 Alois Honek
Alois Honek Zdenka Martinkova
Zdenka Martinkova Jiří Skuhrovec
Jiří Skuhrovec- Functional Diversity Group, Crop Research Institute, Prague, Czechia
From 2014 to 2022, we investigated the length of hibernation in a population of the invasive ladybird Harmonia axyridis (Pallas) (Coleoptera: Coccinellidae) in Central Europe. Adults were collected during the autumn immigration to overwintering sites and placed in an artificial hibernaculum held in an unheated closed shelter. The following spring, the population was monitored for timing of emigration from this hibernaculum. In individual years, massive immigration took place between late September and late October and immigration lasted one to 3 days, each year. The main period of emigration from the hibernaculum (extending from the time that 25% of adults departed the hibernaculum to the time that 75% of the remaining individuals departed) occurred in individual years between mid-April and early May and lasted 8.0 ± 1.3 days (mean ± SE). Massive emigration activity followed a gradual period of increasing temperature. Although the dates of migration in some years, both autumn and spring, ranged over a period of 1 month, the overwintering period (from immigration to emigration of 50% of the population) was similar each year, 185.0 ± 6.3 days (mean ± SE). We hypothesize that the length of overwintering may be fixed by an intrinsic interval timer in H. axyridis adults. The internal timing of the end of hibernation may be advantageous in the cold temperate zone, where H. axyridis adults overwinter in buildings and are thus isolated from external photoperiodic and temperature stimuli that signal the end of the winter season.
Introduction
Overwintering is an important part of the life cycle of ladybirds (Coleoptera: Coccinellidae). Species inhabiting the temperate zone of the Northern Hemisphere overwinter during the adult stage (Nedved, 2020). The course and factors of overwintering have been studied in detail in a number of species (Hodek, 1960; Iperti, 1966). Successful overwintering is conditioned by induction of diapause and, after its termination, by thermal quiescence, which prolongs hibernation until the onset of favorable conditions in the spring. The genetic and neuroendocrine mechanisms governing induction, course and termination of dormancy are well studied (Danilevskii, 1965; Tauber et al., 1986; Danks, 1987). This also applies to many species of coccinellids (Hodek, 2012), including Harmonia axyridis (Pallas) (Roy et al., 2016; Raak-van den Berg et al., 2017). Overwintering of this species has been studied both in its native regions of East Asia (Tanagishi, 1976; Obata, 1986; Sakurai et al., 1993; Wang et al., 2011) and in invaded regions of Europe (Raak-van den Berg et al., 2012a, 2013) and North America (Berthiaume et al., 2003; Labrie et al., 2008). Weather conditions enabling autumn migration flights (Honek et al., 2018), structures attracting adults for hibernation (Nalepa et al., 2005), and survival during the winter were studied (Raak-van den Berg et al., 2012b). However, there are few studies that have investigated the factors that affect the duration of the overwintering period.
We investigated this issue from 2014 to 2022, by monitoring the course and dynamics of overwintering of a local non-native H. axyridis population at Prague-Ruzyně, in Central Europe. Migration to overwintering grounds, length of stay in hibernaculum, and dynamics involved in leaving hibernaculum were studied. This research is important because it provides baseline knowledge on causes of variation in the length of overwintering in a highly invasive species of worldwide importance.
Materials and methods
Study site
Adults of H. axyridis were collected and overwintering experiments were conducted on the grounds of the Crop Research Institute in Prague-Ruzyně (an area of 0.32 km2 centered at 50.0853N, 14.3008E, at 340 m a.s.l.). The central building of the institute was surrounded by an old garden and experimental and production field plots at a distance of more than 800 m away from industrial buildings in an urban area. Garden vegetation and field crop stands were regularly infested with aphid populations, which provided an opportunity for the establishment of ladybird communities, in which the dominant species was H. axyridis (Honek et al., 2019). Meteorological data (available from the authors of the publication) was recorded at a meteorological station located inside the institute’s premises.
Collecting migrants to hibernacula
Adults of H. axyridis used in overwintering experiments were collected during early autumn each year for one to 3 days during their massive migration to hibernacula (Figure 1). Flying adults were collected when they landed on the southern facing wall of the central building on a 15 m long and 2 m high section of facade accessible from a balcony 7 m above the surface of the surrounding garden, at 50.0864592N, 14.3027442E. In all years of the study, individuals were collected by one person (AH) using a standard methodology. This person slowly passed the balcony back and forth and collected all individuals crawling on the facade. The number of captured individuals and their color form were recorded at approximately 15 min intervals (further called the sampling interval). The collected individuals were put in plastic bottles, stored for 2–3 days at 5°C in the dark and then transferred to an artificial hibernaculum.
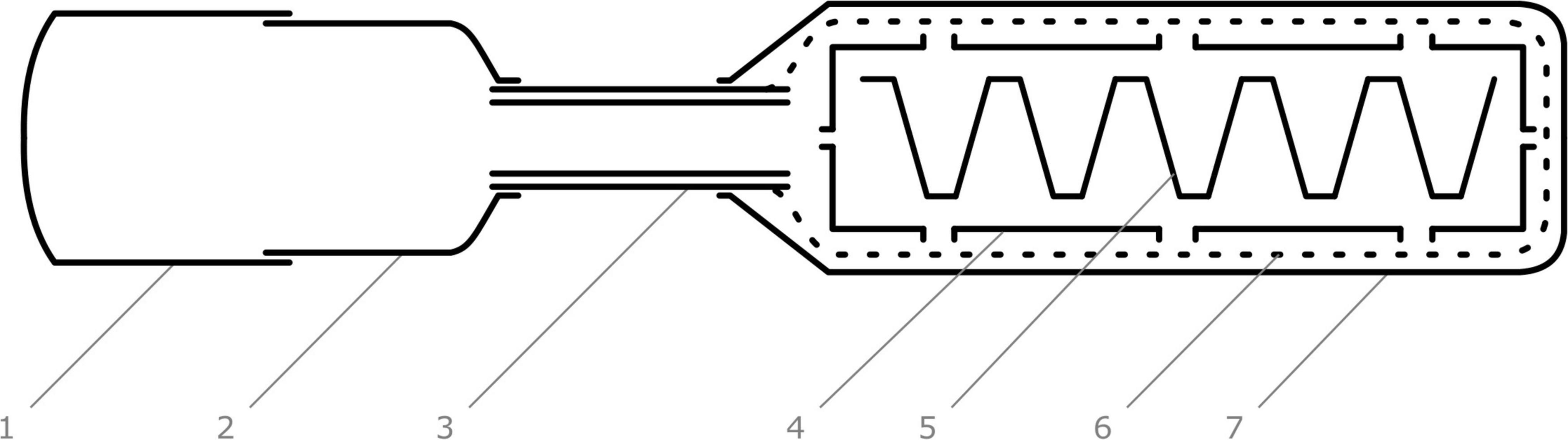
Figure 1. Cross-section of artificial hibernaculum, state at the time when spring emigration of adults was studied. Individual components are marked with labels. 1–collection bottle, lower part, 2–collection bottle, upper part, 3–plastic tube, 4–overwintering box, 5–egg cardboard, 6–gauze bag, 7–opaque plastic bag.
Overwintering in an artificial hibernaculum
To monitor overwintering populations, we used artificial hibernacula because the shelters, which adults entered randomly, were not accessible for observation. This hibernaculum was placed in conditions probably very similar to those that prevailed in the spaces occupied by the adults. We therefore assumed that the behavior of ladybirds in an artificial hibernaculum (during the overwintering season and the dynamics of leaving the hibernaculum in the spring) was the same as the behavior of ladybirds in randomly selected hibernacula. The artificial hibernaculum (Figure 1) consisted of 33 × 19 × 8 cm corrugated cardboard overwintering boxes with inserted egg cardboards, which enlarged the inner surface of the box suitable for adult aggregation. The sides and the lid of the box were punctured with holes of 1 cm diameter, which allowed the adults to leave the box. The box was placed in a gauze bag enabling free ventilation. At the start of the test, 300–500 individuals were placed in each of the two boxes that were replicates of the experiment.
Emigration from hibernacula
Emigration from the artificial hibernaculum was monitored every year (2014–2022) since the beginning of March. Overwintering boxes with gauze bags were placed in opaque plastic bags. Attached to the mouth of the bags was a transparent plastic tube (length 12 cm, diameter 3.5 cm) opening into a transparent plastic collection bottle (length 21 cm, diameter 8.5 cm). Adults that gradually left the overwintering box were collected in the bottle. The lower part of this bottle could be ejected, the collected adults could be removed, and their color form and sex were recorded. The collection bottle was emptied at 2- or 3-day intervals, in the early morning when the adults were not yet able to move extensively due to low temperature. Mortality (percentage of dead individuals found inside the overwintering box out of the total number of individuals introduced into artificial hibernaculum) was determined at the end of May, after emigration of all surviving adults.
Data elaboration
Herein, we defined the overwintering period as the interval between the term immigration (the day of mass flight activity when the experimental adults were collected, Figure 2) and the term emigration (the day when 50% of individuals who that survived hibernation left the artificial hibernaculum). To evaluate flight activity during days of immigration, for each sampling interval, its beginning (initial time Ti), end (final time Tf), and the median time for the given interval (Tm) were recorded, which was calculated as Tm = (Ti + Tf)/2. For each sampling interval, the immigration rate (number of individuals captured in 1 min, Nt) and the percentage of melanic individuals (f. conspicua, f. spectabilis, and f. axyridis together, Pm, calculated only for samples where n ≥ 10) in the total population consisting mainly of f. succinea, further called the light form (Honek et al., 2020), were calculated. The overall abundance of H. axyridis differed among years (Honek et al., 2019). To compensate for these differences, the values of Nt in individual years were standardized by transformation to z scores, which is the distance of a count from the mean value in standard deviation units. Z scores were calculated using the formula z = (x–m)/s, where x is the arithmetic mean of the values of Nt for the day of sampling, m are the values of Nt in particular sampling intervals and s is the standard deviation of the Nt the day sampling. Z scores of Nt and percentage of melanics Pm were plotted against Tm of a given sampling interval. The time trends of the change in immigration rate Nt and the change in percentage of melanics Pm during the day were determined by interpolating the polynomial function of the second degree.

Figure 2. Days on which immigration flight to the overwintering sites took place. Shaded symbols indicate dates when flight activity was intense, open symbols indicate dates when flight activity was small. Asterisked are the dates when adults were collected for experiments with overwintering in artificial hibernacula (in each year one day when flight activity was high). The picture also contains data obtained in 2009–2013, when experiments with artificial hibernacula were not made.
The course of emigration in particular years was determined by plotting the cumulative percentage of individuals who left the hibernaculum against the day of their leaving. Calculations were done on the days in which 5% adults (D0.05), 25% adults (D0.25), 50% adults (D0.50), 75% adults (D0.75), and 95% (D0.95) adults left the hibernaculum. These days were determined by linear interpolation between the percentage for the last day before and the first day after the respective abovementioned percentage criterion were attained. Day D0.50 was considered the term emigration from hibernacula, and its calculation was performed for all individuals, separately for males and females, and separately for light and melanic forms. D0.25 and D0.75 defined the period of mass emigration, and D0.05 and D0.95 characterized the beginning and end of the period of emigration better than the dates of emigration of the first and the last individual leaving hibernaculum.
The agreement between the dynamics of emigration of males and females and the light and melanic forms was tested by the Kolmogorov–Smirnov (K-S) test. Annual differences in the length of hibernation period (set as number of days elapsed from the autumn mass migration to emigration from artificial hibernacula) were tested using analysis of variance (ANOVA) with length of hibernation period as the response variable and years as the factor. Because the normality test on distribution of length of hibernation period in particular years failed, Kruskal–Wallis ANOVA (on ranks) was used. Negative results of the ANOVA indicated the absence of significant differences in the length of the overwintering period among particular winter seasons included in this study. All calculations were performed using Sigma Stat (Systat Software, 2006).
Results
The first date of immigration to hibernacula was observed in different years between 27 September (2017) and 18 October (2021) (Figure 2). Flight activity on the days of autumn mass migration had a characteristic time course. The numbers of individuals landing on the wall of the building during the main flight period were highly variable, somewhat lower at the beginning and end of this period. A meaningful relationship was found between time of day and intensity of immigration (Figure 3). The peak of flight activity was observed at 12:50 Central European Time, when flight activity was above average (z score = +0.44). The ratio of color forms in the samples captured during the daytime was highly variable, and a slightly increased percentage of melanistic forms was found in the samples captured in the morning (Figure 4).
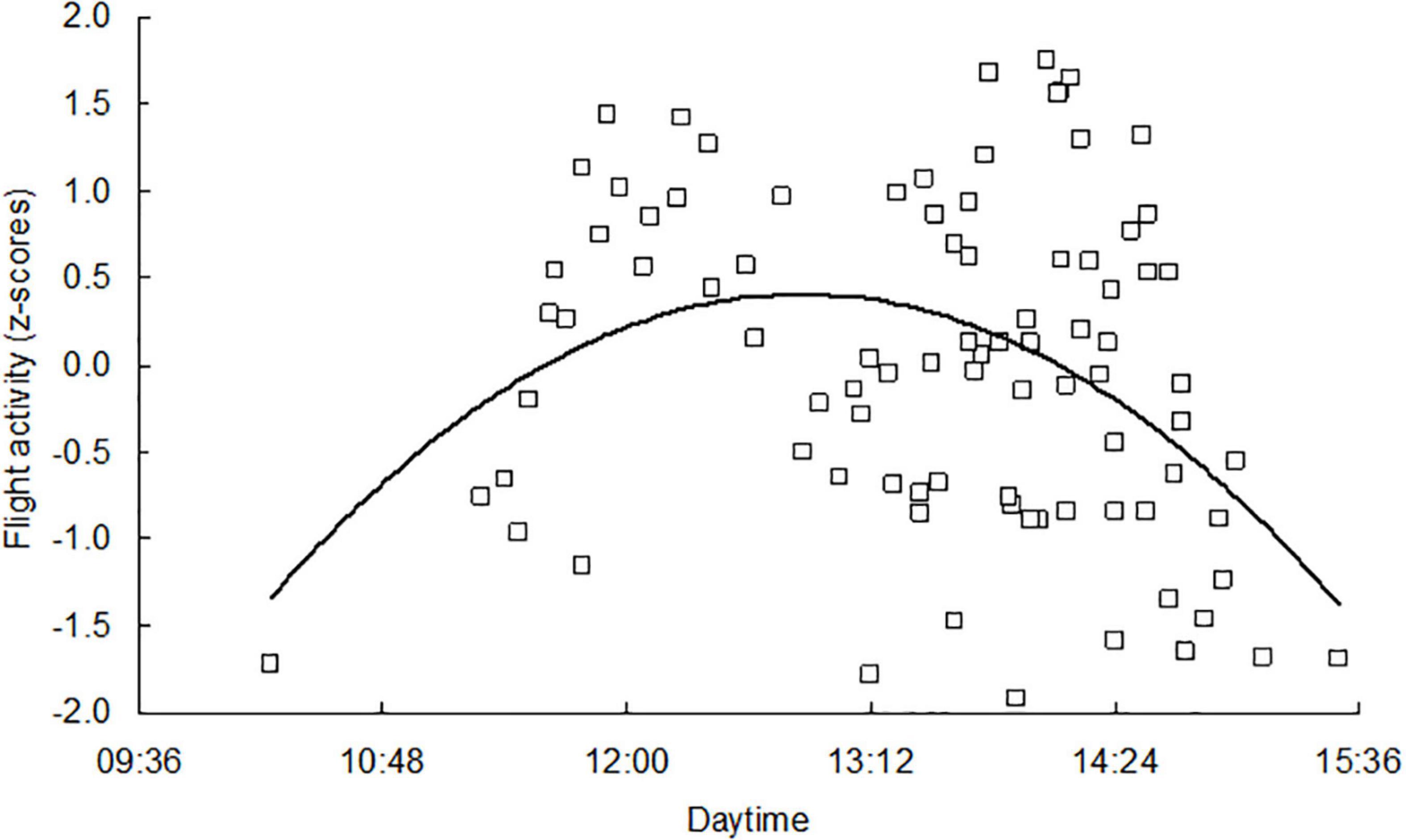
Figure 3. Flight activity of adults immigrating to overwintering sites, aggregate data from 2013 to 2021. Individual symbols indicate the number of adults sampled per 1 min on a standard section of facade (Nt transformed as z scores) on days of mass flight to overwintering sites (shaded symbols in Figure 2). Fitted by second order polynomial y = –147.38x2 + 157.95x–41.91, N = 89, R2 = 0.1537, P = 0.001.
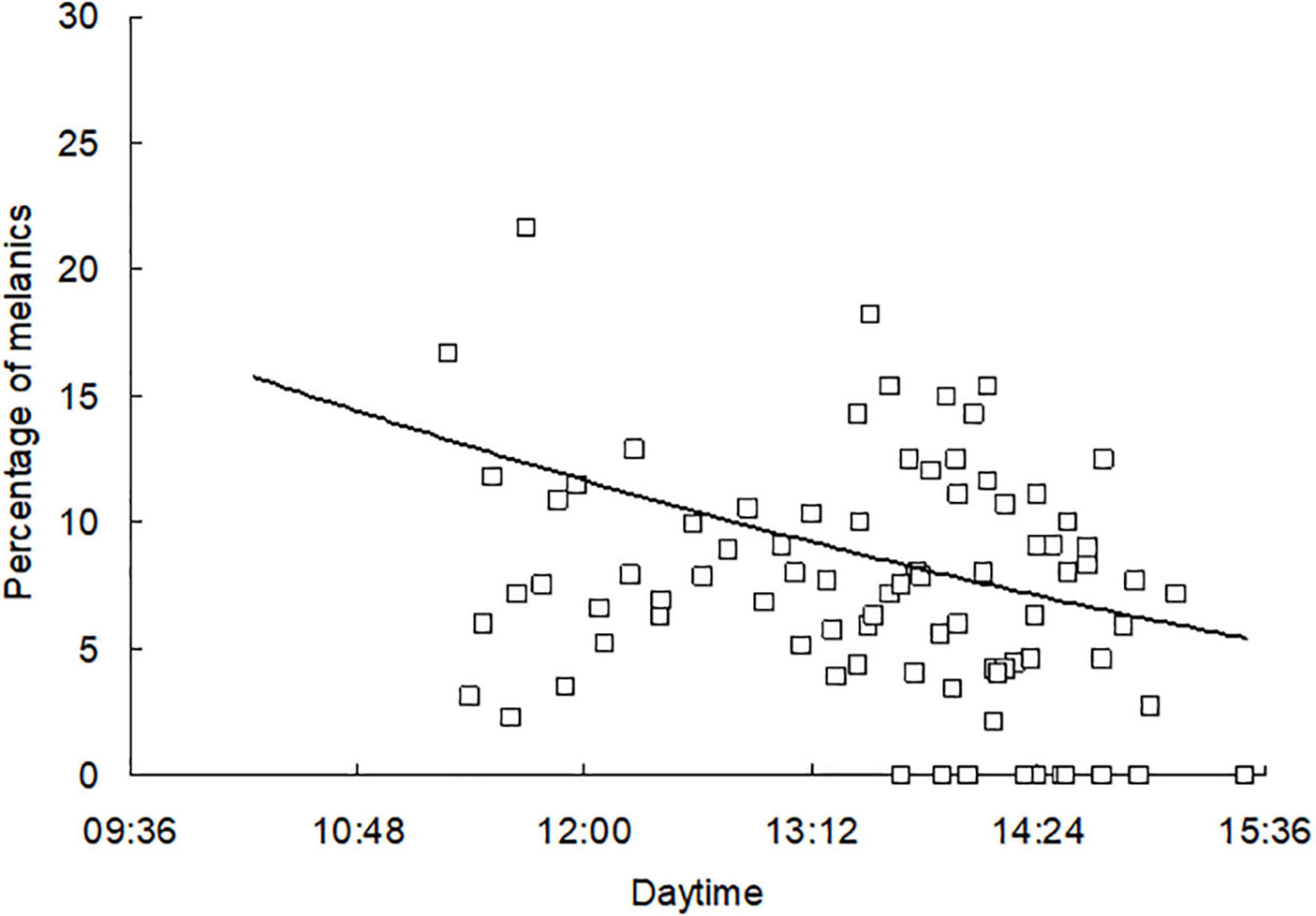
Figure 4. Flight activity of the color forms of adults migrating to overwintering sites; aggregate data from 2013 to 2021. Individual symbols indicate the percentage of melanic adult Pm in samples of adults arriving at the building facade at 15-min intervals on days of mass flight to overwintering sites (shaded symbols in Figure 2). Three values highly exceeding 30% are not shown. The data were fitted by the second-order polynomial y = 60.36x2–112.04x + 52.60, N = 89, R2 = 0.0476, P = 0.050.
The massive flight to the hibernacula took place on a calm day with intense sunshine and increased temperature. This day usually followed a period of weather that was unsuitable for flight. The migration thus took place on a day when the conditions were relatively favorable compared to the conditions of the previous days (Figure 5A). However, it was not possible to find a universal value of temperature (temperature threshold for flight) necessary for starting autumn migration.
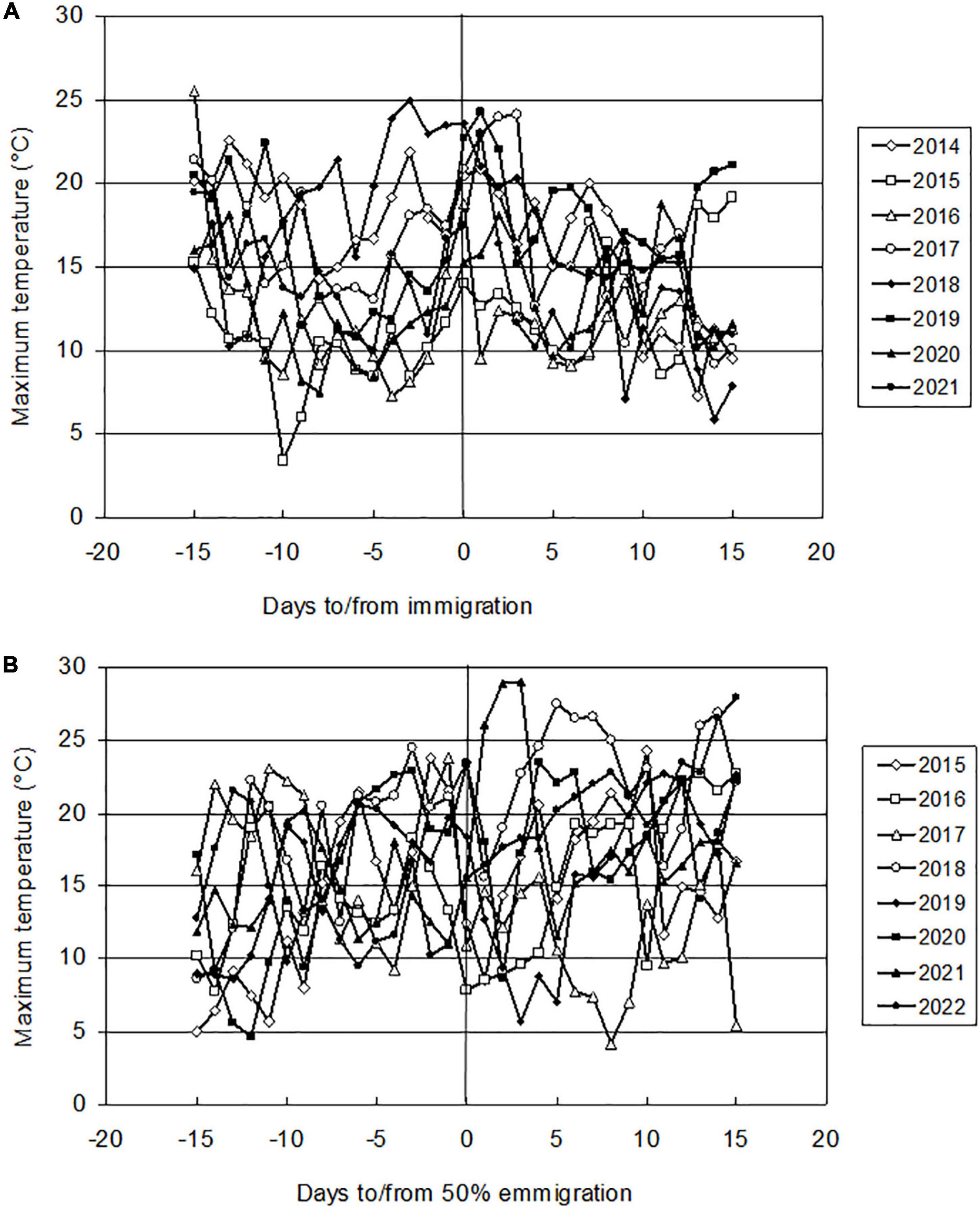
Figure 5. Air temperatures (ordinate: daily maxima measured by the standard meteorological method) during the period of immigration and emigration from wintering grounds. (A) Immigration to the overwintering sites. Abscissa: 0–day of massive migration flight activity, negative values indicate days before this term, positive values indicate days after this term. (B) Emigration from artificial hibernaculum. Abscissa: 0–the day when 50% of individuals left the artificial hibernaculum (D0.50), negative values mean days before D0.50, positive values mean days after D0.50.
The percentage of emigration plotted against time had a characteristic S-shape (Figure 6). Five percent of emigrants (D0.05) left the hibernacula on average on 5 April (±10.1 days), half of the emigrants (D0.50) left the winter quarters on 19 April (±9.8 days), and 95% of emigrants (D0.95) left the winter quarters on 2 May (±8.5 days) (Table 1). The highest rate of emigration (number of individuals leaving hibernacula per day) was during the period between D0.25 and D0.75. This period of massive emigration was interrupted in some years by a cold period that prevented movement of adults (lasting 3 days in 2020, 7 days in 2019, 11 days in 2022, 15 days in 2017). If the length of the emigration period was corrected for these periods of temperature-forced interruptions, the period of massive emigration lasted 4–14 days with a mean ± SE of 8 ± 1.2 days.
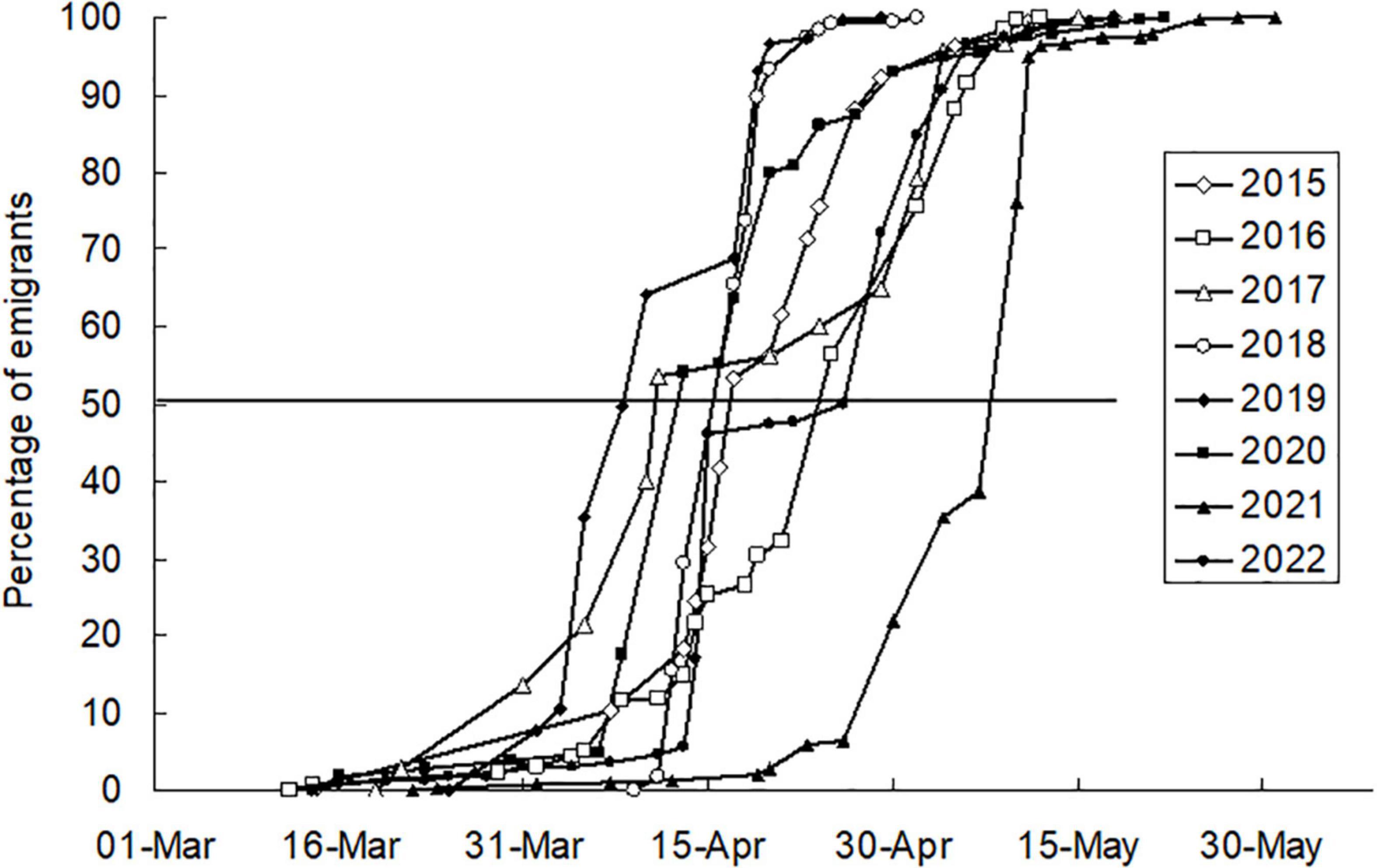
Figure 6. Emigration from an artificial hibernaculum, 2015–2022. Cumulative percentage of individuals who that left the artificial hibernaculum plotted against the date of emigration.
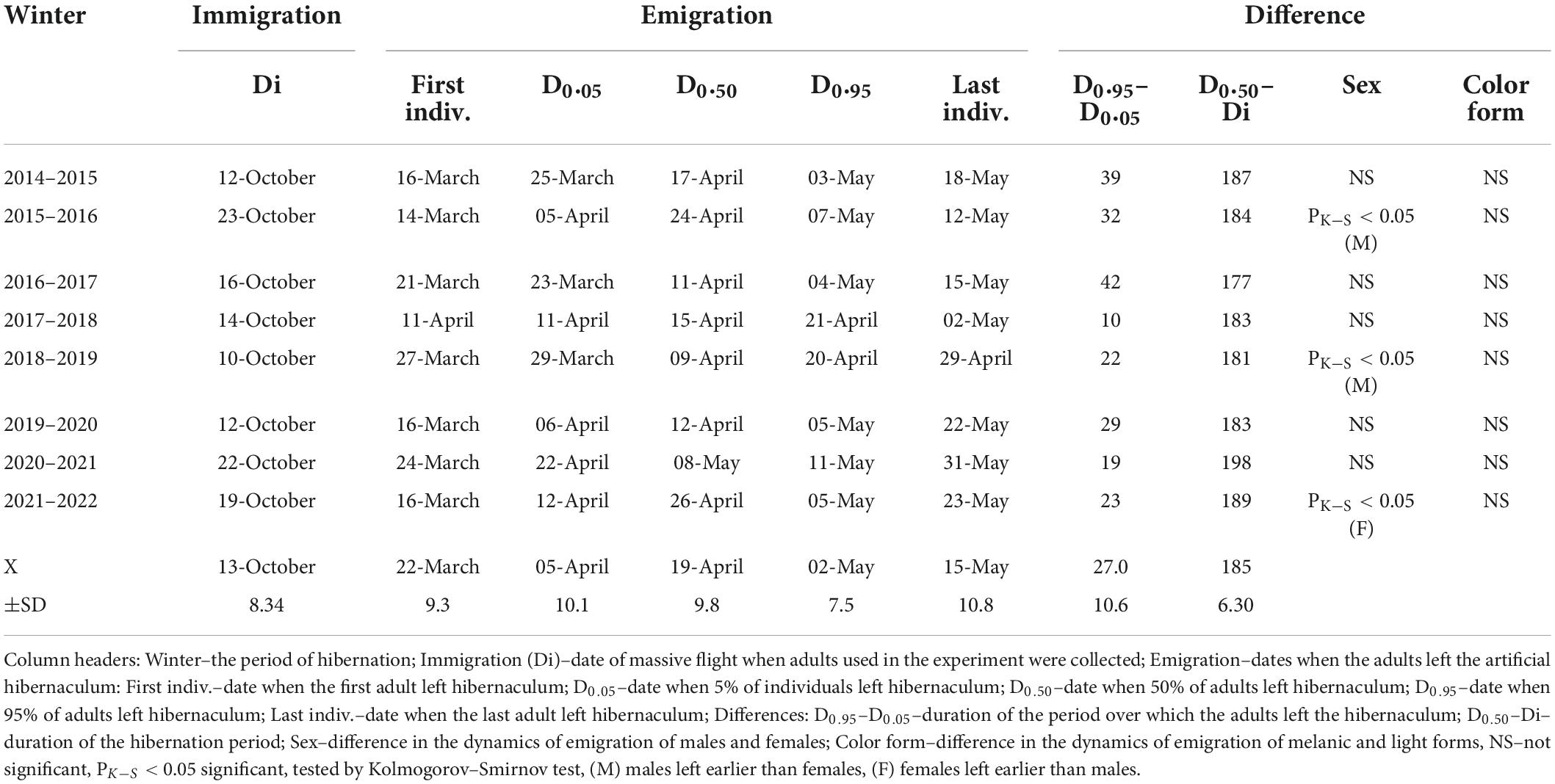
Table 1. Immigration and emigration of adults from artificial hibernaculum.
A significant difference between males and females in the temporal distribution of leaving the hibernaculum was found in 3 years; males left the hibernacula earlier in 2016 and 2019, and females left it earlier in 2022 (Table 1). No significant difference in the temporal distribution of leaving the hibernaculum was found between melanic and non-melanic forms (Table 1).
The period of intensive emigration from the hibernacula usually coincided with a period of increased outdoor temperature (Figure 5B). Due to the large annual variation in dates when mass emigration took place, there were large differences in the temperature conditions of this period. Therefore, it was not possible to unambiguously determine the temperature requirements necessary for emigration.
The time that adults spent in the artificial hibernaculum from the day of immigration flight to the day when half of the emigrants left hibernaculum (D0.50) fluctuated between 177 days (in 2016–2017) and 198 days (in 2020–2021) with a mean ± SE of 185 ± 6.3 days. The low variability in the duration of the period spent in the hibernaculum was because the dates of the beginning (day of massive autumn immigration) and the end (D0.50) of the overwintering period were positively correlated (Figure 7). The results of the analysis of variance showed the absence of a significant difference in the length of stay in the artificial hibernacula in seven of the eight winter periods examined in our study (Kruskal–Wallis test, H = 5.823, df = 6, P = 0.443, median length of the hibernation period between 179 days in 2016–2017 and 189 days in 2021–2022). A significantly longer hibernation period (median–199 days) was found in 2020–2021, when the reason for the delay in emigration from the hibernacula (Figure 6) was the low temperature prevailing from April 1 to 20 (mean ± SE, 5.5 ± 0.85°C). Mortality during the hibernation period (percentage of dead individuals found inside the overwintering box out of the total number of individuals introduced into artificial hibernacula) ranged from 3.4% (in 2021–2022) to 25.2% (in 2014–2015), with a mean ± SE of 14.7 ± 3.57%.
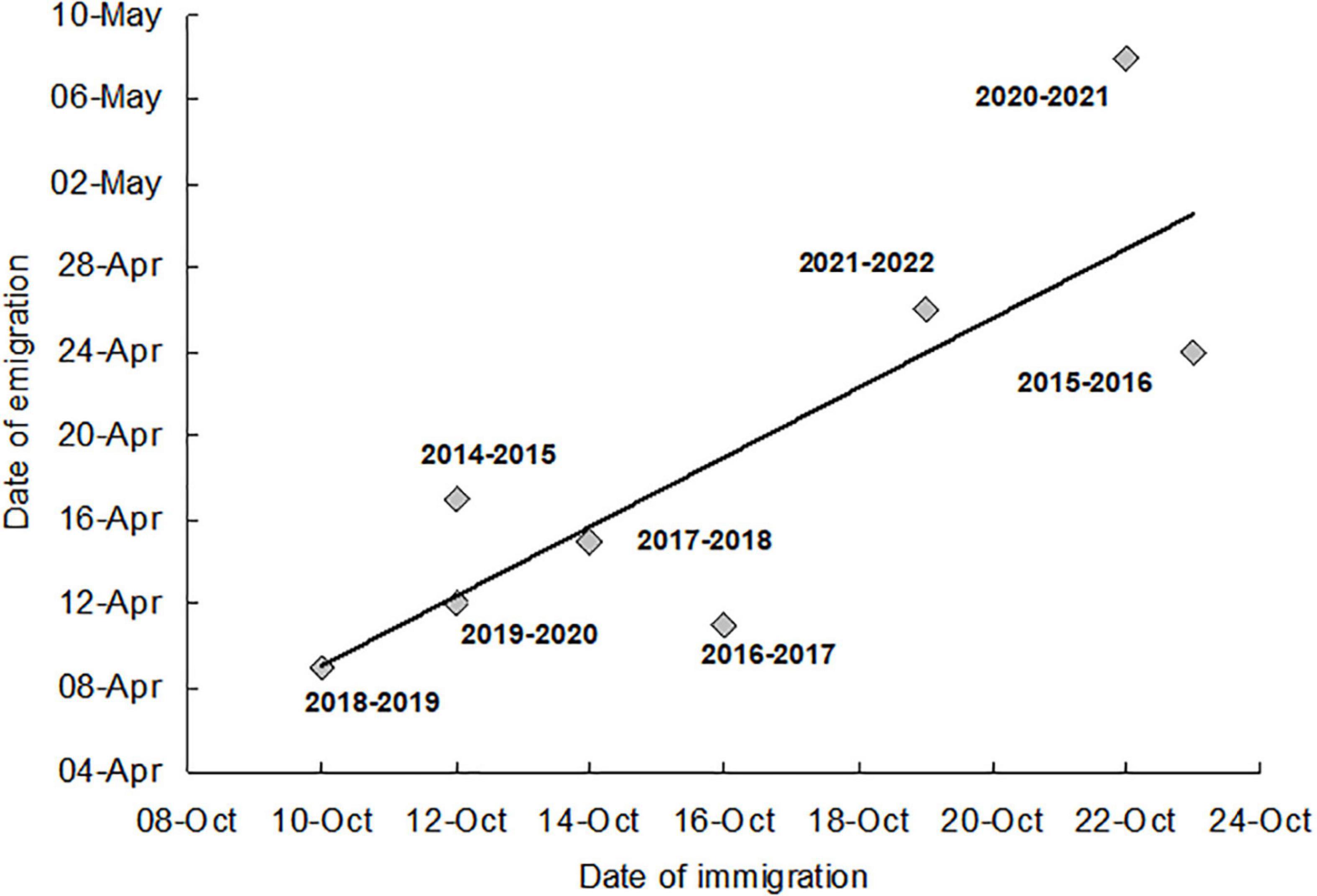
Figure 7. Dates of emigration from the artificial hibernaculum (the day when 50% of individuals left the artificial hibernaculum D0.50, abscissa), plotted against the dates of immigration to overwintering sites (day of massive migratory flight activity, ordinata). The symbols indicate the hibernation period from which the dates of immigration and emigration originate. y = 42.369 + 1.488x, N = 8, R2 = 0.600, P = 0.024.
Discussion
In the zone of cold temperate climate in North America and Europe, a significant portion of the adults in H. axyridis populations overwinter in buildings (Schaefer, 2003) or other artificial structures (Dudek et al., 2015), and their survival in winter conditions depends on these man-made hibernacula (Labrie et al., 2008). The adults fly to buildings and settle inside spaces (Nalepa, 2007). The adults collected during a massive autumn flight on the building facade belonged to this major part of the population overwintering in man-made shelters. The characteristics of the onset, end, and length of the overwintering period established in this study thus likely apply to a large portion of the local H. axyridis population.
The trigger for an immigration flight to hibernacula is the onset of suitable weather. Conditions convenient for flight are high temperatures, calm winds, and intense sunlight (Raak-van den Berg et al., 2012a). Our results (Figure 5A) suggest that a threshold temperature for triggering autumn migration is difficult to determine. The reason for this difficulty is the unsteady timing of the day of massive migration. The dates of the first day of mass migration to hibernacula differed by 26 days. The daily temperature maximum, which was approximately equal to the temperature reached during the period of maximum flight activity, ranged from 14.0°C (23.10.2015) to 15.3°C (22.10.2020) in the years when immigration was late to 22.7°C (27.9.2018 and 12.10.2019) and 23.9°C (10.10.2018) in the years when immigration occurred at an early date. The average temperature of the previous 5 days was lower by 1.1°C (2014) to 6.0°C (2019). The migration therefore took place on a day when the temperature was favorable (adequately to the season of year when the migration took place) compared to the conditions of the previous days. Due to the large variability in weather conditions in particular years, it was not possible to find a universal temperature threshold for migration flight or to define a sequence of conditions that would trigger autumn migration (Figure 5A).
It is even more difficult to establish the influence of weather on emigration from hibernacula. In the hibernation site hidden inside the building, the effect of weather conditions outside the buildings is damped: the amplitude of temperature fluctuations is reduced, the onset of daily minima and maxima is delayed (Havlíček, 1986), and the effect of photoperiod is ruled out. How (and whether at all) changes in external conditions are perceived and evaluated by adults overwintering inside buildings needs further investigation.
During our observation period, immigration dates were positively correlated with emigration dates. The assumption is that this correlation is not a phenomenon that occurs randomly in a series of eight consecutive winter periods (from 2014–2015 to 2021–2022) leading to the conjecture that the length of overwintering is limited by an intrinsic mechanism. Here, we propose the hypothesis that the date of emigration and ipso facto the length of the overwintering period are determined by an intrinsic interval timer. This interval timer is “triggered” by flight to winter quarters and determines the time of leaving the winter quarters. After the completion of the predetermined interval, the adults only wait for the onset of relatively suitable weather conditions. Of course, there is no guarantee that the observed 8-year series of results will continue in the future. However, the possibility of such continuation makes further monitoring of the timing of emigration and eventually the research on the neurophysiological mechanism of interval timer in H. axyridis interesting.
The existence and function of a “long-range” timer of consecutive events in insect life histories is known (Lees, 1966; Saunders, 1976). The interval timer uses information obtained from the environment (usually “daylength”) to synchronize a variety of developmental phenomena in further life to the appropriate season. Interval timers controlling circannual periods of adult molting in Coleoptera (Blake, 1958), diapause induction in Hymenoptera (Reznik and Samartsev, 2015; Reznik and Voinovich, 2021) and the production of morphs in aphids (Brodel and Schaefers, 1979; Campbell and Tregidga, 2006) have been described.
The existence of an interval timer in H. axyridis would be particularly useful for populations in the north cold temperate zones, where adults overwinter mainly in buildings and are dependent on this type of hibernacula. These shelters reduce the information regarding spring weather changes that serve as a signal for emigration. In these circumstances, it is advantageous to have an intrinsic timer of overwintering length that, after completion of a determined period, induces a state of readiness for emigration. The advantage of the interval timer is logical, but its existence requires further evidence.
For most native species of coccinellids, the interval timer of spring emigration from the hibernacula seems less necessary. These species overwinter at sites more exposed to outdoor conditions, such as prominent rock formations (Hodek, 1960), scree slopes and grass tussocks (Honek et al., 2007), leaf litter (Hemptinne, 1988), tree trunks (Parry, 1980), and house facades (Hemptinne, 1985). The interval timer is also probably less necessary for native populations of H. axyridis in East Asia, where adults overwinter on rock formations (Obata, 1986). Moreover, even in the cold temperate zone, a part of the H. axyridis population overwinters outdoors, on conifer branches (Holecova et al., 2018) or in leaf litter.
We conclude that the timing of H. axyridis overwintering is very flexible and allows both the shift of the date of immigration to hibernacula and the date of emigration from the hibernacula. The migration activity follows a relative temperature increase, i.e., an increase against the temperature of the previous period. Despite the variability of immigration and emigration data, the length of hibernation was similar in the seven consecutive overwintering periods investigated in this study. We hypothesize that the date of emigration could be affected by an intrinsic interval timer triggered during the period of immigration. Such a mechanism would be advantageous for non-native populations of H. axyridis in the northern cold temperate zone that overwinter in man-made structures isolated from outdoor conditions. The existence of this interval time requires further experimental verification and research into its neurophysiological mechanism.
Data availability statement
The original contributions presented in this study are included in the article/supplementary material, further inquiries can be directed to the corresponding author.
Author contributions
AH designed the experiments, collected and analyzed the data, and performed the statistical analyses. ZM coordinated all work. All authors wrote and edited the manuscript.
Funding
This work was supported by the program VES19 INTER-COST No. MSMT-15739/2019-6 (MŠMT ČR) and the Ministry of Agriculture of the Czech Republic, institutional support MZE-RO0418.
Acknowledgments
We thank Mrs. Jana Kohoutová, Helena Uhlířová, and Hana Smutná for excellent technical assistance, and Nela Glorikova for digitalization of Figure 1. We also thank the peer reviewers and editors for helpful comments and suggestions to improve this manuscript.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Berthiaume, R., Hebert, C., and Bauce, E. (2003). Impact of temperature and duration of cold exposure on adult survival of the Asian ladybird beetle, Harmonia axyridis (Pallas). Phytoprotection 84, 85–91. doi: 10.7202/007811ar
Blake, G. M. (1958). Diapause and the regulation of development in Anthrenus verbasci (L.) (Col., Dermestidae). Bull. Entomol. Res. 49, 751–775. doi: 10.1017/S0007485300054006
Brodel, C. F., and Schaefers, G. A. (1979). Interval timer for the production of oviparae in Aphis rubicola (Homoptera, Aphididae). Entomol. Exp. Appl. 25, 1–8. doi: 10.1111/j.1570-7458.1979.tb02841.x
Campbell, C. A. M., and Tregidga, E. L. (2006). A transgenerational interval timer inhibits unseasonal sexual morph production in damson hop aphid, Phorodon humuli. Physiol. Entomol. 31, 394–397. doi: 10.1111/j.1365-3032.2006.00531.x
Danilevskii, A. S. (1965). Photoperiodism and seasonal development of insects. London: Edinburgh Oliver and Boyd.
Danks, H. V. (1987). Insect dormancy: An ecological perspective Monograph Series, No 1. Ottawa: Biological Survey of Canada (Terrestrial Arthropods).
Dudek, K., Dudek, M., and Tryjanowski, P. (2015). Wind turbines as overwintering sites attractive to an invasive lady beetle, Harmonia axyridis Pallas (Coleoptera: Coccinellidae). Coleopt. Bull. 69, 665–669. doi: 10.1649/0010-065X-69.4.665
Hemptinne, J. L. (1985). Les sites d’hivernation de la coccinelle Adalia bipunctata (L.) (Col., Coccinellidae) en Belgique. Acta Oecol., Oecol. Appl. 6, 3–13.
Hemptinne, J. L. (1988). Ecological requirements for hibernating Propylea quatuordecimpunctata (L.) and Coccinella septempunctata (Col.: Coccinellidae). Entomophaga 33, 505–515. doi: 10.1007/BF02373186
Hodek, I. (2012). “Diapause/dormancy,” in Ecology and behaviour of the ladybird beetles (Coccinellidae), eds I. Hodek, H. F. van Emden, and A. Honek (Chichester: Wiley-Blackwell), 275–342. doi: 10.1002/9781118223208.ch6
Holecova, M., Zach, P., Holla, K., Sebestova, M., Klesniakova, M., Sestakova, A., et al. (2018). Overwintering of ladybirds (Coleoptera: Coccinellidae) on Scots pine in Central Europe. Eur. J. Entomol. 115, 657–668. doi: 10.14411/eje.2018.065
Honek, A., Brabec, M., Martinkova, Z., Dixon, A. F. G., Pekár, S., and Skuhrovec, J. (2019). Factors determining local and seasonal variation in abundance of Harmonia axyridis (Coleoptera: Coccinellidae) in Central Europe. Eur. J. Entomol. 116, 93–103. doi: 10.14411/eje.2019.011
Honek, A., Brown, P. M. J., Martinkova, Z., Skuhrovec, J., Brabec, M., Burgio, G., et al. (2020). Factors determining variation in colour morph frequencies in invasive Harmonia axyridis populations. Biol. Invasions 22, 2049–2062. doi: 10.1007/s10530-020-02238-0
Honek, A., Martinkova, Z., and Pekar, S. (2007). Aggregation characteristics of three species of Coccinellidae (Coleoptera) at hibernation sites. Eur. J. Entomol. 104, 51–56. doi: 10.14411/eje.2007.008
Honek, A., Martinkova, Z., Dixon, A. F. G., Skuhrovec, J., Roy, H. E., Brabec, M., et al. (2018). Life cycle of Harmonia axyridis (Coleoptera: Coccinellidae) in central Europe. Biocontrol 63, 241–252. doi: 10.1007/s10526-017-9864-2
Iperti, G. (1966). “Voltinism and arrest of development in aphidophagous coccinellids of south-east of France,” in Ecology of Aphidophagous Insects, ed. I. Hodek (Praha: Academia), 105–106.
Labrie, G., Coderre, D., and Lucas, E. (2008). Overwintering strategy of multicolored Asian lady beetle (Coleoptera: Coccinellidae): Cold-free space as a factor of invasive success. Ann. Entomol. Soc. Am. 101, 860–866. doi: 10.1093/aesa/101.5.860
Lees, A. D. (1966). The control of polymorphism in aphids. Adv. Insect Physiol. 3, 207–277. doi: 10.1016/S0065-2806(08)60188-5
Nalepa, C. A. (2007). Harmonia axyridis (Coleoptera: Coccinellidae) in buildings: Relationship between body height and crevice size allowing entry. J. Econ. Entomol. 100, 1633–1636. doi: 10.1093/jee/100.5.1633
Nalepa, C. A., Kennedy, G. G., and Brownie, C. (2005). Role of visual contrast in the alighting behavior of Harmonia axyridis (Coleoptera: Coccinellidae) at overwintering sites. Environ. Entomol. 34, 425–431. doi: 10.1603/0046-225X-34.2.425
Obata, S. (1986). Determination of hibernation site in the ladybird beetle Harmonia axyridis Pallas (Coleoptera: Coccinellidae). Kontyu 54, 218–223.
Parry, W. H. (1980). Overwintering of Aphidecta obliterata (L.) (Coleoptera: Coccinellidae) in North East Scotland. Acta Oecol., Oecol. Appl. 1, 307–316.
Raak-van den Berg, C. L., de Jong, P. W., Hemerik, L., and van Lenteren, J. C. (2013). Diapause and post-diapause quiescence demonstrated in overwintering Harmonia axyridis (Coleoptera: Coccinellidae) in northwestern Europe. Eur. J. Entomol. 110, 585–591. doi: 10.14411/eje.2013.079
Raak-van den Berg, C. L., Hemerik, L., de Jong, P. W., and van Lenteren, J. C. (2012a). Mode of overwintering of invasive Harmonia axyridis in the Netherlands. Biocontrol 57, 71–84. doi: 10.1007/s10526-011-9394-2
Raak-van den Berg, C. L., Stam, J. M., de Jong, P. W., Hemerik, L., and van Lenteren, J. C. (2012b). Winter survival of Harmonia axyridis in the Netherlands. Biol. Control 60, 68–76. doi: 10.1016/j.biocontrol.2011.10.001
Raak-van den Berg, C. L., Hemerik, L., van der Werf, W., de Jong, P. W., and van Lenteren, J. C. (2017). Life history of the harlequin ladybird, Harmonia axyridis: A global meta-analysis. Biocontrol 62, 283–296. doi: 10.1007/s10526-017-9805-0
Reznik, S. Y., and Samartsev, K. G. (2015). Multigenerational maternal inhibition of prepupal diapause in two Trichogramma species (Hymenoptera: Trichogrammatidae). J. Insect Physiol. 81, 14–20. doi: 10.1016/j.jinsphys.2015.06.012
Reznik, S. Y., and Voinovich, N. D. (2021). Influence of temperature on the restoration of the ability of Trichogramma principium to undergo diapause. J. Insect Physiol. 132, 10427. doi: 10.1016/j.jinsphys.2021.104271
Roy, H. E., Brown, P. M. J., Adriaens, T., Berkvens, N., Borges, I., Clusella Trullas, S., et al. (2016). The harlequin ladybird, Harmonia axyridis: Global perspectives on invasion history and ecology. Biol. Invasions 18, 997–1044. doi: 10.1007/s10530-016-1077-6
Sakurai, H., Kumada, Y., and Takeda, S. (1993). Seasonal prevalence and hibernating diapause behaviour in the lady beetle, Harmonia axyridis. Res. Bull. Facul. Agric. Gifu Univ. 1993, 51–55.
Schaefer, P. W. (2003). Winter aggregation of Harmonia axyridis (Coleoptera: Coccinellidae) in a concrete observation tower. Entomol. News 114, 23–28.
Tauber, M. J., Tauber, C. A., and Masaki, S. (1986). Seasonal adaptations of insects. New York, NY: Oxford University Press.
Keywords: artificial hibernaculum, flight, temperature, immigration, emigration
Citation: Honek A, Martinkova Z and Skuhrovec J (2022) Factors influencing hibernation in Harmonia axyridis: Role of the environment and interval timer. Front. Ecol. Evol. 10:994978. doi: 10.3389/fevo.2022.994978
Received: 15 July 2022; Accepted: 02 September 2022;
Published: 27 September 2022.
Edited by:
Eric Wellington Riddick, Agricultural Research Service (USDA), United StatesReviewed by:
Lu Zhoazhi, Qingdao Agricultural University, ChinaJulian Chen, Institute of Plant Protection (CAAS), China
Geetanjali Mishra, University of Lucknow, India
Copyright © 2022 Honek, Martinkova and Skuhrovec. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Jiří Skuhrovec, amlyaXNsYXZza3Vocm92ZWNAZ21haWwuY29t