 David Lindenmayer
David Lindenmayer Elle Bowd
Elle Bowd Lachlan McBurney
Lachlan McBurney- Threatened Species Recovery Hub, National Environmental Science Program, The Fenner School of Environment and Society, The Australian National University, Canberra, ACT, Australia
Birds are high profile elements of the vertebrate biota in almost all terrestrial ecosystems worldwide. Many studies have uncovered evidence of a decline in bird biodiversity, but temporal patterns of change vary among ecosystems and among bird species with different life history traits. Ecosystem-specific, long-term studies are critical for identifying patterns of temporal change in bird biodiversity and the drivers of that change. Here we present a case study of drivers of temporal change in the bird fauna of the Mountain Ash and Alpine Ash eucalypt forests of south-eastern Australia. Using insights from observational studies and experiments conducted over the past 18 years, we discuss the direct and interactive effects of fire and logging on birds. The extent and severity of wildfires have major negative effects on almost all bird species, and have persisted for more than a decade after the last major conflagration (in 2009). Logging has markedly different effects on birds than those quantified for fire, and may have resulted in elevated levels of site occupancy in remaining uncut areas in the landscape. Both fire and logging have led to marked losses in the extent of old growth forest in Mountain Ash and Alpine Ash ecosystems. This is a concern given the strong association of most species of birds with old forest relative to younger age cohorts. Based on an understanding of the effects of fire and logging as drivers of change, we propose a series of inter-related management actions designed to enhance the conservation of avifauna in Mountain Ash and Alpine Ash ecosystems. A particular focus of management must be on increasing the interval between fires and limiting the spatial extent of wildfires and, in turn, significantly expanding the extent of old growth forest. This is because old growth forest is where most bird species are most likely to occur, and in the event of future wildfires, where fire severity will be lowest. Expansion of the old growth estate will require commercial logging operations to be excluded from large parts of Mountain Ash and Alpine Ash forests.
Introduction
Birds are a high profile component of almost all terrestrial ecosystems worldwide (Gill, 1995). Numerous studies have highlighted the extent to which avifauna are declining in many parts of the world [Sanderson et al., 2006; Ceballos et al., 2017; Intergovernmental Science-policy Platform on Biodiversity and Ecosystem Services (IPBES), 2019]. However, patterns of temporal change are not consistent across regions, nor between groups of birds [e.g., large vs. small-bodied species, common vs. rare species (Inger et al., 2014; Lindenmayer et al., 2018c)]. Detailed long-term ecosystem-level studies are therefore essential for quantifying patterns of temporal change in populations of birds and identifying the drivers of those changes. Indeed, such an approach is critical for developing informed conservation programs that are both ecologically effective and cost effective (Sodhi and Ehrlich, 2010; Wintle et al., 2019; Lindenmayer et al., 2020a).
Here we provide a general synthesis of the change in avifauna over the past 18 years in the iconic wet eucalypt Mountain Ash (Eucalyptus regnans) and Alpine Ash (Eucalyptus delegatensis) forest ecosystems in mainland south-eastern Australia. A particular focus of this article is on temporal responses of birds to the drivers of change in these ecosystems, especially high-severity wildfire, clearcut logging, and their interaction. Climate change is also emerging as both a direct and indirect driver of change in the bird fauna of Mountain Ash and Alpine Ash forest ecosystems, especially through its impacts on altered fire regimes (sensu Keeley, 2009; Boer et al., 2020). Given the challenges facing the conservation of bird biodiversity in these ecosystems, the final part of this paper is dedicated to a discussion of management policies and strategies that should be implemented to enhance the protection of the avifauna in Mountain Ash and Alpine Ash forests.
Study Area and Background Datasets
This case study is focused on the Mountain Ash and Alpine Ash forests of the Central Highlands of Victoria, south-eastern Australia (Figure 1A). Mountain Ash forests are dominated by Mountain Ash (Eucalyptus regnans) trees, the tallest flowering plants on earth, which can exceed 90 m in height (Ashton, 1981b). Alpine Ash (Eucalyptus delegatensis) forests are also spectacular with trees approaching 50–60 m in height (Boland et al., 2006). These forests support a diverse understorey consisting of midstorey Acacia species, broad leaved shrubs including Olearia agrophylla, Bedfordia arborescens, tree-ferns, and a mesic ground layer rich in fern and herb species (Blair et al., 2016; Bowd et al., 2018;Bowd et al., in press).
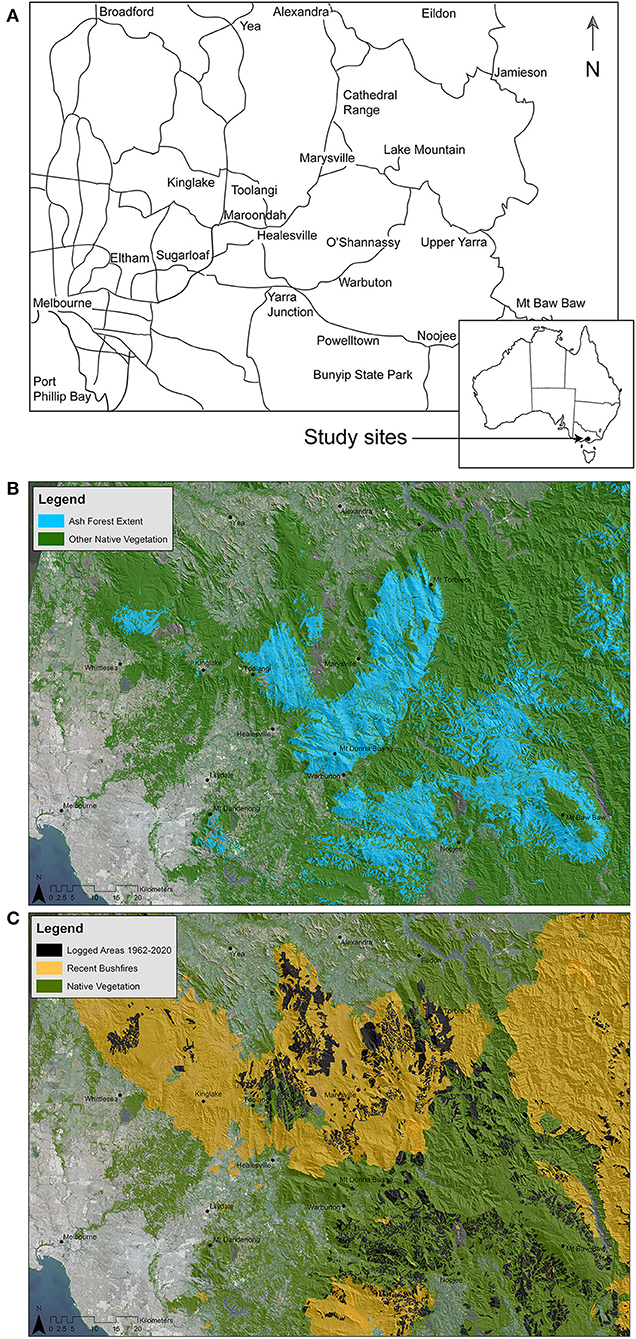
Figure 1. (A) The location of the montane ash (Mountain Ash and Alpine Ash) forests in the Central Highlands of Victoria where a series of long-term studies of birds have been conducted over the past 18 years. (B) The extent of disturbance from wildfire and logging in the Central Highlands of Victoria.
Mountain Ash and Alpine Ash ecosystems support a wide range of bird species (Loyn, 1985) that comprise many functional groups from large nocturnal predators to small diurnal leaf-gleaning species. We have recorded a total of 79 species of birds in repeated field surveys since 2004. The bird assemblages in Mountain Ash and Alpine Ash forests include a number of taxa of conservation concern (Taylor and Lindenmayer, 2019). For example, they are strongholds for species such as the Eastern Yellow Robin (Eopsaltria australis), Flame Robin (Petroica phoenicea), and Crested Shrike-tit (Falcunculus frontatus) which are declining markedly elsewhere in their respective distributions (Montague-Drake et al., 2009; Lindenmayer et al., 2018c). These forests are also important for species such as the Sooty Owl (Tyto tenebricosa) and Masked Owl (Tyto novaehollandiae) which are of conservation concern throughout much of their range in eastern Australia (Debus et al., 2009).
Mountain Ash and Alpine Ash ecosystems sometimes support areas of cool temperate rainforest within, or adjacent to them (Lindenmayer et al., 2000b). However, there are few differences in bird assemblages between ash-type forests and cool temperate rainforest (Lindenmayer et al., 2010) and we do not consider the avifauna of rainforest further in the remainder of this paper.
Fire is the primary form of natural disturbance in Mountain Ash and Alpine Ash forests (Figure 1B). These obligate seeder eucalypts are typically killed by fire, although they require fire to stimulate natural regeneration from canopy stored seed (Smith et al., 2016). While these ecosystems have coevolved with fire, they are vulnerable to decline under short fire return intervals (<every 30 years) that prevent tree species from maturing to an age where they produce viable numbers of seeds (Ashton, 1981a; Bowman et al., 2014; Enright et al., 2015). The natural fire regime in Mountain Ash and Alpine Ash forests is high-intensity and high-severity stand-replacing wildfire that occurs every 75–150 years (McCarthy et al., 1999) (Figure 2A). However, there appears to have been an increase in the frequency of fires (and a reduction in the interval between fires) in the past century (Lindenmayer and Taylor, 2020; Cary et al., in press) with major conflagrations in 1926, 1932, 1939, 1983, and 2009. There have also been less extensive wildfires in 1908, 1918–1919, 1948, and 2019.

Figure 2. Key forms of disturbance in Mountain Ash and Alpine Ash forests: (A) Wildfire. (B) Clearcut logging. (C) Post-fire salvage logging (Photos by David Lindenmayer).
Clearcut logging is the primary form of human disturbance in Mountain Ash and Alpine Ash forests (Figures 1B, 2B) and extensive areas of forest have been harvested in the past 50 years (Taylor and Lindenmayer, 2020). Under such operations, all merchantable trees within stands of approximately 15–40 ha in area are cut with the remaining non-merchantable trees and understorey vegetation then left to dry before being burnt in a high-intensity fire lit to promote the regeneration of a new stand (Flint and Fagg, 2007). In more recent times, there have been attempts to reduce the environmental impacts of logging operations by moving to alternative forms of harvesting to clearcutting, such as by using variable retention harvest systems (sensu Fedrowitz et al., 2014). Under such silvicultural systems, parts of the original stand are retained during logging operations (Lindenmayer et al., 2019a). However, they have remained high-intensity logging operations and continue to resemble conventional clearcuts (see Figure 3).

Figure 3. Logged area subject to variable retention harvesting in Alpine Ash in the Central Highlands of Victoria (Photo by Chris Taylor).
A second form of logging in Mountain Ash and Alpine Ash forests is post-fire “salvage” logging (Noble, 1977; Lindenmayer and Ough, 2006) (Figure 2C). The harvest method for salvage logging is similar to that of clearcutting, except that the sequence of “treatments” is reversed. That is, following a wildfire, all potentially merchantable burnt trees are removed. Sometimes a second (deliberate) burn is applied to remove logging slash (such as tree heads, lateral branches and disturbed understorey vegetation) and, in turn, promote the regeneration of a new stand of young regrowth trees after salvage logging operations have been completed (Lindenmayer et al., 2018d).
Bird Surveys
Mountain Ash and Alpine Ash forests have been targeted for extensive, repeated surveys of birds on an almost annual basis since 2004. The work on birds has entailed both observational studies and true experiments (sensu Cunningham and Lindenmayer, 2016). Details of these investigations are summarized in Table 1. Results from these respective studies have provided the basis for this synthesis article.
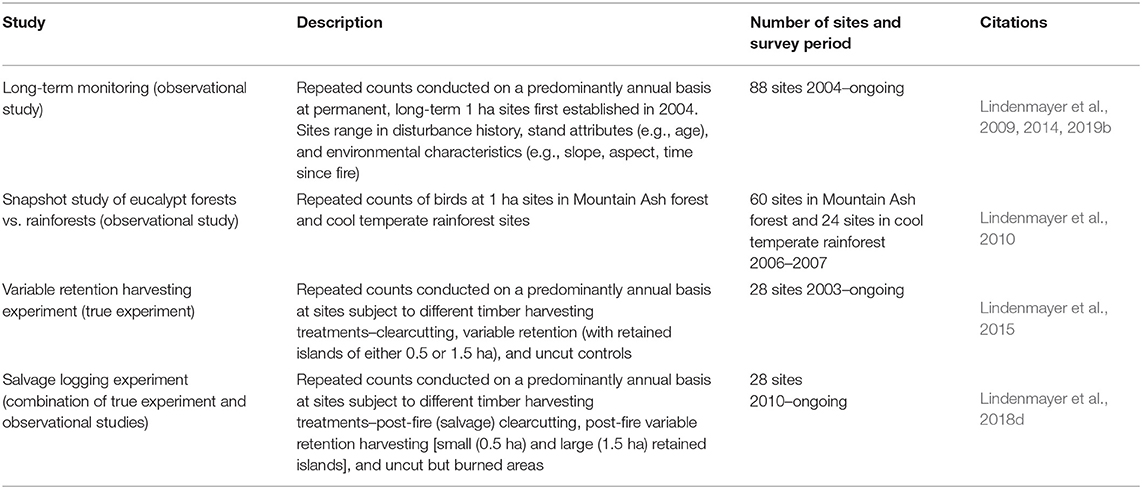
Table 1. Summary of the range of studies of birds conducted in the Mountain Ash and Alpine Ash forests of the Central Highlands of Victoria.
The datasets in the studies outlined in Table 1 have been gathered in broadly the same way, using repeated point interval counts (Pyke and Recher, 1983) along a permanent transect established at a given site. All surveys have been conducted in late November of a given year, which is the time when spring migrants have arrived and are actively calling. Birds are surveyed at least twice by different observers and on different days in an attempt to limit the effects of observer heterogeneity and day effects on datasets (see Cunningham et al., 1999). Broad consistency in bird counting protocols has enabled the integration of datasets to answer particular questions, such as bird responses across a disturbance intensity gradient (Lindenmayer et al., 2018d) and comparisons of bird assemblages between ash-type forests and cool temperate rainforest (Lindenmayer et al., 2010).
Allied with the work on counting birds on long-term sites and experiments, extensive data have also been gathered on the vegetation structure, plant species composition, disturbance history, and stand age at each survey site. Such surveys include measurements of the abundance and condition of large old trees (Lindenmayer et al., 2018a,b) and the abundance and richness of vascular plant species and plant life forms (Blair et al., 2016; Bowd et al., in press). We also have gathered data on spatio-temporal changes in landscape-level forest cover (including the age of the forest) surrounding each site that has resulted from wildfire (at varying levels of severity) and clearcut logging (Lindenmayer et al., 2019b; Taylor and Lindenmayer, 2020). These data have been used to develop sets of covariates for use in analyses of the factors influencing the occurrence of birds (including patterns of temporal change) (e.g., Lindenmayer et al., 2019b). They have also been used to assist in the interpretation of the results of experimental studies (Lindenmayer et al., 2018d).
Temporal Patterns of Change in Site Occupancy
Data gathered from repeated surveys over the past ~20 years indicate that 79 bird species inhabit Mountain Ash and Alpine Ash forests. Of these, approximately half are detected sufficiently frequently (>5% of detections from site x year surveys combined over the duration of studies) to enable detailed statistical analyses. Overall, 24 species have exhibited strong evidence of a decline, with temporal patterns for eight example taxa shown in Figure 4. Examples of other species exhibiting temporal declines (in addition to those in Figure 4) include the Superb Lyrebird (Menura novaehollandiae) and the Pied Currawong (Strepera graculina) (Lindenmayer et al., 2019b). One species, the Flame Robin, exhibited a trend for a strong temporal increase over the duration of our studies.
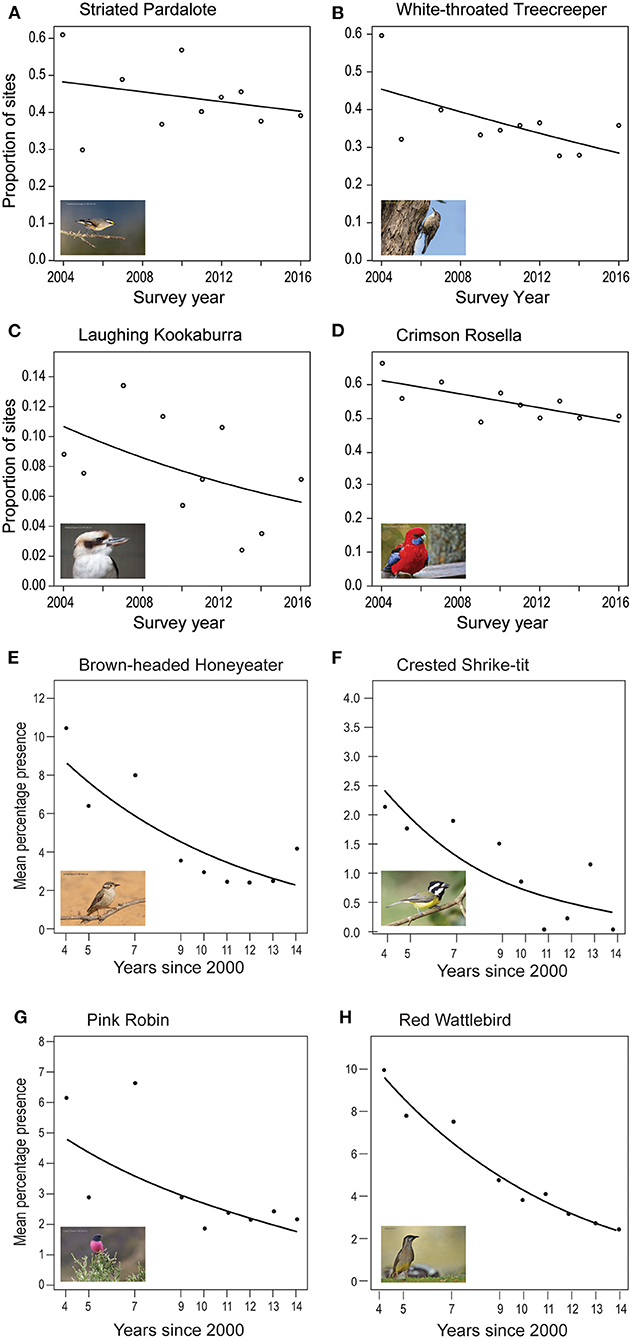
Figure 4. Patterns of temporal change in eight example species of birds in the Mountain Ash forests of the Central Highlands of Victoria. Redrawn from Lindenmayer and Sato (2018) and Lindenmayer et al. (2009). The different measures on the respective axes for the top four vs. bottom four species correspond to differences in the ways data were analyzed in the two different studies.
Many bird species are responding to factors that are changing through time. For example, as outlined in the following section of this article, statistical analyses have revealed complex interactions between the occurrence of bird species, time, and the amount of fire in the landscape (Lindenmayer et al., 2019b). In addition, birds respond to the amount of logging in Mountain Ash and Alpine Ash landscapes (Lindenmayer et al., 2019b), a variable that changes over time with increasing numbers of cutblocks associated with annual harvesting operations. These associations with spatio-temporal disturbance patterns, coupled with data on temporal changes in vegetation and forest age, enable an assessment of the factors influencing temporal changes in bird biodiversity. We present commentary of these drivers in the following section.
Drivers of Change in the Bird Fauna of Mountain Ash and Alpine Ash Forest
Loss of Old Growth Forest
Our analyses of the habitat requirements of birds in Mountain Ash and Alpine Ash forests indicate a strong positive association between the occurrence of most bird species and the presence of old growth forest (Lindenmayer et al., 2019b). That is, almost all bird taxa are more likely to be detected in sites dominated by old growth forest, relative to stands of younger age. Old growth forests appear to be particularly important for small-bodied bird species (Lindenmayer et al., 2019b). Recent studies have documented a significant decline in the amount of old growth Mountain Ash and Alpine Ash forest over the past 25 years (Lindenmayer and Taylor, 2020). Indeed, the extent of old growth Mountain Ash forest is now currently just 1.16% of the ecosystem (Burns et al., 2015). This is 1/30th to 1/60th of what the extent of old growth forests was thought to have been historically (Lindenmayer and McCarthy, 2002). In the case of Alpine Ash forest, 0.47% of the ecosystem is currently old growth, although there are no estimates of historical levels of old growth cover.
As the extent of old growth forest has been declining, populations of key elements of old growth forest such as large old trees have also been in rapid decline (Lindenmayer et al., 2018a). These trees occur both in old growth stands and also as single trees and small clusters of trees embedded within much younger regrowth forest (Lindenmayer et al., 1991). However, rates of loss of such trees are significantly slower in old growth stands than they are in regrowth forest (Lindenmayer et al., 2018a). Losses of individual large old trees are important because they are critical nest sites for a range of species, including iconic birds like the Yellow-tailed Black Cockatoo (Calyptorhynchus funereus) (Nelson and Morris, 1994). Large old living trees also support extensive bark streamers (Lindenmayer et al., 2000a) which can be important foraging substrates for a range of forest birds (Loyn, 1985) such as the Crested Shrike-tit, a species which is declining in many parts of its range (Lindenmayer et al., 2018c), including in Mountain Ash and Alpine Ash forest (see Figure 4). Other forest attributes such as the presence of mistletoe are lost when large old trees are lost from Mountain Ash and Alpine Ash ecosystems. This, in turn, may result in the decline of taxa strongly associated with mistletoe, like the Mistletoebird (Dicaeum hirundinaceum).
Fire
The impacts of wildfire on the bird fauna of Mountain Ash and Alpine Ash forests have been profound. The last major wildfire in the Central Highlands region occurred in 2009, and resulted in depressed levels of bird species richness (Figure 5). Site-level occurrence of the vast majority of species (84%) was negatively associated with the amount of burnt forest in the surrounding landscape (Lindenmayer et al., 2019b). Such declines may be explained by associated habitat losses and limited resources (Whelan, 1995), but also the many individual birds that were likely killed directly by fire (Keith et al., 2002). Just one species, the Flame Robin, has benefitted from the effects of fire and was more likely to occur in landscapes with large amounts of burnt area (Lindenmayer et al., 2019b). Indeed, the Flame Robin is the only early successional specialist that inhabits Mountain Ash and Alpine Ash forests (Lindenmayer et al., 2019b). Notably, 76% of bird species are not exhibiting signs of recovery following the fire in 2009, suggesting that it may require years or even decades for bird biodiversity to return to pre-fire levels. Indeed, data from other obligate seeder vegetation types (Franklin et al., 2002; Gosper et al., 2019a) suggest that a return to pre-fire bird assemblages may take centuries. Nevertheless, some species in Mountain Ash and Alpine Ash forests are responding better to the effects of fire than others. For example, large-bodied birds and migratory taxa are more prevalent in landscapes subject to large amounts of fire relative to species with other life history attributes (Lindenmayer et al., 2019b).
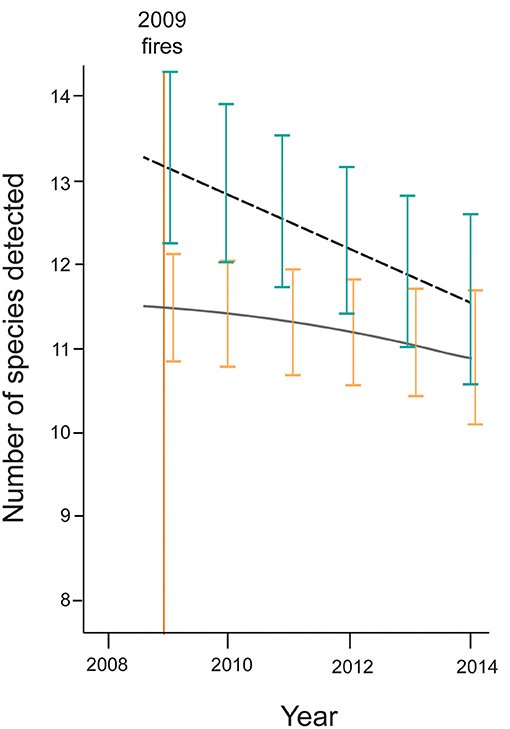
Figure 5. Temporal changes in bird species richness and the amount of fire in 2009 in landscape surrounding long-term field sites in the Mountain Ash and Alpine Ash forests of the Central Highlands of Victoria (modified from Lindenmayer et al., 2019b). The dashed line is for species richness at sites where limited amounts (<5%) of the forest were burnt and the solid line corresponds to sites where ~20% of the landscape surrounding sites was burnt in 2009. The orange vertical line shows the timing of the 2009 wildfires. In many cases, sites surrounded by forest that was burned also were burnt themselves.
An important outcome of studies of fire effects on birds is that current levels of bird species richness are greatest in areas where richness was highest prior to fire (Lindenmayer et al., 2014). We suggest that this effect is occurring because of inherent environmental factors (possibly site productivity) that result in some areas remaining suitable for more bird species, even in the event of major disturbances such as wildfire (Lindenmayer et al., 2014).
Logging
None of our long-term sites have been logged between 2004 and 2019 when surveys were conducted. However, the landscapes surrounding many of our sites have been subject to widespread clearcutting. We have found evidence of relationships between patterns of temporal change for some individual species and the amount of logging in the landscape. Most of these relationships are positive, with birds more likely to be detected in long-term sites with increasing amounts of logging in the surrounding landscape (Lindenmayer et al., 2019b). This is possibly because of a “concentration effect” (sensu Darveau et al., 1995) with animals moving to unlogged areas with increasing amounts of logging in the broader surrounding landscape (Lindenmayer et al., 2019b).
In contrast to work conducted at our long-term sites where timber harvesting is excluded, sites surveyed for birds in the variable retention harvesting experiment and the salvage logging experiment (see Table 1) have been logged. Results from the variable retention harvesting experiment show that logging has negative effects on bird biodiversity (Lindenmayer et al., 2015) (Figure 6). Bird species richness was depressed most markedly in areas subject to clearcutting, relative to unlogged control sites. Clearcutting also significantly altered the composition of bird assemblages with some of the species most negatively affected including the Eastern Spinebill (Acanthorhynchus tenuirostris) and the Gray Fantail (Rhipidura albiscapa) (Lindenmayer et al., 2015). Areas subject to variable retention harvesting supported intermediate levels of bird species richness, relative to clearcutting and unlogged controls (Lindenmayer et al., 2015) (Figure 6). Thus, some of the negative effects of clearcutting can be partially offset by the retention of islands of uncut forest within the boundaries of cutblocks (Lindenmayer et al., 2015).
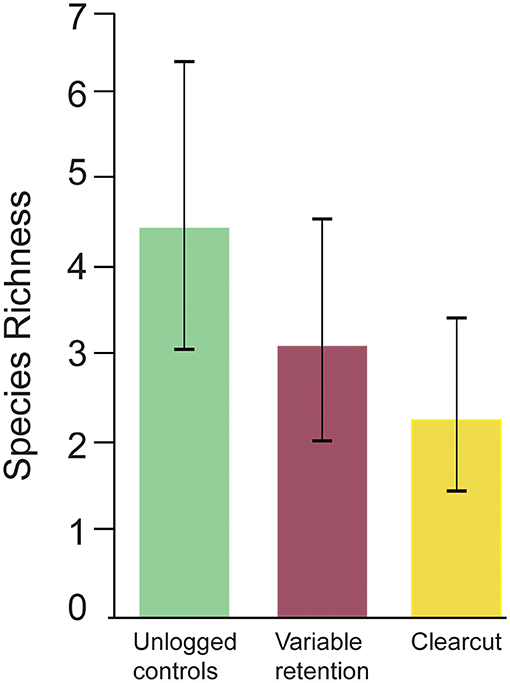
Figure 6. Plot-level mean bird species richness at unlogged control sites, cutblocks subject to variable retention harvesting, and conventional clearcut blocks (redrawn from Lindenmayer et al., 2015).
Logging has other impacts on forest structure that may, in turn, influence patterns of occurrence of birds. For example, clearcutting results in a major reduction in mesic elements of forests such as tree ferns (Blair et al., 2016) and rainforest trees (Lindenmayer et al., 2000a), with some of these key components of stand structure known to be important in the habitat requirements of bird species such as the Eastern Yellow Robin (Eopsaltria australis) (Lindenmayer et al., 2009) and the Pink Robin (Petroica rodinogaster) (Loyn, 1985; Lindenmayer et al., 2010). Logging also can also reduce soil fertility and moisture (Bowd et al., 2019), potentially affecting species that forage extensively in the leaf litter layer such as the Superb Lyrebird (Maisey et al., 2020). Finally, increased logging in the landscape can elevate rates of loss of large old trees in areas that remain uncut areas (Lindenmayer et al., 1997, 2018b) with potentially negative effects on bird species that require access to these trees for nesting sites, for foraging resources, or both. It is likely that changes in the spatial patterns of forest cover as a result of logging lead to greater wind fetch and windspeeds (see Gratkowski, 1956; Miller, 1981) that promote the collapse of living and dead large old trees in uncut areas within wood production landscapes (Lindenmayer et al., 2018b).
Fire and Logging
Post-fire (salvage) logging has been practiced following major conflagrations in Mountain Ash and Alpine Ash forests since the 1940s (Noble, 1977) including after wildfires in 1939, 1983, and 2009 (Lindenmayer et al., 2018d). An experimental study has found more pronounced impacts of salvage logging on bird species than those from conventional (green forest) clearcutting (Lindenmayer et al., 2018d). For example, bird species richness is more severely depressed in areas subject to post-fire salvage logging than in areas subject to wildfire or clearcutting. The impacts of salvage logging on birds can be mitigated to a limited extent by the retention of islands of burnt forest within the perimeter of cutblocks (Lindenmayer et al., 2018d). Nevertheless, negative impacts are more pronounced in salvage logged areas, relative to those of other disturbances in Mountain Ash and Alpine Ash forests, especially wildfire (Lindenmayer et al., 2018d).
Interactions Among Drivers
Salvage logging is an interaction chain (sensu Foster et al., 2016) between fire and subsequent logging that can effect bird biodiversity. However, there are other important interactions between the key drivers of decline in Mountain Ash and Alpine Ash forests. For example, areas that are logged and then regenerated are at elevated risk of burning at higher severity in the event of a subsequent fire (Taylor et al., 2014, 2020). Increased fire severity matters for birds. This is because of established relationships between severity and bird responses (Lindenmayer et al., 2014). High severity fire also results in the rapid loss of large old trees (Lindenmayer et al., 2012), which as outlined above, are important components of habitat for a range of bird species. High severity fire retards the recruitment of new cohorts of old growth, which is detrimental to bird populations given the importance of such age classes of forest for bird biodiversity in Mountain Ash and Alpine Ash forests (Lindenmayer et al., 2019b).
Other stand-age fire severity relationships may have implications for birds. For example, forests that are young when they are burned have fewer biological legacies (sensu Franklin et al., 2000) such as large old trees, relative to when old growth stands are burned (Lindenmayer et al., 2019c). In addition, the regeneration of mesic understorey elements [where birds often nest (Beruldsen, 2003)] can be impaired when young vs. old forests are burned (Bowd et al., in press).
General Discussion
Based on a brief synthesis of key outcomes from several empirical studies over almost two decades, we have presented a case study of the responses of bird fauna to key drivers of change in the Mountain Ash and Alpine Ash forest ecosystems in mainland south-eastern Australia. The work has highlighted the impacts of wildfire, logging, and the interactions between these drivers (e.g., post-fire salvage logging) on bird biodiversity. Such drivers of change are seen in numerous forests globally (Hutto et al., 2016; Sommerfeld et al., 2018) and it is possible that the kinds of responses described in this article may be replicated (at least in part) in some of these ecosystems. The paucity of early successional specialists in Mountain Ash and Alpine Ash forests is consistent with some other obligate seeder vegetation types where such types of species are rare (Gosper et al., 2019b). However, it is in marked contrast to yet other forest types globally where high-intensity and high-severity stand-replacing disturbances are a part of natural fire regimes and early successional species can be prevalent (Burton et al., 2003; Swanson et al., 2011; Hutto et al., 2016).
The empirical studies to date in Mountain Ash and Alpine Ash forests suggest that temporal changes in birds are strongly influenced, not only by the direct effects of key drivers such as fire and logging which can kill animals or trigger concentration effects in undisturbed areas, but also interactions among disturbances which can have compounding effects. These include: (a) wildfire reducing the amount of old growth forest (Lindenmayer and Taylor, 2020), (b) wildfire triggering salvage logging, (c) wildfire and logging driving losses in key biological legacies such as large old trees (Lindenmayer et al., 2012, 2018b), and (d) logging resulting in subsequently elevated fire severity (Taylor et al., 2014). These direct and interacting effects of drivers of change are summarized in Figure 7. An understanding of such relationships, in turn, highlights. Some of the strategies that might be employed to best tackle drivers of change as part of efforts to better conserve forests and the avifauna that it supports (see Table 2).
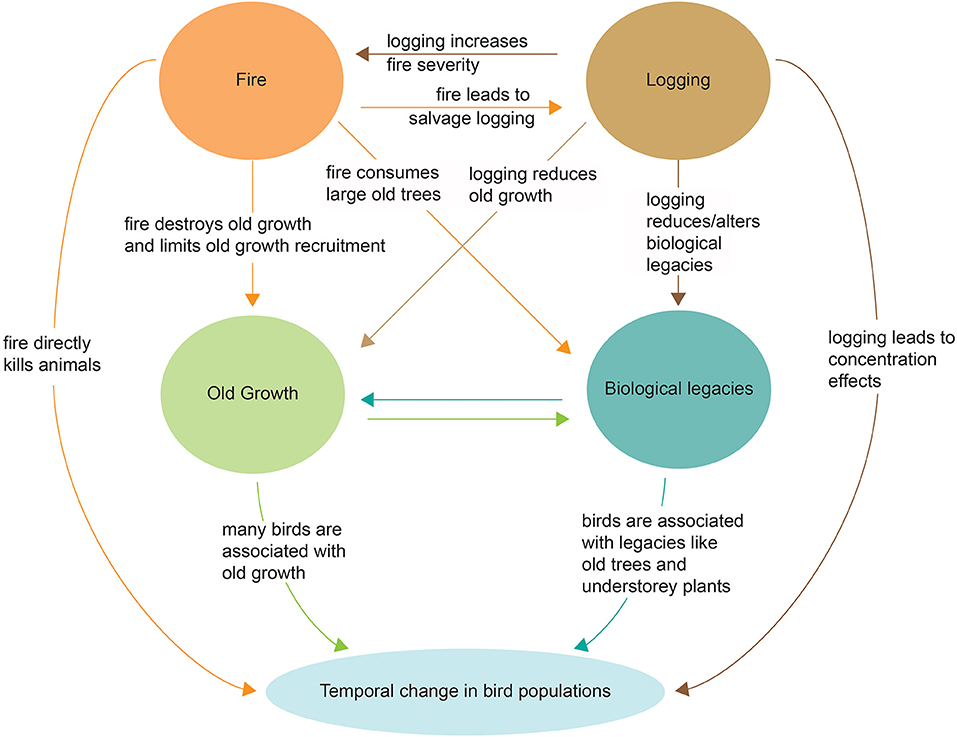
Figure 7. Direct and interacting drivers of temporal change in bird fauna in Mountain Ash and Alpine Ash forests in south-eastern mainland Australia.

Table 2. Summary of possible management actions to promote the conservation of birds in the Mountain Ash and Alpine Ash forests of the Central Highlands of Victoria.
Strategies for Mitigating the Drivers of Temporal Change in Birds
Fire is a key driver of change in Mountain Ash and Alpine Ash ecosystems. Extensive fires have had direct effects on birds [presumably by killing them (Keith et al., 2002; Shine, 2020)], but also indirectly by reducing areas of old growth forest (Lindenmayer and Taylor, 2020), elevating the rate of decline of large old trees (Lindenmayer et al., 2012), and altering the structure and composition of the understorey (Blair et al., 2016; Bowd et al., 2018) (Figure 7). Reducing the extent of wildfire in Australian forests, including those dominated by Mountain Ash and Alpine Ash, is a major challenge. This is, in part, because extreme wildfire is driven by climate and weather (Boer et al., 2020; Jones et al., 2020) and addressing the effects of climate change will require global action. Practices like hazard reduction burning are not a management option to reduce the severity of wildfire in Mountain Ash and Alpine Ash ecosystems. This is because, as obligate-seeding species, Mountain Ash and Alpine Ash are typically killed by fire which would not satisfy the objective of using prescribed fire to reduce fire severity. Indeed, relative to old growth forests, young regrowth stands are highly flammable and at risk of elevated fire severity (Taylor et al., 2014; Zylstra, 2018). Moreover, the environments occupied by Mountain Ash and Alpine Ash are either too wet to enable low-severity burns to be applied or when fires are actually ignited then such forests are at risk of suffering high severity crown burns. In addition, repeated fires at short intervals can eliminate Mountain Ash and Alpine Ash ecosystems altogether (Bowman et al., 2014; Enright et al., 2015). This is because trees fail to reach maturity before they are killed, leading to them being replaced by other ecosystems such as those dominated by Acacia woodland (Lindenmayer et al., 2011). However, there may be a role for hazard reduction burning in drier forest types to occur in areas adjacent to Mountain Ash and Alpine Ash ecosystems, although recurrent fire in these environments can trigger resprout failure (Fairman et al., 2019) and hence elevated tree death. There are also risks associated with a loss of control of prescribed burning operations and them becoming wildfires.
Efforts to reduce the risks of fire in Mountain Ash and Alpine Ash ecosystems will require expanding the currently very limited areas of old growth forest (as this is where fire severity is lowest) (Lindenmayer and Taylor, 2020). This will take considerable time as old growth stands require a prolonged period to develop the key attributes that define them (Lindenmayer et al., 2000a)—in the order of 40–100 years at least. Notably, old growth stands are currently not subject to logging in Victoria, although relatively recent changes in definitions of old growth have made it harder for areas of advanced growth forest to actually be classified as old growth. This is because the age criterion for old growth has been lifted from 120 to 250 years old (Lindenmayer and Taylor, 2020). Given the current scarcity of old growth Mountain Ash and Alpine Ash, coupled with its critical importance for the vast majority of bird species (Lindenmayer et al., 2019b), we strongly suggest that the Victoria Government must revert to the previous definition of old growth (of any stands >120 years old) and ensure that they remain unlogged. An advantage of growing more old growth forest is that fire severity is lowest in these forests (Taylor et al., 2014, 2020) and hence this may reduce the risk of widespread wildfires in the landscape (Lindenmayer et al., 2011). The reasons why fire severity is reduced in old growth forest remain unclear. However, they may be related to: (1) the greater abundance of mesic elements such as rainforest trees and tree ferns in older forests (Lindenmayer et al., 2000a) (Blair et al., 2016), (2) high levels of soil moisture in long undisturbed forests (Bowd et al., 2019), and (3) reduced ladder fuels (that characterize flammable young regenerating forests) (Zylstra et al., 2016). Notably, other forest types around the world, including those dominated by obligate seeding species and where wildfires are typically stand replacing events, are characterized by lower levels of flammability in old growth stands (e.g., Zald and Dunn, 2017; Tiribelli et al., 2018).
Beyond greater protection of stands of old growth forest, there also will be a need to protect some areas of younger forest that have the potential of growing through to become old growth (in the absence of further disturbance). For example, the oldest forests in the Central Highlands region (other than the limited areas of old growth forest), are stands that regenerated following wildfires in 1939. These 80+ year stands are the next nearest old growth and we argue that they should be exempt from logging as part of an old growth restoration strategy in Mountain Ash and Alpine Ash forests. Notably, population viability analyses have suggested that greater levels of protection to eventually expand the extent of the old growth estate will be critical for the long-term conservation of birds of conservation concern such as the Sooty Owl in the Central Highlands region (Taylor et al., 2017).
In the event of future fires, we suggest that the forest policies that result in salvage logging should be abandoned. This is because of the highly detrimental impacts that these forestry operations have on bird biodiversity both directly, and indirectly through impacts on other forest elements including the composition and structure of vegetation communities (Blair et al., 2016; Lindenmayer et al., 2018d). Areas of old growth forest that have been burnt should be a particular target for conservation (and hence the exclusion of salvage logging). This is because the key biological legacies in such areas (like large old fire-scarred trees and dead trees) can be critical habitat elements for post-fire biotic recovery (Lindenmayer et al., 2019c). This may be one of the reasons explaining why sites with high values for birds prior to wildfires remain relatively species rich for birds after fire in Mountain Ash and Alpine Ash forests (Lindenmayer et al., 2014). Therefore, areas of burnt forest should not automatically be placed into areas available for logging under planned harvesting schedules.
Given that old growth Mountain Ash and Alpine Ash forests are rare, and it will take a prolonged period to recruit new cohorts of such age classes, remaining large old trees in regrowth forests are likely to be important for some elements of bird biodiversity. However, they are at risk of collapse when surrounding areas are logged (Lindenmayer et al., 2016, 2018b). On this basis, we suggest a key management strategy should be to promote the protection of these individual large old trees with buffers of unlogged forest to reduce their susceptibility to edge effects such as windthrow.
Caveats and Limitations
The collective set of studies in Mountain Ash and Alpine Ash has focused largely on birds for which sufficient data can be gathered from standardized, routine, repeated surveys to facilitate subsequent statistical analyses. However, some key species of conservation concern like large forest owls (e.g., the Sooty Owl) are not detected by conventional diurnal counting protocols; fit-for-purpose studies are needed to understand both temporal changes in populations and the drivers underpinning those changes (e.g., Milledge et al., 1991). We do note that species such as the Sooty Owl and other large forest owls such as the Masked Owl are sometimes detected in night-time counts of arboreal marsupials and annual records of them are now far less prevalent than 20 years ago (Lindenmayer et al. unpublished data). This is possibly as a result of the steep decline in prey items such as possums and gliders in Mountain Ash and Alpine Ash forests during this time (Lindenmayer et al., 2020b).
The focus of this paper has been on key drivers of change in Mountain Ash and Alpine Ash forests and their implications for bird biodiversity. One major driver that has not been explicitly considered to date is climate change. The direct impacts on birds resulting from likely reduced rainfall and increased temperatures that are likely to manifest in the Central Highlands region have, to date, not been modeled. However, the impacts are likely to be substantial. For example, elevated temperatures and marked rainfall deficits preceding the 2009 wildfires appeared to contribute to widespread death of large old trees in the Central Highlands region (Lindenmayer et al., 2012). Climate and weather are critical drivers of fire regimes in south-eastern Australia (Bradstock, 2010) and climate change has a high likelihood of resulting in more frequent, more extensive, and more severe wildfires in Mountain Ash and Alpine Ash forests (Cary et al., 2012). This will, in turn, drive further losses of old growth forest (Lindenmayer and Taylor, 2020), increase the rate of decline in populations of large old trees (Lindenmayer et al., 2012) (see Figure 7), and alter the suitability of sites and landscapes for birds (Lindenmayer et al., 2019b). Additional fires may also trigger further salvage logging, with corresponding negative impacts on birds (Lindenmayer et al., 2018d). A major current knowledge gap is the extent to which climate change and weather influence the ability of bird assemblages to recover following wildfire and this area of work will be a focus for future research.
Data Availability Statement
The original contributions presented in the study are included in the article/supplementary materials, further inquiries can be directed to the corresponding author.
Ethics Statement
The animal study was reviewed and approved by ANU Animal Experimentation Ethics Committee.
Author Contributions
The idea for the paper was developed by DL and EB who wrote the paper together. DL led the research the paper was drawn from. LM completed the on ground field work and compiled the data. All authors contributed to the article and approved the submitted version.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
Many of the insights summarized in this paper could not have been generated without the significant input of three outstanding statisticians; Professor Ross Cunningham, the late Professor Jeff Wood, and Associate-Professor Wade Blanchard. Tabitha Boyer assisted with many aspects of manuscript preparation. Dr. Chris Taylor assisted with the development of maps in Figure 1. The late Dr. Dave Blair assisted with of research and monitoring co-ordination over the past 10 years.
References
Ashton, D. H. (1981a). “Fire in tall open forests (wet sclerophyll forests),” in Fire and the Australian Biota, eds A. M. Gill, R. H. Groves, and I. R. Noble (Canberra, ACT: Australian Academy of Science), 339–366.
Ashton, D. H. (1981b). “Tall Open Forests,” in Australian Vegetation, ed R. H. Groves (Cambridge: Cambridge University Press), 121–151.
Beruldsen, G. R. (2003). Australian Birds: Their Nests and Eggs. Kenmore Hills, QLD: G and E Beruldsen.
Blair, D., McBurney, L. W., B., Banks, S., and Lindenmayer, D.B. (2016). Disturbance gradient shows logging affects plant functional groups more than fire. Ecol. Appl. 26, 2280–2301. doi: 10.1002/eap.1369
Boer, M. M., Resco de Dios, V., and Bradstock, R. A. (2020). Unprecedented burn area of Australian mega forest fires. Nat. Clim. Change 10, 171–172. doi: 10.1038/s41558-020-0716-1
Boland, D. J., Brooker, M. I., Chippendale, G. M., Hall, N., Hyland, B. P., Johnson, R. D., et al. (2006). Forest Trees of Australia. Melbourne, VIC: CSIRO Publishing.
Bowd, E., Blair, D. P., and Lindenmayer, D. B. (in press). Prior disturbance legacy effects on plant recovery post-high severity wildfire. Ecosphere.
Bowd, E. J., Banks, S. C., Strong, C. L., and Lindenmayer, D. B. (2019). Long-term impacts of wildfire and logging on forest soils. Nat. Geosci. 12, 113–118. doi: 10.1038/s41561-018-0294-2
Bowd, E. J., Lindenmayer, D. B., Banks, S. C., and Blair, D. P. (2018). Logging and fire regimes alter plant communities. Ecol. Appl. 28, 826–841. doi: 10.1002/eap.1693
Bowman, D. M. J. S., Murphy, B. P., Neyland, D. L. J., Williamson, G. J., and Prior, L. D. (2014). Abrupt fire regime change may cause landscape-wide loss of mature obligate seeder forests. Glob. Change Biol. 20, 1008–1015. doi: 10.1111/gcb.12433
Bradstock, R. A. (2010). A biogeographic model of fire regimes in Australia: current and future implications. Glob. Ecol. Biogeogr. 19, 145–158. doi: 10.1111/j.1466-8238.2009.00512.x
Burns, E. L., Lindenmayer, D. B., Stein, J., Blanchard, W., McBurney, L., Blair, D., et al. (2015). Ecosystem assessment of mountain ash forest in the Central Highlands of Victoria, south-eastern Australia. Austral Ecol. 40, 386–399. doi: 10.1111/aec.12200
Burton, P. J., Messier, C., Smith, D. W., and Adamowicz, W. L. (2003). Towards Sustainable Management of the Boreal Forest. Ottawa, ON: National Research Council of Canada.
Cary, G., Blanchard, W., Foster, C. N., and Lindenmayer, D. B. (in press). Effects of altered fire regimes on critical timber production conservation rotations. Int. J. Wildland Fire.
Cary, G. J., Bradstock, R. A., Gill, A. M., and Williams, R. J. (2012). “Global change and fire regimes in Australia,” in Flammable Australia. Fire Regimes, Biodiversity, and Ecosystems in a Changing World, eds R. A. Bradstock, A. M. Gill, and R. J. Williams (Melbourne, VIC: CSIRO Publishing), 149–169.
Ceballos, G., Ehrlich, P. R., and Dirzo, R. (2017). Biological annihilation via the ongoing sixth mass extinction signaled by vertebrate population losses and declines. Proc. Natl. Acad. Sci. U.S.A. 114, E6089–E6096. doi: 10.1073/pnas.1704949114
Cunningham, R., and Lindenmayer, D. B. (2016). Approaches to landscape scale inference and design issues. Curr. Landsc. Ecol. Rep. 2, 42–50. doi: 10.1007/s40823-016-0019-4
Cunningham, R. B., Lindenmayer, D. B., Nix, H. A., and Lindenmayer, B. D. (1999). Quantifying observer heterogeneity in bird counts. Aust. J. Ecol. 24, 270–277. doi: 10.1046/j.1442-9993.1999.00971.x
Darveau, M., Beauchesne, P., Belanger, L., Hout, J., and Larue, P. (1995). Riparian forest strips as habitat for breeding birds in boreal forest. J. Wildl. Manag. 59, 67–78. doi: 10.2307/3809117
Debus, S. J. S., Davies, J. N., and Hollands, D. (2009). The Owls of Australia: A Field Guide to Australian Night Birds. Sydney, NSW: Envirobook in association with Birds Australia.
Enright, N. J., Fontaine, J. B., Bowman, D. M., Bradstock, R. A., and Williams, R. J. (2015). Interval squeeze: altered fire regimes and demographic responses interact to threaten woody species persistence as climate changes. Front. Ecol. Environ. 13, 265–272. doi: 10.1890/140231
Fairman, T. A., Bennett, L. T., and Nitschke, C. R. (2019). Short-interval wildfires increase likelihood of resprouting failure in fire-tolerant trees. J. Environ. Manag. 231, 59–65. doi: 10.1016/j.jenvman.2018.10.021
Fedrowitz, K. F., Koricheva, J., Baker, S. C., Lindenmayer, D. B., Palik, B., Rosenvald, R., et al. (2014). Can retention forestry help conserve biodiversity? A meta-analysis. J. Appl. Ecol. 51, 1669–1679. doi: 10.1111/1365-2664.12289
Flint, A., and Fagg, P. (2007). Mountain Ash in Victoria's State Forests. Silviculture reference manual No. 1. (Melbourne, VIC: Department of Sustainability and Environment).
Foster, C. N., Sato, C. F., Lindenmayer, D. B., and Barton, P. S. (2016). Integrating theory into empirical studied of disturbance interactions to improve management outcomes. Glob. Change Biol. 22, 1325–1335. doi: 10.1111/gcb.13155
Franklin, J. F., Lindenmayer, D. B., MacMahon, J. A., McKee, A., Magnuson, J., Perry, D. A., et al. (2000). Threads of continuity. Conserv. Pract. 1, 8–17. doi: 10.1111/j.1526-4629.2000.tb00155.x
Franklin, J. F., Spies, T. A., van Pelt, R., Carey, A. B., Thornburgh, D. A., Berg, D. R., et al. (2002). Disturbances and the structural development of natural forest ecosystems with silvicultural implications, using Douglas-fir forests as an example. For. Ecol. Manag. 155, 399–423. doi: 10.1016/S0378-1127(01)00575-8
Gosper, C. R., Fox, E., Burbidge, A. H., Craig, M. D., Douglas, T. K., Fitzsimons, J. A., et al. (2019a). Multi-century periods since fire in an intact woodland landscape favour bird species declining in an adjacent agricultural region. Biol. Conserv. 230, 82–90. doi: 10.1016/j.biocon.2018.12.011
Gosper, C. R., Watson, S. J., Fox, E., Burbidge, A. H., Craig, M. D., Douglas, T. K., et al. (2019b). Fire-mediated habitat change regulates woodland bird species and functional group occurrence. Ecol. Appl. 29:e01997. doi: 10.1002/eap.1997
Gratkowski, H. J. (1956). Windthrow around staggered settings in old-growth Douglas-fir. For. Sci. 2, 60–74.
Hutto, R. L., Keane, R. E., Sherriff, R. L., Rota, C. T., Eby, L. A., and Saab, V. A. (2016). Toward a more ecologically informed view of severe forest fires. Ecosphere 7:e01255. doi: 10.1002/ecs2.1255
Inger, R., Gregory, R., Duffy, J. P., Stott, I., Vorisek, P., and Gaston, K. (2014). Common European birds are declining rapidly while less abundant species are rising. Ecol. Lett. 18, 28–36. doi: 10.1111/ele.12387
Intergovernmental Science-policy Platform on Biodiversity and Ecosystem Services (IPBES) (2019). IPBES Global Assessment Summary for Policymakers. New York, NY: United Nations.
Jones, M. W., Smith, A., Betts, R., Canadell, J. G., Prentice, C. I., and Le Quéré, C. (2020). Climate Change Increases Risk of Wildfires. ScienceBrief Review [Online]. Available online at: https://sciencebrief.org/briefs/wildfires (accessed February 3, 2020).
Keeley, J. E. (2009). Fire intensity, fire severity, and burn severity: a brief review and suggested usage. Int. J. Wildland Fire 18, 116–126. doi: 10.1071/WF07049
Keith, D., McCaw, W. L., and Whelan, R. J. (2002). “Fire regimes in Australian heathlands and their effects on plants,” in Flammable Australia. The Fire Regimes and Biodiversity of a Continent, eds R. Bradstock, J. Williams, and A. M. Gill (Cambridge: Cambridge University Press), 199–237.
Lindenmayer, D., Blanchard, W., Blair, D., McBurney, L., Taylor, C., Scheele, B., et al. (2020b). The response of arboreal marsupials to long-term changes in forest disturbance. Anim. Conserv. doi: 10.1111/acv.12634. [Epub ahead of print].
Lindenmayer, D. B. (2017). Conserving large old trees as small natural features. Biol. Conserv. 211, 51–59. doi: 10.1016/j.biocon.2016.11.012
Lindenmayer, D. B., Blair, D., and McBurney, L. (2019a). Variable retention harvesting in Victoria's Mountain Ash (Eucalyptus regnans) forests (southeastern Australia). Ecol. Process. 8:2. doi: 10.1186/s13717-018-0156-2
Lindenmayer, D. B., Blanchard, W., Blair, D., and McBurney, L. (2018a). The road to oblivion—quantifying pathways in the decline of large old trees. For. Ecol. Manag. 430, 259–264. doi: 10.1016/j.foreco.2018.08.013
Lindenmayer, D. B., Blanchard, W., Blair, D., McBurney, L., and Banks, S. C. (2016). Environmental and human drivers of large old tree abundance in Australian wet forests. For. Ecol. Manag. 372, 266–235. doi: 10.1016/j.foreco.2016.04.017
Lindenmayer, D. B., Blanchard, W., Blair, D., McBurney, L., Stein, J., and Banks, S. C. (2018b). Empirical relationships between tree fall and landscape-level amounts of logging and fire. PLoS ONE 13:e0193132. doi: 10.1371/journal.pone.0193132
Lindenmayer, D. B., Blanchard, W., Blair, D., Westgate, M. J., and Scheele, B. C. (2019b). Spatio-temporal effects of logging and fire on forest birds. Ecol. Appl. 29:e01999. doi: 10.1002/eap.1999
Lindenmayer, D. B., Blanchard, W., McBurney, L., Blair, D., Banks, S., Likens, G. E., et al. (2012). Interacting factors driving a major loss of large trees with cavities in a forest ecosystem. PLoS ONE 7:e41864. doi: 10.1371/journal.pone.0041864
Lindenmayer, D. B., Blanchard, W., McBurney, L., Blair, D., Banks, S. C., Driscoll, D. A., et al. (2014). Complex responses of birds to landscape-level fire extent, fire severity, and environmental drivers. Divers. Distrib. 20, 467–477. doi: 10.1111/ddi.12172
Lindenmayer, D. B., Cunningham, R. B., and Donnelly, C. F. (1997). Decay and collapse of trees with hollows in eastern Australian forests: impacts on arboreal marsupials. Ecol. Appl. 7, 625–641. doi: 10.1890/1051-0761(1997)007[0625:DACOTW]2.0.CO;2
Lindenmayer, D. B., Cunningham, R. B., Donnelly, C. F., and Franklin, J. F. (2000a). Structural features of old growth Australian montane ash forests. For. Ecol. Manag. 134, 189–204. doi: 10.1016/S0378-1127(99)00257-1
Lindenmayer, D. B., Cunningham, R. B., Nix, H. A., Tanton, M. T., and Smith, A. P. (1991). Predicting the abundance of hollow-bearing trees in montane ash forests of southeastern Australia. Aust. J. Ecol. 16, 91–98. doi: 10.1111/j.1442-9993.1991.tb01484.x
Lindenmayer, D. B., Hobbs, R. J., Likens, G. E., Krebs, C., and Banks, S. (2011). Newly discovered landscape traps produce regime shifts in wet forests. Proc. Natl. Acad. Sci. U.S.A. 108, 15887–15891. doi: 10.1073/pnas.1110245108
Lindenmayer, D. B., Lane, P., Westgate, M., Scheele, B., Foster, C., Sato, C., et al. (2018c). Tests of predictions associated with temporal changes in Australian bird populations. Biol. Conserv. 222, 212–221. doi: 10.1016/j.biocon.2018.04.007
Lindenmayer, D. B., Mackey, B. G., Cunningham, R. B., Donnelly, C. F., Mullen, I. C., McCarthy, M. A., et al. (2000b). Factors affecting the presence of the cool temperate rain forest tree myrtle beech (Nothofagus cunninghamii) in southern Australia: integrating climatic, terrain, and disturbance predictors of distribution patterns. J. Biogeogr. 27, 1001–1009. doi: 10.1046/j.1365-2699.2000.00443.x
Lindenmayer, D. B., McBurney, L., Blair, D., Wood, J., and Banks, S. C. (2018d). From unburnt to salvage logged: quantifying bird responses to different levels of disturbance severity. J. Appl. Ecol. 55, 1626–1636. doi: 10.1111/1365-2664.13137
Lindenmayer, D. B., and McCarthy, M. A. (2002). Congruence between natural and human forest disturbance: a case study from Australian montane ash forests. For. Ecol. Manag. 155, 319–335. doi: 10.1016/S0378-1127(01)00569-2
Lindenmayer, D. B., and Ough, K. (2006). Salvage logging in the montane ash eucalypt forests of the Central Highlands of Victoria and its potential impacts on biodiversity. Conserv. Biol. 20, 1005–1015. doi: 10.1111/j.1523-1739.2006.00501.x
Lindenmayer, D. B., and Sato, C. (2018). Hidden collapse is driven by fire and logging in a socioecological forest ecosystem. Proc. Natl. Acad. Sci. U.S.A. 115, 5181–5186. doi: 10.1073/pnas.1721738115
Lindenmayer, D. B., and Taylor, C. (2020). Extensive recent wildfires demand more stringent protection of critical old growth forest. Pac. Conserv. Biol. 26, 384–394002E doi: 10.1071/PC20037
Lindenmayer, D. B., Westgate, M. J., Scheele, B. C., Foster, C. N., and Blair, D. P. (2019c). Key perspectives on early successional forests subject to stand-replacing disturbances. For. Ecol. Manag. 454:117656. doi: 10.1016/j.foreco.2019.117656
Lindenmayer, D. B., Woinarski, J., Legge, S., Southwell, D., Lavery, T., Robinson, N., et al. (2020a). A checklist of attributes for effective monitoring of threatened species and threatened ecosystems. J. Environ. Manag. 262:110312. doi: 10.1016/j.jenvman.2020.110312
Lindenmayer, D. B., Wood, J., McBurney, L., Blair, D., and Banks, S. C. (2015). Single large versus several small: the SLOSS debate in the context of bird responses to a variable retention logging experiment. For. Ecol. Manag. 339, 1–10. doi: 10.1016/j.foreco.2014.11.027
Lindenmayer, D. B., Wood, J., McBurney, L., Michael, D., Crane, M., MacGregor, C., et al. (2010). Comparing bird species richness and assemblage composition between montane ash eucalypt forest and cool temperate rainforests–an empirical study from Victoria, south-eastern Australia. Emu 110, 109–117. doi: 10.1071/MU09074
Lindenmayer, D. B., Wood, J., Michael, D., Crane, M., MacGregor, C., Montague-Drake, R., et al. (2009). Are gullies best for biodiversity? an empirical examination of Australian wet forest types. For. Ecol. Manag. 258, 169–177. doi: 10.1016/j.foreco.2009.04.002
Loyn, R. H. (1985). Bird populations in successional forests of Mountain Ash Eucalyptus regnans in Central Victoria. Emu 85, 213–230. doi: 10.1071/MU9850213
Maisey, A. C., Haslem, A., Leonard, S., and Bennett, A. F. (2020). Foraging by an avian ecosystem engineer extensively modifies the litter and soil layer in forest ecosystems. Ecol. Appl. 31:e02219. doi: 10.1002/eap.2219
McCarthy, M. A., Gill, A. M., and Lindenmayer, D. B. (1999). Fire regimes in mountain ash forest: evidence from forest age structure, extinction models, and wildlife habitat. For. Ecol. Manag. 124, 193–203. doi: 10.1016/S0378-1127(99)00066-3
Milledge, D. R., Palmer, C. L., and Nelson, J. L. (1991). “‘Barometers of change’: the distribution of large owls and gliders in Mountain Ash forests of the Victorian Central Highlands and their potential as management indicators,” in Conservation of Australia's Forest Fauna, ed D. Lunney (Sydney, NSW: Royal Zoological Society of N.S.W.), 53–65.
Miller, D. H. (1981). Energy at the Surface of the Earth: An Introduction to the Energetics of Ecosystems. New York, NY: Academic Publishers.
Montague-Drake, R. M., Lindenmayer, D. B., and Cunningham, R. B. (2009). Factors affecting site occupancy by woodland bird species of conservation concern. Biol. Conserv. 142, 2896–2903. doi: 10.1016/j.biocon.2009.07.009
Nelson, J. L., and Morris, B. J. (1994). Nesting requirements of the Yellow-tailed Black Cockatoo Calyptorynchus funereus in Mountain Ash forest Eucalyptus regnans and implications for forest management. Wildl. Res. 21, 257–268. doi: 10.1071/WR9940267
Pyke, G. H., and Recher, H. F. (1983). “Censusing Australian birds: a summary of procedures and a scheme for standardisation of data presentation and storage,” in Methods of Censusing Birds in Australia, ed S. J. Davies (Perth, WA: Proceedings of a symposium organised by the Zoology section of the ANZAAS and the Western Australian Group of the Royal Australasian Ornithologists Union. Department of Conservation and Environment), 55–63.
Sanderson, F. J., Donald, P. F., Pain, D. J., Burfield, I. J., and van Bommel, F. P. J. (2006). Long-term population declines in Afro-Paleartic migrant birds. Biol. Conserv. 131, 93–105. doi: 10.1016/j.biocon.2006.02.008
Shine, J. (2020). Statement Regarding Australian Bushfires [Online]. Canberra, ACT: Australian Academy of Science. Available online at: https://www.science.org.au/news-and-events/news-and-media-releases/australian-bushfires-why-they-are-unprecedented (accessed February 6, 2020).
Smith, A. L., Blanchard, W., Blair, D., McBurney, L., Banks, S. C., Driscoll, D. A., et al. (2016). The dynamic regeneration niche of a forest following a rare disturbance event. Divers. Distrib. 22, 457–467. doi: 10.1111/ddi.12414
Sodhi, N. S., and Ehrlich, P. R. (eds.). (2010). Conservation Biology for All. Oxford: Oxford University Press.
Sommerfeld, A., Senf, C., Buma, B., D'Amato, A. W., Després, T., Díaz-Hormazábal, I., et al. (2018). Patterns and drivers of recent disturbances across the temperate forest biome. Nat. Commun. 9:4355. doi: 10.1038/s41467-018-06788-9
Swanson, M. E., Franklin, J. F., Beschta, R. L., Crisafulli, C. M., DellaSala, D. A., Hutto, R. L., et al. (2011). The forgotten stage of forest succession: early-successional ecosystems on forest sites. Front. Ecol. Environ. 9, 117–125. doi: 10.1890/090157
Taylor, C., Blanchard, W., and Lindenmayer, D. B. (2020). Does forest thinning reduce fire severity in Australian eucalypt forests? Conserv. Lett. doi: 10.1111/conl.12766. [Epub ahead of print].
Taylor, C., Cadenhead, N., Lindenmayer, D. B., and Wintle, B. A. (2017). Improving the design of a conservation reserve for a critically endangered species. PLoS ONE 12:e0169629. doi: 10.1371/journal.pone.0169629
Taylor, C., and Lindenmayer, D. B. (2019). The adequacy of Victoria's protected areas for conserving its forest-dependent fauna. Austral Ecol. 44, 1076–1090. doi: 10.1111/aec.12805
Taylor, C., and Lindenmayer, D. B. (2020). Temporal fragmentation of a critically endangered ecosystem. Austral Ecol. 45, 340–354. doi: 10.1111/aec.12863
Taylor, C., McCarthy, M. A., and Lindenmayer, D. B. (2014). Non-linear effects of stand age on fire severity. Conserv. Lett. 7, 355–370. doi: 10.1111/conl.12122
Tiribelli, F., Morales, J. M., Gowda, J. H., Mermoz, M., and Kitzberger, T. (2018). Non-additive effects of alternative stable states on landscape flammability in NW Patagonia: fire history and simulation modelling evidence. Int. J. Wildland Fire 28, 149–159. doi: 10.1071/WF18073
Wintle, B. W., Cadenhead, N., Morgain, R. A., Legge, S. M., Bekessy, S. A., Possingham, H. P., et al. (2019). Spending to save: what will it cost to halt Australia's extinction crisis. Conserv. Lett. 12:e12682. doi: 10.1111/conl.12682
Zald, S. J., and Dunn, C. (2017). Severe fire weather and intensive forest management increase fire severity in a multi-ownership landscape. Ecosphere 28, 1068–1080. doi: 10.1002/eap.1710
Zylstra, P. (2018). Flammability dynamics in the Australian Alps. Austral Ecol. 43, 578–591. doi: 10.1111/aec.12594
Zylstra, P. J., Bradstock, R. A., Bedward, M., Penman, T. D., Doherty, M. D., Weber, R. O., et al. (2016). Biophysical mechanistic modelling quantifies the effects of plant traits on fire severity: species, not surface fuel loads determine flame dimensions in eucalypt forests. PLoS ONE 11:e0160715. doi: 10.1371/journal.pone.0160715
Keywords: mountain ash forest, logging, fire, landscape change, interacting drivers, old growth forest, birds (Australian terrestrial), Mountain Ash Eucalyptus regnans
Citation: Lindenmayer D, Bowd E and McBurney L (2021) Long-Term Empirical Studies Highlight Multiple Drivers of Temporal Change in Bird Fauna in the Wet Forests of Victoria, South-Eastern Australia. Front. Ecol. Evol. 9:610147. doi: 10.3389/fevo.2021.610147
Received: 25 September 2020; Accepted: 12 January 2021;
Published: 11 February 2021.
Edited by:
Natalia Ocampo-Peñuela, ETH Zürich, SwitzerlandReviewed by:
Dan Cogalniceanu, Ovidius University, RomaniaCarl R. Gosper, Department of Biodiversity, Conservation and Attractions (DBCA), Australia
Copyright © 2021 Lindenmayer, Bowd and McBurney. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: David Lindenmayer, ZGF2aWQubGluZGVubWF5ZXJAYW51LmVkdS5hdQ==