 Yahya Sohrabi1,2,3
Yahya Sohrabi1,2,3 Tatyana Kobets2Valeriya Volkova2Eliška Javorková4Imtissal Krayem2,5Alena Zajícová4Helena Havelková2Milena Svobodová6Vladimír Holáň4Peter Demant7
Tatyana Kobets2Valeriya Volkova2Eliška Javorková4Imtissal Krayem2,5Alena Zajícová4Helena Havelková2Milena Svobodová6Vladimír Holáň4Peter Demant7 Marie Lipoldová1,2,8*†
Marie Lipoldová1,2,8*†- 1Department of Medical Genetics, Third Faculty of Medicine, Charles University, Prague, Czechia
- 2Laboratory of Molecular and Cellular Immunology, Institute of Molecular Genetics, Czech Academy of Sciences, Prague, Czechia
- 3Department of Cardiology I, Coronary, Peripheral Vascular Disease and Heart Failure, University Hospital Münster, University of Münster, Münster, Germany
- 4Department of Toxicology and Molecular Epidemiology, Institute of Experimental Medicine of the Czech Academy of Sciences, Prague, Czechia
- 5Laboratory of Leukocyte Signalling, Institute of Molecular Genetics, Czech Academy of Sciences, Prague, Czechia
- 6Department of Parasitology, Faculty of Science, Charles University, Prague, Czechia
- 7Department of Molecular and Cellular Biology, Roswell Park Comprehensive Cancer Center, Buffalo, NY, United States
- 8Laboratory of Signal Transduction, Institute of Molecular Genetics, Czech Academy of Sciences, Prague, Czechia
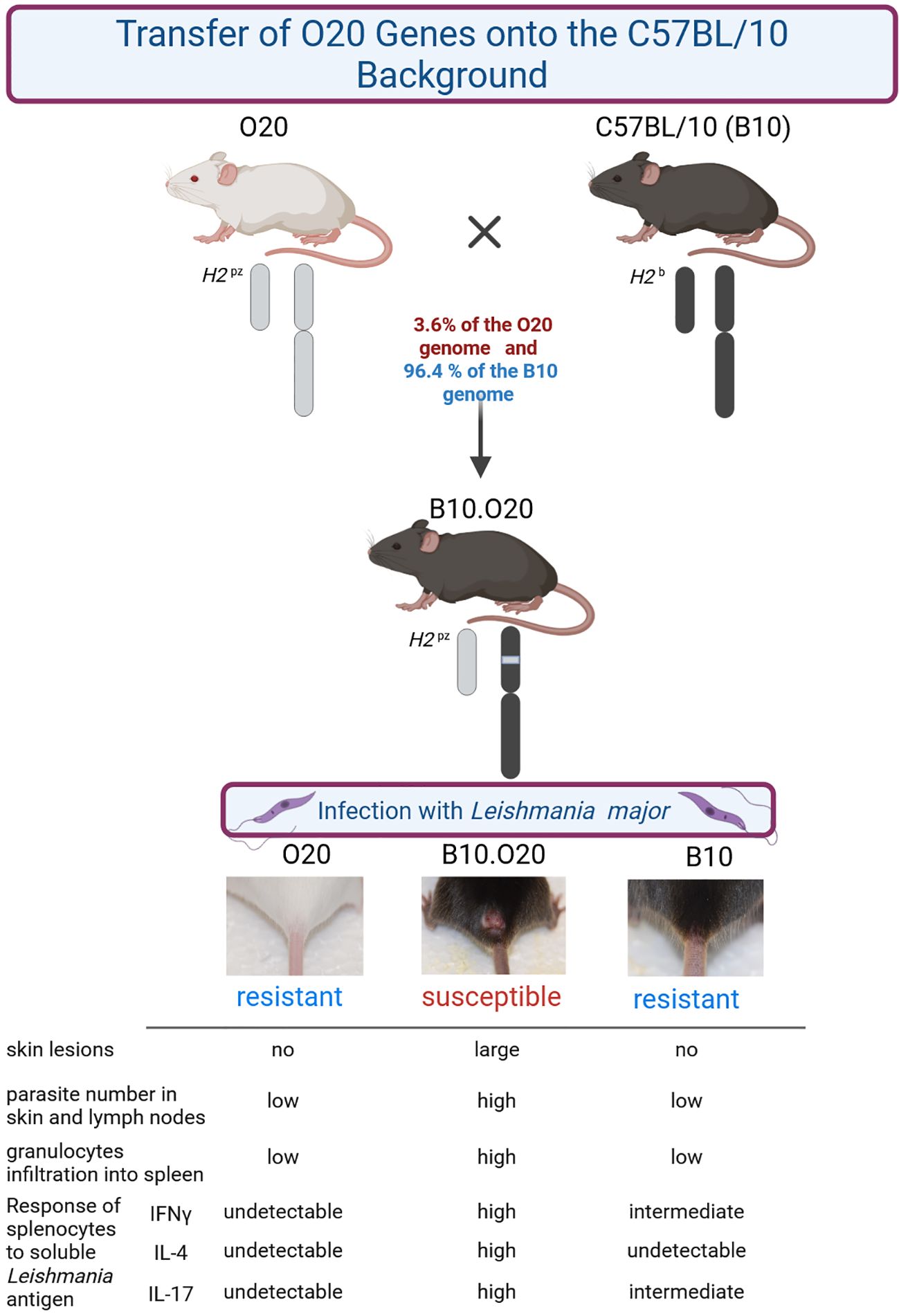
The mouse strain O20 is highly resistant to parasite Leishmania major. O20 mice differed from all resistant strains tested until now, as they harbored parasites in their organs, but upon exposure to soluble Leishmania antigen (SLA) their splenocytes did not respond by cytokine production and their macrophages did not produce NO, suggesting a novel mechanism of resistance. Another resistant strain C57BL/10 (B10) harbors similar numbers of parasites as O20 in its organs and its splenocytes respond to SLA by production of IFNγ, but not IL-4. They also produce IL-2, IL-6, IL-10 and IL-17. Macrophages respond to SLA by NO production. Strain B10.O20 was derived from a cross of these two resistant strains. B10 provided 96.4% of its genome and O20 contributed 3.6% of its genome. Unexpectedly, this very limited difference between the two strains resulted in the very large phenotypic effects. B10.O20 was susceptible to L. major, as it exhibited large skin lesions, high parasite numbers in skin and lymph nodes, and a massive spleen infiltration by CD11b+CD193+ and CD11b+Gr1+ cells. Thus, a small percentage of genes of the resistant strain O20 in the genome of the second resistant strain B10 resulted in high susceptibility to L. major. After stimulation with SLA, splenocytes of B10.O20 produced significantly higher levels of all Th1, Th2 and Th17 cytokines than both its parental strains B10 and O20. This suggested a chronic inflammation with imbalance of several arms of immune response. In summary, the responses of strains B10.O20 and O20 to L. major revealed novel disease phenotypes that have not been observed previously in mice but they were seen in several clinical studies of human leishmaniasis. The studies of heterogeneity of defensive strategies of mouse strains may guide development of effective antileishmanial therapies or vaccine development and it could serve as a basis for investigation of asymptomatic responses to other infectious diseases.
Graphical Abstract.
Introduction
More than 1 billion people living in areas endemic for leishmaniasis are at risk of infection by Leishmania – an obligatory intracellular protozoan parasite (Kinetoplastida) of vertebrates, transmitted by female phlebotomine sandflies (1–4). The principal vertebrate host target cell is the macrophage, where parasites proliferate, but Leishmania parasites can also invade many other cell types (5–7). Three main clinical syndromes of leishmaniasis were described: the cutaneous form of the disease in dermis, which can be localized or diffuse; mucocutaneous leishmaniasis in the mucosa, and the visceral leishmaniasis that results from the spread of infection to the spleen and liver (8, 9). Currently, there is no human vaccine to prevent leishmaniasis and drugs used for treatment have serious side effects (4, 10, 11).
In addition to people with clinical signs of the leishmaniasis, there are at least 120 million people with asymptomatic infection (12). While they do not manifest any symptoms of the disease, their serological, PCR (polymerase chain reaction) or leishmanin skin tests are positive (13). In some areas these subclinical infections represent 80-90% of those infected [reviewed in (14)]. These clinically asymptomatic individuals can serve as reservoirs for transmission of parasites (15, 16). The persistent parasites also pose a risk to asymptomatic individuals, as they can be reactivated in immunocompromised host (17, 18). The resistance or susceptibility to the infection and manifestations of the disease depend on Leishmania species, and on genetic and non-genetic factors such as age, sex, and social and environmental influences (2, 19–25).
It is difficult to control these multiple factors in analyses of human infection, therefore many aspects of infection were analyzed in animal experiments. Especially the mouse studies with large numbers of genetically identical individuals kept in controlled conditions helped to establish many elements of mechanical and genetic control of the disease (19, 26–30). The most extensively studied mouse model has been infection by Leishmania major. This parasite causes cutaneous lesions in humans, but induces both cutaneous and systemic pathology in mouse. Most information about response to infection was obtained using the two paradigmatic strains: susceptible BALB/c and resistant C57BL/6N (B6) (23, 26).
Following inoculation of L. major, susceptible BALB/c mice exhibit uncontrolled growth of the parasite in the primary site of infection, development of large necrotic lesions and dissemination of parasites beyond the local draining lymph node to spleen, liver, bone marrow, and other cutaneous sites (31, 32), whereas resistant B6 mice develop neither cutaneous nor systemic disease (31).
Several early studies suggested that resistance or susceptibility to infection caused by L. major was controlled by the behavior of different subpopulations of helper T cells (33, 34), because the amount of IL-4 produced by T helper 2 (Th2) cells appeared to correlate with progression of leishmaniasis, whereas activation of IFNγ-producing Th1 lymphocytes, which promotes the production of nitric oxide in phagocytes at the site of infection (35), has been associated with resolution of the disease. Subsequent studies revealed that both T and non-T compartments contributed to determination of the susceptibility or resistance (36) and identified additional cells and cytokines involved to the susceptibility (32, 37). They include dendritic cells (38, 39), neutrophils (40, 41), keratinocytes (42), myeloid-derived suppressor cells (43), and microRNA (44).
Multiple regulatory factors revealed in mechanistic studies have been supported by genetic analyses that established a network-like organization of the numerous functionally diverse genes influencing susceptibility to leishmaniasis (19, 23, 27, 45–49).
Despite these insights into the immune response against Leishmania parasites, there are indication that additional responses required for protection against the disease remain to be defined (10, 50). This is supported by the fact that there is currently no safe and effective vaccine that would prevent any form of human leishmaniasis (4, 10, 11, 50, 51).
This might be partly caused by the fact that the majority of analyses had been performed only on a few mouse strains (26, 52), which often originated from common ancestors (53). This does not reflect the multiple genotypes existing in the human population.
In order to develop an additional model for analysis of susceptibility to L. major, we tested response to L. major in strains O20/A (O20), C57BL/10Sn (B10), and compared it to that of C57BL/10-H2pz (B10.O20). The strain B10.O20 is a H2 congenic strain on the B10 background (N8), which carries H2pz haplotype derived from the strain O20 (54). Because the inbreeding of the strain B10.O20 started at the eighth backcross generation, approximately 3.6% of its genome, including H2 region, are derived from the strain O20 (55). Analysis of susceptibility to L. major in the strains O20, B10 and B10.O20 revealed that the combination of genes of two resistant strains, O20 and B10, present in the strain B10.O20 caused its susceptibility. Here we show that this susceptibility of B10.O20 mice is associated with novel immunological and pathological characteristics of leishmaniasis of this strain. We also observed that the strain O20 exhibits a paradoxical combination of high resistance to infection and a defective response to SLA (soluble Leishmania antigen) previously unreported in mice. In order to facilitate the understanding of these novel phenotypes, we give a detailed description of the immunological parameters of uninfected and infected O20, B10 and B10.O20 mice.
Materials and methods
Mice
The first set of experiments comprised: 147 (71 infected and 76 uninfected) female mice of strains O20/A (abbrev. O20) total 57 (23 infected and 34 uninfected), B10.O20/R164/Dem (abbrev. B10.O20) (The Jackson Laboratory Strain 38154/MMRRC: 069936) total 51 (24 infected and 27 uninfected) and C57BL/10Sn (abbrev. B10) total 39 (24 infected and 15 uninfected) that were tested in 5 independent experiments. Age of mice was 12 to 28 weeks (mean 17.8 weeks, median 16.1 weeks) at the time of infection. 3 mice died during experiments and one mouse, which had injury (was bitten), was excluded from calculation.
The second set of experiments: 35 (18 infected and 17 uninfected) female mice of strains BALB/c (6 infected and 6 uninfected), STS (6 infected and 5 uninfected) and O20 (6 infected and 6 uninfected) were tested in 2 independent experiments. Age of mice was 9 to 15 weeks (mean 12.4 weeks, median 13 weeks) at the time of infection.
Mice were euthanized by cervical dislocation after 8 weeks of infection. Blood, skin, spleen, liver and inguinal lymph nodes were collected for later analysis.
Ethical statement
All experimental protocols utilized in this study comply with the Czech Government Requirements under the Policy of Animal Protection Law (No.246/1992) and with the regulations of the Ministry of Agriculture of the Czech Republic (No.207/2004), which are in agreement with all relevant European Union guidelines for work with animals. The study was approved by the Institutional Animal Care Committee of the Institute of Molecular Genetics of the Czech Academy of Sciences and by Departmental Expert Committee for the Approval of Projects of Experiments on Animals of the Czech Academy of Sciences (permission Nr. 93/2015).
Parasite
Leishmania major LV 561 (MHOM/IL/67/LRCL 137 JERICHO II) was maintained in rump lesions of BALB/c females. Amastigotes were transformed to promastigotes using SNB-9 medium (56). 107 promastigotes from the passage number 2 cultivated for six days were inoculated in 50 μl sterile saline s.c. into mouse rump (57).
Disease phenotype
The size of the skin lesions was measured every second week using the Profi LCD Electronic Digital Caliper Messschieber Schieblehre Messer (Shenzhen Xtension Technology Co., Ltd. Guangdong, China), which has accuracy 0.02 mm.
Quantification of parasite load by PCR-ELISA
Parasite load was measured in frozen skin, lymph nodes, spleen, and liver samples using PCR-ELISA according to the previously published protocol (58). Briefly, total DNA was isolated using a TRI reagent (Molecular Research Center, Cincinnati, USA) standard procedure (https://www.mrcgene.com/dna-isolation) or a modified proteinase K procedure (58). For PCR, two primers (digoxigenin-labeled F 5′-ATT TTA CAC CAA CCC CCA GTT-3′ and biotin-labeled R 5′-GTG GGG GAG GGG CGT TCT-3′ (VBC Genomics Biosciences Research, Austria) were used for amplification of the 120-bp conservative region of the kinetoplast minicircle of Leishmania parasite, and 50 ng of extracted DNA was used per each PCR reaction. For a positive control, 20 ng of L. major DNA per reaction was amplified as a highest concentration of standard. A 30-cycle PCR reaction was used for quantification of parasites in lymph nodes, and 35 cycles for skin, spleen, and liver. Under these conditions the amount of PCR product is linearly proportional to the number of parasites (58). PCR product was measured by the modified ELISA (Pharmingen, San Diego, USA). Concentration of Leishmania DNA was determined using the ELISA Reader Tecan and the curve fitter program KIM-E (Schoeller Pharma, Prague, Czech Republic) with least squares-based linear regression analysis.
Preparation of soluble Leishmania antigen
L. major LV 561 (MHOM/IL/67/LRC-L137 JERICHO II) promastigotes were harvested from RPMI 1640 medium supplemented with 10% inactivated fetal calf serum (Sigma-Aldrich, USA), 63.7 μg/ml penicillin (Sigma-Aldrich, USA), and 100 μg/ml streptomycin (Sigma-Aldrich, USA), washed 3 times with PBS (pH 7.2) and used for preparation of soluble Leishmania antigen (SLA) as previously described (59). Briefly, 100 μl of protease inhibitor cocktail enzyme (Sigma, St. Louis, MO, USA) was added to 1 × 109 promastigotes, the parasites were freeze-thawed 10 times followed by sonication at 4°C with two 20-sec blasts. The parasite suspension was then centrifuged at 30,000× g for 20 min at 4°C, the supernatant was collected and re-centrifuged at 100,000× g for 4 h at 4°C, and the resulting supernatant was filtered through a 0.22 μm MILLEX® GP Syringe Filter Unit (Millipore, Carrigtwohill, Co., Cork, Ireland). Protein concentration of SLA was determined using Lowry method, and the SLA preparation was aliquoted and stored at −20°C until used. In first and second sets of experiments (see the above section Mice) were used different batches of SLA.
Stimulation of splenocytes or macrophages by SLA
Single cell suspensions of spleen cells were prepared in RPMI 1640 medium (Sigma Corp., St. Louis, MO) containing 10% of FCS (Sigma), antibiotics (100 U/ml of penicillin, 100 μg/ml of streptomycin), 10 mM HEPES buffer, and 5 x 10-5 2-mercaptoethanol. Spleen cells at a concentration 0.6 x 106 cells/ml were incubated in 48-well tissue culture plates (Corning Inc., Corning, NY) in a final volume of 1 ml of complete RPMI 1640 medium unstimulated or stimulated with 6 µg/ml of soluble Leishmania antigen (SLA). The supernatants were harvested after a 24-h (IL-2 detection), 48-h (IFN-γ detection) or 72-h (IL-4, IL-6, IL-10 and IL-17 detection) incubation at 37°C in an atmosphere of 5% CO2.
To produce and measure IL-1β, IL-12 and NO, peritoneal exudate cells (PEC) containing 20-30% of macrophages (60) obtained by washing the peritoneal cavity of tested mice were adjusted to a concentration of 1 × 106 cells/ml and were stimulated with 6 µg/ml of SLA. The supernatants were harvested after 48-h incubation for detection of NO or after 72 h for quantification of IL-1β and IL-12.
Cytokine detection and quantification by ELISA
The production of IL-1β, IL-2, IL-4, IL-6, IL-10, IL-12, IL-17 and IFNγ was quantified by ELISA. The pairs of cytokine-specific capture and detection monoclonal antibodies (mAb) purchased from R&D Systems, Inc. (Minneapolis, MN) were used for the detection of IL-2 (mAb clones JES6-5H4 and JES6-1A12), IL-6 (mAb clones MP5-20F3 and MP5-32C11) and IFNγ (mAb clones R4-6A2 and XM6-1.2). IL-1β, IL-4, IL-10, IL-12 and IL-17 were measured using ELISA kits (DuoSet ELISA Development System: IL-4 (kit DY404), IL-10 (DY417), IL-17 (DY421), IL-1β (DY401), IL-12 (DY499)) purchased from R & D Systems (Minneapolis, MN), following the instructions of the manufacturer. The reaction was quantified by spectrophotometry using a Sunrise Remote ELISA Reader (Gröding, Austria).
Nitric oxide measurement
Briefly, 100 μl of the cell culture supernatant was incubated with 50 μl of 1% sulfanilamide (in 3% H3PO4) and 50 μl of 0.3% N-1-napthyletylenediamine dihydrochloride (in 3% H3PO4) at room temperature for 5 min. Nitrite was quantified by spectrophotometry at 550 nm using sodium nitrite as a standard (61).
Flow cytometry characterization of spleens from infected and uninfected mice
Single-cell suspensions prepared from spleens of infected and uninfected mice were washed in PBS containing 0.5% BSA and incubated for 30 min on ice with the following anti-mouse mAb (all purchased from BioLegend, San Diego, CA): allophycocyanine (APC)-labeled anti-CD11b (clone M1/70), fluorescein isothiocyanate (FITC)-labeled anti-Ly6G/Ly-6C (Gr-1) (clone RB6-8C5), FITC-labeled anti-CD19 (clone 6D5), Alexa Fluor 647-labeled anti-CD22 (clone OX-97), phycoerythrin-labeled anti-CD193 (clone J073E5), APC-labeled anti-CD3 (clone 17A2), FITC-labeled anti-CD4 (clone GK1.5), PE-labeled anti-CD8 (clone 53-6.7), APC-labeled anti-CD80 (clone 16-10A1), FITC-labeled anti-CD206 (clone C068C2), PE-labeled anti-F4/80 (clone BM8), PE-labeled anti-CD14 (clone Sa 14-2). All samples were also incubated with Pacific Blue-labeled anti-TER-119 (clone TER-119) in order to exclude erythroid cells. Dead cells were stained using Hoechst 33258 fluorescent dye (Invitrogen, Carlsbad, CA) added to the samples 10 min before flow cytometry analysis. Data were collected using an LSRII cytometer (BD Biosciences, Franklin Lakes, NJ) and analyzed using FlowJo software (Tree Star, Ashland, OR). Fifty thousand events from each sample were measured.
Evaluation of histopathological changes in spleen
Organs were fixed in 4% formaldehyde and embedded in paraffin. Histological changes were evaluated in hematoxylin-eosin (H&E) stained 2 μm sections under a light microscope.
Statistical analysis
Differences between strains O20, B10.O20 and B10, and between infected and uninfected mice within tested strains were analyzed by Mann-Whitney test using the program Statistica for Windows 12.0 (StatSoft, Inc., Tulsa, Oklahoma, USA).
Results
Combination of genes of two resistant strains results in a susceptible strain
Previously, H2pz haplotype of the Leishmania-resistant strain O20 was transferred on the background of another resistant strain C57BL/10Sn-H2b (B10) to create a strain C57BL/10-H2pz (B10.O20) (62). We have tested these three strains, B10, O20 and B10.O20, for susceptibility to L. major and found that the combination of genomes of the two resistant strains B10 and O20, gave rise to the strain B10.O20 (Figure 1), which was susceptible and had larger skin lesions (Figure 1A), higher number of parasites in skin (Figure 1B) and in inguinal lymph nodes (Figure 1C) than either resistant parental strain. The presence of high numbers of amastigotes in the skin of B10.O20 was also confirmed by anti-Leishmania staining (Supplementary Figure 1). B10.O20 also developed extensive epithelioid granulomas in the skin (Figure 2C), whereas B10 had only isolated granulomas (Figure 2A) and O20 mice did not develop any epithelioid granulomas (Figure 2E). Skins of uninfected control mice are shown in Figures 2B, D, F.
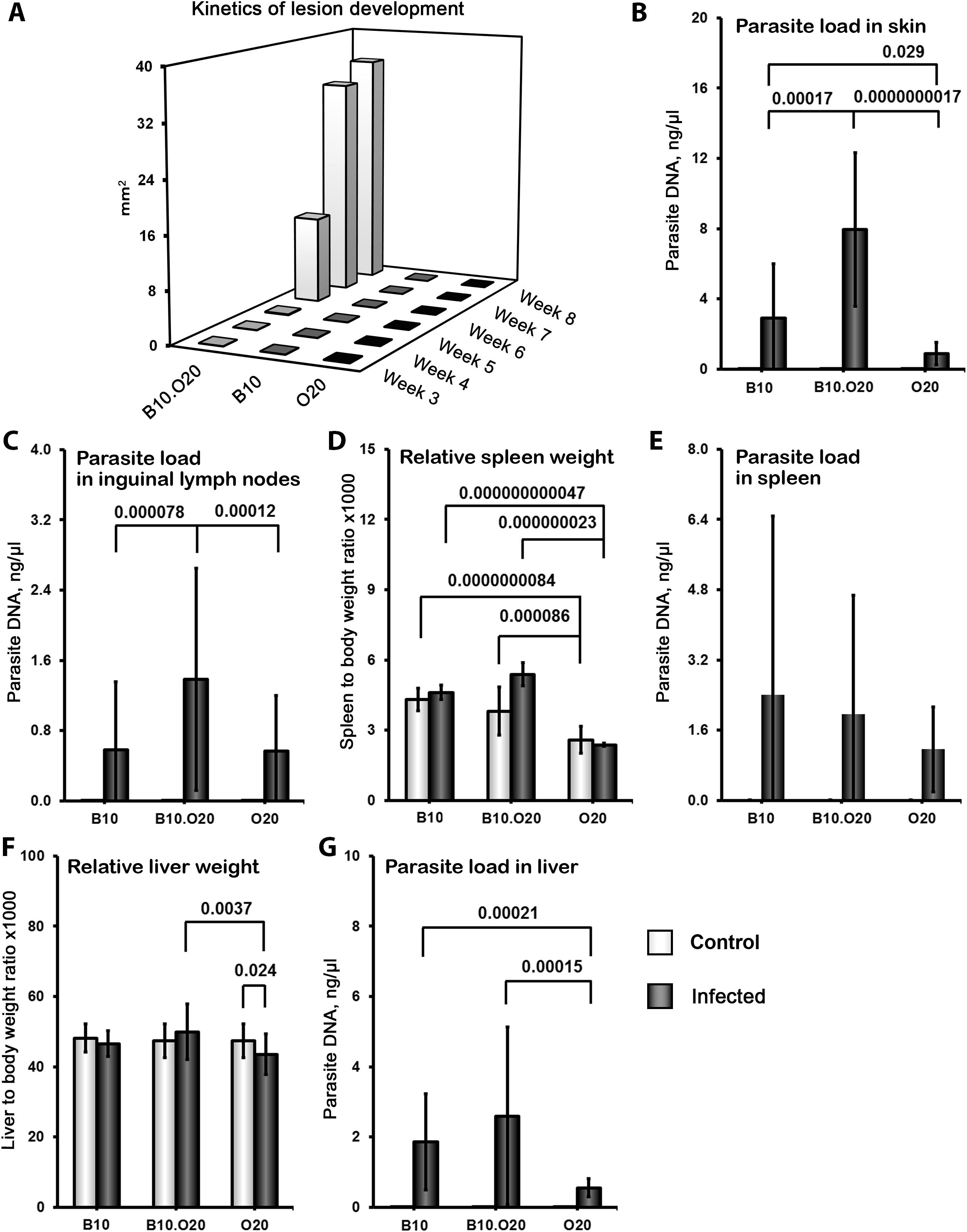
Figure 1. Differences in skin lesions, relative spleen and liver weight and parasite numbers in organs after L. major infection. Female mice of strains O20 (23 infected, 28 uninfected), B10 (23 infected, 11 uninfected) and B10.O20 (21 infected, 23 uninfected) were compared. Animals were subcutaneously inoculated with 107 promastigotes of L. major. Control, uninfected mice were kept in the same animal facility. Both groups were killed after 8 weeks of infection. The data show the medians (A - skin lesions) and the means ± SD (B–G) from five independent experiments.
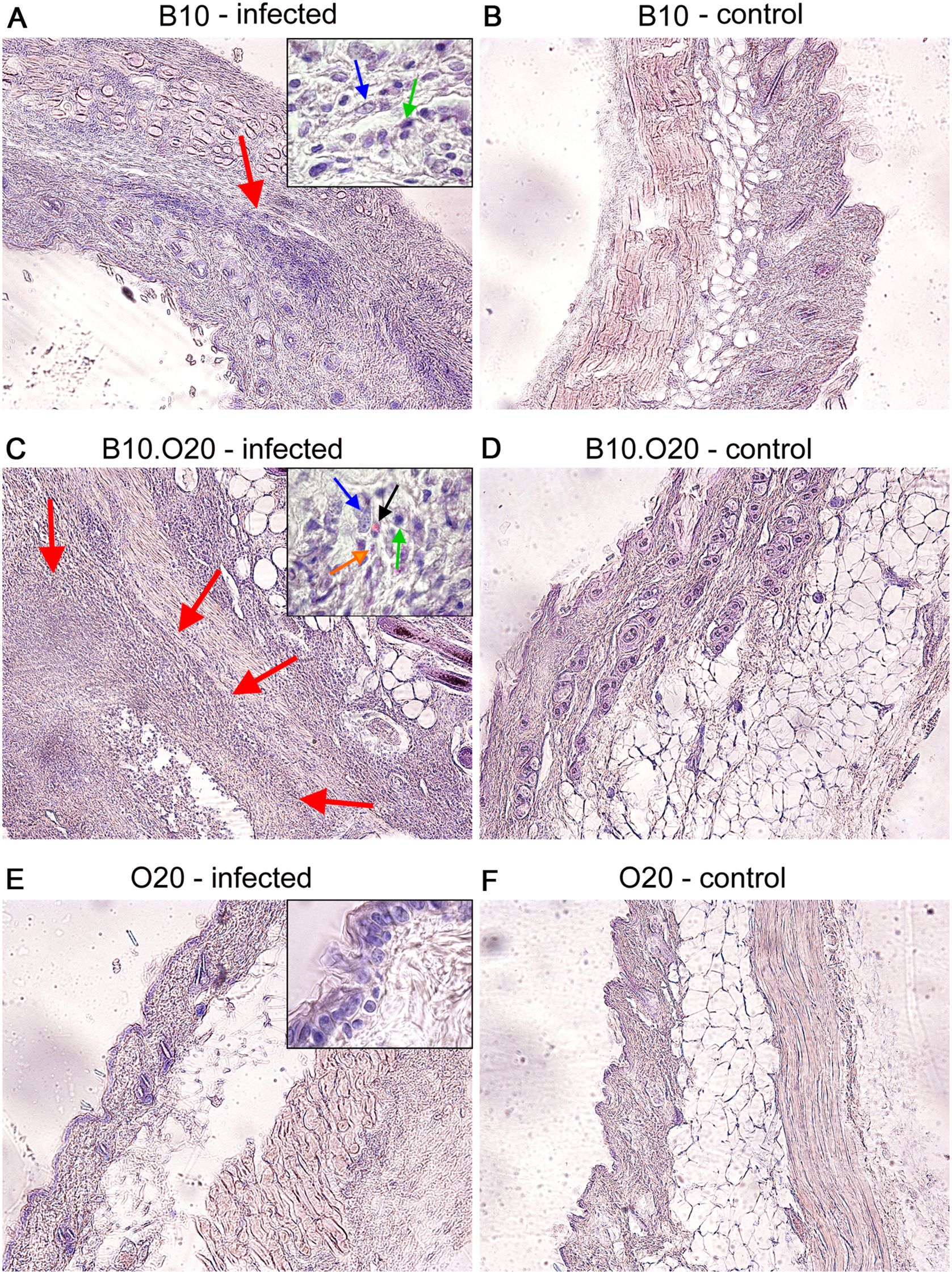
Figure 2. Comparison of the strain influence on architecture of infected skin in O20, B10, and B10.O20 mice. Photos show hematoxylin-eosin (H&E) stained skin sections of infected (A, C, E) and uninfected (B, D, F) female mice at x100 magnification. Red arrows point areas of epithelioid granulomas. Infected mice of susceptible B10.O20 strain developed extensive granulomas with massive infiltration of various immune cells and parasites. In the high magnification inserts, blue, green and black arrows point macrophages, lymphocytes and eosinophils, respectively. Orange arrow shows presence of amastigotes in skin of B10.O20.
In addition, considerable pathological changes were observed also in spleens of B10.O20 mice, but not in B10 and O20 (see the next section).
Spleens of susceptible strain B10.O20 exhibit upon infection extensive disorganization and high myeloid-derived cells infiltration
Strain O20 has a lower relative spleen weight than both B10 and B10.O20 prior infection (Figure 1D). None of the three strains exhibited significant increase of relative spleen weight after infection (Figure 1D) and tested strains also did not significantly differ in parasite load in spleen (Figure 1E). Figure 3 shows microarchitecture of spleens of uninfected (Figures 3B, D, F) and infected (Figures 3A, C, E) mice. Spleens of B10.O20 were extensively disorganized (Figure 3C), having lost the distinction between red and white pulp. They had also a higher frequency of CD11b+Gr1+ (Figures 4A, C) and CD11b+CD193+ (Figures 4B, D) subpopulations than both B10 (P = 0.0012, P = 0.0012, respectively) and O20 (P = 0.00058, P = 0.00058, respectively). The frequency of CD11b+Gr1+ subpopulation in the uninfected strain B10.O20 was higher than in both parental strains, and the L. major infection led to dramatic increase of these cells (Figures 4A, C). Similarly, uninfected B10.O20 had higher frequency of CD11+CD193+ (eosinophils) subpopulation than the strain O20; infection led to increase of CD11+CD193+ in B10.O20, but not in B10 and O20, which led to the increase of the difference between B10.O20 and its parental strains (Figures 4B, C). Higher infiltration of eosinophils in spleens of B10.O20 mice was confirmed also by observation of high numbers of eosinophils in histological examination (Figure 5).
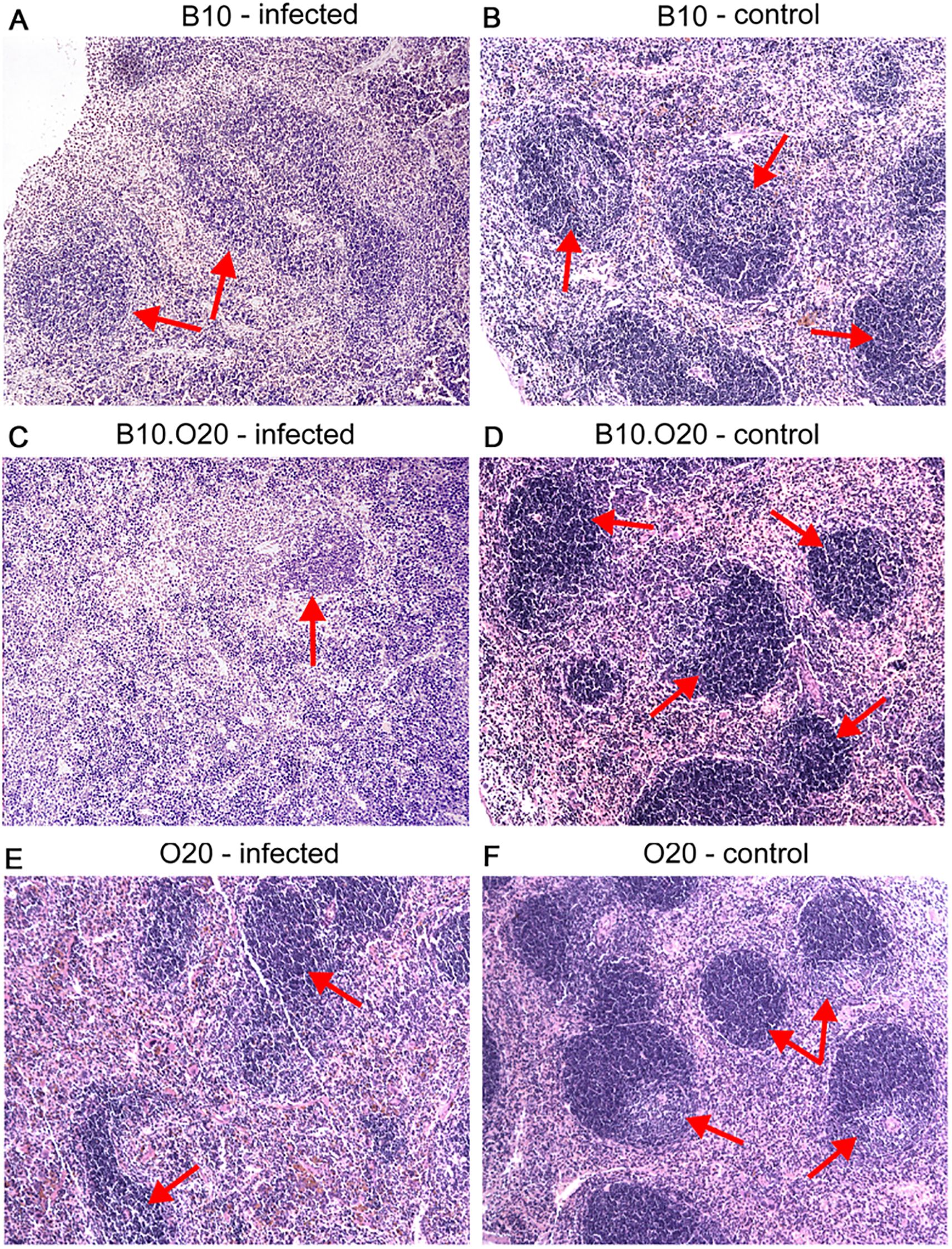
Figure 3. Microarchitecture of the spleen. Photos show microstructure of spleen of infected (A, C, E) and uninfected (B, D, F) female mice at x100 magnification, hematoxylin–eosin stained cuts. Arrows point areas of preserved white pulp. Infected mice exhibit signs of white pulp activation, especially notable in the susceptible B10.O20 strain (C).
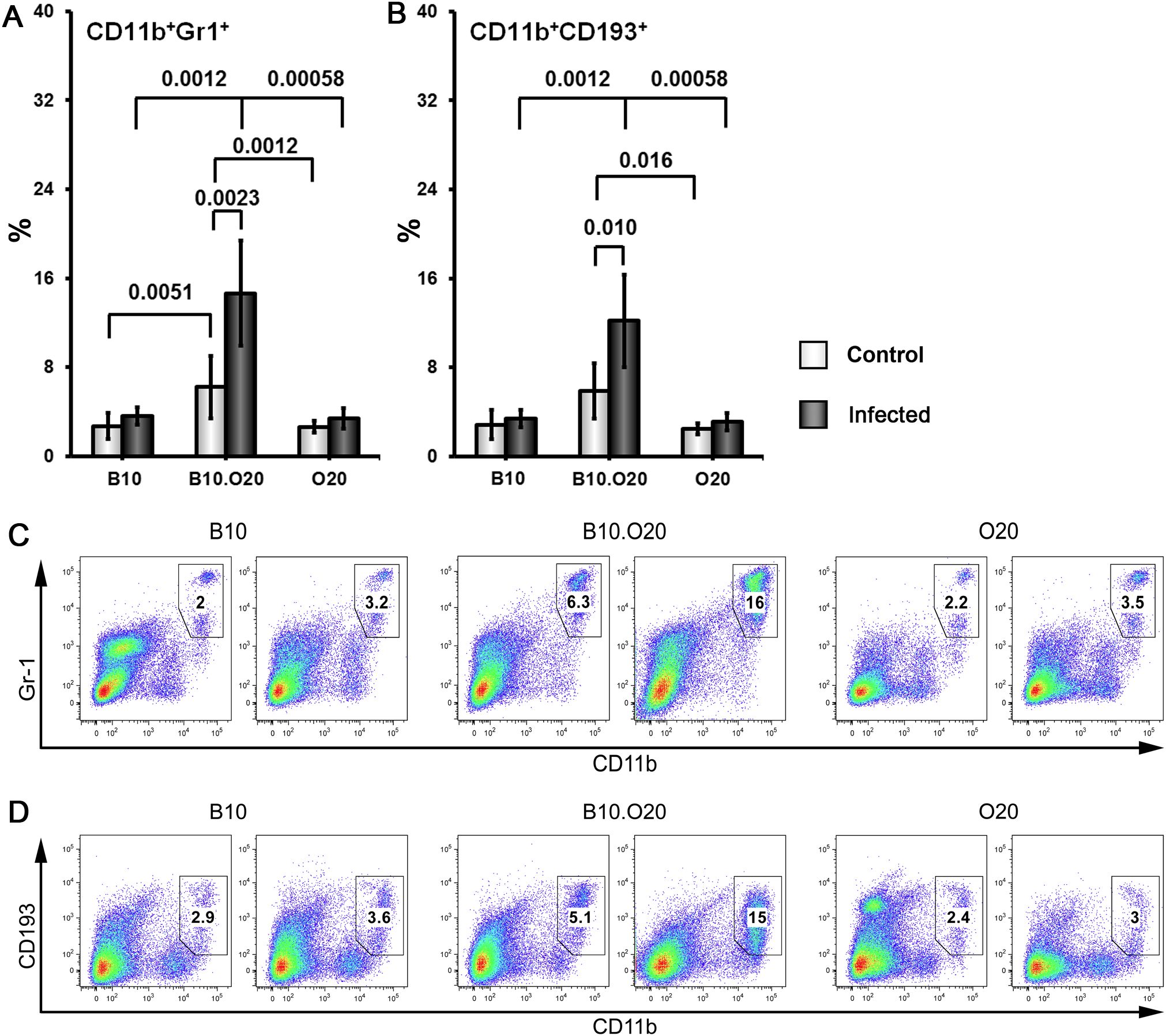
Figure 4. Flow cytometric analysis of freshly obtained mouse spleen cells. Female mice of strains O20 (7 infected, 6 uninfected), B10 (6 infected, 5 uninfected) and B10.O20 (7 infected, 7 uninfected) were compared. Frequencies of CD11b+Gr1+ cells (A, C) and CD11b+CD193+ cells (B, D) are shown. Dead cells were stained using Hoechst 33258 fluorescent dye added to the samples 10 min before flow cytometry analysis. Cells were stained with allophycocyanine (APC)-labeled anti-CD11b (clone M1/70), fluorescein isothiocyanate (FITC)-labeled anti-Ly6G/Ly-6C (Gr-1) (clone RB6-8C5) and phycoerythrin-labeled anti-CD193 (clone J073E5) antibodies as described in Materials and Methods. Selected cell populations were gated on single cell events after exclusion of cell debris and dead cells. Figure shows means ± SD from 4 independent experiments and a representative FACS experiment.
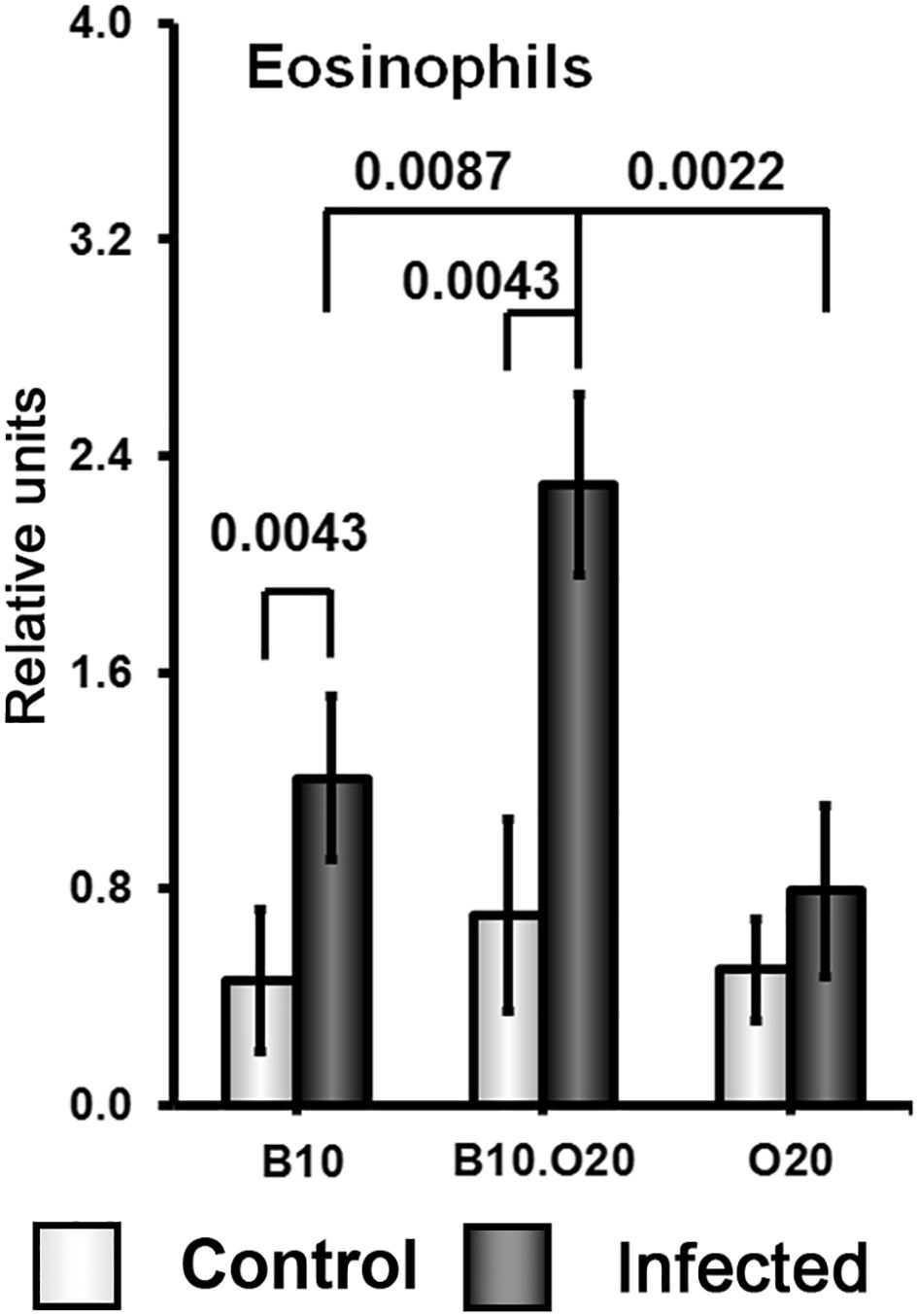
Figure 5. Eosinophil infiltration in spleens of infected and uninfected mice. Female mice of strains O20 (6 infected, 6 uninfected), B10 (6 infected, 6 uninfected) and B10.O20 (6 infected, 5 uninfected) were compared. The data show the means ± SD from two independent experiments. The analysis was performed on H&E sections.
Because of the striking high frequency of CD11b+Gr1+ and CD11+CD193+ subpopulations in the strain B10.O20, we decided to compare it with the most intensively studied susceptible strain BALB/c (Figure 6). We found that uninfected B10.O20 mice have higher frequency of both these subpopulations than uninfected BALB/c and this difference increased after L. major infection.
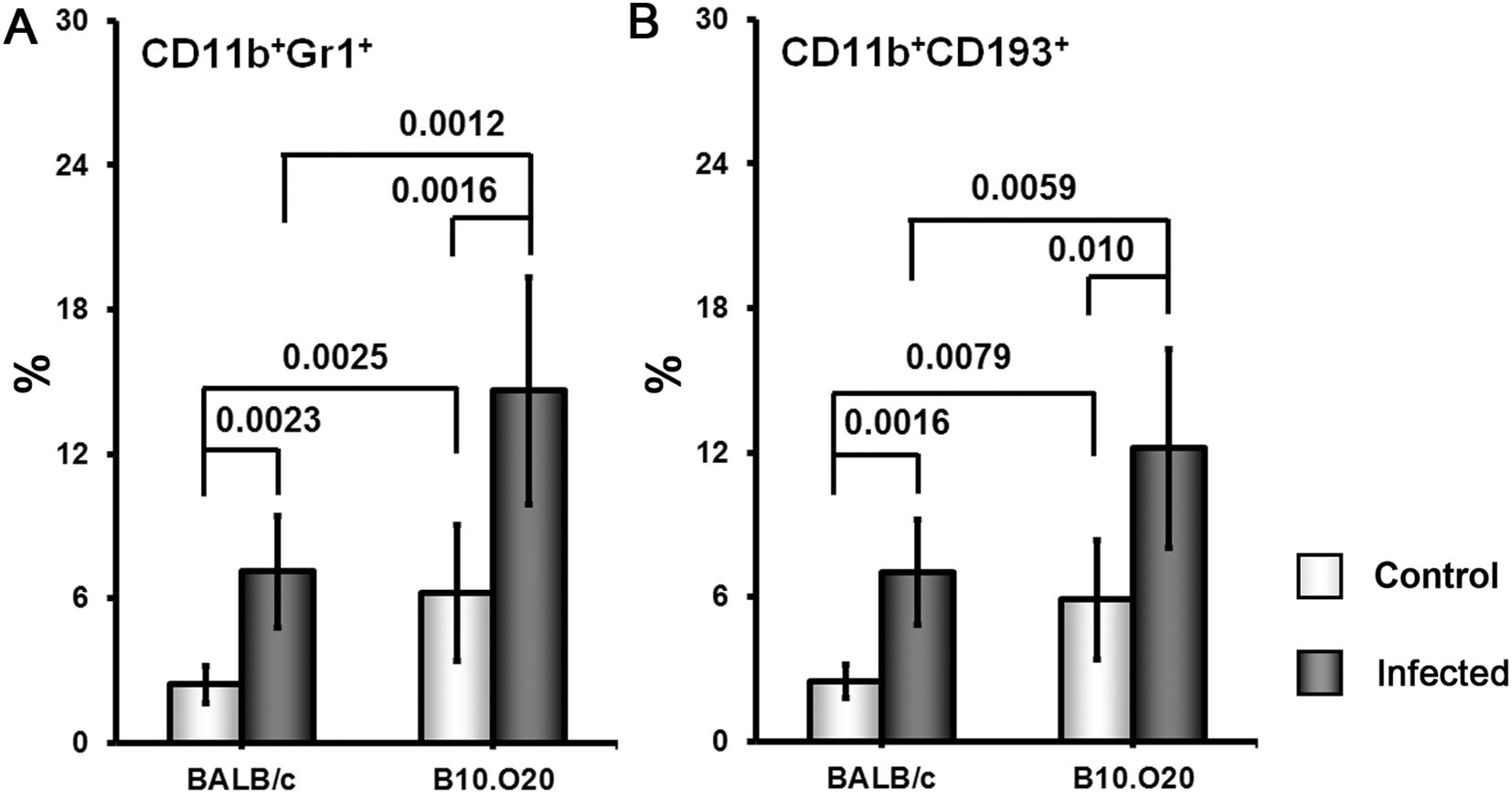
Figure 6. Comparison of frequency of CD11b+Gr1+ (A) and CD11b+CD193+ (B) cells in spleens of the strains B10.O20 and BALB/c. Female mice of strains BALB/c (8 infected, 5 uninfected) and B10.O20 (7 infected, 7 uninfected) were analyzed by flow cytometry. Dead cells were stained using Hoechst 33258 fluorescent dye added to the samples 10 min before flow cytometry analysis. Cells were stained with allophycocyanine (APC)-labeled anti-CD11b (clone M1/70), fluorescein isothiocyanate (FITC)-labeled anti-Ly6G/Ly-6C (Gr-1) (clone RB6-8C5) and phycoerythrin-labeled anti-CD193 (clone J073E5) antibodies as described in Materials and Methods. Selected cell populations were gated on single cell events after exclusion of cell debris and dead cells. Figure shows means ± SD from 2 independent experiments and a representative FACS experiment.
Spleens of infected B10.O20 mice exhibited also higher frequency of monocyte derived CD14+ (Supplementary Figure 2A) and F4/80+ (Supplementary Figure 2B) subpopulations and a lower frequency of T cell helper CD3+CD4+ (Supplementary Figure 2C), T cell cytotoxic CD3+CD8+ (Supplementary Figure 2D) and B lymphocyte CD19+CD22+ (Supplementary Figure 2E) subpopulations than both B10 and O20. B10.O20 spleens display a higher frequency of cells carrying T cell stimulation receptor CD86+ (Supplementary Figure 2F) than O20 and lower than B10. However, these differences were less pronounced than differences in CD11b+Gr1+ and CD11b+CD193+ subpopulations, and except CD19+CD22+ subpopulation were already present in uninfected mice (Supplementary Figure 2).
Infected mice did not show a significant increase of relative liver weight after infection (Figure 1F). The parasite load in liver of B10.O20 exhibited “B10-like” responses. Both B10.O20 (P = 0.00015) and B10 (P = 0.00021) exhibit higher parasite load in liver than O20 (Figure 1G). B10.O20 exhibited “B10-like” responses also in other liver parameters. Both B10 and B10.O20 had larger number and size of granulomas than O20 (Supplementary Figures 3A, B), and more macrophages (Supplementary Figure 3C), eosinophils (Supplementary Figure 3D), neutrophils (Supplementary Figure 3E) and lymphocytes (Supplementary Figure 3F) in granulomas. However, none of the histological liver parameters (except neutrophil content in granulomas, which was higher in infected than in uninfected B10.O20 mice (P = 0.030)) changed after infection and they mirrored differences in granulomas numbers, size and composition already present in uninfected mice (Supplementary Figure 3).
Soluble Leishmania antigen stimulated high cytokine production in the susceptible strain B10.O20 and intermediate in resistant strain B10, which contrasted with low production in the resistant strain O20
Splenocytes of uninfected mice of all strains produced low or undetectable levels of tested cytokines after stimulation with SLA. Infection of the strain B10.O20 resulted in 15-350× increase of splenocytes’ production of IFNγ (Figure 7A) IL-2 (Figure 7B), IL-4 (Figure 7C), IL-6 (Figure 7D), IL-10 (Figure 7E) and IL-17 (Figure 7F). Cells of the strain B10 also exhibit increased production of IFNγ IL-2, IL-6, IL-10 and IL-17, but not to the extent of the strain B10.O20 (Figure 7). The splenocytes of the resistant strain O20 practically do not respond to SLA by production of cytokines. The production of IFNγ, IL-4 and IL-17 by O20 is undetectable and production of IL-2, IL-10 and IL-6 by this strain was very low.
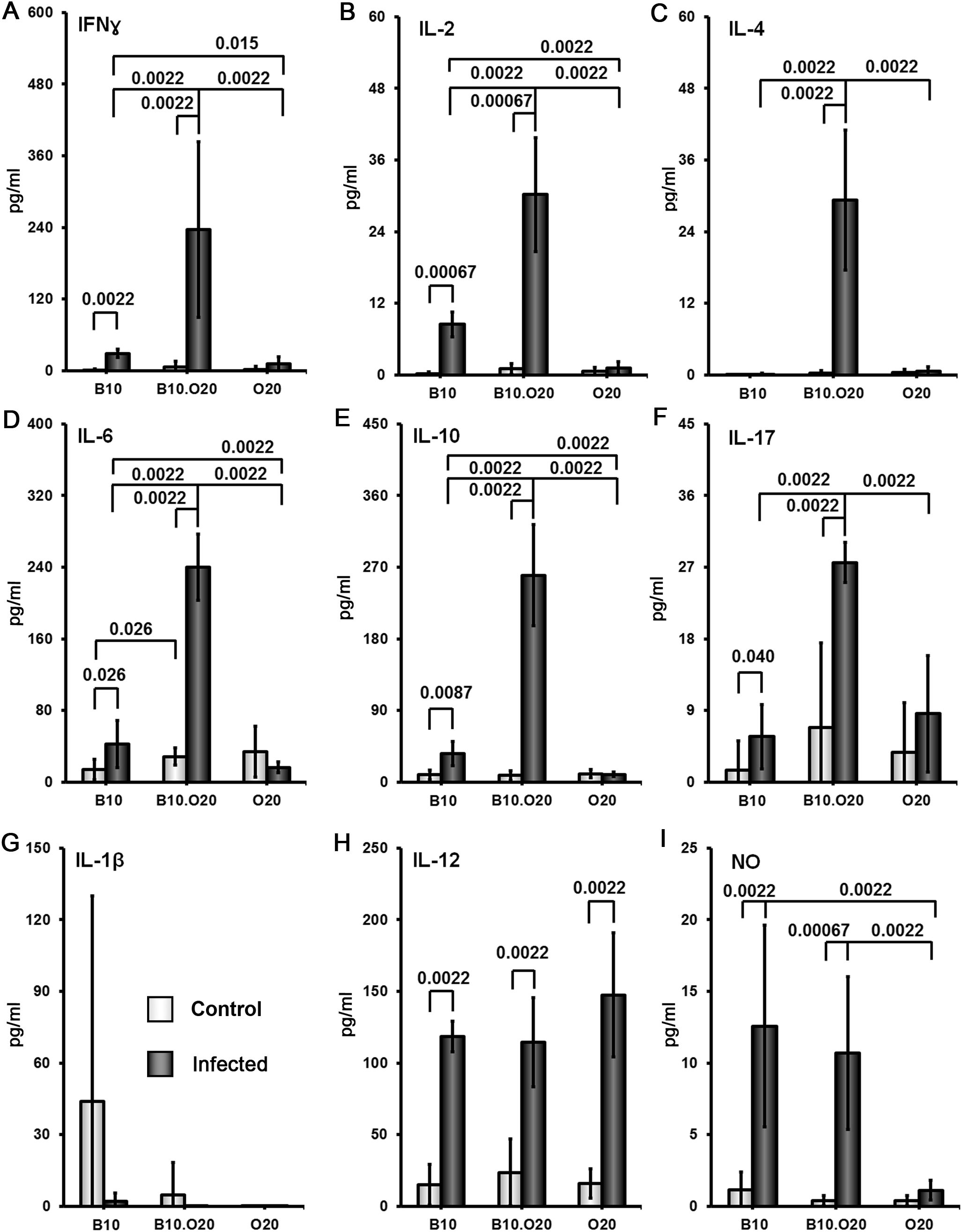
Figure 7. Comparison of cytokine and NO response to the soluble Leishmania antigen (SLA). Female mice of strains O20, B10 and B10.O20 were compared. At least 6 uninfected and 6 infected mice from each strain were analyzed. Production of IFNγ (A), IL-2 (B), IL-4 (C), IL-6 (D), IL-10 (E) and IL-17 (F) by splenocytes and production of IL-1β (G), IL-12 (H) and NO (I) by peritoneal exudate cells was determined. The data show the means ± SD from 5 independent experiments.
After SLA stimulation peritoneal macrophages of O20 mice do not produce NO, in contrast to B10 and B10.O20 macrophages (Figure 7I), but there was no difference in production of IL-1β (Figure 7G) and IL-12 (Figure 7H) by SLA-stimulated macrophages among the tested strains. Macrophages of the resistant strain B10 and susceptible strain B10.O20 did not differ in production of NO. Surprisingly, macrophages of the resistant strain O20 produce IL-12, but not IL-1β and NO (Figures 7G–I).
Surprising lack of cytokine response to SLA by the strain O20 prompted us to compare it with another Leishmania resistant strain STS and the widely studied Leishmania susceptible strain BALB/c. All these strains contain parasites in their spleens (Figure 8). Similarly as in previous tests, splenocytes of uninfected BALB/c, STS and O20 mice produce low or undetectable levels of tested cytokines after stimulation with SLA. Infection of BALB/c and STS mice resulted in 10-500× increase of production of IFNγ (Figure 9A) IL-2 (Figure 9B), IL-4 (Figure 9C), IL-6 (Figure 9D) and IL-10 (Figure 9E) by their splenocytes, whereas strain O20 splenocytes did not produce any of these cytokines except IL-6 (Figure 9D). None of the tested strains showed significant increase in production of IL-17 (Figure 9F) by splenocytes and increase of IL-1β by peritoneal macrophages (Figure 9G). The increase of NO by peritoneal macrophages was observed only in the strain STS (Figure 9H).
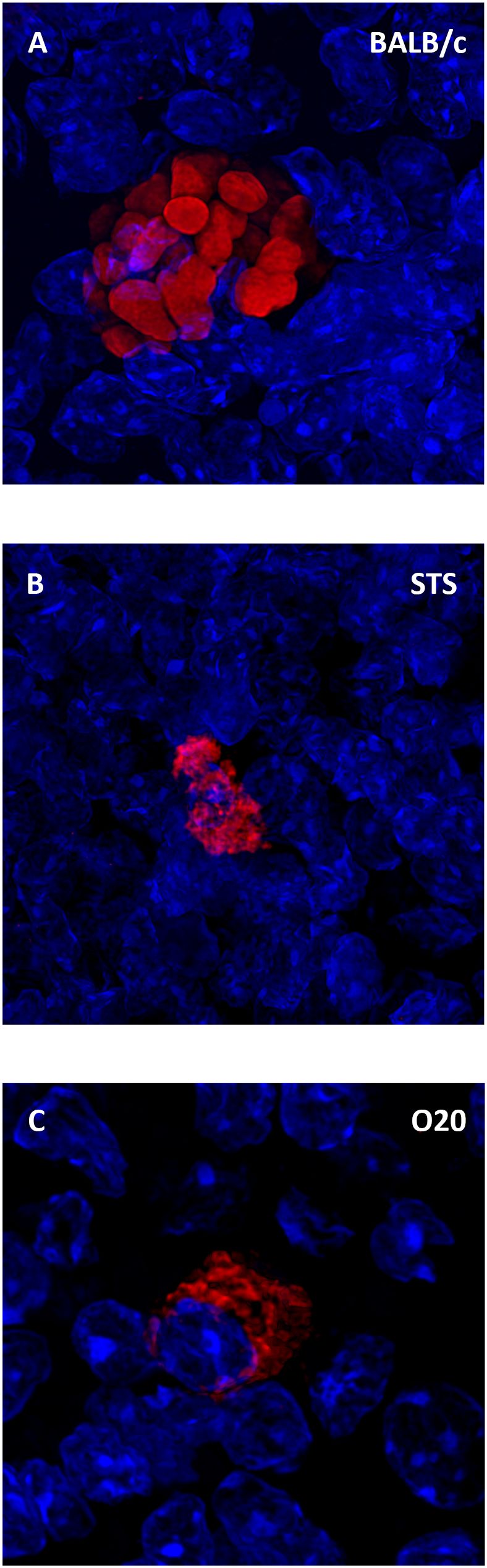
Figure 8. Leishmania major parasites inside the spleen. Slices of spleen tissue of females of BALB/c (A), STS (B) and O20 (C) mice were stained with the anti-Leishmania lipophosphoglycan mouse monoclonal antibody (cat. no. CLP003A, Cedarlane, Hornby, Canada) and TRITC labelled IgM (115-025-020, Jackson ImmunoResearch, West Grove, PA) all diluted 1:500. Nuclei of the cells were stained with Bisbenzimide H33258 (Sigma-Aldrich, St. Louis, MO) 10 mg per 1 ml diluted 1:1000. Images were captured with confocal microscope Leica TCS SP8 objective HC PL APO 63×/1.40 OIL.
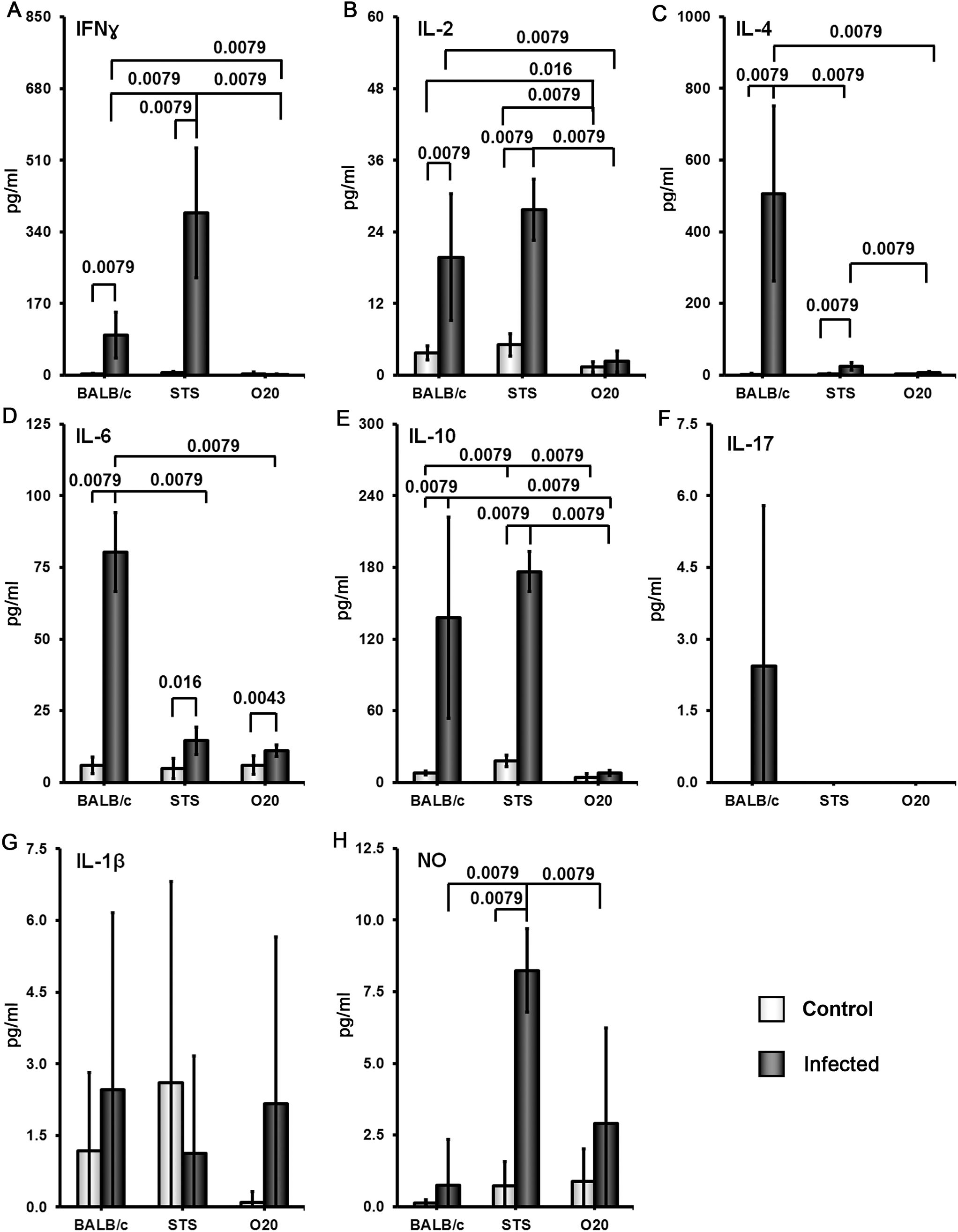
Figure 9. Comparison of cytokine and NO response to the soluble Leishmania antigen (SLA).Female mice of the strains O20, BALB/c and STS were compared. At least 5 uninfected and 6 infected mice from each strain were analyzed. Production of IFNγ (A), IL-2 (B), IL-4 (C), IL-6 (D), IL-10 (E) and IL-17 (F) by splenocytes and production of IL-1β (G) and NO (H) by peritoneal exudate cellswas estimated. The data show the means ± SD from 2 independent experiments.
In summary, we have analyzed two susceptible strains, B10.O20 and BALB/c, and three resistant strains: B10, O20 and STS. These five strains exhibited five different patterns of splenocytes SLA response. B10.O20: high levels of IFNγ, IL-2, IL-4, IL-6, IL-10 and IL-17; BALB/c: high levels of IL-2, IL-4, IL-6, IL-10 and intermediate levels of IFNγ and IL-17; STS: high levels of IFNγ, IL-2 and IL-10, low to intermediate level of IL-4, intermediate level of IL-6 and undetectable level of IL-17, B10: intermediate level of IFNγ, IL-2, IL-6, IL-10 and IL-17 and undetectable level of IL-4; O20 did not contain detectable concentration of tested cytokines with the exception of IL-6.
Discussion
We show that the combination of genes of two resistant strains, O20 and B10, which is present in the strain B10.O20, unexpectedly results in its susceptibility and extremely high numbers of CD11b+Gr1+ cells in B10.O20 mice before and even more after infection. In addition, we observed in strains O20 and B10.O20 several previously unreported phenotypes, namely a high resistance to infection in O20 mice accompanied by non-responsiveness to the SLA antigen. In view of the potential conceptual importance of the responses to L. major in these strains, we report in detail these immunological and immunopathological phenotypes. The parameters that appear to be most closely associated with response to L. major are given in the main body of the paper, those not altered after infection are given in detail in Supplementary Figures.
Strong combinatorial genetic influence on response to Leishmania infection
We have observed that combination of genes of two resistant strains gave rise to a susceptible strain. There might be several causes of this phenomenon: a) epistasis, b) combination of effects of minor “susceptibility” alleles of resistant strains, c) combination of a) and b).
a) Epistasis. Influence of genetic background on phenotypic effects was discovered in studies of many genes disrupted by targeting: for example Igf1r (insulin-like growth factor I receptor) (63), Egfr (epidermal growth factor receptor) (64), Fcgr2b (Fc receptor, IgG, low affinity Iib) (65), as well as on naturally mutated genes Map3k14 (mitogen-activated protein kinase kinase kinase 14) (66) and polymorphic haplotypes H2b, H2d (67), and H2d, H2k (68). These effects are attributable to modifier genes, which act in combination with the causative gene, differ on different genetic backgrounds and were identified only in few cases (68).
We have previously identified loci/genes that function differently on B10 background in control of frequency of CD11b+Gr1+ subpopulation in spleens of uninfected B10.O20 (55). This parameter is controlled by three loci Mydc1 (myeloid-derived cells 1), Mydc2 and Mydc3 on chromosomes 1, 15 and 17, respectively. We identified potential candidate genes for Mydc1 (Smap1) and for Mydc3 (Vps52, Tnxb and Rab44). Mydc1 exhibits single gene effect, whereas the influence of Mydc2 and Mydc3 was observed only in their interaction. In all three loci, the alleles controlling higher frequency of CD11b+Gr1+ cells were of O20 origin (55). Thus, the transfer of O20 alleles on B10 background led to increase of frequency CD11b+Gr1+ cells.
b) A limited number of non-overlapping loci with susceptible alleles in O20 and B10. These two sets of “susceptibility” alleles that became by chance accumulated in B10.O20 strain and their combined effect prevails over “resistant” alleles of B10 and O20. These “susceptibility” alleles can work additively or exhibit genetic epistasis.
Novel L. major response phenotypes in O20 and B10.O20 mice
O20 and B10.O20 mice exhibit resistant and susceptible phenotypes, respectively, that have not been observed before.
The strain O20 does not develop skin lesions (Figure 1A) and it is more resistant to L. major than the widely used resistant strains B6 and 129, which develop localized transient skin lesions that heal spontaneously (69–71), and it is also more resistant than the strain STS, which develops none, or only small localized lesions (45, 57). O20 is also more resistant than the strain B10, as described in detail in Results (Figures 1B, 2A). However, the mechanisms of resistance of O20 are different. Similarly as the B10 and B10.O20, O20 does not exhibit splenomegaly or hepatomegaly and does not differ from these strains in parasite load in spleen (Figures 1D–F), but it has more preserved white pulp (Figures 3A, C, E). Surprisingly, spleen cells of the infected O20 mice practically do not respond to SLA by cytokine production (Figures 7A–F), but its peritoneal macrophages respond to SLA by IL-12 production (Figure 7H). This cytokine response differs from other resistant strains: B6, B10 and 129 mice respond to SLA by high production of IFNγ and low production of IL-4 (69–71 and this paper). However, macrophages of O20 do not respond to SLA by production of NO (Figure 7I). NO induction is more difficult to induce in humans than in most tested rodents (72), thus the strain O20 is more “human-like” in this trait. As mentioned above, the resistant strain B10 responds to SLA with increase of production of IFNγ, but it also exhibits increase of IL-10 and IL-17 that were described to induce immunopathology (32, 73); however, increase of levels of these cytokines was less expressed than increase of IFNγ (Figure 7). Response of the resistant strain STS is also characterized by high and low production of IFNγ (Figure 9A) and IL-4 (Figure 9C), respectively; similarly as B10, it also responds by increase of IL-10 level (Figure 9E). The dichotomy in IFNγ response to SLA in the strain O20, and the strains B6, B10, 129 and STS resembles dichotomy observed in human responses to L. donovani antigens (74). Thus, analysis of more mouse strains might better model the heterogeneity of human responses to leishmaniasis.
The strain B10.O20 is susceptible, but to a lesser degree than the strain BALB/c. In the same experimental design, BALB/c mice suffer from approximately twice as large skin lesions as B10.O20 and develop extensive splenomegaly and hepatomegaly (57). Splenocytes of infected susceptible B10.O20 mice produce Th1, Th2 and Th17 cytokines in response to SLA. They have increased production of Th1 cytokine IFNγ which is associated with resolution of the disease (33), as well as increase of Th2 cytokines IL-4 and IL-10, and Th17 cytokine IL-17 (Figures 7A–F), that were described to correlate with aggravation of disease (32, 33, 73). This mixed cytokine response is similar to responses observed in multiple studies of human cutaneous (75, 76) and visceral (74, 77, 78) leishmaniasis, where patients exhibit both Th1 and Th2 responses.
Another unexpected feature of response of B10.O20 mice to L. major infection is an exceptionally high increase of CD11b+Gr1+ and CD11b+CD193+ subpopulations. The frequencies of these subpopulations were even higher in B10.O20 than in the prototype susceptible BALB/c mice (Figure 6). The myeloid-derived CD11b+Gr1+ subpopulation has been found to play role in cancer, autoimmune diseases, traumatic stress and transplantation [reviewed in (79)], as well as in infectious diseases, including leishmaniasis. Gr1+ monocytes from B6. but not BALB/c mice have been found to kill L. major parasites in the early stages of infection (80). Their role in the chronic disease has to be elucidated. We have also found increased eosinophilic infiltration in spleens of the infected B10.O20 mice (Figure 5). The future analysis of the phenomena described in this subsection needs to comprise also time-course studies, which are beyond the scope of the present paper.
The tested strains differed prior to infection in relative spleen weight (Figure 1D), frequencies in cell subpopulations in the spleen (Figure 4; Supplementary Figure 2) and in liver (Supplementary Figure 3). The heritable differences in gene expression (81) and metabolism (82) present prior to infection may contribute to differential susceptibility to Bordetella pertussis and L. donovani, respectively. Future studies leading to identification of genes controlling baseline parameters and genes determining susceptibility to L. major and their comparison will elucidate the role of baseline differences observed in the present study on response to the infection.
Asymptomatic response
Encounter of host with pathogen can lead to no entrance or clearance of pathogen, to asymptomatic infection when host mounts a response strong enough to control pathogen, and to a symptomatic disease (83). Asymptomatic infections by viruses, bacteria and parasites are widespread. Data about the proportion of asymptomatic individuals from the first years of SARS-Cov19 epidemics varies from 1.2% in Wuhan, China (84) to 17.9% on the board of the Diamond Princess cruise ship (85). Asymptomatic norovirus prevalence was estimated to be about 7% (86). Kendall and coworkers indicated that 30% of people diagnosed through community-based testing were completely asymptomatic for Mycobacterium tuberculosis (87). 37.5% of Plasmodium falciparum infections and 18.5% of P. vivax infections in Brazil were asymptomatic when microscopy was used for diagnosis (88). Infection with Toxoplasma gondii in developed countries is asymptomatic in 80% of newly infected patients (89).
The outcome of an interplay between pathogen and host depends on many factors such as pathogen density and properties, host immunity, underlying medical conditions, as well as on age, immune and hormonal status, sex, nutrition (90, 91), and genetic background of the host (23, 92–94). Mouse experiments in controlled conditions allow to study an influence of these individual factors. In this paper, we show in mouse experiments that two different genotypes (O20 and STS) lead to asymptomatic infection by different mechanisms.
Conclusion
The present report demonstrates the benefits of using more mouse strains to analyze disease susceptibility in order to better reflect the diverse immunological characteristics of the human population. We have observed strong effect of small percentage of genes of the resistant strain O20 on the genetic background of another resistant strain B10 in control of susceptibility to L. major. This is accompanied by novel phenotypes of susceptible and resistant strains exhibiting heterogeneity of defensive strategies, which resemble certain human disease phenotypes. Analysis of these novel phenotypes can help to develop new therapies and vaccines designed to treat different genotypes. Moreover, these studies could serve as a basis for the investigation of asymptomatic responses to leishmaniasis, but also to other infectious diseases.
Data availability statement
The original contributions presented in the study are included in the article/Supplementary Material. Further inquiries can be directed to the corresponding author.
Ethics statement
All experimental protocols utilized in this study comply with the Czech Government Requirements under the Policy of Animal Protection Law (No.246/1992) and with the regulations of the Ministry of Agriculture of the Czech Republic (No.207/2004), which are in agreement with all relevant European Union guidelines for work with animals. The study was approved by the Institutional Animal Care Committee of the Institute of Molecular Genetics of the Czech Academy of Sciences and by Departmental Expert Committee for the Approval of Projects of Experiments on Animals of the Czech Academy of Sciences (permission Nr. 93/2015). The study was conducted in accordance with the local legislation and institutional requirements.
Author contributions
YS: Conceptualization, Data curation, Investigation, Methodology, Writing – review & editing. TK: Data curation, Investigation, Methodology, Visualization, Writing – review & editing. VV: Investigation, Writing – review & editing, Data curation, Methodology, Software. EJ: Data curation, Investigation, Methodology, Writing – review & editing. IK: Data curation, Investigation, Visualization, Writing – review & editing. AZ: Investigation, Methodology, Writing – review & editing. HH: Investigation, Methodology, Writing – review & editing. MS: Investigation, Writing – review & editing, Methodology. VH: Conceptualization, Investigation, Writing – review & editing, Methodology. PD: Conceptualization, Data curation, Investigation, Writing – review & editing. ML: Conceptualization, Data curation, Funding acquisition, Investigation, Project administration, Supervision, Writing – original draft, Writing – review & editing.
Funding
The author(s) declare financial support was received for the research and/or publication of this article. This article is based upon work from the COST Action BM1404 Mye-EUNITER (https://www.cost.eu/actions/BM1404/), supported by COST (European Cooperation in Science and Technology). It was also supported by the Charles University Program COOPERATIO: the scientific project “Medical Diagnostics and Basic Medical Sciences” (the field “Medical Genetics”).
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fimmu.2025.1579257/full#supplementary-material
References
1. Alvar J, Velez ID, Bern C, Herrero M, Desjeux P, Cano J, et al. Leishmaniasis worldwide and global estimates of its incidence. PloS One. (2012) 7:e35671. doi: 10.1371/journal.pone.0035671
2. Burza S, Croft SL, and Boelaert M. Leishmaniasis. Lancet. (2018) 392:951–70. doi: 10.1016/S0140-6736(18)31204-2
3. WHO. Available online at: https://www.who.int/health-topics/leishmaniasistab=tab_1 (Accessed November 12, 2024).
4. Bamorovat M, Sharifi I, Khosravi A, Aflatoonian MR, Agha Kuchak Afshari S, Salarkia E, et al. Global dilemma and needs assessment toward achieving sustainable development goals in controlling leishmaniasis. J Epidemiol Glob Health. (2024) 14:22–34. doi: 10.1007/s44197-024-00190-z
5. Bogdan C. Mechanisms and consequences of persistence of intracellular pathogens: leishmaniasis as an example. Cell Microbiol. (2008) 10:1221–34. doi: 10.1111/j.1462-5822.2008.01146.x
6. Terrazas CA, Terrazas LI, and Gómez-García L. Modulation of dendritic cell responses by parasites: a common strategy to survive. J BioMed Biotechnol. (2010) 2010:357106. doi: 10.1155/2010/357106
7. Schwing A, Pisani DF, Pomares C, Majoor A, Lacas-Gervais S, Jager J, et al. Identification of adipocytes as target cells for Leishmania infantum parasites. Sci Rep. (2021) 11:21275. doi: 10.1038/s41598-021-00443-y
8. Dedet J-P. Current status of epidemiology of leishmaniases. In: Farrel JP, editor. Leishmania. Springer USA, Boston, MA (2002). p. 1–10.
9. Lainson R and Shaw JJ. Evolution, classification and geographical distribution. In: Peters W and Killick-Kendrick R, editors. The leishmaniases in biology and medicine. Volume I. Biology and Epidemiology. Academic Press, London (1987), ISBN: ISBN 0 12 552101 4. p. 120.
10. Kobets T, Grekov I, and Lipoldova M. Leishmaniasis: prevention, parasite detection and treatment. Curr Med Chem. (2012) 19:1443–74. doi: 10.2174/092986712799828300
11. King A. In search of a vaccine for leishmaniasis. Nature. (2023). doi: 10.1038/d41586-023-02580-y
12. Mandell MA and Beverley SM. Continual renewal and replication of persistent Leishmania major parasites in concomitantly immune hosts. Proc Natl Acad Sci. (2017) 114:E801–10. doi: 10.1073/pnas.1619265114
13. Hasker E, Kansal S, Malaviya P, Gidwani K, Picado A, Singh RP, et al. Latent infection with Leishmania donovani in highly endemic villages in Bihar, India. PloS Negl Trop Dis. (2013) 7:e2053. doi: 10.1371/journal.pntd.0002053
14. Singh OP and Sundar S. Whole blood assay and visceral leishmaniasis: Challenges and promises. Immunobiology. (2014) 219:323–8. doi: 10.1016/j.imbio.2014.01.005
15. Singh OP, Hasker E, Sacks D, Boelaert M, and Sundar S. Asymptomatic Leishmania infection: a new challenge for Leishmania control. Clin Infect Dis. (2014) 58:1424–9. doi: 10.1093/cid/ciu102
16. Rock KS, Chapman LAC, Dobson AP, Adams ER, and Hollingsworth TD. The hidden hand of asymptomatic infection hinders control of neglected tropical diseases: A modeling analysis. Clin Infect Dis. (2024) 78:S175–82. doi: 10.1093/cid/ciae096
17. Aebischer T. Recurrent cutaneous leishmaniasis: a role for persistent parasites? Parasitol Today. (1994) 10:25–8. doi: 10.1016/0169-4758(94)90353-0
18. Tuon FF, Bombonatto GM, Battaglin ER, Sakumoto MH, Amato VS, de Camargo RA, et al. Reactivation of mucosal and cutaneous leishmaniasis in a renal transplanted patient. Am J Trop Med Hyg. (2014) 91:81–3. doi: 10.4269/ajtmh.13-0578
19. Lipoldová M and Demant P. Genetic susceptibility to infectious disease: lessons from mouse models of leishmaniasis. Nat Rev Genet. (2006) 7:294–305. doi: 10.1038/nrg1832
20. Oryan A and Akbari M. Worldwide risk factors in leishmaniasis. Asian Pac J Trop Med. (2016) 9:925–32. doi: 10.1016/j.apjtm.2016.06.021
21. Blackwell JM, Fakiola M, and Castellucci LC. Human genetics of Leishmania infections. Hum Genet. (2020) 139:813–9. doi: 10.1007/s00439-020-02130-w
22. Lipoldová M and Demant P. Gene-specific sex effects on susceptibility to infectious diseases. Front Immunol. (2021) 12:712688. doi: 10.3389/fimmu.2021.712688
23. Krayem I and Lipoldová M. Role of host genetics and cytokines in Leishmania infection. Cytokine. (2021) 147:155244. doi: 10.1016/j.cyto.2020.155244
24. Bharati K. Human genetic polymorphism and Leishmaniasis. Infect Genet Evol. (2022) 98:105203. doi: 10.1016/j.meegid.2021.105203
25. Krayem I, Sohrabi Y, Havelková H, Gusareva ES, Strnad H, Čepičková M, et al. Functionally distinct regions of the locus Leishmania major response 15 control IgE or IFNγ level in addition to skin lesions. Front Immunol. (2023) A14:1145269. doi: 10.3389/fimmu.2023.1145269
26. Sacks D and Noben-Trauth N. The immunology of susceptibility and resistance to Leishmania major in mice. Nat Rev Immunol. (2002) 2:845–58. doi: 10.1038/nri933
27. Sakthianandeswaren A, Foote SJ, and Handman E. The role of host genetics in leishmaniasis. Trends Parasitol. (2009) 25:383–91. doi: 10.1016/j.pt.2009.05.004
28. Loeuillet C, Bañuls AL, and Hide M. Study of Leishmania pathogenesis in mice: experimental considerations. Parasit Vectors. (2016) 9:144. doi: 10.1186/s13071-016-1413-9
29. Bogdan C. Macrophages as host, effector and immunoregulatory cells in leishmaniasis: Impact of tissue micro-environment and metabolism. Cytokine X. (2020) 2:100041. doi: 10.1016/j.cytox.2020.100041
30. Lipoldová M and Sohrabi Y. Role of interferon-induced GTPases in leishmaniasis. PloS Negl Trop Dis. (2022) 16:e0010093. doi: 10.1371/journal.pntd.0010093
31. Mock BA, Fortier AH, Potter M, and Nacy CA. Genetic control of systemic Leishmania major infections: dissociation of intrahepatic amastigote replication from control by the Lsh gene. Infect Immun. (1985) 50:588–91. doi: 10.1128/iai.50.2.588-591.1985
32. Sacks D and Anderson C. Re-examination of the immunosuppressive mechanisms mediating non-cure of Leishmania infection in mice. Immunol Rev. (2004) 201:225–38. doi: 10.1111/j.0105-2896.2004.00185.x
33. Heinzel FP, Sadick MD, Holaday BJ, Coffman RL, and Locksley RM. Reciprocal expression of interferon gamma or interleukin 4 during the resolution or progression of murine leishmaniasis. Evidence for expansion of distinct helper T cell subsets. J Exp Med. (1989) 169:59–72. doi: 10.1084/jem.169.1.59
34. Liew FY. Functional heterogeneity of CD4+ T cells in leishmaniasis. Immunol Today. (1989) 10:40–5. doi: 10.1016/0167-5699(89)90302-2
35. Liew FY, Millott S, Parkinson C, Palmer RM, and Moncada S. Macrophage killing of Leishmania parasite in vivo is mediated by nitric oxide from L-arginine. J Immunol. (1990) 144:4794–7. doi: 10.4049/jimmunol.144.12.4794
36. Shankar AH and Titus RG. T cell and non-T cell compartments can independently determine resistance to Leishmania major. J Exp Med. (1995) 181:845–55. doi: 10.1084/jem.181.3.845
37. Nylén S and Gautam S. Immunological perspectives of leishmaniasis. J Glob Infect Dis. (2010) 2:135–46. doi: 10.4103/0974-777X.62876
38. Ashok D and Acha-Orbea H. Timing is everything: dendritic cell subsets in murine Leishmania infection. Trends Parasitol. (2014) 30:499–507. doi: 10.1016/j.pt.2014.08.001
39. Moll H. Epidermal Langerhans cells are critical for immunoregulation of cutaneous leishmaniasis. Immunol Today. (1993) 14:383–7. doi: 10.1016/0167-5699(93)90138-B
40. Charmoy M, Brunner-Agten S, Aebischer D, Auderset F, Launois P, Milon G, et al. Neutrophil-derived CCL3 is essential for the rapid recruitment of dendritic cells to the site of Leishmania major inoculation in resistant mice. PloS Pathog. (2010) 6:e1000755. doi: 10.1371/journal.ppat.1000755
41. Tacchini-Cottier F, Zweifel C, Belkaid Y, Mukankundiye C, Vasei M, Launois P, et al. An immunomodulatory function for neutrophils during the induction of a CD4+ Th2 response in BALB/c mice infected with Leishmania major. J Immunol. (2000) 165:2628–36. doi: 10.4049/jimmunol.165.5.2628
42. Ehrchen JM, Roebrock K, Foell D, Nippe N, von Stebut E, Weiss JM, et al. Keratinocytes determine Th1 immunity during early experimental leishmaniasis. PloS Pathog. (2010) 6:e1000871. doi: 10.1371/journal.ppat.1000871
43. Schmid M, Zimara N, Wege AK, and Ritter U. Myeloid-derived suppressor cell functionality and interaction with Leishmania major parasites differ in C57BL/6 and BALB/c mice. Eur J Immunol. (2014) 44:3295–306. doi: 10.1002/eji.201344335
44. Rashidi S, Mansouri R, Ali-Hassanzadeh M, Ghani E, Barazesh A, Karimazar M, et al. Highlighting the interplay of microRNAs from Leishmania parasites and infected-host cells. Parasitology. (2021) 148:1434–46. doi: 10.1017/S0031182021001177
45. Demant P, Lipoldova M, and Svobodova M. Resistance to Leishmania major in mice. Science. (1996) 274:1392a. doi: 10.1126/science.274.5291.1392a
46. Beebe AM, Mauze S, Schork NJ, and Coffman RL. Serial backcross mapping of multiple loci associated with resistance to Leishmania major in mice. Immunity. (1997) 6:551–7. doi: 10.1016/s1074-7613(00)80343-x
47. Roberts LJ, Baldwin TM, Curtis JM, Handman E, and Foote SJ. Resistance to Leishmania major is linked to the H2 region on chromosome 17 and to chromosome 9. J Exp Med. (1997) 185:1705–10. doi: 10.1084/jem.185.9.1705
48. Slapničková M, Volkova V, Čepičková M, Kobets T, Šíma M, Svobodová M, et al. Gene-specific sex effects on eosinophil infiltration in leishmaniasis. Biol Sex Differ. (2016) 7:59. doi: 10.1186/s13293-016-0117-3
49. Kobets T, Čepičková M, Volkova V, Sohrabi Y, Havelková H, Svobodová M, et al. Novel loci controlling parasite load in organs of mice infected with Leishmania major, their interactions and sex influence. Front Immunol. (2019) 10:1083. doi: 10.3389/fimmu.2019.01083
50. Kedzierski L and Evans KJ. Immune responses during cutaneous and visceral leishmaniasis. Parasitology. (2014) 141(12):1544–62. doi: 10.1017/S003118201400095X
51. Mohan S, Revill P, Malvolti S, Malhame M, Sculpher M, and Kaye PM. Estimating the global demand curve for a leishmaniasis vaccine: A generalisable approach based on global burden of disease estimates. PloS Negl Trop Dis. (2022) 16:e0010471. doi: 10.1371/journal.pntd.0010471
52. Nagill R and Kaur S. Vaccine candidates for leishmaniasis: a review. Int Immunopharmacol. (2011) 11:1464–88. doi: 10.1016/j.intimp.2011.05.008
53. Silver LM. Mouse genetics: concepts and applications. Oxford, New York: Oxford University Press (1995).
54. Stassen AP, Groot PC, Eppig JT, and Demant P. Genetic composition of the recombinant congenic strains. Mamm Genome. (1996) 7:55–8. doi: 10.1007/s003359900013
55. Krayem I, Sohrabi Y, Javorková E, Volkova V, Strnad H, Havelková H, et al. Genetic influence on frequencies of myeloid-derived cell subpopulations in mouse. Front Immunol. (2022) 12:760881. doi: 10.3389/fimmu.2021.760881
56. Grekov I, Svobodová M, Nohýnková E, and Lipoldová M. Preparation of highly infective Leishmania promastigotes by cultivation on SNB-9 biphasic medium. J Microbiol Methods. (2011) 87:273–7. doi: 10.1016/j.mimet.2011.08.012
57. Lipoldová M, Svobodová M, Krulová M, Havelková H, Badalová J, Nohýnková E, et al. Susceptibility to Leishmania major infection in mice: multiple loci and heterogeneity of immunopathological phenotypes. Genes Immun. (2000) 1:200–6. doi: 10.1038/sj.gene.6363660
58. Kobets T, Badalová J, Grekov I, Havelková H, Svobodová M, and Lipoldová M. Leishmania parasite detection and quantification using PCR-ELISA. Nat Protoc. (2010) 5:1074–80. doi: 10.1038/nprot.2010.68
59. Khamesipour A, Nateghi Rostami M, Tasbihi M, Miramin Mohammadi A, Shahrestani T, Sarrafnejad A, et al. Phenotyping of circulating CD8+ T cell subsets in human cutaneous leishmaniasis. Microbes Infect. (2012) 14:702–11. doi: 10.1016/j.micinf.2012.02.006
60. Holan V, Cervena T, Zajicova A, Hermankova B, Echalar B, Palacka K, et al. The impact of metal nanoparticles on the immunoregulatory and therapeutic properties of mesenchymal stem cells. Stem Cell Rev Rep. (2023) 19:1360–9. doi: 10.1007/s12015-022-10500-2
61. Green LC, Wagner DA, Glogowski J, Skipper PL, Wishnok JS, and Tannenbaum SR. Analysis of nitrate, nitrite, and [15N]nitrate in biological fluids. Anal Biochem. (1982) 126:131–8. doi: 10.1016/0003-2697(82)90118-x
62. Démant P and Hart AA. Recombinant congenic strains–a new tool for analyzing genetic traits determined by more than one gene. Immunogenetics. (1986) 24:416–22. doi: 10.1007/BF00377961
63. Xu J, Gontier G, Chaker Z, Lacube P, Dupont J, and Holzenberger M. Longevity effect of IGF-1R(+/-) mutation depends on genetic background-specific receptor activation. Aging Cell. (2014) 13:19–28. doi: 10.1111/acel.12145
64. Threadgill DW, Dlugosz AA, Hansen LA, Tennenbaum T, Lichti U, Yee D, et al. Targeted disruption of mouse EGF receptor: effect of genetic background on mutant phenotype. Science. (1995) 269:230–4. doi: 10.1126/science.7618084
65. Bolland S and Ravetch JV. Spontaneous autoimmune disease in Fc(gamma)RIIB-deficient mice results from strain-specific epistasis. Immunity. (2000) 13:277–85. doi: 10.1016/s1074-7613(00)00027-3
66. Shinzawa M, Maruyama Y, Qin J, Akiyama N, Miyauchi M, Yanai H, et al. Splenic extramedullary hemopoiesis caused by a dysfunctional mutation in the NF-κB-inducing kinase gene. Biochem Biophys Res Commun. (2011) 414:773–8. doi: 10.1016/j.bbrc.2011.10.001
67. Howard JG, Hale C, and Chan-Liew WL. Immunological regulation of experimental cutaneous leishmaniasis. 1. Immunogenetic aspects of susceptibility to Leishmania tropica in mice. Parasite Immunol. (1980) 2:303–14. doi: 10.1111/j.1365-3024.1980.tb00061.x
68. Desrosiers MP, Kielczewska A, Loredo-Osti JC, Adam SG, Makrigiannis AP, Lemieux S, et al. Epistasis between mouse Klra and major histocompatibility complex class I loci is associated with a new mechanism of natural killer cell-mediated innate resistance to cytomegalovirus infection. Nat Genet. (2005) 37:593–9. doi: 10.1038/ng1564
69. Guler R, Afshar M, Arendse B, Parihar SP, Revaz-Breton M, Leitges M, et al. PKCδ regulates IL-12p40/p70 production by macrophages and dendritic cells, driving a type 1 healer phenotype in cutaneous leishmaniasis. Eur J Immunol. (2011) 41:706–15. doi: 10.1002/eji.201040985
70. Heinzel FP, Rerko RM, Hujer AM, and Maier RA Jr. Increased capacity for interleukin-2 synthesis parallels disease progression in mice infected with Leishmania major. Infect Immun. (1998) 66:4537–40. doi: 10.1128/IAI.66.9.4537-4540.1998
71. Muraille E, De Trez C, Brait M, De Baetselier P, Leo O, Carlier Y, et al. Genetically resistant mice lacking MyD88-adapter protein display a high susceptibility to Leishmania major infection associated with a polarized Th2 response. J Immunol. (2003) 170:4237–41. doi: 10.4049/jimmunol.170.8.4237
72. Panaro MA, Brandonisio O, Acquafredda A, Sisto M, and Mitolo V. Evidences for iNOS expression and nitric oxide production in the human macrophages. Curr Drug Targets Immune Endocr Metabol Disord. (2003) 3:210–21. doi: 10.2174/1568008033340216
73. Lopez Kostka S, Dinges S, Griewank K, Iwakura Y, Udey MC, and von Stebut E. IL-17 promotes progression of cutaneous leishmaniasis in susceptible mice. J Immunol. (2009) 182:3039–46. doi: 10.4049/jimmunol.0713598
74. Khalil EA, Ayed NB, Musa AM, Ibrahim ME, Mukhtar MM, Zijlstra EE, et al. Dichotomy of protective cellular immune responses to human visceral leishmaniasis. Clin Exp Immunol. (2005) 140:349–53. doi: 10.1111/j.1365-2249.2005.02768.x
75. Kammoun-Rebai W, Naouar I, Libri V, Albert M, Louzir H, Meddeb-Garnaoui A, et al. Protein biomarkers discriminate Leishmania major-infected and non-infected individuals in areas endemic for cutaneous leishmaniasis. BMC Infect Dis. (2016) 16:138. doi: 10.1186/s12879-016-1458-6
76. Sassi A, Louzir H, Ben Salah A, Mokni M, Ben Osman A, and Dellagi K. Leishmanin skin test lymphoproliferative responses and cytokine production after symptomatic or asymptomatic Leishmania major infection in Tunisia. Clin Exp Immunol. (1999) 116:127–32. doi: 10.1046/j.1365-2249.1999.00844.x
77. Costa AS, Costa GC, Aquino DM, Mendonça VR, Barral A, Barral-Netto M, et al. Cytokines and visceral leishmaniasis: a comparison of plasma cytokine profiles between the clinical forms of visceral leishmaniasis. Mem Inst Oswaldo Cruz. (2012) 107:735–9. doi: 10.1590/s0074-02762012000600005
78. Peruhype-Magalhães V, Martins-Filho OA, Prata A, Silva Lde A, Rabello A, Teixeira-Carvalho A, et al. Mixed inflammatory/regulatory cytokine profile marked by simultaneous raise of interferon-gamma and interleukin-10 and low frequency of tumour necrosis factor-alpha(+) monocytes are hallmarks of active human visceral Leishmaniasis due to Leishmania chagasi infection. Clin Exp Immunol. (2006) 146:124–32. doi: 10.1111/j.1365-2249.2006.03171.x
79. Kong YY, Fuchsberger M, Xiang SD, Apostolopoulos V, and Plebanski M. Myeloid derived suppressor cells and their role in diseases. Curr Med Chem. (2013) 20:1437–44. doi: 10.2174/0929867311320110006
80. Pereira WF, Ribeiro-Gomes FL, Guillermo LV, Vellozo NS, Montalvão F, Dosreis GA, et al. Myeloid-derived suppressor cells help protective immunity to Leishmania major infection despite suppressed T cell responses. J Leukoc Biol. (2011) 90:1191–7. doi: 10.1189/jlb.1110608
81. Banus S, Vandebriel RJ, Pennings JL, Gremmer ER, Wester PW, van Kranen HJ, et al. Comparative gene expression profiling in two congenic mouse strains following Bordetella pertussis infection. BMC Microbiol. (2007) 7:88. doi: 10.1186/1471-2180-7-88
82. Ghosh J, Das S, Guha R, Ghosh D, Naskar K, Das A, et al. Hyperlipidemia offers protection against Leishmania donovani infection: role of membrane cholesterol. J Lipid Res. (2012) 53:2560–72. doi: 10.1194/jlr.M026914
83. Netea MG, Domínguez-Andrés J, van de Veerdonk FL, van Crevel R, Pulendran B, and van der Meer JWM. Natural resistance against infections: focus on COVID-19. Trends Immunol. (2021) 43(2):106–16. doi: 10.1016/j.it.2021.12.001
84. The Novel Coronavirus Pneumonia Emergency Response Epidemiology Team. The epidemiological characteristics of an outbreak of 2019 novel coronavirus diseases (COVID-19) - China, 2020. China CDC Wkly. (2020) 2:113–22.
85. Mizumoto K, Kagaya K, Zarebski A, and Chowell G. Estimating the asymptomatic proportion of coronavirus disease 2019 (COVID-19) cases on board the Diamond Princess cruise ship, Yokohama, Japan, 2020. Euro Surveill. (2020) 25:2000180. doi: 10.2807/1560-7917.ES.2020.25.10.2000180
86. Qi R, Huang YT, Liu JW, Sun Y, Sun XF, Han HJ, et al. Global prevalence of asymptomatic norovirus infection: A meta-analysis. EClinicalMedicine. (2018) 2-3:50–8. doi: 10.1016/j.eclinm.2018.09.001
87. Kendall EA, Kitonsa PJ, Nalutaaya A, Erisa KC, Mukiibi J, Nakasolya O, et al. The spectrum of tuberculosis disease in an urban Ugandan community and its health facilities. Clin Infect Dis. (2021) 72:e1035–e43. doi: 10.1093/cid/ciaa1824
88. da Silva-Nunes M and Ferreira MU. Clinical spectrum of uncomplicated malaria in semi-immune Amazonians: beyond the “ symptomatic “ vs “ asymptomatic “ dichotomy. Mem Inst Oswaldo Cruz. (2007) 102:341–7. doi: 10.1590/s0074-02762007005000051
89. Paris L. 106 - toxoplasmosis. In: Ryan ET, Hill DR, Solomon T, Aronson NE, and Endy TP, editors. Hunter’s Tropical Medicine and Emerging Infectious Diseases (Tenth Edition). Elsevier, London (2020). p. 803–13.
90. Laishram DD, Sutton PL, Nanda N, Sharma VL, Sobti RC, Carlton JM, et al. The complexities of malaria disease manifestations with a focus on asymptomatic malaria. Malar J. (2012) 11:29. doi: 10.1186/1475-2875-11-29
91. Lindblade KA, Steinhardt L, Samuels A, Kachur SP, and Slutsker L. The silent threat: asymptomatic parasitemia and malaria transmission. Expert Rev Anti Infect Ther. (2013) 11:623–39. doi: 10.1586/eri.13.45
92. Missinou MA, Lell B, and Kremsner PG. Uncommon asymptomatic Plasmodium falciparum infections in Gabonese children. Clin Infect Dis. (2003) 36:1198–202. doi: 10.1086/374555
93. Garcia G, Sierra B, Perez AB, Aguirre E, Rosado I, Gonzalez N, et al. Asymptomatic dengue infection in a Cuban population confirms the protective role of the RR variant of the FcgammaRIIa polymorphism. Am J Trop Med Hyg. (2010) 82:1153–6. doi: 10.4269/ajtmh.2010.09-0353
Keywords: leishmaniasis, host-pathogen interaction, mouse model, novel genetic mechanisms of disease susceptibility and resistance, CD11b + Gr1 + cells, asymptomatic leishmaniasis
Citation: Sohrabi Y, Kobets T, Volkova V, Javorková E, Krayem I, Zajícová A, Havelková H, Svobodová M, Holáň V, Demant P and Lipoldová M (2025) Non-additive strong gene interactions cause striking differences in organ pathology and cytokine response in Leishmaniasis. Front. Immunol. 16:1579257. doi: 10.3389/fimmu.2025.1579257
Received: 18 February 2025; Accepted: 13 June 2025;
Published: 14 October 2025.
Edited by:
Maria Carolina Accioly Brelaz de Castro, Federal University of Pernambuco, BrazilReviewed by:
Sara Passos, Century Therapeutics, United StatesAbhik Sen, Department of Health Research, India
Copyright © 2025 Sohrabi, Kobets, Volkova, Javorková, Krayem, Zajícová, Havelková, Svobodová, Holáň, Demant and Lipoldová. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Marie Lipoldová, bGlwb2xkb3ZhQGltZy5jYXMuY3o=; bWFyaWUubGlwb2xkb3ZhQGxmMy5jdW5pLmN6
†ORCID: Marie Lipoldová, orcid.org/0000-0002-2101-0334