 Kazuki M. Matsuda1†
Kazuki M. Matsuda1† Yumi Umeda-Kameyama2
Yumi Umeda-Kameyama2 Kazuhiro Iwadoh1Masashi Miyawaki2Mitsutaka Yakabe2Masaki Ishii2
Kazuhiro Iwadoh1Masashi Miyawaki2Mitsutaka Yakabe2Masaki Ishii2 Sumito Ogawa2Masahiro Akishita2,3
Sumito Ogawa2Masahiro Akishita2,3 Shinichi Sato1
Shinichi Sato1 Ayumi Yoshizaki1*†
Ayumi Yoshizaki1*†- 1Department of Dermatology, The University of Tokyo Graduate School of Medicine, Tokyo, Japan
- 2Department of Geriatric Medicine, The University of Tokyo Graduate School of Medicine, Tokyo, Japan
- 3Tokyo Metropolitan Institute for Geriatrics and Gerontology, Tokyo, Japan
Introduction: Dementia is a neurodegenerative syndrome marked by the accumulation of disease-specific proteins and immune dysregulation, including autoimmune mechanisms involving autoantibodies. Current diagnostic methods are often invasive, time-consuming, or costly.
Methods: This study explores the use of proteome-wide autoantibody screening (PWAbS) for noninvasive dementia diagnosis by analyzing serum samples from Alzheimer's disease (AD), dementia with Lewy bodies (DLB), and age-matched cognitively normal individuals (CNIs). Serum samples from 35 subjects were analyzed utilizing our original wet protein arrays displaying more than 13,000 human proteins.
Results: PWAbS revealed elevated gross autoantibody levels in AD and DLB patients compared to CNIs. A total of 229 autoantibodies were differentially elevated in AD and/or DLB, effectively distinguishing between patient groups. Machine learning models showed high accuracy in classifying AD, DLB, and CNIs. Gene ontology analysis highlighted autoantibodies targeting neuroactive ligands/receptors in AD and lipid metabolism proteins in DLB. Notably, autoantibodies targeting neuropeptide B (NPB) and adhesion G protein-coupled receptor F5 (ADGRF5) showed significant correlations with clinical traits including Mini Mental State Examination scores.
Discussion: The study demonstrates the potential of PWAbS and artificial intelligence integration as a noninvasive diagnostic tool for dementia, uncovering biomarkers that could enhance understanding of disease mechanisms. Limitations include demographic differences, small sample size, and lack of external validation. Future research should involve longitudinal observation in larger, diverse cohorts and functional studies to clarify autoantibodies' roles in dementia pathogenesis and their diagnostic and therapeutic potential.
Introduction
Dementia is a complex neurodegenerative syndrome affecting millions worldwide. Early diagnosis is crucial for timely intervention, yet many current diagnostic methods are either invasive, time-consuming, or expensive. For instance, psychological assessments require significant time and concern to patients themselves, cerebrospinal fluid examination is invasive, and amyloid positron emission tomography (PET) is costly. Consequently, there is a pressing need for a simpler, noninvasive, and cost-effective diagnostic method for dementia (1–3).
Pathologically, dementia is marked by the aggregation of disease-specific proteins in the brain (4). While the pathogenic role of abnormal protein deposition in dementia is well-established, the precise mechanisms behind the initiation and progression of neurodegeneration remain unclear. Meanwhile, emerging evidence has highlighted the role of immune dysregulation in dementia’s pathogenesis. Genome-wide association studies have identified common genetic variations in immune system processes that are associated with neurodegenerative diseases such as Alzheimer’s disease (AD), frontotemporal dementia (FTD), and Parkinson’s disease dementia (5–7).
Autoimmune mechanisms are gaining recognition as a key factor in the pathophysiology of dementia (8–10). Autoantibodies—self-reactive antibodies produced by B cells—play a role in immune tolerance and homeostasis (11). However, due to various genetic and environmental factors, the ability to distinguish “self” from “non-self” deteriorates, leading autoantibodies to trigger and sustain inflammatory processes that cause tissue damage (12, 13). Autoantibodies have been detected in both blood and cerebrospinal fluid of patients with various forms of dementia, including autoimmune dementia and neurodegenerative dementias such as AD, FTD, vascular dementia (VD), and dementia with Lewy bodies (DLB) (14–18). Autoimmune dementia is characterized by progressive cognitive decline with an early onset, atypical clinical presentation, rapid progression, the presence of neural antibodies, cerebrospinal fluid inflammation, brain changes in MRI atypical for neurodegenerative diseases, and a good response to immunotherapy (19). Various neural autoantibodies have been frequently identified in individuals with progressive cognitive decline, targeting cell surface proteins such as the N-methyl-D-aspartate receptor, gamma-aminobutyric acid B receptor, alpha-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid receptor, leucine-rich glioma inactivated protein 1, dipeptidyl-peptidase protein-like 6, vesicular glutamate transporter 2 (20), potassium voltage-gated channel subfamily A member 2 (21), and transcobalamin receptor (22–24). The accumulation of these clinical insights has led to the development of the disease concept termed “neural autoantibodies-associated dementia (25, 26).” However, there is an overlap in the neural autoantibody profiles between autoimmune dementia and neurodegenerative dementias like FTD and DLB, necessitating further research to clarify disease specificity (22).
AD, one of the most well-known forms of dementia, is characterized by the accumulation of amyloid plaques and neurofibrillary tangles in the brain (27). Autoantibodies targeting amyloid-β (Aβ), tau, neurotransmitters, and microglia have been reported in AD patients (28, 29). Specifically, autoantibodies against Aβ are decreased in AD patients (30, 31), suggesting a protective role against Aβ toxicity (32, 33), in line with clinical efficacy of lecanemab, a humanized monoclonal antibody targeting Aβ soluble protofibrils (34). Additionally, increased levels of autoantibodies against glutamate (35), oxidized low-density lipoproteins (36), glial markers such as GFAP and S100B (37), and receptors for advanced glycosylation end products have been observed in AD patients’ serum or cerebrospinal fluid (38). DLB is another progressive neurodegenerative disorder characterized by the presence of Lewy bodies—abnormal aggregates of the protein alpha-synuclein—in the brain (39). Autoantibodies against alpha-synuclein, Aβ, myelin oligodendrocyte glycoprotein, myelin basic protein, S100B, and Rho-GTPase-activating protein 26 have been identified in some DLB patients (40–42). Autoantibodies have been detected even in patients with mild cognitive impairment (MCI), indicating a potential role in disease progression (43–45). Despite the discovery of autoantibodies related to various forms of dementia pathology, further research is needed to assess their potential as diagnostic or prognostic biomarkers and their utility in developing effective immunotherapies for dementia (32).
One promising approach is the use of protein microarrays for autoantibody profiling, which could help identify novel autoantibodies for diagnosing and monitoring MCI and dementia (44, 45). In this pilot study, we utilized a proteome-wide autoantibody screening (PWAbS) technique employing wet protein arrays (WPAs) displaying more than 13,000 human proteins (46, 47). This method has previously been used to develop multiplex measurements for disease-related autoantibodies (48, 49), identify clinically relevant novel autoantibodies (50–53), and investigate epitope spreading during disease progression (54). We have successfully applied this technique to a variety of inflammatory disorders, including systemic sclerosis (52), and identified autoantibodies to membranous antigens like G protein-coupled receptors (GPCRs) using machine learning approaches (53). In this study, we applied PWAbS to serum samples from patients with AD or DLB and age-matched cognitively normal individuals (CNIs) to elucidate the autoantibody landscape in dementia. Our goal was to identify clusters of autoantibodies that may contribute to the pathophysiology of dementia, by integration of artificial intelligence (AI) and omics-based approach. This research aims to uncover novel biomarkers and enhance our understanding of dementia’s pathogenesis.
Materials and methods
Participants
We enrolled 26 dementia participants who were admitted to the Department of Geriatric Medicine, The University of Tokyo Hospital, Tokyo, Japan, for evaluation of cognitive impairment. All participants were diagnosed by experienced geriatricians using DSM-IV criteria for AD (n=18), and Revised 2017 Clinical Diagnostic Criteria for DLB by McKeith et al. (n=8) (39). Nine participants were NCIs who admitted to the Department of Geriatric Medicine, The University of Tokyo Hospital, for other reasons, except acute illness and autoimmune disease. Patients with malignant disorders were excluded. We made precise diagnoses using psychological tests, information from family, laboratory data, brain structural imaging (X-ray computed tomography or nuclear magnetic resonance imaging). We also performed N-isopropyl-p-iodoamphetamine brain perfusion single-photon emission computed tomography (SPECT), metaiodobenzylguanidine scintigraphy, ioflupane dopamine transporter SPECT, amyloid PET, and cerebrospinal fluid (CSF) examination in a subset of participants to confirm biological diagnoses. Clinical metrics included number of comorbidities, Charlson’s Comorbidity Index, Comprehensive Geriatric Assessment-short version (CGA7), Mini Mental State Examination (MMSE), Hasegawa’s Dementia Scale-Revised (HDSR), Barthel Index, Lowton’s Instrumental Activities of Daily Living (IADL) scores, Geriatric Depression Scale 15 (GDS15), and Vitality Index. All procedures were approved by the Ethical Review Board at The University of Tokyo Hospital and The University of Tokyo (approval number 2797). The clinical study guidelines of the University of Tokyo, which conform to the Declaration of Helsinki, were strictly adhered to CNIs, dementia patients and their families. They were provided with detailed information about the study, and all provided written informed consent to participate.
Autoantibody measurement
WPAs were arranged as previously described (48). First, proteins were synthesized in vitro utilizing a wheat germ cell-free system from 13,455 clones of the HuPEX (46). Second, synthesized proteins were plotted onto glass plates (Matsunami Glass, Osaka, Japan) in an array format by the affinity between the GST-tag added to the N-terminus of each protein and glutathione modified on the plates. The WPAs were treated with human serum diluted by 3:1000 in the reaction buffer containing 1x Synthetic block (Invitrogen), phosphate-buffered saline (PBS), and 0.1% Tween 20. Next, the WPAs were washed, and goat anti-Human IgG (H+L) Alexa Flour 647 conjugate (Thermo Fisher Scientific, San Jose, CA, USA) diluted 1000-fold was added to the WPAs and reacted for 1 hour at room temperature. Finally, the WPAs were washed, air-dried, and fluorescent images were acquired using a fluorescence imager (Typhoon FLA 9500, Cytiva, Marlborough, MA, USA). Fluorescence images were analyzed to quantify serum levels of autoantibodies targeting each antigen, following the formula shown below:
AU: arbitrary unit
F autoantigen: fluorescent intensity of autoantigen spot
F negative control: fluorescent intensity of negative control spot
F positive control: fluorescent intensity of positive control spot
Machine learning
We applied supervised machine learning techniques using Python (v3.10.12) with libraries from Scikit-learn and the PyTorch framework to construct classifiers for the diagnosis of dementia based on the autoantibody measurement data. The performance of the classifiers was evaluated with 5-fold cross validation using the “KFold” method from Scikit-learn with “shuffle=True”, using the metrics of area under the receiver operating characteristics curve (ROC-AUC), area under the precision-recall curve, accuracy, precision, recall, and F1-score, with the higher score indicating the better classification performance. Machine learning models from Scikit-learn included simple linear regression, Lasso regression, Ridge regression, logistic regression, support vector machine (SVM), random forest, XGBoost, LightGBM, CatBoost, decision trees, gradient boosting machines and naïve Bayes to conduct binary classification. Hyperparameters of the models were tuned using Optuna (Preferred Networks, Inc., Tokyo, Japan) to ensure optimal performance.
Feature importance scores and feature selection
Linear models such as simple linear regression, Lasso, Ridge, logistic regression, and linear SVM determine feature importance based on the absolute values of their coefficients. In contrast, tree-based models, including decision trees, random forests, XGBoost, LightGBM, CatBoost, and gradient boosting machines, measure feature importance through metrics such as impurity reduction or gain achieved at each split or by counting how frequently a feature is used for splitting. We identified the top 10 features from models that achieved ROC-AUC exceeding 0.96 in the binary classification task (AD vs. the others), evaluated the overlap among these models, and selected autoantibodies consistently highlighted by more than two algorithms for further analyses.
Deep neural network
We developed a deep neural network using PyTorch to classify three types of dementia based on autoantibody-derived features. The detailed architecture and training procedures are described below:
Network Architecture:
● Input Layer: Receives input features derived from autoantibody profiles.
● Hidden Layers: The network includes two fully connected hidden layers. The first hidden layer consists of 8 neurons, and the second hidden layer comprises 4 neurons. Each hidden layer employs the Rectified Linear Unit (ReLU) activation function to introduce non-linearity.
● Output Layer: The final layer contains neurons equal to the number of dementia classes. A softmax activation function is applied during evaluation to convert logits into probability scores for each class.
Training Details:
● Loss Function: CrossEntropyLoss was selected as it effectively handles multi-class classification by combining log-softmax activation with negative log-likelihood loss.
● Optimizer: The Adam optimizer was used with a learning rate set at 0.001, leveraging its adaptive learning rate to facilitate efficient convergence.
● Number of Epochs: Training was conducted for 150 epochs, balancing adequate model learning and avoiding overfitting.
● Mini-Batch Size: A mini-batch size of 16 was employed, with training data shuffled at each epoch to ensure diverse mini-batches and improve generalization.
Evaluation Methodology:
● The performance of the model was evaluated using 3-fold cross-validation, generated using the “KFold” method from Scikit-learn with” shuffle=True”.
● During each fold of the cross-validation process, the model’s performance was continuously monitored through training and validation loss curves. Final evaluations on the independent test sets were performed using confusion matrices, detailed classification reports, ROC curves, and Precision-Recall curves to provide a comprehensive performance analysis.
Statistical analysis
Fisher’s exact test was performed to compare categorical variables. Mann-Whitney U test was performed to compare continuous variables. Spearman correlation test was used for correlation analysis. P values of < 0.05 were considered statistically significant. Data analyses were conducted using R (v4.2.1) and Stata/IC 15 (StataCorp LLC, TX, USA).
Protein functional enrichment analysis
Gene Ontology Analysis using web-based tools targeted the list of the entry clones coding the differentially highlighted autoantigens was performed for gene-list enrichment analysis, gene-disease association analysis, and transcriptional regulatory network analysis with Metascape (55).
Sequence identity analysis
To assess cross-reactivity among proteins that express similar antigen epitopes and are highly correlated, we checked the correlation of the differentially expressed autoantibodies. The corresponding proteins of the highly correlated autoantibodies (Spearman’s r > 0.5) were then aligned with the highly correlated proteins using the Uniprot alignment tool.
Data visualization
Box plots, scatter plots, hierarchical clustering, and correlation matrix were visualized by using R (v4.2.1). Box plots were defined as follows: the middle line corresponds to the median; the lower and upper hinges correspond to the first and third quartiles; the upper whisker extends from the hinge to the largest value no further than 1.5 times the interquartile range (IQR) from the hinge; and the lower whisker extends from the hinge to the smallest value at most 1.5 times the IQR of the hinge.
Results
Demographic and clinical characteristics
Serum samples from 35 subjects, including 18 patients with AD, 8 patients with DLB, and 9 CNIs were served for PWAbS utilizing WPAs. The baseline demographics across the three groups were similar, except that the proportion of females was highest in the AD group and lowest among CNIs (Supplementary Table 1). The proportion of females in the AD, DLB, and CNI groups were 82.4%, 62.5%, and 33.3%. The Hasegawa’s Dementia Scale-Revised (HDSR) scores for the each group were 19.9 ± 5.6, 22.1 ± 5.6, and 27.9 ± 2.0, respectively, while the MMSE scores were 20.2 ± 3.9, 21.1 ± 6.6, and 28.9 ± 1.4.
Sum of autoantibody levels
We defined the sum of autoantibody levels (SAL) as the total serum concentration of all autoantibodies measured in our PWAbS. Although not statistically significant, SAL was higher in patients with AD and DLB compared to CNIs (Figure 1A). This trend persisted across all age groups (Supplementary Figure 1A) and was relatively higher in females than in males (Supplementary Figure 1B).
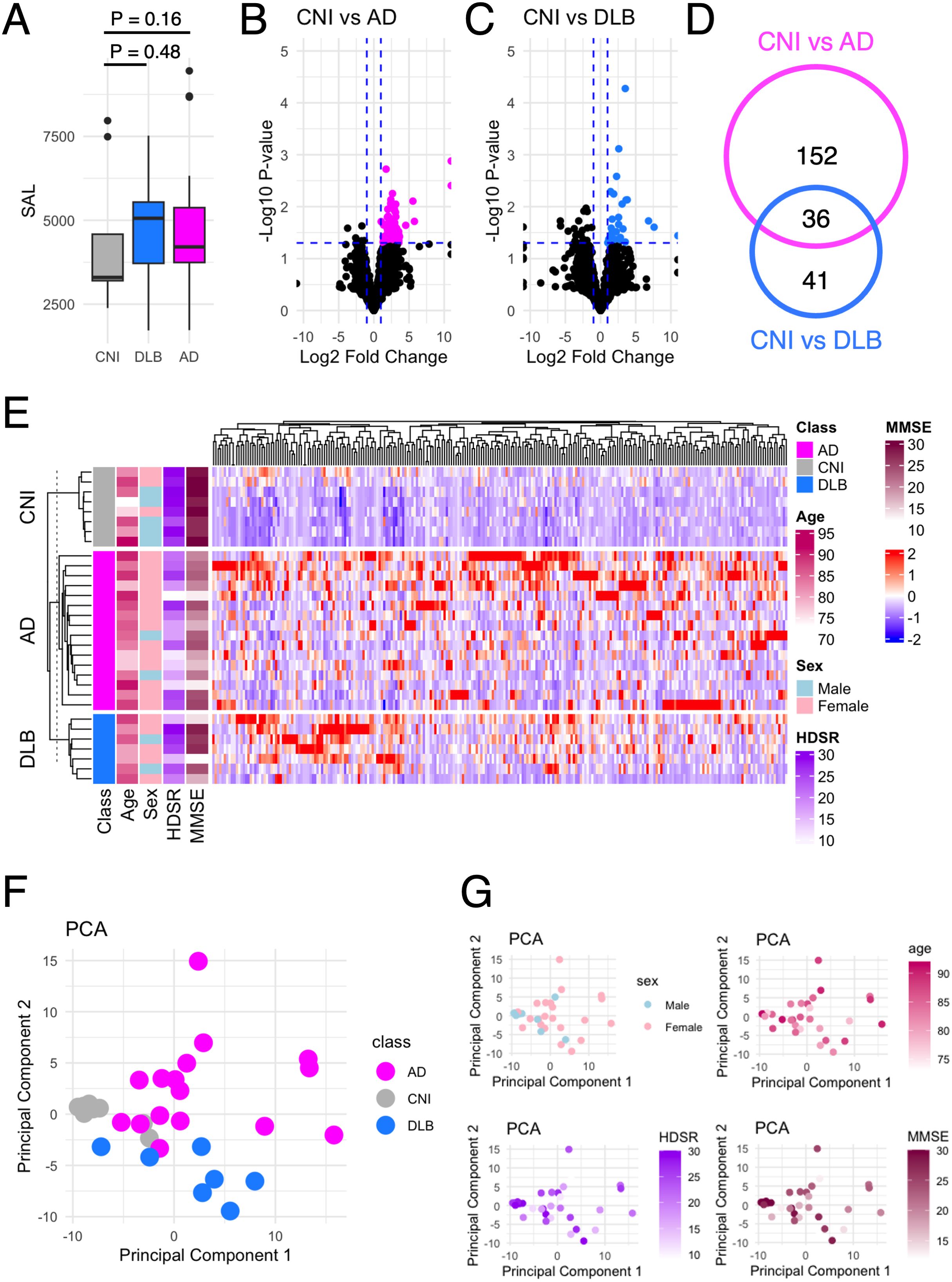
Figure 1. Autoantibodies differentially elevated in dementia. (A) The SAL in AD, DLB, and CNI. (B) Volcano plot that shows autoantibodies differentially elevated in AD compared to NCI. The vertical dash line indicates P = 0.05. The horizontal dash line indicates fold change = ± 2. (C) Volcano plot that shows autoantibodies differentially elevated in DLB compared to NCIs. The vertical dash line indicates P = 0.05. The horizontal dash line indicates fold change = ± 2. (D) Venn diagram that illustrates the inclusion relationship between autoantibodies differentially elevated in AD and/or DLB. (E) Heat map that shows the serum levels of 229 autoantibodies differentially elevated in AD and/or DLB. (F) PCA of 229 autoantibodies differentially elevated in AD and/or DLB. In the scatter plot, individual subjects as points. (G) PCA plots colored by sex, age, HDSR, and MMSE.
Identification of differentially elevated autoantibodies
Next, we focused on identifying autoantibodies with serum levels significantly elevated in AD (Figure 1B) and/or DLB (Figure 1C) compared to CNIs. This analysis revealed 188 autoantibodies elevated in AD and 77 in DLB, with 36 overlapping between the two conditions (Figure 1D), totaling 229 distinct items (Figure 1E). Using these autoantibodies, we performed principal component analysis (PCA), which effectively differentiated AD patients, DLB patients, and CNIs (Figure 1F), regardless of sex, age, or cognitive impairment severity as measured by HDSR and MMSE (Figure 1G).
AI-based 2-class classification
To identify which of the 229 autoantibodies were most strongly associated with disease status, we employed 14 different machine learning frameworks. Logistic regression with normalization or standardization, along with SVM under similar conditions, achieved ROC-AUC exceeding 0.96, indicating near-perfect accuracy in distinguishing AD patients from others (Table 1). We identified the top 10 features from these four models (Figure 2A), assessed their overlap (Figure 2B), and analyzed the serum levels of 12 autoantibodies highlighted in more than two frameworks (Figure 2C).
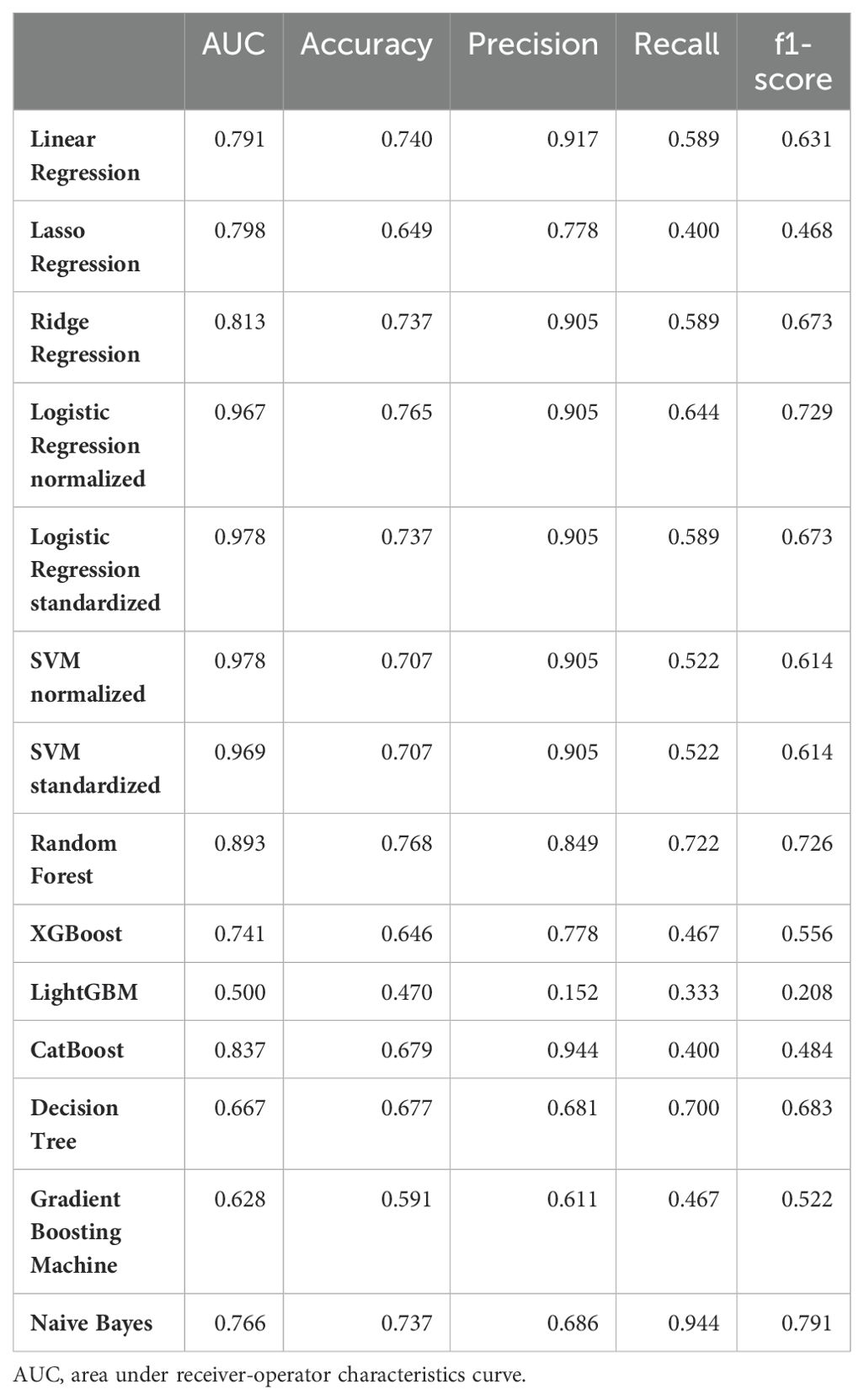
Table 1. Performance of machine learning frameworks for the 2-class classification task.
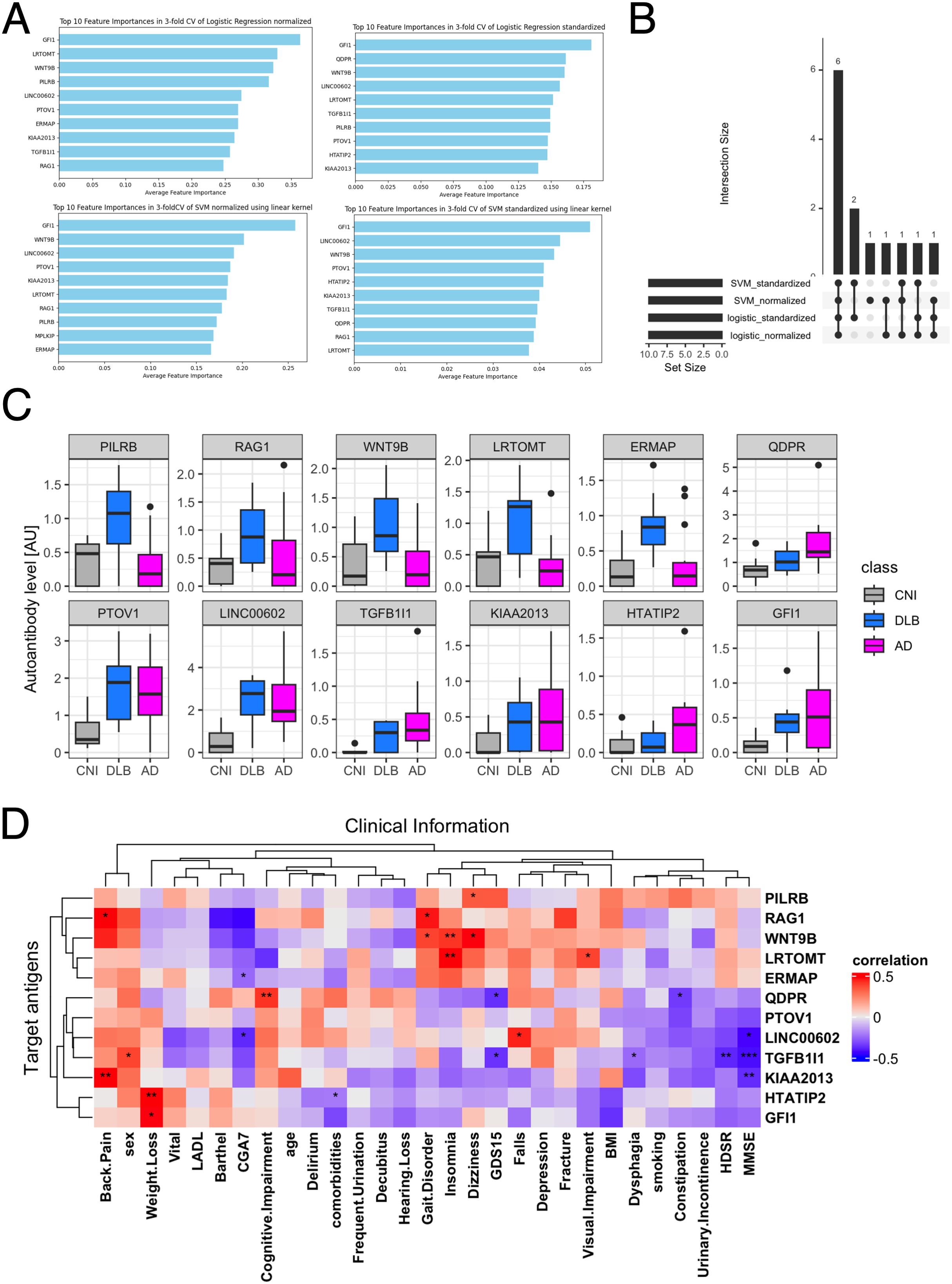
Figure 2. Autoantibodies highlighted in 2-class classification tasks by AI. (A) Autoantibodies that were mostly highlighted according to feature importance by Logistic regression and SVM with standardization or normalization. (B) UpSet plot shows the inclusion relationship of autoantibodies highlighted by the four machine learning frameworks. (C) Box plots describe the serum levels of autoantibodies highlighted by more than two frameworks in AD, DLB, and CNI. (D) Heatmap illustrates correlation between autoantibodies highlighted in machine learning analysis and demographic and clinical characteristics of dementia. The presence of depression and cognitive impairment was initially screened using the Comprehensive Geriatric Assessment 7 (CGA7), which includes the three-item recall test (‘sakura, cat, train’) and the question ‘Do you feel helpless?’. Cognitive impairment was subsequently assessed in more detail using the Hasegawa Dementia Scale-Revised (HDS-R) and the Mini-Mental State Examination (MMSE). Depression severity was further evaluated with the 15-item Geriatric Depression Scale (GDS-15). *P < 0.05, **P < 0.01, ***P < 0.001. P values were calculated by Spearman’s correlation test.
Next, we examined the relationship between these 12 autoantibodies and clinical traits (Figure 2D). This analysis revealed a significant correlation of serum levels of autoantibodies targeting proteins encoded by TGFB1I1 and KIAA2013 with HDSR scores. However, a database search, utilizing the Human Protein Atlas (56), indicated that these two genes are not specifically expressed in the central nervous system (data not shown). Although serum levels of anti-TGFB1I1 antibodies were significantly associated with sex, trends in the distribution of these autoantibodies among three groups were generally similar between both sex (Supplementary Figure 2A). To evaluate cross-reactivity, we performed a correlation analysis on these 12 autoantibodies. Those with moderate to high correlations (Spearman’s r > 0.5) underwent sequence alignment and identity analysis. The correlation matrix (Supplementary Figure 2B) revealed five correlated autoantibodies, and sequence analysis showed that all proteins shared less than 25% identity (Supplementary Figure 2C). Additionally, we investigated the prevalence of these highlighted autoantibodies across a broader spectrum of human disorders using the aUToAntiBody Comprehensive Database (UT-ABCD) (52). Most of these autoantibodies were found to be non-specifically elevated in various pathological conditions (Supplementary Figure 2D).
AI-based 3-class classification
We also explored multi-class classification among AD, DLB, and CNI by training deep neural networks with two hidden layers using the 229-dimensional autoantibody profiles. The optimal number of epochs was determined based on the accuracy and loss trajectories (Figure 3A). The validation loss became consistently lower than the training loss, clearly indicating the absence of overfitting. This approach resulted in high accuracy, with ROC-AUC values reaching up to 0.95 (Figure 3B), as well as high precision and recall (Figure 3C).
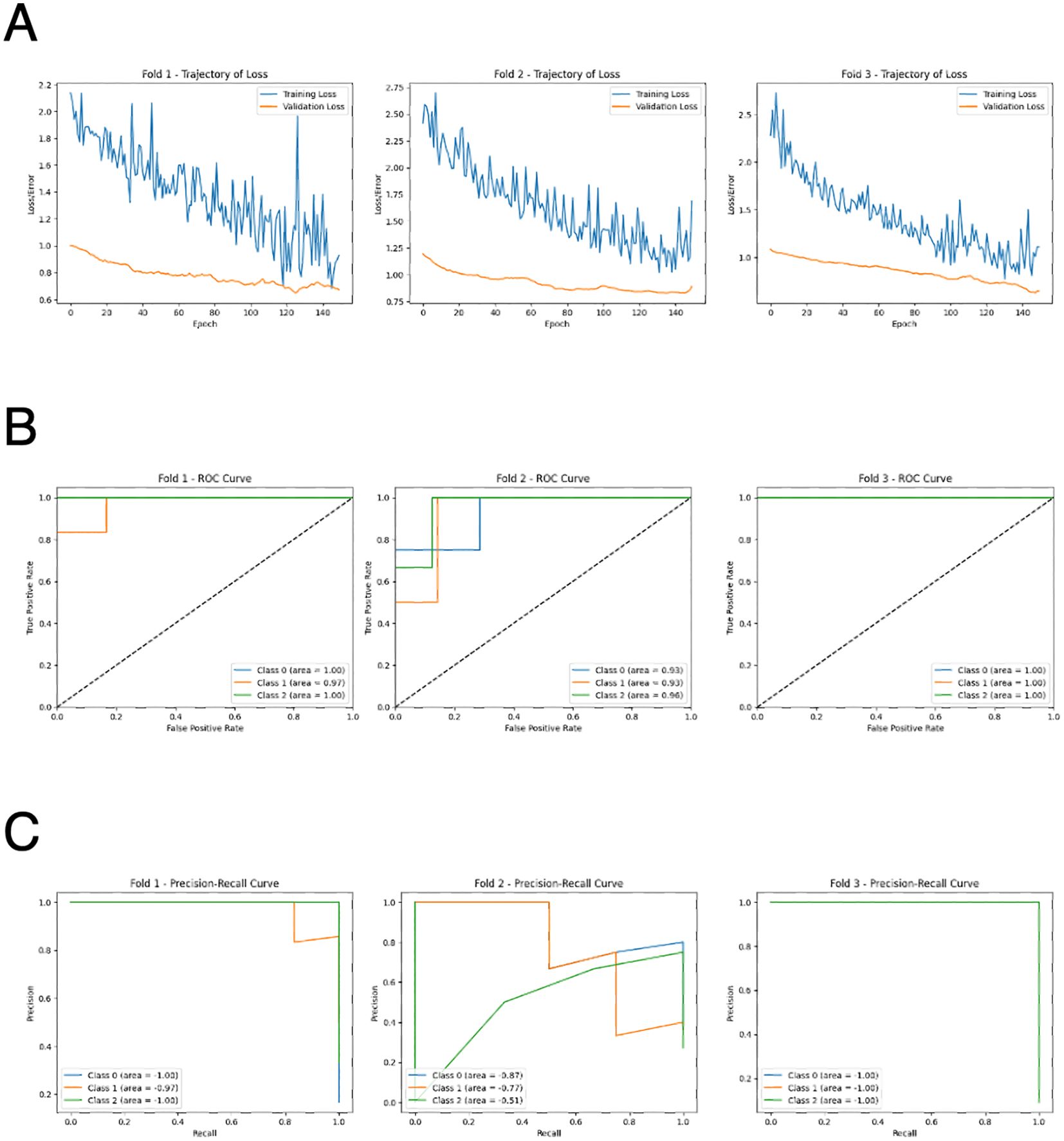
Figure 3. Performance of deep neural network for 3-class classification by AI. (A) Learning curves of the deep neural network model in 3-fold cross validation. (B) ROC curves of the deep neural network model in 3-fold cross validation. Class 1: CNI, class 2: AD, class 3: DLB. (C) Precision-recall curves of the deep neural network model in 3-fold cross validation. Class 1: CNI, class 2: AD, class 3: DLB.
Gene ontology analysis
We aimed to identify autoantibodies with potential pathogenic roles in dementia by conducting gene ontology analysis on the gene lists encoding the 229 autoantigens targeted by differentially elevated autoantibodies in AD and/or DLB (Figure 4). The analysis highlighted the “neuroactive ligand-receptor interaction” pathway in autoantibodies elevated specifically in AD. We also focused on “regulation of lipid metabolic process” highlighted only in DLB, considering recent advances in understanding the role of lipid metabolism in the pathogenesis of DLB, including associations with specific lipid species (57), or genetic polymorphisms (58–60), as well as ultrastructural findings derived directly from Lewy bodies (61).
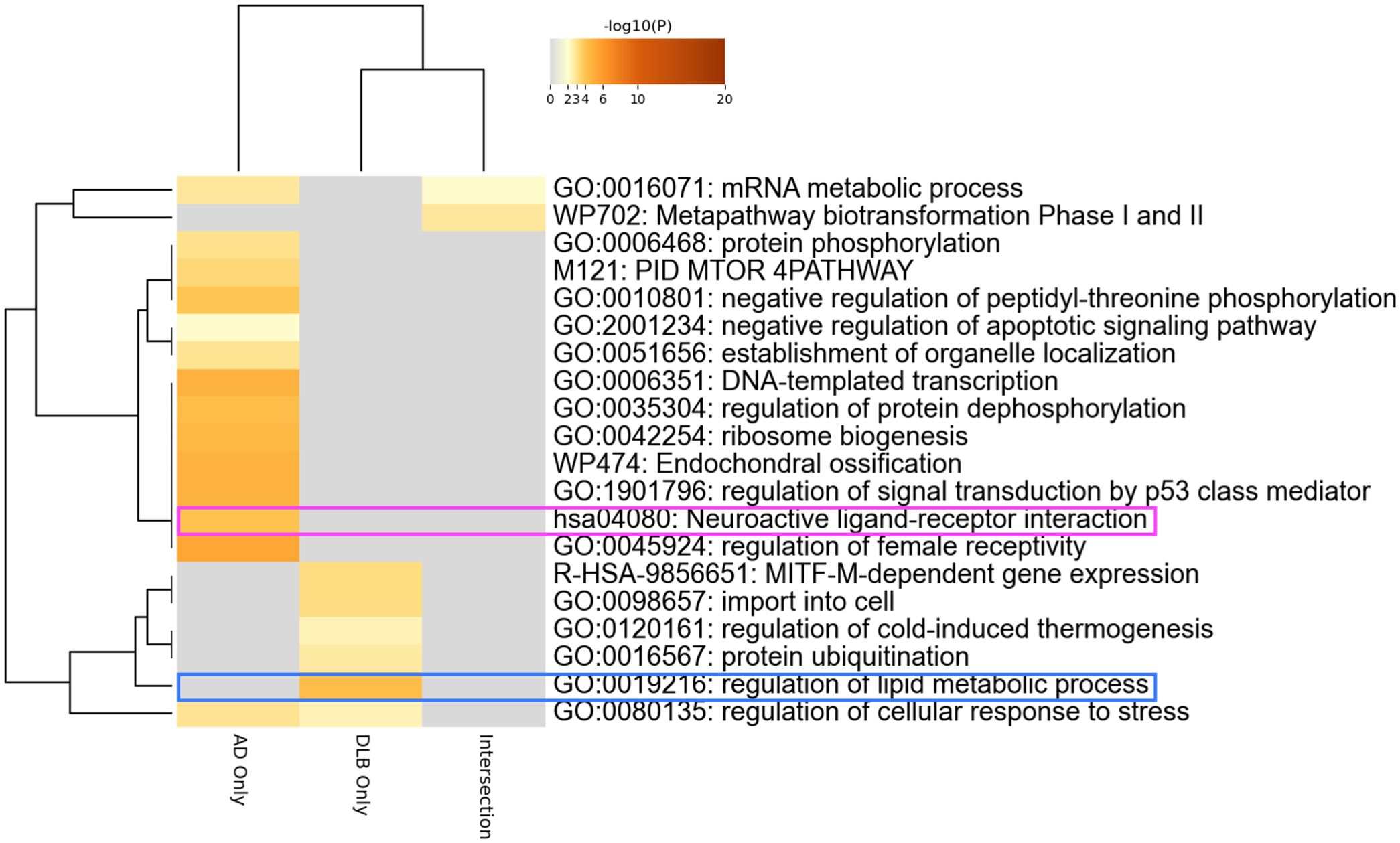
Figure 4. Autoantibodies to neuroactive ligand-receptor interaction-associated proteins. Gene ontology analysis encompassing the genes coding proteins targeted by autoantibodies differentially elevated in AD and/or DLB.
Autoantibodies to neuroactive ligand-receptor interaction-associated proteins
There were exactly 12 autoantibodies associated with neuroactive ligand-receptor interaction, and their serum levels are illustrated in Figure 5A. We examined the relationship between these 12 autoantibodies and clinical traits (Figure 5B), revealing a significant association of the serum levels of autoantibodies targeting neuropeptide B, a protein encoded by NPB, with female sex, presence of back pain, and MMSE scores. However, the trend of elevated serum levels of anti-NPB antibody in dementia was observed in both sex (Supplementary Figure 3A). There was no obvious cross-reactivity among the autoantibodies (Supplementary Figures 3B, C) and showed no disease specificity (Supplementary Figure 3D). To further investigate the potential of anti-NPB antibody to play a role in the pathogenesis of AD, we examined the correlation between serum levels of the autoantibody and all the subscales of MMSE (Supplementary Figure 4). As a result, there was statistically significant correlation in memory-related items (“Registration” and “Recall”). In line with this, a database search indicated that NPB is expressed in the CNS (Supplementary Figure 5A), including the hippocampus (Supplementary Figure 5B). The highest expression was reported in oligodendrocytes (Supplementary Figure 5C).
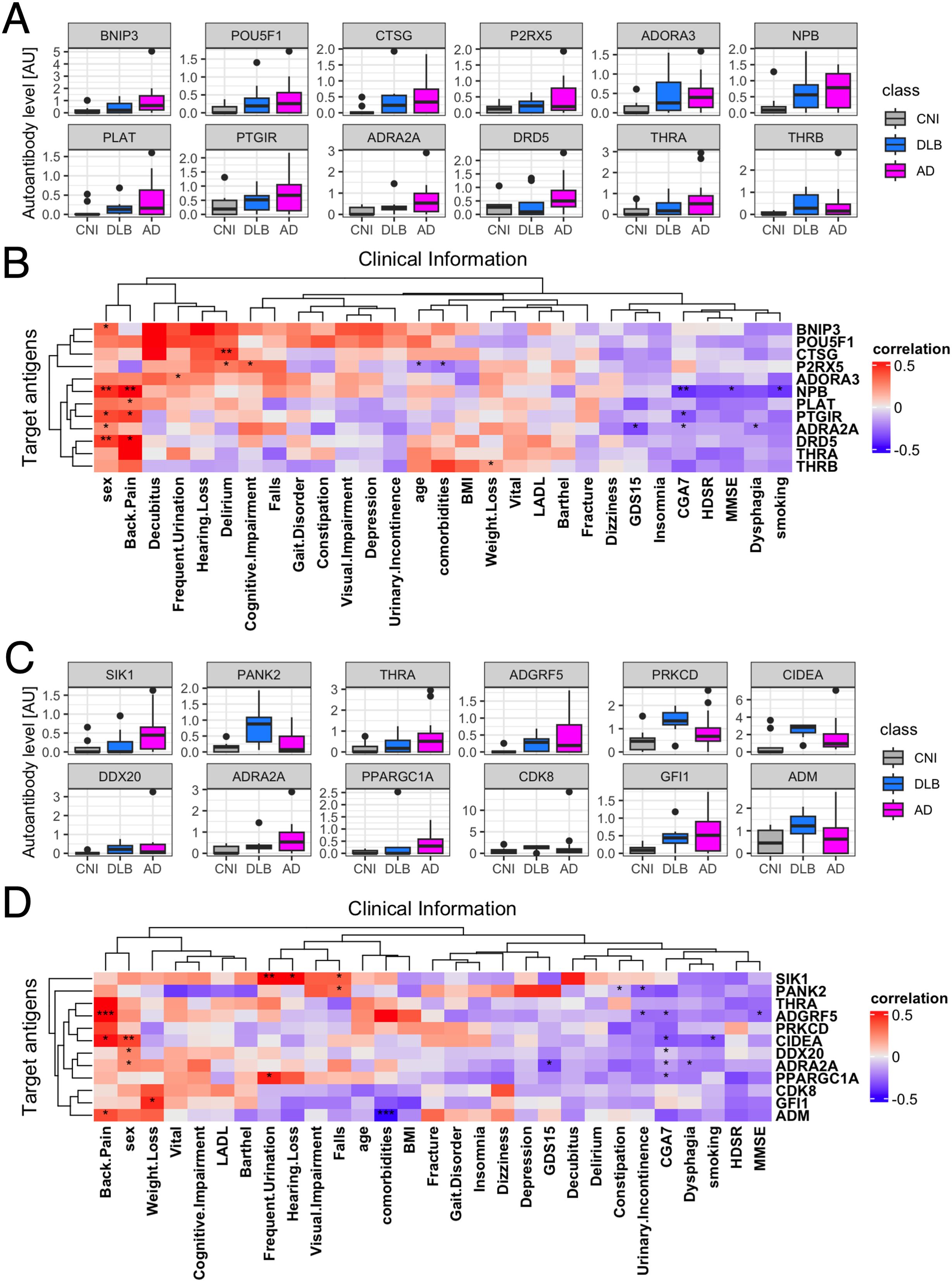
Figure 5. Correlation between autoantibodies highlighted in gene ontology analysis and clinical traits of dementia. (A) Box plots describe the serum levels of autoantibodies to neuroactive ligand-receptor interaction-associated proteins. (B) Heatmap illustrates correlation between autoantibodies to neuroactive ligand-receptor interaction-associated proteins and demographic and clinical characteristics of dementia. (C) Box plots describe the serum levels of autoantibodies to regulation of lipid metabolic process-associated proteins. (D) Heatmap illustrates correlation between autoantibodies to regulation of lipid metabolic process-associated proteins and demographic and clinical characteristics of dementia. The presence of depression and cognitive impairment was initially screened using the Comprehensive Geriatric Assessment 7 (CGA7), which includes the three-item recall test (‘sakura, cat, train’) and the question ‘Do you feel helpless?’. Cognitive impairment was subsequently assessed in more detail using the Hasegawa Dementia Scale-Revised (HDS-R) and the Mini-Mental State Examination (MMSE). Depression severity was further evaluated with the 15-item Geriatric Depression Scale (GDS-15). *P < 0.05, **P < 0.01, ***P < 0.001. P values were calculated by Spearman’s correlation test.
Autoantibodies to lipid metabolism-associated proteins
Finally, we focused on all the autoantibodies targeting lipid metabolism-associated proteins, whose serum levels are illustrated in Figure 5C. We examined the relationship between these 12 autoantibodies and clinical traits (Figure 5D). This analysis revealed a significant association of the serum levels of autoantibodies targeting Adhesion G Protein-Coupled Receptor F5 (ADGRF5) encoded by ADGRF5 with presence of back pain, lower Comprehensive Geriatric Assessment 7 (CGA7) scores, and lower MMSE scores, especially in “Registration” and “Repetition” subscales (Supplementary Figure 6). There was no big difference between both sex (Supplementary Figure 7A), cross-reactivity, nor disease specificity. (Supplementary Figures 7B–D). A database search indicated that the expression of ADGRF5 is ubiquitous across various human tissues (Supplementary Figure 8A), including the CNS (Supplementary Figure 8B), predominantly in microglial cells (Supplementary Figure 8C).
Age and sex-adjusted simple linear regression analysis
Finally, we conducted linear regression analyses to explore potential correlations between MMSE scores, its subscales, and serum anti-NPB and anti-ADGRP5 Ab levels (Supplementary Table 2). The univariate analysis identified statistically significant correlations between MMSE scores, sex, and serum anti-NPB Ab levels, whereas no significant association was found with anti-ADGRP5 Ab levels. However, upon performing multivariate regression analyses adjusting for age, sex, and antibody levels, neither anti-NPB nor anti-ADGRP5 Ab levels remained significantly correlated with MMSE total scores. Notably, multivariate analyses did confirm significant associations between serum anti-ADGRP5 Ab levels and the MMSE subscales “Orientation_Space” and “Recall.
Discussion
In this study, we utilized our proprietary PWAbS technique to analyze serum samples from patients with AD, DLB, and CNIs. Our results showed an increase in the overall levels of autoantibodies in AD and DLB patients compared to CNIs (Figure 1A). We identified 229 autoantibodies that were differentially elevated in AD and/or DLB (Figure 1D), effectively distinguishing between AD, DLB, and CNI groups (Figure 1F). Machine learning applied to these 229 autoantibodies demonstrated high accuracy in differentiating AD patients from others (Table 1), and even achieved success in multi-class classification (Figure 3). Gene ontology analysis highlighted autoantibodies targeting neuroactive ligands and receptors in AD, including anti-NPB antibody, as well as lipid metabolism-associated proteins in DLB, such as anti-ADGRF5 antibody (Figure 4). Both of anti-NPB and anti-ADGRF5 autoantibodies showed significant correlation with total MMSE scores (Figures 5B, D) and memory-related subscale scores (Supplementary Figures 4, 6). Considering the expression of NPB and ADGRF5 in the central nervous system (Supplementary Figures 5, 8), these findings suggest that autoantibodies targeting NPB or ADGRF5 may contribute to the pathogenesis of dementia. Our results underscore the potential of our systems-based approach in developing novel diagnostic tools and propose a new research strategy to explore the autoimmune aspects of dementia.
A key highlight of our analysis is the ability of AI integrated with our multiplex autoantibody measurement to achieve near-perfect accuracy in classifying AD versus other groups (Table 1) and even in multi-class classification tasks (AD, DLB, and CNI; Figure 3). This concept has already been demonstrated in other autoimmune and malignant disorders (52, 53), and is partially available commercially as the Autoantibody Array Assay (A-Cube) (49). Given that blood tests are less invasive than other procedures like cerebrospinal fluid collection and radiological imaging studies and can be conducted without causing undue concern to the patient about suspected cognitive impairment, multiplex measurement of serum autoantibodies using WPAs and AI-based interpretation represents a promising strategy for diagnosing dementia and its subtypes.
In our study, we implemented a comprehensive strategy to mitigate the risk of overfitting that arises from the combination of a small sample size and a high-dimensional feature space. Specifically, we employed a robust k-fold cross-validation framework, ensuring that every subject contributed to both training and evaluation phases, thereby stabilizing performance estimates. Moreover, the use of regularized models such as Lasso and Ridge regression inherently facilitated feature selection by shrinking the coefficients of less informative autoantibodies, effectively reducing dimensionality. Furthermore, decision tree-based models such as Random Forest, XGBoost, LightGBM, CatBoost, and Gradient Boosting Machine inherently possess the capability to perform dimensionality reduction, which can help prevent overfitting. Due to their robustness to data redundancy, they are less likely to capture noise, and their convergence is faster owing to inductive bias. Hyperparameter optimization using Optuna further balanced model complexity and performance, while evaluation of feature importance across multiple models revealed a significant overlap in key autoantibody biomarkers, underscoring the robustness of our findings. Notably, the deep neural network demonstrated stable loss function curves during training and validation, indicating little signs of overfitting and reinforcing the reliability of our methodological approach.
The NPB gene encodes neuropeptide B, a short biologically active peptide that acts as an agonist for GPCRs known as neuropeptide B/W receptors 1 (NPBWR1) and 2 (NPBWR2) (62). Neuropeptide B is believed to play roles in regulating feeding, the neuroendocrine system, memory, learning, and the pain pathway (63). Research by Nagata-Kuroiwa R et al. on NPBWR1 knockout mice revealed increased autonomic and neuroendocrine responses to physical stress and abnormalities in contextual fear conditioning, suggesting a role for NPBWR1 in stress vulnerability and fear memory (64). Histological and electrophysiological studies indicate that NPBWR1 acts as an inhibitory regulator on a subpopulation of GABAergic neurons in the lateral division of the central nucleus of the amygdala, terminating stress responses. Additionally, Watanabe N et al. demonstrated that a single nucleotide polymorphism in NPBWR1, associated with impaired molecular function, affected valence evaluation and dominance ratings in response to seeing angry faces in humans, suggesting NPBWR1’s involvement in social interaction (65). These insights highlight the potential role of autoantibodies affecting the NPB-NPBWR1 signaling system in social behavior, suggesting its potential contribution to the clinical manifestations of AD, particularly its behavioral and psychological symptoms.
Our study also revealed a strong association between serum anti-NPB antibody levels and the presence of back pain, likely due to the role of NPB-NPBWR1 signaling in pain transmission. NPB knockout mice exhibit different responses to pain; they show hyperalgesia to acute inflammatory pain but not to thermal or chemical pain (66). Intrathecal administration of NPB reduced mechanical allodynia via activation of NPBWR1 receptors without affecting thermal hyperalgesia (67). These effects were not inhibited by naloxone, an opioid receptor antagonist, indicating the involvement of a non-opioid analgesic pathway, possibly related to myelin-forming Schwann cells, which express low levels of NPBWR1 under physiological conditions but much higher levels in patients with inflammatory neuropathies. Thus, anti-NPB antibodies may play a role in modulating nociceptive transmission.
ADGRF5, a member of the adhesion GPCR (aGPCR) family, which is the second largest GPCR subfamily, has recently garnered attention for its biological functions, disease relevance, and potential as a drug target (68). Predominantly expressed in the lung and kidney, ADGRF5 may play a crucial role in regulating surfactant protein synthesis acid-base balance in these organs (69–71). DiBlasi et al. identified a single nucleotide polymorphism in the ADGRF5 gene linked to an increased risk of suicide (72), suggesting its psychiatric role. Additionally, Kaur et al. found that plasma levels of ADGRF5 are associated with the APOE genotype (73), a known risk factor for DLB and AD (59, 60). Elevated levels of anti-ADGRF5 antibodies correlated with global geriatric function scores assessed by CGA7 (Figure 5D), and the fact that ADGRF5 expression is not exclusive to the CNS (Supplementary Figure 8), may reflect systemic aspects of DLB affecting multiple organs (74).
It is important to note that not all patients had anti-NPB nor anti-ADGRF5 antibodies, and their serum levels in AD were not specific to the condition (Supplementary Figures 3D, 7D). This suggests that while the presence of these autoantibodies may not explain the entire pathogenesis of dementia, they could influence disease manifestation and progression as bystanders. Further investigation is needed to clarify the role of anti-NPB and anti-ADGRF5 antibodies in the pathophysiology, including functional assays to assess the effects of these antibodies on neurons or glial cells, passive immune challenge in AD animal models by administering anti-NPB or anti-ADGRF5 antibodies, and active immunization of animals with NPB or ADGRF5 antigens.
Our study has several strengths. First, by including multiple types of dementia (AD and DLB), as well as CNIs, we were able to identify autoantibodies that are differentially elevated in each condition and develop machine learning methodologies for distinguishing different types of dementia in a relatively non-invasive way. Second, the use of a wheat-germ in vitro protein synthesis system and the manipulation technique for WPAs allowed for high-throughput expression of a wide range of human proteins, including soluble proteins, on a single platform (46, 47, 75). This enabled our autoantibody measurement to cover an almost proteome-wide range of antigens, allowing the application of omics-based bioinformatics approaches to interpret the data. Third, integration of AI and omics-based approach allowed us to conduct an unbiased and holistic investigation, resulting in novel discoveries.
A major limitation of our study is the demographic differences among the human subjects, particularly in terms of sex (Supplementary Table 1). Moreover, the sample size was modest, lacked external validation, and was cross-sectional. The absence of significant correlations in the multivariate regression analyses may reflect insufficient statistical power due to the small sample size of our study (Supplementary Table 2). Additionally, demographic factors, particularly sex, may introduce confounding effects, complicating the interpretation of serum Ab levels as independent predictors of cognitive impairment. Additionally, biological diagnosis of AD, as opposed to symptomatic diagnosis, was not confirmed in all recruited cases using biomarkers reflecting disease-specific biological processes, such as amyloid PET and CSF examinations, an approach increasingly emphasized in recent advances in AD diagnosis (76, 77). Future studies should target larger, more demographically balanced patient groups with a wider range of dementia types, such as VD and FTD, confirmed by precise biological diagnosis. Recruiting longitudinal specimens and data from elderly individuals before and after the onset of MCI in prospective population-based cohorts would be a valuable challenge to explore the causal relationship between autoantibodies and dementia pathogenesis.
Data availability statement
The raw data supporting the conclusions of this article will be made available by the authors, upon reasonable request.
Ethics statement
The studies involving humans were approved by Ethical Review Board at The University of Tokyo Hospital and The University of Tokyo. The studies were conducted in accordance with the local legislation and institutional requirements. The participants provided their written informed consent to participate in this study.
Author contributions
KM: Data curation, Formal analysis, Investigation, Methodology, Project administration, Resources, Visualization, Writing – original draft, Writing – review & editing. YU: Investigation, Project administration, Resources, Supervision, Writing – review & editing. KI: Investigation, Methodology, Software, Writing – review & editing. MM: Resources, Writing – review & editing. MY: Resources, Writing – review & editing. MI: Resources, Writing – review & editing. SO: Resources, Supervision, Writing – review & editing. MA: Resources, Supervision, Writing – review & editing. SS: Supervision, Writing – review & editing. AY: Conceptualization, Funding acquisition, Project administration, Supervision, Writing – review & editing.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. JSPS KAKENHI 25HP8021.
Acknowledgments
We thank Ms. Maiko Enomoto and her colleagues for their secretarial work. We appreciate K. Yamaguchi, T. Okumura, C. Ono, A. Sato, A. Miya, and N. Goshima from ProteoBridge Corporation for preparing the WPAs. We also acknowledge R. Uchino, Y. Murakami, and H. Matsunaka from TOKIWA Pharmaceuticals Co. Ltd. for providing technical assistance with autoantibody measurement.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fimmu.2025.1537659/full#supplementary-material
Supplementary Figure 1 | Sum of autoantibody levels by age and sex. (A) Box plots show SAL by age groups. (B) Box plots show SAL by sex.
Supplementary Figure 2 | Additional information for autoantibodies highlighted in 2-class classification tasks by AI. (A) Box plots describe the serum levels of autoantibodies highlighted in 2-class classification tasks by sex. (B) A correlation matrix of the autoantibodies highlighted in 2-class classification tasks using Spearman’s correlation. Only statistically significant pairs (P < 0.05) are shown. (C) Identity matrix, generated from aligning the corresponding protein sequences of the highly correlated autoantibodies (Spearman’s r > 0.5). (D) Box plots describe the serum levels of autoantibodies highlighted in 2-class classification tasks in COVID-19, atopic dermatitis, anti-neutrophil cytoplasmic antibody-associated vasculitis, systemic lupus erythematosus, systemic sclerosis, and healthy controls. The data derives from the UT-ABCD.
Supplementary Figure 3 | Additional information for autoantibodies to neuroactive ligand-receptor interaction-associated proteins. (A) Box plots describe the serum levels of autoantibodies to neuroactive ligand-receptor interaction-associated proteins by sex. (B) A correlation matrix of the autoantibodies to neuroactive ligand-receptor interaction-associated proteins using Spearman’s correlation. Only statistically significant pairs (P < 0.05) are shown. (C) Identity matrix, generated from aligning the corresponding protein sequences of the highly correlated autoantibodies (Spearman’s r > 0.5). (D) Box plots describe the serum levels of autoantibodies to neuroactive ligand-receptor interaction-associated proteins in COVID-19, atopic dermatitis, anti-neutrophil cytoplasmic antibody-associated vasculitis, systemic lupus erythematosus, systemic sclerosis, and healthy controls. The data derives from the UT-ABCD.
Supplementary Figure 4 | Correlation between serum levels of anti-NPB antibodies and MMSE subscales.
Supplementary Figure 5 | Expression of the NPB gene in human tissues and single cells. (A) Expression of NPB in multiple human tissues measured by bulk RNA-sequencing from the Human Protein Atlas. (B) Expression of NPB in the CNS from the Human Protein Atlas. (C) Expression of NPB in the CNS evaluated by single-cell RNA-sequencing from the Human Protein Atlas.
Supplementary Figure 6 | Correlation between serum levels of anti-ADGRF5 antibodies and MMSE subscales.
Supplementary Figure 7 | Additional information for autoantibodies to regulation of lipid metabolic process-associated proteins. (A) Box plots describe the serum levels of autoantibodies to regulation of lipid metabolic process-associated proteins by sex. (B) A correlation matrix of the autoantibodies to regulation of lipid metabolic process-associated proteins using Spearman’s correlation. Only statistically significant pairs (P < 0.05) are shown. (C) Identity matrix, generated from aligning the corresponding protein sequences of the highly correlated autoantibodies (Spearman’s r > 0.5). (D) Box plots describe the serum levels of autoantibodies to regulation of lipid metabolic process-associated proteins in COVID-19, atopic dermatitis, anti-neutrophil cytoplasmic antibody-associated vasculitis, systemic lupus erythematosus, systemic sclerosis, and healthy controls. The data derives from the UT-ABCD.
Supplementary Figure 8 | Expression of the ADGRF5 gene in human tissues and single cells. (A) Expression of ADGRF5 in multiple human tissues measured by bulk RNA-sequencing from the Human Protein Atlas. (B) Expression of ADGRF5 in the CNS from the Human Protein Atlas. (C) Expression of ADGRF5 in the CNS evaluated by single-cell RNA-sequencing from the Human Protein Atlas.
References
1. Umeda-Kameyama Y, Mori T, Wada-Isoe K, Kikuchi T, Kojima S, Kagimura T, et al. Development of a novel convenient Alzheimer’s disease assessment scale, the ABC Dementia Scale, using item response theory. Geriatr Gerontol Int. (2019) 19:18–23. doi: 10.1111/ggi.13552
2. Wada-Isoe K, Kikuchi T, Umeda-Kameyama Y, Mori T, Akishita M, Nakamura Y. ABC dementia scale classifies Alzheimer’s disease patients into subgroups characterized by activities of daily living, behavioral and psychological symptoms of dementia, and cognitive function. J Alzheimer’s Dis. (2020) 73:383–92. doi: 10.3233/JAD-190767
3. Umeda-Kameyama Y, Kameyama M, Tanaka T, Son B-K, Kojima T, Fukasawa M, et al. Screening of Alzheimer’s disease by facial complexion using artificial intelligence. Aging (Albany NY). (2021) 13:1765–72. doi: 10.18632/aging.202545
4. Wilson DM, Cookson MR, Van Den Bosch L, Zetterberg H, Holtzman DM, Dewachter I. Hallmarks of neurodegenerative diseases. Cell. (2023) 186:693–714. doi: 10.1016/j.cell.2022.12.032
5. Yokoyama JS, Wang Y, Schork AJ, Thompson WK, Karch CM, Cruchaga C, et al. Association between genetic traits for immune-mediated diseases and Alzheimer disease. JAMA Neurol. (2016) 73:691–7. doi: 10.1001/jamaneurol.2016.0150
6. Witoelar A, Jansen IE, Wang Y, Desikan RS, Gibbs JR, Blauwendraat C, et al. Genome-wide pleiotropy between Parkinson disease and autoimmune diseases. JAMA Neurol. (2017) 74:780–92. doi: 10.1001/jamaneurol.2017.0469
7. Ferrari R, Hernandez DG, Nalls MA, Rohrer JD, Ramasamy A, Kwok JBJ, et al. Frontotemporal dementia and its subtypes: a genome-wide association study. Lancet Neurol. (2014) 13:686–99. doi: 10.1016/S1474-4422(14)70065-1
8. Archelos JJ, Hartung HP. Pathogenetic role of autoantibodies in neurological diseases. Trends Neurosci. (2000) 23:317–27. doi: 10.1016/S0166-2236(00)01575-7
9. D’Andrea MR. Add Alzheimer’s disease to the list of autoimmune diseases. Med Hypotheses. (2005) 64:458–63. doi: 10.1016/j.mehy.2004.08.024
10. Sardi F, Fassina L, Venturini L, Inguscio M, Guerriero F, Rolfo E, et al. Alzheimer’s disease, autoimmunity and inflammation. The good, the bad and the ugly. Autoimmun Rev. (2011) 11:149–53. doi: 10.1016/j.autrev.2011.09.005
11. Elkon K, Casali P. Nature and functions of autoantibodies. Nat Clin Pract Rheumatol. (2008) 4:491–8. doi: 10.1038/ncprheum0895
12. Chatanaka MK, Sohaei D, Diamandis EP, Prassas I. Beyond the amyloid hypothesis: how current research implicates autoimmunity in Alzheimer’s disease pathogenesis. Crit Rev Clin Lab Sci. (2023) 60:398–426. doi: 10.1080/10408363.2023.2187342
13. Wu J, Li L. Autoantibodies in Alzheimer’s disease: potential biomarkers, pathogenic roles, and therapeutic implications. J Biomed Res. (2016) 30:361–72. doi: 10.7555/JBR.30.20150131
14. Gibson LL, McKeever A, Cullen AE, Nicholson TR, Aarsland D, Zandi MS, et al. Neuronal surface autoantibodies in dementia: a systematic review and meta-analysis. J Neurol. (2021) 268:2769–79. doi: 10.1007/s00415-020-09825-0
15. Terryberry JW, Thor G, Peter JB. Autoantibodies in neurodegenerative diseases: antigen-specific frequencies and intrathecal analysis. Neurobiol Aging. (1998) 19:205–16. doi: 10.1016/S0197-4580(98)00049-9
16. Arshad F, Varghese F, Paplikar A, Gangadhar Y, Ramakrishnan S, Chaudhuri JR, et al. Role of autoantibodies in neurodegenerative dementia: an emerging association. Dement Geriatr Cogn Disord. (2021) 50:153–60. doi: 10.1159/000517238
17. Nagele E, Han M, Demarshall C, Belinka B, Nagele R. Diagnosis of Alzheimer’s disease based on disease-specific autoantibody profiles in human sera. PLoS One. (2011) 6:e23112. doi: 10.1371/journal.pone.0023112
18. Reddy MM, Wilson R, Wilson J, Connell S, Gocke A, Hynan L, et al. Identification of candidate IgG biomarkers for Alzheimer’s disease via combinatorial library screening. Cell. (2011) 144:132–42. doi: 10.1016/j.cell.2010.11.054
19. Flanagan EP, McKeon A, Lennon VA, Boeve BF, Trenerry MR, Tan KM, et al. Autoimmune dementia: clinical course and predictors of immunotherapy response. Mayo Clin Proc. (2010) 85:881–97. doi: 10.4065/mcp.2010.0326
20. Hansen N, Teegen B, Hirschel S, Wiltfang J, Schott BH, Bartels C, et al. Case report: Mixed dementia associated with autoantibodies targeting the vesicular glutamate transporter 2. Front Psychiatry. (2023) 14. doi: 10.3389/fpsyt.2023.1227824
21. Hansen N, Teegen B, Hirschel S, Fox J, Fitzner D, Wiltfang J, et al. Longitudinally persisting KCNA2-autoantibodies in mild amnestic dementia with Alzheimer´s pathology – Report and literature review. Behav Brain Res. (2025) 482:115437. doi: 10.1016/j.bbr.2025.115437
22. Hansen N. Current nosology of neural autoantibody-associated dementia. Front Aging Neurosci. (2021) 13:711195. doi: 10.3389/fnagi.2021.711195
23. Banks SA, Sechi E, Flanagan EP. Autoimmune encephalopathies presenting as dementia of subacute onset and rapid progression. Ther Adv Neurol Disord. (2021) 14:1756286421998906. doi: 10.1177/1756286421998906
24. Pluvinage JV, Ngo T, Fouassier C, McDonagh M, Holmes BB, Bartley CM, et al. Transcobalamin receptor antibodies in autoimmune vitamin B12 central deficiency. Sci Transl Med. (2024) 16:eadl3758. doi: 10.1126/scitranslmed.adl3758
25. Hansen N, Juhl AL, Grenzer IM, Hirschel S, Teegen B, Fitzner D, et al. Cerebrospinal fluid total tau protein correlates with longitudinal, progressing cognitive dysfunction in anti-neural autoantibody-associated dementia and Alzheimer’s dementia: A case–control study. Front Immunol. (2022) 13. doi: 10.3389/fimmu.2022.837376
26. Hansen N, Hirschel S, Teegen B, Wiltfang J, Malchow B. Preserved visuoconstruction in patients with Alzheimer’s pathology and anti-neural autoantibodies: A case control study. Front Dement. (2022) 1. doi: 10.3389/frdem.2022.975851
27. Scheltens P, Blennow K, Breteler MMB, de Strooper B, Frisoni GB, Salloway S, et al. Alzheimer’s disease. Lancet (London England). (2016) 388:505–17. doi: 10.1016/S0140-6736(15)01124-1
28. Henstridge CM, Hyman BT, Spires-Jones TL. Beyond the neuron-cellular interactions early in Alzheimer disease pathogenesis. Nat Rev Neurosci. (2019) 20:94–108. doi: 10.1038/s41583-018-0113-1
29. Colasanti T, Barbati C, Rosano G, Malorni W, Ortona E. Autoantibodies in patients with Alzheimer’s disease: pathogenetic role and potential use as biomarkers of disease progression. Autoimmun Rev. (2010) 9:807–11. doi: 10.1016/j.autrev.2010.07.008
30. Du Y, Dodel R, Hampel H, Buerger K, Lin S, Eastwood B, et al. Reduced levels of amyloid beta-peptide antibody in Alzheimer disease. Neurology. (2001) 57:801–5. doi: 10.1212/WNL.57.5.801
31. Weksler ME, Relkin N, Turkenich R, LaRusse S, Zhou L, Szabo P, et al. Patients with Alzheimer disease have lower levels of serum anti-amyloid peptide antibodies than healthy elderly individuals. Exp Gerontol. (2002) 37:943–8. doi: 10.1016/S0531-5565(02)00029-3
32. Britschgi M, Olin CE, Johns HT, Takeda-Uchimura Y, LeMieux MC, Rufibach K, et al. Neuroprotective natural antibodies to assemblies of amyloidogenic peptides decrease with normal aging and advancing Alzheimer’s disease. Proc Natl Acad Sci U S A. (2009) 106:12145–50. doi: 10.1073/pnas.0904866106
33. Paul S, Planque S, Nishiyama Y. Immunological origin and functional properties of catalytic autoantibodies to amyloid beta peptide. J Clin Immunol. (2010) 30 Suppl:S43–9. doi: 10.1007/s10875-010-9414-5
34. van Dyck CH, Swanson CJ, Aisen P, Bateman RJ, Chen C, Gee M, et al. Lecanemab in early Alzheimer’s disease. N Engl J Med. (2023) 388:9–21. doi: 10.1056/NEJMoa2212948
35. Davydova TV, Voskresenskaya NI, Fomina VG, Vetrile LA, Doronina OA. Induction of autoantibodies to glutamate in patients with Alzheimer’s disease. Bull Exp Biol Med. (2007) 143:182–3. doi: 10.1007/s10517-007-0044-8
36. Kankaanpää J, Turunen SP, Moilanen V, Hörkkö S, Remes AM. Cerebrospinal fluid antibodies to oxidized LDL are increased in Alzheimer’s disease. Neurobiol Dis. (2009) 33:467–72. doi: 10.1016/j.nbd.2008.12.001
37. Mecocci P, Voskresenskaya NI, Fomina VG, Vetrile LA, Doronina OA. Serum anti-GFAP and anti-S100 autoantibodies in brain aging, Alzheimer’s disease and vascular dementia. J Neuroimmunol. (1995) 57:165–70. doi: 10.1016/0165-5728(94)00180-V
38. Mruthinti S, Buccafusco JJ, Hill WD, Waller JL, Jackson TW, Zamrini EY, et al. Autoimmunity in Alzheimer’s disease: increased levels of circulating IgGs binding Abeta and RAGE peptides. Neurobiol Aging. (2004) 25:1023–32. doi: 10.1016/j.neurobiolaging.2003.11.001
39. McKeith IG, Boeve BF, Dickson DW, Halliday G, Taylor J-P, Weintraub D, et al. Diagnosis and management of dementia with Lewy bodies: Fourth consensus report of the DLB Consortium. Neurology. (2017) 89:88–100. doi: 10.1212/WNL.0000000000004058
40. Maetzler W, Berg D, Synofzik M, Brockmann K, Godau J, Melms A, et al. Autoantibodies against amyloid and glial-derived antigens are increased in serum and cerebrospinal fluid of Lewy body-associated dementias. J Alzheimers Dis. (2011) 26:171–9. doi: 10.3233/JAD-2011-110221
41. Marchionatti A, Woodhall M, Waters PJ, Sato DK. Detection of MOG-IgG by cell-based assay: moving from discovery to clinical practice. Neurol Sci Off J Ital Neurol Soc Ital Soc Clin Neurophysiol. (2021) 42:73–80. doi: 10.1007/s10072-020-04828-1
42. Hansen N, Rentzsch K, Hirschel S, Wiltfang J, Schott BH, Bartels C, et al. Case report: Anti-ARHGAP26 autoantibodies in atypical dementia with Lewy bodies. Front Dement. (2023) 2. doi: 10.3389/frdem.2023.1227823
43. Di Domenico F, Pupo G, Giraldo E, Lloret A, Badia M-C, Schinina ME, et al. Autoantibodies Profile in Matching CSF and Serum from AD and aMCI patients: Potential Pathogenic Role and Link to Oxidative Damage. Curr Alzheimer Res. (2016) 13:112–22. doi: 10.2174/1567205013666151218131424
44. Ehtewish H, Mesleh A, Ponirakis G, Lennard K, Al Hamad H, Chandran M, et al. Profiling the autoantibody repertoire reveals autoantibodies associated with mild cognitive impairment and dementia. Front Neurol. (2023) 14:1256745. doi: 10.3389/fneur.2023.1256745
45. Shim S-M, Koh YH, Kim J-H, Jeon J-P. A combination of multiple autoantibodies is associated with the risk of Alzheimer’s disease and cognitive impairment. Sci Rep. (2022) 12:1312. doi: 10.1038/s41598-021-04556-2
46. Goshima N, Kawamura Y, Fukumoto A, Miura A, Honma R, Satoh R, et al. Human protein factory for converting the transcriptome into an. vitro-expressed proteome Nat Methods. (2008) 5:1011–7. doi: 10.1038/nmeth.1273
47. Fukuda E, Tanaka H, Yamaguchi K, Takasaka M, Kawamura Y, Okuda H, et al. Identification and characterization of the antigen recognized by the germ cell mAb TRA98 using a human comprehensive wet protein array. Genes to Cells. (2021) 26:180–9. doi: 10.1111/gtc.12832
48. Matsuda KM, Yoshizaki A, Yamaguchi K, Fukuda E, Okumura T, Ogawa K, et al. Autoantibody Landscape Revealed by Wet Protein Array : Sum of Autoantibody Levels Re fl ects Disease Status. Front Immunol. (2022) 13:1–14. doi: 10.3389/fimmu.2022.893086
49. Kuzumi A, Norimatsu Y, Matsuda KM, Ono C, Okumura T, Kogo E, et al. Comprehensive autoantibody profiling in systemic autoimmunity by a highly-sensitive multiplex protein array. Front Immunol. (2023) 14. doi: 10.3389/fimmu.2023.1255540
50. Matsuda KM, Kotani H, Yamaguchi K, Okumura T, Fukuda E. Significance of anti-transcobalamin receptor antibodies in cutaneous arteritis revealed by proteome-wide autoantibody screening. J Autoimmun. (2023) 135:102995. doi: 10.1016/j.jaut.2023.102995
51. Matsuda KM, Kotani H, Yamaguchi K, Okumura T, Fukuda E. Autoantibodies to nuclear valosin-containing protein-like protein: systemic sclerosis-specific antibodies revealed by in vitro human proteome. Rheumatol (Oxford). (2024). doi: 10.1093/rheumatology/keae063
52. Matsuda KM, Kotani H, Yamaguchi K, Ono C, Okumura T, Ogawa K, et al. Proteome-wide autoantibody screening and holistic autoantigenomic analysis unveil COVID-19 signature of autoantibody landscape. medRxiv. (2024) 2024:06.07.24308592. doi: 10.1101/2024.06.07.24308592
53. Matsuda KM, Kawase Y, Iwadoh K, Kurano M, Yatomi Y, Okamoto K, et al. Deciphering autoantibody landscape of systemic sclerosis through systems-based approach: insights from a B-cell depletion clinical trial. medRxiv. (2024). doi: 10.1101/2024.07.30.24311212
54. Kotani H, Matsuda KM, Yamaguchi K, Ono C, Ogawa K, Kobayashi Y, et al. Diversity and epitope spreading of anti-RNA polymerase III antibodies in systemic sclerosis: a potential biomarker for skin and lung involvement. Arthritis Rheumatol. (2025) 77:67–79. doi: 10.1002/art.42975
55. Zhou Y, Zhou B, Pache L, Chang M, Benner C, Chanda SK, et al. Metascape provides a biologist-oriented resource for the analysis of systems-level datasets. Nat Commun. (2019) 10:1523. doi: 10.1038/s41467-019-09234-6
56. Thul PJ, Akesson L, Wiking M, Mahdessian D, Geladaki A, Ait Blal H, et al. A subcellular map of the human proteome. Sci (80-.). (2017) 356:eaal3321. doi: 10.1126/science.aal3321
57. Zhang C, Liu X, Wang J, Zhou G, Geng B, Hu M, et al. Investigating the causal relationships between lipid traits and dementia with lewy bodies: A mendelian randomization study. Parkinsonism Relat Disord. (2025) 0:107312. doi: 10.1016/j.parkreldis.2025.107312
58. García-Sanz P, Aerts JMFG, Moratalla R. The role of cholesterol in α-Synuclein and Lewy body pathology in Parkinson’s disease. Mov Disord. (2021) 36:1070–85. doi: 10.1002/mds.28396
59. Guerreiro R, Ross OA, Kun-Rodrigues C, Hernandez DG, Orme T, Eicher JD, et al. Investigating the genetic architecture of dementia with Lewy bodies: a two-stage genome-wide association study. Lancet Neurol. (2018) 17:64–74. doi: 10.1016/S1474-4422(17)30400-3
60. Tsuang D, Leverenz JB, Lopez OL, Hamilton RL, Bennett DA, Schneider JA, et al. APOE ϵ4 increases risk for dementia in pure synucleinopathies. JAMA Neurol. (2013) 70:223–8. doi: 10.1001/jamaneurol.2013.600
61. Shahmoradian SH, Lewis AJ, Genoud C, Hench J, Moors TE, Navarro PP, et al. Lewy pathology in Parkinson’s disease consists of crowded organelles and lipid membranes. Nat Neurosci. (2019) 22:1099–109. doi: 10.1038/s41593-019-0423-2
62. Singh G, Davenport AP. Neuropeptide B and W: neurotransmitters in an emerging G-protein-coupled receptor system. Br J Pharmacol. (2006) 148:1033–41. doi: 10.1038/sj.bjp.0706825
63. Wojciechowicz T, Billert M, Jasaszwili M, Strowski MZ, Nowak KW, Skrzypski M, et al. The role of neuropeptide B and its receptors in controlling appetite, metabolism, and energy homeostasis. Int J Mol Sci. (2021) 22:6632. doi: 10.3390/ijms22126632
64. Nagata-Kuroiwa R, Furutani N, Hara J, Hondo M, Ishii M, Abe T, et al. Critical role of neuropeptides B/W receptor 1 signaling in social behavior and fear memory. PLoS One. (2011) 6:e16972. doi: 10.1371/journal.pone.0016972
65. Watanabe N, Wada M, Irukayama-Tomobe Y, Ogata Y, Tsujino N, Suzuki M, et al. A single nucleotide polymorphism of the neuropeptide B/W receptor-1 gene influences the evaluation of facial expressions. PLoS One. (2012) 7:e35390. doi: 10.1371/journal.pone.0035390
66. Kelly MA, Beuckmann CT, Williams SC, Sinton CM, Motoike T, Richardson JA, et al. Neuropeptide B-deficient mice demonstrate hyperalgesia in response to inflammatory pain. Proc Natl Acad Sci U S A. (2005) 102:9942–7. doi: 10.1073/pnas.0503795102
67. Yamamoto T, Saito O, Shono K, Tanabe S. Anti-hyperalgesic effects of intrathecally administered neuropeptide W-23, and neuropeptide B, in tests of inflammatory pain in rats. Brain Res. (2005) 1045:97–106. doi: 10.1016/j.brainres.2005.03.027
68. Einspahr JM, Tilley DG. Pathophysiological impact of the adhesion G protein-coupled receptor family. Am J Physiol Physiol. (2022) 323:C640–7. doi: 10.1152/ajpcell.00445.2021
69. Bridges JP, Ludwig M-G, Mueller M, Kinzel B, Sato A, Xu Y, et al. Orphan G protein-coupled receptor GPR116 regulates pulmonary surfactant pool size. Am J Respir Cell Mol Biol. (2013) 49:348–57. doi: 10.1165/rcmb.2012-0439OC
70. Yang MY, Hilton MB, Seaman S, Haines DC, Nagashima K, Burks CM, et al. Essential regulation of lung surfactant homeostasis by the orphan G protein-coupled receptor GPR116. Cell Rep. (2013) 3:1457–64. doi: 10.1016/j.celrep.2013.04.019
71. Abe J, Suzuki H, Notoya M, Yamamoto T, Hirose S. Ig-hepta, a novel member of the G protein-coupled hepta-helical receptor (GPCR) family that has immunoglobulin-like repeats in a long N-terminal extracellular domain and defines a new subfamily of GPCRs *. J Biol Chem. (1999) 274:19957–64. doi: 10.1074/jbc.274.28.19957
72. DiBlasi E, Shabalin AA, Monson ET, Keeshin BR, Bakian AV, Kirby AV, et al. Rare protein-coding variants implicate genes involved in risk of suicide death. Am J Med Genet Part B Neuropsychiatr Genet. (2021) 186:508–20. doi: 10.1002/ajmg.b.v186.8
73. Kaur G, Poljak A, Masters CL, Fowler C, Sachdev P. Impact of APOE ϵ3 and ϵ4 genotypes on plasma proteome signatures in Alzheimer’s disease. bioRxiv. (2022) 2022:1.29.478291. doi: 10.1101/2022.01.29.478291
74. Beach TG, Adler CH, Sue LI, Vedders L, Lue LI, White III, et al. Multi-organ distribution of phosphorylated alpha-synuclein histopathology in subjects with Lewy body disorders. Acta Neuropathol. (2010) 119:689–702. doi: 10.1007/s00401-010-0664-3
75. Sawasaki T, Ogasawara T, Morishita R, Endo Y. A cell-free protein synthesis system for high-throughput proteomics. Proc Natl Acad Sci U S A. (2002) 99:14652–7. doi: 10.1073/pnas.232580399
76. Jack CR Jr., Bennett DA, Blennow K, Carrillo MC, Dunn B, Haeberlein SB, et al. NIA-AA Research Framework: Toward a biological definition of Alzheimer’s disease. Alzheimer’s Dement. (2018) 14:535–62. doi: 10.1016/j.jalz.2018.02.018
Keywords: autoantibody, artificial intelligence, machine learning, dementia, Alzheimer’s disease, Lewy body dementia
Citation: Matsuda KM, Umeda-Kameyama Y, Iwadoh K, Miyawaki M, Yakabe M, Ishii M, Ogawa S, Akishita M, Sato S and Yoshizaki A (2025) Artificial intelligence and omics-based autoantibody profiling in dementia. Front. Immunol. 16:1537659. doi: 10.3389/fimmu.2025.1537659
Received: 01 December 2024; Accepted: 07 April 2025;
Published: 08 May 2025.
Edited by:
Brigitte Vannier, University of Poitiers, FranceReviewed by:
Niels Hansen, University Medical Center Göttingen, GermanyYuting Zhang, The Scripps Research Institute, United States
Copyright © 2025 Matsuda, Umeda-Kameyama, Iwadoh, Miyawaki, Yakabe, Ishii, Ogawa, Akishita, Sato and Yoshizaki. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Ayumi Yoshizaki, YXl1eW9zaGlAbWUuY29t
†ORCID: Kazuki M. Matsuda, orcid.org/0000-0002-6755-8047
Ayumi Yoshizaki, orcid.org/0000-0002-8194-9140