 Caroline Chauché1,2
Caroline Chauché1,2 Francesco Vacca1,3Shin Li Chia1Josh Richards1William F. Gregory1Adefunke Ogunkanbi4Martin Wear5
Francesco Vacca1,3Shin Li Chia1Josh Richards1William F. Gregory1Adefunke Ogunkanbi4Martin Wear5 Henry J. McSorley1,4*
Henry J. McSorley1,4*- 1Centre for Inflammation Research, University of Edinburgh, Edinburgh, United Kingdom
- 2Wellcome Centre for Integrative Parasitology, Institute of Infection, Immunity and Inflammation, University of Glasgow, Glasgow, United Kingdom
- 3Malaghan Institute of Medical Research, Wellington, New Zealand
- 4Division of Cell Signalling and Immunology, School of Life Sciences, University of Dundee, Dundee, United Kingdom
- 5The Edinburgh Protein Production Facility (EPPF), Wellcome Trust Centre for Cell Biology (WTCCB), University of Edinburgh, Edinburgh, United Kingdom
The murine intestinal nematode Heligmosomoides polygyrus releases the H. polygyrus Alarmin Release Inhibitor (HpARI) - a protein which binds to IL-33 and to DNA, effectively tethering the cytokine in the nucleus of necrotic cells. Previous work showed that a non-natural truncation consisting of the first 2 domains of HpARI (HpARI_CCP1/2) retains binding to both DNA and IL-33, and inhibited IL-33 release in vivo. Here, we show that the affinity of HpARI_CCP1/2 for IL-33 is significantly lower than that of the full-length protein, and that HpARI_CCP1/2 lacks the ability to prevent interaction of IL-33 with its receptor. When HpARI_CCP1/2 was applied in vivo it potently amplified IL-33-dependent immune responses to Alternaria alternata allergen, Nippostrongylus brasiliensis infection and recombinant IL-33 injection, in direct contrast to the IL-33-suppressive effects of full-length HpARI. Mechanistically, we found that HpARI_CCP1/2 is able to bind to and stabilize IL-33, preventing its degradation and maintaining the cytokine in its active form. This study highlights the importance of IL-33 inactivation, the potential for IL-33 stabilization in vivo, and describes a new tool for IL-33 research.
Introduction
Heligmosomoides polygyrus is a parasitic nematode that infects the intestines of mice. It has a fecal/oral lifecycle, with infective L3 larvae being ingested, and then rapidly penetrating the epithelium of the proximal duodenum. There, the larvae develop to L4 stage and emerge as adults into the intestinal lumen at around day 10 of infection (1, 2). The transit of the parasite through the intestinal wall is likely to cause epithelial damage and cell death, resulting in the release of alarmins such as IL-33 from stromal cells or mast cells (3), in turn inducing an anti-parasite type 2 immune response (4). In order to negate this response, and allow persistence of the parasite in the host, H. polygyrus secretes multiple immunomodulatory factors, including Hp-TGM, a protein mimic of host TGF-β (5), and microRNA-containing extracellular vesicles (6) which modulate transcription of multiple host genes, including suppression of Suppression of Tumorigenicity 2 (ST2), the IL-33 receptor. Furthermore, our recent work shows that H. polygyrus secretes HpBARI, a protein which binds and blocks ST2 (7). We previously showed that the parasite also secretes the H. polygyrus Alarmin Release Inhibitor (HpARI), which blocks IL-33 responses (8).
IL-33 is an alarmin cytokine constitutively produced by epithelial cells. It is stored preformed in the nucleus and released on necrotic cell death, due to mechanical, protease-mediated or chemical damage to the epithelium (9). On necrotic cell death, proteases from the cell cytoplasm, or those secreted by recruited mast cells, neutrophils or those in allergens can then cleave the cytokine between the N-terminus chromatin-binding domain and the C-terminus receptor binding domain, potently increasing the activity of the cytokine (10–12). The IL-33 receptor-binding domain contains four free cysteine residues, which upon release from the reducing nuclear environment into the oxidizing extracellular environment rapidly form disulphide bonds, changing the cytokine's conformation, rendering it unable to bind to its receptor and effectively inactivating it (13). Proteases can also further degrade IL-33 to smaller, inactive forms (12). Thus, the active form of IL-33 has only a very short half-life, and by 1 h after release the vast majority of IL-33 is inactive or degraded.
HpARI binds to the active reduced form of IL-33 and to genomic DNA. This dual binding tethers IL-33 within the nucleus of necrotic cells, preventing its release, and inhibiting interaction of IL-33 with ST2. The HpARI protein consists of 3 Complement Control Protein domains (CCP1-3), and our previous data showed that HpARI binds IL-33 through the CCP2 domain, while DNA-binding was mediated by the CCP1 domain (8). Here, we further characterize the functions of the CCP domains of HpARI, finding that CCP3 stabilizes the interaction between HpARI and IL-33, increasing its affinity and being required for blockade of IL-33-ST2 interactions. Furthermore, we show that HpARI_CCP1/2 (the HpARI truncation lacking CCP3) is able to stabilize IL-33, increasing its half-life and amplifying its effects.
Materials and Methods
Protein Expression and Purification
Constructs encoding HpARI, HpARI_CCP1/2 and HpARI_CCP2/3 (all with C-terminus myc and 6-His tags) were cloned into the pSecTAG2A expression vector as previously described (8). Purified plasmids were transfected into Expi293F™ cells, and supernatants collected 5 days later. Expi293F™ cells were maintained, and transfections carried out using the Expi293 Expression System according to manufacturer's instructions (ThermoFisher Scientific). Expressed protein in supernatants were purified over a HisTrap excel column (GE Healthcare) and eluted in 500 mM imidazole. Eluted protein was then dialysed to PBS, and repurified on a HiTRAP chelating HP column (GE Healthcare) charged with 0.1 M NiSO4. Elution was performed using an imidazole gradient and fractions positive for the protein of interest were pooled, dialysed to PBS and filter-sterilized. Protein concentration was measured at A280 nM (Nanodrop, ThermoFisher Scientific), using calculated extinction coefficient.
Surface Plasmon Resonance (SPR)
SPR measurements were performed using a BIAcore T200 instrument (GE Healthcare). Ni2+-nitrilotriacetic acid (NTA) sensor chips, 1-ethyl-3-(3-diaminopropyl) carbodiimide hydrochloride (EDC), N-hydroxysuccinimide (NHS) and ethanolamine (H2N(CH2)2OH) were purchased from GE Healthcare. HpARI, HpARI_CCP1/2 or HpARI_CCP2/3 were immobilized and covalently stabilized on an NTA sensor chip essentially as described (14) with the following modifications: following Ni2+ priming (30 sec injection of 500 μM NiCl2 at 5 μl·min−1), dextran surface carboxylate groups were minimally activated by an injection of 0.2 M EDC; 50 mM NHS at 5 μl·min−1 for 240 sec. Respective proteins (at concentrations between 10 and 400 nM), in 10 mM NaH2PO4, pH 7.5; 150 mM NaCl; 50 μM EDTA; 0.05% surfactant P20, were captured via the hexa-His tag and simultaneously covalently stabilized to 400 RU, by varying the contact time. Immediately following the capture/stabilization a single 15 s injection of 350 mM EDTA and 50 mM Imidazole in 10 mM NaH2PO4, pH 7.5; 150 mM NaCl; 50 mM EDTA; 0.05% surfactant P20, at 30 μl·min−1, was used to remove non-covalently bound protein, followed by a 180 sec injection of 1 M H2N(CH2)2OH, pH 8.5 at 5 μl·min−1. Prior to any experiments, the surface was further conditioned with a 600 s wash with 10 mM NaH2PO4, pH 7.5; 150 mM NaCl; 50 μM EDTA; 0.05% surfactant P20 at 100 μl·min−1.
SPR single-cycle kinetic titration binding experiments were performed at 25°C. Three-fold dilution series of mIL-33 (2.47 nM to 200 nM), were injected over the sensor surface, in 10 mM NaH2PO4, pH 7.5; 150 mM NaCl; 50 μM EDTA; 0.05% surfactant P20, at 30 ml.min−1 for 30 s followed by a final 600 s dissociation phase. The on- (k+) and off-rate (k−) constants and the equilibrium dissociation constants were calculated from the double referenced sensorgrams by global fitting of a 1:1 binding model, with mass transport considerations, using analysis software (v2.02) provided with the Biacore T200 instrument.
Immunoprecipitation
Protein G dynabeads (ThermoFisher Scientific) were coated with 1 μg mouse ST2-Fc (Biolegend), and washed on a DynaMag-2 magnet with PBS 0.02% Tween 20. 100 ng recombinant murine IL-33 (Biolegend) was then mixed with 1 μg HpARI, HpARI_CCP1/2 or HpARI_CCP2/3, and incubated at room temperature for 15 min, prior to adding to ST2-Fc-coated protein G dynabeads. Beads were washed and bound IL-33 eluted with 50 mM glycine pH2.8, then ran on 4–12% SDS-PAGE gels (ThermoFisher Scientific) under reducing conditions, and transferred to nitrocellulose membranes for western blotting, probing with anti-IL-33 goat polyclonal antibody (R&D Systems AF3626), rabbit anti-goat IgG-HRP secondary antibody (ThermoFisher Scientific) and detected using WesternSure Premium reagent (Licor). Densitometry was carried out using ImageJ, and expressed as fold change from controls at each timepoint.
Animals
BALB/cAnNCrl and C57BL/6JCrl mice were purchased from Charles River, UK. Heterozygous IL-13eGFP+/GFP mice (15) were bred in-house. All mice were accommodated and procedures performed under UK Home Office licenses with institutional oversight performed by qualified veterinarians.
Alternaria Models
Alternaria alternata allergen was used in vivo as previously described (8, 16). Alternaria allergen (10 μg), OVA (20 μg), HpARI (10 μg) and HpARI_CCP1/2 (10 μg) were intranasally administered to BALB/c mice. Where indicated, the OVA-specific response was recalled by daily intranasal administration of 20 μg OVA protein on days 14, 15, and 16. Tissues were harvested 24 h or 17 days after initial Alternaria allergen administration. Lungs were flushed with 4 washes of 0.5 ml ice-cold PBS to collect bronchoalveolar lavage cells, followed by lung dissection for single cell preparation.
Nippostrongylus brasiliensis Infection
The life cycle of N. brasiliensis was maintained in Sprague-Dawley rats as previously described (17), and infective L3 larvae were prepared from 1 to 3 week rat fecal cultures. C57BL/6 mice were subcutaneously infected with 400 L3 N. brasiliensis larvae, and culled 3 or 6 days later.
Intraperitoneal IL-33 Treatment
Recombinant murine IL-33 (Biolegend) was injected intraperitoneally to C57BL/6 mice (100 ng/mouse). Mice were culled 3 h later and peritoneal lavage cells collected in 3 washes of 3 ml ice-cold RPMI.
Flow Cytometry
Cells were stained with Fixable Blue Live/Dead stain (ThermoFisher Scientific), then blocked with anti-mouse CD16/32 antibody and surface stained with CD3 (FITC, clone 145-2C11), CD5 (FITC, clone 53-7.3), CD11b (FITC, M1/70), CD19 (FITC, clone 6D5), GR1 (FITC, clone RB6-8C5), CD45 (AF700, clone 30-F11), ICOS (PCP, clone 15F9), CD4 (PE-Dazzle, cloneRM4.5), CD11c (AF647, clone N418), Ly6G (PerCP, clone 1A8), CD25 (BV650, clone PC61) (Biolegend); CD49b (FITC, clone DX5), ST2 (APC, clone RMST2-2) (ThermoFisher Scientific); Siglec-F (PE, clone ES22-10D8) (Miltenyi). The lineage stain consisted of CD3, CD5, CD11b, CD19, CD49b and GR1, all on FITC. Samples were acquired on an LSR Fortessa (BD Biosciences) and analyzed using FlowJo 10 (Treestar).
CMT-64 Cell Line
CMT-64 cells (ECACC 10032301) were maintained by serial passage in “complete” RPMI [RPMI 1640 medium containing 10% fetal bovine serum, 2 mM L-glutamine, 100 U/ml Penicillin and 100 μg/ml Streptomycin (ThermoFisher Scientific)] at 37°C, 5% CO2. Cells were seeded into 24- or 96-well plates for Triton-X100 or freeze-thaw treatment, respectively. Cells were grown to 100% confluency prior to 2 washes with PBS. For Triton-X100 treatment, cells were then washed into RPMI 1640 containing 0.1% BSA with or without 0.1% Triton-X100, and incubated at 37°C as indicated, prior to collection of supernatants and measurement of IL-33 by ELISA and western blot. For freeze-thaw assays, cells were then washed into complete RPMI containing 10 μg/ml of HpARI or HpARI_CCP1/2, frozen on dry ice for at least 1 h, then thawed and incubated at 37°C as indicated, prior to collection of supernatants and application to bone marrow cell cultures.
Bone Marrow Cell Culture
Single cell suspensions of bone marrow cells were prepared from C57BL/6 mice, by flushing tibias and femurs with RPMI 1640 medium using a 21 g needle. Cells were resuspended in red blood cell lysis buffer (Sigma) for 5 min at room temperature, prior to resuspension in medium and passing through a 70 μm cell strainer. Cells were cultured in round-bottom 96-well-plates in a final 200 μl volume, containing 0.5 × 106 cells/well. IL-2 and IL-7 were added at 10 ng/ml final concentration, with 50 μl of CMT-64 freeze-thaw supernatant. Cells were then cultured at 37°C, 5% CO2, for 5 days, prior to assessment of responses by cytokine ELISA and flow cytometry.
Cytokine Measurement
ELISAs were carried out to manufacturer's instructions for IL-5, IL-13 (Ready-SET-go, ThermoFisher Scientific) and IL-33 (Duoset, Biotechne). IL-33 was also measured in CMT-64 supernatants by western blot – supernatants were ran on 4–12% NuPAGE gels (ThermoFisher Scientific) under reducing conditions, before transferring to nitrocellulose membrane and probing with goat anti-mIL-33 (Biotechne), and rabbit anti-goat IgG HRP secondary antibody (Thermo Fisher), and detected using WesternSure Premium reagent (Licor).
Statistical Analysis
All data was analyzed using Prism (Graphpad Software Inc.). One-way ANOVA with Dunnet's multiple comparisons post-test was used to compare multiple independent groups, while two-way ANOVA and Tukey's multiple comparison's post-test was used to compare multiple timepoints or concentrations between independent groups. Where necessary, data was log-transformed to give a normal distribution and to equalize variances. ****p < 0.0001, ***p < 0.001, **p < 0.01, *p < 0.05, N.S. = Not significant (p > 0.05).
Results
HpARI CCP2 Binds IL-33, While HpARI CCP3 Is Required to Block IL-33-ST2 Interaction
Constructs encoding full-length HpARI, or truncations lacking CCP3 (HpARI_CCP1/2), or lacking CCP1 (HpARI_CCP2/3) were expressed in Expi293FTM mammalian cells, and purified on 6-His tags. These constructs were then tested for binding to IL-33 in surface plasmon resonance experiments, showing that the affinity for IL-33 of full-length HpARI and HpARI_CCP2/3 were similar (Kd of 1.1 +/− 0.44 nM and 1.4 +/− 0.14 nM, respectively), while HpARI_CCP1/2 had approximately a 10-fold lower affinity for the cytokine (Kd = 9.8 +/−6.7 nM). This difference in affinity was largely due to an approximately 20-fold faster off-rate for HpARI_CCP1/2 (K− of 30 × 10−4 s−1 vs. 1.5 × 10−4 s−1 for HpARI) (Figure 1A).
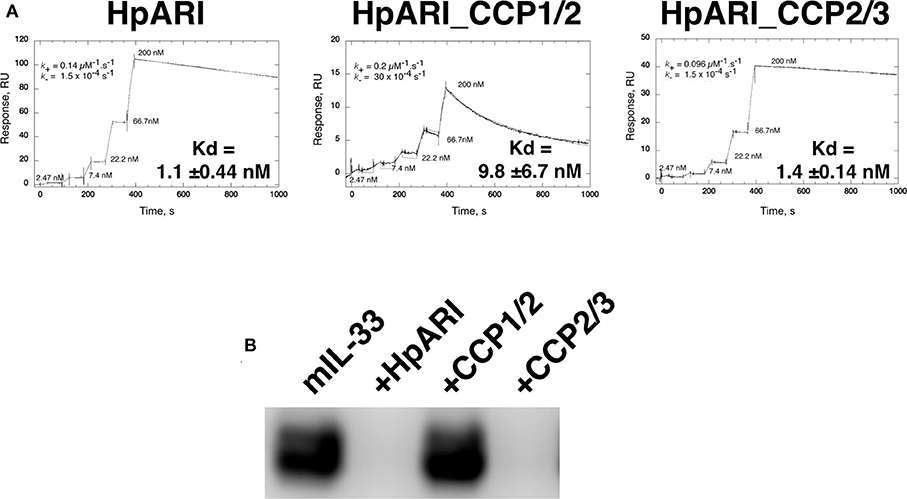
Figure 1. (A) Surface plasmon resonance measurements of IL-33 binding to chip-bound HpARI, HpARI_CCP1/2 and HpARI_CCP2/3. Kd values calculated from 3 replicate experiments, and indicates mean and SD. (B) ST2-Fc was bound to protein G-coated magnetic beads and used to immunoprecipitate murine IL-33 (mIL-33). IL-33 western blot of eluted material shown. Image representative of two independent experiments.
The CCP3 domain also appears important for preventing IL-33-ST2 interactions. While full-length HpARI and HpARI_CCP2/3 were able to prevent IL-33 immunoprecipitation by ST2-Fc, HpARI_CCP1/2 could not (Figure 1B).
HpARI_CCP1/2 Increases Responses to IL-33
We previously showed that HpARI_CCP1/2 was capable of suppressing the release of IL-33 in vivo, 15 min after Alternaria alternata administration (8). To assess whether HpARI_CCP1/2 could replicate the inhibition of IL-33-dependent responses seen with full-length HpARI, we administered HpARI or HpARI_CCP1/2 together with Alternaria allergen and OVA protein and assessed type 2 immune responses after OVA challenge 2 weeks later (Figure 2A). While HpARI suppressed allergic reactivity in this model (as shown previously (8)), HpARI_CCP1/2 had the opposite effect, increasing BAL and lung eosinophil, and lung ILC2 and ICOS+ST2+ Th2 cell numbers (18) (Figure 2A and Supplementary Figure 1).
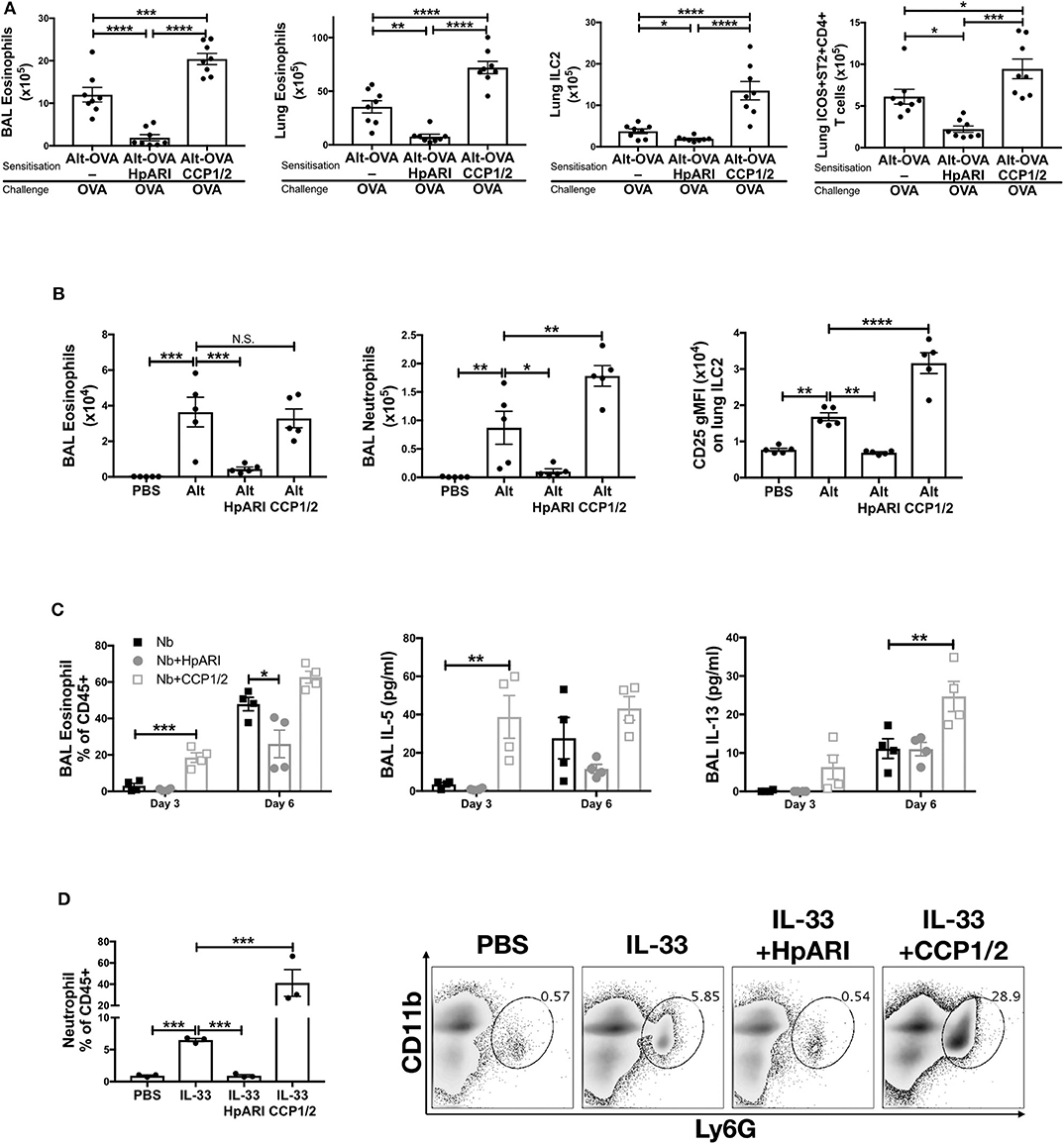
Figure 2. (A) HpARI or HpARI_CCP1/2 (CCP1/2) were co-administered with Alternaria allergen and OVA by the intranasal route, then the OVA-specific response recalled 2 weeks later. BAL and lung eosinophil (Siglecf+CD11c–CD45+), and lung ILC2 (ICOS+lineage–CD45+) and Th2 (ICOS+ST2+CD4+lineage+CD45+) cell numbers shown. Data pooled from 2 repeat experiments each containing 4 mice per group. (B) HpARI_CCP1/2 (CCP1/2) was coadministered with Alternaria allergen by the intranasal route. After 24 h, BAL eosinophil (Siglecf+CD11c–CD45+) and neutrophil (Ly6G+CD11b+Siglecf–CD11c–CD45+) cell numbers, and lung ILC2 CD25 geometric mean fluorescent intensity were assessed by flow cytometry. Data representative of 2 repeat experiments each containing 3–5 mice per group. (C) HpARI or HpARI_CCP1/2 were intranasally administered on days 0, 1, and 2 after infection with Nippostrongylus brasiliensis. BAL eosinophil (Siglecf+CD11c–)% of CD45+ cells, and BAL IL-5 and IL-13 were measured on days 3 and 6 post-infection. Data representative of 3 repeat experiments, each with 4 mice per group. (D) Recombinant IL-33 was intraperitoneally injected with HpARI or HpARI_CCP1/2, and proportions of Ly6G+CD11b+ neutrophils in the CD45+ peritoneal lavage population assessed 3 h post-injection. Representative FACS plots shown of CD45+ live cells. Data representative of 2 repeat experiments, each with 3 mice per group. Error bars show SEM. N.S. = Not significant, *p < 0.05, **p < 0.01, ***p < 0.001, ****p < 0.0001.
When the innate Alternaria-induced immune response was assessed 24 h after initial administration of the allergen to naïve mice, we found that although HpARI_CCP1/2 did not change the eosinophil response compared to Alternaria alone, HpARI_CCP1/2 increased BAL neutrophil numbers. At this timepoint, no ILC2 proliferation has yet occurred, as previously described (19), so total lung ILC2 cell numbers were similar in all groups (data not shown). However, allergen-activated ILC2s showed strong upregulation of CD25 expression, as described previously during activation of ILC2s in this model (20), which was further increased by HpARI_CCP1/2 (Figure 2B).
To exclude the possibility that HpARI_CCP1/2 is interfering with the Alternaria allergen directly, exacerbating the response to it, we used a second model of IL-33-dependent responses (21–23), infecting mice with Nippostrongylus brasiliensis and administering HpARI or HpARI_CCP1/2 to the lungs during the first 3 days of infection. During N. brasiliensis infection, L3 larvae migrate through the lung at days 1–4, enter the intestines as L4 larvae and develop to adults at days 4–10 post-infection (21). Mice were culled at days 3 and 6 post-infection, when parasites were present in the lung and gut, respectively, and the type 2 immune response in the lung was assessed at both timepoints. Again, HpARI suppressed type 2 immune responses as shown previously (8), while HpARI_CCP1/2 increased BAL eosinophilia, IL-5 and IL-13 production (Figure 2C). Neither HpARI nor HpARI_CCP1/2 had any effect on BAL neutrophilia at these timepoints (data not shown), implying that neutrophil recruitment in N. brasiliensis is not IL-33 dependent. Similarly, in Strongyloides venezuelensis lung-stage infection, neutrophil recruitment is IL-33-independent (24).
Finally, we utilized a model of recombinant IL-33 intraperitoneal injection, which induces a mast cell-dependent neutrophilia (25, 26), in contrast to the ILC2-dependent, largely eosinophilic response seen on IL-33 release in the lung. Again, here we found that while HpARI suppressed IL-33 induced neutrophilia, HpARI_CCP1/2 exacerbated it (Figure 2D).
In conclusion, HpARI_CCP1/2 amplifies IL-33-dependent responses in vivo. We hypothesized that this activity was due to stabilization of the cytokine, increasing its effective half-life. To test this hypothesis, we developed an in vitro model of IL-33 release and IL-33 responses.
HpARI_CCP1/2 Maintains IL-33 in Its Active Form
The CMT-64 cell line constitutively produces IL-33, which is released on cellular necrosis (12). Confluent CMT-64 cells were washed into PBS+0.1% BSA, and necrosis induced by addition of 0.1% Triton-X100, in the presence or absence of HpARI or HpARI_CCP1/2. Over a 24 h timecourse following Triton-X100 addition, we assessed IL-33 release by ELISA and western blot. IL-33 ELISA showed that Triton-X100 caused rapid IL-33 release, with high concentrations of the cytokine detected in culture supernatants within 15 min of addition of the detergent in control wells. IL-33 levels then gradually decreased at later timepoints, presumably as the protein was degraded (Figure 3A) (12). HpARI addition ablated the IL-33 signal seen in the ELISA, as shown in our previous study (8): as well as retarding the release of the cytokine, HpARI binding also out-competes the ELISA antibodies, abolishing detection of IL-33. HpARI_CCP1/2 did not abolish detection of IL-33 in the ELISA, but did reduce the IL-33 signal at early timepoints. Moreover, in the presence of HpARI_CCP1/2, IL-33 accumulated over the timecourse and maintained high levels at later timepoints.
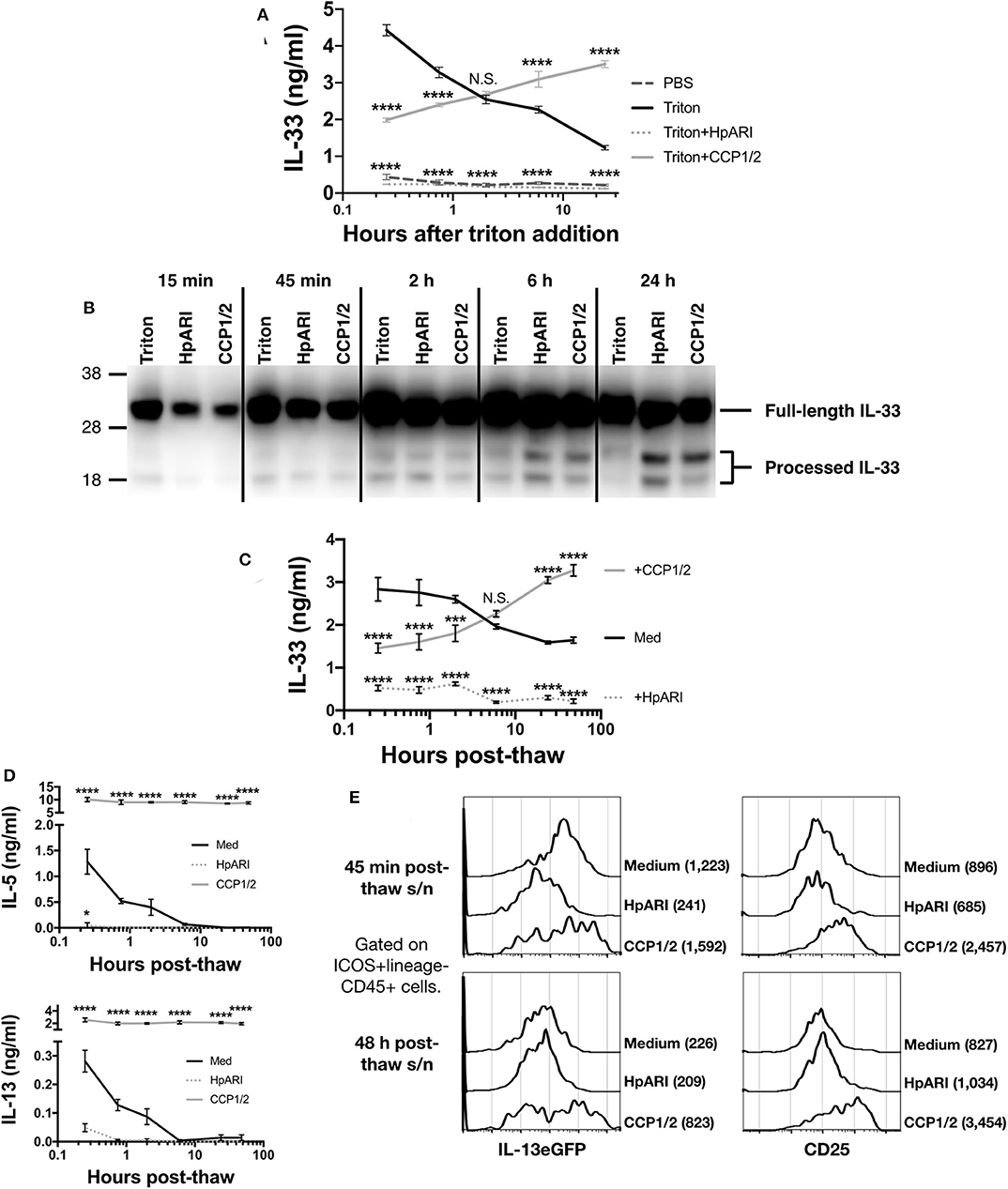
Figure 3. (A) CMT-64 cells were cultured to confluency and treated with 0.1% Triton-X100+0.1% BSA alone, or in the presence of HpARI or HpARI_CCP1/2 (CCP1/2). Supernatants were harvested over a timecourse and IL-33 levels assessed by ELISA. Each measurement contains 4 technical replicates and is representative of 3 repeat experiments. (B) IL-33 western blot of pooled samples from (A). Representative of 3 repeat experiments. (C) CMT-64 cells were cultured to confluency in RPMI+10% FCS, and freeze-thawed in the presence of complete medium (Med), HpARI or HpARI_CCP1/2. After thaw, cultures of necrotic cells were incubated at 37°C, and supernatants taken over a timecourse, and assessed for IL-33 levels by ELISA. Each timepoint shows 4 technical replicates. (D) Supernatants from (C) were applied to IL-13eGFP+/GFP bone marrow cells in the presence of IL-2 and IL-7 and cultured for 5 days. Levels of IL-5 (upper panel) and IL-13 (lower panel) in supernatants were assessed by ELISA. Each timepoint shows 4 technical replicates. (E) Bone marrow cells from (D) after 5 days of culture were pooled, stained, and gated on ICOS+lineage–CD45+ ILC2s, and assessed for IL-13eGFP and CD25 expression. Numbers in parentheses indicate geometric mean fluorescent intensity for each condition. All data from (C–E) is representative of 3 repeat experiments. Error bars show SEM. N.S. = Not significant, *p < 0.05, **p < 0.01, ***p < 0.001, ****p < 0.0001.
In contrast, when IL-33 in the same samples was assessed by western blot, a very strong signal was seen at all timepoints at a size consistent with full-length IL-33 protein (~30 kDa), while a weaker signal was seen at around 18–20 kDa, consistent with processed mature IL-33 (Figure 3B and Supplementary Figures 2A,B). While a strong full-length IL-33 band was seen across all timepoints and treatments, the density of the mature bands were dynamically altered by the presence of each treatment. In control wells, mature IL-33 was present early after Triton-X100 treatment and was degraded at later timepoints. In contrast, in the presence of HpARI_CCP1/2, the mature form was present at lower intensities than in control wells at early timepoints, but accumulated over the timecourse and was strongest at 24 h post Triton-X100 treatment, reflecting ELISA data (Figure 3A). HpARI treatment had a similar effect to HpARI_CCP1/2 when IL-33 was assessed by western blot. Quantification of band intensities by densitometry reflected this increase of mature IL-33 signal in the presence of HpARI or HpARI_CCP1/2 (Supplementary Figure 2C). The difference in IL-33 signal strength between ELISA and western blot in the presence of HpARI was seen in a previous study (8), and is thought to be due to interference with antibody binding to the endogenous IL-33-HpARI complex in ELISA, but in a denaturing western blot proteins from this complex are dissociated and available for antibody detection. Together, this data suggests that binding of IL-33 by HpARI or HpARI_CCP1/2 stabilizes the mature cytokine, protecting it from degradation.
To assess the activity of the cytokine released, we induced necrosis of CMT-64 cells via freeze-thaw treatment. This treatment could be carried out in complete culture medium (without toxic additives such as Triton-X100), allowing downstream assessment of cellular responses to the released cytokine. On thaw, necrotic CMT-64 cells were incubated for up to 48 h at 37°C, and IL-33 levels in supernatants assessed by ELISA. Similarly to Triton-X100-mediated necrosis, we found high levels of IL-33 released rapidly after freeze-thaw necrosis, which gradually decreased over the 48 h timecourse in control wells, while IL-33 levels increased over the timecourse in the presence of HpARI_CCP1/2 (Figure 3C). These supernatants were applied to total bone marrow cells from IL-13eGFP+/GFP reporter mice (15) cultured in the presence of IL-2 and IL-7 (to support ILC2 differentiation), and cytokine responses were assessed 5 days later. As shown in Figure 3D, control freeze-thaw CMT-64 supernatants could only induce bone marrow cell IL-5 and IL-13 production at early timepoints post-thaw, implying that after ~6 h post-thaw, all IL-33 present in the culture medium was inactive. This response appeared IL-33-dependent as HpARI entirely inhibited IL-5 and IL-13 release. In contrast, supernatants from cells freeze-thawed in the presence of HpARI_CCP1/2 were able to maintain high levels of IL-5 and IL-13 stimulation (~10-fold higher than the peak production seen in control wells) and this stimulation was maintained even when supernatants had been incubated for 48 h post-thaw. To specifically assess the ILC2 response within these total bone marrow cell cultures, we used flow cytometry for IL-13eGFP reporter or CD25 expression on ICOS+lineage−CD45+ ILC2s to confirm that these cells were activated by supernatants from medium of freeze-thaw control wells at early (45 min post-thaw), but not late (48 h post-thaw) timepoints, while wells containing HpARI_CCP1/2 remained highly activated throughout the timecourse (Figure 3E and Supplementary Figure 3).
Discussion
HpARI blocks IL-33 responses and is secreted by H. polygyrus, as part of a suite of immunomodulatory effector molecules which act to prevent immune-mediated ejection of the parasite (27). HpARI acts by binding to IL-33 through the HpARI CCP2 domain and to genomic DNA in necrotic cells through the HpARI CCP1 domain, tethering the cytokine within the necrotic cell nucleus and preventing its release (8). Here, we further characterize these interactions, showing that a synthetic, non-natural construct lacking the CCP3 domain (HpARI_CCP1/2) binds IL-33 with an approximately 10-fold lower affinity than the full-length HpARI protein, and lacks the blocking activity of HpARI against IL-33-ST2 interactions. Furthermore, HpARI_CCP1/2 had the surprising effect of stabilizing and amplifying IL-33 responses in vitro and in vivo.
As opposed to HpARI_CCP1/2, HpARI_CCP2/3 showed high affinity binding to IL-33, and prevented ligation of ST2 by IL-33, replicating the IL-33-blocking effects of full-length HpARI. In a previous study (8), we showed that HpARI_CCP2/3 lacked the DNA-binding activity of full-length HpARI and HpARI_CCP1/2, implying that this activity is mediated by the CCP1 domain. We previously also showed that HpARI_CCP2/3 increased, rather than decreased IL-33 levels in the bronchoalveolar lavage of mice 15 min after Alternaria allergen treatment. Our work here supports the hypothesis that this increase in IL-33 is due to HpARI_CCP2/3 preventing the rapid uptake and degradation of bound IL-33 by ST2-expressing immune cells (13, 28, 29), while lacking the DNA-binding (and hence tethering function) of HpARI or HpARI_CCP1/2. Thus, all IL-33 released is retained in the bronchoalveolar lavage, leading to increased IL-33 levels compared to controls.
IL-33 is known to mediate parasite expulsion in a type-2 dependent-manner (22). The HpARI_CCP1/2 truncated protein maintains the activity of IL-33, potentially amplifying its anti-parasitic effects. It is worthwhile emphasizing that this truncated construct is not a protein naturally secreted by the parasite, but rather a synthetic product with an unexpected activity.
As the IL-33 pathway is strongly implicated in human asthma, HpARI, with its unique mechanism of action and strong binding to IL-33, is a potential therapeutic agent. IL-33 is a potently inflammatory cytokine which is kept tightly regulated. Once released, IL-33 undergoes rapid oxidation and degradation, confining its effects to a short time after release (12, 13). Addition of HpARI or HpARI_CCP1/2 prevented degradation of the cytokine and maintained it in its active form, possibly due to steric hinderance of proteases. As HpARI also blocked the interaction of IL-33 with its receptor there was no cellular response to IL-33 in the presence of HpARI, while HpARI_CCP1/2, which lacks this IL-33-ST2 blocking activity, was unable to inhibit responses to IL-33. Furthermore, most surprisingly, HpARI_CCP1/2 was able to maintain the effects of IL-33 over a long timecourse, potently exacerbating IL-33-dependent responses in vivo and in vitro.
The effects of HpARI_CCP1/2 may not be confined to extending the half-life of IL-33 by preventing its degradation, but may prevent the much more rapid oxidation of the cytokine. Partial oxidation of IL-33 occurs in vivo within 15 min of release (13), therefore the activity of released IL-33 in vivo may be less than that of fully active IL-33. Indeed, when a purified wild-type or an oxidation-resistant mutant of human IL-33 were tested in vitro, the mutant form of IL-33 was found to be 30-fold more potent than WT IL-33 (13). In this study, we were not able to measure the difference between reduced and oxidized IL-33, therefore we cannot make definitive statements about this activity of HpARI_CCP1/2. However, inhibition of IL-33 inactivation, either through prevention of oxidation or proteolytic degradation, could be a potent method for amplifying IL-33-dependent responses.
Although IL-33 is strongly implicated in inducing eosinophilic inflammation in anti-parasite or allergic type 2 immune responses (21, 30), the cytokine has also shown protective effects in models of colitis (31), graft-vs.-host disease (32), autoimmunity (33), obesity (34), wound healing and tissue restoration (35, 36). Therefore, treatments which amplify endogenous IL-33 responses could have clinical potential in a range of treatments.
HpARI_CCP1/2 could also be a useful tool for IL-33 research. Modulating IL-33 responses by using HpARI and HpARI_CCP1/2 in parallel allows assessment of the role of IL-33 in a system in the absence of potentially confounding effects of recombinant cytokine administration or genetic manipulation. In addition, the strategy of IL-33 stabilization by HpARI_CCP1/2 may be able to be replicated using a monoclonal antibody-based therapy, with low-affinity or non-blocking antibodies potentially able to amplify IL-33 responses. As anti-IL-33 treatments enter clinical trials (37), this is an important consideration, as sub-optimal antibodies could result in amplification rather than suppression of IL-33 responses.
This study sheds further light on the mechanism of binding of HpARI to IL-33, the function of the domains of HpARI, and the effects of IL-33 degradation and inactivation. Further structural characterization of HpARI–IL-33 binding will be useful in characterizing this interaction and could allow guided design of more effective IL-33-blocking or IL-33-amplifying therapeutic agents.
Data Availability Statement
All datasets presented in this study are included in the article/Supplementary Material.
Ethics Statement
All mice were accommodated and procedures performed under UK Home Office licenses with institutional oversight performed by qualified veterinarians. UK Home Office project license number 70/8733.
Author Contributions
CC, FV, MW, and HM designed and planned experiments. CC, FV, SC, JR, WG, AO, MW, and HM undertook experiments. MW provided guidance on the design of the SPR experiments and carried these out. HM supervised the work and wrote the first version of the paper. CC, FV, and AO were involved in reviewing and revising the paper. All authors have approved the final version.
Funding
This work was funded by awards to HM from LONGFONDS. Accelerate as part of the AWWA project and the Medical Research Council (MR/S000593/1).
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
Flow cytometry data was generated with support from the QMRI Flow Cytometry and cell sorting facility, University of Edinburgh.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fimmu.2020.01363/full#supplementary-material
References
1. Johnston CJ, Robertson E, Harcus Y, Grainger JR, Coakley G, Smyth DJ, et al. Cultivation of Heligmosomoides polygyrus: an immunomodulatory nematode parasite and its secreted products. J Vis Exp. (2015) 98:e52412. doi: 10.3791/52412
2. Reynolds LA, Filbey KJ, Maizels RM. Immunity to the model intestinal helminth parasite Heligmosomoides polygyrus. Semin Immunopathol. (2012) 34:829–46. doi: 10.1007/s00281-012-0347-3
3. Shimokawa C, Kanaya T, Hachisuka M, Ishiwata K, Hisaeda H, Kurashima Y, et al. Mast cells are crucial for induction of group 2 innate lymphoid cells and clearance of helminth infections. Immunity. (2017) 46:863–74 e864. doi: 10.1016/j.immuni.2017.04.017
4. Harris NL, Loke P. Recent advances in type-2-cell-mediated immunity: insights from helminth infection. Immunity. (2017) 47:1024–36. doi: 10.1016/j.immuni.2017.11.015
5. Johnston CJ, Smyth DJ, Kodali RB, White MPJ, Harcus Y, Filbey KJ, et al. A structurally distinct TGF-β mimic from an intestinal helminth parasite potently induces regulatory T cells. Nat Commun. (2017) 8:1741. doi: 10.1038/s41467-017-01886-6
6. Buck AH, Coakley G, Simbari F, McSorley HJ, Quintana JF, Le Bihan T, et al. Exosomes secreted by nematode parasites transfer small RNAs to mammalian cells and modulate innate immunity. Nat Commun. (2014) 5:5488. doi: 10.1038/ncomms6488
7. Vacca F, Chauche C, Jamwal A, Hinchy EC, Heieis G, Webster H, et al. A helminth-derived suppressor of ST2 blocks allergic responses. Elife. (2020) 9:e54017. doi: 10.7554/eLife.54017.sa2
8. Osbourn M, Soares DC, Vacca F, Cohen ES, Scott IC, Gregory WF, et al. HpARI protein secreted by a helminth parasite suppresses interleukin-33. Immunity. (2017) 47:739–51 e735. doi: 10.1016/j.immuni.2017.09.015
9. Johansson K, McSorley HJ. Interleukin-33 in the developing lung-roles in asthma and infection. Pediatr Allergy Immunol. (2019) 30:503–10. doi: 10.1111/pai.13040
10. Cayrol C, Duval A, Schmitt P, Roga S, Camus M, Stella A, et al. Environmental allergens induce allergic inflammation through proteolytic maturation of IL-33. Nat Immunol. (2018) 19:375–85. doi: 10.1038/s41590-018-0067-5
11. Lefrancais E, Roga S, Gautier V, Gonzalez-de-Peredo A, Monsarrat B, Girard JP, et al. IL-33 is processed into mature bioactive forms by neutrophil elastase and cathepsin G. Proc Natl Acad Sci USA. (2012) 109:1673–8. doi: 10.1073/pnas.1115884109
12. Scott IC, Majithiya JB, Sanden C, Thornton P, Sanders PN, Moore T, et al. Interleukin-33 is activated by allergen- and necrosis-associated proteolytic activities to regulate its alarmin activity during epithelial damage. Sci Rep. (2018) 8:3363. doi: 10.1038/s41598-018-21589-2
13. Cohen ES, Scott IC, Majithiya JB, Rapley L, Kemp BP, England E, et al. Oxidation of the alarmin IL-33 regulates ST2-dependent inflammation. Nat Commun. (2015) 6:8327. doi: 10.1038/ncomms9327
14. Wear MA, Walkinshaw MD. Thermodynamics of the cyclophilin-A/cyclosporin-A interaction: a direct comparison of parameters determined by surface plasmon resonance using Biacore T100 and isothermal titration calorimetry. Anal Biochem. (2006) 359:285–7. doi: 10.1016/j.ab.2006.08.038
15. Neill DR, Wong SH, Bellosi A, Flynn RJ, Daly M, Langford TK, et al. Nuocytes represent a new innate effector leukocyte that mediates type-2 immunity. Nature. (2010) 464:1367–70. doi: 10.1038/nature08900
16. McSorley HJ, Blair NF, Smith KA, McKenzie AN, Maizels RM. Blockade of IL-33 release and suppression of type 2 innate lymphoid cell responses by helminth secreted products in airway allergy. Mucosal Immunol. (2014) 7:1068–78. doi: 10.1038/mi.2013.123
17. Lawrence RA, Gray CA, Osborne J, Maizels RM. Nippostrongylus brasiliensis: cytokine responses and nematode expulsion in normal and IL-4-deficient mice. Exp Parasitol. (1996) 84:65–73. doi: 10.1006/expr.1996.0090
18. Yamamoto T, Endo Y, Onodera A, Hirahara K, Asou HK, Nakajima T, et al. DUSP10 constrains innate IL-33-mediated cytokine production in ST2(hi) memory-type pathogenic Th2 cells. Nat Commun. (2018) 9:4231. doi: 10.1038/s41467-018-06468-8
19. Doherty TA, Khorram N, Chang JE, Kim HK, Rosenthal P, Croft M, et al. STAT6 regulates natural helper cell proliferation during lung inflammation initiated by Alternaria. Am J Physiol Lung Cell Mol Physiol. (2012) 303:L577–88. doi: 10.1152/ajplung.00174.2012
20. Bartemes KR, Iijima K, Kobayashi T, Kephart GM, McKenzie AN, Kita H. IL-33-responsive lineage- CD25+ CD44(hi) lymphoid cells mediate innate type 2 immunity and allergic inflammation in the lungs. J Immunol. (2012) 188:1503–13. doi: 10.4049/jimmunol.1102832
21. Filbey KJ, Camberis M, Chandler J, Turner R, Kettle AJ, Eichenberger RM, et al. Intestinal helminth infection promotes IL-5- and CD4(+) T cell-dependent immunity in the lung against migrating parasites. Mucosal Immunol. (2019) 12:352–62. doi: 10.1038/s41385-018-0102-8
22. Hung LY, Lewkowich IP, Dawson LA, Downey J, Yang Y, Smith DE, et al. IL-33 drives biphasic IL-13 production for noncanonical Type 2 immunity against hookworms. Proc Natl Acad Sci USA. (2013) 110:282–7. doi: 10.1073/pnas.1206587110
23. Wills-Karp M, Rani R, Dienger K, Lewkowich I, Fox JG, Perkins C, et al. Trefoil factor 2 rapidly induces interleukin 33 to promote type 2 immunity during allergic asthma and hookworm infection. J Exp Med. (2012) 209:607–22. doi: 10.1084/jem.20110079
24. Yasuda K, Muto T, Kawagoe T, Matsumoto M, Sasaki Y, Matsushita K, et al. Contribution of IL-33-activated type II innate lymphoid cells to pulmonary eosinophilia in intestinal nematode-infected mice. Proc Natl Acad Sci USA. (2012) 109:3451–6. doi: 10.1073/pnas.1201042109
25. Enoksson M, Moller-Westerberg C, Wicher G, Fallon PG, Forsberg-Nilsson K, Lunderius-Andersson C, et al. Intraperitoneal influx of neutrophils in response to IL-33 is mast cell-dependent. Blood. (2013) 121:530–6. doi: 10.1182/blood-2012-05-434209
26. McCarthy PC, Phair IR, Greger C, Pardali K, McGuire VA, Clark AR, et al. IL-33 regulates cytokine production and neutrophil recruitment via the p38 MAPK-activated kinases MK2/3. Immunol Cell Biol. (2019) 97:54–71. doi: 10.1111/imcb.12200
27. Maizels RM, Smits HH, McSorley HJ. Modulation of host immunity by helminths: the expanding repertoire of parasite effector molecules. Immunity. (2018) 49:801–18. doi: 10.1016/j.immuni.2018.10.016
28. Kouzaki H, Iijima K, Kobayashi T, O'Grady SM, Kita H. The danger signal, extracellular ATP, is a sensor for an airborne allergen and triggers IL-33 release and innate Th2-type responses. J Immunol. (2011) 186:4375–87. doi: 10.4049/jimmunol.1003020
29. Zhao J, Wei J, Mialki RK, Mallampalli DF, Chen BB, Coon T, et al. F-box protein FBXL19-mediated ubiquitination and degradation of the receptor for IL-33 limits pulmonary inflammation. Nat Immunol. (2012) 13:651–8. doi: 10.1038/ni.2341
30. Liew FY, Girard JP, Turnquist HR. Interleukin-33 in health and disease. Nat Rev Immunol. (2016) 16:676–89. doi: 10.1038/nri.2016.95
31. Lopetuso LR, De Salvo C, Pastorelli L, Rana N, Senkfor HN, Petito V, et al. IL-33 promotes recovery from acute colitis by inducing miR-320 to stimulate epithelial restitution and repair. Proc Natl Acad Sci USA. (2018) 115:E9362–70. doi: 10.1073/pnas.1803613115
32. Zhang J, Ramadan AM, Griesenauer B, Li W, Turner MJ, Liu C, et al. ST2 blockade reduces sST2-producing T cells while maintaining protective mST2-expressing T cells during graft-versus-host disease. Sci Transl Med. (2015) 7:308ra160. doi: 10.1126/scitranslmed.aab0166
33. Jiang HR, Milovanovic M, Allan D, Niedbala W, Besnard AG, Fukada SY, et al. IL-33 attenuates EAE by suppressing IL-17 and IFN-γ production and inducing alternatively activated macrophages. Eur J Immunol. (2012) 42:1804–14. doi: 10.1002/eji.201141947
34. Mahlakoiv T, Flamar AL, Johnston LK, Moriyama S, Putzel GG, Bryce PJ, et al. Stromal cells maintain immune cell homeostasis in adipose tissue via production of interleukin-33. Sci Immunol. (2019) 4:eaax0416. doi: 10.1126/sciimmunol.aax0416
35. Monticelli LA, Sonnenberg GF, Abt MC, Alenghat T, Ziegler CG, Doering TA, et al. Innate lymphoid cells promote lung-tissue homeostasis after infection with influenza virus. Nat Immunol. (2011) 12:1045–54. doi: 10.1038/ni.2131
36. Rak GD, Osborne LC, Siracusa MC, Kim BS, Wang K, Bayat A, et al. IL-33-dependent group 2 innate lymphoid cells promote cutaneous wound healing. J Invest Dermatol. (2016) 136:487–96. doi: 10.1038/JID.2015.406
Keywords: Heligmosomoides polygyrus, IL-33, allergy, cytokine, ILC2
Citation: Chauché C, Vacca F, Chia SL, Richards J, Gregory WF, Ogunkanbi A, Wear M and McSorley HJ (2020) A Truncated Form of HpARI Stabilizes IL-33, Amplifying Responses to the Cytokine. Front. Immunol. 11:1363. doi: 10.3389/fimmu.2020.01363
Received: 09 April 2020; Accepted: 28 May 2020;
Published: 30 June 2020.
Edited by:
Paul Giacomin, James Cook University, AustraliaReviewed by:
Lauren Webb, University of Washington, United StatesTiffany Bouchery, Monash University, Australia
DeBroski Herbert, University of Pennsylvania, United States
Copyright © 2020 Chauché, Vacca, Chia, Richards, Gregory, Ogunkanbi, Wear and McSorley. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Henry J. McSorley, aG1jc29ybGV5MDAxQGR1bmRlZS5hYy51aw==