Å (2018). Experimentally increased nest temperature affects body temperature, growth and apparent survival in blue tit nestlings.J. Avian Biol.491–14. 10.1111/jav.01620
8
AndrewS. C.AwasthyM.GriffithA. D.NakagawaS.GriffithS. C. (2018). Clinal variation in avian body size is better explained by summer maximum temperatures during development than by cold winter temperatures.Auk135206–217. 10.1642/AUK-17-129.1
9
AndrewS. C.HurleyL. L.MarietteM. M.GriffithS. C. (2017). Higher temperatures during development reduce body size in the Zebra Finch in the laboratory and in the wild.J. Evol. Biol.302156–2164. 10.1111/jeb.13181
10
ArbeiterS.SchulzeM.TammP.HahnS. (2016). Strong cascading effect of weather conditions on prey availability and annual breeding performance in European bee-eaters Merops apiaster.J. Ornithol.157155–163. 10.1007/s10336-015-1262-x
11
ArdiaD. R.PérezJ. H.ChadE. K.VossM. A.ClotfelterE. D. (2009). Temperature and life history: experimental heating leads female tree swallows to modulate egg temperature and incubation behaviour.J. Anim. Ecol.784–13.
12
ArdiaD. R.PérezJ. H.ClotfelterE. D. (2010). Experimental cooling during incubation leads to reduced innate immunity and body condition in nestling tree swallows.Proc. R. Soc. B Biol. Sci.2771881–1888. 10.1098/rspb.2009.2138
13
ArnoldS. J.BürgerR.HohenloheP. A.AjieB. C.JonesA. G. (2008). Understanding the evolution and stability of the g-matrix.Evolution622451–2461. 10.1111/j.1558-5646.2008.00472.x
14
BaillyJ.ScheiflerR.BertheS.Clément-DemangeV.-A.LeblondM.PasteurB.et al (2016). From eggs to fledging: negative impact of urban habitat on reproduction in two tit species.J. Ornithol.157377–392. 10.1007/s10336-015-1293-3
15
BarrettR. T.RikardsenF. (1992). Chick growth, fledging periods and adult mass loss of Atlantic Puffins Fratercula arctica during Years of Prolonged Food Stress.Colon. Waterbirds1524–32. 10.2307/1521351
16
Benowitz-FredericksZ. M.KitayskyA. S.WelckerJ.HatchS. A. (2013). Effects of food availability on yolk androgen deposition in the black-legged kittiwake (Rissa tridactyla), a seabird with facultative brood reduction.PLoS One8:e0062949. 10.1371/journal.pone.0062949
17
BentzA. B.NavaraK. J.SieffermanL. (2013). Phenotypic plasticity in response to breeding density in tree swallows: an adaptive maternal effect?Horm. Behav.64729–736. 10.1016/j.yhbeh.2013.08.009
18
BergmannC. (1847). Ueber die verhältnisse der wärmeökonomie der thiere zu ihrer grösse.Göttinger Studien1595–708.
19
BertramD. F.KaiserG. W.YdenbergR. C. (1991). Patterns in the provisioning and growth of nestling Rhinoceros Auklets.Auk108842–852. 10.1093/auk/108.4.842
20
BonamourS.ChevinL.-M.CharmantierA.TeplitskyC. (2019). Phenotypic plasticity in response to climate change: the importance of cue variation.Philos. Trans. R. Soc. B Biol. Sci.37420180178. 10.1098/rstb.2018.0178
21
BonneaudC.SinsheimerJ. S.RichardM.ChastelO.SorciG. (2009). Mhc polymorphisms fail to explain the heritability of phytohaemagglutinin-induced skin swelling in a wild passerine.Biol. Lett.5784–787. 10.1098/rsbl.2009.0435
22
BonnetT.WandelerP.CamenischG.PostmaE. (2017). Bigger is fitter? Quantitative genetic decomposition of selection reveals an adaptive evolutionary decline of body mass in a wild rodent population.PLoS Biol.15:e1002592. 10.1371/journal.pbio.1002592
23
BortolottiG. R.WiebeK. L.IkoW. M. (1991). Cannibalism of nestling American kestrals by their parents and siblings.Can. J. Zool.691447–1453.
24
BothC.BouwhuisS.LessellsC. M.VisserM. E. (2006). Climate change and population declines in a long-distance migratory bird.Nature44181–83. 10.1038/nature04539
25
BradburyR. B.WilsonJ. D.MoorcroftD.MorrisA. J.PerkinsA. J. (2003). Habitat and weather are weak correlates of nestling condition and growth rates of four UK farmland passerines.Ibis145295–306. 10.1046/j.1474-919X.2003.00142.x
26
Brisson-CuradeauE.PattersonA.WhelanS.LazarusT.ElliottK. H. (2017). Tracking Cairns: biologging improves the use of seabirds as sentinels of the sea.Front. Mar. Sci.4:357. 10.3389/fmars.2017.00357
27
BrookR. W.LeafloorJ. O.AbrahamK. F.DouglasD. C. (2015). Density dependence and phenological mismatch: consequences for growth and survival of sub-arctic nesting Canada geese.Avian Conserv. Ecol.101. 10.5751/ACE-00708-100101
28
BryantD. M. (1978). Environmental influences on growth and survival of nestling House Martins Delichon Urbica.Ibis120271–283. 10.1111/j.1474-919X.1978.tb06788.x
29
BuijR.FolkertsmaI.KortekaasK.De IonghH. H.KomdeurJ. (2013). Effects of land-use change and rainfall in Sudano-Sahelian West Africa on the diet and nestling growth rates of an avian predator.Ibis15589–101. 10.1111/ibi.12006
30
BürknerP.-C. (2017). brms: an R package for bayesian multilevel models using stan.J. Stat. Softw.801–28. 10.18637/jss.v080.i01
31
CairnsD. K. (1988). Seabirds as indicators of marine food supplies.Biol. Oceanogr.5261–271. 10.1080/01965581.1987.10749517
32
CaizerguesA. E.GrégoireA.CharmantierA. (2018). Urban versus forest ecotypes are not explained by divergent reproductive selection.Proc. R. Soc. B Biol. Sci.28520180261. 10.1098/rspb.2018.0261
33
CatryI.CatryT.PattoP.FrancoA. M. A.MoreiraF. (2015). Differential heat tolerance in nestlings suggests sympatric species may face different climate change risks.Clim. Res.6613–25. 10.3354/cr01329
34
CharmantierA.GarantD. (2005). Environmental quality and evolutionary potential: lessons from wild populations.Proc. R. Soc. B Biol. Sci.2721415–1425. 10.1098/rspb.2005.3117
35
CharmantierA.GienappP. (2014). Climate change and timing of avian breeding and migration: evolutionary versus plastic changes.Evol. Appl.715–28. 10.1111/eva.12126
36
CharmantierA.KruukL. E. B.LambrechtsM. M. (2004). Parasitism reduces the potential for evolution in a wild bird population.Evolution58203–206. 10.1554/03-350
37
ChevinL. M.CollinsS.LefèvreF. (2013). Phenotypic plasticity and evolutionary demographic responses to climate change: taking theory out to the field.Funct. Ecol.27967–979. 10.1111/j.1365-2435.2012.02043.x
38
ChiaradiaA.NisbetI. C. T. (2006). Plasticity in parental provisioning and chick growth in Little Penguins Eudyptula minor in years of high and low breeding success.Ardea94254–270.
39
Christensen-DalsgaardS.MayR. F.BarrettR. T.LangsetM.SandercockB. K.LorentsenS. (2018). Prevailing weather conditions and diet composition affect chick growth and survival in the black-leggedd kittiwake.Mar. Ecol. Progr. Ser.604237–249. 10.3354/meps12744
40
ChristiansJ. K. (2002). Avian egg size: variation within species and inflexibility within individuals.Biol. Rev. Camb. Philos. Soc.771–26. 10.1017/s1464793101005784
41
ConradieS. R.WoodborneS. M.CunninghamS. J.McKechnieA. E. (2019). Chronic, sublethal effects of high temperatures will cause severe declines in southern African arid-zone birds during the 21st century.Proc. Natl. Acad. Sci. U.S.A.11614065–14070. 10.1073/pnas.1821312116
42
CooperE. B.KruukL. E. B. (2018). Ageing with a silver-spoon: a meta-analysis of the effect of developmental environment on senescence.Evol. Lett.2460–471. 10.1002/evl3.79
43
CoxA. R.RobertsonR. J.LendvaiA. Z.EverittK.BonierF. (2019). Rainy springs linked to poor nestling growth in a declining avian aerial insectivore (Tachycineta bicolor).Proc. R. Soc. B Biol. Sci.28620190018. 10.1098/rspb.2019.0018
44
CrinoO. L.DriscollS. C.BrandlH. B.BuchananK. L.GriffithS. C. (2020). Under the weather: corticosterone levels in wild nestlings are associated with ambient temperature and wind.Gen. Comp. Endocrinol.285113247. 10.1016/j.ygcen.2019.113247
45
CunninghamS. J.MartinR. O.HojemC. L.HockeyP. A. R. (2013). Temperatures in excess of critical thresholds threaten nestling growth and survival in a rapidly-warming arid savanna: a study of common fiscals.PLoS One8:e74613. 10.1371/journal.pone.0074613
46
DanaN.WaaijE. H.vanderArendonkJ. A. M.van. (2011). Genetic and phenotypic parameter estimates for body weights and egg production in Horro chicken of Ethiopia.Trop. Anim. Health Prod.4321–28. 10.1007/s11250-010-9649-4
47
DauntF.AfanasyevV.SilkJ. R. D.WanlessS. (2006). Extrinsic and intrinsic determinants of winter foraging and breeding phenology in a temperate seabird.Behav. Ecol. Sociobiol.59381–388. 10.1007/s00265-005-0061-4
48
DawsonR. D.BortolottiG. R. (2000). Reproductive success of American kestrels: the role of prey abundance and weather.Condor102814–822. 10.1093/condor/102.4.814
49
DawsonR. D.LawrieC. C.O’BrienE. L. (2005). The importance of microclimate variation in determining size, growth and survival of avian offspring: experimental evidence from a cavity nesting passerine.Oecologia144499–507. 10.1007/s00442-005-0075-7
50
DivokyG. J.LukacsP. M.DruckenmillerM. L. (2015). Effects of recent decreases in arctic sea ice on an ice-associated marine bird.Prog. Oceanogr.136151–161. 10.1016/j.pocean.2015.05.010
51
DobsonF. S.BeckerP. H.ArnaudC. M.BouwhuisS.CharmantierA. (2017). Plasticity results in delayed breeding in a long-distant migrant seabird.Ecol. Evol.73100–3109. 10.1002/ece3.2777
52
DunnE. H. (1975). The timing of endothermy in the development of altrical birds.Condor77288–293. 10.2307/1366224
53
DuRantS. E.HeppG. R.MooreI. T.HopkinsB. C.HopkinsW. A. (2010). Slight differences in incubation temperature affect early growth and stress endocrinology of wood duck (Aix sponsa) ducklings.J. Exp. Biol.21345–51. 10.1242/jeb.034488
54
DyrczA. (1974). Factors affecting the growth rate of nestling great reed warblers and reed warblers at Milicz, Poland.Ibis116330–339. 10.1111/j.1474-919X.1974.tb00128.x
55
EevaT.EspínS.Sánchez-VirostaP.RainioM. (2020). Weather effects on breeding parameters of two insectivorous passerines in a polluted area.Sci. Total Environ.729138913. 10.1016/j.scitotenv.2020.138913
56
EevaT.LehikoinenE.RönkäM.LummaaV.CurrieD. (2002). Different responses to cold weather in two pied flycatcher populations.Ecography25705–713. 10.1034/j.1600-0587.2002.250606.x
57
ElliottK. H.ChiversL. S.BesseyL.GastonA. J.HatchS. A.KatoA.et al (2014). Windscapes shape seabird instantaneous energy costs but adult behavior buffers impact on offspring.Mov. Ecol.217. 10.1186/s40462-014-0017-2
58
EmlenS. T.WregeP. H.DemongN. J.HegnerR. E. (1991). Flexible growth rates in nestling White-Fronted Bee-Eaters: a possible adaptation to short-term food shortage.Condor93591–597. 10.2307/1368191
59
FaceyR. J.VafidisJ. O.SmithJ. A.VaughanI. P.ThomasR. J. (2020). Contrasting sensitivity of nestling and fledgling Barn Swallow Hirundo rustica body mass to local weather conditions.Ibis1621163–1174. 10.1111/ibi.12824
60
FalconerD. S. (1952). The problem of environment and selection.Am. Nat.86293–298.
61
FleischerR. C.JohnstonR. F. (1984). The relationship between winter climate and selection on body size of house sparrows.Can. J. Zool.62405–410. 10.1139/z84-063
62
FreemanM. T.CzenzeZ. J.SchoemanK.McKechnieA. E. (2020). Extreme hyperthermia tolerance in the world’s most abundant wild bird.Sci. Rep.1013098. 10.1038/s41598-020-69997-7
63
GarantD.KruukL. E. B.WilkinT. A.McCleeryR. H.SheldonB. C. (2005). Evolution driven by differential dispersal within a wild bird population.Nature43360–65. 10.1038/nature03051
64
GardnerJ. L.PetersA.KearneyM. R.JosephL.HeinsohnR. (2011). Declining body size: a third universal response to warming?Trends Ecol. Evol.26285–291. 10.1016/j.tree.2011.03.005
65
GaspariniJ.BoulinierT.GillV. A.GilD.HatchS. A.RoulinA. (2007). Food availability affects the maternal transfer of androgens and antibodies into eggs of a colonial seabird.J. Evol. Biol.20874–880. 10.1111/j.1420-9101.2007.01315.x
66
GastonA. J.ElliottK. H. (2014). Seabird diet changes in northern Hudson Bay, 1981-2013, reflect the availability of schooling prey.Mar. Ecol. Prog. Ser.513211–223. 10.3354/meps10945
67
GavriletsS.ScheinerS. M. (1993). The genetics of phenotypic plasticity. V. Evolution of reaction norm shape.J. Evol. Biol.631–48. 10.1046/j.1420-9101.1993.6010031.x
68
Gebhardt-HenrichS. G. (1992). Heritability of growth curve parameters and heritability of final size: a simulation study.Growth Dev. Aging5623–33.
69
Gebhardt-HenrichS. G.NoordwijkA. J. V. (1994). The genetical ecology of nestling growth in the Great Tit. Environmental influences on the expression of genetic variances during growth.Funct. Ecol.8469–476. 10.2307/2390071
70
Gebhardt-HenrichS. G.NoordwijkA. J. V. (1991). Nestling growth in the Great Tit I. Heritability estimates under different environmental conditions.J. Evol. Biol.4341–362. 10.1046/j.1420-9101.1991.4030341.x
71
GeiserS.ArlettazR.SchaubM. (2008). Impact of weather variation on feeding behaviour, nestling growth and brood survival in wrynecks Jynx torquilla.J. Ornithol.149597–606. 10.1007/s10336-008-0305-y
72
GersonA. R.McKechnieA. E.SmitB.WhitfieldM. C.SmithE. K.TalbotW. A.et al (2019). The functional significance of facultative hyperthermia varies with body size and phylogeny in birds.Funct. Ecol.33597–607. 10.1111/1365-2435.13274
73
GhalamborC. K.MartinT. E. (2001). Fecundity-survival trade-offs and parental risk-taking in birds.Science292494–497. 10.1126/science.1059379
74
GhalamborC. K.McKayJ. K.CarrollS. P.ReznickD. N. (2007). Adaptive versus non-adaptive phenotypic plasticity and the potential for contemporary adaptation in new environments.Funct. Ecol.21394–407. 10.1111/j.1365-2435.2007.01283.x
75
GienappP.BrommerJ. E. (2014). “Evolutionary dynamics in response to climate change,” in Quantitative Genetics in the Wild, edsCharmantierA.GarantD.KruukL. E. B. (New York, NY: Oxford University Press), 254–273.
76
GillF. B.PrumR. O. (2019). Ornithology, Fourth Edition.New York, NY: W.H. Freeman & Co., 1022–1025.
77
GjerdrumC.ValléeA. M. J.St. ClairC. C.BertramD. F.RyderJ. L.BlackburnG. S. (2003). Tufted puffin reproduction reveals ocean climate variability.Proc. Natl. Acad. Sci. USA1009377–9382. 10.1073/pnas.1133383100
78
GrémilletD.CharmantierA. (2010). Shifts in phenotypic plasticity constrain the value of seabirds as ecological indicators of marine ecosystems.Ecol. Appl. Publ. Ecol. Soc. Am.201498–1503. 10.1890/09-1586.1
79
GriffithS. C.AndrewS. C.McCowanL. S. C.HurleyL. L.Englert DuursmaD.BuchananK. L.et al (2020). Egg size is unrelated to ambient temperature in the opportunistically breeding zebra finch.J. Avian Biol.511–14. 10.1111/jav.02356
80
GriffithS. C.MainwaringM. C.SoratoE.BeckmannC. (2016). High atmospheric temperatures and “ambient incubation” drive embryonic development and lead to earlier hatching in a passerine bird.R. Soc. Open Sci.3150371. 10.1098/rsos.150371
81
GrimmN. B.FaethS. H.GolubiewskiN. E.RedmanC. L.WuJ.BaiX.et al (2008). Global change and the ecology of cities.Science319756–760. 10.1126/science.1150195
82
GroothuisT. G. G.MüllerW.von EngelhardtN.CarereC.EisingC. (2005). Maternal hormones as a tool to adjust offspring phenotype in avian species.Neurosci. Biobehav. Rev.29329–352. 10.1016/j.neubiorev.2004.12.002
83
GroothuisT. G. G.SchwablH. (2008). Hormone-mediated maternal effects in birds: mechanisms matter but what do we know of them?Philos. Trans. R. Soc. Lond. B. Biol. Sci.3631647–1661. 10.1098/rstb.2007.0007
84
GrossmanM.BohrenB. B. (1985). Logistic growth curve of chickens: heritability of parameters.J. Hered.76459–462. 10.1093/oxfordjournals.jhered.a110145
85
GrzędzickaE. (2019). Impact of rainfall on the offspring PHA-response and body mass in the Eurasian blue tit.Ecol. Res.3485–93. 10.1111/1440-1703.1005
86
HadfieldJ. (2019). MCMCglmm: MCMC Generalised Linear Mixed Models. Available online at: https://CRAN.R-project.org/package=MCMCglmm(accessed February 20, 2020).
87
HadfieldJ. D. (2008). Estimating evolutionary parameters when viability selection is operating.Proc. R. Soc. B Biol. Sci.275723–734. 10.1098/rspb.2007.1013
88
HadfieldJ. D. (2010). MCMC methods for multi-response generalized linear mixed models: the MCMCglmm R package.J. Stat. Softw.331–22. 10.1002/ana.22635
89
HadfieldJ. D.HeapE. A.BayerF.MittellE. A.CrouchN. M. A. (2013). Disentangling genetic and prenatal sources of familial resemblance across ontogeny in a wild passerine.Evolution672701–2713. 10.1111/evo.12144
90
HammersM.RichardsonD. S.BurkeT.KomdeurJ. (2013). The impact of reproductive investment and early-life environmental conditions on senescence: support for the disposable soma hypothesis.J. Evol. Biol.261999–2007. 10.1111/jeb.12204
91
HansenT. F.PélabonC.HouleD. (2011). Heritability is not evolvability.Evol. Biol.38258–277. 10.1007/s11692-011-9127-6
92
HarterB. B. (2007). Black Guillemots as Indicators of Change in the Near-Shore Arctic Marine Ecosystem. Ph. D. Thesis, University of Manitoba, Winnipeg, CA.
93
HaunshiS.ShanmugamM.PadhiM. K.NiranjanM.RajkumarU.ReddyM. R.et al (2012). Evaluation of two Indian native chicken breeds for reproduction traits and heritability of juvenile growth traits.Trop. Anim. Health Prod.44969–973. 10.1007/s11250-011-9994-y
94
HawksleyO. (1957). Ecology of a breeding population of arctic terns.Bird Band2857–92. 10.2307/4510623
95
HaywoodS.PerrinsC. M. (1992). Is clutch size in birds affected by environmental conditions during growth?Proc. R. Soc. Lond. B Biol. Sci.249195–197. 10.1098/rspb.1992.0103
96
HeddA.RyderJ. L.CowenL. L.BertramD. F. (2002). Inter-annual variation in the diet, provisioning and growth of Cassin’s auklet at Triangle Island, British Columbia: responses to variation in ocean climate.Mar. Ecol. Prog. Ser.229221–232. 10.3354/meps229221
97
HeppG. R.KennamerR. A.JohnsonM. H. (2006). Maternal effects in Wood Ducks: incubation temperature influences incubation period and neonate phenotype.Funct. Ecol.20308–314. 10.1111/j.1365-2435.2006.01108.x
98
HiraldoF.VeigaJ. P.MáñezM. (1990). Growth of nestling Black Kites Milvus migrans: effects of hatching order, weather and season.J. Zool.222197–214. 10.1111/j.1469-7998.1990.tb05672.x
99
HohtolaE.VisserH. (1998). “Development of locomotion and endothermy in altricial and precocial birds,” in Avian Growth and Development: Evolution within the Altricial-Precocial Spectrum, edsStarckJ. M.RicklefsR. E. (New York, NY: Oxford University Press), 157–170.
100
HouleD. (1992). Comparing evolvability and variability of quantitative traits.Genetics130195–204.
101
HouslayT. M. (2017). Tutorials. Available online at: https://tomhouslay.com/tutorials/(accessed February 20, 2020).
102
ImlayT. L.NickersonD.HornA. G. (2018). Temperature and breeding success for cliff swallows (Petrochelidon pyrrhonota) nesting on man-made structures: ecological traps?Can. J. Zool.97429–435. 10.1139/cjz-2018-0224
103
Intergovernmental Panel on Climate Change (2013). Climate change 2013: The Physical Science Basis.Geneva: IPCC.
104
Intergovernmental Panel on Climate Change (2018). Global warming of 1.5°C.Geneva: IPCC.
105
JenouvrierS.VisserM. E. (2011). Climate change, phenological shifts, eco-evolutionary responses and population viability: towards a unifying predictive approach.Int. J. Biometeorol.55905–919.
106
JohnstonR. F.SelanderR. K. (1964). House sparrows: rapid evolution of races in North America.Science144548–550.
107
JohnstonR. F.SelanderR. K. (1973). Evolution in the House Sparrow. II. Adaptive differentiation in North American populations.Evolution251–28. 10.2307/2406496
108
JonesB. C.DuValE. H. (2019). Direct and indirect effects of the El Niño Southern Oscillation on development and survival of young of a tropical passerine.Oecologia190485–496. 10.1007/s00442-019-04418-9
109
JongG. D. (1990). Quantitative Genetics of reaction norms.J. Evol. Biol.3447–468. 10.1046/j.1420-9101.1990.3050447.x
110
JordanJ. S. (1953). Effects of starvation on wild mallards.J. Wildl. Manag.17304–311. 10.2307/3797112
111
KaspryzkowskiZ.PolakM.ChylareckiP. (2014). Effects of weather conditions, time of breeding, brood size, and hatching order on Eurasian Bittern nestling growth in a food-rich fishpond habitat.Ann. Zool. Fenn.51477–487. 10.5735/086.051.0602
112
KellerL. F.NoordwijkA. J. V. (1993). A method to isolate environmental effects on nestling growth, illustrated with examples from the great tit (Parus major).Funct. Ecol.7493–502. 10.2307/2390037
113
KellerL. F.NoordwijkA. J. V. (1994). Effects of local environmental conditions on nestling growth.Ardea82349–362. 10.5167/uzh-53659
114
KhaliqI.HofC.PrinzingerR.Böhning-GaeseK.PfenningerM. (2014). Global variation in thermal tolerances and vulnerability of endotherms to climate change.Proc. R. Soc. B Biol. Sci.28120141097. 10.1098/rspb.2014.1097
115
KonarzewskiM.TaylorJ. R. E. (1989). The influence of weather conditions on growth of Little Auk Alle alle chicks.Ornis Scand.20112–116. 10.2307/3676877
116
KosickiJ. Z.IndykiewiczP. (2011). Effects of breeding date and weather on nestling development in White Storks Ciconia ciconia.Bird Study58178–185. 10.1080/00063657.2011.554531
117
KoubaM.BartošL.KorpimäkiE.ZárybnickáM. (2015). Factors affecting the duration of nestling period and fledging order in Tengmalm’s Owl (Aegolius funereus): effect of wing length and hatching sequence.PloS ONE10:e0121641. 10.1371/journal.pone.0121641
118
KristM. (2011). Egg size and offspring quality: a meta-analysis in birds.Biol. Rev.86692–716. 10.1111/j.1469-185X.2010.00166.x
119
KruukL. E. B. (2004). Estimating genetic parameters in natural populations using the “animal model.”.Philos. Trans. R. Soc. B Biol. Sci.359873–890. 10.1098/rstb.2003.1437
120
KruukL. E. B.OsmondH. L.CockburnA. (2015). Contrasting effects of climate on juvenile body size in a Southern Hemisphere passerine bird.Glob. Change Biol.212929–2941. 10.1111/gcb.12926
121
KuepperN. D.MarekC.CoriaN.LibertelliM. M.QuillfeldtP. (2018). Facultative hypothermia as a survival strategy during snowstorm induced food shortages in Antarctic storm-petrel chicks.Comp. Biochem. Phys. A22474–83. 10.1016/j.cbpa.2018.06.018
122
KunzEkman (2000). Genetic and environmental components of growth in nestling blue tits (Parus caeruleus).J. Evol. Biol.13199–212. 10.1046/j.1420-9101.2000.00158.x
123
LackE.LackD. (1951). The breeding biology of the Swift Apus Apus.Ibis93501–546. 10.1111/j.1474-919X.1951.tb05457.x
124
LanghamN. P. E. (1968). The Comparative Biology of Terns, Sterna spp. Doctoral dissertation, Durham University, Durham.
125
LarsonE. R.EastwoodJ. R.BuchananK. L.BennettA. T. D.BergM. L. (2015). How does nest-box temperature affect nestling growth rate and breeding success in a parrot?Emu115247–255. 10.1071/MU14081
126
LarsonE. R.EastwoodJ. R.MicallefS.WehbeJ.BennettA. T. D.BergM. L. (2018). Nest microclimate predicts bill growth in the Adelaide rosella (Aves: Psittaculidae).Biol. J. Linn. Soc.124339–349. 10.1093/biolinnean/bly058
127
LindströmJ. (1999). Early development and fitness in birds and mammals.Trends Ecol. Evol.14343–348. 10.1016/S0169-5347(99)01639-0
128
LloydJ. D.MartinT. E. (2004). Nest-site preference and maternal effects on offspring growth.Behav. Ecol.15816–823. 10.1093/beheco/arh085
129
LoveO. P.WilliamsT. D. (2008). Plasticity in the adrenocortical response of a free-living vertebrate: the role of pre- and post-natal developmental stress.Horm. Behav.54496–505. 10.1016/j.yhbeh.2008.01.006
130
LoyauT.BedraniL.BerriC.Métayer-CoustardS.PraudC.CousthamV.et al (2015). Cyclic variations in incubation conditions induce adaptive responses to later heat exposure in chickens: a review.Animal976–85. 10.1017/S1751731114001931
131
MachínP.Fernándex-ElipeJ.KlaassenR. H. G. (2018). The relative importance of food abundance and weather on the growth of a sub-arctic shorebird chick.Behav. Ecol. Sociobiol.7242. 10.1007/s00265-018-2457-y
132
MachmerM. M.YdenbergR. C. (1990). Weather and Osprey foraging energetics.Can. J. Zool.6840–43. 10.1139/z90-007
133
MainwaringM. C.HartleyI. R. (2016). Local weather conditions have complex effects on the growth of blue tit nestlings.J. Therm. Biol.6012–19. 10.1016/j.jtherbio.2016.05.005
134
ManessT. J.AndersonD. J. (2013). Predictors of juvenile survival in birds.Ornithol. Monogr.781–55. 10.1525/om.2013.78.1.1
135
MarietteM. M.BuchananK. L. (2016). Prenatal acoustic communication programs offspring for high posthatching temperatures in a songbird.Science353812–814. 10.1126/science.aaf7049
136
MarksH. L. (1990). “Genetics of growth and meat production in other Galliforms (Japenese Quail, Guinea fowl, and ring-necked pheasants),” in Poultry Breeding and Genetics, ed.CrawfordR. D. (Amsterdam: Elsevier Science Publishers), 677–690.
137
MarksH. L. (1996). Long-term selection for body weight in japanese quail under different environments.Poult. Sci.751198–1203. 10.3382/ps.0751198
138
McCartyJ. P.WinklerD. W. (1999). Relative importance off environmental variables in determining the growth off nestling Tree Swallows Tachycineta bicolor.Ibis141286–296. 10.1111/j.1474-919X.1999.tb07551.x
139
McCleeryR. H.PerrinsC. M.SheldonB. C.CharmantierA. (2008). Age-specific reproduction in a long-lived species: the combined effects of senescence and individual quality.Proc. R. Soc. B Biol. Sci.275963–970. 10.1098/rspb.2007.1418
140
McKechnieA. E.WolfB. O. (2010). Climate change increases the likelihood of catastrophic avian mortality events during extreme heat waves.Biol. Lett.6253–256. 10.1098/rsbl.2009.0702
141
McKinnonL.NolE.JuilletC. (2013). Arctic-nesting birds find physiological relief in the face of trophic constraints.Sci. Rep.31816. 10.1038/srep01816
142
McKinnonL.PicotinM.BolducE.JuilletC.BêtyJ. (2012). Timing of breeding, peak food availability, and effects of mismatch on chick growth in birds nesting in the High Arctic.Can. J. Zool.90961–971. 10.1139/z2012-064
143
MeriläJ.KruukL. E. B.SheldonB. C. (2001). Cryptic evolution in a wild bird population.Nature41276–79. 10.1038/35083580
144
MerklingT.PerrotC.HelfensteinF.FerdyJ.-B.GaillardL.LefolE.et al (2016). Maternal effects as drivers of sibling competition in a parent–offspring conflict context? An experimental test.Ecol. Evol.63699–3710. 10.1002/ece3.1777
145
MerrillL.GrindstaffJ. L. (2018). Early life stress strengthens trait covariance: a plastic response that results in reduced flexibility.Am. Nat.192593–604. 10.1086/699839
146
Mignon-GrasteauS. (1999). Genetic parameters of growth curve parameters in male and female chickens.Br. Poult. Sci.4044–51. 10.1080/00071669987827
147
MillienV.Kathleen LyonsS.OlsonL.SmithF. A.WilsonA. B.Yom-TovY. (2006). Ecotypic variation in the context of global climate change: revisiting the rules.Ecol. Lett.9853–869. 10.1111/j.1461-0248.2006.00928.x
148
MitchellA. E.BoersmaJ.AnthonyA.KitayamaK.MartinT. E. (2020). Experimental amelioration of harsh weather speeds growth and development in a tropical montane songbird.Am. Nat.196E110–E118. 10.1086/710151
149
MockD. W.ParkerG. A. (1997). The Evolution of Sibling Rivalry.Oxford: Oxford University Press.
150
MockD. W.ParkerG. A. (1998). Siblicide, family conflict, and the evolutionary limits of selfishness.Anim. Behav.561–10.
151
MorrisseyM. B.LieftingM. (2016). Variation in reaction norms: statistical considerations and biological interpretation.Evolution.701944–1959. 10.1111/evo.13003
152
MuellerA. J.MillerK. D.BowersE. K. (2019). Nest microclimate during incubation affects posthatching development and parental care in wild birds.Sci. Rep.95161. 10.1038/s41598-019-41690-4
153
MüllerM.GroothuisT. G. (2013). Within-clutch variation in yolk testosterone as an adaptive maternal effect to modulate avian sibling competition; evidence from a comparative study.Am. Nat.181125–136.
154
MurphyM. T. (1985). Nestling eastern kingbird growth: effects of initial size and ambient temperature.Ecology66162–170. 10.2307/1941316
155
Naef-DaenzerB.KellerL. F. (1999). The foraging performance of great and blue tits (Parus major and P. caeruleus) in relation to caterpillar development, and its consequences for nestling growth and fledging weight.J. Anim. Ecol.68708–718. 10.1046/j.1365-2656.1999.00318.x
156
NagerR. G.NoordwijkA. J. V. (1992). Energetic limitation in the egg-laying period of great tits.Proc. R. Soc. B Biol. Sci.249259–263. 10.1098/rspb.1992.0112
157
NdithiaH. K.BakariS. N.MatsonK. D.MuchaiM.TielemanB. I. (2017). Georgraphical and temporal variation in environmental conditions affects nestling growth but not immune function in a year-round breeding equatorial lark.Front. Zool.14:28. 10.1186/s12983-017-0213
158
N’DriA. L.Mignon-GrasteauS.SellierN.Tixier-BoichardM.BeaumontC. (2006). Genetic relationships between feed conversion ratio, growth curve and body composition in slow-growing chickens.Br. Poult. Sci.47273–280. 10.1080/00071660600753664
159
NichelmannM.TzschentkeB. (2002). Ontogeny of thermoregulation in precocial birds.Comp. Biochem. Physiol. A. Mol. Integr. Physiol.131751–763. 10.1016/S1095-6433(02)00013-2
160
NilssonJ. F.StjernmanM.NilssonJ. -Å (2008). Experimental reduction of incubation temperature affects both nestling and adult blue tits Cyanistes caeruleus.J. Avian Biol.39553–559. 10.1111/j.0908-8857.2008.04199.x
161
NoordwijkA. J. V. (1982). Variation in body weight of the great tit, heritability and condition.Verh. Kon. Ned. Akad. Wetensch. Afd. Natuurk. Tweede Reeks799–12.
162
NoordwijkA. J. V.MarksH. L. (1998). “Genetic aspects of growth,” in Avian Growth and Development: Evolution within the Altricial-Precocial Spectrum, edsStarckJ. M.RicklefsR. E. (New York: Oxford University Press), 305–319.
163
NordA.GiroudS. (2020). Lifelong effects of thermal challenges during development in birds and mammals.Front. Physiol11:419. 10.3389/fphys.2020.00419
164
NordA.NilssonJ. -Å (2011). Incubation temperature affects growth and energy metabolism in blue tit nestlings.Am. Nat.178639–651. 10.1086/662172
165
NordA.NilssonJ. -Å (2019). Heat dissipation rate constrains reproductive investment in a wild bird.Funct. Ecol.33250–259.
166
NyeP. (1964). Heat loss in wet ducklings and chicks.Ibis106189–197. 10.1111/j.1474-919X.1964.tb03695.x
167
OkeO. E.AloE. T.OkeF. O.OyebamijiY. A.IjaiyaM. A.OdefemiM. A.et al (2020). Early age thermal manipulation on the performance and physiological response of broiler chickens under hot humid tropical climate.J. Therm. Biol.88102517. 10.1016/j.jtherbio.2020.102517
168
PatrickS. C.BearhopS.GrémilletD.LescroëlA.GrecianW. J.BodeyT. W.et al (2014). Individual differences in searching behaviour and spatial foraging consistency in a central place marine predator.Oikos12333–40. 10.1111/j.1600-0706.2013.00406.x
169
PeckD. R.SmithersB. V.KrockenbergerA. K.CongdonB. C. (2004). Sea surface temperature constrains wedge-tailed shearwater foraging success within breeding seasons.Mar. Ecol. Prog. Ser.281259–266. 10.3354/meps281259
170
PérezJ. H.ArdiaD. R.ChadE. K.ClotfelterE. D. (2008). Experimental heating reveals nest temperature affects nestling condition in tree swallows Tachycineta bicolor.Biol. Lett.4468–471. 10.1098/rsbl.2008.0266
171
PérezJ. H.KrauseJ. S.ChmuraH. E.BowmanS.McGuiganM.AsmusA. L.et al (2016). Nestling growth rates in relation to food abundance and weather in the Arctic.Auk133261–272. 10.1642/AUK-15-111.1
172
PiattJ. F.ParrishJ. K.RennerH. M.SchoenS. K.JonesT. T.ArimitsuM. L.et al (2020). Extreme mortality and reproductive failure of common murres resulting from the northeast Pacific marine heatwave of 2014-2016.PLoS One15:e0226087. 10.1371/journal.pone.0226087
173
PinaudD.CherelY.WeimerskirchH. (2005). Effect of environmental variability on habitat selection, diet, provisioning behaviour and chick growth in yellow-nosed albatrosses.Mar. Ecol. Prog. Ser.298295–304. 10.3354/meps298295
174
PostmaE.BrommerJ. E. (2014). “Four decades of estimating heritabilities in wild vertebrate populations,” in Quantitative Genetics in the Wild, edsCharmantierA.GarantD.KruukL. E. B. (New York, NY: Oxford University Press), 16–33.
175
PriceE. R.DzialowskiE. M. (2018). Development of endothermy in birds: patterns and mechanisms.J. Comp. Physiol.188373–391. 10.1007/s00360-017-1135-0
176
ReedT. E.WaplesR. S.SchindlerD. E.HardJ. J.KinnisonM. T. (2010). Phenotypic plasticity and population viability: the importance of environmental predictability.Proc. R. Soc. B Biol. Sci.2773391–3400. 10.1098/rspb.2010.0771
177
ReidJ. M.MonaghanP.RuxtonG. D. (2000). Resource allocation between reproductive phases: the importance of thermal conditions in determining the cost of incubation.Proc. R. Soc. B Biol. Sci.26737–41. 10.1098/rspb.2000.0963
178
ReynoldsP. S. (1996). Brood reduction and siblicide in Black-billed magpies (Pica pica).Auk113189–199.
179
RicklefsR. E. (1968). Patterns of Growth in Birds.Ibis110419–451. 10.1111/j.1474-919X.1968.tb00058.x
180
RivkinL. R.SantangeloJ. S.AlbertiM.AronsonM. F. J.de KeyzerC. W.DiamondS. E.et al (2019). A roadmap for urban evolutionary ecology.Evol. Appl.12384–398. 10.1111/eva.12734
181
RobinsonB. G.FrankeA.DerocherA. E. (2017). Weather-mediated decline in prey delivery rates causes food-limitation in a top avian predator.J. Avian Biol.48748–758. 10.1111/jav.01130
182
RodríguezS.BarbaE. (2016a). Effects of cool nest microclimates on nestling development: an experimental study with Mediterranean great tits Parus major.Ardeola.63251–260. 10.13157/arla.63.2.2016.ra2
183
RodríguezS.BarbaE. (2016b). Nestling growth is impaired by heat stress: an experimental study in a Mediterranean great tit population.Zool. Stud.55e40. 10.6620/zs.2016.55-40
184
RoffD. A.WilsonA. J. (2014). “Quantifying genotype-by-environment interactions in laboratory systems,” in Genotype–Environment Interactions and Sexual Selection, edsHuntJ.HoskenD. J. (Oxford: John Wiley & Sons), 101–136.
185
RomanoffA. L. (1944). Avian spare yolk and its assimilation.Auk61235–241. 10.2307/4079367
186
RuuskanenS.GroothuisT. G. G.SchaperS. V.DarrasV. M.de VriesB.VisserM. E. (2016). Temperature-induced variation in yolk androgen and thyroid hormone levels in avian eggs.Gen. Comp. Endocrinol.23529–37. 10.1016/j.ygcen.2016.05.026
187
SalaberriaC.CelisP.López-RullI.GilD. (2014). Effects of temperature and nest heat exposure on nestling growth, dehydration and survival in a Mediterranean hole-nesting passerine.Ibis156265–275. 10.1111/ibi.12121
188
SalewskiV.WattC. (2017). Bergmann’s rule: a biophysiological rule examined in birds.Oikos126161–172. 10.1111/oik.03698
189
SætherB.-E.BakkeØ (2000). Avian life history variation and contribution of demographic traits to the population growth rate.Ecology81642–653.
190
ScheinerS. M. (1993). Genetics and evolution of phenotypic plasticity.Annu. Rev. Ecol. Syst.2435–68. 10.1146/annurev.es.24.110193.000343
191
SchekkermanH.RoomenM. W. J.UnderhillL. G. (1998). Growth, behaviour of broods and weather-related variation in breeding productivity of Curlew Sandpipers Calidris Ferruginea.Ardea86153–168.
192
SchewW. A.RicklefsR. E. (1998). “Developmental plasticity,” in Avian Growth and Development: Evolution within the Altricial-Precocial Spectrum, edsStarckJ. M.RicklefsR. E. (New York, NY: Oxford University Press), 288–300.
193
SchwablH. (1996). Maternal testosterone in the avian egg enhances postnatal growth.Comp. Biochem. Physiol. A Physiol.114271–276. 10.1016/0300-9629(96)00009-6
194
ShinderD.LugerD.RusalM.RzepakovskyV.BreslerV.YahavS. (2002). Early age cold conditioning in broiler chickens (Gallus domesticus) : thermotolerance and growth responses.J. Therm. Biol.27517–523. 10.1016/S0306-4565(02)00025-6
195
SicurellaB.CaffiM.CaprioliM.RuboliniD.SainoN.AmbrosiniR. (2014). Weather conditions, brood size and hatching order affect Common Swift Apus apus nestlings’ survival and growth.Bird Study6264–77. 10.1080/00063657.2014.989193
196
SiikamäkiP. (1996). Nestling growth and mortality of Pied Flycatchers Ficedula hypoleuca in relation to weather and breeding effort.Ibis138471–478. 10.1111/j.1474-919X.1996.tb08067.x
197
SingsenE. P.NagelJ.PatrickS. G.MattersonL. D. (1964). The effect of a lysine deficiency on body weight and age at sexual maturity of meat-type pullets.Poult. Sci.43786–787. 10.3382/ps.0430786
198
SmithH. G.WettermarkK. J. (1995). Heritability of nestling growth in cross-fostered European Starlings Sturnus vulgaris.Genetics141657–665.
199
SmithersB. V.PeckD. R.KrockenbergerA. K.CongdonB. C. (2003). Elevated sea-surface temperature, reduced provisioning and reproductive failure of wedge-tailed shearwaters (Puffinus pacificus) in the southern Great Barrier Reef, Australia.Mar. Freshw. Res.54973–977. 10.1071/mf02137
200
Snell-RoodE. C.KobielaM. E.SikkinkK. L.ShephardA. M. (2018). Mechanisms of plastic rescue in novel environments.Annu. Rev. Ecol. Evol. Syst.49331–354. 10.1146/annurev-ecolsys-110617-062622
201
StarckJ. M.RicklefsR. E. (1998). Avian Growth and Development: Evolution within the Altricial-Precocial Spectrum.Oxford: Oxford University Press on Demand.
202
StearnsS. C. (1989). Trade-offs in life-history evolution.Funct. Ecol.3259–268. 10.2307/2389364
203
SteignerJ. W.NestorK. E.LilburnM. S. (1992). Growth and development of lines of Japanese quail, Coturnix coturnix japonica, divergently selected for body weight at four weeks of age.Comp. Biochem. Physiol.102389–393. 10.1016/0300-9629(92)90152-g
204
StensethN. C.MysterudA.OttersenG.HurrellJ. W.ChanK. S.LimaM. (2002). Ecological effects of climate fluctuations.Science2971292–1296. 10.1126/science.1071281
205
StevensG. C. (1989). The latitudinal gradient in geographical range: how so many species coexist in the tropics.Am. Nat.133240–256.
206
TapperS.NoceraJ. J.BurnessG. (2020). Heat dissipation capacity influences reproductive performance in an aerial insectivore.J. Exp. Biol.223jeb222232. 10.1242/jeb.222232
207
TaylorI. R. (1983). Effect of wind on the foraging behaviour of Common and Sandwich Terns.Ornis Scand.1490–96. 10.2307/3676011
208
TeplitskyC.MillienV. (2014). Climate warming and Bergmann’s rule through time: is there any evidence?Evol. Appl.7156–168. 10.1111/eva.12129
209
ThackerayS. J.HenrysP. A.HemmingD.BellJ. R.BothamM. S.BurtheS.et al (2016). Phenological sensitivity to climate across taxa and trophic levels.Nature535241–245. 10.1038/nature18608
210
ThomsonC. E.BayerF.CrouchN.FarrellS.HeapE.MittellE.et al (2017). Selection on parental performance opposes selection for larger body mass in a wild population of blue tits.Evolution71716–732. 10.1111/evo.13169
211
ThomsonC. E.HadfieldJ. D. (2017). Prenatal maternal effects appear to be insensitive to experimental or natural environmental variation.Funct. Ecol.312008–2020. 10.1111/1365-2435.12896
212
TiganoA.FriesenV. L. (2016). Genomics of local adaptation with gene flow.Mol. Ecol.252144–2164. 10.1111/mec.13606
213
TjørveE.TjørveK. M. C. (2010). A unified approach to the Richards-model family for use in growth analyses: why we need only two model forms.J. Theor. Biol.267417–425. 10.1016/j.jtbi.2010.09.008
214
TjørveK. M. C.TjørveE. (2017). Modelling avian growth with the Unified-Richards: as exemplified by wader-chick growth.J. Avian Biol.48770–784. 10.1111/jav.00992
215
TorralbaV.Doblas-ReyesF. J.Gonzalez-ReviriegoN. (2017). Uncertainty in recent near-surface wind speed trends: a global reanalysis intercomparison.Environ. Res. Lett.12114019. 10.1088/1748-9326/aa8a58
216
van de PolM.BaileyL. D.McLeanN.RijsdijkL.LawsonC. R.BrouwerL. (2016). Identifying the best climatic predictors in ecology and evolution.Methods Ecol. Evol.71246–1257. 10.1111/2041-210X.12590
217
Van de VenT. M. F. N. (2017). Implications of Climate Change on the Reproductive Success of the Southern Yellow-billed Hornbill, Tockus leucomelas. Doctoral dissertation, University of Cape Town, Cape Town.
218
van GilsJ. A.LisovskiS.LokT.MeissnerW.OzarowskaA.de FouwJ.et al (2016). Body shrinkage due to Arctic warming reduces red knot fitness in tropical wintering range.Sci. Rep.352819–822. 10.1126/science.aad6351
219
VedderO.BouwhuisS.SheldonB. C. (2013). Quantitative assessment of the importance of phenotypic plasticity in adaptation to climate change in wild bird populations.PLoS Biol.11:e1001605. 10.1371/journal.pbio.1001605
220
VerbovenN.MonaghanP.EvansD. M.SchwablH.EvansN.WhitelawC.et al (2003). Maternal condition, yolk androgens and offspring performance: a supplemental feeding experiment in the lesser black-backed gull (Larus fuscus).Proc. Biol. Sci.2702223–2232. 10.1098/rspb.2003.2496
221
VergauwenJ.GoerlichV.GroothuisT. G. G.EensM.MullerW. (2012). Food conditions affect yolk testosterone deposition but not incubation attendance.Gen. Comp. Endocrinol.176112–119.
222
ViaS.LandeR. (1985). Genotype-environment interaction and the evolution of phenotypic plasticity.Evolution39505–522. 10.1111/j.1558-5646.1985.tb00391.x
223
VisserG. H. (1998). “Development of temperature regulation,” in Avian Growth and Development: Evolution within the Altricial-Precocial Spectrum, edsStarckJ. M.RicklefsR. E. (New York: Oxford University Press), 117–150.
224
VisserM. E.HollemanL. J. M.GienappP. (2006). Shifts in caterpillar biomass phenology due to climate change and its impact on the breeding biology of an insectivorous bird.Oecologia147164–172. 10.1007/s00442-005-0299-6
225
WalshB.LynchM. (2018). Evolution and Selection of Quantitative Traits.Oxford, New York: Oxford University Press.
226
WebbD. R. (1987). Thermal tolerance of avian embryos: a review.Condor89874–888. 10.2307/1368537
227
WeimerskirchH.ZimmermannL.PrinceP. A. (2001). Influence of environmental variability on breeding effort in a long-lived seabird, the Yellow-nosed Albatross.Behav. Ecol.1222–30. 10.1093/oxfordjournals.beheco.a000374
228
WhitfieldM. C.SmitB.McKechnieA. E.WolfB. O. (2015). Avian thermoregulation in the heat: scaling of heat tolerance and evaporative cooling capacity in three southern African arid-zone passerines.J. Exp. Biol.2181705–1714. 10.1242/jeb.121749
229
WhittowG. C.TazawaH. (1991). The early development of thermoregulation in birds.Physiol. Zool.641371–1390.
230
WileyE. M.RidleyA. R. (2016). The effects of temperature on offspring provisioning in a cooperative breeder.Anim. Behav.117187–195. 10.1016/j.anbehav.2016.05.009
231
WillhamR. L. (1972). The role of maternal effects in animal breeding. 3. Biometrical aspects of maternal effects in animals.J. Anim. Sci.351288–1293.
232
WilliamsT. (2012). Physiological Adaptations for Breeding in Birds.Woodstock: Princeton University Press.
233
WilsonA. J.PilkingtonJ. G.PembertonJ. M.ColtmanD. W.OverallA. D. J.ByrneK. A.et al (2005). Selection on mothers and offspring: whose phenotype is it and does it matter?Evolution59451–463. 10.1111/j.0014-3820.2005.tb01003.x
234
WilsonA. J.RéaleD.ClementsM. N.MorrisseyM. M.PostmaE.WallingC. A.et al (2010). An ecologist’s guide to the animal model.J. Anim. Ecol.7913–26. 10.1111/j.1365-2656.2009.01639.x
235
WooK. J.ElliottK. H.DavidsonM.GastonA. J.DavorenG. K. (2008). Individual specialization in diet by a generalist marine predator reflects specialization in foraging behaviour.J. Anim. Ecol.771082–1091. 10.1111/j.1365-2656.2008.01429.x
236
WoodC. W.BrodieE. D. (2016). Evolutionary response when selection and genetic variation covary across environments.Ecol. Lett.191189–1200. 10.1111/ele.12662
237
ZuidhofM. J.SchneiderB. L.CarneyV. L.KorverD. R.RobinsonF. E. (2014). Growth, efficiency, and yield of commercial broilers from 1957, 1978, and 2005.Poult. Sci.932970–2982. 10.3382/ps.2014-04291
Summary
Keywords
environmental change, development, heritability, life-history, maternal effects, natural selection, plasticity
Citation
Sauve D, Friesen VL and Charmantier A (2021) The Effects of Weather on Avian Growth and Implications for Adaptation to Climate Change. Front. Ecol. Evol. 9:569741. doi: 10.3389/fevo.2021.569741
Received
05 June 2020
Accepted
04 January 2021
Published
22 January 2021
Volume
9 - 2021
Edited by
Andreas Nord, Lund University, Sweden
Reviewed by
Simon Charles Griffith, Macquarie University, Australia; Kate Buchanan, Deakin University, Australia
Updates
Copyright
© 2021 Sauve, Friesen and Charmantier.
This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Drew Sauve, 0as69@queensu.ca; sauve.drew@gmail.com
This article was submitted to Behavioral and Evolutionary Ecology, a section of the journal Frontiers in Ecology and Evolution
Disclaimer
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.