 David S Berman1
David S Berman1 Stuart S. Sumida
Stuart S. Sumida Amy C. Henrici
Amy C. Henrici- 1Section of Vertebrate Paleontology, Carnegie Museum of Natural History, Pittsburgh, PA, United States
- 2Department of Biology, California State University, San Bernardino, CA, United States
- 3Research Associate, Section of Vertebrate Paleontology, Carnegie Museum of Natural History, Pittsburgh, PA, United States
- 4Department of Biology, University of Toronto Mississauga, Mississauga, ON, Canada
A comprehensive description of the holotype skeleton is presented here for the first time of the lower Permian (Artinskian) reptile Eudibamus cursoris from the Bromacker locality of Germany since the brief description of the holotype in 2000. The holotype is essentially complete and is the only known bolosaurid represented by a well-preserved articulated skeleton. Included in the description here is a superbly preserved, partial, articulated second specimen of E. cursoris discovered at the same locality that includes a short portion of the vertebral column associated with the pelvis and right hindlimb. Descriptions of the holotype and new specimen add substantially to features of the skull and postcranium that not only confirm a bolosaurid assignment, but also add significantly to an already long list of structural features supporting an ability unique among Paleozoic vertebrates to reach relatively high bipedal and quadrupedal running speeds employing a parasagittal stride and digitigrade stance with the limbs held in a near vertical posture. Structural differences between the two specimens are restricted to the tarsi and are attributed to different ontogenetic stages of ossification, with the holotype representing a more juvenile individual, and the larger second specimen representing a more mature animal.
Introduction
Bolosauridae was first formally proposed by Cope in 1878 to include the sole representative Bolosaurus striatus Cope, 1878, which was based on several poorly preserved skulls, jaw fragments, and a few postcranial fragments, all recovered from a single lower Permian locality in Texas. However, it was long afterward that these specimens were described in detail by Watson (1954), who suggested that B. striatus represented a highly specialized member of the diadectids based on its highly unique heterodont dentition of anterior procumbent incisiform-like teeth and greatly expanded posterior molariform-like cheek teeth with bulbous crowns. In addition to the dentition, other features of the skull have undoubtedly contributed to B. striatus being assigned or considered closely related to widely divergent taxa, including Cotylosauria (Case, 1911), ‘Pelycosauria’ (Watson, 1917), and captorhinomorphs (Carroll and Gaskill, 1971). Most likely the non-dental skull features of B. striatus that influenced this taxonomic confusion included: a temporal fenestra bordered dorsally by the jugal and squamosal and ventrally by the quadratojugal; a long, narrow, boomerang-shaped postorbital that borders the posterodorsal corner of the orbit; and a high broad-based coronoid eminence of the lower jaw. With the descriptions of the nearly complete articulated holotypic skeleton (MNG 8852) of Eudibamus cursoris Berman et al. (2000), from the lower Permian of Germany (Berman et al., 2000) and more recently superbly preserved skulls of the bolosaurid Belebey vegrandis Ivakhnemko, 1973, from the middle Permian of Russia (Reisz et al., 2007), the list of unique, bolosaurid skull features was expanded to also include: palate devoid of teeth; greatly reduced transverse flange of the pterygoid that lies in the same horizontal plane as the palate; and a medial ventrally expanded flange of the alveolar shelf of the dentary that nearly reaches the level of the ventral margin of the mandible and is extensively covered medially by a long anterior extension of the prearticular. A highly unique dentition has not only allowed unmistakable identification of isolated jaw elements as indeterminate bolosaurid, but also reference either previously described or new species.
Although bolosaurid occurrences are relatively rare, they have greatly expanded geographic and temporal histories. To date, three genera comprising seven species of Bolosauridae have been described: Bolosaurus striatus from the lower Permian of Texas (Watson, 1954) and New Mexico (Lucas et al., 2005); Eudibamus cursoris from the lower Permian of Germany (Berman et al., 2000); Bolosaurus grandis Reisz et al. (2002), from the lower Permian of Oklahoma (Reisz et al., 2002); Belebey vegrandis Ivakhnemko (1973), and Belebey maxima Ivakhnemko and Tverdochlebova (1987), from the middle Permian of Russia (Ivakhnemko, 1973; Ivakhnemko and Tverdochlebova, 1987; Reisz et al., 2007); Belebey chengi Müller et al., 2008, from the middle Permian of China (Müller et al., 2008) and Belebey augustodunensis Falconnet (2012), from the Permo-Pennsylvanian of France (Falconnet, 2012). Additionally, isolated bolosaurid-like teeth have been described from the upper Pennsylvanian of New Mexico (Harris et al., 2004).
Eudibamus cursoris is a member of a diverse tetrapod assemblage, which to date consists of twelve species, ranging from fully terrestrial to semi-aquatic (Berman et al., 2020). All were collected from the lower Permian Tambach Formation of the Bromacker locality, an area of small, abandoned, sandstone quarries scattered over an area of less than 0.5 km2 in the midregion of the Thuringian Forest, approximately 1.5 km north of the village of Tambach-Dietharz and 20 km south of the town of Gotha, central Germany. Outcrops of the Tambach Formation are restricted to an area of about 50 km2 that were deposited in a small, internally drained, upland basin termed the Tambach Basin with an original aerial extent of approximately 200–300 km2. All but one of the Bromacker species were collected from an extensively quarried site, the Bromacker quarry, centrally located in the Tambach Basin. Detailed stratigraphic, sedimentological, and paleogeographic data of the Bromacker locality and quarry are summarized by Eberth et al. (2000).
Presented here is the first comprehensive description of the essentially complete, articulated skeleton of the holotype of Eudibamus cursoris (Figure 1) since its brief original description (Berman et al., 2000). It is of special importance in being the only known bolosaurid specimen described to date that includes a well-preserved postcranial skeleton. Most significantly, its postcranial anatomy documents the earliest occurrence of a vertebrate capable of employing a cursorial quadrupedal or bipedal digitigrade locomotion with the limbs held in a vertical erect posture and swung pendulum-like in a parasagittal plane. Otherwise, it is not until 60 or more million years later with the appearance of the Late Triassic archosaurs that this type of cursorial gait is again attributable to any group of terrestrial tetrapods. Although several very plausible examples of Permian bipedal reptiles have been described (Carroll, 1973; Carroll and Thomson, 1982; Spindler et al., 2019), in none has this ability been coupled with evidence of both limbs occupying a vertical parasagittal plane and swung in a fore and aft pendulum-like style during locomotion.
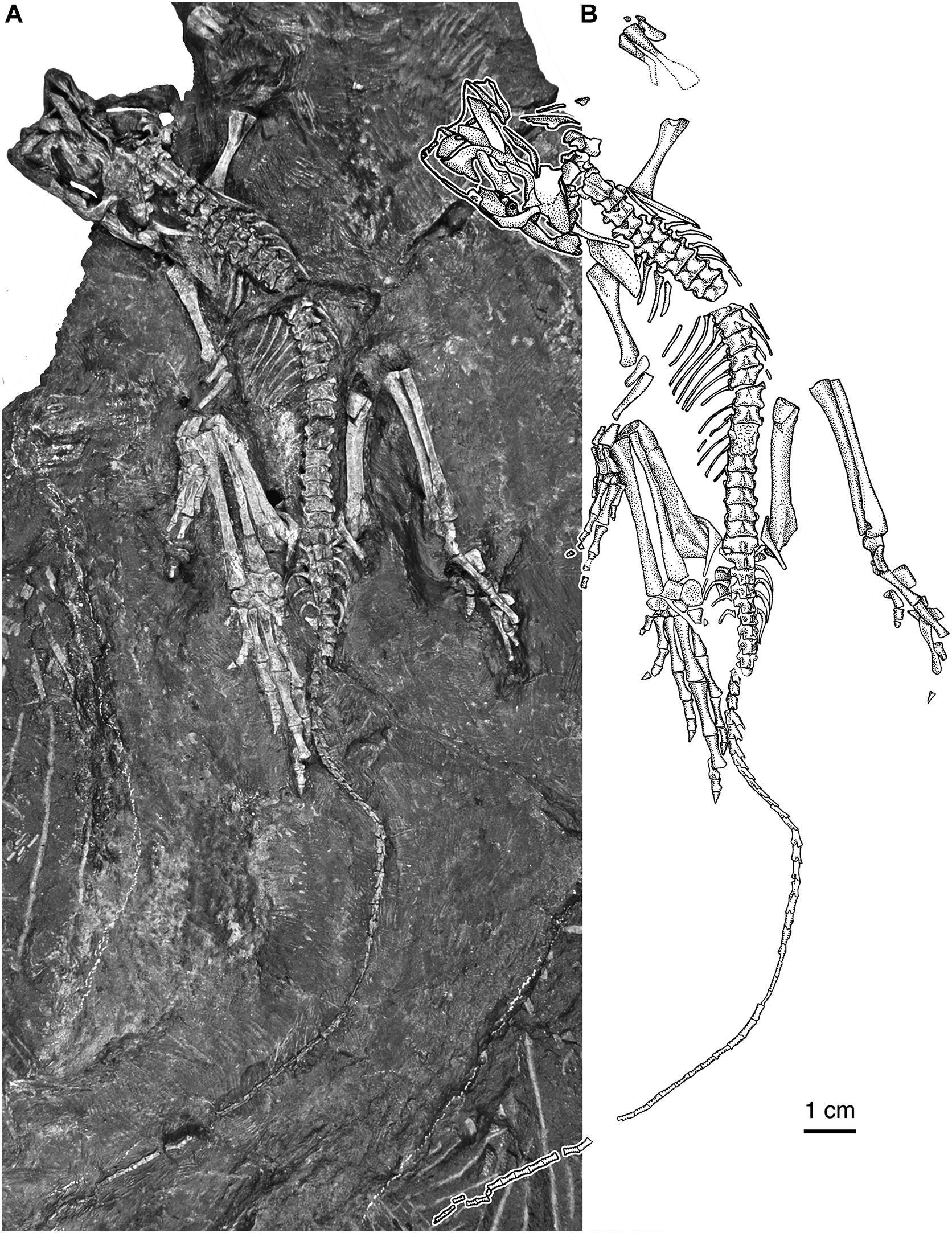
Figure 1. Eudibamus cursoris, holotype (MNG 8852). Essentially complete skeleton exposed mainly in dorsal view. (A) Photograph and (B) illustration (modified after Berman et al., 2000).
From the same Bromacker locality as the holotype a second well-preserved, partial articulated skeleton of Eudibamus cursoris (MNG 12895) was subsequently discovered. It consists of an estimated articulated string of eleven vertebrae extending approximately from the posteriormost four presacrals to the anteriormost three caudals, along with the pelvis and right hindlimb with pes (Figure 2) that adds significantly to the morphology of the holotype of E. cursoris.
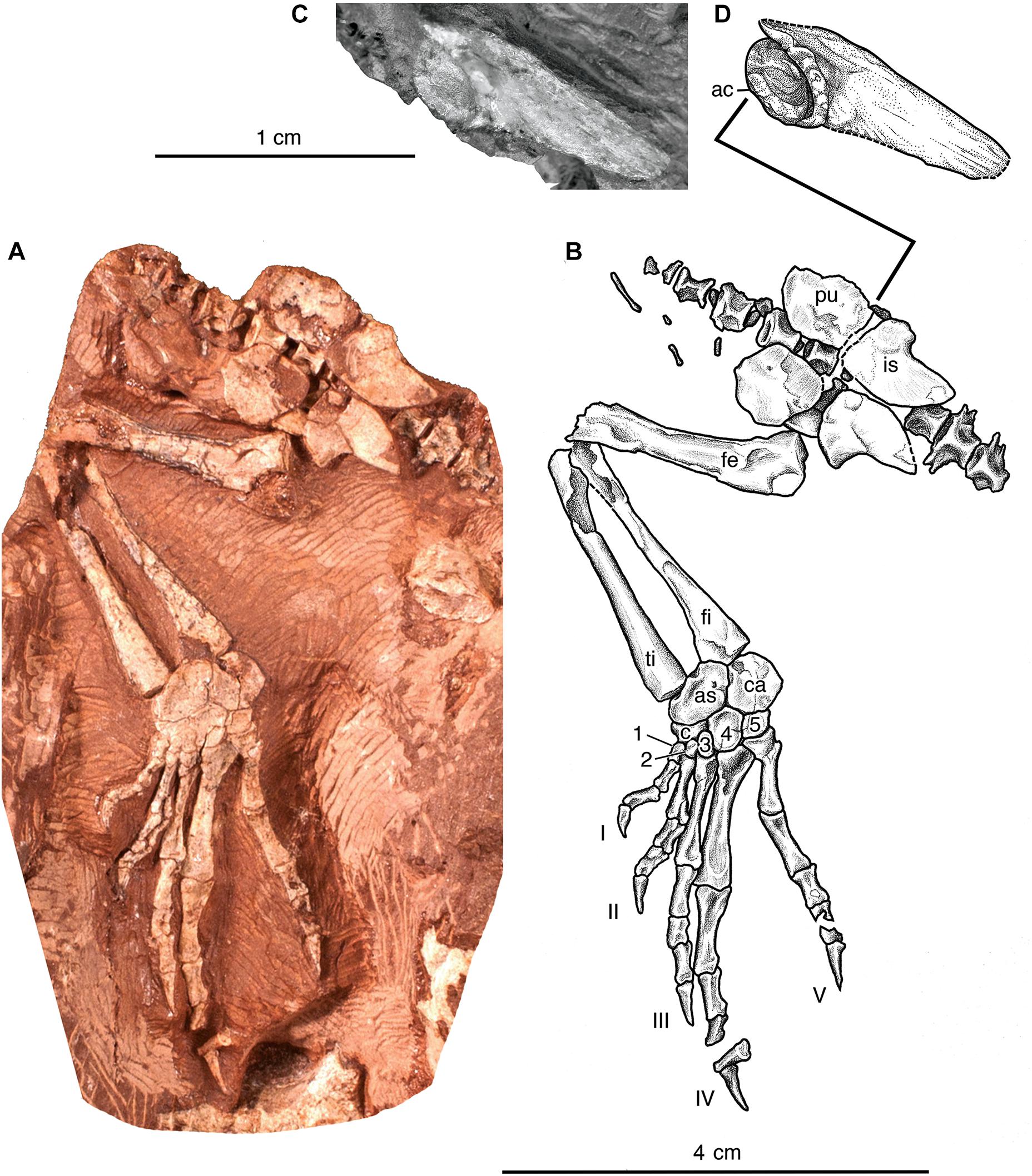
Figure 2. Eudibamus cursoris, referred specimen (MNG 12895). (A) Photograph and (B) illustration of entire articulated specimen in ventral view, consisting of partial vertebral column spanning from approximately posteriormost four presacral vertebrae to the anteriormost three caudal vertebrae, pelvis, and right hindlimb. (C) Photograph and (D) illustration of partial left ilium shown isolated in lateral view.
In the original description of Eudibamus cursoris it was recognized for the first time by Reisz that bolosaurids are not only parareptiles, but also the basalmost and oldest member of the Bolosauridae clade (Reisz et al., 2007; Müller et al., 2008). The phylogenetic analysis by Berman et al. (2000) that posits E. cursoris as the oldest known member of the Parareptilia has been challenged by two recent descriptions of Carboniferous genera. On the basis of a single, nearly complete articulated skeleton from the latest Carboniferous of Prince Edward Island, Canada, Modesto et al. (2015), described a new genus and species of parareptile, Erpetonyx arsenaultorum Modesto et al., 2015. In the phylogenetic analysis of Parareptilia presented by Modesto et al. (2015) E. arsenaultorum is recognized as the sister taxon and closest relative of Bolosauridae and together they comprise Bolosauria. Bolosauria was also retrieved in a more recent analysis of Parareptilia by Cisneros et al. (2021), and Eudibamus remained as the sister taxon to Belebey and Bolosaurus, as in Müller et al. (2008). Notably, their phylogenetic analysis included data for Belebey based on B. vegrandis only, and not the other two species. The second parareptile described from the Carboniferous, Carbonodraco lundi Mann et al., 2019, was diagnosed as a cleistorhinid parareptile, on the basis of two specimens consisting of disarticulated cranial elements from the middle Pennsylvanian Linton, Ohio (Mann et al., 2019), making it the oldest known member of Parareptilia currently known. The description presented here emphasizes those structures of Eudibamus cursoris that supports its specialized cursorial ability and envisioned as an agile herbivore (Hotton et al., 1997) that relied on a unique ability to employ bipedal and quadrupedal gaits presumably to speedily escape predators.
Abbreviations
The acronym MNG refers to the Museum der Natur, Gotha, Germany.
Key to abbreviations in figures: ac, acetabulum; an, angular; ar, articular; as, astragalus; atl, atlantal elements; ana, axial neural arch; c, centrale; ca, calcaneum; co, coracoid plate (co-ossified anterior and posterior coracoids); cd, coronoid; cl, clavicle; cth, cleithrum; d, dentary; eo, exoccipital; f, frontal; fe, femur; fi, fibula; h, humerus; i, intermedium; ic, interclavicle; il, ilium; is, ischium; j, jugal; l, lacrimal; lc, lateral centrale; m, maxilla; mf, mandibular foramen; mc, medial centrale; n, nasal; o, orbital rim; op, opisthotic; p, parietal; pal, palatine; pf, postfrontal; pi, pineal foramen; pm, premaxilla; pra, prearticular; prf, prefrontal; ps, parasphenoid; pt, pterygoid; pu, pubis; qj, quadratojugal; r, radius; ra, radiale; sa, surangular; sc, scapula; so, supraoccipital; sp, splenial; sq, squamosal; t, tabular; ti, tibia; u, ulna; ul, ulnare; I-V, digits; 1-5, distal carpals and tarsals; ∗, isolated tooth.
Description
Skull Roof
Description of the skull of Eudibamus cursoris is based on its dorsal, ventral, and lateral exposures in Figures 1, 3, 4. Despite almost all the skull bones being either disarticulated, partially overlapped by neighboring bones, or marginally incomplete, in most cases they can be described as they would have appeared in the articulated skull. Unfortunately, numerous critical structural uncertainties prevent a confident reconstruction of the skull roof. Although the skull is crushed severely dorsoventrally, and the skull roof is incompletely preserved, with a single exception all the paired standard roofing bones are represented by one or both exposed in external view. As in Belebey vegrandis (Reisz et al., 2007), E. cursoris does not appear to possess a supratemporal, which was incorrectly illustrated as present in B. striatus by Watson (1954) as interposed between the parietal and squamosal.
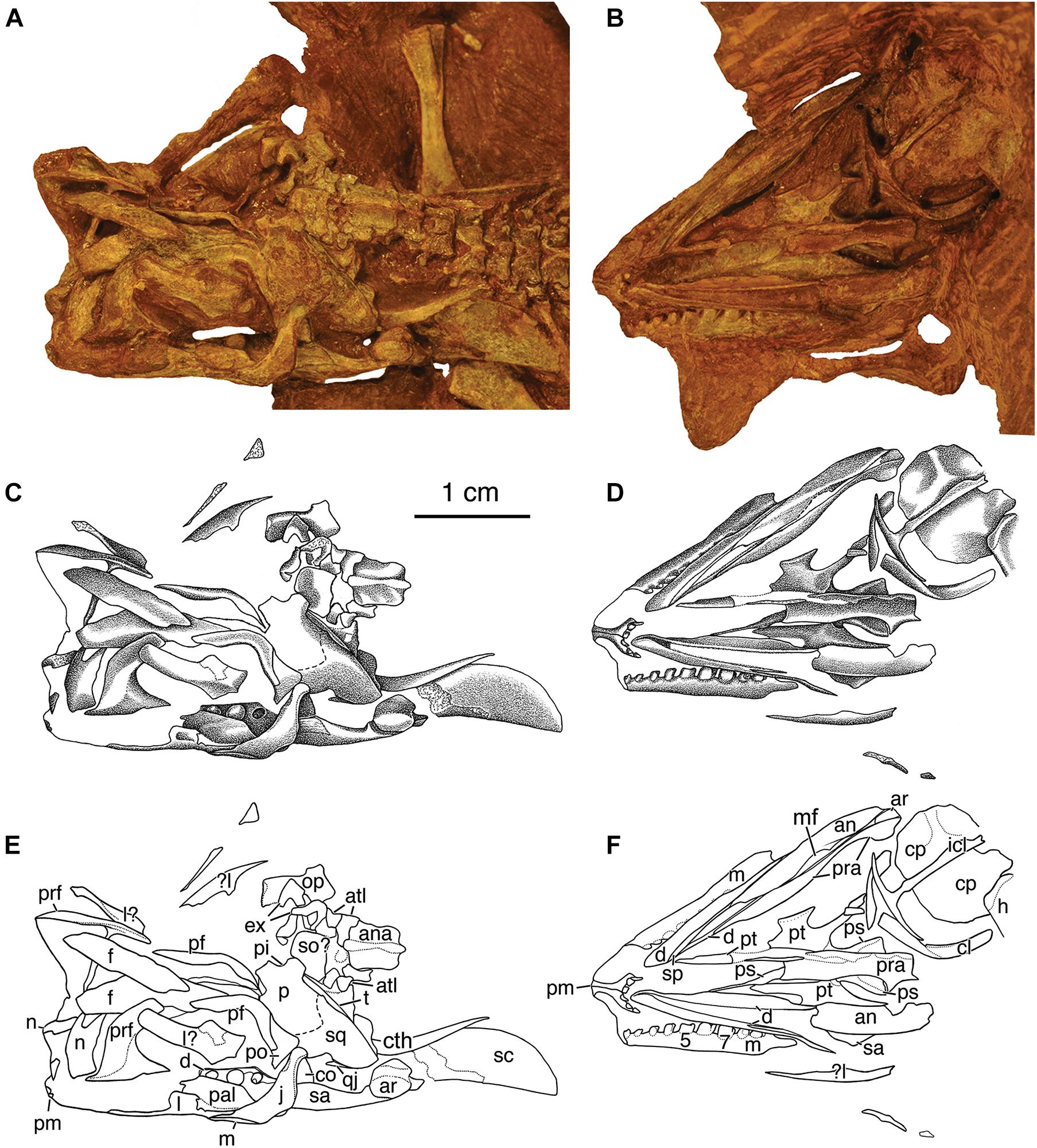
Figure 3. Skull of Eudibamus cursoris, holotype (MNG 8852). (A,B) Photographs, (C,D) illustrations, and (E,F) labeled outline drawings of elements of dorsoventrally crushed skull in (A,C,E), dorsal and (B,D,F), ventral views. Dorsal views of skull (A,C,E) also include disarticulated elements of the braincase and atlas-axis complex, dorsal portion of laterally exposed left scapula and cleithrum, and right humerus. Ventral views of skull (B,D,F) also include disarticulated elements of the dorsal portions of laterally exposed left scapula and cleithrum and partially exposed ventral view of shoulder girdle. Abbreviations in this and following figures as listed in text.
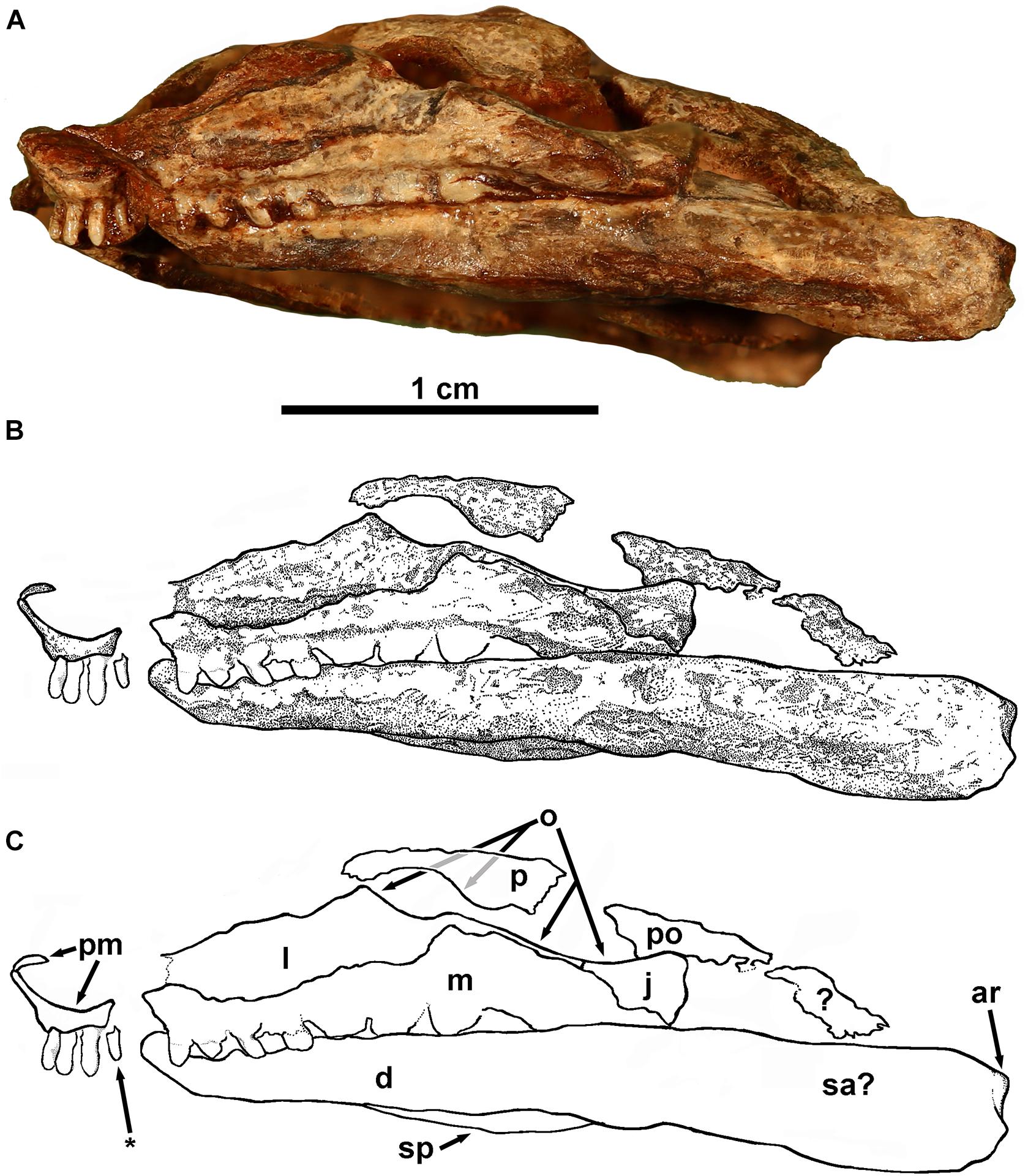
Figure 4. Eudibamus cursoris, holotype (MNG 8852). Partial left lateral view of skull. (A) Photograph, (B) illustration, and (C) labeled outlines drawings of elements. *Indicates visible tooth of right premaxilla.
The premaxillae are complete and united along the midline. They are relatively small slender elements, consisting of the two recognized standard components, an anterior dorsal process and a ventral posteriorly directed tooth bearing process. Together they form a deeply concave anteroventral margin of the external naris. The dorsal processes are slightly inclined anterodorsally and distally barely reach the skull roof, where they presumably inserted between the anterior ends of the paired nasals. In ventral view of the skull the ventral processes curve posterolaterally from their midline contact, ending immediately posterior to the dentition without evidence of a distinct maxillary process. The premaxillae each possess three tightly spaced teeth with both series meeting at the midline without a gap. The teeth are narrowly cone-shaped, decrease serially in size slightly posteriorly from a maximum diameter of about 0.3 mm, and are oriented vertically, rather than being procumbent as in Bolosaurus striatus (Watson, 1954) and B. vegrandis (Reisz et al., 2007).
Both maxillae are exposed in the ventral exposure of the skull with the left also exposed in the lateral view of the skull. The disarticulated right maxilla is exposed laterally and missing only a small portion posterior to the dental series, whereas the left is complete and not only narrowly exposed ventrolaterally, but also fully exposed in the lateral view of the skull. In the latter view it is articulated with the lacrimal and jugal, but narrowly separated from contacting the distal end of the ventral process of the premaxilla due to marginally incomplete preservation. The maxilla is greatly elongated with its full length contributing to the ventral margin of the skull from the distal end of the ventral process of the premaxilla to the level of the posterior orbital margin. The anterior two-thirds of the maxilla form a narrow elongate rectangular process-like structure that contacts the ventral margin of the lacrimal. The shorter expanded posterior portion of the maxilla narrows as it curves posteroventrally in contact with the anteroventral margin of the jugal to end in a blunt point well below the level of the alveolar shelf. The posterior portion of the maxilla dorsally overlaps the lateral surface of the lacrimal except for an extremely thin suborbital exposure that posteriorly contacts an equally narrow but much shorter anterior suborbital process of the jugal. Thus, the maxilla is narrowly excluded from the orbit by a lacrimal-jugal contact. There is no evidence of an anterior premaxillary process of the maxilla, and the spacing of the teeth at the union of the premaxillary and maxillary dentitions is the same as elsewhere in the series.
The dentition of the right maxilla is well-exposed laterally, extends posteriorly to the midlength level of the orbit, and is complete with ten teeth. The teeth gradually increase serially in size posteriorly to the seventh tooth, with the first two teeth being slightly larger than those of the premaxilla and the last three teeth being markedly reduced to about the size of the third and fourth. Although exposures of the teeth are limited mainly to the lateral surfaces, they exhibit a rudimentary development of the distinctive molariform features of other bolosaurids, particularly the larger posterior teeth. The teeth are bulbous and expanded anteroposteriorly, and the crown is divided unequally into distinct anterior and posterior portions. The much larger anterior portion of the crown has the form of a heel-like structure with a very slightly shallow basin-like dorsal surface, whereas the much smaller posterior portion of the crown rises dorsally into a very small, pointed cusp. The posterior surface of the cusp is an extension of the rounded tooth base, whereas the anterior surface is sharply defined by a flattened or possibly a slightly vertically concave surface, giving the cusp a half dome-like appearance.
Only the left jugal is preserved, but it is complete and fully exposed in the dorsal view of the skull, where it borders the concave posteroventral orbital margin between contacts with the postorbital dorsally and the maxilla ventrally. The midlength portion of the jugal is broadly expanded posteroventrally with a slightly concave anteroventral margin. The narrow dorsal extension of the jugal is posteriorly inclined and inserts between ventrally diverging margins of the postorbital and squamosal. Presumably a short dorsal portion of the slightly concave posterior margin of the dorsal extension enclosed the anterior margin of the temporal fenestra.
Description of the lacrimal relies on the left bone exposed in the lateral view of the skull, which is complete except for a very small loss at its narrow anterior end that undoubtedly contributed to the posterior margin of the naris. A small triangular dorsal process of the lacrimal forming the anteroventral margin of the orbit roughly divides the lacrimal into subequal posterior and anterior portions. Anterior to the dorsal process the lacrimal has an elongate subrectangular outline that narrows slightly anteriorly. Its dorsal margin is believed to have been contacted by the incomplete disarticulated ventral margin of the prefrontal.
All that remains of the right nasal is a small remnant, but the left is fully exposed and appears complete, but undoubtedly has rotated anteroventrally about 90o from its contact with the anterior margin of the frontal. Correctly oriented it has roughly the outline of an anteroposteriorly elongated rectangle, but with an unusually short length equal to about 40% of the frontal length, suggesting an abbreviated snout. A deep V-shaped cleft in the posterior margin of the left nasal presumably received a complementary pointed anterior end of a more completely preserved frontal than those described here. The anterior third of the nasal is angled slightly anteroventrally from the midline, presumably to accommodate the dorsal process of the premaxilla. The vertically truncated anterior margin likely contributed to the posterodorsal margin of the external naris.
Both frontals are present but disarticulated along their midline contact, and only the left bone appears mostly complete. It is greatly elongate anteroposteriorly, perhaps equaling more than 50% of the midline length of the skull. The left frontal preserves a short poorly defined lateral orbital process that intervenes between the prefrontal and postfrontal, dividing the frontal into anterior and posterior processes, with the anterior process being about twice as long.
Only the left parietal can be accounted for, which due to dorsoventral crushing of the skull roof has resulted in it occupying a single plane, rather than originally being folded longitudinally into a dorsal skull table exposure and a ventral lateral cheek exposure. Although the anterior margin of the skull table portion is incomplete, the midline length of the parietal was obviously relatively short, particularly when compared to that of the frontal, which appears to be about three times as long. The midline margin of the parietal is interrupted by what appears to be the lateral half of a moderate-sized pineal foramen. Together the ventral lateral cheek exposure of the parietal and the squamosal form a very broad, vertically elongate rectangular plate that is slightly bowed laterally and directed strongly posteroventrally well beyond the level of the posterior margin of the skull table. The contact between the parietal and squamosal can be only tentatively placed. The anteroventral margin of the united pair is contacted by a posteroventral succession of the postfrontal, postorbital, and jugal bones, which border the posterior half of the concave orbital margin.
The squamosal extends ventrally to nearly the ventral margin of the skull roof, where it would have laterally overlapped the articular condyle of the quadrate. The anterior ventral margin of the squamosal is incised dorsally by a shallow step-wise emargination that bordered the posterodorsal corner of the temporal fenestra. A short distance ventral from the skull midline the dorsal half of the posterior margin of the united parietal-squamosal is narrowly recessed and inflected medially as an occipital flange that is overlapped by the tabular.
All that remains of the quadratojugals is a small, pointed remnant of the right bone projecting anteriorly from beneath the posteroventral corner of the ventral expansion of the squamosal. Presumably the complete quadratojugal would have extended anteriorly as a long narrow bar that ventrally bordered a narrowly elongated temporal fenestra and contacted the posterior margin of the jugal.
Both prefrontals are preserved, but exposed in contrasting views that confusingly appear to portray radically different morphologies. The laterally exposed left prefrontal appears to be essentially complete and preserved in proper orientation within the skull and relative to surrounding elements. Ventrally it consists of a narrow longitudinally elongated base with a horizontally straight ventral margin, which likely contacted the dorsal margin of the anterior half of the ventrally displaced lacrimal. Extending dorsally from the base is a dorsal blade that distally curves posterodorsally and apparently is disarticulated from the lateral margin of the anterior process of the frontal. The posterior margin of the prefrontal is deeply concave to form the anterior orbital margin. The right prefrontal also appears complete and, although narrowly disarticulated, occupies its near correct relationship with the frontal but with a ventrolateral rotation of about 90o that exposes it in internal view as T-shaped with a long thin ventral stem. The horizontal dorsal bar of the T is narrowly exposed dorsally and is divided by the stem into posterior and anterior processes of unequal lengths. The medial margin of the shorter narrower posterior process is narrowly separated from its contact with the lateral margin of the anterior process of the frontal, whereas the medial margin of the longer slightly wider anterior process undoubtedly contacted the lateral margin of the nasal. The ventral stem is believed to have extended along the anterior orbital margin in contact with the posterior margin of the lacrimal.
The postfrontals are complete but only the left is exposed fully and retains its marginal contacts. It has a long narrow boomerang shape that wraps around the concave posterodorsal margin of the orbit as it extends from the dorsal skull table to the lateral temporal area of the skull roof. Successive contacts along its non-orbital margin include the lateral margin of the posterior process of the frontal, a short midlength portion of the anterior margin of the parietal, and the greater dorsal portion of the anterior margin of postorbital.
Of the postorbital, the smallest of the skull roofing bones, only the left can be accounted for but appears complete. It has an outline of a broad-based triangle with the longer, vertical anterior base contributing to the postorbital margin, although greatly restricted due to the narrow ventral extension of the postfrontal contacting most of the dorsal portion of its anterior margin. The posteriorly converging margins of its triangular outline extend between contacts with the united parietal-squamosal dorsally and ventrally the jugal. The arrangement of the orbital bones suggests that the orbit had a longitudinally expanded oval outline.
Palate
Of the dermal palatal bones only the pterygoids are preserved (Figure 3), but neither is fully exposed or complete. The well-exposed left transverse flange exhibits several differences from the standard morphology of the majority of Paleozoic amniotes (Reisz et al., 2007). It is exceptionally small and subrectangular in outline with the longitudinal length being slightly greater than the transverse width. The flange is constricted anteroposteriorly by concave margins, and the deeper posterior concavity likely received the posterior end of the ectopterygoid as in Belebey vegrandis (Reisz et al., 2007). The ventral surface of the flange is flat, and occupies the same horizontal plane as that of the pterygoid proper, and, although devoid of teeth, there is a low narrow ridge bordering the lateral distal margin. Only the ventral margin of the palatal ramus of the left pterygoid is exposed, and, although possibly incomplete distally, its anterior projection is close to the midline, suggesting a very narrow interpterygoid vacuity. The flange-like quadrate process of the pterygoid extends posterolaterally to the level of the posterior margin of the parasphenoid. As preserved in the right process the medial surface is slightly concave with a narrowly thickened rounded ventral margin that extends the full length of the process. The disarticulated right prearticular obscures a ventral exposure of both basicranial articulations. A partially exposed basal process of the left pterygoid, however, indicates a posteromedially facing line of contact with the basipterygoid process of the braincase. Although tightly articulated, at least some mobility between the palate and braincase may have been possible.
Braincase
The only definitely identifiable element of the braincase is the parasphenoid, although disarticulated elements are tentatively identified as the supraoccipital, opisthotic, and exoccipitals (Figure 3). The parasphenoid, which presumably is indistinguishably fused dorsally with the basisphenoid, is complete, but the right prearticular obscures a wide area along its midline. Immediately posterior to the basipterygoid processes the parasphenoid expands bilaterally into a broad triangular plate with the lateral margins thickened into smooth ventral ridges of the cristae ventrolaterales. What may be the cultriform process is a poorly preserved thin anteriorly narrowing bone that extends far anteriorly from dorsally above the anterior broken margin of the prearticular. A short distance medial to the left parietal are two small disarticulated poorly preserved bones that have the U-shaped outline shape expected of the exoccipitals with their internal margins presumably forming the lateral margins of the foramen magnum. Adjacent to the exoccipitals is an isolated element with the outline and size expected of an opisthotic, believed to be the right. It consists of two rectangular portions joined at a right angle. The smaller of the two portions would have contributed to the braincase wall, whereas the longer portion, which expands slightly distally, would be the paroccipital process. A rectangular plate-like element that is relatively large compared to the surrounding elements and adjacent to the anteroposterior corner of the parietal is suggestive of the supraoccipital.
Mandible
The left mandible is complete and exposed in both the ventral and dorsal exposures of the skull (Figure 3). The entire length of the ventral surface of the mandible is preserved with all the elements tightly articulated. The few preserved bones of the right mandible are widely disarticulated, mostly fragmentary, and variously exposed. Fortunately, the ventrally exposed right dentary clearly indicates the unique bolosaurid feature of the Meckelian canal being bordered by narrowly separated medial and lateral flanges. Unfortunately, however, what are not visible are the flanges originating as ventral extensions of the medial and lateral margins of the alveolar shelf. Presumably, if fully exposed the flanges would extend well below the alveolar shelf, giving the entire structure an inverted U-shaped appearance in transverse section with the Meckelian canal opening ventrally (Reisz et al., 2002, 2007). All that is visible of the medial flange of the dentary is a narrow ventral margin that extends from a short distance posterior to the symphysis to about the midlength of the mandible. The anterior half of the flange margin exposure is bordered ventrally by a moderate dorsal expansion of the prearticular, suggesting that it may have extensively overlapped the medial surface of the medial flange of the dentary, a feature unique to bolosaurids (Reisz et al., 2002).
In the laterally exposed articulated left mandible an extremely thin posteriorly narrowing splint-like splenial contacts approximately the anterior half of the ventral margin of the dentary. Anteriorly it is marginally excluded from the symphysis, whereas posteriorly it contacts the anterior dorsal margin of the angular. Unique to bolosaurids and as described in Belebey vegrandis (Reisz et al., 2007) and inferred in Bolosaurus grandis (Reisz et al., 2002) the splenial ventrally floored the Meckelian canal. There is no observable evidence to confirm this feature in Eudibamus.
Exposure of the dentary dentition is limited to the posteriormost three positions dorsally exposed at the posterior end of the left dentary. The second tooth is greatly enlarged over the anterior tooth and presumably the posterior tooth is represented by an empty socket. Features of the crowns are too indistinct to describe accept to say that they are bulbous with a circular cross-sectional base.
The only extensive view of the surangular is a dorsolaterally exposed posterior portion of the left bone that includes the upper portion of the lateral wall of the adductor fossa with a slightly concave dorsal margin. Immediately posterior to the adductor fossa rim the surangular expands slightly into a concave articulation with the upper surface of the articular. Anteriorly its contact with the posterior margin of the dentary is obscured by the overlying left jugal. At the anterior border of the adductor fossa the dorsal margin of the surangular contacts the ventral margin of the dorsally directed coronoid process. Although a narrow border of its anterior margin is obscured by the left jugal, it otherwise appears to be free of contacts and to narrow dorsally to a pointed apex of a cone-shaped structure.
Exposures of the angular, prearticular, and articular are limited essentially to the ventral surface of the left mandible. Here the angular is greatly elongated anteriorly from the jaw joint to about 60% of the ventral midline length of the mandible before ending anteriorly in what appears to be an irregular broken margin. Presumably, if continued anteriorly it would have occupied an anteriorly narrowing space that reaches to within a very short distance of the jaw symphysis. As such, the angular would have been bordered laterally by the splenial and medially by the ventral medial flange of the dentary that medially borders the Meckelian canal. The posterior half of the angular is folded longitudinally with the dorsal half consisting of a wide vertical expansion that curves dorsally to form the ventral portion of the lateral wall of the adductor fossa (not exposed). As preserved the anterior half of the angular consists of a broad ventrally exposed expansion that extends between the ventral margins of the lateral and medial flanges of the dentary to floor the Meckalian canal. For most of its posterior length the angular medially contacts the ventral margin of the prearticular in nearly a straight margin except for two small openings. Posteriorly the two bones narrowly diverge to expose the ventral surface of the articular, and a short distance anteriorly there is narrow opening described in Belebey vegrandis by Reisz et al. (2007) as the mandibular foramen. The angular borders the laterally concave margin of the mandibular foramen, whereas the prearticular forms a straight medial margin.
The prearticular is greatly elongated and narrows as it extends anteriorly in a straight ventral medial margin from the jaw articulation to about three fourths the length of the mandible. The shorter dorsally exposed posterior portion forms the medial margin of the adductor fossa with a slightly concave dorsal margin. It ends posteriorly in a low dorsal process that thickens slightly posteriorly as it cups the convex medial surface of the articular. The dorsal surface of the articular is well-exposed and divided into longitudinally elongated lateral and medial oval condyles with the former being much larger and more strongly developed. The articular does not possess a retroarticular process.
Axial Skeleton
Description of the axial skeleton is based almost entirely on the well-preserved, essentially complete, and mostly articulated column of the holotype MNG 8852 (Figures 1, 3) that includes twenty-six presacrals, two sacrals, and fifty-five caudals. The referred specimen MNG 12895 (Figure 2) includes an estimated string of eleven vertebrae, although approximately three are obscured by the pelvis. Otherwise, the series is believed to include the posteriormost four presacrals or their remnants, followed likely by two sacrals, a gap that held about three caudals, and three additional caudals.
Nearly the entire precaudal vertebral series in the holotype is well- preserved and exposed dorsally except those of the cervical region, including elements of the atlas-axis complex (Figure 3). The atlas is represented by identical bilateral series of three extremely small, loosely associated elements directed anteriorly from adjacent to the anterior margin of the axial neural arch. They presumably represent a bipartite atlas, but cannot be individually identified. The axial neural arch has a longitudinally elongated rectangular outline in dorsal view, the length of which is nearly twice those of the postcervical dorsals. A low ridge-like neural spine extends the full length of the arch. The postaxial presacral neural arches are greatly expanded laterally and slightly swollen dorsally. The zygapophyses are widely spaced from the midline and appear to articulate in a horizontal plane. Small neural spines have a longitudinally elongate oval outline in dorsal view and are positioned close to the posterior margin of the neural arch. The postaxial presacral transverse processes are most prominently exposed in the cervical region, where they have a distally short rectangular outline and are directed anterolaterally from a level immediately posterior to the anterior zygapophyses. The neural arch of the first sacral is like that of the dorsals except for the transverse process being greatly expanded in width, slightly more so at its distal articulation with the ilium, and indistinguishably fused to the centrum. The neural arch of the second sacral is greatly reduced compared to the first, and the transverse processes, although not well exposed, are extremely narrow and project anterolaterally; it cannot be determined whether they independently contacted the ilium or provided support to that of the first sacral.
In the ventrally exposed partial column of the referred specimen MNG 12895 the ventral surfaces of the dorsal and caudal centra are strongly pinched bilaterally to produce a midline keel that is rounded in transverse section and widens slightly at either end to merge with the rounded articular lips of the centrum. Well-developed transverse processes of the caudals project far laterally and bifurcate into prominent processes presumably for the divided head of the rib. Remnants of intercentra are associated with the dorsal vertebrae.
The first six pairs of cervical ribs in the holotype are either lost or not exposed; otherwise the ribs extend without break to the twenty-second presacral vertebra. All are unusually short, extremely slender, and only modestly curved ventrally. The varying shapes of most of the rib heads are difficult to discern except for those of the cervical region, which are narrowly separated from the centrum and along with the shafts, have been rotated slightly posterodorsally. The heads clearly exhibit a triangular outline, and the capitulum and tuberculum areas are well defined along the fore and aft margins by thickened rounded margins separated by a shallow channel. The longer anterior process-like capitulum is directed ventromedially to contact presumably the ventral portion of the anterior margin of the centrum, whereas the tuberculum is a near continuation of the shaft that contacted the transverse process. The rib heads exhibit a gradual serial reduction in size posteriorly, whereas the rib shafts gradually increase serially in length posteriorly to about sixteenth vertebrae, followed by a marked decrease in length and slight decrease in width to the end of the series.
The holotype possesses two pairs of sacral ribs. The first sacral rib is broad and extends straight laterally to the anterior part of the iliac blade. The second rib is quite slender and extends anterolaterally to the iliac blade, and its position and orientation suggests that it made contact with the iliac blade immediately posterior to the first sacral rib. The first four vertebrae of the tail possess ribs that are narrowly separated from the transverse processes. The rib heads are short, flat, and triangular in dorsal outline and their possible division into capitular and tubercular processes cannot be discerned. The shafts on the right side are complete, subequal in length, and strongly curved posteriorly. A fragment of the fifth caudal rib is preserved on the right side.
The tail of the holotype is essentially complete and except for the possible loss or incompleteness of the last few vertebrae is exceptionally long with a calculated length equal to 64.3% of the full axial length (Berman et al., 2000) and, yet, it includes only the approximate standard number of about 55 or more vertebrae. Approximately the first eight caudal vertebrae are exposed dorsally and are identical to the dorsals except for being noticeably smaller and exhibiting a serial reduction in size posteriorly. Continuing posteriorly the caudal neural arches are serially reduced in size whereas in contrast the centra uniquely persist as relatively elongate structures of subequal lengths, although with a slight length reduction of the last six or more centra. The relative lengthening of the caudals accounts in great part for the extraordinary length of the tail in Eudibamus, although there is a slight length reduction of the last six or more caudal centra.
The neural arches of the caudals exhibit several, marked, structural changes from about the ninth to the thirtieth serial position: (1) they gradually become reduced in size to little or no evidence of their presence; (2) the neural spines are spike-like and project strongly posterodorsally; (3) the pre- and postzygapophyses become greatly narrowed in width and barely separated from one another along the midline with the articular facets steeply tilted ventromedially; and (4) despite the centra being nearly equal in length, the transverse dimensions of the neural arches become reduced. In the remainder of the tail the vertebrae consist of simple rod-shaped centrum elements with slightly concave dorsal and ventral surfaces.
Shoulder Girdle
The holotypic shoulder girdle is well represented by possessing all the standard bones, most of which are well exposed but narrowly disarticulated (Figures 1, 3, 5, 6). The scapulae are probably nearly complete but only partially exposed in the dorsal view of the skull. Exposure of the right blade is limited to its anterior edge, whereas the greater dorsal portion of the left is laterally exposed but ends ventrally in a broken margin. From a thin broadly convex anterior margin the blade thickens toward a straight posterodorsally inclined posterior margin. Closely associated with both scapular blades is a splint-like cleithrum, which tapers to a sharp point as it ascends the ventral half of the anterior margin of the blade.
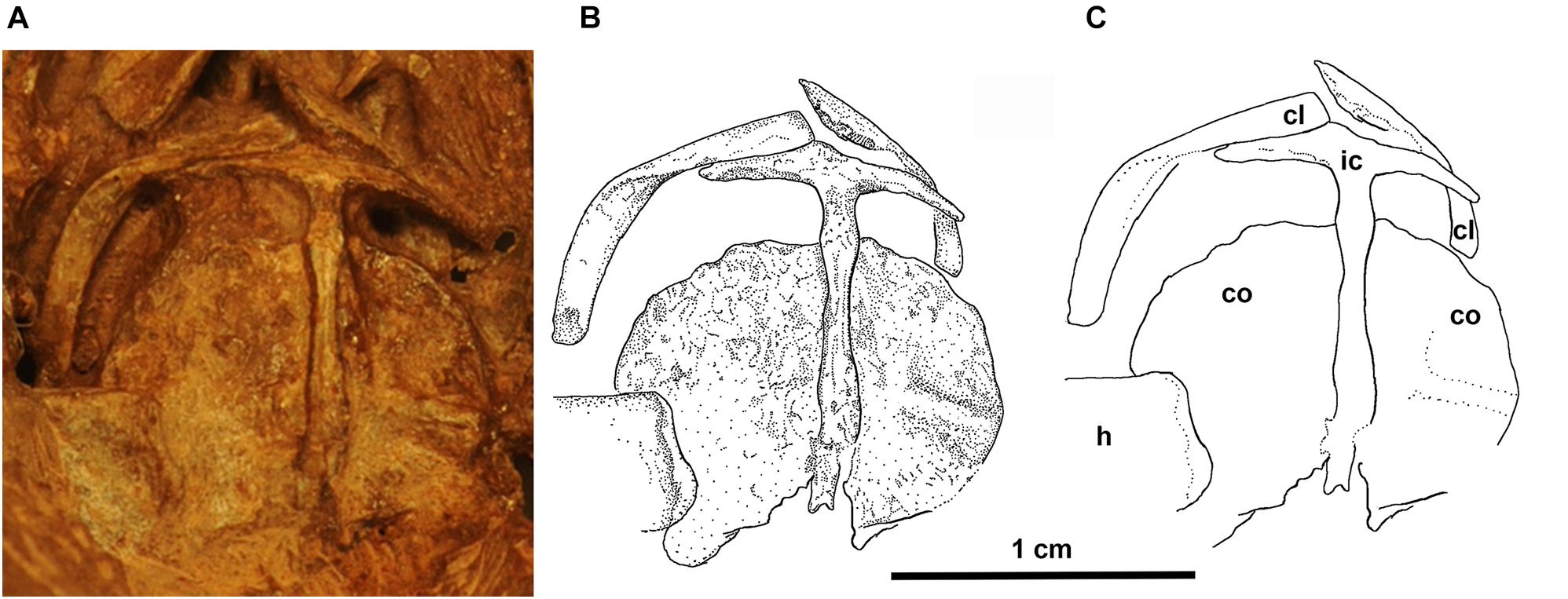
Figure 5. Eudibamus cursoris, holotype (MNG 8852). (A) Photograph, (B) illustration, and (C) outlines of individual elements of shoulder girdle in ventral view.
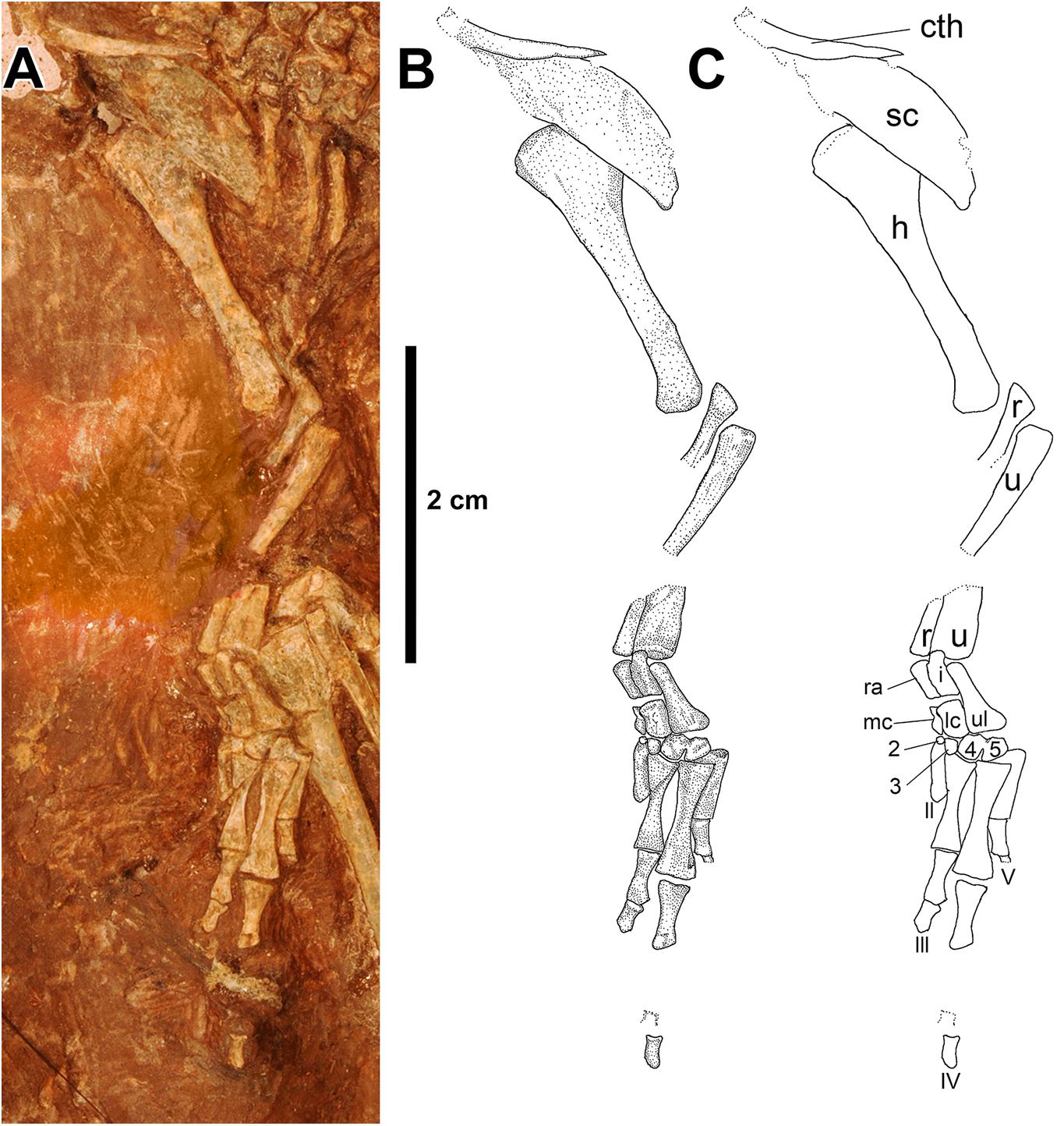
Figure 6. Eudibamus cursoris, holotype (MNG 8852). (A) Photograph, (B) illustration, and (C) labeled outlines of elements of forelimb and shoulder girdle in dorsal view.
Exposures of the clavicles, interclavicle, and coracoid plates (co-ossified anterior and posterior coracoids) are limited to ventral exposure on the ventral surface of the skull. The clavicles are strap-like and divided into subequal lengths by an approximately 90o twist about their long axis. The twist produces the standard division into an anteromedially directed cranial portion and a posterolaterally directed caudal portion. The cranial portion expands slightly as it nearly reaches the midline and overlaps the anterior surface of the lateral wing of the interclavicle. The distal half of the clavicle is slightly bowed laterally to match its contact with the anterolateral margin of the dorsal surface of the scapulocoracoid complex. The lateral wings of the T-shaped interclavicle are extremely narrow, taper to a distal point, and curve slightly posteriorly in contact with the anterolateral marginal contour of the coracoid plate. A long narrow posterior stem approximates the length to the midline contact of the coracoid plates. It exhibits little change in width except for two minor swellings, one near either end of its extent.
The coracoid plates are hemi-circular in outline, flaring laterally from presumably a straight mutual midline contact dorsal to the interclavicle stem. A low poorly defined ridge transversely divides the ventral surface of the left coracoid plate into unequal portions, which may mark the line of fusion of the anterior and posterior coracoids.
Forelimb
To avoid confusion in describing the bones of the fore- and hindlimbs (defined here as including the manus and pes, respectively) the anatomical views used are based on a posture in which the limbs occupy a parasagittal plane that extends directly anterior from the girdles. Description of the forelimb relies almost entirely on the mostly articulated left limb (Figure 6), which is missing short midlength sections of the radius and ulna shafts, and entirely distal carpal 1, metacarpal I, and all but a few proximal phalanges of digits III-V. Preservation, however, is complete enough recognize its pronounced elongation with an estimated length equal to 82.7% of the trunk length (Berman et al., 2000). In contrast, the right forelimb is limited to the humerus, distal portions of the radius and ulna, and a few unidentifiable isolated bones. The humerus is not of the typical primitive tetrahedral structure common to late Paleozoic tetrapods in which greatly expanded proximal and distal heads occupy different planes and are joined by a short shaft, but instead has the unique appearance that is more typical of a late Paleozoic tetrapod femur. The left humerus has a greatly elongated slender morphology except for an expansive deltopectoral crest that projects medially from the proximal head and gradually narrows as it extends distally along shaft to nearly its midlength. Although most of the deltopectoral crest is obscured by an overlying scapular blade, in the ventral view of the shoulder girdle a narrow margin of the proximal end of the humerus is slightly disarticulated from the glenoid, exposing the combined terminal articular margins of the humerus and crest as being transversely continuous. The distal head of the humerus is divided into two well-developed closely adjacent condyles that increase in size distally to vertically elongated and slightly convex terminal facets in which presumably the medial condyle contacted the radius and the lateral condyle the ulna. The medial and lateral condyles represent a radical restructuring of the ectepicondyle and entepicondyle condyles of the typical Paleozoic tetrapod humerus, which also accounts for the absence of the ectepicondylar and entepicondylar foramina.
The mid-shaft sections of the left epipodials are missing, but the otherwise articulated nature of the forelimb allows confident interpretation of their lengths and structure. Both bones can be described as being straight, very slender, and closely spaced to one another with lengths approximately equal to 78% of the humerus length. The shafts exhibit gradual narrowing from the slightly expanded proximal and distal heads with vertical transversely flat terminal facets. The ulna lacks an olecranon process and semi-lunar or sigmoid notch, which presumably is related to its slender structure and end-to-end contact with the lateral condyle of the humerus.
Identifiable elements of the carpus include the radiale, intermedium, ulnare, medial and lateral centralia, and distal carpals 2–5 (distal carpal 1 is either lost or not yet ossified). The ulnare, the largest of the carpals, is flask-shaped, narrowing proximally from an expanded distally flat base that contacts distal carpals 4 and 5 to a narrow stem-like structure of uniform width that contacts the distal end of the ulna. Medially the ulnare contacts the lateral margins of the intermedium proximally and the lateral centrale distally, the combined lengths of which match that of the ulnare.
The remaining four proximal carpals are uniquely elongated longitudinally and articulated in end-to-end pairs of subequal widths, which contribute to the relative lengthening of the manus. Furthermore, the proximal carpals are unusual in being tightly compacted mediolaterally with their proximal and distal articular facets aligning in transverse vertical planes. The radiale and medial centrale extend between contacts with the radius proximally and the distal carpal 2 distally. Laterally the pair contacts the medial margins of the intermedium proximally and the lateral centrale distally of much greater widths and extend between contacts with the ulna proximally and distal carpals 3 and 4 distally.
An articulated series of four distal carpals is visible and believed to include 2–5 based on their contacts with the proximal carpals, although conceivably they could represent 1–4. Distal carpals 4 and 5, the largest of the series, are subrectangular in outline, whereas 2 and 3 are greatly reduced to small spheres. The mutual contact between 4 and 5 is fused proximally and regarded as an anomaly. Metacarpal I is absent, II is incomplete, and III-V are complete and well preserved. The digits of the manus are tightly compacted mediolaterally and greatly elongated with the longest fourth equaling about 28% of the trunk length (Berman et al., 2000). Metacarpals III-V are greatly elongated, particularly IV, which is about 20 and 50% longer than III and V, respectively. The proximal and distal heads are greatly expanded with vertical, essentially transversely flat terminal facets and narrow to very slender midlength shafts.
Pelvic Girdle
The holotypic pelvic girdle is very likely complete, but exposure is limited to dorsal views of the ilia (Figure 1). Fortunately, the pelvic girdle in the referred specimen MNG 12895 (Figure 2) is nearly complete with the lateral surfaces of the bilaterally paired pubes and ischia of the puboischiadic plates exposed ventrally, and a lateral exposure of the left ilium that lacks a large portion immediately anterior to the acetabulum. The pelvic bones are well-defined by narrowly open sutures, suggesting that even this individual was not fully mature. For clarity, each element of the pelvic girdle is described in the universally accepted perspective of occupying a vertical parasagittal plane, which here is laterally exposed. However, it is recognized that in life the puboischiadic plates extended dorsolaterally from their midline union with the ilium directed dorsally.
Several unique features distinguish the ilium from those of other Paleozoic tetrapods, foremost among which is the absence of a distinct dorsal blade. The ilia in the holotype are complete and occupy their correct relationship with the sacrum but appear to be barely disarticulated from the sacral transverse processes. However, in both the holotype and referred specimen MNG 12895 the typical broad dorsal iliac blade is absent and in its place elongated anterior and posterior processes of subequal lengths unite above the acetabulum in a narrow band with an uninterrupted horizontal dorsal margin. In the holotype the processes taper gradually and curve slightly laterally as they extend to a distal point. The anterior process of the laterally exposed left ilium in MNG 12895 is almost completely broken away, but the posterior process is complete and well preserved. Anteriorly the posterior process borders the posterior half of a complete acetabulum, whereas posteriorly it gradually narrows to a blunt point as it extends to the posteriormost level of the puboischiadic plate. Enough of the anterodorsal portion of the posterior process remains to indicate that the acetabulum was narrowly separated from the dorsal margin of the ilium, and, therefore, eliminating the presence of the short neck-like constriction of the ilium that characteristically separates the two structures in most Paleozoic tetrapods. An equally unique feature of the ilium is a deep circular cup-shaped acetabulum that is completely bordered by a thick extremely rugose rounded ridge and contained completely within the ilium immediately above its ventral contact with the puboischiadic plate; that is, neither the ischium nor the pubis contributed to the acetabulum. A nearly identical morphology was figured by Watson (1954) in an isolated left ilium of Bolosaurus striatus.
The paired pubes and ischia of the puboischiadic plates in MNG 12895 are essentially complete except for some marginal loss of the right pubis and are well-defined by narrowly open sutures. At a level shortly anterior to the midlength of the puboischiadic plates the vertical contact between the ischium and pubis ends dorsally in a wide shallow V-shaped sutural notch for the ventral margin of the ilium. The lateral surfaces of the pubes and ischia are flat except, as is typical in Paleozoic tetrapods, the portion of the ischium bordering the iliac notch being thickened into a smoothly rounded ridge that curves a short distance posteroventrally where it is continued as a much thinner slightly convex dorsal margin that extends the remaining length of the bone. The ventral margin of the ischium is broadly convex and contacts its bilateral counterpart for about half their anterior lengths. Posteriorly the ventral margin curves strongly upward away from the midline to a pointed union with the slightly convex posterior portion of the dorsal margin. Although the pubes are considerably shorter anteriorly than the ischia, in outline they are roughly similar to one another. There is no evidence of a pubic tubercle or an obturator foramen seen in some Paleozoic tetrapods.
Hindlimb
Considered together the holotype and referred specimen (MNG 12895) allow a thorough description of the hindlimb. In the holotype (Figure 7) an essentially complete articulated left hindlimb is exposed dorsally except for the femur being ventrally exposed. In contrast, in the right hindlimb the dorsally exposed femur and crus are narrowly disarticulated, and the few remaining elements of the pes are mostly disarticulated. The referred specimen MNG 12895 (Figure 2) includes an articulated ventrally exposed right hindlimb that lacks only the distal head of the femur. The two specimens differ strikingly in a few features in their tarsus. Whereas the forelimb has been characterized as relatively elongate for a late Paleozoic tetrapod at over 87% the length of the presacral column, the hindlimb is dramatic in this regard at nearly 135% its length.
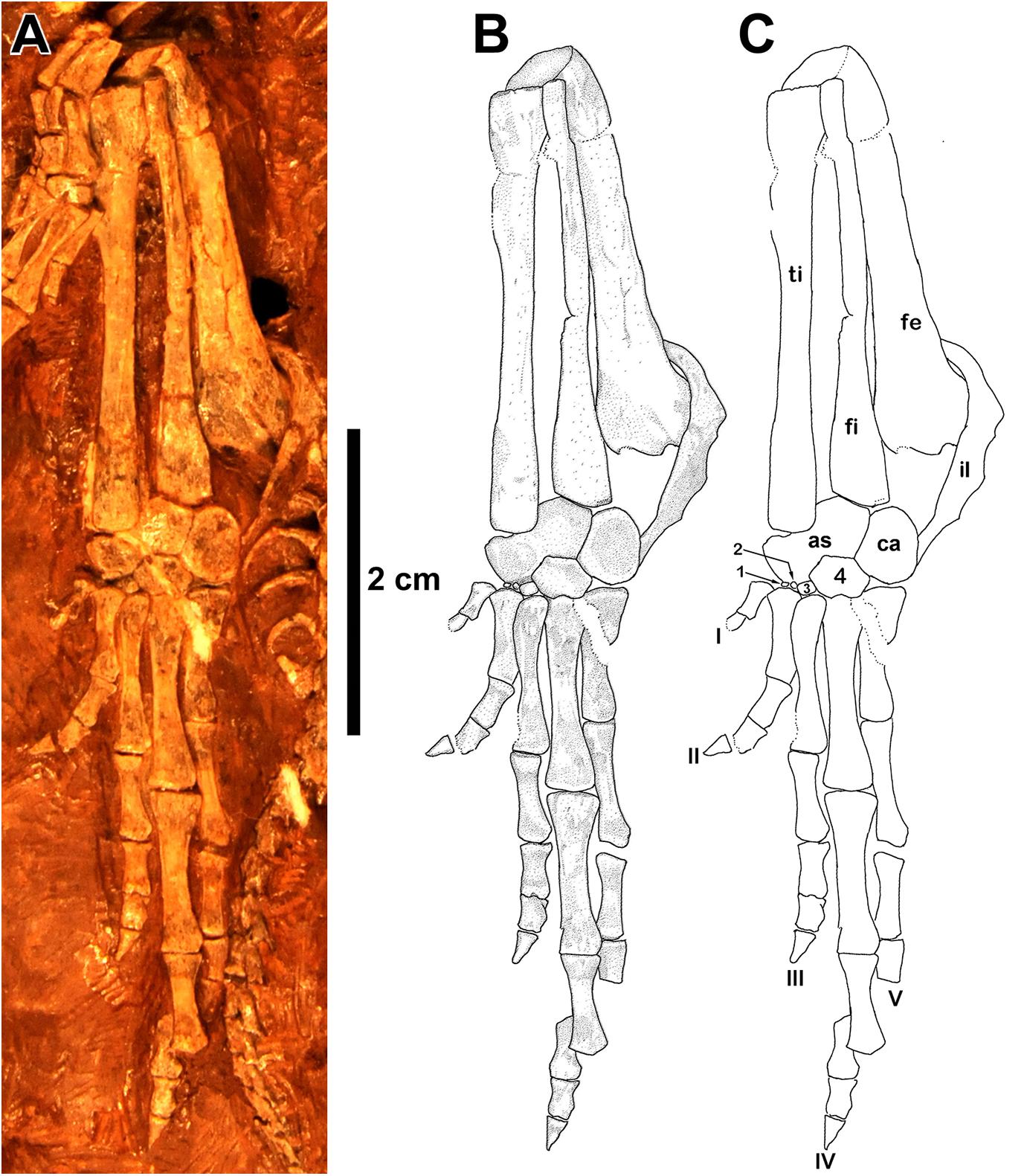
Figure 7. Eudibamus cursoris, holotype (MNG 8852). (A) Photograph showing left hindlimb in dorsal view except for ventral view of femur. (B) Illustration and (C) outlines of labeled elements of femur, crus, and pes.
As with the humerus the femur of Eudibamus is much more gracile in build than those of late Paleozoic tetrapods. The well-preserved holotypic left femur is slender, nearly straight, and gradually expands proximally and distally from its midlength region, nearly doubling in width at its transversely flat terminal margins. Projecting medially from the proximal head of the femur is a prominent triangular internal trochanter that narrows medially to a sharply pointed angle. There is no obvious feature demarking the division between the two structures, and together they contribute to an uninterrupted narrow transverse terminal facet. The distal head of the femur curves slightly ventrally to terminate in a transversely oval, essentially flat facet that is shared disproportionably by contacts of the proximal heads of the epipodials, that of the tibia facet being much larger. Consistent with its gracile build, the distal head of the femur is atypical of late Paleozoic tetrapods in not being divided into dominant separate condyles that contacted the epipodials.
The epipodials in the holotype and referred specimen MNG 12895 are greatly elongated, extraordinarily slender, straight, subequal in length, and closely adjacent to one another. The crurofemoral length is slightly greater than half the total length of the limb. For most their midlength region the epipodials are circular in cross section but gradually expand slightly transversely as they extend to the proximal and distal heads. Whereas the terminal facet of the tibia for the astragalus is transversely oval, the distal end of the fibula is divided subequally into two distinct flat facets, one facing distomedially for the astragalus and the other facing distolaterally for the calcaneum.
The tarsi of the holotype and referred specimen MNG 12895 are not a perfect match. The referred specimen exhibits the standard Paleozoic tetrapod complement of tarsal bones that includes the astragalus, calcaneum, centrale, and distal tarsals 1–5. The holotype, on the other hand, differs most prominently from the referred specimen in the absence of the centrale and distal tarsal 5, and greatly reduced sizes of distal tarsals 1–3, all of which are features most reasonably attributable to an earlier ontogenetic stage of ossification than that of the referred specimen. In both specimens the astragalus conforms in general to the standard reversed L-shaped configuration of Paleozoic tetrapods except mainly for the internal angle being slightly greater than a right angle. Both limbs of the astragalus are roughly rectangular in outline, but they differ in the proximally directed limb being narrower and shorter than the medially directed limb. The proximal surface of the longitudinal limb slopes distolaterally in contact with the medial facet of the fibula. The tibia-astragalus contact is atypical among Paleozoic tetrapods in that the tibia contacts the proximal surface of the transverse limb rather than, as is standard, the distal terminal surface. Also, possibly unique, the facet for the tibia, as revealed in the ventral view of the referred specimen MNG 12895, is expanded ventrally by a narrow but prominent shelf-like structure.
In the referred specimen the medial half of the distal surface of the transverse limb of the astragalus contacts the proximal surface of the centrale in a distally convex suture, whereas the lateral half of the distal surface of the astragalus contacts a proximomedial facet of distal tarsal 4. In both the holotype and referred specimen the longitudinal length and transverse width dimensions of the calcaneum are subequal, although the width is maximized by a convex expansion of the lateral margin. As is standard in Paleozoic tetrapods the astragalus and calcaneum in both specimens contact one another in a longitudinally straight suture, which at about midlength in the ventrally exposed referred specimen exhibits evidence of a foramen for the perforating artery. A distomedially sloping proximal margin of the calcaneum contacts the lateral facet of the fibula. The distal margin of the calcaneum is divided into two distinct adjacent facets of subequal lengths, a distomedially facet for distal tarsal 4 and a transverse facet for distal tarsal 5. In the absence of distal tarsal 5 in the holotype the distal margin of the calcaneum is narrowly separated from contacting the proximal margin of metatarsal V. The centrale of the referred specimen is transversely elongate with a slightly convex distal margin that is contacted by distal tarsals 1–3, whereas laterally it contacts the medial margin of distal tarsal 4. Distal tarsals 1–3 are extremely small compared to 4 and 5 with narrow proximodistal elongate outlines; the slightly larger 3 contacts the greater ventral portion of the medial margin of 4. Distal tarsal 4, by far the largest of the series, is pentagonal in outline. The proximal margin is divided into two equal, distally diverging facets, one contacting the distolateral corner of the astragalus and the other the distomedial margin of the calcaneum. Distal tarsal 5 has roughly a square outline with a transverse proximal margin that contacts the distal margin of the calcaneum in a transverse suture and the medial margin contacts the greater ventral portion of the lateral margin of distal tarsal 4. The distal articular margins of the distal tarsals are slightly convex and restricted to their respective metatarsals.
The pes lengths of the holotype and referred specimen MNG 12895 are nearly equal and remarkable in their great lengths of about 43% that of the presacral column in the holotype and approximately 46% that of the second specimen based on comparison of vertebral lengths. As is typical in late Paleozoic tetrapods digits I–IV in both specimens greatly lengthen serially, but rather than V being typically substantially shorter than III, it is subequal in length. Most noticeable, however, in the serial lengthening of the digits there is a marked shift in the lengths from the typically short I and II to the extraordinarily long III–V. The elongation of digits III–V is due in great part to a unique disproportionate lengthening of their first phalanx. In the holotype the length of the first phalanx of digits III–V is equal to about 59, 83, and 96% of their respective metatarsal lengths, and in the referred specimen the values for the same proportions are 67, 61, and 83%. In the seven basal synapsid pedes illustrated by Romer and Price (1940, Figure 41) the same proportions yield greatly reduced values that rarely exceed 50% and then only by a slight percentage, otherwise the great majority of values range in the 30–40 percentiles. A similar pattern is seen in basal eureptilian captorhinid reptiles (Sumida, 1989; Holmes, 2003). As a result of reapportioning the lengths of digits III–V, in particular the unusual lengthening of V, they uniquely display a strong degree of bilateral symmetry.
The distal tarsals in MNG 12895, in strong contrast to those in the holotype, are strongly ossified with well-defined marginal contacts, including a distally convex contact with their respective metatarsals. Accounting for the missing ungual of digit I in the holotype, both specimens retain the primitive phalangeal count of 2-3-4-5-4. The unguals, best exposed in MNG 12895, are slightly longer than the penultimate phalanx, extremely narrow, and sharply pointed with little ventral curvature.
Structures Supporting a Parasagittal Stride and Digitigrade Stance of the Limbs
General
In the original description of Eudibamus cursoris (Berman et al., 2000) several structural and proportional skeletal features were described, almost all appendicular, that clearly indicate a unique ability to attain relatively high speeds during quadrupedal and bipedal locomotion. This was achieved by both limbs being greatly elongated and employing a parasagittal digitigrade stance and gait with the fore- and hindlimbs occupying near vertical postures and swung fore and aft in a pendulum-like arc. In Eudibamus this was affected in great part by three structural and proportional features that are widely and frequently encountered in its skeleton. Foremost is a pronounced lengthening of the limb bones. This served to increase stride length, and, therefore, running speed, as it is stride length and not stride frequency that running speeds are most greatly increased in vertebrates (e.g., Snyder, 1954, 1962; Irschick and Jayne, 1999). Secondly, maximum extension between terminal articulations of elongate limb bones during rapid locomotion was made structurally possible by hinge-type joints in which flat opposing facets meet in a transverse plane perpendicular to the long axis of the limb. This would have constrained limb orientation to a near vertical parasagittal plane regardless of the degree of flexion or extension of the joint. Lastly, the vertical posture of the limbs and mediolateral compactness of the longitudinally elongated bones about the long axis of the limb permitted a slender structure, which reduced the weight of the limbs without sacrificing their structural support of the trunk. In addition to the above structural and proportional adaptations, others are recognized and described below under their respective major skeletal subdivisions.
Axial Skeleton
(1) The uniquely short slender trunk ribs reflect a narrow trunk that would have allowed the limbs to be brought closer to the body midline, providing not only greater support of the trunk, but also increased the ease and efficiency of the fore and aft swing of the extended limbs during rapid quadrupedal or bipedal locomotion.
(2) The uniquely long tail (about 64.3% of the entire axial length, Berman et al., 2000) would have had the effect of greatly increasing its relative weight. Presumably this would have affected a significant caudal displacement of the body center of mass posteriorly beyond the pelvis to the anterior region of the tail, a condition that has been suggested as a feature of the earliest reptilian bipeds (Clemente, 2014). The result would be an increased length of the moment arm of the tail, the distance between the body center of mass and the femur-hip joint and an increase in the rotational torque at the hip joint, which would aid in lifting the front of the body into a bipedal stance and maintaining it during bipedal locomotion (Clemente and Wu, 2018).
Pelvis
(1) The thick rounded, rugose ridge bordering the circular margin of the acetabulum presumably served to prevent dislocation of the femur head, particularly critical during parasagittal bipedal gait with the hindlimb held in a nearly vertical stance.
(2) The deep circular cup-like acetabulum is uniquely contained completely within the ilium immediately above its ventral contact with the puboischiadic plate. This is very likely in response to the hindlimbs having shifted from a sprawling to a vertical posture positioned directly beneath the trunk. In a sprawling posture the force of the femur is directed mainly medially, impacting the three pelvic bones radiating from the acetabulum, and thus provided minimal support of the trunk. However, with the hindlimbs held in a vertical posture beneath the trunk and close to the midline, the need for much greater support of the body mass is provided structurally that is otherwise lacking in a sprawling posture.
Forelimb
(1) The forelimb is relatively greatly elongated and slender with a fully extended length equal to 82.7% of the trunk length (Berman et al., 2000).
(2) The humerus is unusually slender with almost no expansions of the proximal and distal heads in marked contrast to the primitive tetrahedral structure common to late Paleozoic tetrapods in which the heads are enormously expanded.
(3) The distal head of the humerus is divided into two, well-developed, tightly adjacent, and longitudinally elongated condyles of subequal size that distally contacted the epipodials in a resultant hinge joint, thus restricting the limb bones of the elbow to a vertical parasagittal plane regardless of their degree of flexion or extension.
(4) The slender structure of the humerus presumably accounts for the absence of both the ectepicondylar and entepicondylar foramina.
(5) The epipodials are straight, very slender with only slight expansions of the proximal and distal heads, closely spaced parallel to one another about the longitudinal axis of the limb, and subequal in length.
(6) The absence of an olecranon process and semi-lunar or sigmoid notch of the ulna is presumably related to its slender structure and terminal margin contact with the lateral condyle of the humerus.
(7) Several features of the proximal carpals uniquely distinguish the carpus among Paleozoic tetrapods. They are tightly compacted mediolaterally about the long axis of the limb, and with the exception of the ulnare, the four other proximal carpals have outline shapes of elongated rectangles of subequal widths and lengths that articulate longitudinally in end-to-end pairs. The ulnare is greatly elongated, matching the length of the paired proximal carpals, and collectively the proximal carpals contribute to a uniquely lengthened manus.
(8) The proximal and distal proximal terminal articular facets of the carpals for the epipodials and distal carpals, respectively, are of the hinge type and align transversely, allowing maximum flexion and extension of the manus restricted to a vertical parasagittal plane.
(9) The digits are tightly compacted mediolaterally with the length of the fourth being greatly elongated to equal about 28% of the trunk length (Berman et al., 2000).
Hindlimb
(1) In both the holotype and referred specimen the hindlimb is extraordinarily elongated with an estimated length in the former equal to 134.7% of the trunk length (Berman et al., 2000). Additionally, although the forelimb may be characterized as elongated relative to those of other late Paleozoic tetrapods, it is relatively shorter than the hind limb, a feature characteristic of other bipedal reptiles (Snyder, 1954, 1962).
(2) In both specimens the bones of the knee and ankle articulate longitudinally in simple hinge joints. In a digitigrade stance this confines the limb to a narrow vertical parasagittal plane regardless of the degree of flexion or extension at the joints. Furthermore, the fully extended hindlimb provided maximum limb length and, therefore, also stride length and running speed. It is assumed that as high running speeds were reached a concomitant lengthening stride of the long hindlimbs could only be accommodated without interfering with the much shorter strides of the forelimbs was by assuming a somewhat more erect posture and flexion of the elbow and wrist joints.
(3) The digits in both specimens are tightly compacted mediolaterally about the long axis of the limb for maximum support of the tarsus. Digits I and II are greatly reduced in length, especially relative to digits III–V, which prevented them from contacting the ground during extended digitigrade locomotion. Conversely, digits III–V are atypical in being greatly elongated and conforming strongly to a bilaterally symmetrical pattern. The lengthening of the digits is due in large part to a unique substantial lengthening of the first phalanx in digits III–V relative to the respective metatarsal length of each. Digits III–V therefore, presumably provided the entire vertical support of the hindlimbs during bipedal digitigrade locomotion. A more erect limb posture is correlated with an increase in effective mechanical advantage, and an elongation in metapodial limb segments to a digitigrade posture (Reilly et al., 2007). The hypothesized increase in stride length proposed here for Eudibamus agrees with models for correlations of stride length and speed increase as documented by work on extant reptiles (e.g., Snyder, 1954, 1962; Irschick and Jayne, 1999).
(4) In both specimens the tibia atypically contacts the proximal margin of the horizontal limb of the astragalus rather than the distal terminal margin, as is more standard in other late Paleozoic amniotes. In ventral view of the referred specimen the contact is expanded ventrally by a narrow but pronounced lip-like extension, which would have enhanced the bipedal digitigrade stance and locomotion in two ways: enhanced the support of the contact between the crus and tarsus and, therefore, also the vertical posture of the limb. The tibia is drawn close to the fibula, resulting in a compactness of the epipodials about the long axis of the limb that permitted a relatively long slender crus without sacrificing the vertical structural support of the trunk it provided.
(5) A complete circular, cup-like acetabulum is clearly contained completely within the ilium immediately above its ventral sutural margin with the puboischiadic plate. This is likely in response to the hindlimbs having shifted from a horizontally sprawling posture to a vertical posture positioned beneath the trunk. In a sprawling posture the force of the hindlimb is directed medially and is far less effective in supporting the weight of the trunk than if contained in a vertical parasagittal plane position beneath the trunk.
Conclusion
Eudibamus cursoris is truly a unique reptile for its times. Not only is it among the few oldest known bipedal reptiles, it is also the oldest known late Paleozoic reptile capable of achieving relatively high running speeds during quadrupedal and bipedal locomotion. This was accomplished by uniquely employing a parasagittal stride and digitigrade posture with the limbs positioned in a near vertical stance beneath the trunk and swung fore and aft in a pendulum-like style. This mode of locomotion is further supported by numerous structural features of both the fore and hind limbs. However, the possession of the unique locomotor adaptations by E. cursoris as early as the lower Permian raises the very plausible narrative that undiscovered terrestrial reptiles of approximately the same age also may not have been restricted to a sprawling quadrupedal locomotion style in their early history.
This skeletal description of the holotype skeleton of the lower Permian (Artinskian) bolosaurid parareptile Eudibamus cursoris from the Bromacker locality of Germany is augmented by, a well preserved, partial, articulated second more mature specimen that consists of a short portion of the vertebral column associated with the pelvis and right hindlimb with pes. This comprehensive description and interpretation of the anatomy of Eudibamus not only confirms a bolosaurid assignment, but also reinforces the previous interpretation that this small agile herbivore had the unique ability among Paleozoic vertebrates to reach relatively high bipedal and quadrupedal running speeds employing a parasagittal stride and digitigrade stance with the limbs held in a near vertical posture and swung fore and aft pendulum style (Figure 8). This is a surprising combination of features and locomotory abilities, and in strong contrast to the relatively slow, short limbed, massive herbivores of the Paleozoic. Interestingly, the anatomy of the older parareptiles Erpetonyx, considered to be the sister taxon to bolosaurids like Eudibamus, exhibits neither the dental nor the skeletal characteristics of the latter. This raises the possibility that the adaptation of bolosaurids to a herbivorous feeding strategy occurred in concert with their surprising adaptations to rapid bipedal and quadrupedal locomotion. We therefore interpret this as a mechanism that evolved not for food capture but for escape from the numerous predators that were common in the early Permian terrestrial vertebrate communities.
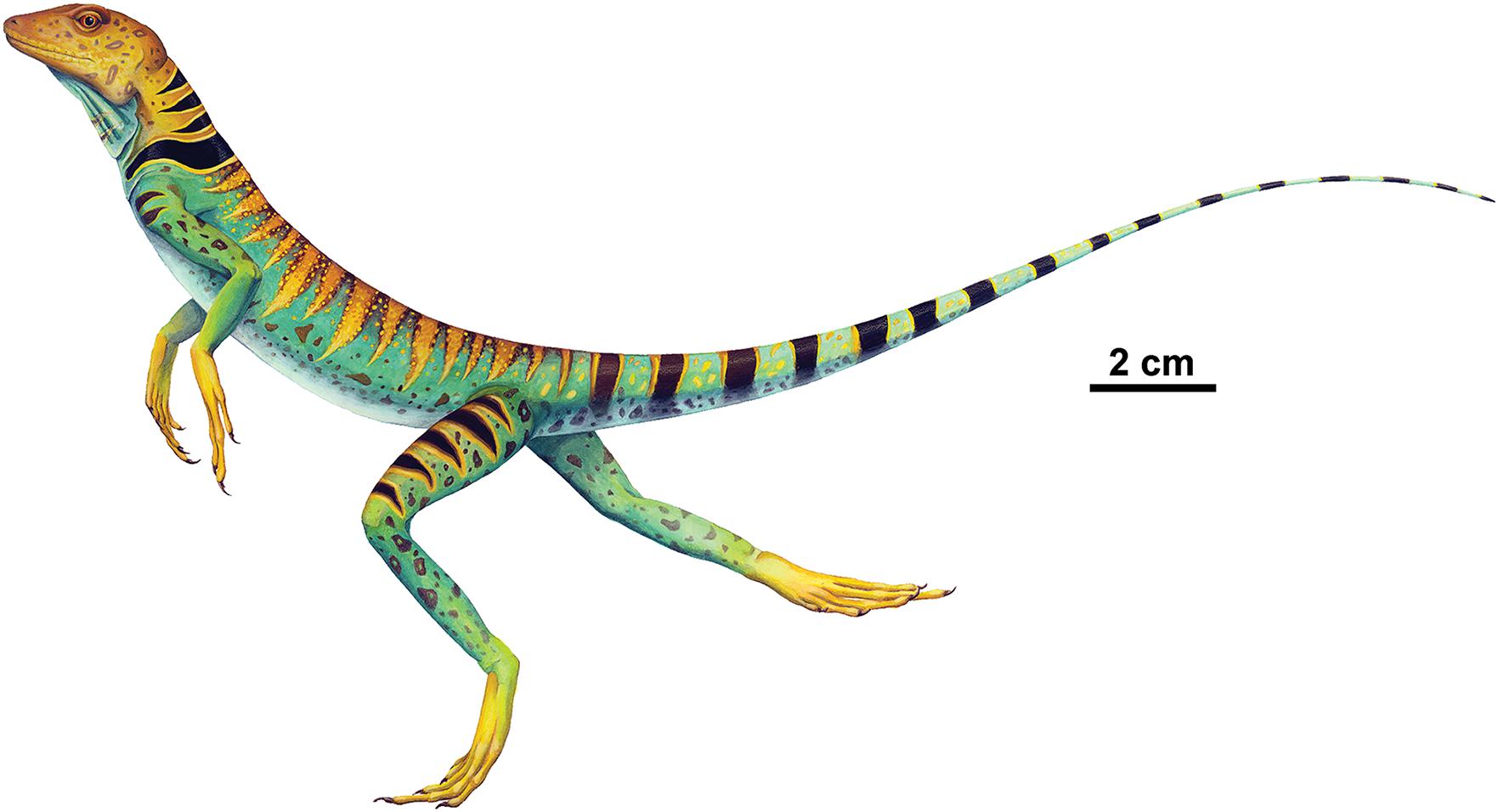
Figure 8. Whole-body restoration of Eudibamus cursoris running bipedally based on the holotype (MNG 8852). Illustration by Sandra Budd.
Data Availability Statement
The original contributions presented in the study are included in the article/supplementary materials, further inquiries can be directed to the corresponding author/s.
Ethics Statement
Ethical review and approval was not required for the animal study because animals studied were fossils.
Author Contributions
AH, DB, SS, and TM performed the fieldwork that resulted in recovery of the specimens described. AH performed most of the paleontological preparation on the specimens described, with DS contributing to detailed preparation. DB wrote the manuscript. AH, DS, RR, and SS reviewed and edited the manuscript drafts. DS and SS prepared the figures. RR played an important role in identifying cranial elements and interpreting their structural relationships with neighboring bones. All authors contributed to the article and approved the submitted version.
Funding
Research for this project was supported by grants from the National Geographic Society and North Atlantic Treaty Organization (NATO) (to AH, DB, and SS), Edward O’Neil Endowment Fund and M. Graham Netting Research Fund, Carnegie Museum of Natural History (to DB), Deutsche Forschungsgemeinschaft (DFG), Museum der Natur Gotha (to TM), Office of the College of Natural Sciences, California State University San Bernardino (to SS), and Natural Sciences and Engineering Research Council of Canada (to RR).
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
Special thanks are due to Andrew McAfee for his expertise in illustrating, composing, and formatting illustrations of Eudibamus cursoris. We are especially grateful to Sandra Budd for allowing us to reproduce her whole-body reconstruction of the holotype running bipedally in Figure 8. We also express our thanks to the reviewers whose careful constructive comments and criticisms had considerable impact on the improvement of the manuscript.
References
Berman, D. S., Maddin, H. C., Henrici, A. C., Sumida, S. S., Scott, D., and Reisz, R. R. (2020). New primitive caseid (Synapsida, Caseasauria) from the Lower Permian of Germany. Ann. Carneg. Museum 86, 43–75. doi: 10.2992/007.086.0103
Berman, D. S., Reisz, R. R., Scott, D., Henrici, A. C., Sumida, S. S., and Martens, T. (2000). Early Permian bipedal reptile. Science 290, 969–972. doi: 10.1126/science.290.5493.969
Carroll, R. L. (1973). The early differentiation of diapsid reptiles. Colloques Internationaux du Centre National de la Recherche Scientifique, no 218, Problemés actuels de Paleontologie. Evol. des Verteb. 1973, 433–449.
Carroll, R. L., and Gaskill, P. A. (1971). Captorhinomorph reptile from the Lower Permian of Europe. J. Paleont. 45, 450–463.
Carroll, R. L., and Thomson, P. (1982). A bipedal lizard-like reptile from the Karroo. J. Paleont. 56, 1–10.
Case, E. C. (1911). A revision of the Cotylosauria of North America. Carneg. Instit. Washing. Public. 145, 121.
Cisneros, J. C., Kammerer, C. F., Angielczyk, K. D., Fröbisch, J., Marsicano, C., and Smith, R. M. H. (2021). A new reptile from the lower Permian of Brazil (Karutia fortunata gen. et sp. Nov.) and the interrelationships of Parareptilia. J. Sys. Palaeont. 18, 1939–1959. doi: 10.1080/14772019.2020,1863487
Clemente, C. J. (2014). The evolution of bipedal running in lizards suggests a consequential origin may be exploited in later lineages. Evolution 68, 2171–2183. doi: 10.1111/evo.12447
Clemente, C. J., and Wu, N. C. (2018). Body and tail-assisted pitch control facilitates bipedal locomotion in Australian agamid lizards. J. Roy. Soc. 15:20180276. doi: 10.1098/rsif.2018.0276
Eberth, D. A., Berman, D. S., Sumida, S. S., and Hopf, H. (2000). Lower Permian terrestrial paleoenvironments and vertebrate paleoecology of the Tambach Basin (Thuringia, central Germany): the ‘Uplands’ holy grail. Palaios 15, 293–313. doi: 10.1669/0883-13512000015<0293:LPTPAV<2.0.CO;2
Falconnet, J. (2012). First evidence of a bolosaurid parareptile in France (latest Carboniferous-earliest Permian of the Autun basin) and the spatiotemporal distribution of the Bolosauridae. Bull. Soc. Geol. France 183, 495–508. doi: 10.2113/gssgfbull.183.6.495
Harris, S., Lucas, S. G., Berman, D. S., and Henrici, A. C. (2004). “Vertebrate fossil assemblage from the Upper Pennsylvanian Red Tanks Member of the Bursum Formation, Lucero Uplift, central New Mexico,” in Carboniferous-Permian Transition, New Mexico Museum of Natural History and Science, Bulletin 25, ed. S. G. Lucas, 267–284.
Holmes, R. (2003). The hindlimb of Captorhinus aguti and the step cycle of basal amniotes. Canadian J. Earth Sci. 40, 515–526. doi: 10.1139/e02-039
Hotton, N., Olson, E. C., and Beerbower, R. (1997). “Amniote origins and the discovery of herbivory,” in Amniote Origins, Completing the Transition to Land, eds. S. S. Sumida and K. L. M. Martin (Academic Press: San Diego), 207–264.
Irschick, D. J., and Jayne, B. C. (1999). Comparative three-dimensional kinematics of the hindlimb for high-speed bipedal and quadrupedal locomotion of lizards. J. Exp. Biol. 202, 1047–1065. doi: 10.1242/jeb.202.9.1047
Ivakhnemko, M. F., and Tverdochlebova, G. I. (1987). A revision of the Permian bolosauromorphs of Eastern Europe. Paleontologicheskii Zhurnal. 1987, 98–106.
Lucas, S. G., Berman, D. S., Henrici, A. C., and Hunt, A. P. (2005). “Bolosaurus from the Lower Permian at Arroyo del Agua, New Mexico, and its stratigraphic significance,” in The Permian of central New Mexico. New Mexico Museum of Natural Science Bulletin, 31, 125, eds S. G. Lucas, K. E. Ziegler, and J. A. Spielmann.
Mann, A., McDaniel, E. J., McColville, E. R., and Maddin, H. C. (2019). Carbonodraco lundi gen et sp. nov., the oldest parareptile, from Linton, Ohio, and new insights into the early radiation of reptiles. Roy. Soc. Open Sci. 6:191191. doi: 10.1098/rsos.191191
Modesto, S. P., Scott, D. M., MacDougall, M. J., Sues, H.-D., Evans, D. C., and Reisz, R. R. (2015). The oldest parareptile and the early diversification of reptiles. Proc. R. Soc. B 282:20141912. doi: 10.1098/rspb.2014.1912
Müller, J., Li, J.-L., and Reisz, R. R. (2008). A new bolosaurid parareptile, Belebey chengi sp. nov., from the Middle Permian of China and its paleogeographic significance. Naturwissenschaften 95, 1–6. doi: 10.1007/s00114-008-0438-0
Reilly, S. M., McElroy, E. J., and Biknevicius, A. R. (2007). Posture, gait, and the ecological relevance of locomotor costs and energy-saving mechanisms in tetrapods. Zoology 110, 271–289. doi: 10.1016/j.zool.2007.01.003
Reisz, R. R., Barkas, V., and Scott, D. (2002). A new Early Permian bolosaurid reptile from the Richards Spur Dolese Brothers Quarry, near Fort Sill, Oklahoma. J. Vert. Paleo 22, 23–28. doi: 10.1671/0272-46342002022[0023:ANEPBR]2.0.CO;2
Reisz, R. R., Müller, J., Tsuji, L., and Scott, D. (2007). The cranial osteology of Belebey vegrandis (Parareptilia: Bolosauridae), from the Middle Permian of Russia, and its bearing on reptilian evolution. Zool. J. Linn. Soc. 151, 191–214. doi: 10.1111/j.1096-3642.2007.00312.x
Romer, A. S., and Price, L. I. (1940). Review of the Pelycosauria. Geolog. Soc. Am. Special Paper 28, 1–538. doi: 10.1130/spe28-p1
Snyder, R. C. (1954). The anatomy and function of the pelvic girdle and hindlimb in lizard locomotion. Am. J. Anat. 95, 1–46. doi: 10.1002/aja.1000950102
Snyder, R. C. (1962). Adaptations for bipedal locomotion of reptiles. Am. Zool. 2, 191–203. doi: 10.1093/icb/2.2.191
Spindler, F., Werneburg, R., and Schneider, J. W. (2019). A new mesenosaurine from the lower Permian of Germany and the postcrania of Mesenosaurus: implications for early amniote comparative osteology. PalZ 93, 303–344. doi: 10.1007/s12542-018-0439-z
Sumida, S. S. (1989). The appendicular skeleton of the Early Permian genus Labidosaurus (Reptilia, Captorhinomorha, Captorhinidae) and the hind limb musculature of captorhinid reptiles. J. Vert Paleo. 9, 295–313. doi: 10.1080/02724634.1989.10011764
Watson, D. M. S. (1917). A sketch classification of the pre-Jurassic tetrapod vertebrates. Proc. Zool. Soc. London 1917, 167–186. doi: 10.1111/j.1096-3642.1917.tb02055.x
Keywords: Bromacker locality, cursorial features, Bolosauridae, Parareptilia, Bipedality
Citation: Berman DS, Sumida SS, Henrici AC, Scott D, Reisz RR and Martens T (2021) The Early Permian Bolosaurid Eudibamus cursoris: Earliest Reptile to Combine Parasagittal Stride and Digitigrade Posture During Quadrupedal and Bipedal Locomotion. Front. Ecol. Evol. 9:674173. doi: 10.3389/fevo.2021.674173
Received: 28 February 2021; Accepted: 11 May 2021;
Published: 20 July 2021.
Edited by:
Sean P. Modesto, Cape Breton University, CanadaReviewed by:
Juan Carlos Cisneros, Federal University of Piauí, BrazilMark Joseph MacDougall, Museum of Natural History in Berlin (MfN), Germany
Copyright © 2021 Berman, Sumida, Henrici, Scott, Reisz and Martens. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Stuart S. Sumida, c3N1bWlkYUBjc3VzYi5lZHU=