 Valdirene Leão Carneiro1†
Valdirene Leão Carneiro1† Hugo Bernardino Ferreira da Silva2†Gerson de Almeida Queiroz2
Hugo Bernardino Ferreira da Silva2†Gerson de Almeida Queiroz2 Rafael Valente Veiga3
Rafael Valente Veiga3 Pablo Rafael Silveira Oliveira3,4Norma Vilany Queiroz Carneiro2Anaque de Oliveira Pires2Raimon Rios da Silva2
Pablo Rafael Silveira Oliveira3,4Norma Vilany Queiroz Carneiro2Anaque de Oliveira Pires2Raimon Rios da Silva2 Flavia Sena2
Flavia Sena2 Emilia Belitardo2
Emilia Belitardo2 Regina Nascimento2
Regina Nascimento2 Milca Silva2
Milca Silva2 Cintia Rodrigues Marques5Ryan dos Santos Costa2Neuza Maria Alcantra-Neves2
Cintia Rodrigues Marques5Ryan dos Santos Costa2Neuza Maria Alcantra-Neves2 Mauricio L. Barreto3,6
Mauricio L. Barreto3,6 Philip J. Cooper7,8
Philip J. Cooper7,8 Camila Alexandrina Figueiredo2*
Camila Alexandrina Figueiredo2*- 1Department of Life Sciences, State University of Bahia, Salvador, Brazil
- 2Institute of Health Sciences, Federal University of Bahia, Salvador, Brazil
- 3Center for Data and Knowledge Integration for Health, Fiocruz, Salvador, Brazil
- 4Institute of Biological Sciences, Federal University of Bahia, Salvador, Brazil
- 5Multidisciplinary Health Institute, Federal University of Bahia, Salvador, Brazil
- 6Institute of Collective Health, Federal University of Bahia, Salvador, Brazil
- 7School of Medicine, International University of Ecuador, Quito, Ecuador
- 8St. George's University of London, London, United Kingdom
Genetic and epigenetic factors are considered to be critical for host-parasite interactions. There are limited data on the role of such factors during human infections with Ascaris lumbricoides. Here, we describe the potential role of genetic factors as determinants of the Th2 immune response to A. lumbricoides in Brazilian children. Stool samples were collected from the children to detect A. lumbricoides by microscopy and peripheral blood leukocytes (PBLs) were cultured in whole blood cultures for detection of cytokines (IL-5, IL-10, and IL-13) in vitro. Levels of anti-A. lumbricoides IgE and IgG4 were measured in plasma. DNA was extracted from PBLs and genotyped using Illumina 2.5 Human Omni Beadchip. Candidate genes associated with A. lumbricoides responses were identified and SNVs in these selected genes associated with the Th2 immune response to A. lumbricoides. Haplotype, gene expression, and epigenetic analyses were done to identify potential associations with Th2 immune responses. GWAS on samples from 1,189 children identified WSB1 as a candidate gene, and IL-21R was selected as a biologically relevant linked gene for further analysis. Variants in WSB1 and IL21R were associated with markers of Th2 immune responses: increased A. lumbricoides-specific IgE and IL-5/IL-13 by PBLs from infected compared to uninfected individuals. In infected children, WSB1 but not IL21R gene expression was suppressed and increased methylation was observed in the WSB1 promoter region. This is the first study to show an association between genetic variants in WSB1 and IL21R and Th2 immune responses during A. lumbricoides infections in children. WSB1/IL21R pathways could provide a potential target for the treatment of Th2-mediated diseases.
Introduction
A quarter of the world's population is estimated to be infected with soil-transmitted helminth (STH) parasites. The highest prevalence occurs among children living in rural areas of the tropics in conditions of poverty with limited access to treated water and sanitation (1, 2). Among STH infections, Ascaris lumbricoides infection is estimated to infect 820 millions causing a significant burden of morbidity and mortality, the latter generally being associated with intestinal obstruction (3–5). Chronic infections in children, particularly among those with high parasite burdens, can impair host nutrition leading to growth stunting and diminished cognitive development (1, 6).
Ascaris lumbricoides infection induces strong Th2-type immune responses in infected humans leading to the production of high circulating levels of total and parasite-specific IgE, generally targeted at larvae that undergo a phase of extra-intestinal migration through the lungs. Th2-induced host protective mechanisms against A. lumbricoides parasites include eosinophil-mediated killing of larvae in the tissues, mast-cell degranulation in the tissues and intestinal tract, and increased intestinal mucus production through goblet-cell hyperplasia (7, 8).
WSB1, IL21, and IL21R genes are important regulators of the IgE response. The WSB1 gene has a role in the regulation and maturation of the interleukin-21 receptor (IL-21R) (9). The WSB1 gene was initially described through its relationship with the suppressor-protein-signaling box (SOCS) cytokine family (10, 11). The IL21R gene is constitutively expressed on T and B lymphocytes and NK cells (12) and has effects that vary according to the stage of cell differentiation. B cell proliferation and differentiation into plasma cell in vitro appear to occur via IL-21 signaling (13) and IL-21R knock-out mice have high levels of IgE and reduced IgG1. In mice, IL-21 inhibits IgE responses through the IL-21 receptor on B cells, triggering IL-4-independent signaling of STAT3 (14). IL21/IL21R binding activates STAT-3 and production of interferon-gamma by T cells and NK cells that counteracts the effects of IL-4 on IgE production (13). In contrast, IL-21 activates STAT-3 in human B cells and acts synergistically with IL-4 to increase the secretion of IgE (14). Other studies in humans have shown that IL-21 can suppress IgE synthesis, indicating that effects of the IL-21/IL-12R pathway on IgE production may be affected by host genetics: genetic variants in the IL21R gene associated with IgE production have been identified by GWAS (15, 16). IL-21R may have a critical role in the control of allergic responses and helminth infections (17, 18).
The host immune response, during the course of a helminth infection such as A. lumbricoides, involves the induction of complex immune responses that include protective Th2-mediated protective mechanisms. Host genetics is likely to play a key role in resistance and susceptibility to A. lumbricoides (19). Loci shown to be associated with susceptibility to helminth infection include 5q31-q33, signal transducer and transcriptional activator 6 (STAT6) and ligase 4 (LIG4) (20–22). To date, no genome-wide association studies have addressed the role of host genetics in Th2 responses to A. lumbricoides infection and limited candidate-genes studies have been done (22–24). Recently, positive associations between epigenetic alterations of increased histone acetylation and type 2 immune responses including IgE have been observed among individuals infected with A. lumbricoides (25).
In the present study, we used a variety of strategies to study genetic determinants of the host Th2 immune response during A. lumbricoides infection in children that included gene discovery using a genome-wide approach and a candidate gene approach based on the findings of the former. This was followed by expression quantitative trait loci and epigenetic analyses to explore how genetic variations in candidate genes are linked to host Th2 immune response during A. lumbricoides infection.
Methods
Characterization of the Reference Population
This study was done among children and adolescents in the city of Salvador, Brazil, that has a population of 2.8 millions. The study sample has been described in detail elsewhere (2, 26, 27). Briefly, 1,445 children were recruited in early childhood into a prospective study to measure the impact of a sanitation program in the city of Salvador on child morbidity (28). Data were collected from children born between 1994 and 2001, who lived in sentinel areas of the city. Standardized questionnaires were applied to the legal guardian of each child between 1997 and 2003 (baseline) to collect data on demographic and social variables, as well as on the domestic environment. In 2000, fecal samples were collected for detection of geohelminth parasites by microscopy. The children were surveyed again in 2005 to obtain stool and blood samples for laboratory tests and extraction of genomic DNA.
Ethics
The Brazilian National Research Ethics Committee approved the study protocol and informed written consent was obtained from the legal guardian of each child/adolescent (Resolution Number: 15895).
Blood Collection and Cell Culture
Blood samples were collected in heparinized tubes and peripheral blood leukocytes (PBLs) were cultured in whole blood at a dilution of 1:4 in RPMI medium (Gibco, Auckland, New Zealand), supplemented with 10 mmol/L glutamine (Sigma-Aldrich, St Louis, USA) and 100 μg/ml gentamicin (Sigma-Aldrich, St Louis, USA). PBLs were cultured within 6 h of collection in the presence of A. lumbricoides antigen (10 ug/mL, endotoxin-free), pokeweed mitogen (2.5 ug/mL), or no stimulant, in a humidified environment at 37°C with 5% CO2 for 5 days. Supernatant fluids were harvested for 24 h (IL-10) or 5 days of cultures (IL-5 and IL-13) (2, 29).
IL-10, IL-13, and IL-5 Measurements
Concentrations of IL-5, IL-10, and IL-13 in cell culture supernatant were measured using commercial sandwich ELISAs following the manufacturer's instructions (BD PharMingen, San Diego, CA, USA). Cytokine concentrations were dichotomized into responders and non-responders using the lowest detection level for each cytokine. Low/high detection limits (in pg/ml) were 15.6/500 for IL-5, 62.5/4,000 for IL-13 and 31.25/500 for IL-10. The number of individuals evaluated for IL-5 and IL-13 production were 67 and 73, respectively.
Parasitological Analysis
Two fecal samples were collected from each child, separated by a 2-week interval, and analyzed for A. lumbricoides infection using sedimentation (30) and Kato-Katz methods (31) as described (21). Positive children were defined by the presence of A. lumbricoides eggs detected by either method. All positive children were treated with appropriate anthelmintics (26).
IgE and IgG4 Anti-A. lumbricoides Antibodies Serum Concentrations
The ImmunoCAP assay (Phadia Diagnostics AB, Uppsala, Sweden) was used for determination of specific IgE serum concentrations against Ascaris and positive samples had ≥0.35 kU/L of anti–A. lumbricoides IgE. Anti–A. lumbricoides IgG4 was detected using an indirect ELISA as described previously (32).
Genotyping and Quality Control
Genotyping was performed using the Illumina BeadChip Human Omni2.5-8 Kit (www.illumina.com), by the Consortium EPIGEN-Brazil (https://epigen.grude.ufmg.br/). One individual was excluded due to inconsistency between registered and genetic sex, based on X chromosome SNVs and 61 were removed based on kinship coefficients (≥0.1, to include second-degree relatives) between pairs of individuals (33). SNVs excluded from the analysis were: on X, Y and mitochondrial chromosomes; genotyping call rate <0.98; and deviance in the Hardy-Weinberg equilibrium with a P-value <10−4 and Minor Allele Frequency (MAF) <1% (34). After quality control, 1,857,191 autosomal SNVs were included. A total of 636 individuals had detectable values of IgE and/or IgG4 and were included in the analysis. Linear regression was done using ln-transformed ratio of anti-A. lumbricoides IgE to IgG4 (35). Through this genome-wide analysis, we selected SNVs in the WSB1 gene pathway for a candidate gene approach based on biological role in immune response from among the top 20 hits. The closely linked gene pathway for IL21R was selected also. Genotype information for these two genes was extracted from the chip at the following regions: WSB1 from 27294080 to 27315926 (location: NC_000017.11) position at chromosome 17. IL21R from 27402162 to 27452043 (location: NC_000016.10) position at chromosome, 16 and a candidate-gene analysis was done for both genes. For quality control, the following filters were applied: genotyping call rate >90%, imbalance of Hardy-Weinberg equilibrium with P < 0.05 and the Minor Allele Frequency (MAF) >1% (34). A total of 12 markers on WSB1 and 35 markers on IL21R were analyzed after quality control. These data are deposited in the European Nucleotide Archive [PRJEB9080 (ERP010139) Genomic Epidemiology of Complex Diseases in Population-Based Brazilian Cohorts], Accession No. EGAS00001001245, under EPIGEN Committee Controlled Access mode.
Real-Time Quantitative Polymerase Chain Reaction (qRT-PCR)
To evaluate the expression levels of WSB1 and IL21R genes, RNA was isolated from PBL cultures using RNeasy Mini Kit (Qiagen, Hamburg, Germany) and 0.3 μg of total RNA from each sample was reverse transcribed into cDNA using 200 U of Superscript III Reverse Transcriptase (Life Technologies, Carlsbad, CA, USA) and 500 ng of Oligo (dT) (Life Technologies, Carlsbad, CA, USA), as described previously (36). Pre-synthesized Taqman® Gene Expression Assays (Applied Biosystems, Foster City, CA, USA) were used to amplify WSB1 (Hs00373204_m1), IL21R (Hs00222310_m1) and β-actin (Hs01060665_g1). cDNA was detected using QuantStudio 12K Sequence Detection System (Applied Biosystems, Foster City, CA, USA). Each qRT-PCR assay was performed with 10 ng of cDNA in 10 μL of Taqman-PCR Master mix 2X (Applied Biosystems, Foster City, CA, USA) and 1 μL of primer/probe set and purified using deionized H2O q.s. 20 μL. Gene expression was normalized to β-actin levels. Relative quantification was performed using the comparative threshold cycle (ΔΔCT) method (37–39).
In silico Functional Analysis
RegulomeDB (regulomedb.org) is a database for interpretation of regulatory variants in the human genome. It includes high-throughput, experimental datasets from ENCODE (Encyclopedia of DNA Elements) and other sources. A score ranging from 1 to 6 is attributed for each SNV; the lower the score, the greater the presumed involvement in regulatory processes (40).
DNA Methylation Assessment
We used an epigenetic approach to determine the level of methylation on the promoter region WSB1 following infection with A. lumbricoides using OneStep qMethyl kit (Zymo Research). Primers within the CpG rich (promoter) region of WSB1 were: forward, 5′-CAG GCC TTT GCA ATG TTT AGG-3′; reverse, 5′-AGC CAG CAG GTT TTA GGA AGG-3′. Methylation percentages were obtained using 20 ng of DNA in duplicate in test and reference reaction mixes. Reactions were done using a QuantStudio 12K Sequence Detection System (Applied Biosystems, Foster City, CA, USA) as follows: 2 h 37°C; 10 min 95°C; 40 cycles 30 s 95°C, 1 min 54°C, 1 min 72°C followed by an dissociation stage to check specificity of PCR products. The Ct values obtained were used to calculate ΔCt values Ct (test) and Ct (reference). Methylation percentages were calculated as the product of 100 × 2–ΔCt.
Statistical Analysis
For GWAS, linear regressions were done to evaluate the association between SNVs and lg (anti-A. lumbricoides IgE/ anti-A. lumbricoides IgG4) using additive models. Power for genetic association analyses depends on effects of individual polymorphisms (depending on both allelic frequency and associated OR/beta), sample size, and type I error. In the context of GWAS, it is common to consider two levels of significance: a more stringent level such as 5 × 10−8 which may allow a conclusion of statistical significance, and a less stringent level such as 1 × 10−5 to identify potentially suggestive associations. Using an additive model and a type I error of 1 × 10−5, our sample of 996 uninfected and 189 infected children had a power of 80% to detect a polymorphism with beta of 0.3 and frequency >0.15. The statistical power calculation was done using Quanto software (v1.2.4). Models were controlled for confounding by population stratification by inclusion of the first three components of a principal components analysis (PCA) of ancestry informative markers (AIMs) as described (41). In addition, the genomic inflation factor (λ) was estimated to visualize and avoid inflated test statistics (42). Quantile-quantile (Q-Q) plots were used to evaluate the overall significance of the genome-wide association results (Supplementary Figure 2). Associations between polymorphisms in WSB1 or IL21R and A. lumbricoides infection, and IL-5, IL-13, and IL-10 cytokine production by PBLs stimulated with A. lumbricoides and anti-A. lumbricoides IgE and IgG4 were done using logistic regression model in which multivariate models were adjusted for sex, age, and ancestry (first 2 components of PCA analysis of AIMs). Principal components (PC1 and PC2) have categorized individuals according to their ethnic characteristics. Additive models were used in all analyses. Adaptive permutations were also done in adjusted and unadjusted analyses. A computationally intensive procedure based on 1,000,000 permutations was used to estimate the statistical significance of multiple correlation tests in the genetic association analysis (43). Haplotype and genetic risk score analysis were performed using SNPStats program (https://www.snpstats.net/start.htm) (44). Linkage disequilibrium (LD) analysis was done for selected SNVs. Haploview 4.2 software was used to calculate the degree of confidence in the R2-value. Mann-Whitney or Kruskal-Wallis-tests were used to compare continuous variables and the Chi-squared-test to compare frequencies of categorical variables. Except as specified for GWAS, statistical significance was inferred by P < 0.05. Statistical analyses were done using PLINK 1.9 software (www.cog-genomics.org/plink/1.9/), R Statistical Software (Foundation for Statistical Computing, Vienna, Austria), and Prism software version 6 (GraphPad Inc., San Diego, CA).
Results
Characteristics of the Study Population
Of 1,246 children eligible, 61 did not have stool data for A. lumbricoides infection and were excluded from the analysis, leaving 1,189 (996 non-infected and 189 A. lumbricoides infected) children with complete data. Baseline characteristics of the analysis sample are shown in Table 1. Levels of anti-A. lumbricoides IgE and IgG4, and levels of IL-5 produced by A. lumbricoides-stimulated PBLs were greater among infected than non-infected children (P < 0.001).
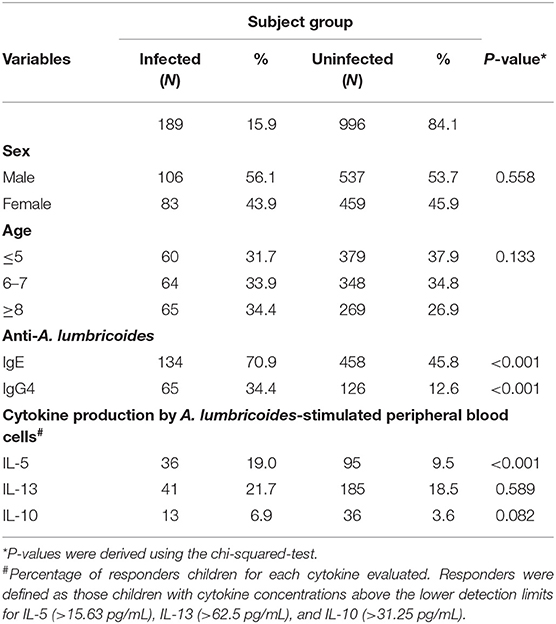
Table 1. Baseline characteristics and immunological markers of A. lumbricoides infection among 1,189 children, stratified by A. lumbricoides infection.
Genome Wide Association Study for SNVs Linked to Parasite-Specific IgE/IgG4 Responses
The Manhattan plot for the genome wide analysis of SNVs associated with anti-A. lumbricoides IgE/IgG4 are shown in Figure 1 and the top 20 SNVs identified are listed in Table 2 with results of a mapping analysis for these provided in Supplementary Table 1. Among identified SNVs, rs7212516 (Beta: 0.33, CI: 0.18–0.47, P = 6.675 × 10−06), is an intronic variant located in the WSB1 (WD repeat and SOCS box containing 1) gene and plays an important role in IgE production (9). Further analyses were focused on WSB1 and the linked gene, IL21R. In addition, SNV rs3093406 in IL21R was significantly associated with IgE/IgG4 ratio in GWAS (P = 0.0222).
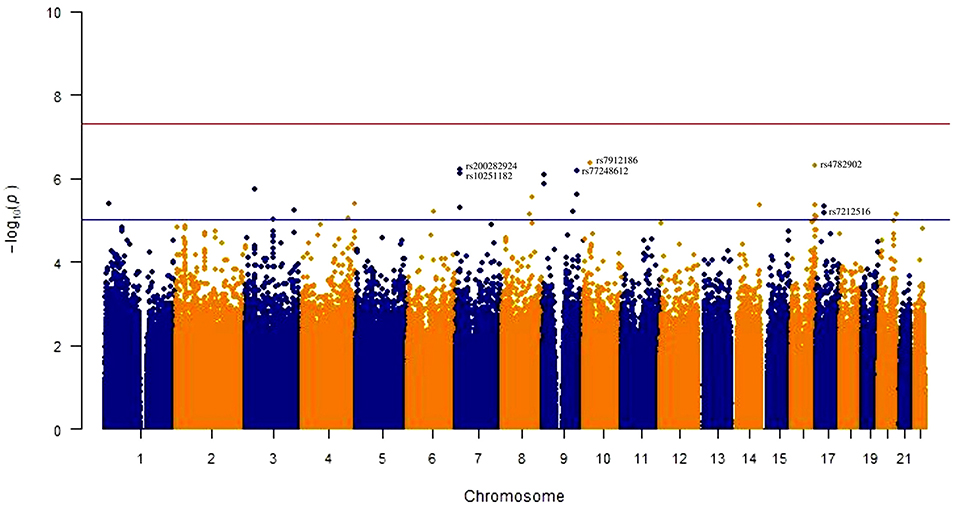
Figure 1. Manhattan plot for genome-wide association with anti-A. lumbricoide IgE/IgG4 ratio for 636 subjects. Each single nucleotide variants (SNVs) was tested for association by linear regression using an additive regression model, adjusted for ancestry markers. The red line indicates genome-wide significance (p = 5 × 10−8) and blue line suggestive significance (P = 1 × 10−5).
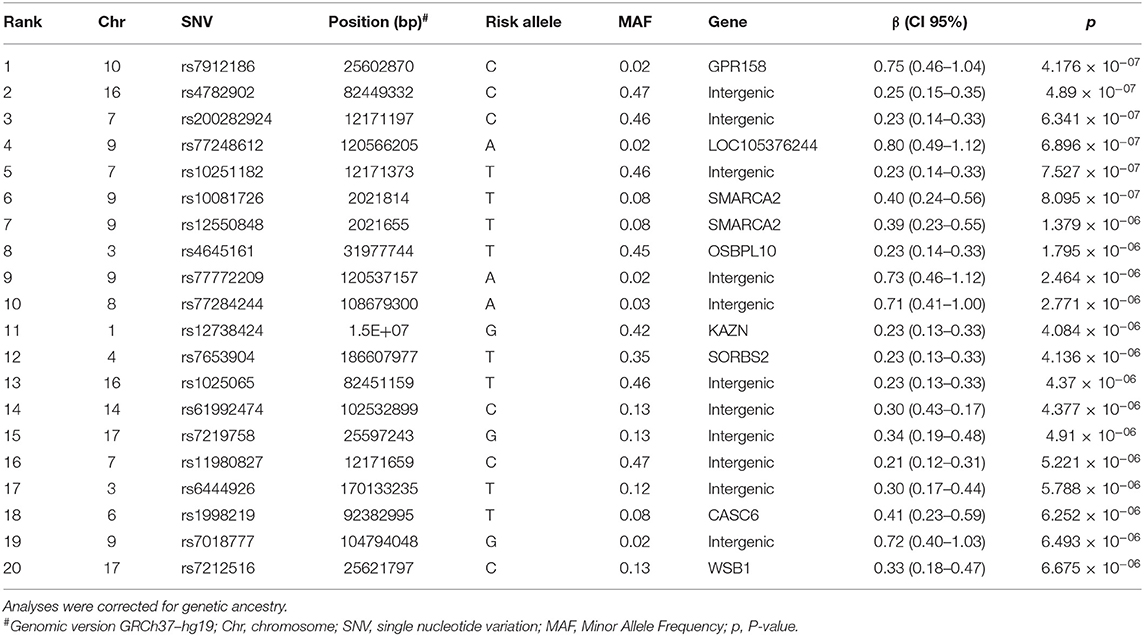
Table 2. The results for the 20 best hits in the genome wide association with study anti-A. lumbricoide IgE/IgG4.
Using log-transformed anti-A. lumbricoides IgE and IgG4 as continuous variables: IgG4 was associated with immune response genes such as PRKCA (a kinase that participates in macrophage differentiation induced by macrophage colony-stimulating factor) (P-values ranging 10−5 and 10−6); and continuous IgE was associated with several genes including NKAI2, a sodium-potassium transporter ATPase in T cells (P-values ranging 10−5 to 10−6).
Associations Between WSB1 and IL21R Variants and Parameters of Host Immune Response to A. lumbricoides
The associations between 12 variants in WSB1 and infection with A. lumbricoides and the host immune response to the parasite were studied (Table 3). P-values refer to the permutational test. With respect to levels of anti-A. lumbricoides specific IgE, SNVs rs7213148, and rs8065359 were positively associated, while rs1060618 and rs9867 were negatively associated. rs9867 was associated with lower production of IL-5 in A. lumbricoides antigen-stimulated PBL cultures. With respect to levels of anti-A. lumbricoides specific IgG4, rs6505199 and rs9303634 were inversely associated (rs6505199 and rs9303634 are in total LD- see Supplementary Figure 1A) and rs7212516 was positively associated.
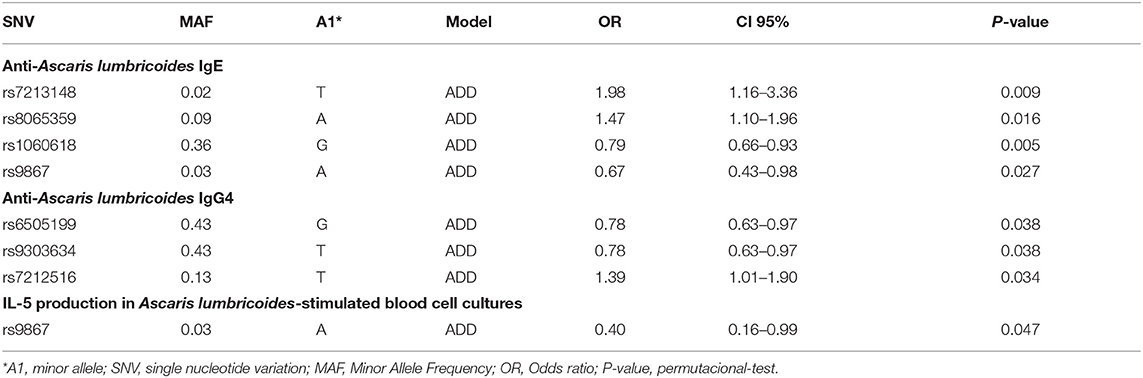
Table 3. Significant associations between SNVs on WSB1 and parameters of the host immune response to A. lumbricoides including specific IgE and IgG4, and IL-5 production by A. lumbricoides antigen-stimulated PBLs.
Because WSB1 is functionally related to IL-21R activation, we scanned the IL21R gene for SNVs associated to A. lumbricoides. Table 4 shows significant associations between IL21R polymorphisms with the presence of measured parameters of infection and immune response to A. lumbricoides. The SNVs rs9938401 and rs3093406 were positively associated with a presence of active A. lumbricoides infection. SNVs rs3093412 and rs179763 were inversely associated with levels of anti-A. lumbricoides IgG4. Four SNVs were inversely associated with levels of anti A. lumbricoides IgE (T allele rs76678990; T allele rs58579343; A allele rs11074859; and T allele rs4140673), while two SNVs were positively associated (T allele rs3093308; A allele, rs115350516). These latter two SNVs were associated also with higher IL-5 production by PBLs stimulated with parasite antigens (T allele, rs3093308; A allele, rs115350516) as were rs3093319 and rs77718993. The SNV rs3091236 was positively associated with IL-10 production while rs9930086 was negatively associated. A high degree of linkage disequilibrium was seen between rs58579343 and rs11074859 (see Supplementary Figure 1B).
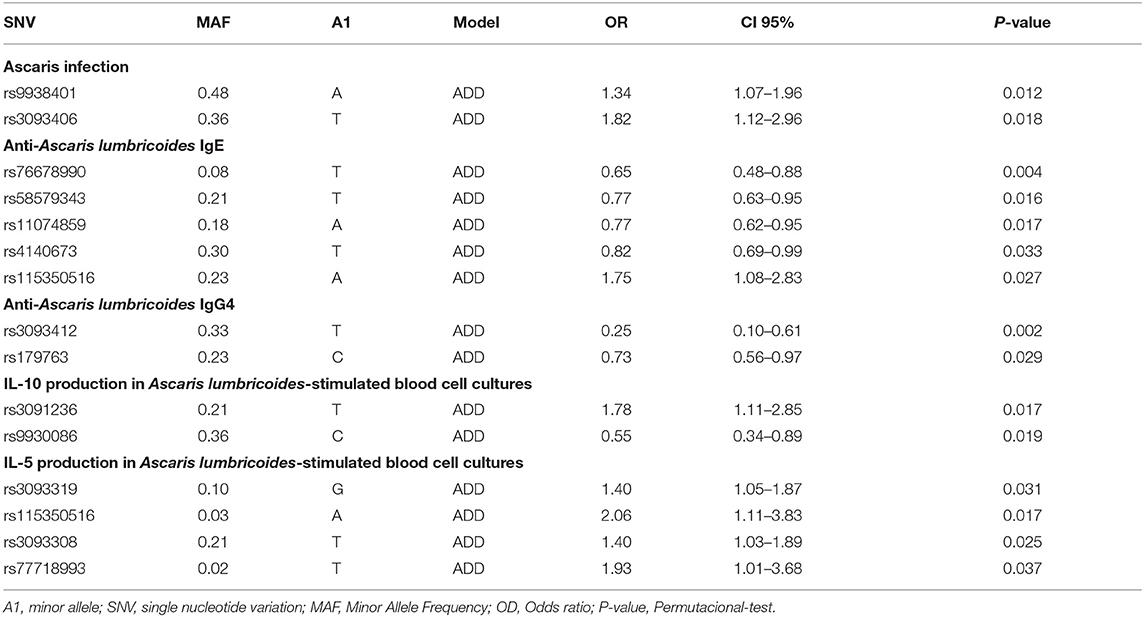
Table 4. Significant associations between SNVs on IL21R and infection with A. lumbricoides and levels of anti-Ascaris lumbricoides IgE and IgG4 and parasite antigen induced production of IL-5 and IL-10 by PBLs.
Haplotype analysis for IL21R SNVs rs115350516 and rs3093308 showed that PBLs from individuals with haplotypes GT and AC produced greater levels of IL-5 when stimulated with parasite antigen (Table 5). Individuals with haplotype AC produced greater levels of anti-A. lumbricoides IgE.

Table 5. Associations between haplotypes for rs115350516 and rs3093308 in the IL21R gene and levels of anti-A. lumbricoides IgE and IL-5 produced by A. lumbricoides-stimulated PBLs.
SNV rs3093308 in IL21R Is Associated With Type 2 Cytokine Production
The T allele of SNV rs3093308 was associated with elevated levels of anti-A. lumbricoides IgE and parasite antigen induced IL-5 production, both indicators of a strong Th2 response and potential resistance to A. lumbricoides infection (45). The presence of one T allele of rs3093308 was associated with higher levels of Th2 cytokines (IL-5 and IL-13, Figures 2A,B) by mitogen-stimulated PBLs among infected individuals (P < 0.01). None of the other SNVs studied were associated with alterations in in vitro cytokine production by infection status (data not shown). However, IL21R gene expression by PBLs was not significantly different between the two alleles (Figure 2C).
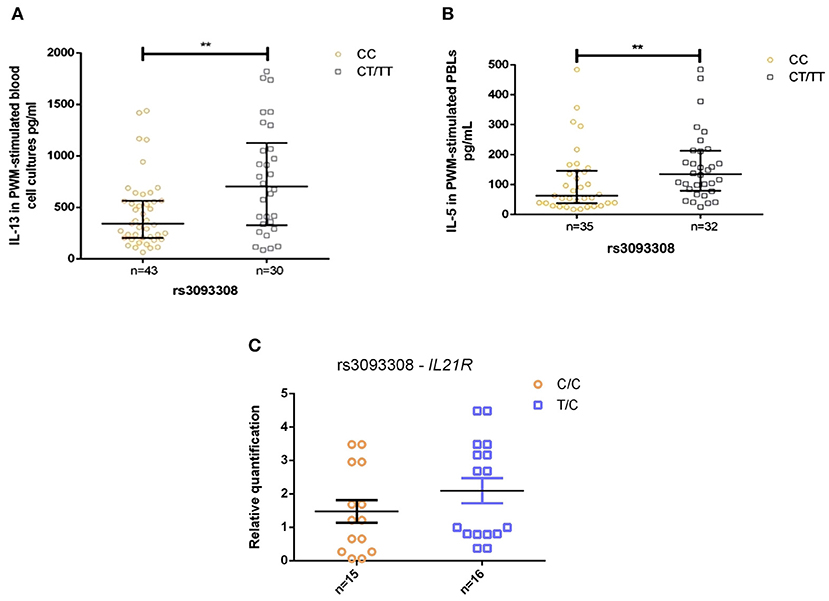
Figure 2. Levels (pg/ml) of IL-5 and IL-13 in A. lumbricoides infected subjects produced by parasite antigen stimulated PBLs, by allele for SNV rs3093308 in IL21R: (A) IL-13, (B) IL-5, and (C) gene expression analysis for IL21R in PBLs (non-significant). CC, genotype CC; CT/TT, genotype CT/TT. **p < 0.01.
Expression of WSB1 and IL21R in Ascaris-Infected and Uninfected Individuals
Figure 3 shows the expression levels of the WSB1 and IL21R genes in A. lumbricoides-infected and uninfected individuals. Expression of WSB1 was lower in infected (N = 15) subjects compared to uninfected (N = 16) subjects (P = 0.0207; Figure 3A). No difference was observed between the two groups for IL21R gene expression (Figure 3B).
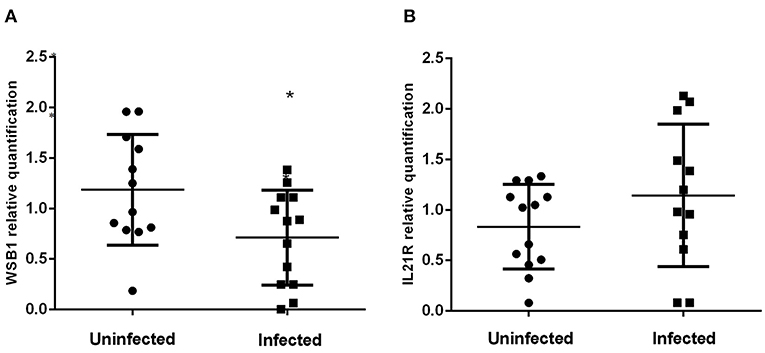
Figure 3. Levels of WSB1 (A) and IL21R (B) gene expression in Ascaris lumbricoides- infected (N = 15) and uninfected (N = 16) subjects in peripheral blood leukocytes. *P ≤ 0.05.
Methylation of WSB1 Promoter Region
Figure 4 shows the percent methylation of WSB1 gene in PBLs from A. lumbricoides-infected (N = 9) and uninfected (N = 8) individuals. Infection with A. lumbricoides was associated with increased WSB1 methylation (P = 0.031). rs7212516 is in the first intron (position 692 bp). The region included in the methylation analysis is in the promoter region (position −771 bp to −443 bp) containing 14 CpG sites and 4 restriction sites (according to NCBI, Gene ID: 26118). The other SNVs are in the position above 6,000 bp. The amplified region for DNA methylation was analyzed in the reference populations of the 1,000 genome project and found 13 SNVs in that region, two of them with a frequency >1% in Africans and African Americans (https://www.ncbi.nlm.nih.gov/variation/tools/1000genomes/?assm=GCF_000001405.25). These two SNVs do not alter or create restriction sites for the enzymes present in the OneStep qMethyl kit (Zymo Research). In addition, none of the 13 SNVs identified in the 1000 genomes project belong to Illumina BeadChip Human Omni2-8-8 Kit, used here for genotyping as well.
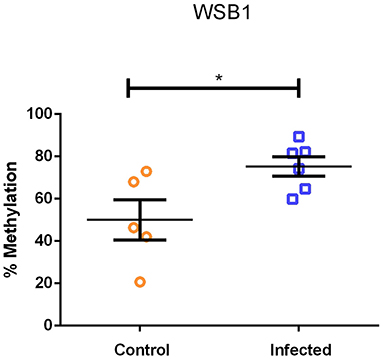
Figure 4. Percentage of DNA methylation in the WSB1 promoter region in Ascaris lumbricoides-infected (N = 9) and uninfected (N = 8) individuals (*P = 0.031, Mann-Whitney-test).
Discussion
Previous studies suggest that the balance between helminth specific IgE and IgG4 might determine resistance or susceptibility to helminth infections, showing that levels of specific IgE have been correlated with resistance to infection, whereas levels of IgG4 have been associated with susceptibility (34–36). In this manuscript, we have conducted two distinct approaches to determine genetic markers associated with A. lumbricoides infection. First, we conducted a GWAS for anti-A. lumbricoides IgE/IgG4 ratio for A. lumbricoides infection in a cohort of an admixture population to determine if there are common genetic variants contributing to susceptibility to A. lumbricoides infection and as a second phase, based on the GWAS pieces of evidence, we focused our attention to WSB1/IL21R pathway, which revealed associations with markers of exposure and cytokine responses to A. lumbricoides. For the best of our knowledge, these associations have never been reported before.
We did not identify any novel SNVs meeting genome-wide significance but did identify several SNVs below the genome-wide threshold as being of potential interest: (1) rs7912186 in the GPR158 gene, described as being linked to plasma membrane scaffold protein in retinal bipolar neurons, contributing to the pathophysiology of steroid-induced ocular hypertension and glaucoma, (46, 47) and also involved in the regulation of the pre-frontal cortex with a potential role chronic stress and depression (48); (2) rs10081726 and rs12550848, located in SMARCA2, that plays a role in the development of lung cancer, hepatocellular carcinoma and esophageal adenocarcinoma (49, 50); and (3) rs7212516, located in the WSB1 gene on chromosome 17 that is known to be involved in IgE regulation (9).
Previous epidemiological studies have shown inverse associations between levels of anti-Ascaris IgE and parasite burden with A. lumbricoides indicating a potential role for IgE in resistance to infection (51, 52). WSB-1, a IL-21 receptor binding molecule, enhances the maturation of IL-21 receptor. WSB1 gene plays an important role in the regulation and maturation of the IL21R, and both genes are important for IgE production (9). For this reason, we included the IL21R as a gene of biological relevance in our candidate gene analysis. There are several lines of evidence showing that IL-21/IL-21R signaling plays a clear role modulating Type 2 cytokines production (9, 11). Mice deficient for IL21R had reduced airways eosinophilia in a model of mite-induced asthma (18), and knock-out mice for IL21R expressed higher levels of IgE and lower levels of IgG1 than normal mice after mite antigen exposure (17).
We explored if genetic variants in WSB1/IL21R might influence Th2-associated immune responses during A. lumbricoides infection using immunological markers of susceptibility and resistance to infection including production of Th2 cytokines in vitro. Our results show that variants in these two genes are associated with such markers of the host Th2 response during this helminth infection. SNVs in WSB1 (rs7213148 and rs8065359) and IL21R [rs115350516 (A allele) and rs3093308 (T allele)] were associated with increased production of A. lumbricoides-specific IgE (Tables 4, 5) and could be potentially linked to greater resistance to infection. The same two SNVs in IL21R SNVs were associated with greater parasite antigen-induced IL-5 production that has been linked to resistance to geohelminth infections (45). These SNVs have not been linked previously to helminth infection or Th2-driven inflammatory conditions.
The T allele of rs3093308 in IL21R was associated also with increased production of Th2 cytokines (IL-5 and IL-13) by mitogen-induced PBLs among infected compared to uninfected children (Figures 2A,B), and the same SNV tended to increase IL21R gene expression (Figure 2C). These findings could be indicative of a stronger protective immune response against A. lumbricoides infection. No previous studies have reported a role for this SNV. In a study evaluating IL21/IL21R signaling in murine model of intestinal inflammation, Th2 responses (IL-4 and IL-5 by CD4+ T cells) were markedly suppressed in IL21R deficient compared to wild-type mice (53).
Our data can explain, at least in part, findings from previous studies showing elevated Ascaris-specific IgE levels to be associated with decreased worm burden and increased resistance to infections with this helminth (51, 52). Other studies have shown significant associations between locus 13q33 that includes the genes, LIG4, ABHD13, and TNFSF13B, with Ascaris-specific IgE levels (22, 54, 55). Thus, consistent with our findings, genetic regulation of IgE production may play an essential role in susceptibility to Ascaris infection.
In our population, the G and T alleles of SNVs rs6505199 and rs9303634, respectively, in WSB1 (see Table 3), were associated with reduced Ascaris-specific IgG4 levels. These results favor increased production of IgE relative to IgG4, the latter known to be a marker of susceptibility to infection (51, 56). Both SNVs were in high linkage disequilibrium (r2 = 1.00), (see Supplementary Figure 1A). Conversely, the T allele of SNV rs7212516 was positively associated with Ascaris-specific IgG4 and perhaps greater susceptibility to the infection.
We also did haplotype analyses in WSB1 and IL21R genes for anti-A. lumbricoides IL-5 and IgE production. Two SNVs IL21R (rs115350516 and rs3093308) and their haplotypes, especially the AC haplotype, were associated with increased production of IL-5 by Ascaris-stimulated PBLs. This same haplotype showed a positive association with anti-A. lumbricoides IgE levels. Interestingly, in regression analyses these same SNVs were associated with increased anti-A. lumbricoides IgE and IL-5 by Ascaris-stimulated PBLs which could be linked to a more effective protective immune response against the parasite. Previous studies have analyzed levels of WSB1 expression in the brain, spleen, kidney and placenta, primarily with research focusing on cancer development (9, 57). There is no previous study describing the role of WSB1 in helminth infections or any other Th2-driven condition.
In our gene expression assay, the WSB1 had lower expression levels in infected subjects when compared with non-infected subjects (Figure 3A). This result allows us to hypothesize that low levels of WSB1 expression in infected subjects may be related to epigenetic regulation as we have demonstrated increased methylation of the WSB1 promoter region in infected individuals (Figure 4). Although cell populations within whole blood cultures may differ between cases and controls [infected subjects had greater total leukocyte and eosinophil counts than uninfected subjects (data not shown)], our findings indicated that infection was associated with greater methylation but lower gene expression. However, further studies are required to support a potential effect of A. lumbricoides infections on WSB1 gene hypermethylation. Epigenetic events, such as post-transcriptional modifications of DNA at CPG sites, regulate gene transcription activity, thereby determining the kinetics and final expression (58, 59). On the other hand, there was no statistical difference in gene expression levels for the IL21R (Figure 3B). Persistent helminth infections appear to induce changes in DNA methylation in CD4+ cells from helminth-infected individuals. Other epigenetic mechanisms may also be involved in the expression of key genes in the type 2 immune response. A study evaluated histone acetylation in individuals exposed to A. lumbricoides found that histone acetylation levels in IL-4 and IL-13 genes were altered by infection (25).
This study has a number of potential limitations including: a relatively small sample size limiting power using the GWAS genome strategy; we were unable to do a replication analysis because of a lack of previous studies collecting data on the same variables (e.g., anti-A. lumbricoides IgE or IgG4); and use of whole blood cultures rather than more homogeneous lymphocyte populations due to logistical issues inherent to a population-based study such as ours (26).
Our results, therefore, provide novel mechanistic insights into how helminth infections that affect immune response regulation may modulate also epigenetic processes. Further studies are needed to improve our understanding on how such regulation may occur and the consequences for Th2-driven inflammatory conditions.
Data Availability Statement
The genetic data sets generated in this study can be found online upon request at https://cidacs.bahia.fiocruz.br/en/platform/epigen-genomic-epidemiology-of-brazilian-cohorts/.
Ethics Statement
The studies involving human participants were reviewed and approved by Institute of Collective Health, Federal University of Bahia, Salvador, Brazil. Written informed consent to participate in this study was provided by the participants' legal guardian/next of kin.
Author Contributions
VC, HS, GQ, RV, PO, NC, AP, and RS have conducted the field work, managed the database, and performed the genetic experiments. FS, EB, CM, and RC were responsible for the supervision of the field work and/or laboratory experiments. VC and HS were responsible for data analysis and writing of the manuscript. MB, NA-N, and PC drafting the paper and revising it critically. CF substantial contributions to research design, or the acquisition, analysis or interpretation of data. All authors have actively participated of the analysis of research results and approve the version that is being submitted.
Funding
This study was conducted through the SCAALA (Social change, Asthma and Allergy in Latin America) initiative, funded by the Wellcome Trust, Grant No. 072405/Z/03/Z. It was also supported by the Brazilian Ministry of Health (Department of Science and Technology from the Secretaria de Ciência, Tecnologia e Insumos Estratégicos) through Financiadora de Estudos e Projetos, Fundação de Amparo a Pesquisa do Estado da Bahia (FAPESB)/CNPq Grant Nos. PRONEX- PNX00001/2014 and PRONEM-PNE0003/2014 and Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq) for the doctoral fellowship granted.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fimmu.2021.622051/full#supplementary-material
References
1. Bethony J, Brooker S, Albonico M, Geiger SM, Loukas A, Diemert D, et al. Soil-transmitted helminth infections: ascariasis, trichuriasis, and hookworm. Lancet. (2006) 367:1521–32. doi: 10.1016/S0140-6736(06)68653-4
2. Figueiredo CA, Alcântara-Neves NM, Veiga R, Amorim LD, Dattoli V, Mendonça LR, et al. Spontaneous cytokine production in children according to biological characteristics and environmental exposures. Environ Health Perspect. (2009) 117:845–9. doi: 10.1289/ehp.0800366
3. Hotez PJ, Molyneux DH, Fenwick A, Kumaresan J, Sachs SE, Sachs JD, et al. Control of neglected tropical diseases. N Engl J Med. (2007) 357:1018–27. doi: 10.1056/NEJMra064142
4. Ekpenyong EA, Eyo EJ. Prevalence of intestinal helminths infections among schooling children in tropical semi urban communities. Anim Res Int. (2008) 5:804–10. doi: 10.4314/ari.v5i1.48715
5. Pullan RL, Smith JL, Jasrasaria R, Brooker SJ. Global numbers of infection and disease burden of soil transmitted helminth infections in 2010. Parasites Vectors. (2014) 7:37. doi: 10.1186/1756-3305-7-37
6. Dold C, Holland CV. Ascaris and ascariasis. Microb Infect. (2011) 13:632–7. doi: 10.1016/j.micinf.2010.09.012
7. Masure D, Vlaminck J, Wang T, Chiers K, Van den Broeck W, Vercruysse J, et al. A role for eosinophils in the intestinal immunity against infective Ascaris suum larvae. PLoS Negl Trop Dis. (2013) 7:e2138. doi: 10.1371/journal.pntd.0002138
8. Coakley G, Volpe B, Bouchery T, Shah K, Butler A, Geldhof P, et al. Immune serum-activated human macrophages coordinate with eosinophils to immobilize Ascaris suum larvae. Parasite Immunol. (2020) 42:e12728. doi: 10.1111/pim.12728
9. Nara H, Onoda T, Rahman M, Araki A, Juliana FM, Tanaka N, et al. WSB-1, a novel IL-21 receptor binding molecule, enhances the maturation of IL-21 receptor. Cell Immunol. (2011) 269:54–9. doi: 10.1016/j.cellimm.2011.03.010
10. Nicholson SE, Hilton DJ. The SOCS proteins: a new family of negative regulators of signal transduction. J Leukoc Biol. (1998) 63:665–8. doi: 10.1002/jlb.63.6.665
11. Haque M, Kendal JK, MacIsaac RM, Demetrick DJ. WSB1: from homeostasis to hypoxia. J Biomed Sci. (2016) 23:1–10. doi: 10.1186/s12929-016-0270-3
12. Mehta DS, Wurster AL, Grusby MJ. Biology of IL-21 and the IL-21 receptor. Immunol Rev. (2004) 202:84–95. doi: 10.1111/j.0105-2896.2004.00201.x
13. Kuchen S, Robbins R, Sims GP, Sheng C, Phillips TM, Lipsky PE, et al. Essential role of IL-21 in B cell activation, expansion, and plasma cell generation during CD4+ T cell-B cell collaboration. J Immunol. (2007) 179:5886–96. doi: 10.4049/jimmunol.179.9.5886
14. Avery DT, Ma CS, Bryant VL, Santner-Nanan B, Nanan R, Wong M, et al. STAT3 is required for IL-21-induced secretion of IgE from human naive B cells. Blood. (2008) 112:1784–93. doi: 10.1182/blood-2008-02-142745
15. Potaczek DP, Kabesch M. Current concepts of IgE regulation and impact of genetic determinants. Clin Exp Allergy. (2012) 42:852–71. doi: 10.1111/j.1365-2222.2011.03953.x
16. Pène J, Guglielmi L, Gauchat J-F, Harrer N, Woisetschläger M, Boulay V, et al. IFN-γ-mediated inhibition of human IgE synthesis by IL-21 is associated with a polymorphism in the IL-21R gene. J Immunol. (2006) 177:5006–13. doi: 10.4049/jimmunol.177.8.5006
17. Ozaki K, Spolski R, Feng CG, Qi C-F, Cheng J, Sher A, et al. A critical role for IL-21 in regulating immunoglobulin production. Science. (2002) 298:1630–4. doi: 10.1126/science.1077002
18. Coquet JM, Schuijs MJ, Smyth MJ, Deswarte K, Beyaert R, Braun H, et al. Interleukin-21-producing CD4+ T cells promote type 2 immunity to house dust mites. Immunity. (2015) 43:318–30. doi: 10.1016/j.immuni.2015.07.015
19. Mangano VD, Modiano D. Host genetics and parasitic infections. Clin Microbiol Infect. (2014) 20:1265–75. doi: 10.1111/1469-0691.12793
20. Peisong G, Mao XQ, Enomoto T, Feng Z, Gloria-Bottini F, Bottini E, et al. An asthma-associated genetic variant of STAT6 predicts low burden of ascaris worm infestation. Genes Immun. (2005) 6:70. doi: 10.1038/sj.gene.6364148
21. Moller M, Gravenor MB, Roberts SE, Sun D, Gao P, Hopkin JM. Genetic haplotypes of Th-2 immune signalling link allergy to enhanced protection to parasitic worms. Hum Mol Genet. (2007) 16:1828–36. doi: 10.1093/hmg/ddm131
22. Acevedo N, Mercado D, Vergara C, Sanchez J, Kennedy MW, Jiménez S, et al. Association between total immunoglobulin E and antibody responses to naturally acquired Ascaris lumbricoides infection and polymorphisms of immune system-related LIG4, TNFSF13B and IRS2 genes. Clin Exp Immunol. (2009) 157:282–90. doi: 10.1111/j.1365-2249.2009.03948.x
23. Quinnell RJ. Genetics of susceptibility to human helminth infection. Int J Parasitol. (2003) 33:1219–31. doi: 10.1016/S0020-7519(03)00175-9
24. Zamanian M, Cook DE, Zdraljevic S, Brady SC, Lee D, Lee J, et al. Discovery of genomic intervals that underlie nematode responses to benzimidazoles. PLoS Negl Trop Dis. (2018) 12:e0006368. doi: 10.1371/journal.pntd.0006368
25. Zakzuk J, Acevedo N, Harb H, Eick L, Renz H, Potaczek DP, et al. IgE levels to Ascaris and house dust mite allergens are associated with increased histone acetylation at key type-2 immune genes. Front Immunol. (2020) 11:756. doi: 10.3389/fimmu.2020.00756
26. Barreto ML, Cunha SS, Alcântara-Neves N, Carvalho LP, Cruz ÁA, Stein RT, et al. Risk factors and immunological pathways for asthma and other allergic diseases in children: background and methodology of a longitudinal study in a large urban center in Northeastern Brazil (Salvador-SCAALA study). BMC Pulm Med. (2006) 6:15. doi: 10.1186/1471-2466-6-15
27. Rodrigues LC, Newcombe PJ, Cunha SS da, Alcantara-Neves NM, Genser B, Cruz AA, et al. Early infection with Trichuris trichiura and allergen skin test reactivity in later childhood. Clin Exp Allergy. (2008) 38:1769–77. doi: 10.1111/j.1365-2222.2008.03027.x
28. Barreto ML, Genser B, Strina A, Assis AMO, Rego RF, Teles CA, et al. Effect of city-wide sanitation programme on reduction in rate of childhood diarrhoea in northeast Brazil: assessment by two cohort studies. Lancet. (2007) 370:1622–8. doi: 10.1016/S0140-6736(07)61638-9
29. Figueiredo CA, Barreto ML, Rodrigues LC, Cooper PJ, Silva NB, Amorim LD, et al. Chronic intestinal helminth infections are associated with immune hyporesponsiveness and induction of a regulatory network. Infect Immun. (2010) 78:3160–7. doi: 10.1128/IAI.01228-09
30. Hoffman WA, Pons JA, Janer JL. The Sedimentation-Concentration Method in Schistosomiasis Mansoni, (1934).
31. Carvalho O dos S, Massara CL, Silveira Neto HV, Alvarenga AG, Vidigal TH, Guerra HL, et al. Schistosomiasis mansoni in the region of the Triangulo Mineiro, State of Minas Gerais, Brazil. Memórias do Instituto Oswaldo Cruz. (1994) 89:509–12. doi: 10.1590/S0074-02761994000400003
32. Figueiredo CA, Amorim LD, Alcantara-Neves NM, Matos SMA, Cooper PJ, Rodrigues LC, et al. Environmental conditions, immunologic phenotypes, atopy, and asthma: new evidence of how the hygiene hypothesis operates in Latin America. J Allergy Clin Immunol. (2013) 131:1064–8.e1. doi: 10.1016/j.jaci.2013.01.016
33. Thornton T, Tang H, Hoffmann TJ, Ochs-Balcom HM, Caan BJ, Risch N. Estimating kinship in admixed populations. Am J Hum Genet. (2012) 91:122–38. doi: 10.1016/j.ajhg.2012.05.024
34. Laurie CC, Doheny KF, Mirel DB, Pugh EW, Bierut LJ, Bhangale T, et al. Quality control and quality assurance in genotypic data for genome-wide association studies. Genet Epidemiol. (2010) 34:591–602. doi: 10.1002/gepi.20516
35. Transformation of Summary Statistics from Linear Mixed Model Association on All-or-None Traits to Odds Ratio|Genetics. Available online at: https://www.genetics.org/content/208/4/1397.abstract (accessed October 5, 2020).
36. Rios R, da Silva HBF, Carneiro NVQ, de Oliveira Pires A, Carneiro TCB, dos Santos Costa R, et al. Solanum paniculatum L. decreases levels of inflammatory cytokines by reducing NFKB, TBET and GATA3 gene expression in vitro. J Ethnopharmacol. (2017) 209:32–40. doi: 10.1016/j.jep.2017.07.014
37. Orlando C, Pinzani P, Pazzagli M. Developments in quantitative PCR. Clin Chem Lab Med. (1998) 36:255–69. doi: 10.1515/CCLM.1998.045
38. Vandesompele J, De Preter K, Pattyn F, Poppe B, Van Roy N, De Paepe A, et al. Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genome Biol. (2002) 3:research0034-1. doi: 10.1186/gb-2002-3-7-research0034
39. Hellemans J, Mortier G, De Paepe A, Speleman F, Vandesompele J. qBase relative quantification framework and software for management and automated analysis of real-time quantitative PCR data. Genome Biol. (2007) 8:1–14. doi: 10.1186/gb-2007-8-2-r19
40. Boyle AP, Hong EL, Hariharan M, Cheng Y, Schaub MA, Kasowski M, et al. Annotation of functional variation in personal genomes using RegulomeDB. Genome Res. (2012) 22:1790–7. doi: 10.1101/gr.137323.112
41. Costa GN, Dudbridge F, Fiaccone RL, da Silva TM, Conceição JS, Strina A, et al. A genome-wide association study of asthma symptoms in Latin American children. BMC Genet. (2015) 16:141. doi: 10.1186/s12863-015-0296-7
42. Price AL, Patterson NJ, Plenge RM, Weinblatt ME, Shadick NA, Reich D. Principal components analysis corrects for stratification in genome-wide association studies. Nat Genet. (2006) 38:904–9. doi: 10.1038/ng1847
43. Doerge RW, Churchill GA. Permutation tests for multiple loci affecting a quantitative character. Genetics. (1996) 142:285–94. doi: 10.1093/genetics/142.1.285
44. Solé X, Guinó E, Valls J, Iniesta R, Moreno V. SNPStats: a web tool for the analysis of association studies. Bioinformatics. (2006) 22:1928–9. doi: 10.1093/bioinformatics/btl268
45. Cooper PJ, Ayre G, Martin C, Rizzo JA, Ponte EV, Cruz AA. Geohelminth infections: a review of the role of IgE and assessment of potential risks of anti-IgE treatment. Allergy. (2008) 63:409–17. doi: 10.1111/j.1398-9995.2007.01601.x
46. Orlandi C, Posokhova E, Masuho I, Ray TA, Hasan N, Gregg RG, et al. GPR158/179 regulate G protein signaling by controlling localization and activity of the RGS7 complexes. J Cell Biol. (2012) 197:711–9. doi: 10.1083/jcb.201202123
47. Patel N, Itakura T, Gonzalez JM Jr, Schwartz SG, Fini ME. GPR158, an orphan member of G protein-coupled receptor Family C: glucocorticoid-stimulated expression and novel nuclear role. PLoS ONE. (2013) 8:e57843. doi: 10.1371/journal.pone.0057843
48. Sutton LP, Orlandi C, Song C, Oh WC, Muntean BS, Xie K, et al. Orphan receptor GPR158 controls stress-induced depression. Elife. (2018) 7:e33273. doi: 10.7554/eLife.33273
49. Korpanty GJ, Eng L, Qiu X, Faluyi OO, Renouf DJ, Cheng D, et al. Association of BRM promoter polymorphisms and esophageal adenocarcinoma outcome. Oncotarget. (2017) 8:28093. doi: 10.18632/oncotarget.15890
50. Pasic I, Wong KM, Lee JJ, Espin-Garcia O, Brhane Y, Cheng D, et al. Two BRM promoter polymorphisms predict poor survival in patients with hepatocellular carcinoma. Mol Carcinog. (2018) 57:106–13. doi: 10.1002/mc.22736
51. Turner JD, Faulkner H, Kamgno J, Kennedy MW, Behnke J, Boussinesq M, et al. Allergen-specific IgE and IgG4 are markers of resistance and susceptibility in a human intestinal nematode infection. Microb Infect. (2005) 7:990–6. doi: 10.1016/j.micinf.2005.03.036
52. Hagel I, Cabrera M, Sánchez P, Rodríguez P, Lattouf JJ. Role of the low affinity IgE receptor (CD23) on the IgE response against Ascaris lumbricoides in Warao Amerindian children from Venezuela. Investig Clin. (2006) 47:241–51.
53. Wang Y, Jiang X, Zhu J, Yue D, Zhang X, Wang X, et al. IL-21/IL-21R signaling suppresses intestinal inflammation induced by DSS through regulation of Th responses in lamina propria in mice. Sci Rep. (2016) 6:31881. doi: 10.1038/srep31881
54. Williams-Blangero S, VandeBerg JL, Subedi J, Aivaliotis MJ, Rai DR, Upadhayay RP, et al. Genes on chromosomes 1 and 13 have significant effects on Ascaris infection. Proc Natl Acad Sci. (2002) 99:5533–8. doi: 10.1073/pnas.082115999
55. Acevedo N, Bornacelly A, Mercado D, Unneberg P, Mittermann I, Valenta R, et al. Genetic variants in CHIA and CHI3L1 are associated with the IgE response to the ascaris resistance marker ABA-1 and the Birch pollen allergen Bet v 1. PLoS ONE. (2016) 11:e0167453. doi: 10.1371/journal.pone.0167453
56. King E-M, Kim HT, Dang NT, Michael E, Drake L, Needham C, et al. Immuno-epidemiology of Ascaris lumbricoides infection in a high transmission community: antibody responses and their impact on current and future infection intensity. Parasite Immunol. (2005) 27:89–96. doi: 10.1111/j.1365-3024.2005.00753.x
57. Archange C, Nowak J, Garcia S, Moutardier V, Calvo EL, Dagorn J-C, et al. The WSB1 gene is involved in pancreatic cancer progression. PLoS ONE. (2008) 3:e2475. doi: 10.1371/journal.pone.0002475
58. Buecker C, Wysocka J. Enhancers as information integration hubs in development: lessons from genomics. Trends Genet. (2012) 28:276–84. doi: 10.1016/j.tig.2012.02.008
Keywords: Ascaris lumbricoides, immunity, polymorphism, WSB1, IL21R
Citation: Carneiro VL, da Silva HBF, Queiroz GdA, Veiga RV, Oliveira PRS, Carneiro NVQ, Pires AdO, da Silva RR, Sena F, Belitardo E, Nascimento R, Silva M, Marques CR, Costa RdS, Alcantra-Neves NM, Barreto ML, Cooper PJ and Figueiredo CA (2021) WSB1 and IL21R Genetic Variants Are Involved in Th2 Immune Responses to Ascaris lumbricoides. Front. Immunol. 12:622051. doi: 10.3389/fimmu.2021.622051
Received: 27 October 2020; Accepted: 29 January 2021;
Published: 22 February 2021.
Edited by:
Petra Ina Pfefferle, University of Marburg, GermanyReviewed by:
Daniel Piotr Potaczek, Philipps University Marburg, GermanyNathalie Acevedo, University of Cartagena, Colombia
Copyright © 2021 Carneiro, da Silva, Queiroz, Veiga, Oliveira, Carneiro, Pires, da Silva, Sena, Belitardo, Nascimento, Silva, Marques, Costa, Alcantra-Neves, Barreto, Cooper and Figueiredo. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Camila Alexandrina Figueiredo, Y2F2ZmlndWVpcmVkb0BnbWFpbC5jb20=
†These authors have contributed equally to this work