 Karen Greig
Karen Greig Nicolas J. Rawlence
Nicolas J. Rawlence- 1Southern Pacific Archaeological Research, Archaeology Programme, University of Otago, Dunedin, New Zealand
- 2Otago Palaeogenetics Laboratory, Department of Zoology, University of Otago, Dunedin, New Zealand
The pre-human Aotearoa New Zealand fauna was dominated by avian and reptilian species. Prior to first human settlement by East Polynesian colonists, the top predators were two giant raptorial birds. Aside from humans themselves, colonisation also resulted in the simultaneous introduction of two novel mammalian predators into this naive ecosystem, the kiore (Pacific rat) and kurī (Polynesian dog). While the ecological impacts of kiore are relatively well understood, those of kurī are difficult to assess, and as such kurī have frequently been disregarded as having any meaningful impact on New Zealand’s biodiversity. Here we use the archaeological and palaeoecological record to reassess the potential impacts of kurī on this ecosystem. We argue that far from being confined to villages, kurī could have had a significant widespread but relatively localised impact on New Zealand’s avian, reptilian and marine mammal (seals and sea lions) fauna as a novel predator of medium-sized species. In this way, kurī potentially amplified the already significant impacts of Polynesian colonists and their descendants on New Zealand’s ecosystem, prior to European arrival. As such, kurī should be included in models of human impact in addition to over-hunting, environmental modification and predation by kiore.
Background
Isolated island ecosystems are evolutionary microcosms typically exhibiting high levels of faunal endemicity (Mendelson and Shaw, 2005; Shaw and Gillespie, 2016). They are also often the last places to have been colonised by modern humans, as recently as hundreds to a few thousand years ago (Wilmshurst et al., 2011; Hansford et al., 2021), at times of relative climatic stability (Wanner et al., 2008; Waters et al., 2017). In island ecosystems modern human colonisation is frequently associated with widespread faunal extinctions and environmental modification (Perry et al., 2014; Hansford et al., 2021; Louys et al., 2021; Nogue et al., 2021).
Aotearoa New Zealand presents a unique opportunity to test for anthropogenic impacts on insular biodiversity. Its flora and fauna were shaped by tens of millions of years of isolation and regular long-distance dispersal (Wallis and Jorge, 2018), and exhibit a range of island adaptions including gigantism, flightlessness or flight-reduced terrestrial lifestyles, and slow K-selected breeding strategies (Worthy and Holdaway, 2002). At the time of East Polynesian colonisation in the late thirteenth to early fourteenth centuries CE (Wilmshurst et al., 2008, 2011), the New Zealand fauna was dominated by birds (>223 species), reptiles and marine mammals, specifically pinnipeds (four species of seals and sea lions). The avian fauna ranged from extinct birds such as nine species of giant flightless moa (Dinornithiformes; Bunce et al., 2009) to medium-sized ground-dwelling taxa including moa chicks or juveniles, giant goose (Cnemiornis spp.), adzebill (Aptornis spp.), moho (Porphyrio mantelli) and other rails, and several waterfowl (e.g., Finsch’s, blue-billed, musk and pink-eared ducks, merganser, and poūwa swan), as well as extant terrestrial birds including takahē (Porphyrio hochstetteri), kākāpō (Strigops habroptilus) and kiwi (Apteryx spp.; Worthy and Holdaway, 2002; Tennyson and Martinson, 2007). The top predators in the pre-human ecosystem were also birds including Haast’s eagle (Aquila moorei) and Eyles’ harrier (Circus eylesi; Tennyson and Martinson, 2007). Within reptiles, there were at least 110 species of Eugongylinae skinks and Diplodactylid geckos, seven Leiopelmatid frogs and one tuatara (Sphenodon punctatus) (Easton et al., 2017; Gemmell et al., 2020; Scarsbrook et al., 2021).
The arrival of Polynesians resulted in the widespread human-driven extinction of around 50% of the vertebrate biodiversity (Figure 1) as a result of hunting (Anderson, 1989; Holdaway et al., 2014; Perry et al., 2014) and anthropogenic environmental modification (McWethy et al., 2014), in addition to biological turnover events (Rawlence et al., 2017a), range-contractions (Salis et al., 2016), population bottlenecks (Rawlence et al., 2015a), and significant changes in indigenous forest cover through widespread anthropogenic burning (McWethy et al., 2014; McConnell et al., 2021). Within a few hundred years of human colonisation, the New Zealand ecosystem had been fundamentally transformed, which has left a clear signature in the archaeological and subfossil record (Holdaway and Worthy, 1996; Tennyson and Martinson, 2007; Holdaway et al., 2014; Perry et al., 2014; Walter et al., 2017; Waters et al., 2017; Rawlence et al., 2019). Polynesians also simultaneously introduced two exotic predators (Figure 1), the kiore (Pacific rat, Rattus exulans) and kurī (Polynesian dog, Canis familiaris) (Wilmshurst et al., 2008; Greig et al., 2018). Bones and teeth from both are found in colonisation-era archaeological sites throughout the three main islands, and on some but not all offshore islands (e.g., kurī have not been documented in archaeological sites on Rēkohu Chatham Islands ∼780 km east of mainland New Zealand but have been inferred, based on chewed bone, to have been present on the sub-Antarctic Auckland Islands ∼480 km south of the mainland; Davidson, 1987; Anderson, 2005; Greig and Walter, 2021). Well-documented archaeozoological analysis of early archaeological sites containing kurī bones also shows the presence of numerous extinct prey taxa (e.g., Anderson et al., 1996; Furey, 2002; Worthy and Holdaway, 2002; Scofield et al., 2003). Kurī were also a source of meat and industrial materials (e.g., kahu kurī dog skin cloaks, bone for tools and ornaments) for Māori, the direct descendants of those East Polynesian immigrants (Anderson, 1981, 1989; Davidson, 1987; Hartnup et al., 2011).
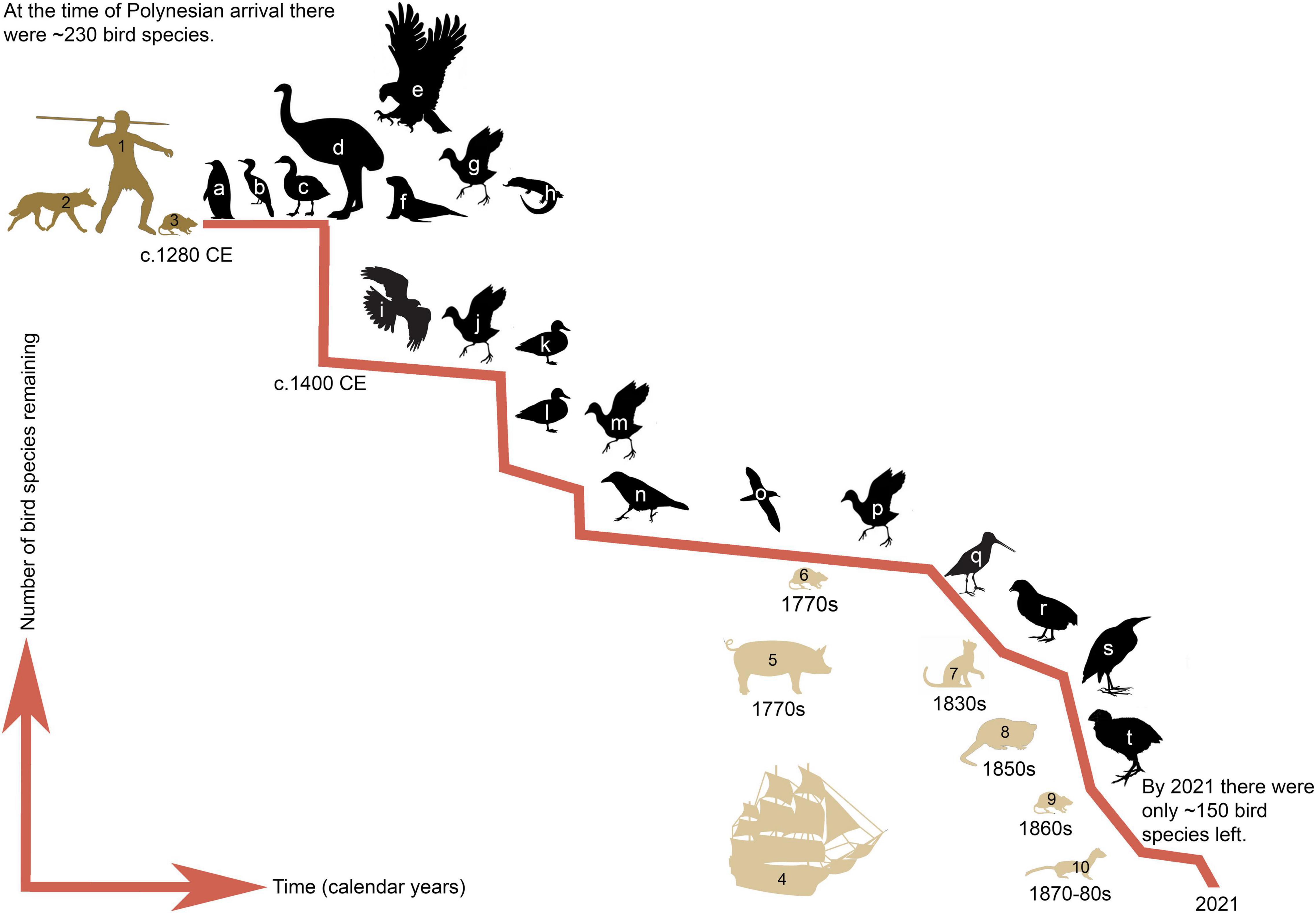
Figure 1. Schematic of medium-large sized vertebrate extinctions in Aotearoa New Zealand and the introduction of novel mammalian predators since East Polynesian, and latter European, settlement. Extinct vertebrates include, but are not limited to, (a) penguins; (b) shags; (c,k,l) waterfowl including geese, swans, and ducks; (d) large flightless moa; (e) eagles; (f) pinnipeds such as rāpoka sea lion and ihupuku southern elephant seals; (g,j,m,p,t) rails; (h) reptiles and frogs; (i) harriers; (n) ravens; (o) shearwaters; (q) snipe; (r) quails; and (s) bitterns. Predators introduced by East Polynesians (1) include Polynesian dog or kurī (2), and Pacific rat or kiore (3); and those introduced by Europeans (4), which included pigs (5), Norway rats (6), cats (7), brushtail possums (8), ship rats (9), and mustelids including ferrets, stoats and weasels (10). Figure adapted from Tennyson and Martinson (2007) with the inclusion of additional species from Worthy (1991); Boessenkool et al. (2009), Collins et al. (2013), Rawlence et al. (2015b, 2017a, 2017b), Easton et al. (2017).
Much archaeological and palaeoecological research has focused on the direct impacts of humans (i.e., hunting, environmental modification; Anderson, 1989; Holdaway et al., 2014; Perry et al., 2014; Nogue et al., 2021) and predation by kiore - the latter based on modern ecological (Rayner et al., 2007; Ismar et al., 2014) and palaeoecological (Wilmshurst and Higham, 2004; Wilmshurst et al., 2008) studies. In contrast, the potential impacts of kurī in particular on the pre-European contact ecosystem have largely been overlooked as they are difficult to assess, despite the potential of dogs to be a major novel predator in recently colonised ecosystems (e.g., Koungoulos and Fillios, 2020; Hixon et al., 2021). Fleming (1962) stated “the simplest explanation is to attribute all late Holocene extinction to the profound ecological changes brought about by man with fire, rats and dogs,” while Anderson (1981, 1989) suggested kurī probably contributed to the extinction of moa. In contrast, Holdaway (1999) argued nearly all the known extinctions were due to people and kiore, and not kurī, which “probably had little effect on the biota.” Worthy and Holdaway (2002) took this idea one step further, boldly stating “the Polynesian dog can be exonerated: it was kept so close to camps that it is not a factor.”
Our aim is to discuss and reassess the potential ecological impacts of kurī on the pre-European New Zealand ecosystem, including their role in human-driven extinctions, and highlight future research directions that need to be addressed to fully understand the impact that dogs can have on insular island faunas.
Kurī (Polynesian Dogs)
An investigation of the impacts of kurī on Aotearoa New Zealand’s biota requires an understanding of kurī biology, behaviour and ecology. This is problematic, as by the mid-nineteenth century CE kurī were no longer identifiable as a distinct dog type in Aotearoa New Zealand, due to interbreeding and replacement by European dogs. Dogs were brought to New Zealand around the late thirteenth century CE by East Polynesian migrants, the end point of a major trajectory of human colonisation of the islands of the Pacific (Greig and Walter, 2021). Dogs were successfully transported and established on many islands of the region during these migrations. Molecular genetic studies show that Pacific dogs possess a distinctive mitochondrial genetic signature, distinguishable from other lineages, most likely originating from southern China (Oskarsson et al., 2012; Greig et al., 2018; Zhang et al., 2021). In the absence of extant populations, the bones, teeth and coprolites (i.e., desiccated faeces) of kurī recovered from archaeological sites now comprise a valuable source of information. Archaeozoological studies and emerging biomolecular techniques can provide data about kurī physical characteristics, diet, mobility, and genetic history. In addition to archaeological remains, there are numerous observations about kurī in early European historical literature, although most accounts focus on physical descriptions, sometimes with a brief comment about the uses of dogs by Māori (for a review see Colenso, 1877), rather than ecological information. In the absence of this type of information, modern studies of free-ranging dog populations and ethnographic data can assist with developing hypotheses for aspects of kurī behaviour.
Kurī feature in Māori mythology, oral histories and art forms, demonstrating their importance in Māori culture (Potts et al., 2013). Legendary kurī are associated with colonising voyages to New Zealand, historical events and the naming of landmarks. In everyday life kurī were kept as companions, watch dogs and hunting dogs, and as a source of meat and industrial materials (bones, teeth, and pelts) (Davidson, 1987). Written descriptions in early European accounts tell of a small, fox-like dog, with pricked ears and a bushy tail (Colenso, 1877). Morphometric analysis of skeletal remains indicates an adult shoulder height of slightly under 40 centimetres, and a body weight of 13–15 kilograms (Clark, 1997). This suggests a body shape similar to a small border collie, but more robust and with shorter legs. There is very little geographic and temporal variation in kurī skeletal remains, suggesting that there was no deliberate selection for particular morphological characteristics, such as size, that can be observed in the skeleton (Clark, 1997), despite their importance as a source of food and raw materials. Information about kurī life history, such as reproductive behaviour is extremely limited. Kurī are thought to have reached sexual maturity around 6 to 8 months of age, similar to modern dog breeds (Clark, 1995).
It appears that sizable kurī populations were able to be sustained in early Māori settlements (Figures 2A,B). The results of archaeozoological analysis of faunal assemblages from early Māori archaeological sites such as Wairau Bar, Shag River Mouth, Kaupokonui and Houhora, for example, have documented substantial numbers of dog bones (Anderson, 1981, 1989; Greig et al., 2018). Some later Māori settlements dating from the sixteenth and seventeenth centuries also appear to have supported high numbers of kurī, for example, Kohika (Horrocks et al., 2002, 2003) and the Masonic Tavern site (Wood et al., 2016). During Captain Cook’s second voyage to New Zealand, while anchored in Queen Charlotte Sound, both he and the ship’s scientist Forster commented that they saw plenty of dogs, including those travelling with people in canoes (Colenso, 1877).
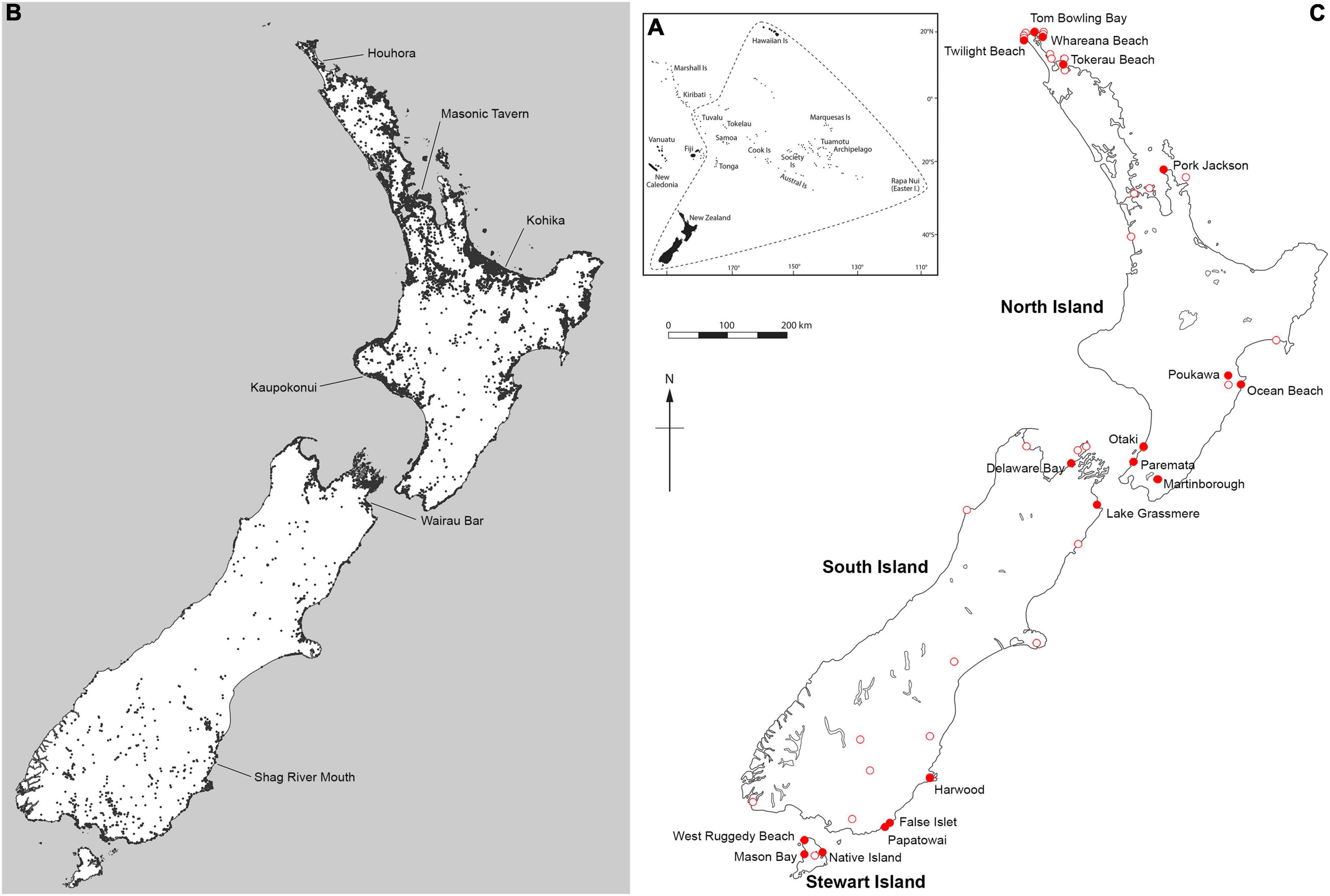
Figure 2. (A) The Polynesian triangle and Aotearoa New Zealand’s place within it. (B) Distribution of pre-European contact Māori archaeological sites (n = ∼54,000) in Aotearoa New Zealand as a proxy for the human-dependant distribution of the Polynesian dog or kurī (Canis familiaris). Each black dot or shaded area represents an archaeological site(s) (Source: ArchSite.org.nz). (C) Distribution of dog specimens reported in natural history collections (open red circles), including from sites containing dog and prey remains, whose ages overlap with Polynesian colonisation (closed red circles). See also Table 1 for further details.
Dogs are omnivorous generalists, capable of consuming and surviving on a wide variety of food types, ranging from human-derived garbage to animals that may be several times their body mass (Vanak and Gompper, 2009; Hughes and MacDonald, 2013). Kurī seen by early European travellers were not restrained or tethered (Clark, 1997) and had the freedom to roam throughout settlements and scavenge for food. This freedom would have enabled kurī to forage beyond the close confines of Māori villages (contra Worthy and Holdaway, 2002). Nineteenth century Māori villages often were observed with fenced enclosures or houses to prevent entry by dogs and pigs (the latter introduced by Europeans) (Earle, 1832; Dieffenbach, 1843). Many travellers described storehouses on poles or stilts several feet above the ground, used to protect food such as seed potatoes, or dried fish from roaming animals (Best, 1916). Left-over food was kept between meals in baskets on poles, for the same purpose (Earle, 1832). Stages or platforms were also documented to keep important objects away from kurī and kiore (Best, 1916).
Macroscopic analysis of kurī coprolites from archaeological sites suggests a varied diet consistent with the range of fauna commonly found in correspondingly dated Māori middens (rubbish heaps). Components include small bird, moa, fish, mollusc shell, as well as charcoal and other plant remains that often do not survive in middens (Clark, 1995, 1997; Irwin, 2004). A recent study of coprolites from the Masonic Tavern site in Auckland used both microscopic and ancient DNA methods, and identified fish, terrestrial and marine mollusc shell, charcoal, and wild and cultivated plant taxa (Wood et al., 2016). The contribution of faeces to the diet has also been noted for modern free-ranging dog populations (Butler et al., 2018). Preliminary isotopic analyses of kurī bones using carbon and nitrogen stable dietary isotopes further support a widely varied diet (Leach et al., 2003; Kinaston et al., 2013; Wood et al., 2017) incorporating terrestrial (i.e., similar isotopic compositions to herbivorous moa) and marine components.
To date, the only published evidence of kurī skeletal remains is associated with archaeological evidence for human activities (Figure 2B) rather than natural sites (e.g., caves, pitfalls, swamps, dunes), potentially creating a taphonomic sampling bias in available data. It is not clear whether kurī formed feral populations, although this is a possibility. There are few truly feral self-sustaining dog populations in the world today, where dogs live in a wild state independently of people. One example is in Australia, where dogs with a similar genetic ancestry to kurī were introduced around 4,000 years ago and subsequently became the dingo (Smith, 2015). Dingoes are an apex predator, and their prey includes a wide variety of Australian fauna and flora, including kangaroos, small mammals, birds, reptiles, fish, crabs, frogs, insects, and seeds, as well as introduced domestic species, including sheep (Smith, 2015). Kurī may have had a similar potential in New Zealand to form feral self-sustaining populations, particularly in warmer northern regions – indeed, recent feral dog populations have been observed in Northland (Piper, 2021) and “wild dogs” were observed subsisting on kākāpō, weka and ground dwelling birds in the nineteenth century (Anderson, 1981). The large proportion of dog bones in “natural” sites (i.e., no clear archaeological context) in northern New Zealand (Table 1) potentially supports this hypothesis (however, see Breadth and intensity of impacts below). Feral populations may have been at low abundance as seen with apex predators (Smith, 2015). Potts et al. (2013) raise the possibility that the monstrous or aggressive dogs in some Māori oral traditions may reflect concerns about kurī that had gone wild and beyond human control.
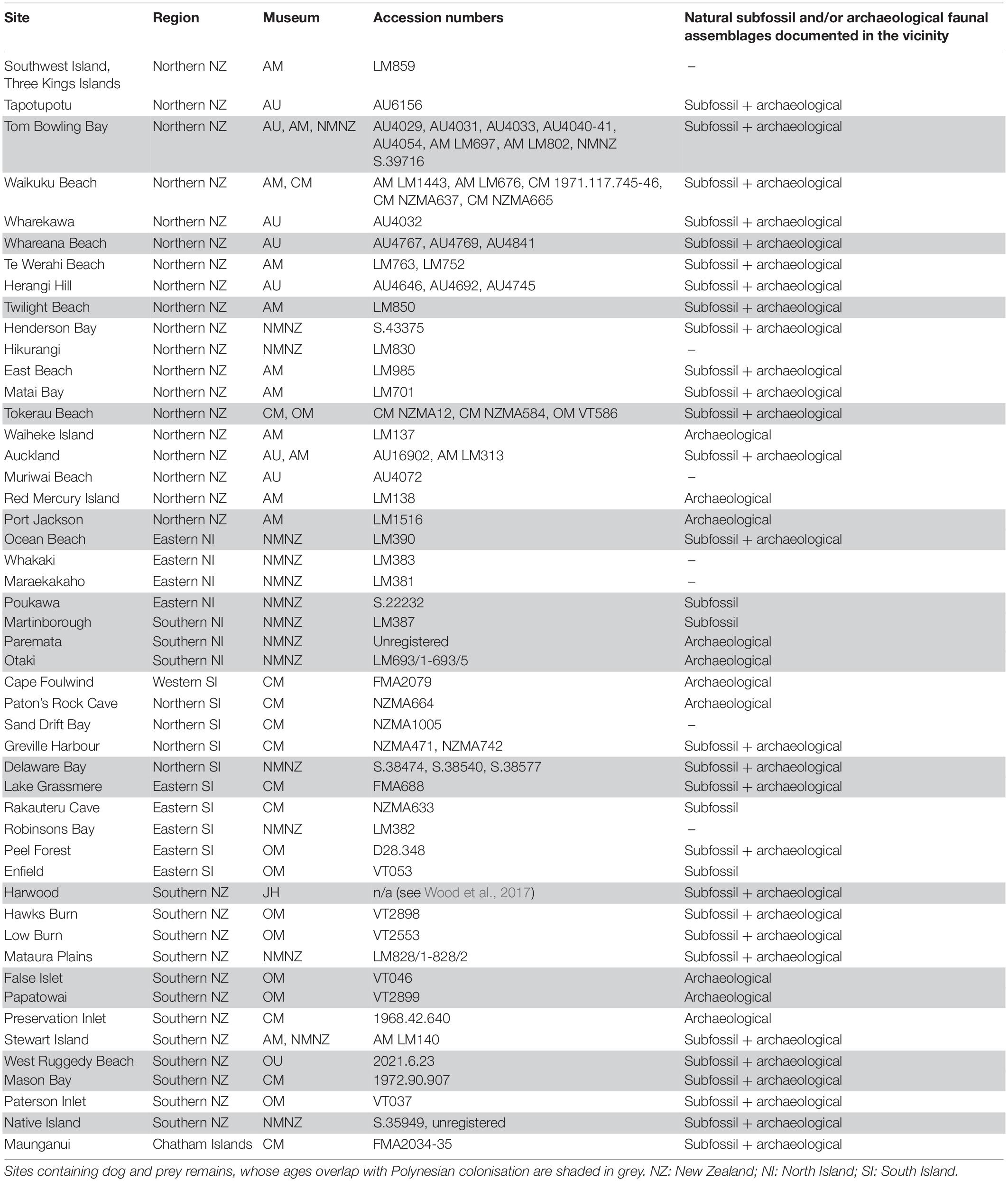
Table 1. Sites where dog remains have been found that are housed in the vertebrate fossil natural history collections at the University of Auckland Geology Department (AU), Auckland Museum (AM), the National Museum of New Zealand Te Papa Tongarewa (NMNZ), Canterbury Museum (CM), Otago Museum (OM), the University of Otago Zoology Department (OU), and the Jill Hamel private collection (JH).
By the 1830s, with the introduction of European breeds to New Zealand, overall dog numbers appear to have increased to the point of being considered a nuisance. Dogs had been brought to New Zealand by Europeans as early as Captain Cook’s voyages in the late eighteenth century (Clark, 1995). Hunting dogs in particular were subsequently introduced relatively quickly by sealers and whalers (Wakefield, 1845). Earle (1832) considered dogs to be the worst introduction by Europeans, due to their rapid increase and the injuries they caused to other animals. Packs of wild dogs present in the mid-19th century appear to be European dogs (e.g., Thomson, 1859) which in the South Island coincide with the arrival of flocks of sheep in central regions. Dieffenbach (1843) observed that a native dog could not bring down a sheep (presumably because of its small size), but that cross-breeds and introduced dogs would do so. By the mid-1800s, kurī were no longer recognisable as a distinct breed, having been subsumed within the burgeoning European dog population (Clark, 1995) through interbreeding and genetic swamping.
Potential Impacts of Kurī
Predation
Studies of free-ranging dog populations from around the world have identified the importance of food derived from human activities in the dogs’ diet (Vanak and Gompper, 2009). This includes deliberate feeding, and scavenged food refuse (plants and animals), remains of livestock and other carcasses, and human (and potentially other dogs, and seal and sea lion) faeces. Some studies have documented dogs killing and feeding on wildlife, but as Vanak and Gompper (2009) point out these studies are generally focused on the effects of predation on the prey species, rather than the overall picture of dog foraging ecology. They argue that reliance on human-derived materials is typical, even when wildlife is also killed and consumed. These human-derived food subsidies can result in long-term negative impacts on direct prey species, which in turn may have indirect flow on effects on the abundance of other species within the ecosystem (Ritchie et al., 2014; Newsome et al., 2015). In New Zealand, kurī were a recent novel predator into a naive ecosystem (Figure 3). Subsidising of their food resources through “moa-hunting” activities no doubt resulted in a rapid population increase of kurī, as evidenced by their presence in early archaeological sites across New Zealand (Davidson, 1987; Figure 2B). The loss of big game species like moa, seals, and sea lions within a few hundred years of Polynesian arrival (Figure 1) would have potentially exacerbated the effects of kurī on small and medium-sized birds and reptiles, hastening their extinction.

Figure 3. The Polynesian dog or kurī (Canis familiaris) (A) and its potential prey (B–L): Herpetofauna including geckos, skinks, frogs, and tuatara (Sphenodon punctatus, B); flightless palaeognathous birds such as kiwi (Apteryx spp., C) and extinct flightless moa chicks or juveniles (D); large flightless rails like the extinct adzebill Aptornis spp. (E) and the extant takahē/extinct moho Porphyrio spp. (F); flightless parrots like the kākāpō Strigops habroptilus (G); waterfowl including several species of extinct duck (e.g., Scarlett’s duck Malacorhynchus scarletti, H) and the flightless extinct Cnemiornis goose (I); seabirds such as cavity or burrow nesting (e.g., extant tawaki Fiordland crested penguin (J) Eudyptes pachyrhynchus) or sedentary species (e.g., extant/extinct blue-eyed shags (K) Leucocarbo spp.); pinnipeds (especially pups or females) including kekeno fur seals (L) Arctocephalus forsteri, and extinct lineages of rāpoka sea lions and ihupuku southern elephant seals. Artwork by Paul Martinson from Tennyson and Martinson (2007) © Te Papa CC BY-NC-ND 4.0. Image of moa chick © Te Papa CC BY 4.0. Image of blue-eyed shag by Philip Griffin (New Zealand Birds Online).
Even with a possible foraging focus on human-derived food, the presence of flightless or flight-reduced ground-living birds, lizards, and seals and sea lions with no “fight or flight” response in New Zealand is likely to have provided a tempting target for kurī (Figure 3). New Zealand birds evolved in the absence of mammalian predators, which hunt by smell. Instead, the top predators in the pre-human ecosystem were raptorial birds (e.g., Haast’s eagle and Eyles’ harrier) that hunt by sight. Consequently, many New Zealand birds have camouflage plumage (e.g., Rawlence et al., 2009) and freeze when confronted by predators (e.g., kākāpō). In addition, on predator-free islands flighted birds often spend a significant amount of time foraging on the ground (e.g., tīeke saddleback on Tiritiri Matangi Island, kōkako on Hauturu/Little Barrier Island, and kererū New Zealand pigeon on Kapiti Island; NJR and Alan Tennyson pers. obs.). Modern anecdotal evidence from single events suggests the potential severity of the impact of dogs on naive avifauna. Taborsky (1988) documented the devastating consequences of a single unrestrained dog in the Waitangi State Forest, which killed at least 23 kiwi over a six-month period. The total number killed is thought to have been as many as 500, which represented half of the total population at that time. In Tasmania, a single attack in 2008 by a dog or dogs resulted in the death of 30 little blue penguins (Holderness-Roddam and McQuillan, 2014). Doherty et al.’s (2017) review of the impacts of dogs on threatened species found that they have contributed to 11 vertebrate extinctions and are a threat to at least 188 species worldwide. Predation is the most frequently reported impact, with the Pacific islands being one of the regions with the most species affected. Outside of canids, the introduction/translocation of novel mammalian predators to island ecosystems can result in the extinction of local burrowing seabird colonies (e.g., Tasmanian devil introduction to Maria Island; Lu, 2021).
The vulnerability of animals to predation may change throughout their lifetimes. Ground-dwelling kiwi are susceptible to predation by dogs (Figure 3) across all life history stages (eggs, chicks, juveniles, and adults), however, a modern study of introduced mammalian predation of brown kiwi and roa great spotted kiwi suggests that the greatest predation of kiwi by dogs takes place on adult birds (McLennan et al., 1996). As with the Waitangi State Forest event, predation by dogs in the forest was found to be unpredictable and episodic, but with drastic results on the breeding population. The rate of dog predation on eggs and adult birds was also found to be much lower than that of chicks and juveniles, although the latter were decimated by mustelids (McLennan et al., 1996). In pinnipeds (seals and sea lions), attacks by dogs are common, even resulting in death (e.g., Boren, 2008; Department of Conservation, 2014; Houseman, 2020; Kerr-Lazenby, 2021). While male kekeno fur seals, rāpoka sea lions and ihupuku southern elephant seals would no doubt have been outside the prey size range for kurī, pups and females (especially while nursing) would have been particularly vulnerable to predation (Figure 3). If a female sea lion is killed, it’s unborn and dependant pups will also die. Modelling shows these slow-breeding pinnipeds could not withstand even low levels of subsistence hunting or predation resulting in their rapid extinction (Rawlence et al., 2016a; Waters et al., 2017).
As well as immediate fatalities, predatory behaviour can also result in chasing or seabird colony disturbance or abandonment (especially for disturbance prone seabirds), survivable injuries or severe injuries that ultimately result in death sometime after the predation attempt. Studies have demonstrated the negative effects of survivable chasing events, such as behavioural changes and physiological stress (e.g., Lima, 1998; Clinchy et al., 2013).
Elsewhere in the world dogs also compete with other carnivores directly for prey and as scavengers for carrion (e.g., Schlacher et al., 2015; Hansford et al., 2021). The pre-human New Zealand situation is different as there were no terrestrial mammalian carnivores, with their roles replaced by flighted and flightless birds including Haast’s eagle, Eyles’ harrier, adzebill, and the New Zealand raven Corax antipodum (e.g., Tennyson and Martinson, 2007; Scofield et al., 2017). It is possible that kurī competed with these species for prey and carrion.
While it has been hypothesised that kiore had the greatest impact on New Zealand’s small avian (e.g., wrens; Tennyson and Martinson, 2007) and reptilian fauna, with human hunting initially focusing on the large megafauna (e.g., moa), it is probable that kurī filled the ecological niche of a predator of medium sized ground-dwelling birds (and in some cases opportunistic predation of ground-foraging volant birds) such as the diverse waterfowl assemblage (goose, ducks, mergansers, swan), but also moa chicks and juveniles (and potentially the smaller males of some moa species), adzebill, takahē/moho (and other rails), kākāpō, kiwi, and ground-nesting seabirds like penguins, shags and burrowing petrels (Figure 3). This probably also included predation of the eggs of ground nesting birds. It is likely that kurī contributed to the extinction of at least three seabirds including Waitaha penguin Megadyptes waitaha (Boessenkool et al., 2009), Kōhatu shag Leucocarbo septentrionalis (Rawlence et al., 2017b) and Scalett’s shearwater Puffinus spelaeus (Holdaway and Worthy, 1994). Given the presence of reptiles in the stomach contents of introduced mammalian predators like stoats (McAulay et al., 2020), it is probable that kurī could have also preyed on skinks, geckos, frogs and tuatara (Figure 3) – there are records from the nineteenth century of dogs killing escaped captive tuatara (Cree, 2014). At least one species of giant skink (Worthy, 1991) and three frogs are known to have gone extinct prior to European colonisation, with several species only surviving on offshore islands (e.g., Scarsbrook et al., 2021). Indeed, tuatara were effectively extinct on the mainland before European arrival and now only survive on islands without kiore and dogs (Cree, 2014).
Habitat Disturbance
Habitat disturbance by potential predators can also result in behavioural changes in animals, which may ultimately result in lowered reproductive success and negative consequences at a population level (for a review see Twardek et al., 2017). The intensity, frequency and duration of disturbance may result in different types and severity of impacts (Hill et al., 1997). Low level continuous noise, for example, may be tolerated by birds, and seals and sea lions, over time, while episodic “startling” events can result in displacement and ultimately avoidance of a location.
Impacts documented internationally from the presence of modern dogs include increased nest vigilance in coots (Randler, 2006), and changes in spatial distribution of pudus (Silva-Rodriguez and Sieving, 2012) and bandicoots (Carthey and Banks, 2012). Even the activity of dog-walkers passing through a woodland area resulted in a 35% decrease in bird diversity and a 41% decrease in bird abundance over time (Banks and Bryant, 2007). Many of New Zealand’s seabird, seal and sea lion colonies are prone to disturbance, which can result in colony abandonment and failure of breeding seasons (e.g., Lord et al., 2001; McConkey et al., 2002; Rawlence et al., 2016a, b; Allott, 2021).
Pathogen Transmission
Throughout history, human movement around the world has been associated with the spread of zoonotic diseases, whether viral, bacterial, microbial or parasitic (Spyrou et al., 2019). As humans and commensal animals move into new areas, the chance of zoonotic disease transfer is heightened [e.g., pre-Columbian seal tuberculosis in Amerindian populations (Bos et al., 2014) or the extinction of the endemic Christmas Island rat due to an introduced trypanosome parasite from black rats; Wyatt et al., 2008]. Recent research on the age and transmission of tuberculosis in New Zealand suggests that it may have been introduced prior to European arrival, and vectors for transmission could include humans, commensal and wild animals (McDonald et al., 2020). To date, eggs from two parasitic helminth taxa (Caprillaria cf. hepatica and Toxocara canis), whose intermediate hosts also include rats (e.g., Dubinsky et al., 1995), have been found in kurī coprolites (Irwin, 2004).
Assistance With Human Hunting
The involvement of dogs in human hunting activities has been documented ethnographically in many parts of the world, and this collaborative behaviour may have been part of a complex set of circumstances that contributed to early dog domestication. Dogs have been used to increase hunting effectiveness across a variety of environments and to target a broad spectrum of prey species, from small rodents, reptiles, ratites, monkeys, marsupials, deer, and boar, to other carnivores [see Koungoulos and Fillios (2020) for a review]. Ethnographic accounts of dogs and human hunting often assume a priori that the presence of dogs has a positive effect on hunting success, but this is not demonstrated empirically (Lupo, 2017). In addition, some of this ethnographic data relates to hunting dogs that have been bred over the last 500–600 years for particular characteristics to improve hunting success, such as scent or sight hounds. Although these characteristics may be selectively bred, dogs’ hunting performance is also affected by their life experience (Udell et al., 2014). Not all dogs will possess these specialised characteristics. Nonetheless, Koungoulos and Fillios’s (2020) review of the use of dingoes for hunting found that although they were not specifically trained to hunt, their general instincts were still beneficial. Hunting with dogs is often a non-selective method – dogs may target prey indiscriminately, including non-desirable animals, juveniles or females with young (Koster, 2008; Koster and Noss, 2014).
Lupo’s (2017) review of ethnographic evidence for the use of dogs for hunting and the implications for productivity suggests that although dogs can influence hunting productivity in some circumstances, overall this is highly variable and does not apply to all prey types. Dogs’ senses and abilities can, however, complement those of human hunters, resulting in encounter rates that differ from those of hunters working without dogs, particularly the detection of nocturnal or burrowing prey (Koster and Noss, 2014). The circumstances where dogs perform best appear to be where dogs are introduced as novel predators (as in New Zealand; Figures 1, 3), used in packs, or in association with a new technology such as firearms (Lupo, 2017).
Any advantages to human hunting conferred by the presence of dogs in New Zealand are impossible to disentangle, as both species arrived in New Zealand at the same time (Figure 1). It is possible, however, that kurī did increase human hunting productivity, by decreasing search costs and improving encounter rates by finding and flushing out or holding at bay ground-based species (Lupo, 2017), such as kiwi, weka, kākāpō, seals and sea lions, and possibly reptiles. It may have been more energetically efficient for smaller kurī to hunt these prey than humans, and in turn, they would obtain a greater nutritional benefit in terms of relative body size than larger humans. Archaeological evidence from faunal remains certainly suggests Māori hunted the entire ontogenetic size range of sea lions (Rawlence et al., 2016a), and may have utilised kurī for assistance. Indeed, there is ethnographic evidence dating from the second-half of the nineteenth century for the use of dogs to hunt ground birds in New Zealand (e.g., weka, pūkeko, kākāpō; Beattie, 1920, 1939) but the antiquity of this practice is not clear. The use of kurī in moa-hunting activities has also been hypothesised (Anderson, 1981), but as this behaviour leaves little or no trace in the archaeological record the investigation of such hypotheses is challenging. Interestingly, Koungoulos and Fillios (2020) document the use of dingoes in Australia as part of game-drives to hunt large prey such as kangaroos, emu and wallaby. These were collaborative hunts, involving men, woman and children which resulted in large numbers of animals being caught.
Breadth and Intensity of Impacts
Unlike kiore, which spread rapidly throughout the New Zealand environment (e.g., presence of rat-gnawed seeds in natural palaeoenvironmental archives; Wilmshurst and Higham, 2004) and have been found in natural subfossil sites (e.g., caves, laughing owl roost sites; Holdaway and Worthy, 1996; Wilmshurst et al., 2008), the distribution of kurī appears to be highly correlated with human settlements and activities (Figure 2B). Furthermore, kiore consume a wide range of different flora and fauna, including seeds, plants, invertebrates and birds’ eggs, and are hypothesised to have had a much broader impact across the New Zealand ecosystem (Rayner et al., 2007; Tennyson and Martinson, 2007; Ismar et al., 2014). The intensity of impacts of kurī on native biota is therefore likely to have been more restricted than kiore, and to have varied across the country in accordance with underlying patterns of human movement, settlement and land use (Figure 2B). As with their human counterparts, kurī may have undergone a period of adaptation to temperate New Zealand from their tropical East Polynesian homeland. There is evidence from macro- and micro-scopic, and genetic analysis of coprolites, for the consumption of cultivated plants, fish and birds by kurī (Clark, 1997; Irwin, 2004; Wood et al., 2016).
During the first human colonisation-era, settlements in New Zealand were predominantly situated on the coast, often in association with river and estuary mouths (Figure 2B). Major river valleys were also used to access inland areas in southern New Zealand. Dog bones are numerous in these early archaeological sites (that also contain the remains of prey species), suggesting sizable kurī populations (Davidson, 1987; Greig et al., 2018). The results of biomolecular analyses of ancient mitochondrial genomes indicate that there was a limited introduction of dogs to New Zealand, but that these dogs and their descendants were transported rapidly around the country (Greig et al., 2018). The distribution of pre-European archaeological sites, as a proxy for the geographic range of kurī (Figure 2B), effectively overlaps with the distribution of potential prey species (Worthy and Holdaway, 2002). Numerous natural subfossil sites contain faunal remains (including moa, medium-sized ground-dwelling birds, seals and sea lions, and tuatara) whose ages overlap with the arrival of Polynesians in New Zealand (e.g., Worthy, 1998a; Brook, 2000; Collins et al., 2013; Rawlence et al., 2017b; Wood et al., 2017; Verry et al., 2021; Table 1 and Figure 2C). However, the co-occurrence or not of dog and prey species within a single site should not be used as evidence for and against potential predation (e.g., Davidson, 2012 cf. Surovell and Grund, 2012), given taphonomic biases (e.g., Seersholm et al., 2018), and the geographically widespread overlap of kurī and prey species.
The impact of kurī is likely to have been intense in the vicinity of the early villages, and possibly along inland routes (i.e., widespread but relatively localised impact compared to widespread non-localised impact by kiore). Habitat modification through human-induced changes to indigenous forest cover from widespread burning, particularly in southern New Zealand, with a commensurate increase in grasslands (McWethy et al., 2014) may have also affected the intensity of kurī predation. Habitat changes, including forest fragmentation and increased edge habitats, can facilitate easier access for dogs to hunt in more complex environments (Lacerda et al., 2009; Ritchie et al., 2014). Following the extinction of moa (Figure 1) human population density in southern Aotearoa New Zealand was markedly reduced until the late 1700s, and human presence was structured around seasonal resource acquisition (Jacomb et al., 2010; Rawlence et al., 2015a; Waters et al., 2017).
By the mid-nineteenth century packs of European, and possibly mixed ancestry, feral dogs were becoming a problem in New Zealand (Clark, 1995). To date, no genetically and morphologically verified kurī remains have been reported outside of archaeological contexts. However, our review of New Zealand natural history collections (specifically fossil vertebrate collections at the University of Auckland Geology Department, Auckland Museum, the National Museum of New Zealand Te Papa Tongarewa, Canterbury Museum, Otago Museum, and the University of Otago Zoology Department) presented here has identified numerous locations where dog specimens have been found without clear archaeological context (see Figure 2C and Table 1). It is not known whether these dog remains are from European or mixed-breed dogs or kurī. The locations comprise predominantly time-averaged sand dune deposits and some cave deposits (not pitfall traps; Worthy and Holdaway, 2002), the majority of which could be partially associated with nearby archaeological middens or occupation sites (e.g., Twilight Beach, Ocean Beach, Delaware Bay, and Greville Harbour).
It would be expected that if kurī lived independently of humans in self-sustaining packs, as occurred in Australia with the dingo, that their remains could be found in natural subfossil sites. However, potential feral populations may be cryptic in the recent fossil record given hypothesised low population densities (cf. sizeable human-dependant populations), especially in isolated areas where taphonomic settings are not conducive to preservation (e.g., high rainfall Fiordland), compared to widespread kiore-gnawed seeds in sediment cores and kiore bones in natural subfossil deposits indicative of high abundance populations (e.g., Holdaway and Worthy, 1996; Wilmshurst and Higham, 2004; Wilmshurst et al., 2008; Wilmshurst and Carpenter, 2020). It may be that in these isolated, rugged areas where human population density was lower (e.g., Jacomb et al., 2010; Waters et al., 2017) that the combined effects of kurī, potentially exhibiting pack hunting behaviour, and kiore, had a greater impact on native species than human hunting ever did.
Future Challenges
Our review has identified a variety of potential impacts to indigenous fauna; direct predation, competition, habitat disturbance, pathogen transmission, and assistance with human hunting. We hypothesise that, given the close association between kurī and Māori, these impacts would have been widespread but relatively localised around Māori settlement or movement patterns (Figure 2B) (though we cannot discount the possibility of pre-European feral kurī populations), and would have significantly amplified the impact of humans on New Zealand’s biodiversity. Furthermore, unlike kiore, kurī impacts would have been focused on a sub-set of New Zealand’s avifauna, herpetofauna, and seals and sea lions, particularly mid-sized taxa (Figure 3). To obtain a full picture this impact, several avenues of future research need to be undertaken.
(1) Coprolites, attributed to “kurī” on the basis of size (and in some cases due to the presence of chewed bone fragments), have been found in numerous midden contexts. However, these coprolites could also be potentially attributed to other carnivores such as sea lions or omnivores like humans, especially given the dietary overlap between kurī and people (e.g., Horrocks et al., 2002, 2003; Wood et al., 2016). The use of ancient DNA is increasingly highlighting its utility as a genetic tool for the study of palaeodiet (e.g., Bon et al., 2012; Wood et al., 2020). Multidisciplinary analyses of genetically confirmed kurī coprolites (ancient DNA, macro- and micro-scopic, isotopic, palaeoproteomics) should be used to test hypotheses about geographical and temporal changes in kurī diet (and as a proxy for human diet), and how this reflects wider patterns in the previously recognised ecosystem change in New Zealand. These analyses should be conducted in conjunction with archaeozoological analysis of midden assemblages (including bulk bone metabarcoding of non-diagnostic material; e.g., Seersholm et al., 2018) and regional pre-human palaeofaunal surveys (e.g., Worthy, 1998b), especially of understudied taxa (e.g., small birds, herpetofauna), and modelling of Māori movement across the environment through time (e.g., is there a differential decline in ground-nesting birds?). Because kurī are closely associated with people, it may always be difficult to distinguish between hunting versus scavenging.
(2) Ancient DNA analysis of kurī bones and coprolites, and sedimentary archives from archaeological sites, focusing on the morphological and genetic identification of zoonoses (e.g., Irwin, 2004; Wood et al., 2013; Witt et al., 2021), could potentially shed light on this understudied aspect of faunal impact (e.g., did dogs bring zoonoses to Aotearoa, and were kurī a vector between seals, sea lions and people for tuberculosis?).
(3) To resolve the debate of whether there were feral pre-European kurī populations, and whether and when kurī had an independent impact of New Zealand’s biodiversity, multiproxy morphometric, genetic, radiocarbon and isotopic analysis of “natural” dog remains in sites with no clear archaeological context should be undertaken to determine if these are kurī or European dogs (or even hybrids), and feral or human-dependant dogs (e.g., are there differences in diet?) (e.g., Koungoulos, 2020; Runge et al., 2021; Witt et al., 2021; on dogs in other geographical contexts). Palaeontological excavations across New Zealand should also be aware of the potential for dogs in natural (i.e., non-cultural) subfossil sites. Integrated approaches incorporating palaeoenvironmental DNA techniques to detect ancient dog DNA in sedimentary archives should also be investigated (Rawlence et al., 2014; Dussex et al., 2021; Mitchell and Rawlence, 2021). It may not be possible to resolve this question but addressing the status of these dogs will allow more informed hypotheses to be made about the potential impact of feral kurī.
Conclusion
Kurī have largely been overlooked in contributing to the ecological consequences of Polynesian settlement of New Zealand as these impacts have been difficult to scientifically assess, with the majority of the research focusing on over-hunting, habitat destruction, and predation from kiore. Far from being “exonerated” we argue that kurī had the potential for a significant, widespread but relatively localised (cf. kiore) impact on New Zealand’s fauna. Given the behavioural characteristics of much of New Zealand’s avifauna (i.e., terrestrial, flight-reduced or flightless, ground nesting, slow breeding), they would have been highly vulnerable to predation by kurī. In this way, independent predation and as part of hunting trips, kurī mirrored and amplified the impact of people, especially we hypothesise for medium sized birds, herpetofauna, and seals and sea lions. Opportunities for future research will help resolve the remaining gaps in our knowledge of the impact of kurī on New Zealand biodiversity and on insular island ecosystems in general.
Data Availability Statement
The original contributions presented in the study are included in the article/supplementary material, further inquiries can be directed to the corresponding author/s.
Author Contributions
KG and NJR devised the study, analysed the data, and wrote the manuscript. Both authors contributed to the article and approved the submitted version.
Funding
NJR is funded by a Royal Society of New Zealand Marsden FastStart grant (16-UOO-045).
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Acknowledgments
We thank the University of Auckland Geology Department (Neville Hudson), Auckland Museum (Matt Rayner), the National Museum of New Zealand Te Papa Tongarewa (Alan Tennyson), Canterbury Museum (Sarah Murray, Johnathon Ridden, and Paul Scofield), and Otago Museum (Kane Fleury and Gerard O’Regan) for providing a list of dog remains in their fossil vertebrate collections. We also thank Alan Tennyson, Richard Walter, Robert Poulin, Mathew Campbell, Phil Seddon, and Yolanda van Heezik for helpful discussions and comments.
References
Allott, A. (2021). Suspected Dog Attack Causes 500 Critically Endangered Gulls to Abandon Nests. Wellington: STUFF.
Anderson, A. (1981). Pre-European hunting dogs in the South Island, New Zealand. New Zealand J. Arch. 3, 15–20.
Anderson, A., Allingham, B., and Smith, I. (1996). Shag River Mouth: The Archaeology of an Early Southern Maori Village. Research Papers in Archaeology and Natural History 27. Australia: Australian National University.
Banks, P. B., and Bryant, J. V. (2007). Four-legged friend or foe? Dog walking displaces native birds from natural areas. Biol. Lett. 3:20070374. doi: 10.1098/rsbl.2007.0374
Beattie, H. (1939). Tikao Talks: Traditions and Tales Told by Teone Taare Tika to Herries Beattie. Edinburgh: Dunedin.
Best, E. (1916). Maori Storehouses and Kindred Structures. Houses, Platforms, Racks, and Pits used for Storing Food, Etc. (No. 5). South Africa: Government Printer.
Boessenkool, S., Austin, J. J., Worthy, T. H., Scofield, R. P., Cooper, A., Seddon, P. J., et al. (2009). Relict or colonizer? Extinction and range expansion of penguins in southern New Zealand. Proc. Biol. Sci. 276, 815–821. doi: 10.1098/rspb.2008.1246
Bon, C., Berthonaud, V., Maksud, F., Labadie, K., Poulain, J., Artiguenave, F., et al. (2012). Coprolites as a source of information on the genome and diet of the cave hyena. Proc. Biol. Sci. 279, 2825–2830. doi: 10.1098/rspb.2012.0358
Boren, L. J. (2008). Seal callouts in the Kaikoura region involving the Department of Conservation. DOC Res. Dev. Ser 297, 1–26.
Bos, K. I., Harkins, K. M., Herbig, A., Coscolla, M., Weber, N., Comas, I., et al. (2014). Pre-Columbian mycobacterial genomes reveal seals as a source of New World human tuberculosis. Nature 514, 494–497. doi: 10.1038/nature13591
Brook, F. J. (2000). Prehistoric predation of the landsnail Placostylus ambagiosus (Stylommatophora: Bulimulidae), and evidence for the timing of establishment of rats in northernmost New Zealand. J. R. Soc. N. Z. 30, 227–241.
Bunce, M., Worthy, T. H., Phillips, M. J., Holdaway, R. N., Willerslev, E., Haile, J., et al. (2009). The evolutionary history of the extinct ratite moa and New Zealand Neogene paleogeography. Proc. Natl. Acad. Sci. U. S. A. 106, 20646–20651. doi: 10.1073/pnas.0906660106
Butler, J., Brown, W., and du Toit, J. T. (2018). Anthropogenic food subsidy to a commensal carnivore: the value and supply of human faeces in the diet of free-ranging dogs. Animals 8:67. doi: 10.3390/ani8050067
Carthey, A. J., and Banks, P. B. (2012). When does an alien become a native species? A vulnerable native mammal recognizes and responds to its long-term alien predator. PLoS One 7:31804. doi: 10.1371/journal.pone.0031804
Clark, G. R. (1995). The Kuri in Prehistory: a Skeletal Analysis of the Extinct Maori Dog. Ph.D. thesis. Dunedin: University of Otago.
Clark, G. R. (1997). Maori subsistence change: zooarchaeological evidence from the prehistoric dog of New Zealand. Asian Perspect. 36, 200–219.
Clinchy, M., Sheriff, M. J., and Zanette, L. Y. (2013). Predator-induced stress and the ecology of fear. Funct. Ecol. 27, 56–65.
Colenso, W. (1877). Notes, chiefly historical, on the ancient dog of the New Zealanders. Trans. Proc. R. Soc. N. Z. 1868:196.
Collins, C. J., Rawlence, N. J., Worthy, T. H., Scofield, R. P., Tennyson, A. J. D., Smith, I., et al. (2013). Pre-human New Zealand sea lion (Phocarctos hookeri) rookeries on mainland New Zealand. J. R. Soc. N. Z. 44, 1–16.
Cree, A. (2014). Tuatara: Biology and Conservation of a Venerable Survivor. Christchurch: Canterbury University Press.
Davidson, I. (2012). Peopling the last new worlds: the first colonisation of Sahul and the Americas. Quat. Int. 285, 1–29. doi: 10.1016/j.quaint.2012.09.023
Department of Conservation (2014). Dog Kills Seal Pup on Canterbury Beach. Wellington: Te Papa Atawhai.
Dieffenbach, E. (1843). Travels in New Zealand With Contributions to the Geography, Geology, Botany, and Natural History of That Country. London: J. Murray.
Doherty, T. S., Dickman, C. R., Glen, A. S., Newsome, T. M., Nimmo, D. G., Ritchie, E. G., et al. (2017). The global impacts of domestic dogs on threatened vertebrates. Biol. Conserv. 210, 56–59.
Dubinsky, P., Havasiova-Reiterova, K., Petko, B., Hovorka, I., and Tomasovicova, O. (1995). Role of small mammals in the epidemiology of toxocariasis. Parasitology 110, 187–193. doi: 10.1017/s0031182000063952
Dussex, N., Bergfeldt, N., Anca Prado, V., Dehasque, M., Molino, D. D., Ersmark, E., et al. (2021). Integrating multi-taxon palaeogenomics sedimentary ancient DNA to study past ecosystem dynamics. Proc. Biol. Sci. 288:20211252. doi: 10.1098/rspb.2021.1252
Earle, A. (1832). A Narrative of a Nine Months’ Residence in New Zealand, in 1827; Together With a Journal of a Residence in Tristan D’acunha, an Island Situated Between South America and the Cape of Good Hope. London: Longman.
Easton, L. J., Rawlence, N. J., Worthy, T. H., Tennyson, A. J. D., Scofield, R. P., Easton, C. J., et al. (2017). Testing species limits of New Zealand’s leiopelmatid frogs through morphometric analyses. Zool. J. Linn. Soc. 183, 431–444. doi: 10.1093/zoolinnean/zlx080
Fleming, C. A. (1962). The extinction of moas and other animals during the Holocene period. Notornis 10, 113–117.
Furey, L. (2002). Houhora: a Fourteenth Century Maori Village in Northland. Bulletin of the Auckland Museum 19. Auckland: Auckland War Memorial Museum.
Gemmell, N. J., Rutherford, K., Prost, S., Tollis, M., Winter, D., Macey, J. R., et al. (2020). The tuatara genome reveals ancient features of amniote evolution. Nature 584, 403–409.
Greig, K., Boocock, J., Allen, M. S., Matisoo-Smith, E., and Walter, R. (2018). Ancient DNA evidence for the introduction and dispersal of dogs (Canis familiaris) in New Zealand. J. Pac. Archaeol. 9, 1–10. doi: 10.30607/kvj.449025
Greig, K., and Walter, R. (2021). A re-assessment of the role of dogs (Canis familiaris) in early Aotearoa New Zealand. Hum. Ecol. doi: 10.1007/s10745-021-00285.2
Hansford, J. P., Lister, A. M., Weston, E. M., and Turvey, S. T. (2021). Simultaneous extinction of Madagascar’s megaherbivores correlates with late Holocene human-caused landscape transformation. Quat. Sci. Rev. 263:106996. doi: 10.1016/j.quascirev.2021.106996
Hartnup, K., Huynen, L., Te Kanawa, R., Shepherd, L. D., Miller, C. D., and Lambert, D. M. (2011). Ancient DNA recovers the origins of Maori feather cloaks. Mol. Biol. Evol. 28, 2741–2750. doi: 10.1093/molbev/msr107
Hill, D., Hockin, D., Price, D., Tucker, G., Morris, R., and Treweek, J. (1997). Bird disturbance: improving the quality and utility of disturbance research. J. Appl. Ecol. 34, 275–288. doi: 10.2307/2404876
Hixon, S. W., Douglass, K. G., Godfrey, L. R., Eccles, L., Crowley, B. E., Rakotozafy, L. M. A., et al. (2021). Ecological consequences of a millennium of introduced dogs on Madagascar. Front. Ecol. Evol. 9:689559. doi: 10.3389/fevo.2021.689559
Holdaway, R. N. (1999). “Introduced predators and avifaunal extinction in New Zealand,” in Extinctions in Near Time, ed. R. D. E. MacPhee (Berlin: Springer), 189–238. doi: 10.1007/978-1-4757-5202-1_9
Holdaway, R. N., Allentoft, M. E., Jacomb, C., Oskam, C. L., Beavan, N. R., and Bunce, M. (2014). An extremely low-density human population exterminated New Zealand moa. Nat. Commun. 5:5436. doi: 10.1038/ncomms6436
Holdaway, R. N., and Worthy, T. H. (1994). A new fossil species of shearwater Puffinus from the late Quaternary of the South Island, New Zealand, and notes on the biogeography and evolution of the Puffinus gavia superspecies. Emu 94, 201–215.
Holdaway, R. N., and Worthy, T. H. (1996). Diet and biology of the laughing owl Sceloglaux albifacies (Aves: Strigidae) on Takaka Hill, Nelson, New Zealand. J. Zool. 239, 545–572. doi: 10.1111/j.1469-7998.1996.tb05942.x
Holderness-Roddam, B., and McQuillan, P. B. (2014). Domestic dogs (Canis familiaris) as a predator and disturbance agent of wildlife in Tasmania. Aust. J. Environ. Manag. 21, 441–452.
Horrocks, M., Irwin, G. J., McGlone, M. S., Nichol, S. L., and Williams, L. J. (2003). Pollen, phytoliths and diatoms in prehistoric coprolites from Kohika, Bay of Plenty, New Zealand. J. Arch. Sci. 30, 13–20. doi: 10.1006/jasc.2001.0714
Horrocks, M., Jones, M. D., Beever, R. E., and Sutton, D. G. (2002). Analysis of plant microfossils in prehistoric coprolites form Harataonga Bay, Great Barrier Island, New Zealand. J. R. Soc. N. Z. 32, 617–628.
Hughes, J., and MacDonald, D. W. (2013). A review of the interactions between free-roaming domestic dogs and wildlife. Biol. Conserv. 157, 341–351. doi: 10.1111/zph.12142
Irwin, G. (ed.) (2004). Kohika: The Archaeology of a Late Māori Lake Village in the Ngāti Awa Rohe, Bay of Plenty, New Zealand. Auckland: Auckland University Press.
Ismar, S. M. H., Baird, K. A., Gaskin, C. P., Taylor, G. A., Tennyson, A. J. D., Rayner, M. J., et al. (2014). A case of natural recovery after the removal of invasive predators – community assemblage changes in the avifauna of Burgess Island. Notornis 61, 188–195.
Jacomb, C., Walter, R., and Jennings, C. (2010). Review of the archaeology of Foveaux Strait, New Zealand. J. Poly. Soc. 119, 25–59.
Kerr-Lazenby, M. (2021). DOC Warns Owners to Control Dogs as Seal Season Sets in. Wellington: STUFF.
Kinaston, R. L., Walter, R. K., Jacomb, C., Brooks, E., Tayles, N., Halcrow, S. E., et al. (2013). The first New Zealanders: patterns of diet and mobility revealed through isotope analysis. PLoS One 8:e64580. doi: 10.1371/journal.pone.0064580
Koster, J., and Noss, A. (2014). “Hunting dogs and the extraction of wildlife as a resource,” in Free-ranging dogs and wildlife conservation, ed. M. E. Gompper (Oxford: Oxford University Press), 265–285. doi: 10.7717/peerj.9906
Koster, J. M. (2008). Hunting with dogs in Nicaragua: an optimal foraging approach. Curr. Anthropol. 49, 935–944.
Koungoulos, L. (2020). Old dogs, new tricks: 3D geometric analysis of cranial morphology supports ancient population substructure in the Australian dingo. Zoomorphology 139, 263–275.
Koungoulos, L., and Fillios, M. (2020). Hunting dogs down under? On the Aboriginal use of tame dingoes in dietary game acquisition and its relevance to Australian prehistory. J. Anthropol. Archaeol. 58, 101146.
Lacerda, A. C. R., Tomas, W. M., and Marinho-Filho, J. (2009). Domestic dogs as an edge effect in the Brasília National Park, Brazil: interactions with native mammals. Anim. Conserv. 12, 477–487.
Leach, F., Quinn, C., Morrison, J., and Lyon, G. (2003). The use of multiple isotope signatures in reconstructing prehistoric human diet from archaeological bone from the Pacific and New Zealand. New Zeal. J. Arch. 23, 31–98.
Lima, S. L. (1998). Stress and decision making under the risk of predation: recent developments from behavioral, reproductive, and ecological perspectives. Adv. Stud. Behav. 27, 215–290.
Lord, A., Waas, J. R., Innes, J., and Whittingham, M. J. (2001). Effects of human approaches to nests of northern New Zealand dotterels. Biol. Conserv. 98, 233–240. doi: 10.1016/s0006-3207(00)00158-0
Louys, J., Braje, T. J., Chang, C. H., Cosgrove, R., Fitzpatrick, S. M., Fujita, M., et al. (2021). No evidence for widespread island extinctions after Pleistocene hominin arrival. Proc. Natl. Acad. Sci. U. S. A. 118:e2023005118. doi: 10.1073/pnas.2023005118
Lu, D. (2021). Tasmanian Devils Wipe out Thousands of Penguins on Tiny Australian Island. Kings Place: The Guardian.
Lupo, K. D. (2017). When and where do dogs improve hunting productivity? The empirical record and some implications from early Upper Paleolithic prey acquisition. J. Anthropol. Archaeol. 47, 139–151. doi: 10.1016/j.jaa.2017.05.003
McAulay, J., Seddon, P. J., Wilson, D. J., and Monks, J. M. (2020). Stable isotope analysis reveals variable diets of stoats (Mustela erminea) in the alpine zone of New Zealand. New Zeal. J. Ecol. 44, 3409.
McConkey, S. D., McConnell, H., Lalas, C., Heinrich, S., Ludmerer, A., McNally, N., et al. (2002). A northward spread in the breeding distribution of the New Zealand sea lion (Phocarctos hookeri). Aust. Mammal. 24, 97–106.
McConnell, J. R., Chellman, N. J., Mulvaney, R., Eckhardt, S., Stohl, A., Plunkett, G., et al. (2021). Hemispheric black carbon increase after the 13th-century Maori arrival in New Zealand. Nature 598, 82–85. doi: 10.1038/s41586-021-03858-9
McDonald, S. K., Matisoo-Smith, E. A., Buckley, H. R., Walter, R. K., Aung, H. L., Collins, C. J., et al. (2020). ‘TB or not TB’: the conundrum of pre-European contact tuberculosis in the Pacific. Philos. Trans. R. Soc. Lond. B Biol. Sci. 375:20190583. doi: 10.1098/rstb.2019.0583
McLennan, J. A., Potter, M. A., Robertson, H. A., Wake, G. C., Colbourne, R., Dew, L., et al. (1996). Role of predation in the decline of kiwi, Apteryx spp. in New Zealand. New Zeal. J. Ecol. 20, 27–35.
McWethy, D. B., Wilmshurst, J. M., Whitlock, C., Wood, J. R., and McGlone, M. S. (2014). A high-resolution chronology of rapid forest transitions following Polynesian arrival in New Zealand. PLoS One 9:e111328. doi: 10.1371/journal.pone.0111328
Mendelson, T. C., and Shaw, K. L. (2005). Sexual behaviour: rapid speciation in an arthropod. Nature 433, 375–376. doi: 10.1038/433375a
Mitchell, K. J., and Rawlence, N. J. (2021). Examining natural history through the lens of palaeogenomics. Trends Ecol. Evol. 36, 258–267. doi: 10.1016/j.tree.2020.10.005
Newsome, T. M., Dellinger, J. A., Pavey, C. R., Ripple, W. J., Shores, C. R., Wirsing, A. J., et al. (2015). The ecological effects of providing resource subsidies to predators. Glob. Ecol. Biogeogr. 24, 1–11. doi: 10.1111/geb.12236
Nogue, S., Santos, A. M. C., Birks, H. J. B., Bjorck, S., Castilla-Beltran, A., Connor, S., et al. (2021). The human dimension of biodiversity changes on islands. Science 372, 488–491. doi: 10.1126/science.abd6706
Oskarsson, M. C. R., Klutsch, C. F. C., Boonyaprakob, U., Wilton, A., Tanabe, Y., and Savolainen, P. (2012). Mitochondrial DNA data indicate an introduction through Mainland Southeast Asia for Australian dingoes and Polynesian domestic dogs. Proc. Biol. Sci. 279:20111395. doi: 10.1098/rspb.2011.1395
Perry, G. L. W., Wheeler, A. B., Wood, J. R., and Wilmshurst, J. M. (2014). A high-precision chronology for the rapid extinction of New Zealand moa (Aves, Dinornithiformes). Quat. Sci. Rev. 105, 126–135. doi: 10.1016/j.quascirev.2014.09.025
Potts, A., Armstrong, P., and Brown, D. (2013). A New Zealand book of beasts: animals in our culture, history and everyday life. Auckland: Auckland University Press.
Randler, C. (2006). Disturbances by dog barking increase vigilance in coots Fulica atra. Eur. J. Wild. Res. 52, 265–270.
Rawlence, N. J., Kardamaki, A., Easton, L. J., Tennyson, A. J. D., Scofield, R. P., and Waters, J. M. (2017a). Ancient DNA and morphometric analysis reveal extinction and replacement of New Zealand’s unique black swan. Proc. Biol. Sci. 284:20170876. doi: 10.1098/rspb.2017.0876
Rawlence, N. J., Till, C. E., Easton, L. J., Spencer, H. G., Schuckard, R., Melville, D. S., et al. (2017b). Speciation, range contraction and extinction in the endemic New Zealand King Shag complex. Mol. Phylogenet. Evol. 115, 197–209. doi: 10.1016/j.ympev.2017.07.011
Rawlence, N. J., Kennedy, M., Anderson, C. N. K., Prost, S., Till, C. E., Smith, I. W. G., et al. (2015a). Geographically contrasting biodiversity reductions in a widespread New Zealand seabird. Mol. Ecol. 24, 4605–4616. doi: 10.1111/mec.13338
Rawlence, N. J., Perry, G. L. W., Smith, I. W. G., Scofield, R. P., Tennyson, A. J. D., Matisoo-Smith, E. A., et al. (2015b). Radiocarbon-dating and ancient DNA reveal rapid replacement of extinct prehistoric penguins. Quat. Sci. Rev. 112, 59–65. doi: 10.1016/j.quascirev.2015.01.011
Rawlence, N. J., Lowe, D. J., Wood, J. R., Young, J. M., Churchman, G. J., Huang, Y. T., et al. (2014). Using palaeoenvironmental DNA to reconstruct past environments: progress and prospects. J. Quat. Sci. 29, 610–626. doi: 10.1002/jqs.2740
Rawlence, N. J., Scofield, R. P., McGlone, M. G., and Knapp, M. (2019). History repeats: large scale synchronous biological turnover in avifauna from the Plio-Pleistocene and Late Holocene of New Zealand. Front. Ecol. Evol. 7:158. doi: 10.3389/fevo.2019.00158
Rawlence, N. J., Collins, C. J., Anderson, C. N. K., Maxwell, J. J., Smith, I. W. G., Robertson, B. C., et al. (2016a). Human-mediated extirpation of the unique Chatham Islands sea lion and implications for the conservation management of remaining New Zealand sea lion populations. Mol. Ecol. 25, 3950–3961.
Rawlence, N. J., Scofield, R. P., Spencer, H. G., Lalas, C., Easton, L. J., Tennyson, A. J. D., et al. (2016b). Genetic and morphological evidence for two species of Leucocarbo shag (Aves, Pelecaniformes, Phalacrocoracidae) from southern South Island of New Zealand. Zool. J. Linn. Soc. 177, 676–694. doi: 10.1111/zoj.12376
Rawlence, N. J., Wood, J. R., Armstrong, K. N., and Cooper, A. (2009). DNA content and distribution in ancient feathers and potential to reconstruct the plumage of extinct avian taxa. Proc. Biol. Sci. 276, 3395–3402. doi: 10.1098/rspb.2009.0755
Rayner, M. J., Hauber, M. E., Imber, M. J., Stamp, R. K., and Clout, M. N. (2007). Spatial heterogeneity of mesopredator release within an oceanic island system. Proc. Natl. Acad. Sci. U. S. A. 104, 20862–20865. doi: 10.1073/pnas.0707414105
Ritchie, E. G., Dickman, C. R., Letnic, M., Vanak, A. T., and Gommper, M. (2014). “Dogs as predators and trophic regulators,” in Free-ranging dogs and wildlife conservation, ed. M. E. Gompper (Oxford: Oxford University Press), 55–68. doi: 10.1098/rspb.2015.1602
Runge, A. K. W., Hendy, J., Richter, K. K., Masson-MacLean, E., Britton, K., Mackie, M., et al. (2021). Palaeoproteomic analyses of dog palaeofaeces reveal a preserved dietary and host digestive proteome. Proc. Biol. Sci. 288:20210020. doi: 10.1098/rspb.2021.0020
Salis, A. T., Easton, L. J., Robertson, B. C., Gemmell, N., Smith, I. W. G., Weisler, M. I., et al. (2016). Myth or relict: does ancient DNA detect the enigmatic upland seal? Mol. Phylogenet. Evol. 97, 101–106. doi: 10.1016/j.ympev.2015.12.012
Scarsbrook, L., Sheratt, E., Hitchmough, R. A., and Rawlence, N. J. (2021). Skeletal variation in extant species enables systematic identification of New Zealand’s large, subfossil diplodactylids. BMC Ecol. Evol. 21:67. doi: 10.1186/s12862-021-01808-7
Schlacher, T. A., Weston, M. A., Lynn, D., Schoeman, D. S., Huijbers, C. M., Olds, A. D., et al. (2015). Conservation gone to the dogs: when canids rule the beach in small coastal reserves. Biodivers. Conserv. 24, 493–509. doi: 10.1007/s10531-014-0830-3
Scofield, P., Worthy, T., and Schlumpf, H. (2003). What birds were New Zealand’s first people eating? Wairau Bar’s avian remains re-examined. Rec. Canterb. Mus. 17, 17–35.
Scofield, R. P., Mitchell, K. J., Wood, J. R., De Pietri, V. L., Jarvie, S., Llamas, B., et al. (2017). The origin and phylogenetic relationships of the New Zealand ravens. Mol. Phylogenet. Evol. 106, 136–143. doi: 10.1016/j.ympev.2016.09.022
Seersholm, F. V., Cole, T. L., Grealy, A., Rawlence, N. J., Greig, K., Knapp, M., et al. (2018). Subsistence practices, past diversity, and anthropogenic impacts revealed by New Zealand-wide ancient DNA survey. Proc. Natl. Acad. Sci. U. S. A. 115, 7771–7776. doi: 10.1073/pnas.1803573115
Shaw, L. K., and Gillespie, R. G. (2016). Comparative phylogeography of oceanic archipelagos: hotspots for inferences of evolutionary processes. Proc. Natl. Acad. Sci. U. S. A. 113, 7986–7993. doi: 10.1073/pnas.1601078113
Silva-Rodriguez, E. A., and Sieving, K. E. (2012). Domestic dogs shape the landscape-scale distribution of a threatened forest ungulate. Biol. Conserv. 150, 103–110. doi: 10.1016/j.biocon.2012.03.008
Spyrou, M. A., Bos, K. I., Herbig, A., and Krause, J. (2019). Ancient pathogen genomics as an emerging tool for infectious disease research. Nat. Rev. Genet. 20, 323–340. doi: 10.1038/s41576-019-0119-1
Surovell, T. A., and Grund, B. S. (2012). The associational critique of Quaternary overkill and why it is largely irrelevant to the extinction debate. Am. Antiq. 77, 672–687.
Taborsky, M. (1988). Kiwis and dog predation: observations in Waitangi State Forest. Notornis 35, 197–202.
Thomson, A. S. (1859). The Story of New Zealand: Past and Present, Savage and Civilised Vol. I. London: J. Murray.
Twardek, W. M., Peiman, K. S., Gallagher, A. J., and Cooke, S. J. (2017). Fido, Fluffy, and wildlife conservation: the environmental consequences of domestic animals. Environ. Rev. 25, 381–395.
Udell, M. A. R., Ewald, M., Dorey, N. R., and Wynne, C. D. L. (2014). Exploring breed differences in dogs (Canis familiaris): does exaggeration or inhibition of predatory response predict performance on human-guided tasks? Anim. Behav. 89, 99–105. doi: 10.1016/j.anbehav.2013.12.012
Vanak, A. T., and Gompper, M. E. (2009). Dogs Canis familiaris as carnivores: their role and function in intraguild competition. Mammal. Rev. 39, 265–283.
Verry, A. J. F., Schmidt, M., and Rawlence, N. J. (2021). A partial skeleton provides evidence for the former occurrence of moa populations on Rakiura Stewart Island. New Zeal. J. Ecol. 46:3458.
Wakefield, E. J. (1845). Adventure in New Zealand. From 1839 to 1844. With Some Account of the Beginning of the British Colonization of the Islands Vol. I and II. Zealand Books. London: J. Murray.
Wallis, G. P., and Jorge, F. (2018). Going under down under? Lineage ages argue for extensive survival of the Oligocene marine transgression on Zealandia. Mol. Ecol. 27, 4368–4396. doi: 10.1111/mec.14875
Walter, R., Buckley, H., Jacomb, C., and Matisoo-Smith, L. (2017). Mass migration and the Polynesian settlement of New Zealand. J. World. Prehist. 30, 351–376. doi: 10.1007/s10963-017-9110-y
Wanner, H., Beer, J., Butikofer, J., Crowley, T. J., Cubasch, U., Fluckiger, J., et al. (2008). Mid- to Late Holocene climate change: an overview. Quat. Sci. Rev. 27, 1791–1828.
Waters, J. M., Fraser, C. I., Maxwell, J. J., and Rawlence, N. J. (2017). Did interaction between human pressure and Little Ice Age drive biological turnover in New Zealand? J. Biogeogr. 44, 1481–1490. doi: 10.1111/jbi.12991
Wilmshurst, J. M., Anderson, A. J., Higham, T. F. G., and Worthy, T. H. (2008). Dating the late prehistoric dispersal of Polynesians to New Zealand using the commensal Pacific rat. Proc. Natl. Acad. Sci. U. S. A. 105, 7676–7680. doi: 10.1073/pnas.0801507105
Wilmshurst, J. M., and Carpenter, J. K. (2020). Rodent detection and monitoring for conservation on islands: gnawed seeds provide reliable indicator of rodent presence. New Zeal. J. Ecol. 44:3398.
Wilmshurst, J. M., and Higham, T. F. G. (2004). Using rat-gnawed seeds to independently date the arrival of Pacific rats and humans in New Zealand. Holocene 14, 801–806. doi: 10.1191/0959683604hl760ft
Wilmshurst, J. M., Hunt, T. L., Lipo, C. P., and Anderson, A. J. (2011). High-precision radiocarbon dating shows recent and rapid human colonization in East Polynesia. Proc. Natl. Acad. Sci. U. S. A. 108, 1815–1820. doi: 10.1073/pnas.1015876108
Witt, K. E., Yarlagadda, K., Allen, J. M., Bader, A. C., Simon, M. L., Kuehn, S. R., et al. (2021). Integrative analysis of DNA, macroscopic remains and stable isotopes of dog coprolites to reconstruct community diet. Sci. Rep. 11:3113.
Wood, J. R., Crown, A., Cole, T. L., and Wilmshurst, J. M. (2016). Microscopic and ancient DNA profiling of Polynesian dog (kurī) coprolites from northern New Zealand. J. Arch. Sci. Rep. 6, 496–505. doi: 10.1016/j.jasrep.2016.03.020
Wood, J. R., Richardson, S. J., McGlone, M. S., and Wilmshurst, J. M. (2020). The diets of moa (Aves: Dinornithiformes). New Zeal. J. Ecol. 44:3397.
Wood, J. R., Scofield, R. P., Hamel, J., Lalas, C., and Wilmshurst, J. M. (2017). Bone stable isotopes indicate a high trophic position for New Zealand’s extinct South Island adzebill (Aptornis defossor) (Gruiformes: Aptornithidae). New Zeal. J. Ecol. 41, 240–244.
Wood, J. R., Wilmshurst, J. M., Rawlence, N. J., Bonner, K. I., Worthy, T. H., Kinsella, J. M., et al. (2013). A megafauna’s microfauna: gastrointestinal parasites of New Zealand’s extinct moa (Aves: Dinornithiformes). PLoS One 8:e57315. doi: 10.1371/journal.pone.0057315
Worthy, T. H. (1991). Fossil skink bones from Northland, New Zealand, and description of a new species of Cyclodina, Scincidae. J. R. Soc. New Zeal. 21, 329–348. doi: 10.1080/03036758.1991.10420831
Worthy, T. H. (1998a). A remarkable fossil and archaeological avifauna from Marfells Beach, Lake Grassmere, South Island, New Zealand. Rec. Canterb. Mus. 12, 79–176.
Worthy, T. H. (1998b). Quaternary fossil faunas of Otago, South Island, New Zealand. J. R. Soc. New Zeal. 28, 421–521. doi: 10.1080/03014223.1998.9517573
Worthy, T. H., and Holdaway, R. N. (2002). Lost World of the Moa. Christchurch: Canterbury University Press.
Wyatt, K. B., Campos, P. F., Gilbert, M. T. P., Kolokotronis, S. O., Hynes, W. H., DeSalle, R., et al. (2008). Historical mammal extinction on Christmas Island (Indian Ocean) correlates with introduced infectious disease. PLoS One 3:3602. doi: 10.1371/journal.pone.0003602
Keywords: birds, diet, dog, habitat disturbance, hunting, Māori, predation, Polynesia
Citation: Greig K and Rawlence NJ (2021) The Contribution of Kurī (Polynesian Dog) to the Ecological Impacts of the Human Settlement of Aotearoa New Zealand. Front. Ecol. Evol. 9:757988. doi: 10.3389/fevo.2021.757988
Received: 13 August 2021; Accepted: 08 November 2021;
Published: 15 December 2021.
Edited by:
G. Lynn Wingard, United States Geological Survey (USGS), United StatesReviewed by:
Christopher Richard Dickman, The University of Sydney, AustraliaCarlo Meloro, Liverpool John Moores University, United Kingdom
Copyright © 2021 Greig and Rawlence. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Karen Greig, a2FyZW4uZ3JlaWdAb3RhZ28uYWMubno=