 Amy V. Vasquez
Amy V. Vasquez Eric M. Wood
Eric M. Wood- 1Department of Biological Sciences, California State University Los Angeles, Los Angeles, CA, United States
- 2Department of Earth and Planetary Sciences, Johns Hopkins University, Baltimore, MD, United States
Urban parks provide amenities that support both human and animal communities. However, parks are often unevenly distributed within cities. One metric used to assess the distribution of parks to the public is termed the Park Score. The Park Score is an approach to measure access, acreage, investment, and amenities, and is designed to understand a city’s needs for greenspace, with a major focus on public health. In addition to issues related to public health, a disparity in the distribution of urban parks may pose a barrier for wildlife, such as birds. Yet, this remains unclear. We designed a study to quantify the role of parks in providing a refuge for birds across a park-needs gradient in Greater Los Angeles (LA), a metropolis with one of the lowest park scores in the United States. We had two objectives to address our goal. First, we quantified patterns in habitat features and avian communities within and adjacent to parks. Second, we analyzed relationships among habitat features within and adjacent to parks on avian abundance. We sampled birds and habitat features at 48 parks across a park-needs gradient in L.A. from October to March of 2017/2018 and 2018/2019. We found three lines of evidence supporting the refugia effect of parks. First, habitat features within parks were similar between low- and high-needs areas of LA, and this likely influenced avian abundance patterns, which were also alike. Second, avian communities were generally similar across the park-needs gradient, where parks in high-needs areas harbored birds affiliated with forest and shrub ecosystems. Third, bird abundance patterns were related to numerous habitat features within parks, regardless of where parks occurred in the city. The patterns we uncovered were opposite to what is found in residential areas (i.e., luxury effect), suggesting that parks provide important habitat for birds, whether in high- or low-needs sections of LA. Our results stress the role of parks as refugia in park-poor areas because they provide habitat in otherwise inhospitable urban conditions. Continued investment in park development in high-needs areas can thus potentially be a win-win when considering the benefits to people and birds.
Introduction
Urban ecosystems are densely populated, human-dominated environments embedded within a mosaic of natural and anthropogenically modified landscapes (Cadenasso and Pickett, 2008; Grimm et al., 2008). Cities and other urban environments are the primary living areas of humans, containing approximately 55% of the world’s population (United Nations, 2018). In addition to providing conditions amiable to people, urban ecosystems also support varying levels of biodiversity (Aronson et al., 2014; Lepczyk et al., 2017). For example, cities tend to harbor an unusually high diversity of plants and a lower, more homogenous diversity of wildlife (Helden and Leather, 2004; Alvey, 2006; Colding and Folke, 2009; Beninde et al., 2015; Talal and Santelmann, 2019). As cities continue to develop and sprawl to accommodate increasingly dense human populations, there is a growing concern about the degradation of green spaces within the urban landscape (Vallejo et al., 2009; Wu, 2010; Xu et al., 2018). Given the loss and fragmentation of habitat, and the increase in land-cover change across the globe, biodiversity must either adapt or risk extirpation in the face of urbanization (McKinney, 2002; Seress and Liker, 2015; La Sorte et al., 2018; Young et al., 2019).
Urban planners often intentionally, or unintentionally, design and include features that benefit biodiversity and the environment. One such feature that is prominent in urban areas is urban parks. The collection of managed amenities in parks, such as trees, shrubs, and lawn cover, is often positively correlated with wildlife (Hermy and Cornelis, 2000; Khera et al., 2009; Nielsen et al., 2014). Nevertheless, parks are usually unevenly distributed across cityscapes. One metric that cities in the U.S. use to assess the value of their parks to the public is the Park Score (Trust for Public Land, 2021). The Park Score measures access, acreage, investment, and amenities, and is designed to understand a city’s needs for greenspace, with a major focus on public health (Trust for Public Land, 2021). Cities with high park scores often have parks distributed equitably across the urban environment, which carries over to benefit the human population. On the other hand, cities with low park scores face the opposite patterns, with large swaths of a metropolis being park-poor, often in lower-income residential communities (Trust for Public Land, 2021). The negative effects of low park scores are correlated with a host of public health issues in low-income communities ranging from higher rates of diabetes and obesity to increased crime and lack of access to nature (Lovasi et al., 2013; Han et al., 2018). Further, given the disparity in habitat conditions across socioeconomic gradients in urban areas (e.g., Wood and Esaian, 2020), cities with low park scores likely also face considerable challenges in providing habitat for wildlife throughout their boundaries.
The ‘luxury effect’ is a socio-ecological hypothesis that states that the amount and diversity of vegetation and wildlife in the urban environment follow general wealth patterns (Hope et al., 2003; Leong et al., 2018; Schell et al., 2020; Magle et al., 2021). Evidence of the luxury effect has been found in many cities (Luck et al., 2009; Clarke et al., 2013; Jenerette et al., 2013; Avolio et al., 2015; Schwarz et al., 2015) and in a variety of green spaces across the urban landscape, such as community gardens (Clarke and Jenerette, 2015) and residential areas (Wang et al., 2015). The luxury effect has similarly been shown to predict patterns of wildlife diversity in cities, with low-income areas being less biodiverse than wealthier counterparts (Kinzig et al., 2005; Strohbach et al., 2009; Lerman and Warren, 2011; Davis et al., 2012). While the luxury effect is not present in every city, often as a result of distinct development and social histories (Kendal et al., 2012; Chamberlain et al., 2019), the phenomenon is typically linked with the segregation of greenspaces (e.g., Venter et al., 2020), which is characterized by the park-score metric (Trust for Public Land, 2021). While parks are public features of cities, concerted efforts in investment are required at the city and community levels to develop and maintain parks. Thus, the luxury effect may also explain patterns of urban park biodiversity. However, this remains untested.
The overarching goal of our study was to understand the role of parks in providing a refuge for birds throughout Greater Los Angeles, California (LA) across a park-needs gradient. LA has one of the lowest park scores of the major cities in the United States (Trust for Public Land, 2021). With a clear understanding of the hurdles this poses to its population, the city (and region) has been investing heavily to meet this challenge (City of Los Angeles Department of Recreation and Parks, 2019). Nevertheless, there remains a lack of information on whether parks provide suitable habitat for birds in LA, especially when considering the variation in parks across the cityscape (Trust for Public Land, 2021). Given that avifauna varies strongly in residential areas across an income gradient throughout LA (Wood and Esaian, 2020), we sought to examine whether parks can buffer the negative effects of urbanization in areas of the metropolis that comparably lack green space. Thus, we designed a study set along a park-needs gradient to understand how variations in park features and urban habitat surrounding parks influence their avifauna. To address our overarching goal, we had two objectives.
First, we analyzed variations in habitat features and avian communities in parks across a park-needs gradient. We predicted a refugia effect of parks, following from refugia effects in the conservation literature (Rojas et al., 2022), where parks with higher stressors surrounding their boundaries would have a higher abundance of birds and a distinct avifaunal community than those with low stresses. In our system, we assumed that neighboring stresses of parks were related to the amount and extent of urbanization, e.g., high impervious surface cover. Since high-park needs areas of LA are generally situated in low-income areas that tend to be less vegetated (Avolio et al., 2018), we expected that parks in these areas would be more beneficial for birds than parks in low-needs areas (high income), where birds may utilize the largely vegetated residential areas (Wood and Esaian, 2020). Further, we predicted that parks surrounded by higher impervious surface cover would harbor a greater abundance of synanthropic species than birds that typically reside in natural areas (Aronson et al., 2016), a pattern found in protected areas across the United States (Wood et al., 2014, 2015).
Second, we analyzed relationships among habitat features within and adjacent to parks on avian abundance. Our intention with the relationship analysis concerning refugia effects was generally to understand whether habitat features within or adjacent to parks are influential in describing avifaunal patterns and whether these patterns vary across the park-needs gradient. Given how parks may functionally act as ‘islands’ in the cityscape, we predicted that larger parks near natural areas would have a greater abundance of birds following from other urban systems and also from the predictions of island biogeography (Donnelly and Marzluff, 2004; Molles and Sher, 2018; La Sorte et al., 2020). Additionally, we predicted that synanthropic birds affiliated with urban habitat features would be positively related to impervious surfaces, both within and adjacent to parks, and other features resembling dense urban form, e.g., less tree cover surrounding parks (Johnston, 2001). Further, we predicted that birds affiliated with shrubs, trees, and other natural amenities would be positively related to similar features within and adjacent to parks (Wood and Esaian, 2020).
Materials and methods
Study area and sampling design
We studied habitat and bird communities in 48 urban parks throughout LA The LA metropolitan area has a population of over ten million people and spans an area of approximately 10,510 (U.S. Census Bureau, 2019). The region is characterized by a Mediterranean climate and experiences hot and dry summers contrasted with cool and wet winters. LA, which primarily covers the San Fernando and San Gabriel Valleys, the LA Plain, and the foothills of various hills and mountains ranges of the region, was formerly a diverse mosaic of wetlands, riparian forests, oak (Quercus spp.) and walnut (Juglans spp.) woodlands, coastal sage scrub, chaparral, and grassland ecosystems before intense development throughout the 20th century (Stein et al., 2007). The region is now mainly urbanized, dominated by a composite of large, medium, and small municipalities with extensive suburbs, numerous urban cores, and few natural green spaces distributed throughout. The greater metropolitan area is primarily bounded by the Pacific Ocean to the South and West and the Transverse and Peninsular Mountain Ranges to the North and East.
We initially selected a random sample of 60 managed urban parks set throughout LAWe identified these 60 parks using a polygon shapefile of the parks and open spaces in Los Angeles County, which we acquired from the Los Angeles County GIS repository (Los Angeles County, 2016a). To categorize the 60 urban parks based on park needs, we utilized the ‘Park Needs Assessment Detailed (Hosted–Public)’ geodatabase (Los Angeles County, 2016b). The geodatabase provides a spatial layer intended to highlight the 2016 needs assessment, that quantified the needs for parks and recreation resources and estimated the potential cost of meeting the need across the County (Los Angeles County, 2016a). The needs assessment grouped locations of L.A. into six categories: very high, high, moderate, low, very low, and not participating. The majority of Los Angeles County falls under the categories of very high (32.2%), high (20.4%), and moderate needs (26.2%). Low (16.5%), and very low needs (4.6%) make up a considerably smaller percentage (Los Angeles County, 2016b). We used a spatial join to merge the 60 managed urban parks with the needs assessment spatial layer. We only retained parks with at least 20% of tree cover in the final sample because we assumed that parks composed primarily of grass, bare ground, or impervious surfaces, which typically were those dominated by ball fields or courts, would have fewer birds due to lack of habitat. Thus, all parks in the study were generally typical of urban parks in LA, with grassy fields, play areas, and trees (Figure 1). The final sample consisted of 48 parks: 10 in very high and 10 in high, which we merged into a ‘high needs category (20), 11 in moderate, and 17 in low (Figure 2; Supplementary Figure S1).

Figure 1. An example of an urban park included in this study (Villa Parke, City of Pasadena, Los Angeles County, California, United States). Photo credit, E. Wood.
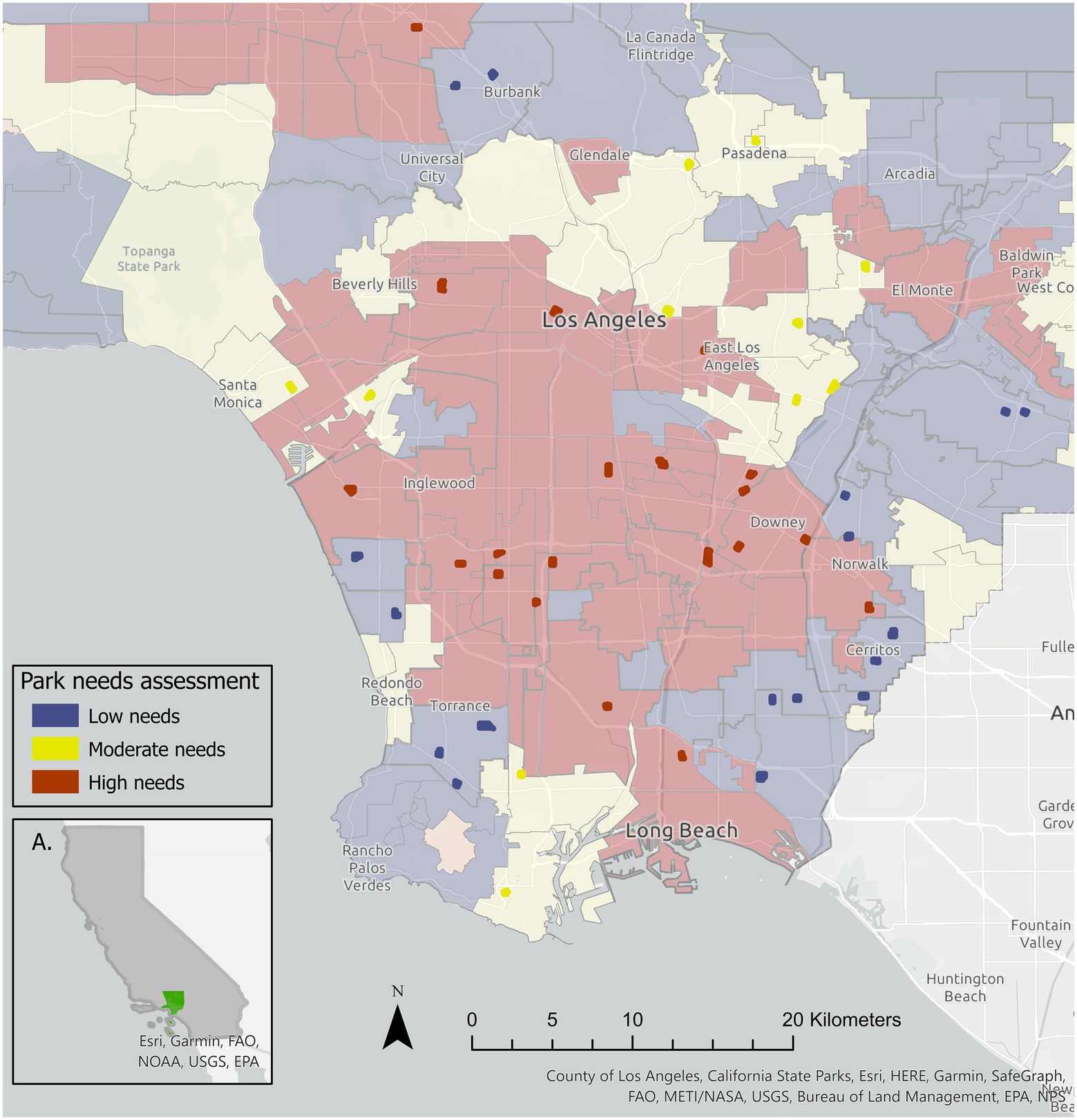
Figure 2. Locations of the 48 urban parks included in this study. The red, yellow, and blue color gradient indicates designations of Greater LA categorized by park need based on the Los Angeles Countywide Comprehensive Parks and Recreation Needs Assessment report (LA County, Department of Los Angeles County, 2016b). We grouped very high, and high needs parks into a ‘high needs’ group (red, n = 20), parks in We then included parks categorized within sections of the city as ‘moderate’ (yellow, n = 11) and ‘low’ (blue, n = 17) needs.
Area search bird surveys
We surveyed parks using an area search method to quantify bird abundance (Loyn, 1986). We favored area search surveys over point counts because nearly all parks were small enough to be sampled in their entirety. We surveyed each park three times over two field seasons with one visit during the winter of 2018/2019 and two additional visits during the winter period of 2019/2020. We surveyed during the winter months from late October to late March as it is a time of year when wintering migratory birds are abundant in southern California (Garrett et al., 2012; Higgins et al., 2019). Surveys typically involved an observer walking on a set route throughout parks, identifying and counting each bird that was seen or heard within park boundaries. We used the Gaia GPS© mobile app to record the initial survey routes that we followed during each subsequent park visit. Care was taken to not double-count birds, especially flocking birds that would frequently move throughout a park during a survey. If an observer encountered a flock of birds, we recorded the number of species and abundance. If a similar composition of birds within a flock was encountered within 100-m of our previous observation, we omitted those from the survey to avoid double counting birds. Birds flying over parks or outside park boundaries were not recorded, as we were only interested in analyzing bird communities within park boundaries during the duration of a survey. We counted raptors and waterbirds in our survey, but they were not included in our analysis (see bird guilds below). Surveys began within an hour after sunrise and were concluded by 1,100 h. to capture the prime activity of birds. The length of each survey varied between parks to account for the variability in park size and tree cover. Larger parks with greater tree cover generally took longer to survey.
N-mixture abundance calculations and bird guilds
To account for detection probability, which is a concern with wildlife count data (MacKenzie et al., 2017), we calculated N-mixture models, which are hierarchical models that incorporate spatial replicates of raw abundance count data (Royle and Nichols, 2003). The model estimates a detection probability for a given species, which is then utilized to adjust abundance estimates from a predicted model using an appropriate error distribution (e.g., Poisson, Royle and Nichols, 2003). We fitted the intercept-only N-mixture model, using the ‘pcount’ function in the R package ‘unmarked,’ for 33 bird species (Table 1). We then estimated the posterior distribution of latent abundance from the N-mixture models for the 33 candidate bird species at each park using empirical Bayes methods from the unmarked package (function, ‘ranef’; Fiske and Chandler, 2011). Therefore, when we present the results of bird abundance, we refer to the estimated abundances from the N-mixture models. We assumed observer bias in our survey was minimal, as one observer (AV) collected nearly all data (92% of observations), with EW and a handful of students occasionally completing surveys. Further, because we sampled over two seasons, we varied detection probability by season to account for potential year-to-year effects. Lastly, a critical assumption for estimating detection probability within a season is ‘closure’ (MacKenzie et al., 2017). While birds move frequently during the non-breeding period, we assumed that the focal species of this study were present and available during the winter months for detection throughout our surveys.
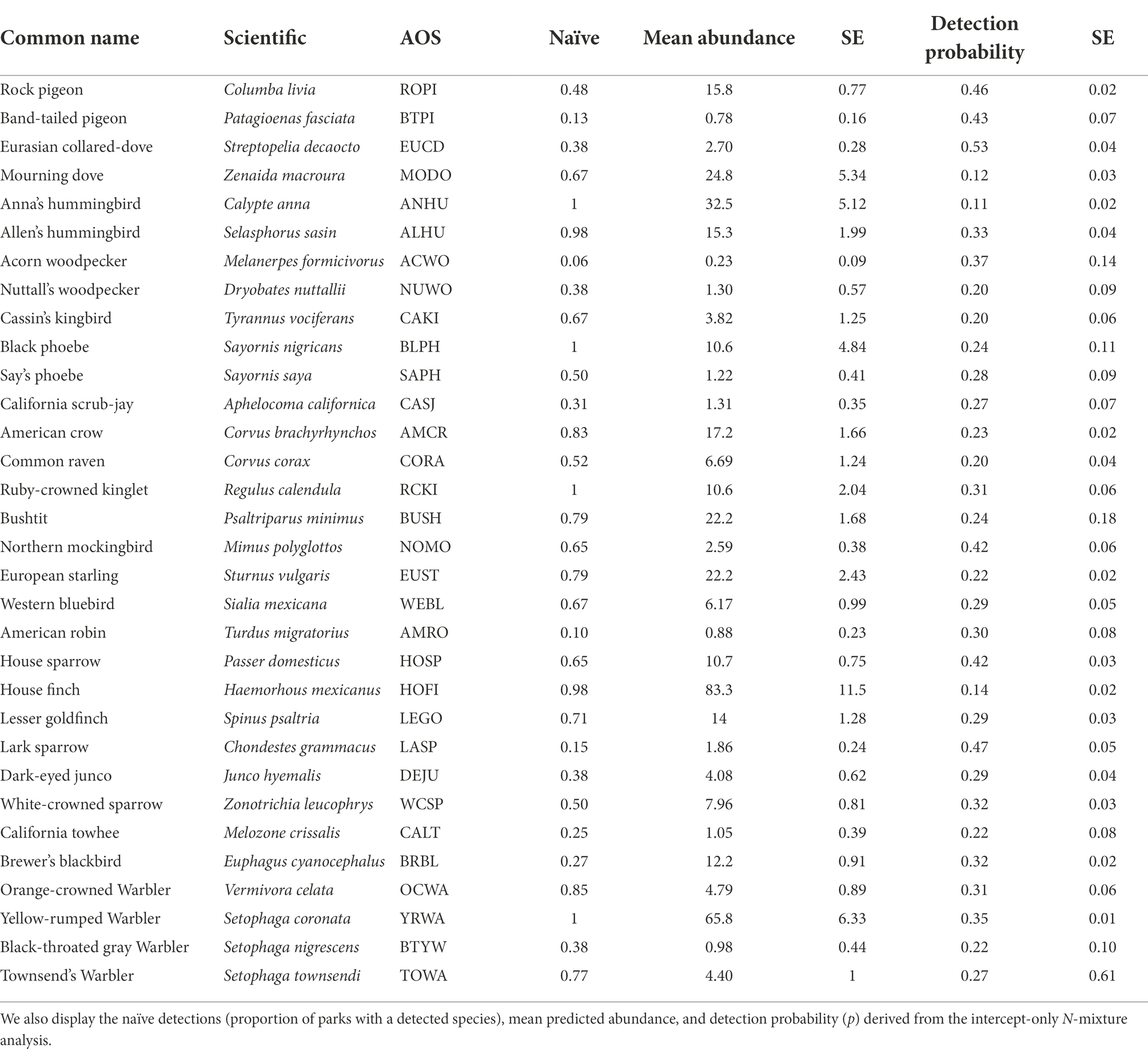
Table 1. Common, scientific names, and the American Ornithological Society (AOS) alpha 4-letter bird codes (Chesser et al., 2021) for 33 species included in the N-mixture modeling analysis.
To focus our analysis on bird species that may have variable responses to park and urban habitat features, we created four bird habitat guilds. These included birds affiliated with forest and open woodland, shrublands (shrub), grassland, or urban ecosystems (urban) during the breeding season, assuming their habitat associations would be similar during the winter months (Supplementary Table S1; Clark, 2017; Billerman et al., 2021). The ‘urban’ birds are species often categorized as synanthropes (Supplementary Table S1). We also created a migratory and resident bird guild, which included species that either depart the L.A. area during the summer for breeding duties or stay within the region (Supplementary Table S1). For each guild, we summed the estimated abundance of each bird within a guild to quantify a guild-specific estimated abundance value, which we used as dependent variables. We also summed the total migratory and resident bird groups for a total abundance group within each park (Supplementary Table S1).
Habitat variables, remote sensing and spatial analysis
We used data from remote sensing platforms coupled with spatial processing to characterize habitat features within and adjacent to parks. We used a pixel-based image classification to derive habitat features within parks and within a 0.8 km (0.5 miles) buffer around each park. The purpose for characterizing habitat features in the 0.8 km buffers surrounding parks was to capture adjacent landscape characteristics and their effect on park avian communities, standard practice when performing landscape-extent analyses (Jimenez et al., 2022). We used a 2016 National Agriculture Inventory Project (NAIP) 4-band image data acquired from the Los Angeles County GIS Database for the classification (Los Angeles County, 2009). The image was taken 2 years before our sampling. However, we assumed that any potential differences in built structures surrounding parks, or infrastructure within parks, e.g., tree removal, would be negligible over the 2 years. Before performing the classification, we extracted the near-infrared, red, and blue bands from the NAIP image. We chose these bands because of their ability to distinguish between vegetation, manufactured objects, and other urban features (Wood et al., 2013).
To further differentiate between vegetation and other urban land-cover features, we created a Normalized Difference Vegetation Index (NDVI) layer (Pettorelli et al., 2011). NDVI measures vegetation ‘greenness’, ranging from a scale of − 1 (least green) to 1 (most green). NDVI is derived with the following equation:
The green vegetation in our study area was typically characterized by high, positive NDVI values; dead vegetation, e.g., grass in the winter months, had low, positive values (~ 0.05). Bare ground had values closest to zero, and impervious surfaces all had negative values.
To increase the classification accuracy, we acquired raw LiDAR point cloud data for the study area from a National Oceanic and Atmospheric Administration (NOAA) data repository (OCM Partners, 2022). We used the first-return LiDAR data points to create a Digital Surface Model (DSM) and the ground return data points to create a Digital Terrain Model (DTM). Using these two layers, we created a Normalized Digital Surface Model (nDSM) that depicts features elevated from the ground, such as trees and buildings. nDSM is derived by subtracting the DSM with the DTM layer. We created the nDSM layer to differentiate trees from grass by their height differences. Finally, we combined the nDSM, NDVI, and the 3-band NAIP image to create a new 5-band image layer used as the raster input for the final classification. We used a Support Vector Machine (SVM) pixel-based classification to classify tree cover, grass cover, bare ground, and impervious surfaces within and around each park. Water and shadows were also classified since these features were common throughout the landscape. However, we did not include water or shadows in our final analysis assuming they had a small effect on the landbirds of our study.
To assess the accuracy of the remote sensing classification, we used 200 assessment points and computed a confusion matrix that revealed a classification accuracy of approximately 86% (Supplementary Table S2). We then used the Tabulate Area tool in ArcMap to calculate the proportion of each feature type within and around each park. Additionally, we used the Near tool in ArcGIS Pro to calculate the Euclidian distance between each park and the nearest natural area. We designated areas as ‘natural’ if classified as ‘protected areas’ or ‘open spaces’ within LA County park’s polygon shapefile (Los Angeles County, 2009). Last, we determined the median income of census tracts where parks were situated using spatial data organized by Southern California Association of Governments (2016). We used the median income data as our indication of potential luxury-effect patterns based on our sampling design (Leong et al., 2018; Schell et al., 2020). These income data were from 2016 and based on projections from the United States 2010 census (U.S. Census Bureau/American FactFinder, 2010). We used these 2016 data as we assumed they approximated income levels in the sections of the city that were comparable to the time we collected data in 2017/2018 and 2018/2019. All remote sensing and spatial analyses were completed using ArcMap and ArcGIS Pro (ESRI, 2020).
Statistical analysis
Objective 1: Patterns of habitat features and avian communities
We completed three analyses to characterize patterns of habitat features and bird abundance in parks across the income gradient. First, to quantify differences in bird guild abundances, park features, and landscape characteristics across socioeconomic statuses, we performed a series of one-way analysis of variance tests (ANOVA). The categorical fixed factor for each model was the park-needs category (low, moderate, high). If tests were significant, we employed a Tukey’s HSD test. As we were making three comparisons among income levels for a particular variable, we used a Bonferroni correction of the alpha value, α = 0.05/3 = 0.02 to assess significance.
Second, to identify the degree of dissimilarity in the bird community concerning the park-needs categories, we conducted a one-way analysis of similarities test (ANOSIM; Oksanen et al., 2019), using the Bray–Curtis dissimilarity of the square-root transform of the 33 species from the N-mixture analysis grouped among high, moderate, and low park needs groups. If an ANOSIM test was significant at the alpha value of 0.05, we calculated pairwise comparisons by performing an ANOSIM analysis of either low-high, low-moderate, or moderate-high. Like the ANOVA analysis, as we were making three comparisons among income levels for a particular variable, we used a Bonferroni correction of the alpha value, α = 0.05/3 = 0.02 to assess significance.
Third, to further assess differences in the avian community across the park-needs gradient, we computed a non-metric multidimensional scaling (NMDS) analysis. We again used the Bray–Curtis dissimilarity of the square-root transform of the 33 avian species from the N-mixture analysis. We created an ordination graph of the 2-D representation of the avian community using the vegan package in R, and we overlaid habitat vectors on the ordination using the ‘envfit’ function in vegan (Oksanen et al., 2019). The envfit function assesses the correlation of both habitat and avian species vectors with the first two axes of the ordination (Oksanen et al., 2019) and thus provides a measure of continuous change of the avian community concerning habitat variables across the park needs gradient.
Objective 2: Relationships among habitat features and bird abundance
To understand the relative effects of local and landscape habitat features on avian park communities, we used a model selection approach, where we fitted a series of generalized linear models (GLMs), regressing the independent habitat variables against the seven bird guilds, which were the dependent variables in the analysis. Because our data were based on counts (abundance), we used Negative Binomial GLMs with a log-link function. We used Negative Binomial models to account for overdispersion in the Poisson distributed count data, which was evident based on calculating the ratio of the residual deviance to the residual degrees of freedom for each model (Zuur et al., 2011). We developed seven distinct model sets, with one for total bird abundance, and then six others for the bird guilds (forest and woodland, shrubland, grassland, urban, migratory, and resident) regressed against 11 independent variables and the intercept-only model. We fitted all models as univariate combinations of an independent and dependent variable. We did not explore multi-variable models or interactions primarily because we were interested in the general correlation of a given independent variable with a dependent variable. Further, numerous independent variables were moderately to highly correlated, thus making fitting multiple variable models challenging (Supplementary Figure S2). For organization purposes, we grouped our independent variables based on whether they were related to the luxury effect (median income), island biogeography (park size and distance to the nearest natural area); park composition (the % cover of impervious surface, trees, grass, and bare ground); and urban habitat features surrounding parks (the % cover of impervious surface, trees, grass, and bare ground). Each independent variable in our analysis was either biologically relevant to the avifauna of our study (e.g., % tree cover), or commonly used in urban ecology studies as a means for understanding potential conservation and habitat associations (e.g., % bare ground). Therefore, each model had biological or management significance.
We used an Akaike’s Information Criterion (AIC) model selection framework to determine which variable was the most important predictor of bird abundance in parks within each of the three analysis extents. We determined ‘top models’ as those with a ΔAIC < 2 (Anderson and Burnham, 2002). We also computed R2 values based on the Kullback–Leibler-divergence ) generated from calculating the likelihood ratio index of a fitted model (Cameron and Windmeijer, 1997). We completed all analyses using the R programming language (R Core Team, 2017), with code and figures run and created using ‘rmarkdown’ (Allaire et al., 2022). We used Adobe Illustrator to finalize the figures (Adobe Inc., 2019).
Results
The average detection probability for the 33 species included in the N-mixture analysis was 0.30 (Table 1), with the Eurasian Collared-Dove (Streptopelia decaocto) having the highest detection probability (p = 0.52) and the Anna’s Hummingbird (Calypte anna) having the lowest (p = 0.11). Four bird species were detected at every park (Naïve detection), including the Anna’s Hummingbird, the Black Phoebe (Sayornis nigricans), the Ruby-crowned Kinglet (Regulus calendula), and the Yellow-rumped Warbler (Setophaga coronata; Table 1). The average mean-estimated abundance for all species was 12.49 individuals per park (Table 1). The most abundant birds in our study were the House Finch (Haemorhous mexicanus; mean estimated abundance across parks = 83.3) and the Yellow-rumped Warbler (65.8), and the rarest species were the Acorn Woodpecker (Melanerpes formicivorus; mean estimated abundance across parks = 0.23), the Band-tailed Pigeon (Patagioenas fasciata; 0.78), the American Robin (Turdus migratorius; 0.88), and the Black-throated Gray Warbler (Setophaga nigrescens; 0.98), all with mean abundances of less than one per park (Table 1).
Objective 1: Patterns of habitat features and avian communities
Overall, there were few differences in habitat characteristics in parks and the surrounding urban environment across the low-, moderate-, and high-needs gradient of our study (Table 2). Notable variables that varied included the median income of the residential areas surrounding parks (value of p < 0.01), which was 30% higher in low than high needs areas; the distance to natural areas, where parks in high-needs areas were over twice as far from natural areas as parks in high-income areas (value of p < 0.01), which indicated their general position in the center of the metropolis; and the proportions of tree cover in the urban environment surrounding parks, which was 25% greater in low than high needs areas of the city (value of p = 0.02; Figure 3; Table 2). Interestingly, the cover of trees within parks did not vary across the park-needs gradient (Figure 3; Table 2), suggesting that the luxury-effect phenomenon of tree cover within residential zones does not apply to urban parks e.g., (see adjacent tree cover results). There were no differences in the avian guilds across the park-needs gradient also suggesting that the luxury effect does not apply to explaining park avifauna in L.A (Table 2).
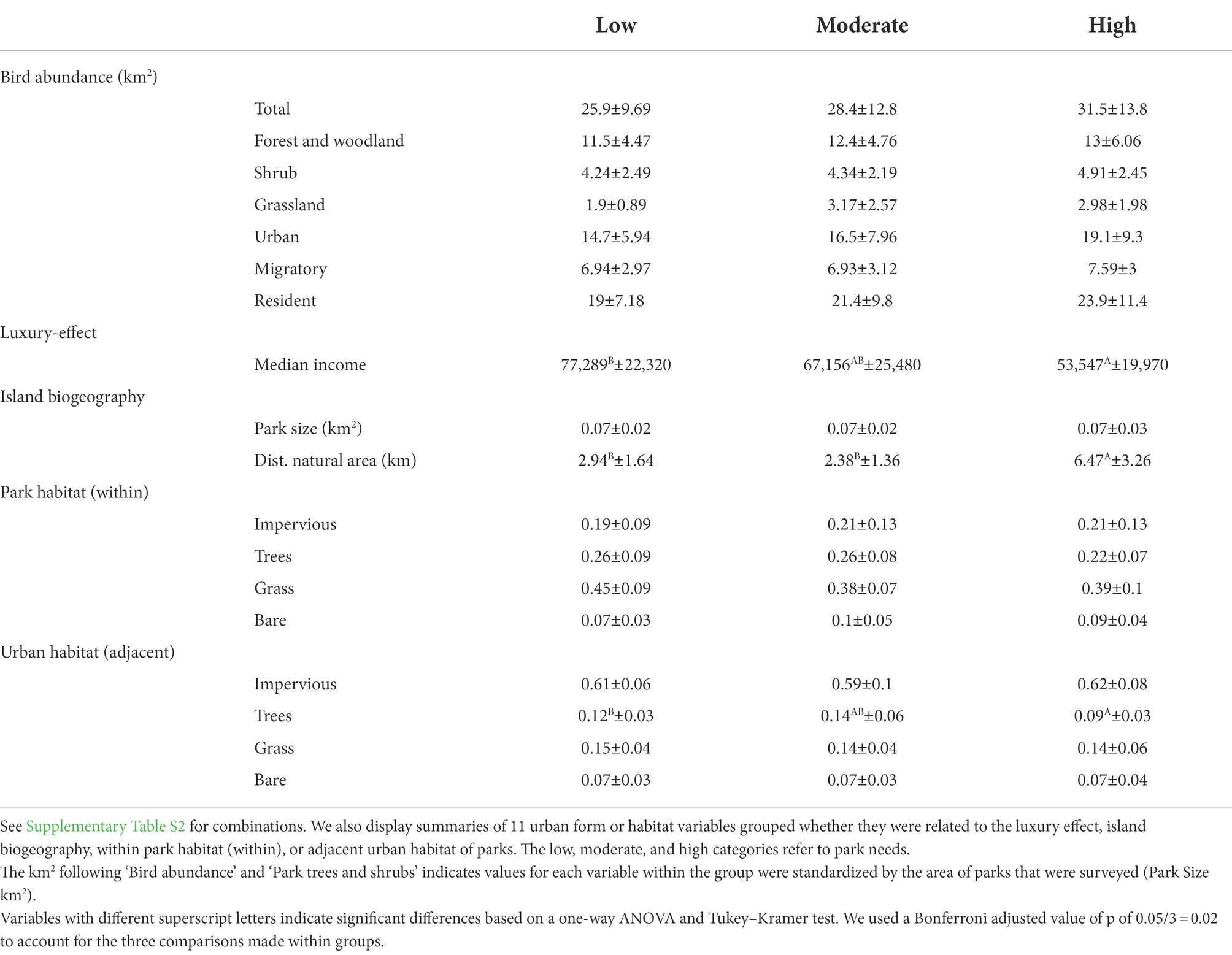
Table 2. Mean ± S.E. summaries of the total abundance of birds, and six additional groups indicating combinations of bird species associated with forest, shrub, grassland, or urban ecosystems during the breeding period, or whether species are wintering migratory birds (migratory) or resident to the L.A. study area.
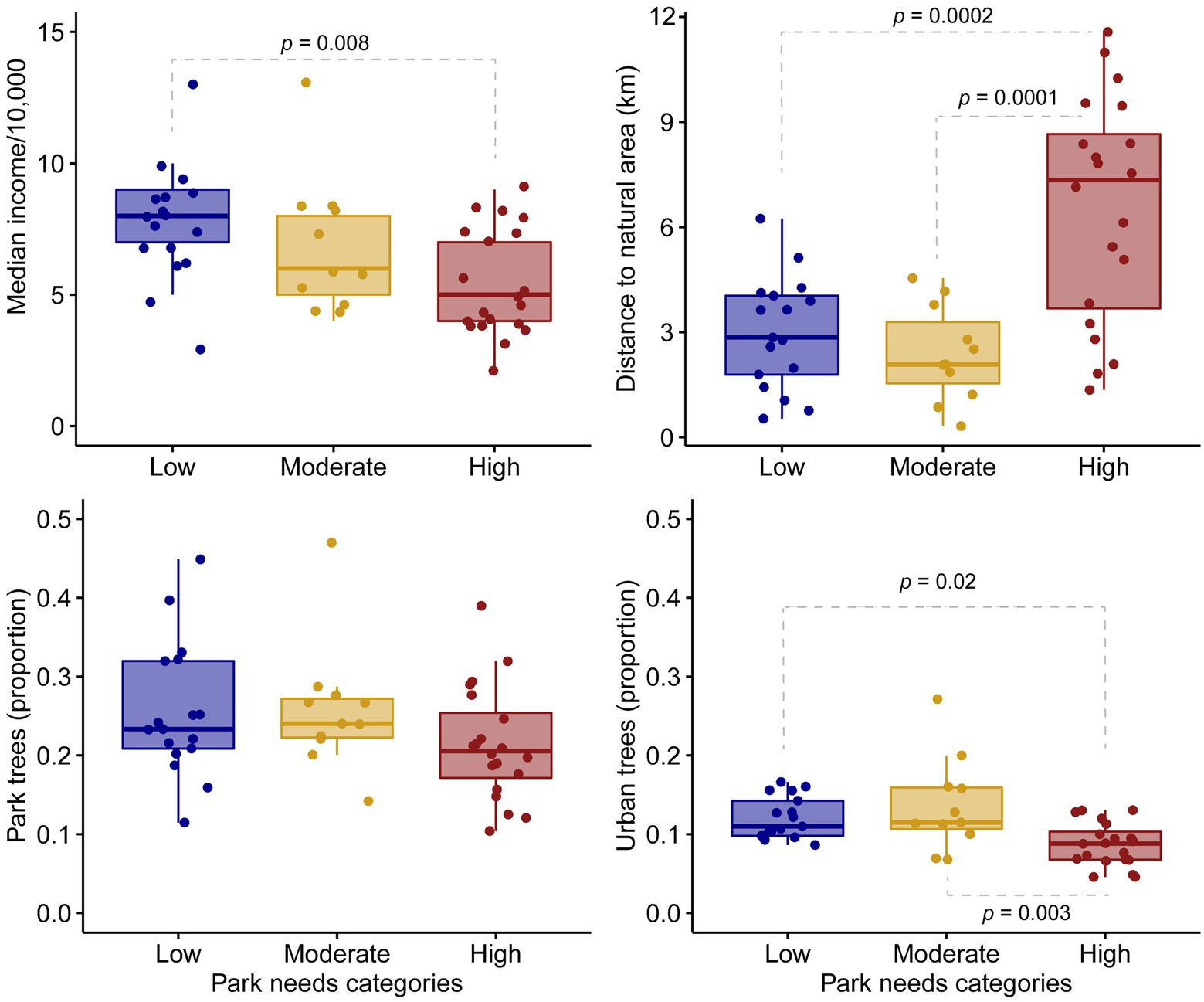
Figure 3. Box plots depicting patterns of median income, distance to the closest large natural area (km), the proportion of tree cover within parks [Park trees (proportion)], and the proportion of tree cover adjacent to parks [Urban trees (proportion)] across a gradient of low-, moderate-, and high-park needs. Dotted gray lines linking boxes indicate significant differences based on a one-way ANOVA and Tukey–Kramer test, or a Kruskal–Wallis rank-sum test followed by a nonparametric multiple-comparisons procedure, based on relative contrast effects. We used a Bonferroni adjusted value of p of 0.05/3 = 0.02 to account for the three comparisons made within groups.
Similar patterns were also evident when analyzing the dissimilarities of avian communities among the park-needs categories. There was slight evidence of dissimilarities in communities for total bird abundance (ANOSIM R = 0.07, value of p = 0.07) and forest and woodland bird abundance (R = 0.08 value of p = 0.05). However, there were no significant differences in dissimilarities when analyzing pairwise comparisons. ANOSIM values range from −1 to 1, with values closer to zero indicating no dissimilarities across groups. Thus, the effects were weak for total and forest and woodland abundance. The only group that did show evidence of dissimilarity across the park-needs groupings was resident birds (R = 0.07, value of p = 0.04), which displayed a trend in dissimilarity between parks in high- and low-needs areas (R = 0.06, value of p = 0.06), and between parks in moderate- and low-needs areas (ANOSIM R = 0.11, value of p = 0.06). However, we note the pairwise comparisons were not significant at the Bonferroni adjusted alpha value of 0.02, again suggesting weak dissimilarities. All other avian guilds were similar across the park-needs categories, with value of ps ranging from 0.13 (urban), 0.19 (grassland), 0.39 (shrub), and 0.42 (migratory).
The NMDS analysis had reached a stress solution of 0.22 suggesting modest confidence in the outputs. Nevertheless, the analysis revealed a few important distinctions in habitat characteristics and avian communities across the continuous park-needs gradient. There were five important habitat vectors identified, including urban trees surrounding parks, bare ground within parks, distance to the nearest natural area, park size, and median income (Figure 4). Median income was positively correlated with axis 1, whereas park bare ground cover was negatively correlated with axis 1 (Figure 4). The distance to the nearest natural area and park size were positively correlated with axis 2, while urban trees surrounding parks was negatively correlated with axis 2 (Figure 4). Avian communities from parks in low-needs areas were weakly positively associated with income and negatively with bare ground cover within parks (Figure 4). Parks in high-needs areas were weakly aligned with distance to a natural area and park size, again indicating the location of the high-needs areas in the center of the city, further from the surrounding natural areas where few large parks occurred (Figure 4). Birds in parks in low-needs areas of LA were aligned with income (positively) and bare ground cover (negatively) within parks (Figure 4).
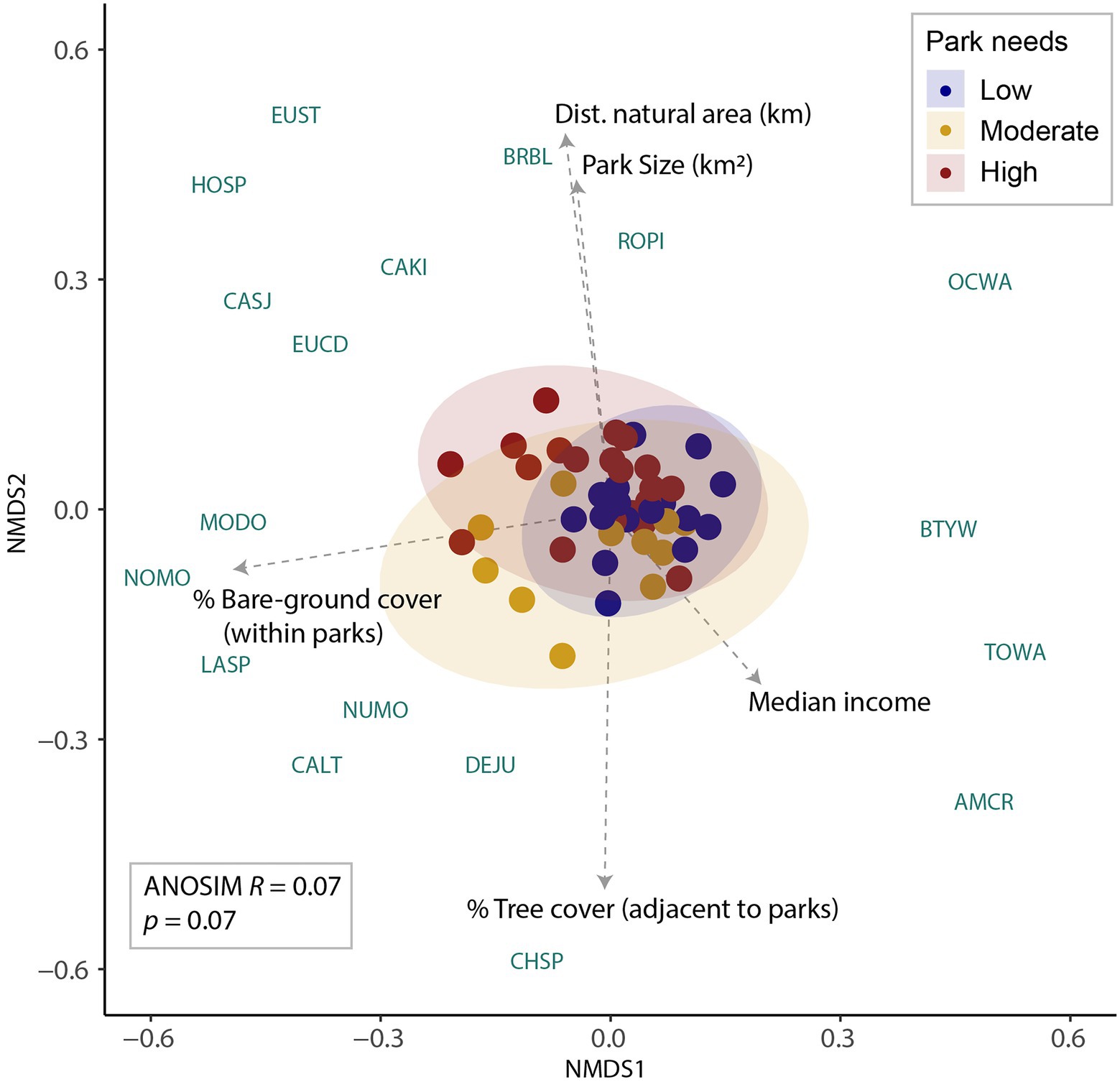
Figure 4. 2-D non-metric, multidimensional scaling ordination (NMDS) plot of avian species composition (N-mixture estimated abundance per species) within parks situated across a ‘park needs’ assessment gradient in Greater Los Angeles (LA). The ordination successfully converged with a stress value of 0.22. The ellipses are the bivariate confidence interval assuming a Student’s-t distribution and characterize the potential of the avian community within a park needs assessment categories. The dotted lines represent vectors of environmental variables that were significantly associated with the ordination scores. Axis one is thus weakly positively associated with median income and negatively with bare ground cover within parks. Axis two is correlated with a positive distance to the nearest natural area (km), and park size (km2) and negatively with the surrounding cover of trees in the urban landscapes adjacent to parks. The four-letter codes are the plotted Bray–Curtis dissimilarity distances of 18 avian species (see Table 1) that were significantly correlated with axes one or two.
Objective 2: Relationships among habitat features and bird abundance
The most influential variables explaining bird abundance varied for each of the bird guilds but were generally aligned with island biogeography variables, and then components of the parks and not the surrounding landscape (Table 3; Appendix S1). Forest and woodland birds and migratory birds were best explained by distance to the nearest natural area (negative and positive association, respectively; Table 3; Figures 5A,E), whereas shrub and grassland bird abundance was best described by park size (Table 3; Figures 5B,C). The park-size finding suggests a species-area effect, which typically explains richness patterns. However, in our case, park size characterized avifaunal abundance. The cover of bare ground was positively related to total, urban, and resident bird abundance and highly competitive with grassland bird abundance (ΔAICc = 0.10; Table 3; Figures 5D,F). There were fewer relationships between the urban environment habitat variables and birds within parks (Table 3). These results indicate that birds will likely use parks as habitat throughout the city depending on the specific management of the parks themselves, and not necessarily due to drivers from the adjacent habitat.
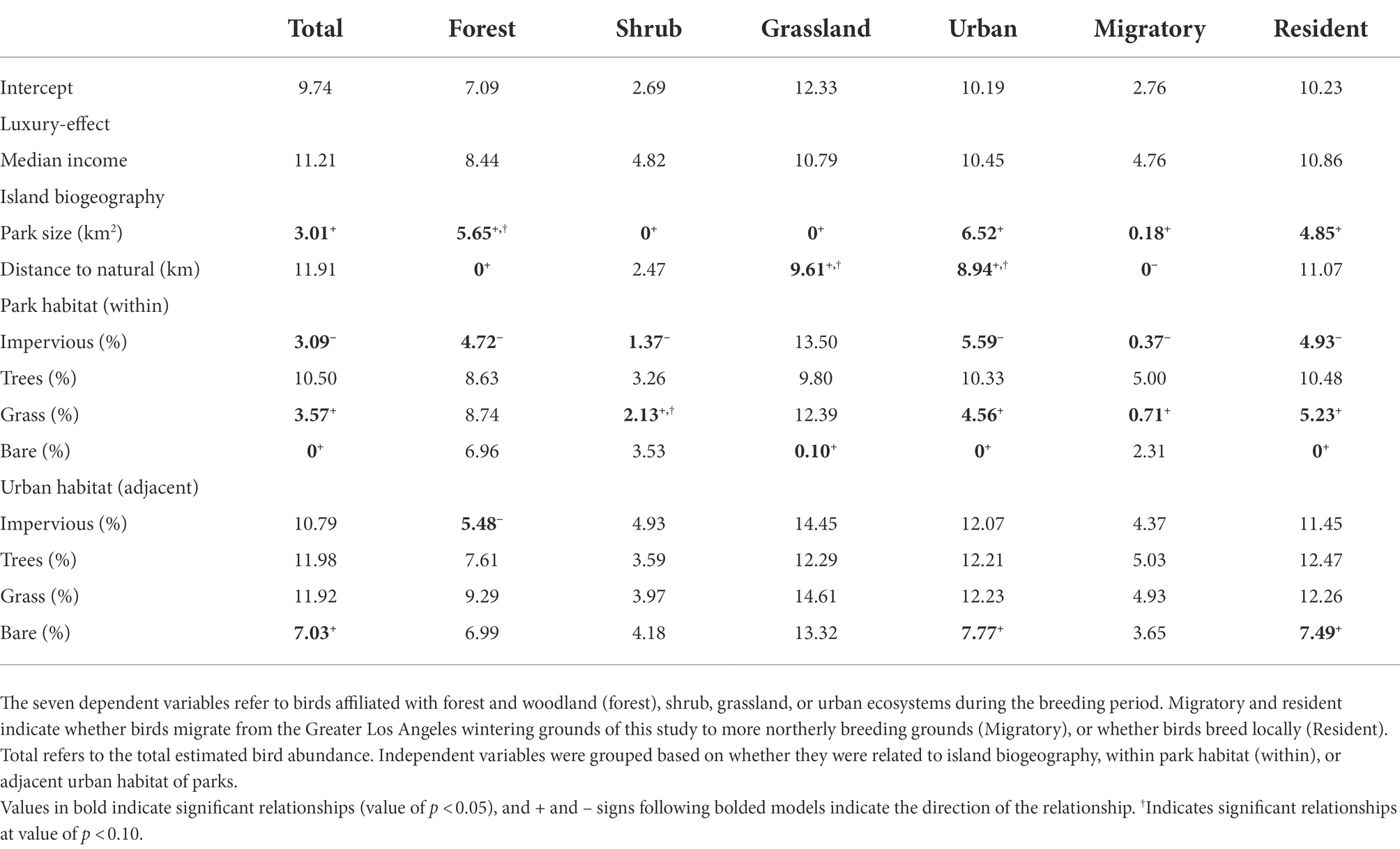
Table 3. ΔAICc values based on a model selection routine for bird abundance regressed against 11 independent variables and the intercept-only model.
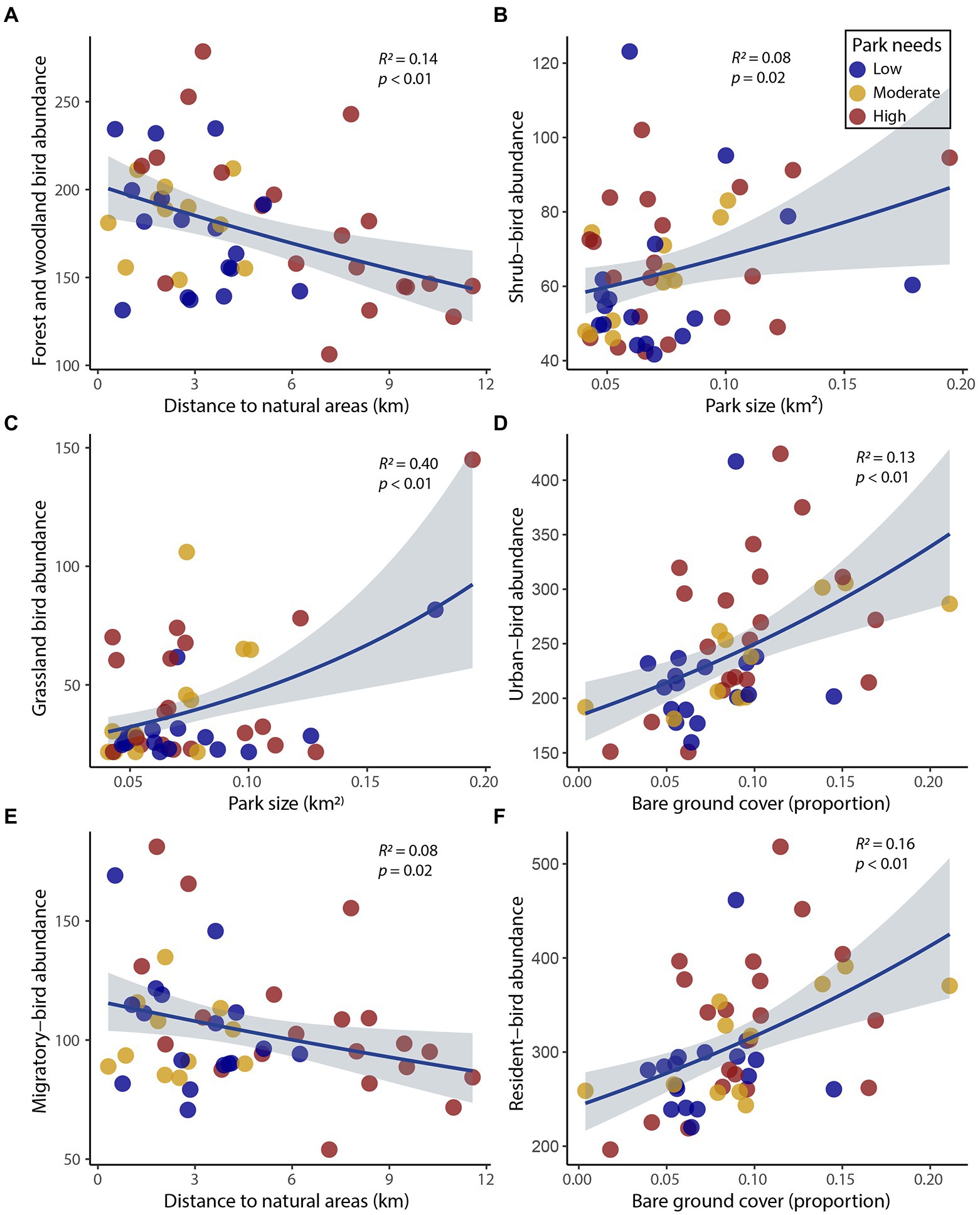
Figure 5. Scatterplots depicting the relationships of the top models for (A) forest and woodland, (B) shrub, (C) grassland, (D) urban, (E) migratory, and (F) resident bird abundance with independent variables. We generated the fitted line and confidence interval (gray shading) based on a negative binomial generalized linear model analysis. The R2 values are Kullback–Leibler-divergence-based values generated from calculating the likelihood ratio index of a fitted model (Cameron and Windmeijer, 1997).
Discussion
Our results indicated that parks are a refuge for avifauna in park-poor sections of cities. We uncovered three lines of evidence to support our main conclusion. First, we initially predicted a refugia effect, where parks with higher stressors surrounding their boundaries would have higher individuals than those with low stresses. Our analysis found support for this prediction. Bird abundance patterns for species affiliated with forest, shrub, and woodland ecosystems were generally similar across the park-needs gradient of LA, indicating that in areas of the metropolis with high stresses (low income and high-park needs) birds utilize parks in relatively high frequencies. Interestingly, this pattern is generally opposite to what is found outside of parks, where forest-affiliated birds are far denser in high-income residential areas of LA (low park needs) than in low-income areas (high park needs; Wood and Esaian, 2020). This result suggests that birds typical of natural ecosystems surrounding L.A. use parks in otherwise inhospitable areas of the city at comparable levels to locations that have abundant greenery outside of park boundaries. In a similar line of evidence, avian communities varied slightly among parks in high and low-needs areas of the city, with few habitat variables weakly associated with avifaunal community structure, including median income, bare ground cover within parks, urban tree cover surrounding parks, park size, and distance to natural areas. Though the patterns were weak, these results, especially for median income and urban tree cover surrounding parks, provided some support that the surrounding cityscape may indeed filter the species pool found within parks (e.g., Aronson et al., 2016). However, we again stress that the patterns we uncovered in parks are far weaker than the filtering effects found outside of parks in residential areas (Wood and Esaian, 2020), again providing support for their refugia potential in dense urban conditions. Lastly, bird abundance patterns were related to numerous island biogeographic and habitat variables within parks, which were stronger than habitat variables surrounding parks. These findings suggested the important role parks have in providing habitat for birds, regardless of whether they are in high- or low-park needs areas of the city. Overall, in addition to the benefits to people, our work suggests park development in park-poor areas of L.A. would also have positive effects on birds.
The luxury effect, parks, birds, and their habitat
Among the many known drivers of biodiversity in cities, the luxury effect posits that vegetation cover and wildlife biodiversity follows patterns of wealth (Leong et al., 2018). The Park Score Index clearly describes the luxury effect highlighting the disparity in the distribution of parks in high- and low-income areas of LA Because the luxury effect is a prevalent and defining feature of biodiversity in residential areas and other greenspaces of LA (Clarke et al., 2013; Wood and Esaian, 2020), we were interested in testing the luxury effect based on avifaunal patterns found within parks across the park-needs gradient. We found evidence for the luxury effect of the tree cover surrounding parks, which has been repeatedly documented in LA and many other cities (e.g., Schwarz et al., 2015). But we did not find support for the luxury-effect hypothesis for other habitat conditions, especially within parks. While our findings did not match our expectations, our results were in line with a handful of other studies from around the world. For example, in Sydney, Australia, topography was the strongest predictor of plant abundance within parks rather than income (Zivanovic and Luck, 2016). Further, in Phoenix, Arizona, the income of the surrounding residential areas was not a strong predictor of park vegetation. Instead, the median year of development and whether residents had a graduate degree best-explained park vegetation abundance and richness (Martin et al., 2004). The patterns from Sydney and Phoenix along with our own suggest that urban parks are likely built and managed similarly across a cityscape regardless of the surrounding socioeconomic patterns.
The similarity in habitat conditions within parks across the park-needs gradient carried over to influence birds, which, also indicated a lack of support for the luxury-effect hypothesis in parks in LA In other areas of the world, habitat features that were similar between high- and low-income areas of cities supported similar biodiversity patterns. For example, there was no evidence of the luxury effect when considering bird diversity patterns in greenspaces throughout Johannesburg, South Africa (Howes and Reynolds, 2021). Rather, the use of water bodies, which were historically implemented to segregate white and black populations of the city now buffer bird diversity patterns in low-income areas of Johannesburg (Howes and Reynolds, 2021). As Leong et al. (2018) suggested, correlations between bird populations and socioeconomics may be directly attributed to differences in vegetation cover across income. Given the similarity of habitats in LA parks, it appears parks can buffer avian communities across income gradients, which again suggests a refugia benefit of parks in LA for urban avifauna.
Parks as habitats for birds
Many studies from around the world have indicated the importance of urban greenspaces, including parks, to birds, which our study strongly supports (e.g., Blair, 1996; Jokimäki and Suhonen 1998; Fernández-Juricic and Jokimäki, 2001; Cornelis and Hermy, 2004; Colding and Folke, 2009; Carbó-Ramírez and Zuria, 2011; Ikin et al., 2013; Zivanovic and Luck, 2016; Amaya-Espinel et al., 2019; Villaseñor and Escobar, 2019; Zhang and Huang, 2020). Nevertheless, our study uncovered some potentially interesting patterns that merit discussion regarding the potential of parks as habitats for birds. For example, bird species such as the Townsend’s (Setophaga townsendi), Orange-crowned (Vermivora celata), and Black-throated Gray Warblers (Setophaga nigrescens) were aligned with parks in low-needs areas of the city. Parks in these areas were embedded within affluent zones of LA with high tree cover surrounding the boundaries of parks (Wood and Esaian, 2020). While park features in low- and high-needs areas of the city were similar, the surrounding tree cover was dissimilar. The three wood-warblers (Parulidae) are forest and woodland breeding species and are common during the nonbreeding period in parts of LA with high tree cover (Wood and Esaian, 2020). Thus, these results suggest there are potentially important neighborhood-level filters in high-income areas attracting birds to affluent sections of the city that carry over to use the parks (Aronson et al., 2016). This pattern is similar to what is found in residential areas in LA (Wood and Esaian, 2020), but, as we previously discussed, the effects were far weaker within parks.
We also uncovered similar filtering effects when examining distribution patterns of birds that require open areas within parks (e.g., bare ground), where species such as the Lark Sparrow (Chondestes grammacus), Cassin’s Kingbird (Tyrannus vociferans), and California Towhee (Melozone crissalis) were generally more abundant. The Lark Sparrow and Cassin’s Kingbird are species of grassland and savanna-type conditions (Billerman et al., 2021), conditions that parks superficially and structurally resemble (Figure 1). The bare ground could be a surrogate for these open conditions where birds may capture insects by flying out from the perches of trees. Or it may be possible that these bird species are attracted to other resources associated with the bare ground, e.g., shrubs planted next to ballfields, seeds, or dust for bathing. Large swaths of the valleys of LA were formerly grassland and shrubland (Ethington et al., 2020), so there could be a historic signal for birds requiring these ecosystem types to use parks in an otherwise heavily urbanized landscape.
We also desired to understand the relationships of habitat adjacent to parks in influencing avifaunal patterns within parks. Surprisingly, we found few important relationships when examining the effect of the surrounding cityscape on avian abundance patterns. The exception was bare ground cover surrounding parks, which was positively related to total, urban, and resident bird abundance. Bare ground in the surrounding landscape was generally associated with construction sites or vacant lots. Unlike roads and buildings (i.e., impervious surfaces), which isolate and limit the movement of birds within the urban landscape (Fernández-Juricic and Jokimäki, 2001; Tremblay and St Clair, 2011). Bare ground may affect birds at a more local scale, for example, providing habitat for a species such as a Mourning Dove (Zenaida macroura). Regardless, what is clear from our results is that habitat within parks generally had stronger effects than habitat adjacent to parks on birds in LA.
Parks as islands in the cityscape
The theory of island biogeography has been well documented in many natural systems around the world and has been extensively tested in anthropogenic systems under the assumption that larger patches near the ‘mainland’ will harbor greater biodiversity (Molles and Sher, 2018). Our study suggested that parks function as island systems within the urban landscape, which supports previous investigations on this theme (e.g., Fernández-Juricic and Jokimäki, 2001). Larger parks generally had greater bird abundance than smaller ones, which indicates a modification of the classic species-area curve (theoretically focused on richness) suggesting larger parks will harbor more individuals (Zhang and Huang, 2020). Amaya-Espinel et al. (2019), also reported greater bird abundance with the increasing size of urban parks in Santiago, Chile, as did Kang et al., 2015) in remnant urban forest patches of Seoul, Korea. The opposing effects of distance to natural areas on bird compositional patterns implies that the definition of a ‘mainland’ is not uniform for all birds in the urban context. While we defined a mainland as any natural area (i.e., protected areas and open spaces), the mainland for synanthropic species is likely the city itself, as evidenced by the distinct compositional patterns of urban birds in parks further from natural areas (Appendix S1). Taken together, our findings provide strong support that island biogeographic effects explain a significant amount of the variability in bird community patterns within parks throughout LA.
Income inequality, park avifauna, and the virtuous cycle
Like many cities across the world, LA’s park-poor areas are generally embedded within low-income areas of the metropolis. These areas are characterized by high-building density, vast stretches of impervious surface, and little green infrastructure, all of which unsurprisingly provide little habitat for birds that are not synanthropic. Moreover, park-poor areas of LA also have some of the lowest densities of city parks per capita (Wolch et al., 2005), presenting a disproportionate public health concern for human communities (de Vries et al., 2003). Our work details the value of parks in buffering avian communities in park-poor areas and points towards a potential win-win situation when also considering the public health crisis that is prevalent in disadvantaged communities in United States cities. A conceptual approach that highlights this win-win scenario is via a framework for socio-ecological virtuous cycles in conservation (Morrison, 2016). The framework suggests a series of linked objectives that follow a particular intervention to improve conditions for biodiversity, which are interrelated with benefits to individual people and their communities. A potential intervention based on our results is simply park development, which is aggressively being pursued in LA, especially in underserved communities (City of Los Angeles Department of Recreation and Parks, 2019). Given the many known benefits of nature on public health, park development could carry over to improve the well-being of people in cities (e.g., Brown and Grant, 2005). The benefit of parks could then inspire a continued desire for change and improvements within a community. This may, theoretically, lead to sustained benefits to the individuals of a community and the community as a whole. While the application of the conceptual nature of a virtuous cycle is infinitely more complicated in practice, L.A. is providing a model case study for the benefits of park development on its biodiversity and people. Our work strongly supports the benefits of parks to birds in park-poor communities. Follow-up work should blend biodiversity research with the people who utilize parks, including their feelings or beliefs, cultural preferences, and desires for future greening initiatives in their communities to quantify the win-win potential of parks.
Data availability statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Ethics statement
Ethical review and approval was not required for the animal study because this work is fully observational, and no birds were handled during any duration of the study.
Author contributions
AV and EW conceived the research, designed the study, analyzed data and produced figures, and wrote and edited the manuscript. AV conducted fieldwork and data development. All authors contributed to the article and approved the submitted version.
Funding
This work was supported by the Society for Conservation Biology, the Santa Monica Bay Audubon Society, and the California State University Minority Opportunities in Research (MORE), #R25 GM061331.
Acknowledgments
The authors thank A. Trigueros, G. Sercel, N. Garcia, E. Pedroza, C. Benitez, P. Larramendy, and M. Puche for their help with data collection and project establishment. S. Wright and K. Fisher provided valuable feedback on the manuscript. Two reviewers provided helpful comments.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fevo.2022.958572/full#supplementary-material
References
Adobe Inc. (2019). Adobe Illustrator. Available at https://adobe.com/products/illustrator (Accessed June 9, 2022).
Allaire, J., Xie, Y., McPherson, J., Luraschi, J., Ushey, K., Atkins, A., et al. (2022). Rmarkdown: dynamic documents for R. R Package Version 2:14. Available at: https://rmarkdown.rstudio.com/docs/authors.html#citation
Alvey, A. A. (2006). Promoting and preserving biodiversity in the urban forest. Urban For. Urban Green. 5, 195–201. doi: 10.1016/j.ufug.2006.09.003
Amaya-Espinel, J. D., Hostetler, M., Henríquez, C., and Bonacic, C. (2019). The influence of building density on Neotropical bird communities found in small urban parks. Landsc. Urban Plan. 190. doi: 10.1016/j.landurbplan.2019.05.009
Anderson, D. R., and Burnham, K. P. (2002). Avoiding pitfalls when using information-theoretic method. J. Wildl. Manag. 66, 912–918. doi: 10.2307/3803155
Aronson, M. F. J., La Sorte, F. A., Nilon, C. H., Katti, M., Goddard, M. A., Lepczyk, C. A., et al. (2014). A global analysis of the impacts of urbanization on bird and plant diversity reveals key anthropogenic drivers. Proc. R. Soc. B Biol. Sci. 281:20133330. doi: 10.1098/rspb.2013.3330
Aronson, M. F. J., Nilon, C. H., Lepczyk, C. A., Parker, T. S., Warren, P. S., Cilliers, S. S., et al. (2016). Hierarchical filters determine community assembly of urban species pools. Ecology 97, 2952–2963. doi: 10.1002/ecy.1535
Avolio, M. L., Pataki, D. E., Gillespie, T. W., Jenerette, G. D., McCarthy, H. R., Pincetl, S., et al. (2015). Tree diversity in southern California’s urban forest: the interacting roles of social and environmental variables. Front. Ecol. Evol. 3:73. doi: 10.3389/fevo.2015.00073
Avolio, M. L., Pataki, D. E., Trammell, T. L., and Endter-Wada, J. (2018). Biodiverse cities: the nursery industry, homeowners, and neighborhood differences drive urban tree composition. Ecol. Monogr. 88, 259–276. doi: 10.1002/ecm.1290
Beninde, J., Veith, M., and Hochkirch, A. (2015). Biodiversity in cities needs space: a meta-analysis of factors determining intra-urban biodiversity variation. Ecol. Lett. 18, 581–592. doi: 10.1111/ele.12427
Billerman, S. M., Keeney, B. K., Rodewald, P. G., and Schulenberg, T. S. (2021). Birds of the World. Ithaca, NY: Cornell Laboratory of Ornithology.
Blair, R. B. (1996). Land use and avian species diversity along an urban gradient. Ecol. Appl. 6, 506–519. doi: 10.2307/2269387
Brown, C., and Grant, M. (2005). Biodiversity and human health: what role for nature in healthy urban planning? Built Environ. 31, 326–338. doi: 10.2148/benv.2005.31.4.326
Cadenasso, M. L., and Pickett, S. T. A. (2008). Urban principles for ecological landscape design and management: scientific fundamentals. Cities Environ. 1, 1–16. doi: 10.15365/cate.1242008
Cameron, A. C., and Windmeijer, F. A. G. (1997). An R-squared measure of goodness of fit for some common nonlinear regression models. J. Econ. 77, 329–342. doi: 10.1016/S0304-4076(96)01818-0
Carbó-Ramírez, P., and Zuria, I. (2011). The value of small urban greenspaces for birds in a Mexican city. Landsc. Urban Plan. 100, 213–222. doi: 10.1016/j.landurbplan.2010.12.008
Chamberlain, D. E., Henry, D. A. W., Reynolds, C., Caprio, E., and Amar, A. (2019). The relationship between wealth and biodiversity: a test of the luxury effect on bird species richness in the developing world. Glob. Chang. Biol. 25, 3045–3055. doi: 10.1111/gcb.14682
Chesser, R. T., Billerman, S. M., Burns, K. J., Cicero, C., Dunn, J. L., Hernández-Baños, B. E., et al. (2021). Sixty-second supplement to the American ornithological Society’s check-list of North American birds. Auk 138:ukab037. doi: 10.1093/ornithology/ukab037
City of Los Angeles Department of Recreation and Parks. (2019). Available at https://www.laparks.org/measure-aprojects (Accessed January 9, 2022).
Clark, C. J. (2017). eBird records show substantial growth of the Allen’s hummingbird (Selasphorus sasin sedentarius) population in urban Southern California. Condor 119, 122–130. doi: 10.1650/CONDOR-16-153.1
Clarke, L. W., and Jenerette, G. D. (2015). Biodiversity and direct ecosystem service regulation in the community gardens of Los Angeles, CA. Landsc. Ecol. 30, 637–653. doi: 10.1007/s10980-014-0143-7
Clarke, L. W., Jenerette, G. D., and Davila, A. (2013). The luxury of vegetation and the legacy of tree biodiversity in Los Angeles, CA. Landsc. Urban Plan. 116, 48–59. doi: 10.1016/j.landurbplan.2013.04.006
Colding, J., and Folke, C. (2009). The role of golf courses in biodiversity conservation and ecosystem management. Ecosystems 12, 191–206. doi: 10.1007/s10021-008-9217-1
Cornelis, J., and Hermy, M. (2004). Biodiversity relationships in urban and suburban parks in Flanders. Landsc. Urban Plan. 69, 385–401. doi: 10.1016/j.landurbplan.2003.10.038
Davis, A. Y., Belaire, J. A., Farfan, M. A., Milz, D., Sweeney, E. R., Loss, S. R., et al. (2012). Green infrastructure and bird diversity across an urban socioeconomic gradient. Ecosphere 3:art105. doi: 10.1890/ES12-00126.1
de Vries, S., Verheij, R. A., Groenewegen, P. P., and Spreeuwenberg, P. (2003). Natural environments - healthy environments? An exploratory analysis of the relationship between greenspace and health. Environ. Plan. A 35, 1717–1731. doi: 10.1068/a35111
Donnelly, R., and Marzluff, J. M. (2004). Importance of reserve size and landscape context to urban bird conservation. Conserv. Biol. 18, 733–745. doi: 10.1111/j.1523-1739.2004.00032.x
ESRI. (2020). ArcGIS pro (version 2.5). Environmental Systems Research Institute: Redlands, CA, USA.
Ethington, P. J., MacDonald, B., Stein, G., Deverell, W., and Longcore, T. (2020). Historical ecology of the Los Angeles River watershed and environs. Infrastructure for a comprehensive analysis. Report to the John Randolph Haynes and Dora Haynes foundation. Available at https://cawaterlibrary.net/wp-content/uploads/2020/06/ethingtonhaynesfinalreportcompressed-1.pdf (Accessed July 25, 2022).
Fernández-Juricic, E., and Jokimäki, J. (2001). A habitat island approach to conserving birds in urban landscapes: case studies from southern and northern Europe. Biodivers. Conserv. 10, 2023–2043. doi: 10.1023/A:1013133308987
Fiske, I. J., and Chandler, R. B. (2011). Unmarked: an R package for fitting hierarchical models of wildlife occurrence and abundance. J. Stat. Softw. 43, 1–23. doi: 10.18637/jss.v043.i10
Garrett, K. L., Dunn, J. L., and Small, B. E. (2012). Birds of Southern California. Olympia, Washington, United States: R. W. Morse Company.
Grimm, N. B., Faeth, S. H., Golubiewski, N. E., Redman, C. L., Wu, J., Bai, X., et al. (2008). Global change and the ecology of cities. Science 319, 756–760. doi: 10.1126/science.1150195
Han, B., Cohen, D. A., Derose, K. P., Li, J., and Williamson, S. (2018). Violent crime and park use in low-income urban neighborhoods. Am. J. Prev. Med. 54, 352–358. doi: 10.1016/j.amepre.2017.10.025
Helden, A. J., and Leather, S. R. (2004). Biodiversity on urban roundabouts-Hemiptera, management and the species-area relationship. Basic Appl. Ecol. 5, 367–377. doi: 10.1016/j.baae.2004.06.004
Hermy, M., and Cornelis, J. (2000). Towards a monitoring method and a number of multifaceted and hierarchical biodiversity indicators for urban and suburban parks. Landsc. Urban Plan. 49, 149–162. doi: 10.1016/S0169-2046(00)00061-X
Higgins, L. M., Pauly, G. B., Goldman, J. G., and Hood, C. (2019). Wild LA: Explore the Amazing Nature in and Around Los Angeles. Portland, Oregon: Timber Press.
Hope, D., Gries, C., Zhu, W., Fagan, W. F., Redman, C. L., Grimm, N. B., et al. (2003). Socioeconomics drive urban plant diversity. Proc. Natl. Acad. Sci. 100, 8788–8792. doi: 10.1007/978-0-387-73412-5_21
Howes, C., and Reynolds, C. (2021). Absence of a luxury effect on bird alpha diversity in a rapidly developing African city, but surrounding landscape is key. Landsc. Urban Plan. 213:104095. doi: 10.1016/j.landurbplan.2021.104095
Ikin, K., Beaty, R. M., Lindenmayer, D. B., Knight, E., Fischer, J., and Manning, A. D. (2013). Pocket parks in a compact city: how do birds respond to increasing residential density? Landsc. Ecol. 28, 45–56. doi: 10.1007/s10980-012-9811-7
Jenerette, G. D., Miller, G., Buyantuev, A., Pataki, D. E., Gillespie, T. W., and Pincetl, S. (2013). Urban vegetation and income segregation in drylands: a synthesis of seven metropolitan regions in the southwestern United States. Environ. Res. Lett. 8, 1–9. doi: 10.1088/1748-9326/8/4/044001
Jimenez, M. F., Pejchar, L., Reed, S. E., and McHale, M. R. (2022). The efficacy of urban habitat enhancement programs for conserving native plants and human-sensitive animals. Landsc. Urban Plan. 220:104356. doi: 10.1016/j.landurbplan.2022.104356
Johnston, R. F. (2001). “Synanthropic birds of North America”, in Avian Ecology and Conservation in an Urbanizing World. eds. J. M. Marzluff, R. Bowman, R. Donnelly Boston, MA: Springer.
Jokimäki, J., and Suhonen, J. (1998). Distribution and habitat selection of wintering birds in urban environments. Landsc. Urban Plan. 39, 253–263.
Kang, W., Minor, E. S., Park, C. R., and Lee, D. (2015). Effects of habitat structure, human disturbance, and habitat connectivity on urban forest bird communities. Urban Ecosyst. 18, 857–870. doi: 10.1007/s11252-014-0433-5
Kendal, D., Williams, N. S. G., and Williams, K. J. H. (2012). Drivers of diversity and tree cover in gardens, parks and streetscapes in an Australian city. Urban For. Urban Green. 11, 257–265. doi: 10.1016/j.ufug.2012.03.005
Khera, N., Mehta, V., and Sabata, B. C. (2009). Interrelationship of birds and habitat features in urban greenspaces in Delhi, India. Urban For. Urban Green. 8, 187–196. doi: 10.1016/J.UFUG.2009.05.001
Kinzig, A. P., Warren, P., Martin, C., Hope, D., and Katti, M. (2005). The effects of human socioeconomic status and cultural characteristics on urban patterns of biodiversity. Ecol. Soc. 10, NIL_181–NIL_193. doi: 10.5751/ES-01264-100123
La Sorte, F. A., Aronson, M. F. J., Lepczyk, C. A., and Horton, K. G. (2020). Area is the primary correlate of annual and seasonal patterns of avian species richness in urban green spaces. Landsc. Urban Plan. 203:103892. doi: 10.1016/j.landurbplan.2020.103892
La Sorte, F. A., Lepczyk, C. A., Aronson, M. F. J., Goddard, M. A., Hedblom, M., Katti, M., et al. (2018). The phylogenetic and functional diversity of regional breeding bird assemblages is reduced and constricted through urbanization. Divers. Distrib. 24, 928–938. doi: 10.1111/ddi.12738
Leong, M., Dunn, R. R., and Trautwein, M. D. (2018). Biodiversity and socioeconomics in the city: a review of the luxury effect. Biol. Lett. 14:20180082. doi: 10.1098/rsbl.2018.0082
Lepczyk, C. A., La Sorte, F. A., Aronson, M. F. J., Goddard, M. A., Macgregor-Fors, I., Nilon, C. H., et al. (2017). “Global patterns and drivers of urban bird diversity,” in Ecology and Conservation of Birds in Urban Environments. eds. E. Murgui, M. Hedblom Cham: Springer.
Lerman, S. B., and Warren, P. S. (2011). The conservation value of residential yards: linking birds and people. Ecol. Appl. 21, 1327–1339. doi: 10.1890/10-0423.1
Los Angeles County. (2009). Geographic information systems. Available at https://planning.lacounty.gov/gis. (Accessed July 1, 2018).
Los Angeles County. (2016b). Los Angeles countywide comprehensive parks and recreation needs assessment. Available at https://lacountyparkneeds.org/final-report/ (Accessed on February 1, 2020).
Los Angeles County. (2016a). Countywide parks and open space. Available at https://geohub.lacity.org/datasets/0eb692c964734202860a211fa6bbbee8_12/explore?location=34.158028%2C-118.042778%2C10.86 (Accessed June 15, 2018).
Lovasi, G. S., Schwartz-Soicher, O., Quinn, J. W., Berger, D. K., Neckerman, K. M., Jaslow, R., et al. (2013). Neighborhood safety and green space as predictors of obesity among preschool children from low-income families in new York City. Prev. Med. 57, 189–193. doi: 10.1016/J.YPMED.2013.05.012
Loyn, R. H. (1986). The 20 minute search - a simple method for counting forest birds. Corella 10, 58–60.
Luck, G. W., Smallbone, L. T., and O’Brien, R. (2009). Socio-economics and vegetation change in urban ecosystems: patterns in space and time. Ecosystems 12, 604–620. doi: 10.1007/s10021-009-9244-6
MacKenzie, D. I., Nichols, J. D., Pollock, K. H., Royle, J. A., Bailey, L. L., and Hines, J. E. (2017). Occupancy Estimation and Modeling: Inferring Patterns and Dynamics of Species Occurrence. San Diego, CA: Academic Press.
Magle, S. B., Fidino, M., Sander, H. A., Rohnke, A. T., Larson, K. L., Gallo, T., et al. (2021). Wealth and urbanization shape medium and large terrestrial mammal communities. Glob. Chang. Biol. 27, 5446–5459. doi: 10.1111/gcb.15800
Martin, C. A., Warren, P. S., and Kinzig, A. P. (2004). Neighborhood socioeconomic status is a useful predictor of perennial landscape vegetation in residential neighborhoods and embedded small parks of Phoenix, AZ. Landsc. Urban Plan. 69, 355–368. doi: 10.1016/j.landurbplan.2003.10.034
McKinney, M. L. (2002). Urbanization, biodiversity, and conservation. Bioscience 52, 883–890. doi: 10.1641/0006-3568(2002)052[0883:UBAC]2.0.CO;2
Molles, M. C., and Sher, A. (2018). Ecology: Concepts and Applications (8th Edn.). New York, NY: McGraw-Hill Education.
Morrison, S. A. (2016). Designing virtuous socio-ecological cycles for biodiversity conservation. Biol. Conserv. 195, 9–16. doi: 10.1016/j.biocon.2015.12.022
Nielsen, A. B., van den Bosch, M., Maruthaveeran, S., and van den Bosch, C. K. (2014). Species richness in urban parks and its drivers: a review of empirical evidence. Urban Ecosyst. 17, 305–327. doi: 10.1007/s11252-013-0316-1
Oksanen, J., Guillaume, B. F., Friendly, M., Kindt, R., Legendre, P., McGlinn, D., et al. (2019). Vegan: community ecology package. R Package Version 2, 5–4.
Pettorelli, N., Ryan, S., Mueller, T., Bunnefeld, N., Jedrzejewska, B., Lima, M., et al. (2011). The normalized difference vegetation index (NDVI): unforeseen successes in animal ecology. Clim. Res. 46, 15–27. doi: 10.3354/cr00936
R Core Team (2017). R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna.
Rojas, I. M., Jennings, M. K., Conlisk, E., Syphard, A. D., Mikesell, J., Kinoshita, A. M., et al. (2022). A landscape-scale framework to identify refugia from multiple stressors. Conserv. Biol. 36, e13834–e13811. doi: 10.1111/cobi.13834
Royle, J. A., and Nichols, J. D. (2003). Estimating abundance from repeated presence-absence data or point counts. Ecology 84, 777–790. doi: 10.1890/0012-9658(2003)084[0777:EAFRPA]2.0.CO;2
Schell, C. J., Dyson, K., Fuentes, T. L., Roches, S. D., Harris, N. C., Miller, D. S., et al. (2020). The ecological and evolutionary consequences of systemic racism in urban environments. Science 369:eaay4497. doi: 10.1126/SCIENCE.AAY4497
Schwarz, K., Fragkias, M., Boone, C. G., Zhou, W., Mchale, M., Grove, J. M., et al. (2015). Trees grow on money. Urban Tree Canopy Cover Environ. Justice 10, 1–17. doi: 10.1371/journal.pone.0122051
Seress, G., and Liker, A. (2015). Habitat urbanization and its effects on birds. Acta Zool. Acad. Sci. Hung. 61, 373–408. doi: 10.17109/AZH.61.4.373.2015
Southern California Association of Governments. (2016). 2016 median household income (MHI) in dollars. Available at https://maps.scag.ca.gov/scaggis/rest/services/ATDB/ATDB_ATP_Requirements/MapServer/layers (Accessed October 5, 2022).
Stein, E. D., Dark, S., Longcore, T., Hall, N., Beland, M., Grossinger, R., et al. (2007). Historical Ecology and Landscape Change of the San Gabriel River and Floodplain. Costa Mesa, California.
Strohbach, M. W., Haase, D., and Kabisch, N. (2009). Birds and the City: urban biodiversity, land use, and socioeconomics. Ecol. Soc. 14, 1–16. doi: 10.5751/ES-03141-140231
Talal, M. L., and Santelmann, M. V. (2019). Plant community composition and biodiversity patterns in urban parks of Portland. Oregon. Front Ecol Evol 7, 1–16. doi: 10.3389/fevo.2019.00201
Tremblay, M. A., and St Clair, C. C. (2011). Permeability of a heterogeneous urban landscape to the movements of forest songbirds. J. Appl. Ecol. 48, 679–688. doi: 10.1111/j.1365-2664.2011.01978.x
Trust for Public Land. (2021). Park score index. Available at https://www.tpl.org/parkscore (Accessed October 3, 2022).
U.S. Census Bureau (2019). Los Angeles County, CA. Quick Facts. Available at: https://www.census.gov/quickfacts/losangelescountycalifornia (Accessed October 3, 2022).
U.S. Census Bureau/American FactFinder. (2010). U.S. Census Bureau, 2010.Web. Available at http://factfinder2.census.gov (Accessed January 1, 2013).
Vallejo, B. M., Aloy, A. B., and Ong, P. S. (2009). The distribution, abundance and diversity of birds in Manila’s last greenspaces. Landsc. Urban Plan. 89, 75–85. doi: 10.1016/j.landurbplan.2008.10.013
Venter, Z. S., Shackleton, C. M., Van Staden, F., Selomane, O., and Masterson, V. A. (2020). Green Apartheid: Urban green infrastructure remains unequally distributed across income and race geographies in South Africa. Landsc. Urban Plan. 203:103889.
Villaseñor, N. R., and Escobar, M. A. H. (2019). Cemeteries and biodiversity conservation in cities: how do landscape and patch-level attributes influence bird diversity in urban park cemeteries? Urban Ecosyst. 22, 1037–1046. doi: 10.1007/s11252-019-00877-3
Wang, H.-F., Qureshi, S., Knapp, S., Friedman, C. R., and Hubacek, K. (2015). A basic assessment of residential plant diversity and its ecosystem services and disservices in Beijing, China. Appl. Geogr. 64, 121–131. doi: 10.1016/J.APGEOG.2015.08.006
Wolch, J., Wilson, J. P., and Fehrenbach, J. (2005). Parks and park funding in Los Angeles: an equity-mapping analysis. Urban Geogr. 26, 4–35. doi: 10.2747/0272-3638.26.1.4
Wood, E. M., and Esaian, S. (2020). The importance of street trees to urban avifauna. Ecol. Appl. 30, e02149–e02120. doi: 10.1002/eap.2149
Wood, E. M., Pidgeon, A. M., Radeloff, V. C., Helmers, D., Culbert, P. D., Keuler, N. S., et al. (2014). Housing development erodes avian community structure in U.S. protected areas. Ecol. Appl. 24, 1445–1462. doi: 10.1890/12-1992.1
Wood, E. M., Pidgeon, A. M., Radeloff, V. C., Helmers, D. P., Culbert, P. D., Keuler, N. S., et al. (2015). Long-term avian community response to housing development at the boundary of US protected areas: effect size increases with time. J. Appl. Ecol. 52, 1227–1236. doi: 10.1111/1365-2664.12492
Wood, E. M., Pidgeon, A. M., Radeloff, V. C., and Keuler, N. S. (2013). Image texture predicts avian density and species richness. PLoS One 8:e63211. doi: 10.1371/journal.pone.0063211
Wu, J. (2010). Urban sustainability: an inevitable goal of landscape research. Landsc. Ecol. 25, 1–4. doi: 10.1007/s10980-009-9444-7
Xu, X., Xie, Y., Qi, K., Luo, Z., and Wang, X. (2018). Detecting the response of bird communities and biodiversity to habitat loss and fragmentation due to urbanization. Sci. Total Environ. 624, 1561–1576. doi: 10.1016/j.scitotenv.2017.12.143
Young, J. K., Golla, J., Draper, J. P., Broman, D., Blankenship, T., and Heilbrun, R. (2019). Space use and movement of urban bobcats. Animals 9:275. doi: 10.3390/ani9050275
Zhang, Z., and Huang, G. (2020). How do urban parks provide bird habitats and birdwatching service? Evidence from Beijing. China. Remote Sens (Basel) 12:3166. doi: 10.3390/rs12193166
Zivanovic, A. J., and Luck, G. W. (2016). Social and environmental factors drive variation in plant and bird communities across urban greenspace in Sydney, Australia. J. Environ. Manag. 169, 210–222. doi: 10.1016/j.jenvman.2015.11.052
Keywords: avifauna, biodiversity, LiDAR, park score, remote sensing, socioeconomic, luxury effect
Citation: Vasquez AV and Wood EM (2022) Urban parks are a refuge for birds in park-poor areas. Front. Ecol. Evol. 10:958572. doi: 10.3389/fevo.2022.958572
Edited by:
Zoltan Elek, Hungarian Academy of Sciences (MTA), HungaryReviewed by:
Jana Růžičková, ELKH-ELTE-MTM Integrative Ecology Research Group, HungaryZsolt Végvári, Hungarian Academy of Science, Hungary
Copyright © 2022 Vasquez and Wood. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Eric M. Wood, ZXJpY213b29kQGNhbHN0YXRlbGEuZWR1