 Emily DeJaegher
Emily DeJaegher Pedro A. Quijón
Pedro A. Quijón Patricia A. Ramey-Balci
Patricia A. Ramey-Balci- 1Department of Biological Sciences, University of Manitoba, Winnipeg, MB, Canada
- 2Coastal Ecology Laboratory, Department of Biology, University of Prince Edward Island, Charlottetown, PE, Canada
Behaviours such as boldness (the willingness to take risks) vary within and among species and can influence fitness by indirectly affecting resource competition, mortality risk, reproductive success, and dispersal. As such, many studies have investigated boldness in decapod crustaceans, a group of considerable ecological and economic importance. An initial review of these studies suggested outcome inconsistencies that warrant an examination of the approaches used to measure boldness. Boldness is often quantified by measuring behaviours such as latency to emerge from a shelter, exploration of novel environments, or activity following a threat. Hence, we provide an overview of the growth of research and taxonomic representation and analyse the gaps in the methodological approaches for studies examining boldness in decapods over 20 years (2004 – 2024). An examination of 78 studies indicates steady growth that has been narrow in terms of subject taxonomy and methodologies to measure boldness. The outcomes of these studies are often affected by design choices such as the behaviours measured (some widely used, like shelter use, others more controversial, such as exploratory behaviours), the sex, age, condition, and origin of the subjects, and the experimental or rearing conditions (e.g., acclimation times, density, feeding regime, and temperature). Understanding how methodological choices influence decapod boldness is necessary to improve temporal consistency, ensure reproducibility and reliable comparisons among studies, thereby facilitating meta-analyses. Otherwise, inconsistent reporting of design choices may limit the accuracy and feasibility of such meta-analyses, hindering the synthesis of results.
1 Introduction
Communities are largely shaped by complex interactions between organisms, for which behaviour can play a key role (Gilman et al., 2010; Nagelkerken and Munday, 2016). Behavioural tendencies that vary among individuals in a manner that is consistent over time and across contexts are referred to as personality traits (Gosling, 2001; Biro and Stamps, 2008). Several personality traits have been identified, including exploration, aggressiveness, activity, sociability (Gosling, 2001; Rádai et al., 2022), and boldness or the willingness to take risks (Ward et al., 2004). Consistent differences in behaviour can be found not only among individuals but also among related species (Linzmaier et al., 2018). So unsurprisingly, traits like boldness have been widely studied due to their implications for foraging (Maskrey et al., 2018), reproductive success (Gruber et al., 2019), and survival (often in a trade off with foraging or mating benefits) (Belgrad and Griffen, 2016). Boldness has also been linked to agonistic interactions such as aggression, known to be important in both natural and man-made aquaculture settings (Su et al., 2022a, b; Pintor et al., 2008; Sun et al., 2024). In the context of biological invasions, boldness is also relevant since bold behaviour can be associated with dispersal tendencies that might favour invasiveness (Fraser et al., 2001; Damas-Moreira et al., 2019).
Understanding behavioural traits such as boldness is important considering that it may reflect wider ecological and evolutionary divergence among lineages. However, the study of boldness in groups like decapod crustaceans is rather recent (Gherardi et al., 2012). In fact, between 2002 and 2011, boldness was studied in only five crustacean species: European hermit crabs (Pagurus bernhardus), fiddler crabs (Austruca mjobergi and Leptuca pugilator), and crayfish (Astacus astacus and Pacifastacus leniusculus) (Pratt et al., 2005; Pintor et al., 2008; Gherardi et al., 2012). This is surprising considering the importance of a group that includes a variety of keystone species, ecosystem engineers, and invasive species (Pintor et al., 2008; Reisinger and Lodge, 2016; Kabalan et al., 2024). For example, some crab species are keystone or top predators in coastal food webs (Kotta et al., 2018), exerting direct and indirect effects on associated benthic communities (Silliman and Bertness, 2002; Quijón and Snelgrove, 2005a, b; Kotta et al., 2018; Young and Elliott, 2020). Similarly, various invasive crabs alter ecosystems worldwide by out-competing or consuming native species (Epifanio, 2013; Ens et al., 2022) or by disrupting habitats or habitat-forming-species (Bissett et al., 2025).
Despite the existing body of research on boldness, there is a growing number of inconsistent outcomes (Watanabe et al., 2012; Hills and Webster, 2022) that warrant an examination of the methodological approaches being used to study this behavioural trait. We address this by examining published studies on boldness in decapod crustaceans over a relevant period (20 years, 2004 – 2024), focusing on the growth of this research, the subjects (species) used, their characteristics and origin, and the experimental conditions of the behavioural trials. We examined 78 studies, focusing on the caveats that could prevent comparisons among species and studies and impede efforts to draw general conclusions about the influence of boldness on decapod ecology.
2 Search approach
We explored the literature on boldness in decapods by focusing on primary research articles published between January 1, 2004 and November 8, 2024 (20 years). The articles were obtained by searching the Web of Science database, using the following generic search terms: [Topic = (risk-taking OR bold* OR shy*) AND (decapod* OR crab OR lobster OR crayfish OR prawn OR shrimp) AND behaviour*] AND [Publication Date = 2004-01-01 to 2024-11-08]. An initial set of articles included 163 studies, but those that did not directly (explicitly) quantify boldness or risk-taking behaviours (n = 85) were subsequently excluded. A total of 78 articles were therefore retained from this process and thoroughly examined in the review.
3 An overview of boldness in decapod crustaceans
3.1 The growth, taxonomic focus and measurement of boldness in decapods
The number of studies published on boldness in decapods has increased consistently since 2004, reaching 78 articles by November 2024 (Figure 1A). Such growth likely reflects the parallel increase in the number of studies in the broader field of animal personality traits, for which some of the species referred to below have been used as models. To-date, research on boldness in decapods has focused on crayfish (21 articles), Anomuran crabs (24 articles), Brachyuran crabs (26 articles), and shrimp and prawns (7 articles) (Figure 1B), from a variety of freshwater, marine, brackish, and terrestrial habitats (Decker and Griffen, 2012; Reisinger et al., 2020; Zhu et al., 2022; Sakich et al., 2023). Hermit crabs (Anomura in Figure 1B) have accounted for nearly 31% of these studies, with most of these (~22% of total) focusing on one species, the marine European hermit crab (Pagurus bernhardus). Crayfish and Brachyuran crabs were also commonly studied groups, accounting for approximately 27% and 33% of the studies, respectively. Notably, our analysis confirms no boldness studies on lobsters exist as of November 2024, despite the ecological and commercial importance of this diverse group of crustaceans (Boudreau and Worm, 2012). The lack of boldness studies on lobsters likely relates to handling constraints and space feasibility, as large, highly mobile animals require bigger tanks or mesocosms, challenging the measurement of natural behavioural responses (see Polverino et al., 2016).
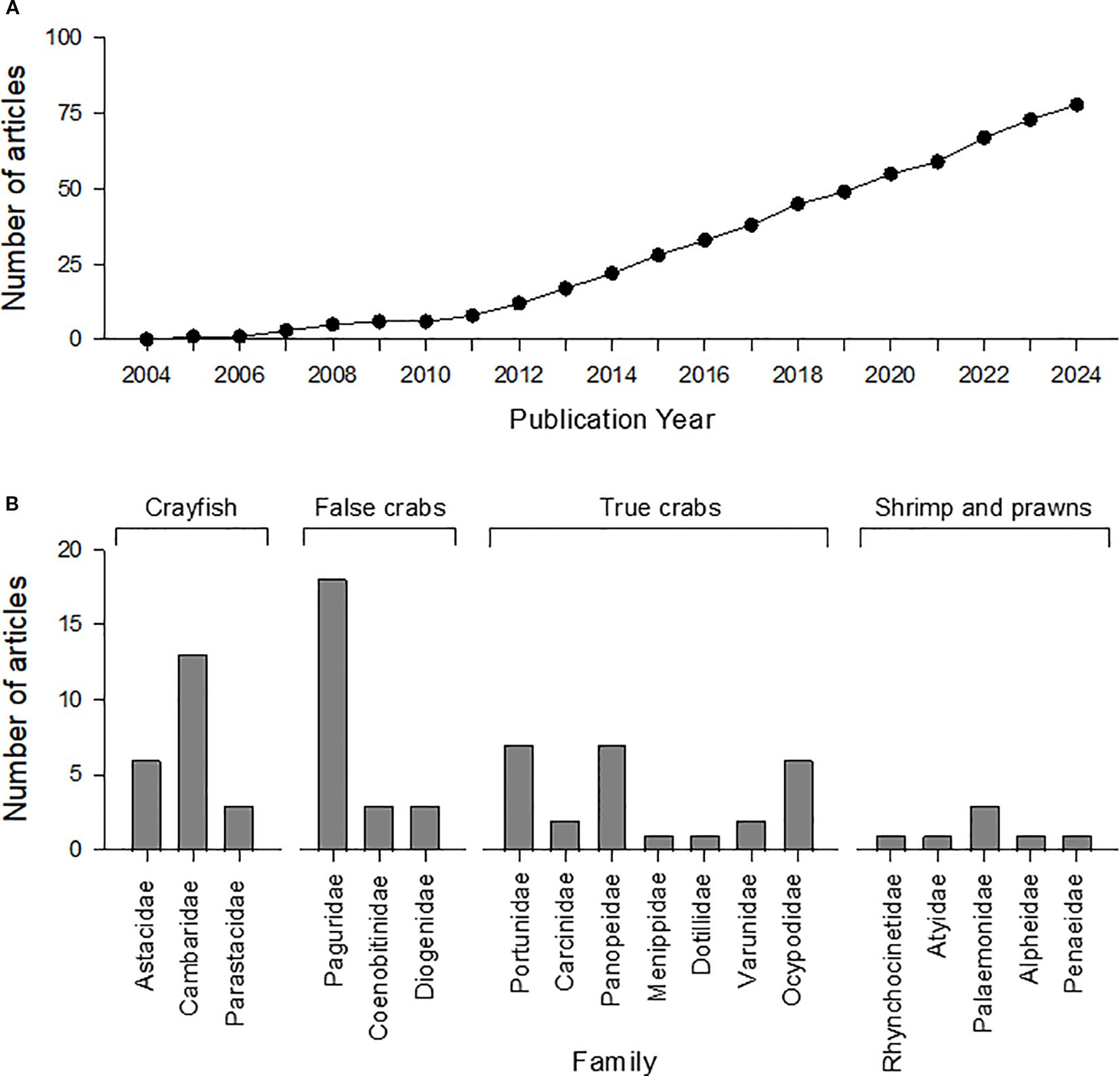
Figure 1. (A) Cumulative number of primary research articles on boldness in decapod species over two decades (January 1, 2004 to November 8, 2024). (B) Allocation of articles on boldness in decapods across families and common groups.
Boldness has most often (~81% of studies) been assessed by measuring shelter use (Tables 1, 2), using the underlying assumption that reduced shelter use is bold behaviour since it entails an increase in mortality risk (Belgrad and Griffen, 2018). Measures like latency to emerge from shelter or the proportion of time spent using shelter, particularly following a threat, are among the most popular. For example, all studies on hermit crabs measured shelter emergence (i.e., startle response duration) to assess boldness (Table 2), since the crabs’ own shell acts as a shelter. Furthermore, minimally invasive shelter use tests can be conducted using existing shelters in the field (e.g., burrows for fiddler crabs; Reaney, 2007). These measures are also simple, repeatable, and applicable on a wide diversity of species. However, the species’ natural behavioural patterns should be considered when selecting a boldness measurement. Measurements of latency to emerge may overestimate boldness in species with less shelter dependence, such as some swimming crab species, or overestimate it in less active species that may sit for longer periods in a shelter, regardless of any direct response to a threat.
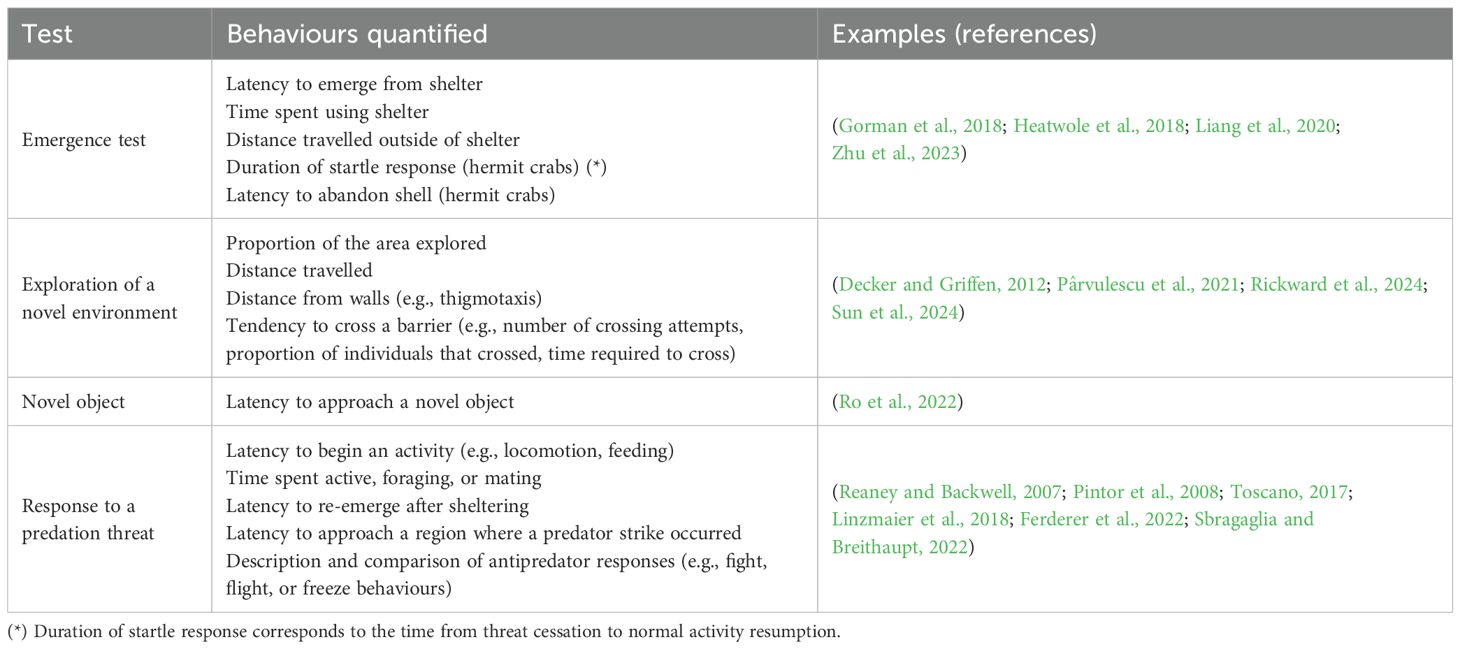
Table 1. Summary of tests used to quantify and compare boldness using decapod species as subjects.
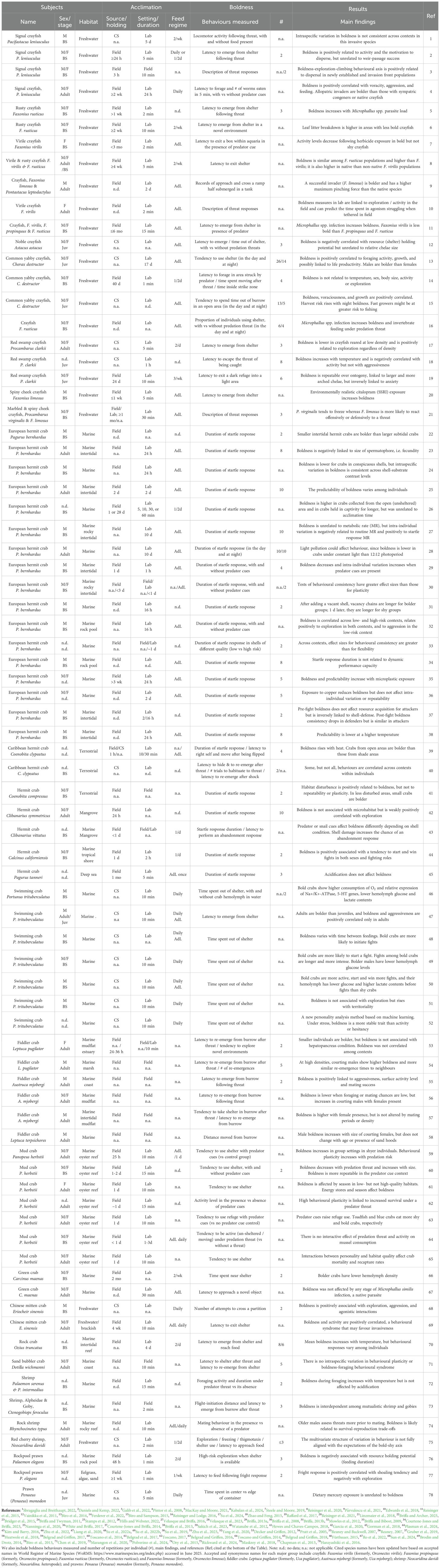
Table 2. Summary of the literature examined, including subjects (common and species names arranged by main groups illustrated in Figure 1, namely crayfish, Anomura and Brachyura, and shrimp and prawns), sex (M: male; F: female) and life stage (Adult; Juv: juvenile; BS: body size if provided instead of life stage), habitat of subject’s origin, and acclimation (pre-trial) conditions, including source of animals (CS: commercial supplier or aquaculture; Field collected), holding time of field-collected subjects, setting and duration of acclimation in experimental set up and pre-experimental feeding regimen.
Even though most studies have relied on shelter tests to evaluate boldness (Table 2), the use of alternative methods seems wise for further research. Circumventing the issue of shelter dependence, boldness has been less frequently quantified using measurements of exploratory tendencies (~8%; e.g., Maskrey et al., 2018), by comparing threat responses (~4% of studies; e.g., claw raising behaviour in crayfish, Galib et al., 2022), or by measuring activity such as foraging, mating, or locomotion, either during or after a threat (e.g., approach by a human or encountering predator cues; ~10% of studies; e.g., Ory et al., 2015; Marangon et al., 2020; Sbragaglia and Breithaupt, 2022). Though exploratory tendencies are well represented in animal personality research (e.g., Galib et al., 2022; Ferderer et al., 2022; Su et al., 2024), they could appear infrequently in our review partly because some of these studies do not explicitly describe it as boldness. Many studies acknowledge an exploration-avoidance axis as a distinct personality trait from a bold-shy axis, describing a subject’s behavioural response to new environments or objects (Réale et al., 2007). However, these novel areas and objects could pose unknown threats, such that exploration is deemed risky and also considered boldness (Réale et al., 2007).
Out of all studies examined, only ~17% used multiple behavioural indices to quantify boldness (Table 2). For example, in studies using male fiddler crabs (L. pugilator), latency to re-emerge from a burrow following a threat (a human waving a towel) was positively correlated with the number of re-emergences after repeated threats before failing to re-emerge (Pratt et al., 2005). Though boldness can be temporally consistent within a given context (Sakich et al., 2023; Su et al., 2024), there is evidence that risk-taking might vary across distinct contexts (Decker and Griffen, 2012). For instance, if exploration in the absence of a threat is considered boldness, then its relationship with threat-related boldness measures can be used to test the context dependency of boldness. To this end, studies with female fiddler crabs (L. pugilator) showed that the latency to re-emerge from a burrow following a threat (i.e., human approach) was unrelated to the tendency to explore a novel environment (Decker and Griffen, 2012). Likewise, relationships between boldness and exploration were not detected in crayfish (Cherax destructor; Ferderer et al., 2022) or swimming crabs (Portunus trituberculatus; Zhu et al., 2023). However, these findings are equivocal given that in the European hermit crab (Mowles et al., 2012) and a crayfish (Procambarus clarkii; Su et al., 2024), exploration and latency to re-emerge from shelter were correlated across individuals.
Overall, these findings suggest the need for multidimensional approaches to evaluate boldness, including considering the context of the behavioural measure (e.g., threatening vs non-threatening contexts). Measuring multiple risk-taking behaviours using standard tests (e.g., emergence latency and response to novel objects) might also be necessary to ensure that what is being measured as boldness is a consistent personality trait that is discernible across multiple contexts. Until further, more compelling, evidence can be gathered, it is reasonable to propose such approach as a standard recommendation: Future studies should aim at assessing boldness across both (threat and non-threat) contexts to ensure reliability and ecological validity. In this regard, researchers must also be mindful that variability among contexts may be influenced by the subject’s ontogeny or energy reserves as discussed in the next section.
3.2 The influence of subjects’ size, stage and condition
Size and life stage are widely known to influence the behaviour of organisms (Toscano et al., 2014; Liang et al., 2020). Nonetheless, nearly 13% of the studies reviewed failed to report the size of the subjects under study, which risks the gathering of inconsistent results among otherwise comparable studies. The subjects’ sizes may influence their risk perception or locomotor performances, indirectly affecting boldness scores. For example, in male swimming crabs, adults exhibited a shorter latency to exit a shelter than juveniles (Liang et al., 2020). Similarly, in mud crabs (Panopeus herbstii), the time spent sheltering was negatively correlated with body size, likely because larger individuals are generally less susceptible to predation (Toscano et al., 2014). These examples suggest that minimum reporting standards are clearly needed, as proposed in more detail below (see Section 4.2). Unlike those studies, larger European hermit crabs were less bold (i.e., displayed longer startle response duration) than smaller individuals. In this case, however, larger individuals occupied a sedimentary subtidal habitat with less refuge compared to the intertidal habitat used by juveniles, that has seaweeds and other complex microhabitats (Briffa and Archer, 2023).
In other species, however, there is opposing evidence that body size is unrelated to boldness measurements. For instance, body size does not affect foraging behaviour following a predator strike in crayfish (C. destructor; Ferderer et al., 2022), startle response duration in the hermit crab Clibanarius symmetricu (Garcia et al., 2020), sheltering behaviour in green crabs (Carcinus maenas) and mitten crabs (Eriocheir sinensis) (Brodin and Drotz, 2014; Fürtbauer, 2015), or the distance that fiddler crabs (Leptuca terpsichores) move away from burrows when mating (Heatwole et al., 2018). Furthermore, the relationship between body size and boldness may vary depending on the specific risk-taking behaviour being analyzed. For example, two measures of boldness in fiddler crabs (L. pugilator), latency to re-emerge and the number of re-emergences following repeated threats, were unrelated to body size (Pratt et al., 2005), even though small females of the same species spent less time sheltering and were more exploratory than larger individuals (Decker and Griffen, 2012). The effect of body size on boldness thus seems to be species, context, and sex- dependent, so studies should always aim to accurately report subject life stage, size range, and sex.
The condition of experimental subjects is also critical to the evaluation of boldness, and aspects like feeding regimes or parasite burden (see below) are directly relevant to condition. While some studies fed subjects ad libitum and omitted a fasting period (Biro and Sampson, 2015; Bridger et al., 2015), others interrupted feeding (often for 24 hours) prior to measuring boldness (Toscano and Griffen, 2014; Belgrad et al., 2017; Liang et al., 2020). Unfortunately, approximately 36% of studies did not clearly report on how they standardized hunger levels prior to trials, either by providing food ad libitum or having a standardized fasting period. In the male swimming crab, measurements of boldness (i.e., proportion of time spent using a shelter) varied with time between feedings (or hunger level; Su et al., 2022a). Compared to crabs fed daily, those fed every 3 days exhibited higher boldness and those fed every 6 days were the least bold (Su et al., 2022a). Furthermore, well-fed Asian shore crabs (Hemigrapsus sanguineus) are known to feed less in bright light compared to dark conditions, likely to avoid predation (Spilmont et al., 2015). However, individuals fasted for 7 days did not avoid light (Spilmont et al., 2015), suggesting that they were bolder. Even though hunger levels might affect boldness, there is still little research addressing the impact of now “standard” fasting periods on the measurement of boldness.
Behavioural studies avoid using individuals with visible parasites (e.g., Bridger et al., 2015) because infected subjects (hosts) can change their behaviour and likely introduce variability in the measurement of boldness (Reisinger et al., 2015; Reisinger and Lodge, 2016). Many decapods, however, have parasites that are not externally visible (Martin et al., 2024). For instance, crayfish species serve as intermediate hosts of trematodes of the genus Microphallus, which encyst in their hepatopancreas (Sargent et al., 2014). The load of Microphallus parasites has been positively correlated with boldness (i.e., latency to emerge from shelter after alarm cue exposure) in the rusty crayfish (Faxonius rusticus) (MacKay and Moore, 2021). Microphallus spp. infections have also prompted the northern clearwater crayfish (Orconectes propinquus) and the virile crayfish (O. virilis) to reduce shelter use by ~40% and 15%, respectively, while increasing shelter affinity in the rusty crayfish by 11% (Reisinger et al., 2015). The same parasites increased the boldness (i.e., reduced latency to emerge from shelter under predation threat) of male crayfish of three species (O. propinquus, O. virilis, and F. rusticus). The only known counterexamples to parasite-induced behavioural change come from the invasive green crab, a species in which the infection by Microphallus similis did not alter the latency to approach a novel object (Ro et al., 2022), nor its time using a shelter or its foraging rates (Blakeslee et al., 2015). These authors suggested that since the green crab has a very limited coevolutionary history with the parasite, this could explain why behaviour was not affected.
The existing research suggests that parasite-driven changes in behaviour (specifically boldness) has the potential to reshape how infected individuals (hosts) and related species interact (Friesen et al., 2020). For example, larger and bolder crayfish infected with trematodes tend to leave sheltered habitats more frequently, a behaviour that may increase exposure to infection (by increasing contact rates; see Lafferty and Shaw, 2013). This may, in turn, increase the efficacy of crayfish as an intermediate host for parasite transmission, with indirect effects on community dynamics. Behavioural interactions likely vary depending on the load or severity of the parasite infection and the ability of the host to manage those parasites. Despite its importance, nearly 74% of the studies compiled in Table 2 did not mention screening for parasites. At a minimum, future studies on behaviour in general − and boldness in particular − should include non-invasive screening for parasites (e.g., Courtene-Jones and Briffa, 2023), especially for taxa known to be at high-risk of infection. When feasible, a post-trial assessment of parasite burden should also be conducted (e.g., Reisinger et al., 2015). The integration of parasite detection protocols into behavioural studies will warrant more accurate results and advance our understanding of the impact of parasites on coexisting species and communities.
3.3 The influence of subjects’ origin, status and holding time
Whether the subjects used in boldness studies are wild-caught or reared in captivity may have an influence on the outcome of boldness measurements, but this has not been thoroughly examined. Linzmaier et al. (2018) studied the response of the marbled crayfish (Procambarus virginalis) to a threat (i.e., the approach of a human hand), and found that wild-caught crayfish were less likely to flee (37%), more likely to fight (15%), and more likely to freeze (47%) than the aquarium-reared crayfish (62, 0, and 37%, respectively) (Linzmaier et al., 2018). This is relevant because differences in threat responses can be used to measure boldness in crayfish (Galib et al., 2022). The underlying causes of these differences may be selective forces that act upon behaviour in wild versus captive populations, due to the lack of natural predation threats in the latter (Huntingford, 2004). Animals bred for generations in captivity can develop differences from wild populations, which may be referred to as conditioning or behavioural plasticity but may also have a genetic basis (see Säisä et al., 2003; Blanchet et al., 2008). Whether the differences are ultimately linked to phenotypic plasticity or potential genetic differences likely depends on the species and rearing conditions. Regardless, these differences introduce an additional source of variation in the subject’s behaviour that may be challenging to interpret but cannot be ignored.
The source population from which wild-caught individuals come from is another key aspect of their origin that could affect boldness, particularly for species with invasive populations (Pintor et al., 2008). This is the case for the signal crayfish (P. leniusculus), in which boldness has been associated with behaviours like raising their claws in response to a threat (Galib et al., 2022). Individuals collected from populations at the frontline of their invasion were more likely to be classified as bold (~72% of individuals, n = 90) than individuals collected from newly established (~53%, n = 90) or fully established populations (~42%, n = 130) (Galib et al., 2022). These differences might be due to a link between boldness and the tendency of these animals to disperse, which is more pronounced in those at the front of an invasion process (Galib et al., 2022). Furthermore, invasive signal crayfish which were geographically apart from the native Shasta crayfish (P. fortis) were found to be bolder (i.e., exhibited shorter latency to forage under a predation threat) than those co-occurring with the Shasta crayfish, and even those found in their original distribution range (Pintor et al., 2008). These authors suggested that environmental differences, such as variations in prey availability, could lead to among-population differences in behavioural phenotypes. The presence of alternate predators, such as invasive or native counterparts, under low prey availability may also be a habitat -mediated selection pressure shaping personality traits (Pintor et al., 2008).
The habitat, or its resource quality, and the microhabitat (those found within a same, larger habitat) from where subjects are collected is another relevant consideration for boldness studies (Hills and Webster, 2022). For example, Sakich et al. (2023) measured boldness in Caribbean hermit crabs (Coenobita clypeatus) collected from either open and sunny microhabitats or shaded microhabitats within the same beaches. Hermit crabs from open microhabitats were bolder, exhibiting shorter startle responses and shorter times to self-flip and move after being placed upside down. Similar results were reported in European hermit crabs, as those collected from open microhabitats were bolder (i.e., shorter startle response durations) than those collected from beneath rocks or seaweeds (Hills and Webster, 2022). If bolder individuals spend less time using sheltered microhabitats, collecting experimental subjects from more visible, easily accessible (open) microhabitats could bias boldness measurements by excluding shyer individuals from the sample population. The quality of a given habitat is also known to affect the abundance of a species, likely causing local density-dependent changes in boldness (Belgrad and Griffen, 2017). Studies should therefore aim to sample individuals haphazardly across the species’ habitat, including multiple microhabitats. Providing detailed descriptions of the methods used to collect subjects, rather than just a location, could also help determine if this is a potential source of subsequent conflicting results between studies.
Indirectly related to the collection of wild-caught subjects, is the amount of time they are held in captivity prior to boldness measurements, an aspect that approximately 15% of the studies examined failed to report (Table 2). This is relevant because this may bias boldness assessments (Hills and Webster, 2022). For example, in wild-caught European hermit crabs, individuals held in captivity for 28 days had a shorter startle response duration than those held for only 1 day (Hills and Webster, 2022). To our surprise, no other study examined had directly addressed the effect of time in captivity on decapod boldness, and so, many studies seemed to assume a lack of any effects. These studies have reported that boldness within a context is consistent over time, implying that any impacts of holding time on behaviour were negligible. For example, the startle response duration in two hermit crabs (Clibanarius symmetricus and P. bernhardus) remained consistent at a given temperature, when tested daily for 10 and 8 days, respectively (Briffa et al., 2013; Garcia et al., 2020; see also Courtene-Jones and Briffa, 2023). Although it remains to be examined, the striking difference between these times and the 28 days lapse used by Hills and Webster (2022), may explain why boldness was not affected.
3.4 The influence of temperature and other experimental conditions
As ectotherms, ambient temperatures have large impacts on the biological processes of decapods and are well-known to influence behaviour (Briffa et al., 2013; Velasque and Briffa, 2016). Temperature is therefore a relevant component of the context in which boldness studies take place, yet approximately 32% of studies failed to report temperature records in some of their experiments. Empirical evidence on the impact of temperature on boldness is variable. Some studies indicate that an increase in temperature leads to an increase in boldness, whereas others have observed no effect (Biro et al., 2013; Marangon et al., 2020; Reisinger et al., 2020). Temperature may also affect intra-specific variation in boldness, as in European hermit crabs, for which the individual’s duration of startle response was more variable at 15°C than at 10°C (Briffa et al., 2013). Since temperature affects the metabolic rate of ectotherms, changes in metabolic processes and energy needs might explain the effects of temperature on boldness as well (Briffa et al., 2013; Marangon et al., 2020). However, Velasque and Briffa (2016) reported that a measure of boldness (startle response duration) was not affected by metabolic rate in European hermit crabs. Future research should focus on disentangling the mechanisms by which temperature affects boldness in decapods, aiming to explain how this behavioural response changes (in direction, strength, or variation) with temperature alterations.
The density of subjects used in behavioural trials (individual versus multiple subjects) may further affect the outcome of boldness studies (Pratt et al., 2005; Belgrad and Griffen, 2017). For instance, in the mud crab (P. herbstii), shy individuals (i.e., those spending more time under a refuge) increased their boldness when conspecifics were present, though the refuge use of bold crabs was consistent regardless of the presence or absence of conspecifics (Belgrad and Griffen, 2017). Similarly, male fiddler crabs (L. pugilator) were also more likely to emerge from their burrows, along with neighbouring males, when population densities were high (Pratt et al., 2005). In these cases, population density seems to be associated with reduced predation risk, which could prompt increased risk-taking in the subjects (Pratt et al., 2005). Higher densities may also increase competition among subjects and prompt shy individuals to act more boldly to compete for limited resources or mates (Reaney, 2007). Regarding the latter (mates), boldness has been shown to increase in males of two species of fiddler crabs, L. pugilator and A. mjobergi, during conditions linked to increased female activity and mating opportunities, though intra-individual variation in boldness in U. pugilator was consistent regardless of these conditions (Pratt et al., 2005; Reaney, 2007).
Density of subjects prior to experiments, such as during rearing, could also influence boldness (Su et al., 2024). For instance, individual crayfish (Procambarus clarkii) reared at high population densities were generally bolder at sexual maturity (i.e., exhibited a shorter latency to emerge from a shelter), than those reared at low density, possibly because boldness could improve an individual’s competitive ability under increased intraspecific competition (Su et al., 2024). A contrasting case regarding the influence of density on boldness was provided by Gruber et al. (2019). These authors reported that population densities of male fiddler crabs (A. mjobergi) did not have an impact on boldness, measured as the tendency to take shelter and latency to re-emerge from burrows following a threat. Future research should focus on the cues used by decapods such as fiddler crabs to assess the presence of counterparts or their density, to attempt to standardize protocols while measuring boldness (Gruber et al., 2019).
As stated above, the core issue prompting our review was the growing number of inconsistent results among boldness studies (see e.g., Watanabe et al., 2012; Hills and Webster, 2022). As a result of the literature examined in sections 3.2 – 3.4 above, we offer an initial model (Figure 2) that attempts to integrate some key methodological variables and their interactions. While not comprehensive (many specific interactions are likely missing), the model illustrates context-dependent outcomes, that explain contrasting results among different subjects. This initial model may help researchers to identify which variables require concurrent controls in their experimental designs, for the species and conditions they choose to use in their trials.
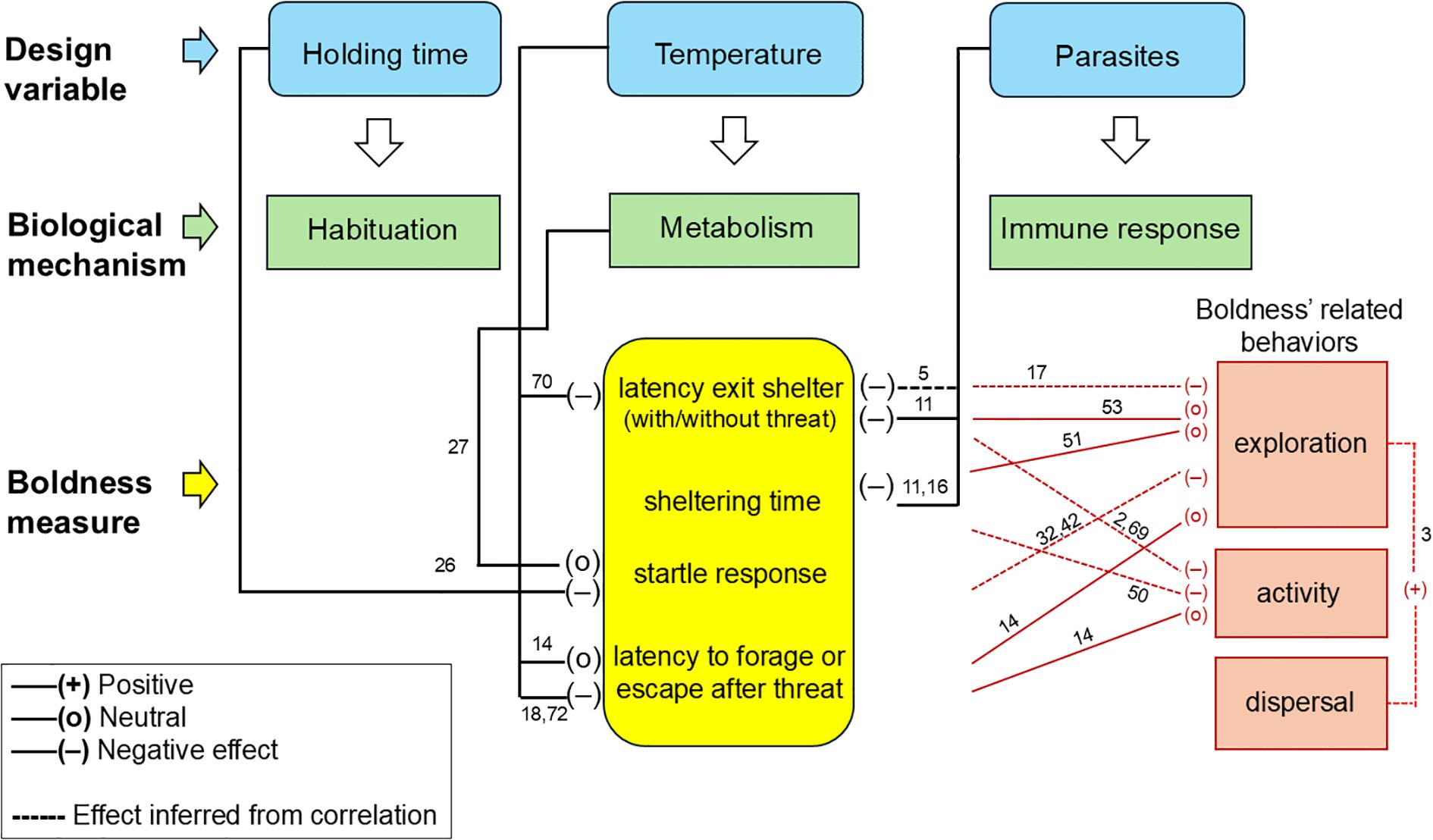
Figure 2. An illustrative model depicting the relationships between some of the main design variables, biological mechanisms, and commonly used boldness measures (two forms of latency, sheltering and startle response). The relationships between boldness measures and other behaviours (in red) are also illustrated. While the model is not comprehensive, it highlights the complexity of different effects (including those inferred from correlations). Numbers along effects refer to the references listed in Table 2.
4 Caveats and takeaways of two decades of boldness studies
4.1 Most common caveats
A better understanding of boldness is critical given the importance of this personality trait on species fitness and ecological interactions (Belgrad and Griffen, 2016; Gruber et al., 2019; Sun et al., 2024). On a more applied context, for farmed species such as the Giant tiger prawn (Penaeus monodon), standardizing feeding protocols could shed light on key growth-cannibalism trade-offs, which are closely associated with boldness. Yet, this and other aspects discussed below were often not clearly reported (see Table 2). While the significance of this personality trait has been increasingly recognized, as evidenced by a steady growth in the number of studies on boldness (Figure 1A), there has not been a parallel increase in the diversification in the subjects (species) under study. As a result, most existing research is still confined to studies focused on a handful of species of e.g., hermit crabs and crayfish (see Table 2 for a compilation of studies). Although the use of a few model species may favor comparability among studies, it also limits our ability to understand boldness more comprehensibly (Voelkl et al., 2020), by exploring a more diverse array of subjects across decapod taxa. For example, commercial species such as lobsters and crabs remain underrepresented, even though understanding boldness in some of these species may provide much insight into the key ecological roles they play in many communities (e.g., Silliman and Bertness, 2002; Kotta et al., 2018; Young and Elliott, 2020).
Beyond the diversity of subjects used, boldness has been measured in a variety of ways. However, an obvious caveat is the heavy focus on measurements of shelter use, and a lack of studies using two or more boldness indices concurrently. It is important to reiterate that different measures of boldness are only in some instances well correlated (Decker and Griffen, 2012; Watanabe et al., 2012), so an increase in the number of studies using multiple boldness measures will shed considerable insight on outcomes that until now appear inconsistent. Likewise, there is little doubt that the subject’s sex, size, stage or condition (including hunger levels) has a strong influence on its (or their) behaviour and therefore on measured boldness levels. Yet, data on these aspects are still missing or not explicitly presented in many of the studies examined (Table 2). For example, 18% of the studies reviewed did not report subjects’ sex, 13% of the studies did not report body size, and 9% of them did not clearly specify the subject life stage (either by stating adult vs juvenile or by reporting body size). The same applies to the subjects’ origin, habitat or microhabitat of origin, or when applicable, the ecological (invasion) status, all of which may influence the outcome of boldness tests (see e.g., Decker and Griffen, 2012; Biro et al., 2013; Hills and Webster, 2022).
Since environmental factors (including rearing conditions) can drive variation in boldness (Sakich et al., 2023), they can also bias our interpretation and possible expectations regarding the interaction of these species with other individuals or species in natural settings. Yet, important details such as the time subjects were held in captivity prior to boldness tests was not explicitly stated in 15% of the studies, and the use of standardized hunger levels was not reported in 36% of the studies (Table 2). These interactions include but are not limited to competition (Liang et al., 2020) and aggressiveness or predation (e.g., Rossong et al., 2011, 2012). Factors like temperature (32% of the studies omitted reporting this in at least one of their sets of experiments) or the presence and number of conspecifics are also important conditions in an experimental setup measuring boldness. Therefore, more research should attempt to assess the influence of these factors before attempting to compare results among studies or among species. Recognizing what causes variation in boldness is critical to further understand the ecological and evolutionary consequences of risk-taking behaviours. Such behaviours, as stated before, dictate some of the main intra- and inter-specific relationships (e.g., Belgrad and Griffen, 2016; Maskrey et al., 2018) that ultimately contribute to coexistence and the structuring of communities. This also holds true for applied settings. For example, in studies quantifying boldness in aquaculture practices, boldness may become relevant for selective breeding, and in studies of invasive decapods, boldness may help to predict local impacts or prepare management measures.
4.2 The takeaway: suggestions for a standardized measurement framework
Based on the existing evidence and the caveats discussed above, a brief set of guidelines can be proposed for future studies. While details on subjects and conditions would likely be species-specific, a general checklist on the reporting of the following parameters would clearly improve the quality of these studies and their comparability. As a minimum, boldness studies should consider: (a) Body size (average and range) and sex ratio of all experimental subjects; (b) An explicit statement of the application (or lack) of a starvation period prior to boldness measurement and its duration; (c) Parasite screening methods or a brief justification as to why this is not being applied; (d) A description of the relevant aspects of the habitats and microhabitat(s) from which the subjects were collected; (e) Acclimation times and the temperatures (or other physical parameters deemed important depending on subject species) used after collection and for experimental trials, and (f) Density in which subjects were raised or maintained before or during the boldness trials.
While generic, these guidelines should improve cross-study comparability, and more importantly, should remove the typical barriers associated with the partial reporting of the important methodological aspects that we have priorly identified as caveats. This may be especially useful if one of our recommendations – to diversify the number of species and types of species used as subjects – is given serious consideration. A broadening on the number of subjects (to include groups such as lobsters, to use an example already discussed), will require added information on the adaptation of methodologies most often used for smaller crustaceans (such as hermit crabs). This may prevent the continued omission of some of the basic methodological parameters identified here. Due to their connections to other fields, research on boldness in crustaceans is expected to continue to drive the interest ecologists and behavioural biologists. An increased rigor on the reporting of the methodological parameters used in boldness studies would only strengthen its growth over the next couple of decades.
Author contributions
ED: Formal analysis, Investigation, Writing – original draft, Data curation, Writing – review & editing. PQ: Writing – review & editing, Data curation, Formal analysis. PR-B: Writing – review & editing, Supervision, Methodology, Formal analysis, Data curation, Investigation, Funding acquisition, Writing – original draft.
Funding
The author(s) declare financial support was received for the research and/or publication of this article. ED was supported by the Faculty of Science, Undergraduate Student Research Award (USRA 2025), University of Manitoba, during the preparation of the manuscript. PR-B thanks the University of Manitoba for start-up funds/seed grant and research study leave. PAQ thanks the support provided by the Natural Sciences and Engineering Research Council, Canada (NSERC), during the preparation and editing of the manuscript.
Acknowledgments
We thank the valuable feedback received by two reviewers on earlier versions of this manuscript.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The author(s) declared that they were an editorial board member of Frontiers, at the time of submission. This had no impact on the peer review process and the final decision.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Belgrad B. A. and Griffen B. D. (2016). Predator–prey interactions mediated by prey personality and predator hunting mode. Proc. R. Soc. B: Biol. Sci. 283, 20160408. doi: 10.1098/rspb.2016.0408
Belgrad B. A. and Griffen B. D. (2017). Habitat quality mediates personality through differences in social context. Oecologia 184, 431–440. doi: 10.1007/s00442-017-3886-4
Belgrad B. A. and Griffen B. D. (2018). Personality interacts with habitat quality to govern individual mortality and dispersal patterns. Ecol. Evol. 8, 7216–7227. doi: 10.1002/ece3.4257
Belgrad B. A., Karan J., and Griffen B. D. (2017). Individual personality associated with interactions between physiological condition and the environment. Anim. Behav. 123, 277–284. doi: 10.1016/j.anbehav.2016.11.008
Biro P. A., Adriaenssens B., and Sampson P. (2014). Individual and sex-specific differences in intrinsic growth rate covary with consistent individual differences in behaviour. J. Anim. Ecol. 83, 1186–1195. doi: 10.1111/1365-2656.12210
Biro P., O’Connor J., Pedini L., and Gribben P. E. (2013). Personality and plasticity: consistent responses within-, but not across-temperature situations in crabs. Behaviour 150, 799–811. doi: 10.1163/1568539X-00003081
Biro P. A. and Sampson P. (2015). Fishing directly selects on growth rate via behaviour: implications of growth-selection that is independent of size. Proc. R. Society. B Biol. Sci. 282, 20142283. doi: 10.1098/rspb.2014.2283
Biro P. A. and Stamps J. A. (2008). Are animal personality traits linked to life-history productivity? Trends Ecol. Evol. (Amsterdam) 23, 361–368. doi: 10.1016/j.tree.2008.04.003
Bissett W. G., Ramey-Balci P. A., and Quijón P. A. (2025). Undermining the foundation: A brief overview of the effects of a widespread invader on coastal ecosystem engineers. Front. Mar. Sci. 12. doi: 10.3389/fmars.2025.1614368
Blakeslee A. M. H., Keogh C. L., Fowler A. E., and Griffen B. D. (2015). Assessing the effects of trematode infection on invasive green crabs in Eastern North America. PloS One 10, e0128674. doi: 10.1371/journal.pone.0128674
Blanchet S., Páez D. J., Bernatchez L., and Dodson J. J. (2008). An integrated comparison of captive-bred and wild Atlantic salmon (Salmo salar): implications for supportive breeding programs. Biol. Conserv. 141, 1989–1999. doi: 10.1016/j.biocon.2008.05.014
Boudreau S. A. and Worm B. (2012). Ecological role of large benthic decapods in marine ecosystems: a review. Mar. Ecol. Prog. Ser. 469, 195–213. doi: 10.3354/meps09862
Bridger D., Bonner S. J., and Briffa M. (2015). Individual quality and personality: Bolder males are less fecund in the hermit crab Pagurus bernhardus. Proc. R. Soc. B: Biol. Sci. 282, 20142492. doi: 10.1098/rspb.2014.2492
Briffa M. (2013a). Plastic proteans: reduced predictability in the face of predation risk in hermit crabs. Biol. Lett. 9, 20130592. doi: 10.1098/rsbl.2013.0592
Briffa M. (2013b). The influence of personality on a group-level process: shy hermit crabs make longer vacancy chains. Ethology 119, 1014–1023. doi: 10.1111/eth.12148
Briffa M. and Archer R. (2023). Size specific boldness associated with differences in resource requirements and habitat use: a cross-sectional study in hermit crabs. Curr. Zoology 69, 360–366. doi: 10.1093/cz/zoac049
Briffa M. and Bibost A. L. (2009). Effects of shell size on behavioural consistency and flexibility in hermit crabs. Can. J. Zoology 87, 597–603. doi: 10.1139/Z09-047
Briffa M., Bridger D., and Biro P. A. (2013). How does temperature affect behaviour? Multilevel analysis of plasticity, personality and predictability in hermit crabs. Anim. Behav. 86, 47–54. doi: 10.1016/j.anbehav.2013.04.009
Briffa M., Rundle S. D., and Fryer A. (2008). Comparing the strength of behavioural plasticity and consistency across situations: animal personalities in the hermit crab Pagurus bernhardus. Proc. R. Soc. B: Biol. Sci. 275, 1305–1311. doi: 10.1098/rspb.2008.0025
Briffa M. and Twyman C. (2011). Do I stand out or blend in? Conspicuousness awareness and consistent behavioural differences in hermit crabs. Biol. Lett. 7, 330–332. doi: 10.1098/rsbl.2010.0761
Brodin T. and Drotz M. K. (2014). Individual variation in dispersal associated behavioral traits of the invasive Chinese mitten crab (Eriocheir sinensis, H. Milne Edwards 1854) during initial invasion of Lake Vänern, Sweden. Curr. Zool. 60, 410–416. doi: 10.1093/czoolo/60.3.410
Chapman B. B., Hegg A., and Ljungberg P. (2013). Sex and the syndrome: individual and population consistency in behaviour in rock pool prawn Palaemon elegans. PloS One 8, e59437. doi: 10.1371/journal.pone.0059437
Chen P. Z., Su T. L., and Lim S. S. L. (2019). To hide or to feed: an evaluation of personality traits in the sand bubbler crab, Dotilla wichmanni, when responding to environmental interference. Behav. Processes 164, 123–132. doi: 10.1016/j.beproc.2019.05.002
Courtene-Jones W. and Briffa M. (2014). Boldness and asymmetric contests: role- and outcome-dependent effects of fighting in hermit crabs. Behav. Ecol. 25, 1073–1082. doi: 10.1093/beheco/aru085
Courtene-Jones W. and Briffa M. (2023). Boldness is not associated with dynamic performance capacity in hermit crabs. Biol. Lett. 19, 20230224. doi: 10.1098/rsbl.2023.0224
Damas-Moreira I., Riley J. L., Harris D. J., and Whiting M. J. (2019). Can behaviour explain invasion success? A comparison between sympatric invasive and native lizards. Anim. Behav. 151, 195–202. doi: 10.1016/j.anbehav.2019.03.008
Daniels J. A. and Kemp P. S. (2022). Personality-dependent passage behaviour of an aquatic invasive species at a barrier to dispersal. Anim. Behav. 192, 63–74. doi: 10.1016/j.anbehav.2022.07.005
Decker R. A. and Griffen B. D. (2012). Correlating context-specific boldness and physiological condition of female sand fiddler crabs (Uca pugilator). J. Ethology 30, 403–412. doi: 10.1007/s10164-012-0338-9
Edwards D. D., Rapin K. E., and Moore P. A. (2018). Linking phenotypic correlations from a diverse set of laboratory tests to field behaviors in the crayfish, Orconectes virilis. Ethology 124, 311–330. doi: 10.1111/eth.12734
Ens N. J., Harvey B., Davies M. M., Thomson H. M., Meyers K. J., Yakimishyn J., et al. (2022). The Green Wave: Reviewing the environmental impacts of the invasive European green crab (Carcinus maenas) and potential management approaches. Environ. Rev. 30, 306–322. doi: 10.1139/er-2021-0059
Epifanio C. E. (2013). Invasion biology of the Asian shore crab Hemigrapsus sanguineus: a review. J. Exp. Mar. Biol. Ecol. 441, 33–49. doi: 10.1016/j.jembe.2013.01.010
Ferderer A., Davis A. R., and Wong M. Y. L. (2022). Temperature and body size influence personality and behavioural syndromes in an invasive crayfish. Anim. Behav. 190, 187–198. doi: 10.1016/j.anbehav.2022.06.009
Fraser D. F., Gilliam J. F., Daley M. J., Le A. N., and Skalski G. T. (2001). Explaining leptokurtic movement distributions: intrapopulation variation in boldness and exploration. Am. Nat. 158, 124–135. doi: 10.1086/321307
Friesen O. C., Goellner S., Poulin R., and Lagrue C. (2020). Parasites shape community structure and dynamics in freshwater crustaceans. Parasitology 147, 182–193. doi: 10.1017/S0031182019001483
Fürtbauer I. (2015). Consistent individual differences in haemolymph density reflect risk propensity in a marine invertebrate. R. Soc. Open Sci. 2, 140482. doi: 10.1098/rsos.140482
Galib S. M., Sun J., Twiss S. D., and Lucas M. C. (2022). Personality, density and habitat drive the dispersal of invasive crayfish. Sci. Rep. 12, 1114. doi: 10.1038/s41598-021-04228-1
Garcia F. A. C., Moura R. R., Ogawa C. Y., Zanette L. R. S., Silva J. R. F., and Rezende C. F. (2020). Never forget where you came from: microhabitat of origin influences boldness and exploration in the hermit crab Clibanarius symmetricus (Diogenidae). J. Exp. Mar. Biol. Ecol. 527, 151365. doi: 10.1016/j.jembe.2020.151365
Gherardi F., Aquiloni L., and Tricarico E. (2012). Behavioral plasticity, behavioral syndromes and animal personality in crustacean decapods: an imperfect map is better than no map. Curr. Zoology 58, 567–579. doi: 10.1093/czoolo/58.4.567
Gilman S. E., Urban M. C., Tewksbury J., Gilchrist G. W., and Holt R. D. (2010). A framework for community interactions under climate change. Trends Ecol. Evol. 25, 325–331. doi: 10.1016/j.tree.2010.03.002
Gorman D., Ragagnin M. N., McCarthy I. D., and Turra A. (2018). Risk-taking and risk-avoiding behaviors by hermit crabs across multiple environmental contexts. J. Exp. Mar. Biol. Ecol. 506, 25–29. doi: 10.1016/j.jembe.2018.05.006
Gosling S. D. (2001). From mice to men: what can we learn about personality from animal research? psychol. Bull. 127, 45–86. doi: 10.1037/0033-2909.127.1.45
Gruber J., Kahn A., and Backwell P. R. Y. (2019). Risks and rewards: balancing costs and benefits of predator avoidance in a fiddler crab. Anim. Behav. 158, 9–13. doi: 10.1016/j.anbehav.2019.09.014
Harayashiki C. A. Y., Reichelt-Brushett A. J., Liu L., and Butcher P. (2016). Behavioural and biochemical alterations in Penaeus monodon post-larvae diet-exposed to inorganic mercury. Chemosphere 164, 241–247. doi: 10.1016/j.chemosphere.2016.08.085
Heatwole S. J., Christy J. H., and Backwell P. R. Y. (2018). Taking a risk: how far will male fiddler crabs go? Behav. Ecol. Sociobiology 72, 82. doi: 10.1007/s00265-018-2500-z
Hewes M. E. and Chaves-Campos J. (2018). Boldness related to size in the hermit crab Coenobita compressus at undisturbed, but not disturbed beach. Ethology 124, 570–578. doi: 10.1111/eth.12766
Hills A. and Webster M. M. (2022). Sampling biases and reproducibility: experimental design decisions affect behavioural responses in hermit crabs. Anim. Behav. 194, 101–110. doi: 10.1016/j.anbehav.2022.09.017
Huntingford F. A. (2004). Implications of domestication and rearing conditions for the behaviour of cultivated fishes. J. Fish Biol. 65, 122–142. doi: 10.1111/j.0022-1112.2004.00562.x
Kabalan B. A., Reisinger A. J., Pintor L. M., Scarasso M. A., and Reisinger L. S. (2024). Intraspecific variation in crayfish behavioral traits affects leaf litter breakdown in streams. Oecologia 205, 515–531. doi: 10.1007/s00442-024-05593-0
Kim T. W. and Barry J. P. (2016). Boldness in a deep sea hermit crab to simulated tactile predator attacks is unaffected by ocean acidification. Ocean Sci. J. 51, 381–386. doi: 10.1007/s12601-016-0034-8
Kotta J., Wernberg T., Jänes H., Kotta I., Nurkse K., Pärnoja M., et al. (2018). Novel crab predator causes marine ecosystem regime shift. Sci. Rep. 8, 1–7. doi: 10.1038/s41598-018-23282-w
Lafferty K. D. and Shaw J. C. (2013). Comparing mechanisms of host manipulation across host and parasite taxa. J. Exp. Biol. 216, 56–66. doi: 10.1242/jeb.073668
Liang Q., Su X., Wang F., Zhu B., and He M. (2020). The developmental plasticity of boldness and aggressiveness in juvenile and adult swimming crab (Portunus trituberculatus). Front. Mar. Sci. 7. doi: 10.3389/fmars.2020.608565
Linzmaier S. M., Goebel L. S., Ruland F., and Jeschke J. M. (2018). Behavioral differences in an over-invasion scenario: marbled vs. spiny-cheek crayfish. Ecosphere 9, e02385. doi: 10.1002/ecs2.2385
MacKay R. N. and Moore P. A. (2021). Parasites differentially impact crayfish personality in different contexts. Behaviour 158, 921–943. doi: 10.1163/1568539X-bja10101
Marangon E., Goldenberg S. U., and Nagelkerken I. (2020). Ocean warming increases availability of crustacean prey via riskier behavior. Behav. Ecol. 31, 287–291. doi: 10.1093/BEHECO/ARZ196
Martin S., Cheslett D., Georgieva S., O’Connor I., Swords F., and O’Dwyer K. (2024). Microparasite screening across four species of decapod crustaceans (Decapoda: Brachyura, Astacidea) in Ireland. J. Crustacean Biol. 44, 29. doi: 10.1093/jcbiol/ruae029
Maskrey D. K., White S. J., Wilson A. J., and Houslay T. M. (2018). Who dares does not always win: risk-averse rockpool prawns are better at controlling a limited food resource. Anim. Behav. 140, 187–197. doi: 10.1016/j.anbehav.2018.04.023
Mowles S. L., Cotton P. A., and Briffa M. (2012). Consistent crustaceans: the identification of stable behavioural syndromes in hermit crabs. Behav. Ecol. Sociobiology 66, 1087–1094. doi: 10.1007/s00265-012-1359-7
Nagelkerken I. and Munday P. L. (2016). Animal behaviour shapes the ecological effects of ocean acidification and warming: moving from individual to community-level responses. Global Change Biol. 22, 974–989. doi: 10.1111/gcb.13167
Nanninga G. B., Horswill C., Lane S. M., Manica A., and Briffa M. (2020). Microplastic exposure Increases predictability of predator avoidance strategies in hermit crabs. J. Hazardous Materials Lett. 1, 100005. doi: 10.1016/j.hazl.2020.100005
Ory N. C., van Son T. C., and Thiel M. (2015). Mating rock shrimp hedge their bets: old males take greater risk, but only after careful assessment of the investment scenario. Behav. Ecol. Sociobiology 69, 1975–1984. doi: 10.1007/s00265-015-2009-7
Pârvulescu L., Stoia D. I., Miok K., Ion M. C., Puha A. E., Sterie M., et al. (2021). Force and boldness: cumulative assets of a successful crayfish invader. Front. Ecol. Evol. 9, 581247. doi: 10.3389/fevo.2021.581247
Pintor L. M., Sih A., and Bauer M. L. (2008). Differences in aggression, activity and boldness between native and introduced populations of an invasive crayfish. Oikos 117, 1629–1636. doi: 10.1111/j.1600-0706.2008.16578.x
Plasman M., Burciaga L. M., and Alcaraz G. (2024). Sex differences in aggression: female hermit crabs initiate few fights against males and lose most of those. Biol. Bull. 245, 139–151. doi: 10.1086/732257
Polverino G., Lehtonen T. K., Geschke A., Callahan T., Urbancic J., and Wong B. B. M. (2024). Size dependent antipredator responses in a fish–shrimp mutualism. Biol. Lett. 20, 20230285. doi: 10.1098/rsbl.2023.0285
Polverino G., Ruberto T., Staaks G., and Mehner T. (2016). Tank size alters mean behaviours and individual rank orders in personality traits of fish depending on their life stage. Anim. Behav. 115, 127–135. doi: 10.1016/j.anbehav.2016.03.013
Pratt A. E., McLain D. K., and Berry A. S. (2005). Variation in the boldness of courting sand fiddler crabs (Uca pugilator). Ethology 111, 63–76. doi: 10.1111/j.1439-0310.2004.01047.x
Quijón P. A. and Snelgrove P. V. R. (2005a). Differential regulatory roles of crustacean predators in a sub-arctic, soft-sediment system. Mar. Ecol. Prog. Ser. 285, 137–149. doi: 10.3354/meps285137
Quijón P. A. and Snelgrove P. V. R. (2005b). Predation regulation of sedimentary faunal structure: potential effects of a fishery-induced switch in predators in a Newfoundland sub-Arctic fjord. Oecologia 144, 125–136. doi: 10.1007/s00442-005-0017-4
Rádai Z., Kiss J., Nagy N. A., Somogyi A.Á., Fülöp A., Tóth Z., et al. (2022). State and physiology behind personality in arthropods: a review. Behav. Ecol. Sociobiology 76, 150. doi: 10.1007/s00265-022-03259-6
Raffard A., Lecerf A., Coet J., Buoro M., Lassus R., and Cucherousset J. (2017). The functional syndrome: Linking individual trait variability to ecosystem functioning. Proc. R. Soc. B: Biol. Sci. 284. doi: 10.1098/rspb.2017.1893
Réale D., Reader S. M., Sol D., McDougall P. T., and Dingemanse N. J. (2007). Integrating animal temperament within ecology and evolution. Biol. Rev. Cambridge Philos. Soc. 82, 291–318. doi: 10.1111/j.1469-185X.2007.00010.x
Reaney L. T. (2007). Foraging and mating opportunities influence refuge use in the fiddler crab, Uca mjobergi. Anim. Behav. 73, 711–716. doi: 10.1016/j.anbehav.2006.05.022
Reaney L. T. and Backwell P. R. Y. (2007). Risk-taking behavior predicts aggression and mating success in a fiddler crab. Behav. Ecol. 18, 521–525. doi: 10.1093/beheco/arm014
Reisinger L. S., Glon M. G., and Pintor L. M. (2020). Divergence in foraging and predator avoidance behavior across the geographic range of native and non-native crayfish. Hydrobiologia 847, 803–818. doi: 10.1007/s10750-019-04139-3
Reisinger L. S. and Lodge D. M. (2016). Parasites alter freshwater communities in mesocosms by modifying invasive crayfish behavior. Ecology 97, 1497–1506. doi: 10.1890/15-1634.1
Reisinger L. S., Petersen I., Hing J. S., Davila R. L., and Lodge D. M. (2015). Infection with a trematode parasite differentially alters competitive interactions and antipredator behaviour in native and invasive crayfish. Freshw. Biol. 60, 1581–1595. doi: 10.1111/fwb.12590
Reisinger A. J., Reisinger L. S., Richmond E. K., and Rosi E. J. (2021). Exposure to a common antidepressant alters crayfish behavior and has potential subsequent ecosystem impacts. Ecosphere 12, e03527. doi: 10.1002/ecs2.3527
Rickward R. A., Santostefano F., and Wilson A. J. (2024). Among-individual behavioural variation in the ornamental red cherry shrimp, Neocaridina heteropoda. Ecol. Evol. 14, e11049. doi: 10.1002/ece3.11049
Ro H., Fowler A. E., Wood C. L., and Blakeslee A. M. H. (2022). Trematode parasites have minimal effect on the behavior of invasive green crabs. Aquat. Invasions 17, 238–258. doi: 10.3391/AI.2022.17.2.07
Rossong M. A., Quijón P. A., Snelgrove P. V. R., Barrett T. J., McKenzie C. H., and Locke A. (2012). Regional differences in foraging behaviour of invasive green crab (Carcinus maenas) populations in Atlantic Canada. Biol. Invasions 14, 659–669. doi: 10.1007/s10530-011-0107-7
Rossong M. A., Quijón P. A., Williams P. J., and Snelgrove P. V. R. (2011). Foraging and shelter behavior of juvenile American lobster (Homarus americanus): the influence of a non-indigenous crab. J. Exp. Mar. Biol. Ecol. 403, 75–80. doi: 10.1016/j.jembe.2011.04.008
Säisä M., Koljonen M., and Tähtinen J. (2003). Genetic changes in Atlantic salmon stocks since historical times and the effective population size of a long-term captive breeding programme. Conserv. Genet. 4, 613–627. doi: 10.1023/A:1025680002296
Sakich N. B., Bartel P. C., Richards M. H., and Tattersall G. J. (2023). Hot crabs with bold choices: temperature has little impact on behavioural repeatability in Caribbean hermit crabs. Behav. Processes 210, 104916. doi: 10.1016/j.beproc.2023.104916
Sargent L. W., Baldridge A. K., Vega-Ross M., Towle K. M., and Lodge D. M. (2014). Trematode parasite alters growth, feeding behavior, and demographic success of invasive rusty crayfish (Orconectes rusticus). Oecologia 175, 947–958. doi: 10.1007/s00442-014-2939-1
Sbragaglia V. and Breithaupt T. (2022). Daily activity rhythms, chronotypes, and risk-taking behavior in the signal crayfish. Curr. Zoology 68, 177–184. doi: 10.1093/cz/zoab023
Silliman B. R. and Bertness M. D. (2002). A trophic cascade regulates salt marsh primary production. Proc. Natl. Acad. Sci. 99, 10500–10505. doi: 10.1073/pnas.162366599
Spilmont N., Gothland M., and Seuront L. (2015). Exogenous control of the feeding activity in the invasive Asian shore crab Hemigrapsus sanguineus (De Haan 1835). Aquat. Invasions 10, 327–332. doi: 10.3391/ai.2015.10.3.07
Stamps J. A., Briffa M., and Biro P. A. (2012). Unpredictable animals: individual differences in intraindividual variability (IIV). Anim. Behav. 83, 1325–1334. doi: 10.1016/j.anbehav.2012.02.017
Steele A. N. and Moore P. A. (2019). Express yourself: Individuals with bold personalities exhibit increased behavioral sensitivity to dynamic herbicide exposure. Ecotoxicology Environ. Saf. 179, 272–281. doi: 10.1016/j.ecoenv.2019.04.069
Su L., Lu L., Si M., Ding J., and Li C. (2024). Effect of population density on personality of crayfish (Procambarus clarkii). Animals 14, 1486. doi: 10.3390/ani14101486
Su X., Sun Y., Liu D., Wang F., Liu J., and Zhu B. (2019). Agonistic behaviour and energy metabolism of bold and shy swimming crabs Portunus trituberculatus. J. Exp. Biol. 222, jeb188706. doi: 10.1242/jeb.188706
Su X., Zhu B., and Wang F. (2022a). Feeding strategy changes boldness and agonistic behaviour in the swimming crab (Portunus trituberculatus). Aquaculture Res. 53, 419–430. doi: 10.1111/are.15583
Su X., Zhu B., Ren Z., and Wang F. (2022b). Differences in agonistic behavior and energy metabolism between male and female swimming crab Portunus trituberculatus based on the analysis of boldness. Animals 12, 2363. doi: 10.3390/ani12182363
Sun J., Zhang D., Hong Y., Weng C., Pang Y., Cheng Y., et al. (2024). The personality traits and interactive behavior of Chinese mitten crab (Eriocheir sinensis). Fishes 9, 408. doi: 10.3390/fishes9100408
Toscano B. J. (2017). Prey behavioural reaction norms: response to threat predicts susceptibility to predation. Anim. Behav. 132, 147–153. doi: 10.1016/j.anbehav.2017.08.014
Toscano B. J., Gatto J., and Griffen B. D. (2014). Effect of predation threat on repeatability of individual crab behavior revealed by mark-recapture. Behav. Ecol. Sociobiology 68, 519–527. doi: 10.1007/s00265-013-1666-7
Toscano B. J. and Griffen B. D. (2014). Trait-mediated functional responses: predator behavioural type mediates prey consumption. J. Anim. Ecol. 83, 1469–1477. doi: 10.1111/1365-2656.12236
Vainikka A., Rantala M. J., Niemelä P., Hirvonen H., and Kortet R. (2011). Boldness as a consistent personality trait in the noble crayfish, Astacus astacus. Acta Ethologica 14, 17–25. doi: 10.1007/s10211-010-0086-1
Velasque M. and Briffa M. (2016). The opposite effects of routine metabolic rate and metabolic rate during startle responses on variation in the predictability of behaviour in hermit crabs. Behaviour 153, 1545–1566. doi: 10.1163/1568539X-00003371
Velasque M., Denton J. A., and Briffa M. (2023). Under the influence of light: how light pollution disrupts personality and metabolism in hermit crabs. Environ. pollut. 316, 120594. doi: 10.1016/j.envpol.2022.120594
Voelkl B., Altman N. S., Forsman A., Forstmeier W., Gurevitch J., Jaric I., et al. (2020). Reproducibility of animal research in light of biological variation. Nat. Rev. Neurosci. 21, 384–393. doi: 10.1038/s41583-020-0313-3
Ward A. J. W., Thomas P., Hart P. J. B., and Krause J. (2004). Correlates of boldness in three-spined sticklebacks (Gasterosteus aculeatus). Behav. Ecol. Sociobiology 55, 561–568. doi: 10.1007/s00265-003-0751-8
Watanabe N. M., Stahlman W. D., Blaisdell A. P., Garlick D., Fast C. D., and Blumstein D. T. (2012). Quantifying personality in the terrestrial hermit crab: different measures, different inferences. Behav. Processes 91, 133–140. doi: 10.1016/j.beproc.2012.06.007
White S. J. and Briffa M. (2017). How do anthropogenic contaminants (ACs) affect behaviour? Multi-level analysis of the effects of copper on boldness in hermit crabs. Oecologia 183, 391–400. doi: 10.1007/s00442-016-3777-0
Yang C., Su X., Liu D., Guo Z., Wang F., and Lu Y. (2020). A new method of aquatic animal personality analysis based on machine learning (PAML): taking swimming crab Portunus trituberculatus as an example. Front. Mar. Sci. 7. doi: 10.3389/fmars.2020.00032
Young A. M. and Elliott J. A. (2020). Life history and population dynamics of green crabs (Carcinus maenas). Fishes 5, 4. doi: 10.3390/fishes5010004
Zhao D. and Feng P. (2015). Temperature increase impacts personality traits in aquatic non-native species: implications for biological invasion under climate change. Curr. Zool. 61, 966–971. doi: 10.1093/czoolo/61.6.966
Zhu B., Su X., Yu W., and Wang F. (2022). What forms, maintains, and changes the boldness of swimming crabs (Portunus trituberculatus)? Animals 12, 1618. doi: 10.3390/ani1213161
Keywords: personality traits, behavioural trial, risk-taking, shelter use, shyness
Citation: DeJaegher E, Quijón PA and Ramey-Balci PA (2025) Measuring boldness in decapod crustaceans: an overview of methodological approaches and potential caveats. Front. Ecol. Evol. 13:1651164. doi: 10.3389/fevo.2025.1651164
Received: 21 June 2025; Accepted: 08 August 2025;
Published: 24 September 2025.
Edited by:
Lucille Chapuis, La Trobe University, AustraliaReviewed by:
Hengtong Liu, Huaian Academy of Nanjing Agricultural University, ChinaNuwandi Pathirana, University of Peradeniya, Sri Lanka
Copyright © 2025 DeJaegher, Quijón and Ramey-Balci. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Patricia A. Ramey-Balci, UGF0cmljaWEuUmFtZXktQmFsY2lAdW1hbml0b2JhLmNh