 Mia Svensk1,2*
Mia Svensk1,2* Marco Pittarello3
Marco Pittarello3 Ginevra Nota3
Ginevra Nota3 Manuel K. Schneider4
Manuel K. Schneider4 Eric Allan2
Eric Allan2 Pierre Mariotte1
Pierre Mariotte1 Massimiliano Probo1
Massimiliano Probo1- 1Grazing Systems, Agroscope, Nyon, Switzerland
- 2Department of Ecology and Evolution, University of Bern, Bern, Switzerland
- 3Department of Agricultural, Forest and Food Sciences (DISAFA), University of Torino, Turin, Italy
- 4Forage Production and Grassland, Agroscope, Zurich, Switzerland
Green alder (Alnus viridis) is a shrub species that has expanded over former pastures in Central Europe due to land abandonment, leading to negative agri-environmental impacts, such as a reduction in forage yield and quality and an increase in nitrate leaching. Robust livestock breeds such as Highland cattle could be used to control A. viridis encroachment. The objectives of this study were to investigate the impact of A. viridis encroachment on plant community composition and diversity and to map the spatial distribution of Highland cattle in A. viridis-encroached pastures. During the summer of 2019, three different Highland cattle herds were placed along an A. viridis encroachment gradient. A total of 58 botanical surveys were carried out before grazing to assess plant community composition, pastoral value, and ecological indicator values. The spatial distribution of cattle was studied during the whole grazing period by monitoring six to eight cows equipped with GPS collars in each herd. Plant species associated with higher pastoral values of the vegetation were found in areas with lower A. viridis cover, while highly encroached areas were dominated by a few nitrophilus and shade-tolerant broad-leaved species and by ferns. Cattle spent more time in areas with higher pastoral value but did not avoid areas with high cover of A. viridis, on steep slopes or far from water. These results show that Highland cattle are able to tolerate harsh environmental conditions and that they can exploit A. viridis-encroached pastures. This suggests that they have a high potential to reduce A. viridis encroachment in the long-term.
Introduction
Throughout the last decades, socioeconomic changes have led to a reduction in agro-pastoral activities in European mountain areas, triggering profound landscape modifications, and widespread natural reforestation and shrub encroachment in former pastures (Estel et al., 2015). For instance, from 1985 to 2013, shrublands and forests have increased by 10.6% across Switzerland, with the largest increases in the Alps (Abegg et al., 2020). Shrublands now cover an area of 679 km2 in Switzerland, with about 70% of them dominated by pure stands of green alder [Alnus viridis (Chaix.) D. C.], which is the most rapidly expanding shrub species in Central Europe (Anthelme et al., 2007). A. viridis is a pioneer species, which lives in symbiosis with the N2-fixing actinomycete Frankia alni (Huss-Daniel, 1997), and its presence is determined by land use intensity (Caviezel et al., 2017). Encroachment by A. viridis can have several negative agri-environmental impacts: in particular, A. viridis-dominated shrublands have lower plant and animal diversity and forage quality than open grazed areas (Anthelme et al., 2001). A recent study showed that A. viridis encroachment resulted in a rapid decline in plant species richness, with dense A. viridis patches hosting 62% less species than adjacent open pastures (Zehnder et al., 2020). Moreover, there is high nitrate and dissolved organic carbon leaching from A. viridis shrublands, which results in water pollution, soil acidification, hydrological drawbacks, and gaseous nitrogen losses (Bühlmann et al., 2016; van den Bergh et al., 2018). It is therefore important to find methods that control A. viridis encroachment in order to reverse biodiversity losses and to protect key ecosystem services.
Controlling shrub encroachment is challenging because mechanical removal is extremely difficult and expensive in harsh and marginal mountain conditions due to steep slopes and a lower density of road network. Moreover, many shrub species are not palatable to grazing animals, since their foliage has low digestibility, mostly due to high tannin concentrations (Waghorn, 2008). However, some robust livestock species and breeds can forage on shrubs and digest their leaves, thanks to ruminal bacterial populations, which can degrade lignified material and which have high resistance to tannins (Manousidis et al., 2016). Highland cattle are a robust breed that are increasingly being reared in different European alpine countries due to their ability to forage on poor quality and shrub-encroached pastures (Pauler et al., 2020a,b). As a consequence, the particular grazing behavior of Highland cattle can also result in distinct pasture botanical composition, with reduced woody plant species cover in the long term (Pauler et al., 2019). However, the impact of robust livestock on shrub encroachment may depend on environmental and management conditions, such as slope, pastoral value of the vegetation, and distance to water sources and attractive supplements for livestock (Probo et al., 2014; Homburger et al., 2015; Pittarello et al., 2016). It is therefore important to evaluate the conditions under which robust livestock breeds are able to effectively reduce shrub encroachment in alpine pasture.
However, to the best of our knowledge, no study has assessed in detail the drivers related to plant community composition and diversity in A. viridis-encroached pastures nor the potential of Highland cattle to reduce A. viridis encroachment. For this reason, in the present study, we aimed (i) to disentangle the topographic and ecological drivers of plant community composition and diversity in A. viridis-encroached pastures, from Northern to Southern Alps and (ii) to measure the spatial distribution of Highland cattle in A. viridis-encroached pastures to determine whether they are attracted to or avoid A. viridis patches. Here, we hypothesized that spatial distribution of Highland cows would vary depending on A. viridis cover and other factors such as distance to water, slope, and the pastoral value of the vegetation. More specifically, we expected cows to spend more time in areas around water sources and in more valuable pasture and to avoid steep slopes.
Methods
To investigate vegetation features and cattle spatial distribution, three Highland cattle herds were placed in four different A. viridis-encroached paddocks in the Swiss and Italian Alps in the summer of 2019. Three paddocks were located in Switzerland: Bovonne 1 and Bovonne 2 (in the canton of Vaud) and Champlong (in the canton of Valais). Bovonne 1 and Bovonne 2 were adjacent paddocks, grazed one after the other by the same herd. The fourth paddock was located in Val Vogna (in the province of Vercelli) in the Italian Alps (Table 1). Highland cattle grazed each paddock for approximately 1 month: in July (Bovonne 1), from mid-July to late-August (Champlong), and in August (Bovonne 2 and Val Vogna). All the herds included suckler cows, heifers, and calves, varying in age from 6 months to 17 years. The paddocks had similar environmental conditions in terms of elevation and slope, were grazed at comparable livestock stocking rates, and were representative of a gradient of A. viridis encroachment, i.e., with A. viridis cover values ranging from 20 to 70% (Table 1). A water trough was installed in each of the two paddocks in Bovonne, while many natural streams were present both in Val Vogna and Champlong paddocks.
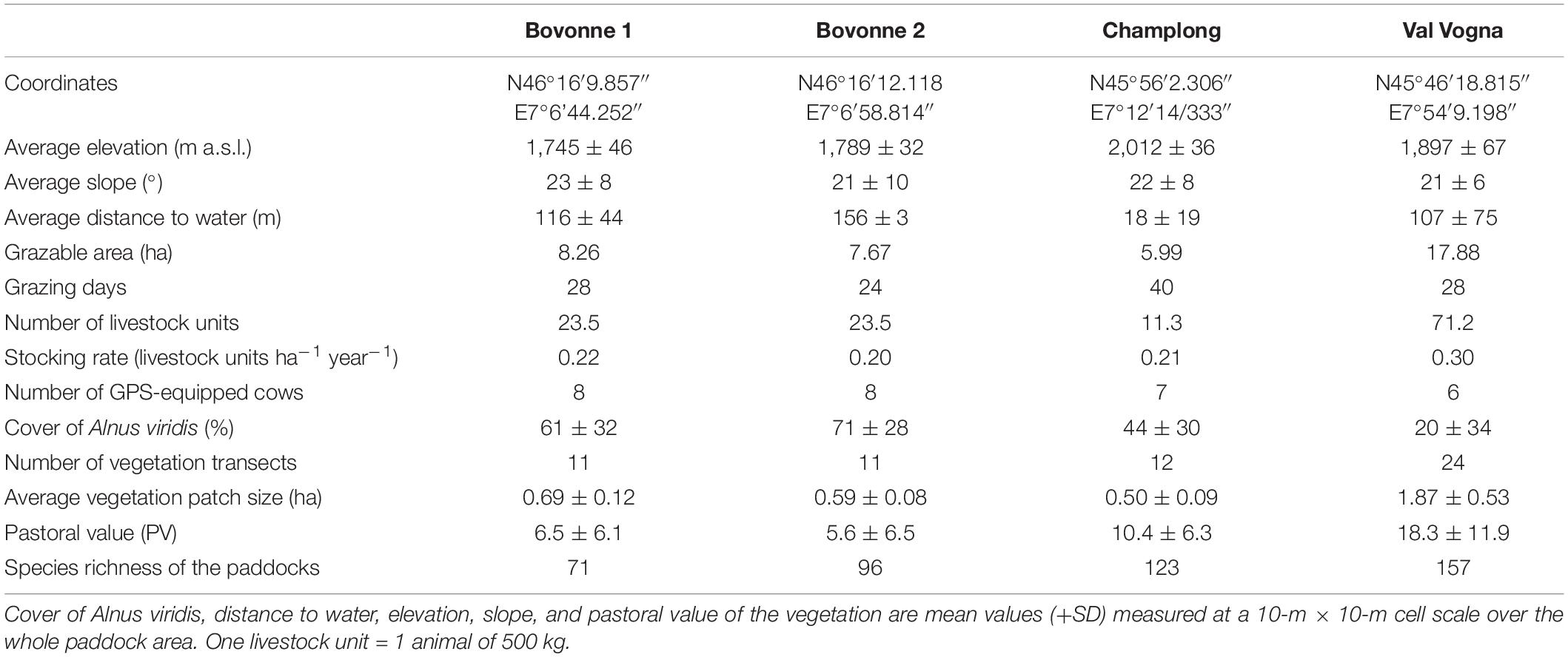
Table 1. Topographical, management, and vegetation characteristics of the four A. viridis-encroached paddocks.
In order to characterize the drivers of plant community composition and diversity within each paddock, the botanical composition was determined along 12.5-m transects. Transects were placed in patches (1.08 ± 0.22 ha, Table 1) with homogeneous botanical composition and vegetation structure, and the vertical point-quadrat method was used to record plant species (Daget and Poissonet, 1971). At 50-cm intervals along each transect, the plant species touching a steel needle were identified and recorded (i.e., 25 points per transect). Since rare species are often missed by this method, all other species within a 1-m buffer area around the transect were also recorded (Kohler et al., 2004). A total of 58 vegetation transects were surveyed in the four paddocks (Table 1). Plant species nomenclature followed Aeschimann et al. (2004). The relative abundance of each plant species was computed by dividing species frequency of occurrence by the sum of frequency of occurrence values for all species in the transect and multiplying it by 100. A species relative abundance value of 0.3% was assigned to all the species found only in the buffer zone and not in the main transect, following Pittarello et al. (2016). We then calculated the pastoral value (PV) of the vegetation, based on the species composition. The PV is a synthetic index summarizing forage yield, quality, and palatability for livestock (Pittarello et al., 2018). To estimate PV, we attributed each species an index of specific quality (Cavallero et al., 2007). The index of specific quality depends on the preference, morphology, structure, and productivity of the plant species, and it ranges from 0 (low) to 5 (high). The PV, which ranges from 0 to 100, was calculated as follows (Daget and Poissonet, 1971):
where SRAi is the species relative abundance, and ISQi is the index of specific quality value of the species i.
Moreover, we used the indicator values of Landolt et al. (2010) for each plant species. These indicate the environmental conditions in the habitats in which the given species occurs and are a measure of its environmental preferences. We used the light (L), soil moisture (F), and nutrient value (N) (nitrogen) indicators. We then determined the mean ecological conditions of each vegetation patch, as the mean Landolt value for L, F, and N by multiplying each species indicator value by its relative abundance. For each vegetation transect, two plant diversity indexes were computed: species richness and Shannon diversity index H′ (Magurran, 1998). A. viridis canopy cover (%) was visually estimated within each vegetation patch.
Each paddock was subdivided into 10- × 10-m grid cells, and the distance to water sources (i.e., streams and water troughs), the aspect, elevation, and slope were calculated for the centroid of each cell using (QGIS/ArcView/SAGA/R). Moreover, A. viridis cover and PV were calculated for each grid cell using the values of the corresponding vegetation patch. If a cell covered more than one vegetation patch, the patch accounting for most of the cell area was considered. To avoid issues with circular variables (where high and low values are close together), aspect was transformed into southness (180 − | aspect − 180|) (Chang et al., 2004). In each herd, six to eight cows were equipped with GPS collars (Followit© Tellus GPS collars) that recorded their position every 10 min during the whole grazing period, with an average accuracy of 2–5 m (Table 1). The number of GPS fixes was calculated for each grid cell.
All statistical analyses were performed using R version 3.4.4. A partial canonical correspondence analysis (CCA, vegan package), taking into account the spatial distance between botanical surveys, was performed to describe how PV, biodiversity indices, ecological indicators (i.e., Landolt indexes), and topography (i.e., slope and aspect) were related to plant community composition (after Hellinger transformation). Moreover, relationships between A. viridis cover and diversity indexes and PV were analyzed with generalized linear mixed models with Poisson (species richness and PV) and quasi-Poisson (Shannon diversity index H′) distributions, with paddock considered as a random factor (lme4 package). The spatial distribution of cattle calculated from GPS data (i.e., the number of GPS fixes counted in each grid cell) was investigated separately for three distinct periods within each paddock, i.e., the beginning (P1), middle (P2), and end (P3) of the grazing period. These periods had equal lengths within each paddock and varied between 1 and 2 weeks depending on the length of the grazing period (Table 1). The impacts of slope, A. viridis cover, distance to water, and PV on the number of GPS fixes were assessed using linear regression models with zero-inflated negative binomial likelihood, as this error distribution accounted for overdispersion in the response variable (tested using the AER package). Homburger et al. (2015) found this likelihood structure to be appropriate for GPS counts in zones incompletely exploited by cattle. In order to account for the spatial autocorrelation of data, a spatial structure error term (using a two-dimensional random walk of second order) was included, and the model was fitted by Integrated Nested Laplace Approximation (INLA).
Results
We found a total of 252 plant species in all the paddocks, with the highest species richness detected in Val Vogna (157 species, Table 1). The results of the CCA analysis highlighted different ecological, topographical, and vegetation impacts on plant community composition (Figure 1), which significantly explained the variation in community composition across the paddocks (CCA 1 = 20.9%, CCA2 = 19.0%, CCA3 = 12.9%, CCA 4 = 10.9%, and total = 84.6%). Vegetation patches within the two Bovonne paddocks were characterized by higher A. viridis cover and nitrogen enrichment (i.e., higher values for N indicator). In contrast, the Champlong paddock had many open pasture patches (with higher light availability), while Val Vogna patches were more heterogeneous. Four patches in Val Vogna were well distinguished and corresponded to vegetation dominated by dwarf shrubs, such as Vaccinium vitis-idaea, Juniperus nana, and Rhododendron ferrugineum, with low PV (Figure 1B). A. viridis cover had a significant negative effect on plant species richness, Shannon diversity index H′, and PV (p < 0.001). Indeed, plant species associated with higher PV (e.g., Festuca pratensis and Trifolium pratense) were often found in the most open areas, characterized by a lower A. viridis cover, as well as typical small sized pasture species (e.g., Medicago lupulina and Polygala alpestris), which were associated with the highest light availability and biodiversity (both in terms of species richness and Shannon diversity index H′). Conversely, areas with high levels of A. viridis encroachment and with high indicator values for N were dominated by a few tall broad-leaved species, such as Adenostyles alliariae and Cicerbita alpina, together with ferns (mainly Athyrium filix-femina and Dryopteris dilatata).
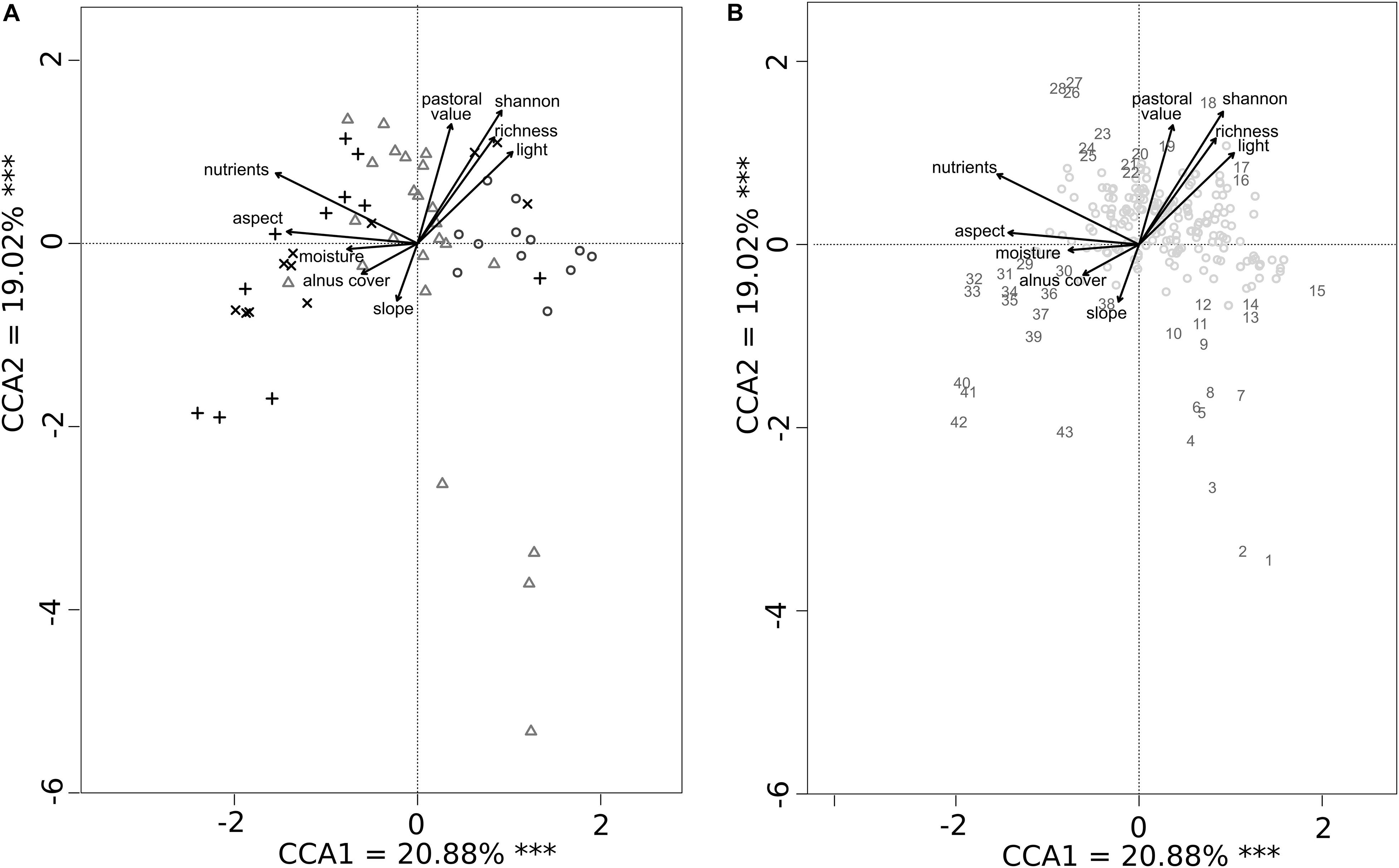
Figure 1. Partial canonical correspondence analysis (CCA) showing plant community composition explained by Alnus cover, slope, aspect, pastoral value, species richness, Shannon index, and ecological indicators for nutrients, light, and moisture (Landolt indexes). Both dimensions presented are significant (∗∗∗p < 0.001). (A) Botanical surveys are represented for Bovonne 1 (+), Bovonne 2 (x), Champlong (o), and Val Vogna (Δ). (B) Plant species are represented by numbers (detailed below) for the most important species and by dots for the remaining species. (1) Vaccinium vitis-idaea, (2) Juniperus nana, (3) Cryptogramma crispa, (4) Gymnocarpium Dryopteris, (5) Astrantia minor, (6) Rhododendron ferrugineum, (7) Arnica montana, (8) Avenella flexuosa, (9) Festuca scabriculmis, (10) Rosa pendulina, (11) Vaccinium myrtillus, (12) Calamagrostis villosa, (13) Melampyrum sylvaticum, (14) Plantago lanceolata, (15) Epilobium montanum, (16) Medicago lupulina, (17) Thlaspi montanum, (18) Polygala alpestris, (19) Molinia arundinacea, (20) Hypericum richeri, (21) Festuca pratensis, (22) Trifolium pratense, (23) Arabis ciliata, (24) Rumex alpestris, (25) Campanula latifolia, (26) Capsella bursa-pastoris, (27) Chenopodium bonus-henricus, (28) Trifolium badium, (29) Cardamine resedifolia, (30) Adenostyles alliariae, (31) Acer pseudoplatanus, (32) Rubus saxatilis, (33) Thalictrum aquilegiifolium, (34) Saxifraga rotundifolia, (35) Cicerbita alpina, (36) Urtica dioica, (37) Dryopteris dilatata, (38) Rubus idaeus, (39) Athyrium filix-femina, (40) Prenanthes purpurea, (41) Sorbus aucuparia, (42) Picea abies, and (43) Oxalis acetosella.
Estimates of linear regression models showed that livestock spatial distribution was often positively influenced by PV. Cattle spent more time in high PV areas in Bovonne 1 (during periods 1 and 2), Champlong (period 3), and Val Vogna (periods 2 and 3) (Figure 2). In Bovonne 2, the PV did not influence cattle spatial distribution. The cover of A. viridis, slope, and distance to water sources did not generally affect livestock spatial distribution. A. viridis cover and slope only had significant effects in the Champlong paddock during the third and the first grazing period, respectively; in both cases, cattle spent more time in areas with high A. viridis cover and on steeper slopes. Distance to water had a negative effect only in periods 1 and 2 in Bovonne 1 paddock, where water sources were less abundant, meaning cattle spent more time close to water.
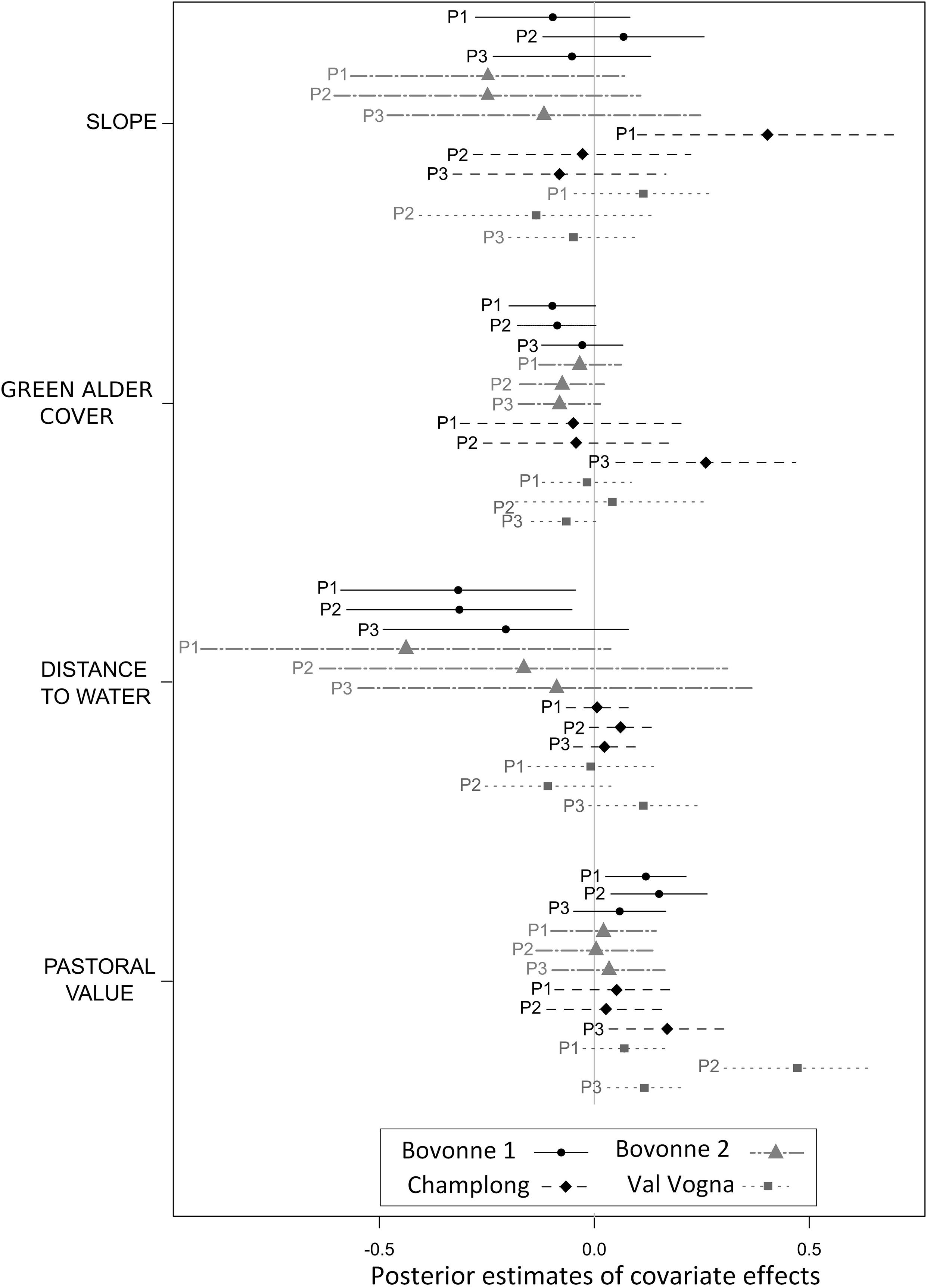
Figure 2. Effects of four covariates on livestock spatial distribution. The dots show the mean estimated effects and the line the 95% confidence interval, in the paddocks of Bovonne 1 (black line), Bovonne 2 (gray dashed line), Champlong (black dashed line), and Val Vogna (gray dotted line) for the three grazing periods (P1, P2, and P3).
Discussion
Overall, plant communities with higher A. viridis cover had lower plant diversity and were dominated by a few broad-leaved species, like A. alliariae and C. alpina, which prefer N-enriched conditions. These results support the findings of Anthelme et al. (2001) and Zehnder et al. (2020), who also showed a decline in alpine plant diversity with A. viridis encroachment. The few species that survive in A. viridis-dominated areas are able to take advantage of the N-enrichment and increased shading, and they replace many typical pasture species that are highly light demanding. Interestingly, other than broad-leaved species, the understory vegetation of many A. viridis patches was dominated by ferns (in particular, A. filix-femina and D. dilatata). These ferns are also highly shade tolerant, but they typically prefer areas with lower N levels, compared to the broad-leaved species (Landolt et al., 2010). They may therefore be more abundant in areas recently invaded by A. viridis where N levels have not increased so much. These results show that encroachment by A. viridis threatens the diversity of Alpine pastures and leads to a loss of typical pasture species.
The Highland cattle did not avoid patches with a high density of A. viridis. In fact, they were able to graze in most A. viridis-covered areas, therefore indicating that they could be valuable in grazing and reducing A. viridis patches. In contrast to many other breeds (e.g., Homburger et al., 2015), Highland cattle were also capable of grazing on the steepest slopes and were able to spend time in areas further from water sources, as they were not significantly influenced by these harsh conditions. This is in line with the observations of Pauler et al. (2020a) who found that Highland cattle used space more evenly and avoid steep slopes less than other breeds. Moreover, cattle even preferred A. viridis patches and steep slopes in Champlong, during certain grazing periods, which shows that cattle are able to move to areas with the most unfavorable conditions and can stay there for relatively long periods (approximately 2 weeks). Cattle also responded differently to the various factors during the time they were on the paddocks. This indicates that the herds may explore the paddock intensively at the beginning of the grazing period before selecting particular areas later on. However, despite the fact that Highland cattle graze less selectively on plants compared to other breeds (Pauler et al., 2020b), they were still mostly attracted to valuable pastures. Thus, providing a mosaic of open pasture areas with higher quality forage in A. viridis-encroached paddocks may be a good solution to respect grazing preferences while increasing grazing pressure on A. viridis-encroached patches. Moreover, this type of setting would enhance the potential for seed translocation through endo- and epi-zoochory by Highland cattle (Mouissie, 2004; Cosyns et al., 2005). Indeed, in the study of Pauler et al. (2019), plant species relying on epizoochory were significantly more frequent in pastures grazed by Highland cattle than in paddocks grazed by other breeds, probably because the long fur of this breed is ideal for transporting seeds. Animal movements might thus provide additional seed translocation fluxes from pastures to encroached areas to facilitate the restoration of typical pasture vegetation in the long-term (Pittarello et al., 2016). However, future research is needed to assess the simultaneous effects of cattle grazing, trampling, and seed transportation on the restoration of pasture vegetation in A. viridis-encroached areas. We observed considerable impact of the cattle on vegetation within the encroached areas (data not shown), including intense defoliation of A. viridis shrubs and a reduction of their canopy cover, due to browsing, as well as damage to branches due to cattle movement and scratching. Moreover, an increase in bare soil due to livestock trampling and a considerable number of dung pats was observed within A. viridis-encroached areas. If repeated for several years, these effects of the cattle could provide favorable conditions for the recolonization of typical pasture vegetation in formerly encroached areas. The repetition of the botanical surveys along the fixed vegetation transects installed during this study could allow us to assess the medium- and long-term impacts produced by Highland cattle on vegetation. In addition, testing improved techniques to attract livestock to the most encroached areas, such as the placement of specific attractive points (Pittarello et al., 2016; Bailey et al., 2019), might be relevant to further increase the grazing pressure and thus livestock impacts on A. viridis-encroached subalpine pastures. In conclusion, our results show that Highland cattle can graze in harsh environmental conditions and exploit A. viridis-encroached pastures. This suggests that they have a high potential to reduce A. viridis encroachment in the long-term.
Data Availability Statement
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found below: http://doi.org/10.5281/zenodo.4420882.
Ethics Statement
The animal study was reviewed and approved by Direction générale de l’agriculture, de la viticulture et des affaires vétérinaires du canton de Vaud. Written informed consent was obtained from the owners for the participation of their animals in this study.
Author Contributions
MS: data collection, formal analysis, and writing. MPi and GN: methodology, data collection, formal analysis, and writing. MKS: methodology, formal analysis, and writing. EA: supervision and writing. PM: supervision, methodology, data collection, and writing. MPr: conceptualization, funding acquisition, project administration, methodology, supervision, data collection, and writing. All authors contributed to the article and approved the submitted version
Funding
This study is part of the RobustAlps project funded by the Swiss National Science Foundation (SNF), grant no. 184925.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We are grateful to the Société d’économie alpestre de Bex, and Millasson, Dorsaz, Reggiani, and Puttalaz families for the provision of cattle and their precious help and collaboration. We would also like to thank Elisa Perotti and Davide Barberis for the help on botanical surveys and Luc Stévenin and Marin Tromme for setting up and monitoring the paddocks fencing.
References
Abegg, M., Brändli, U., Cioldi, F., Fischer, C., Herold-Bonardi, A., Huber, M., et al. (2020). Swiss National Forest Inventory NFI. Result Tables and Maps of the NFI Surveys 1983-2017 (NFI1, NFI2, NFI3, NFI4) on the Internet. Birmensdorf: Swiss Federal Research Institute WSL.
Anthelme, F., Grossi, J.-L., Brun, J.-J., and Didier, L. (2001). Consequences of green alder expansion on vegetation changes and arthropod communities removal in the northern French Alps. For. Ecol. Manag. 145, 57–65. doi: 10.1016/S0378-1127(00)00574-0
Anthelme, F., Villaret, J., and Brun, J. (2007). Shrub encroachment in the Alps gives rise to the convergence of sub-alpine communities on a regional scale. J. Veg. Sci. 18, 355–362. doi: 10.1111/j.1654-1103.2007.tb02547.x
Bailey, D. W., Mosley, J. C., Estell, R. E., Cibils, A. F., Horney, M., Hendrickson, J. R., et al. (2019). Synthesis paper: targeted livestock grazing: prescription for healthy rangelands. Rangel. Ecol. Manag. 72, 865–877. doi: 10.1016/j.rama.2019.06.003
Bühlmann, T., Körner, C., and Hiltbrunner, E. (2016). Shrub expansion of Alnus viridis drives former montane grassland into nitrogen saturation. Ecosystems 19, 968–985. doi: 10.1007/s10021-016-9979-9
Cavallero, A., Aceto, P., Gorlier, A., Lombardi, G., Lonati, M., Martinasso, B., et al. (2007). I Tipi Pastorali Delle Alpi Piemontesi [Pasture Types of the Piedmont Alps]. Ozzano dell’Emilia: Alberto Perdisa Editore Bologna.
Caviezel, C., Hunziker, M., and Kuhn, N. J. (2017). Green alder encroachment in the European Alps: the need for analyzing the spread of a native-invasive species across spatial data. CATENA 159, 149–158. doi: 10.1016/j.catena.2017.08.006
Chang, C.-R., Lee, P.-F., Bai, M.-L., and Lin, T.-T. (2004). Predicting the geographical distribution of plant communities in complex terrain – a case study in Fushian Experimental Forest, northeastern Taiwan. Ecography 27, 577–588. doi: 10.1111/j.0906-7590.2004.03852.x
Cosyns, E., Claerbout, S., Lamoot, I., and Hoffmann, M. (2005). Endozoochorous seed dispersal by cattle and horse in a spatially heterogeneous landscape. Plant Ecol. 178, 149–162. doi: 10.1007/s11258-004-2846-3
Daget, P., and Poissonet, J. (1971). Une méthode d’analyse phytologique des prairies. Ann. Agron. 22, 5–41.
Estel, S., Kuemmerle, T., Alcántara, C., Levers, C., Prishchepov, A., and Hostert, P. (2015). Mapping farmland abandonment and recultivation across Europe using MODIS NDVI time series. Rem. Sens. Environ. 163, 312–325. doi: 10.1016/j.rse.2015.03.028
Homburger, H., Lüscher, A., Scherer-Lorenzen, M., and Schneider, M. K. (2015). Patterns of livestock activity on heterogeneous subalpine pastures reveal distinct responses to spatial autocorrelation, environment and management. Mov. Ecol. 3:35. doi: 10.1186/s40462-015-0053-6
Huss-Daniel, K. (1997). Tansley Review No. 93. Actinorhizal symbioses and their N2 fixation. New Phytol. 136, 375–405. doi: 10.1046/j.1469-8137.1997.00755.x
Kohler, F., Gillet, F., Gobat, J. -M., and Buttler, A. (2004). Seasonal vegetation changes in mountain pastures due to simulated effects of cattle grazing. J. Veg. Sci. 15, 143–150. doi: 10.1111/j.1654-1103.2004.tb02249.x
Landolt, E., Bäumler, B., Ehrhardt, A., Hegg, O., Klötzli, F., Lämmler, W., et al. (2010). Flora Indicativa – Ökologische Zeigerwerte und biologische Kennzeichen zur Flora der Schweiz und der Alpen. Bern: Haupt.
Magurran, A. E. (1998). Ecological Diversity and Its Measurement. Princeton, NJ: Princeton University Press.
Manousidis, T., Kyriazopoulos, A. P., Parissi, Z. M., Abraham, E. M., Korakis, G., and Abas, Z. (2016). Grazing behavior, forage selection and diet composition of goats in a Mediterranean woody rangeland. Small Rumin. Res. 145, 142–153. doi: 10.1016/j.smallrumres.2016.11.007
Mouissie, A. (2004). Seed Dispersal by Large Herbivores: Implications for the Restoration of Plant Biodiversity. Ph.D. thesis, University of Groningen, Netherlands.
Pauler, C. M., Isselstein, J., Berard, J., Braunbeck, T., and Schneider, M. K. (2020a). Grazing allometry: anatomy, movement, and foraging behavior of three cattle breeds of different productivity. Front. Vet. Sci. 7:494. doi: 10.3389/fvets.2020.00494
Pauler, C. M., Isselstein, J., Braunbeck, T., and Schneider, M. K. (2019). Influence of Highland and production-oriented cattle breeds on pasture vegetation: a pairwise assessment across broad environmental gradients. Agric. Ecosyst. Environ. 284:106585. doi: 10.1016/j.agee.2019.106585
Pauler, C. M., Isselstein, J., Suter, M., Berard, J., Braunbeck, T., and Schneider, M. K. (2020b). Choosy grazers: influence of plant traits on forage selection by three cattle breeds. Funct. Ecol. 34, 980–992. doi: 10.1111/1365-2435.13542
Pittarello, M., Lonati, M., Gorlier, A., Perotti, E., Probo, M., and Lombardi, G. (2018). Plant diversity and pastoral value in alpine pastures are maximized at different nutrient indicator values. Ecol. Indic. 85, 518–524. doi: 10.1016/j.ecolind.2017.10.064
Pittarello, M., Probo, M., Lonati, M., Bailey, D. W., and Lombardi, G. (2016). Effects of traditional salt placement and strategically placed mineral mix supplements on cattle distribution in the Western Italian Alps. Grass Forage Sci. 71, 529–539. doi: 10.1111/gfs.12196
Probo, M., Lonati, M., Pittarello, M., Bailey, D. W., Garbarino, M., Gorlier, A., et al. (2014). Implementation of a rotational grazing system with large paddocks changes the distribution of grazing cattle in the south-western Italian Alps. Rangel. J. 36, 445–458. doi: 10.1071/RJ14043
van den Bergh, T., Körner, C., and Hiltbrunner, E. (2018). Alnus shrub expansion increases evapotranspiration in the Swiss Alps. Reg. Environ. Chang. 18, 1375–1385. doi: 10.1007/s10113-017-1246-x
Waghorn, G. (2008). Beneficial and detrimental effects of dietary condensed tannins for sustainable sheep and goat production–progress and challenges. Anim. Feed Sci. Technol. 147, 116–139. doi: 10.1016/j.anifeedsci.2007.09.013
Keywords: biodiversity, GPS tracking, grassland, robust livestock, INLA, Alps, green alder, grazing
Citation: Svensk M, Pittarello M, Nota G, Schneider MK, Allan E, Mariotte P and Probo M (2021) Spatial Distribution of Highland Cattle in Alnus viridis Encroached Subalpine Pastures. Front. Ecol. Evol. 9:626599. doi: 10.3389/fevo.2021.626599
Received: 06 November 2020; Accepted: 18 January 2021;
Published: 19 February 2021.
Edited by:
Balázs Deák, Centre for Ecological Research, Hungarian Academy of Science, HungaryReviewed by:
Victor Rolo, University of Extremadura, SpainBrigitta Erschbamer, University of Innsbruck, Austria
Copyright © 2021 Svensk, Pittarello, Nota, Schneider, Allan, Mariotte and Probo. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Mia Svensk, bWlhLnN2ZW5za0BhZ3Jvc2NvcGUuYWRtaW4uY2g=; bWlhLnN2ZW5zazJAZ21haWwuY29t