 Alisson Amaral Da-Rocha1,2
Alisson Amaral Da-Rocha1,2 Júlio Souza Dos-Santos1,2Igor Santos Bittencourt1,2Douglas B. De-Almeida1,2Naiara Carla dos Santos Manhães1,2
Júlio Souza Dos-Santos1,2Igor Santos Bittencourt1,2Douglas B. De-Almeida1,2Naiara Carla dos Santos Manhães1,2 Hozany Praxedes1,2
Hozany Praxedes1,2 João Victor Paiva Romano1,2
João Victor Paiva Romano1,2 Elias Barbosa da Silva-Junior3Antônio José da Silva-Gonçalves4
Elias Barbosa da Silva-Junior3Antônio José da Silva-Gonçalves4 Marcia Pereira Oliveira4
Marcia Pereira Oliveira4 Luciana Polaco Covre5
Luciana Polaco Covre5 Daniel Claudio de Oliveira Gomes5
Daniel Claudio de Oliveira Gomes5 Alda Maria Da-Cruz4,6
Alda Maria Da-Cruz4,6 Alessandra Marcia da Fonseca-Martins1,2
Alessandra Marcia da Fonseca-Martins1,2 Herbert Leonel de Matos Guedes1,2*
Herbert Leonel de Matos Guedes1,2*- 1Instituto de Microbiologia Paulo de Góes – Universidade Federal do Rio de Janeiro, Rio de Janeiro, Rio de Janeiro, Brazil
- 2Laboratório de Imunologia Clínica, Instituto Oswaldo Cruz, Fundação Oswaldo Cruz, Rio de Janeiro, Brazil
- 3Instituto de Biofísica Carlos Chagas Filho – Universidade Federal do Rio de Janeiro, Rio de Janeiro, Rio de Janeiro, Brazil
- 4Laboratório Interdisciplinar de Pesquisas Médicas, Instituto Oswaldo Cruz, Fundação Oswaldo Cruz, Rio de Janeiro, Brazil
- 5Núcleo de Doenças Infecciosas – Universidade Federal do Espírito Santo, Vitória, Espírito Santo, Brazil
- 6Disciplina de Parasitologia, Faculdade de Ciências Médicas, UERJ, Rio de Janeiro, RJ, Brazil
Mucosal Leishmaniasis is one of the most aggressive clinical manifestations of Leishmania infection disease, characterized by the destruction of nasal and oral tissues. The mechanisms by which this disease occurs are still not well understood due to the lack of effective experimental models. Mucosal leishmaniasis is associated with inflammatory response, especially Th17 response. Based on that, in this work, the immunopathological aspects of the experimental infection of BALB/c mice by Leishmania amazonensis in the mucosa site were evaluated as this mice presents high susceptibility with increased Th17 mediated pathology. Three infection modes were performed and compared according to the injection site. Six weeks post infection, mice presented edema in the nasal and premaxillary region, with progressive growth until twelve weeks. The micro-Computerized Tomography and the histology images demonstrated that the parasite inoculation led to destruction of squamous and transitional tissues in NC and NB groups, with several cells harboring amastigotes. Mice infected in the mucosa tissues had higher parasite load and IgG, IgM antibody levels and increased production of cytotoxic mediators such as CD107, granzyme b and perforin, inflammatory cytokines as IFN-γ; and IL-17, but lower frequencies of CD4+ IL-10+ cells compared to ear dermis. Taken together, our data shows that L. amazonensis parasites are more proliferative in nasal mucosa and the infection leads to an increased inflammatory response compared to ear dermis, suggesting an imbalance between the inflammatory and regulatory response in the mucosa as occurs in human MCL which point this model as an interesting approach to understand some features of the disease immunopathology. Further studies are being performed to understand the Th1-mediated tissue destruction. This study was conducted in accordance with the local legislation and institutional requirements being approved by the Ethics Committee on the Animal Use in Experimentation - under the protocol CEUA No. 133/23 of the Health Sciences Center (CCS) from the University of Rio de Janeiro.
Introduction
Leishmaniasis is a group of neglected tropical diseases caused by infection by protozoan parasites belonging to the genus Leishmania. The disease affects the most deprived populations in the world, being endemic in more than 98 countries, with cases distributed across Asia, Africa, the Middle East, and South and Central America (1). Estimates suggest that around 1 million new cases of leishmaniasis occur every year globally, with a population of 1 billion people living in areas at risk of contracting the disease (2). Of the different clinical manifestations of leishmaniasis, mucocutaneous leishmaniasis (MCL) stands out because it can generate a severe deformity, generating destruction of the tissues of the nose, upper lips, palate, pharynx, larynx, perforation of the septum and loss of bone tissue (3, 4). Mucocutaneous Leishmaniasis (MCL) is a consequence of Leishmania’s tropism for mucosal tissues, involving the respiratory mucosa of the upper tract and the oral cavity. This clinical manifestation is typically a result of infection with New World species such as L. braziliensis, L. panamensis, L. guyanensis and L. amazonensis (5). It is estimated that 3 to 5% of cases of cutaneous leishmaniasis caused by these species evolve into the mucosal form (6). The cure rate for MCL treatment is lower compared to LCL (7, 8), which highlights the importance of a better understanding of development and immune response in this clinical manifestation (9, 10).
Therefore, despite being a more severe manifestation and even with a wide range of Leishmania species that can cause MCL and capable of infecting small rodents such as hamsters and mice, there is a lack of experimental models for MCL (5), and the immune response associated with this clinical manifestation is still poorly characterized. Using Leishmania amazonensis infection on BALB/c mice that is very susceptible to infection, we hypothesized that the parasite could establish the infection directly to the nose inducing manifestation similar to mucosal leishmaniasis. Previous attempts were made in the literature using subcutaneous infection in the paw leading to mucosal metastasis, however, they are very long (approximately 8 months after infection) diverse (some animals do not develop the disease) and several mice don’t survive during the prolonged infection and aging process. It is urgent a model that facilitates the study of mucosal leishmaniasis (11). In the context of the disease, mucosal leishmaniasis is associated with a Th1, but also a Th17, neutrophil infiltrate immune response (12). Meanwhile, BALB/c dermal pathology is associated with Th2 (13) but also with a strong Th17 (14, 15), making BALB/c a model to exploit the role of those immune axis to the pathology of mucosal leishmaniasis. In this way, here in we established a L. amazonensis mucosal leishmaniasis mice model using three infections sites to understand the immunopathology of the disease.
Methods
Leishmania culture
In this work, parasites of the species L. amazonensis (MHOM/BR/75/JOSEFA) were used. The promastigotes were maintained in 25cm² culture flasks with M199 medium (pH 7.2), supplemented with 10% fetal bovine serum, 5µg/mL bovine hemin, 50U/mL penicillin, and 50µg/mL streptomycin.
Animals
Mice from BALB/c lineage, females aged 8–12 weeks, were used. The mice came from the Biotério Central de Camundongos of the Centro de Ciências da Saúde (CCS) from the Universidade Federal do Rio de Janeiro, mice were maintained under the Protocol of the Ethics Committee on the Use of Experimental Animals No. 133/23 of the CCS.
Infection and lesion development
The parasites were cultivated until the beginning of the stationary phase. The culture containing the promastigote forms were collected, washed with phosphate buffer saline (PBS); and centrifuged for 10 minutes at 4°C at 1000 x G force. After centrifugation, the supernatant was discarded, and the cells were washed with PBS. The procedure was repeated twice. Then the cells were counted and adjusted to a concentration of 2x108 parasites/mL. The animals were sedated with ketamine and xylazine and infected with 2x106 parasites using Hamilton® syringes coupled with 34G needles. Different infection methods were implemented: intradermal in the ear performed at an angle of 20°; Cutaneous Nose, with the inoculum carried out at the apex of the mouse’s nose; Septum, with the inoculum occurring in the mouse septum and “Nasobasal”, with the inoculum being carried out at an angle of 60° towards the lower “floor” part of the nose. The lesion was monitored with photos. Weekly measurements of the mice’s ears were taken using Mitutoyo™ thickness gauges.
Micro computerized tomography, 3D reconstruction and lesion analysis
Animals were sedated and placed in the LabPET8 Flex Triumph Gamma Medica™ tomography system at CENABIO, UIPA. The equipment settings included 60kV, 480µA, 1024 projections, 8-minute acquisition, and 30x magnification with a 39.46mm field of view, focusing on the snout and mucous membrane. After acquisition, data was reconstructed and analyzed using 3D Slicer™ software, with the Scissors tool applied to generate the snout area.
Histology analysis
Tissues from animal snouts were fixed in 4% formaldehyde in PBS for 15 days. Decalcification was performed for 28 days using 10% EDTA. After fixation and decalcification, tissues were dehydrated through a series of ethanol and xylene solutions, then embedded in paraffin. Sections (5µm thick) were cut, rehydrated, and stained with Hematoxylin & Eosin (H&E). Tissue processing was done by the Histotechnology Platform at IOC - FIOCRUZ. Histology was performed with n = 5 per group.
Parasite load quantification
At the end of each experiment, parasite load was assessed using the limiting dilution method (LDA). Animals were euthanized with an overdose of Ketamine and Xylazine, and tissues (snout, nose tip, premaxilla, ear, cervical lymph node, ear lymph node, NALT, and spleen) were collected, weighed, and macerated in 1mL of M199. Serial 1:4 dilutions were made, and the results were plotted. Three experiments (n=3-5) were conducted, with two examining the entire snout and one focusing on segmented parts, 12 weeks post-infection.
Flow cytometry
Flow cytometric analysis of cells from cervical and ear lymph nodes of infected or naïve animals was performed. Tissues were macerated, and cells counted with 0.1% trypan blue dye. 1x106 cells were plated in a 96-well plate, centrifuged, and stimulated with PMA (20ng/mL), ionomycin (1µg/mL), and monensin (2.5µg/mL) in Complete RPMI for 4 hours at 37°C. Cells were blocked with anti-FcR, surface stained, fixed using the eBioscience™ FoxP3 kit, and intracellularly labeled. The samples were analyzed on a BD LSR Fortessa X-20 cytometer. Supplementary Table S1 denotes the cytometer configuration. Supplementary Table S2 shows the antibodies (BioLegend™) used and their concentrations. Supplementary Figure S1 denotes the performed gating strategy.
Statistics
Results are represented by Standard Error of the Mean (SEM). Statistical significance was performed by unpaired two tailed Student t test with 95% confidence interval (p<0,05) for the respective data: Lesion profile by tomography, parasite load quantification and cytometry. While One Way ANOVA with 95% interval of confidence (p<0,05) was performed using Tukey post-test, for the ELISA OD Sum data. Statistically significant differences were defined as * for p<0,05; ** for p<0,005; *** for p<0,0005.
Results
Direct inoculation of Leishmania amazonensis in nasal mucosa led to injury but the lesion profile depends on the type of inoculum performed
To assess Leishmania infection in the nasal mucosa, 2x106 promastigotes of L. amazonensis were directly inoculated into different nasal sites of mice. After 6 weeks, the Nose Cutaneous (NC) and Nasobasal (NB) groups developed visible lesions with edema and redness (Figure 1A). These lesions progressed and became necrotic by 12 weeks in some cases. The muzzle volume was significantly larger in the NC and NB groups (387mm³ and 616mm³) compared to the Sham group (Figure 1B), resembling the progressive ear infection, which reached 1.8mm in thickness (Figures 1C, D). In contrast, the Septum group showed no visible lesions throughout the study (Figure 1A). These results indicate that nasal inoculation can cause tissue damage, depending on the infection site.
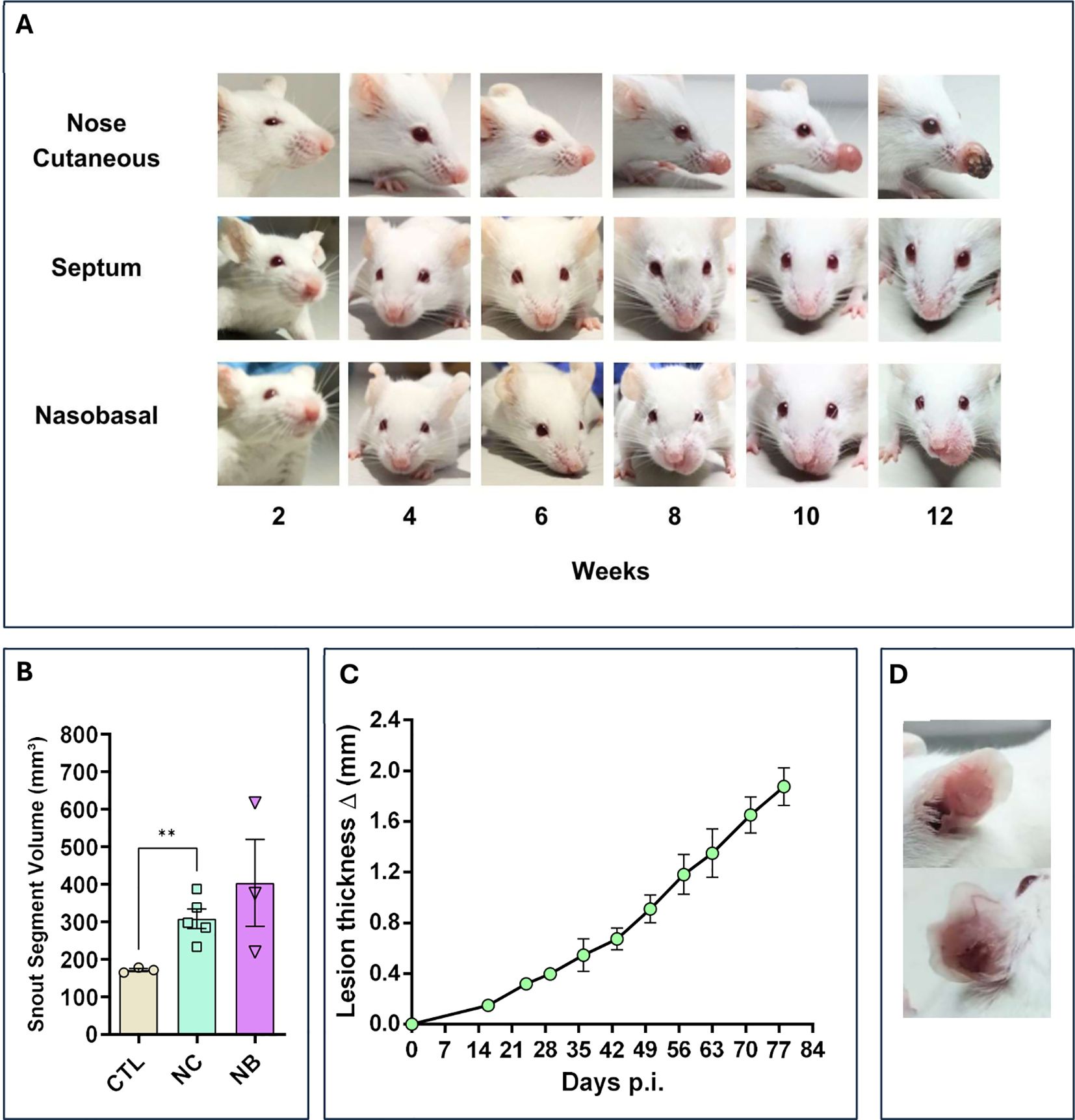
Figure 1. Nasal mucosa lesion profile. The modes of nasal mucosa inoculation with L. amazonensis. Nose cutaneous (NC) group was inoculated in the nostril nose dermis; Septum (ST) was inoculated in the septum tissue, “Nasobasal” (NB) was inoculated in the nasal floor. In all cases was used an 34G needle. (A) Lesion profile of the three groups, during a period of 12 weeks post infection (p.i), showing a continuous development of edema and erythema starting at 6 to 8 weeks for the NC and NB group. ST group does not show any sign of lesion. (B) Snout volume quantification 12 weeks p.i (Standard Error of the Mean – (SEM)), using micro–Computerized Tomography (micro-CT). NC and NB groups have greater volume compared to control groups. (C) Ear dermis lesion thickness of mice infected in the ear as an infection control group (SEM). The thickness continuous increases during the experiment reaching around 2.0mm in 12 weeks p.i. (D) Representative ear of a mice from control infection group showing edema and erythema. (B) Representative data of 3 independent experiments (5 animals per group). (C) Data from one experiment 3–5 animals per group. (D) Representative data from 2 independent experiments 5 animals were used. Statistics: unpaired t-test ** p<0,005.
The nasal mucosa model showed a higher parasite load than ear dermis infection.
Parasite load in infected mice ranged from 106 to 108 parasites per gram of tissue in the Nose Cutaneous (NC) and Nasobasal (NB) groups, while the Septum (ST) group showed only 104 parasites, with Only 20% of animals testing positive (Figure 2A). Ear dermis infections had 106 parasites per gram (Figure 2B). Parasites were mostly concentrated in the edema sites, such as the nose tip and pre-maxillary regions (Figures 2C, D). Some animals in the NB group had around 105 parasites in NALT (Figure 2E). Only one animal in the NC group showed spleen infection (Figure 2F). The cervical lymph nodes of NC and NB mice had 106 parasites (Figure 2G), while the ear lymph nodes had 105 parasites (Figure 2H). The Septum group showed parasitic load in the cervical lymph node but not the nasal cavity (Figure 2G). These results indicate that mucocutaneous L. amazonensis infection leads to a higher parasite content in nasal mucosa infection compared to ear dermis, with the cervical lymph node being the primary draining node.
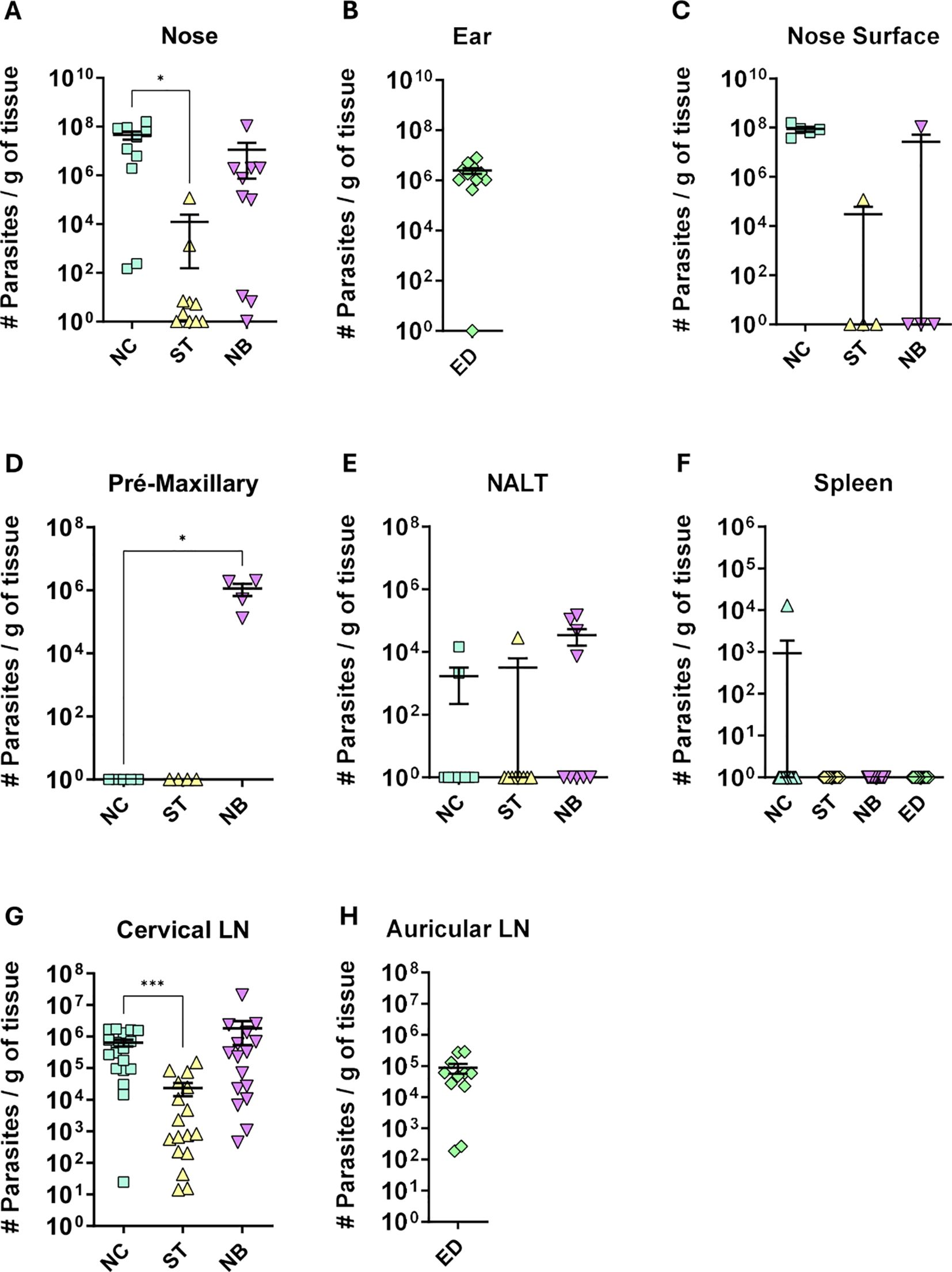
Figure 2. Parasite load profile. Profile of parasite load per gram of tissue, of the different groups of infection. NC, ST, NB, ED means respectively to Nose Cutaneous, Septum, “Nasobasal” and Ear dermis infection groups. The following tissues are represented in the figure: (A) Nose = the entire snout containing the nasal tissues; (B) Ear; (C) Nose Surface = only the visible nose; (D) mice premaxillary region; (E) Nasal Associated Lymphoid Tissue (NALT); (F) Spleen; (G) Cervical lymph node; (H) auricular lymph node. Parasite load was detected by the limiting dilution method (LDA) carried out at the end of the experiment (approximately 12 weeks of infection). (A) Data from 3 independent experiments; (B–D) 1 independent experiment; and (E), Data from 4 independent experiments; (G, H), 2 independent experiments. N = 5 animals per group. Data represented with the Standard Error of The Mean (SEM). Statistics: unpaired t-test *p < 0,05, ***p < 0,0005.
Nasal infection induced nasal swelling, but no septum perforation or new nasal cavity formation took place
MicroCT analysis was performed to assess nasal cavity impairment, focusing on the NC and NB groups, as the ST group showed no visible lesions or significant parasite load. NC-infected mice typically had lesions at the nose apex, while NB-infected mice had lesions affecting the mucosa and premaxilla, extending to the upper lips (Figures 3A, B). No nasal cavity enlargement, perforations, or deformation were observed compared to controls (Supplementary Figure S2), but tissue swelling was evident in both NC and NB groups (Figures 3A-D). Axial sections showed noticeable edema on the outer nasal cavity (Figures 3C, D). These results indicate that lesion profiles depend on the inoculation site, mainly affecting the nasal tissues initial portions.
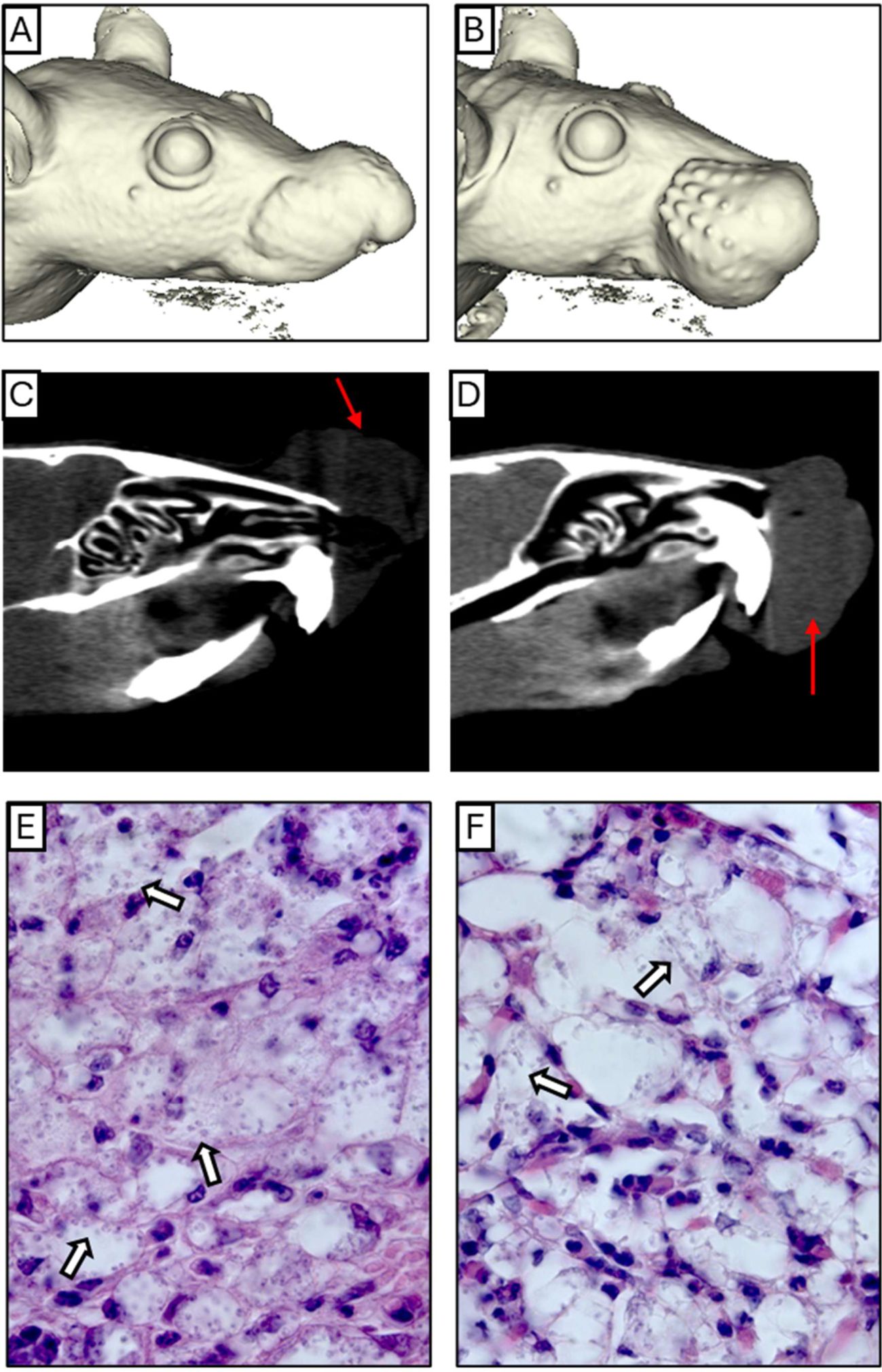
Figure 3. Anatomical and histological profile. (A, B) A complete three-dimensional reconstruction of NC and NB mice model respectively. (C, D) Coronal Section planes showing edema and partial nose obstruction of NC and NB mice respectively. (E, F) Intense inflammatory infiltrate containing infected cells, below the epithelium of NC and NB mice respectively. The red arrows indicate the region of edema, and the white arrows show examples of cells bearing amastigotes. Magnification: (E) 1000x and (F) 1000x.
Nasal infection promoted destruction and deconfiguration of squamous and transitional tissues
Histological analysis of the nasal cavity revealed significant inflammation and parasite presence. In the NC group, edema with a neutrophilic infiltrate and vacuolated macrophages containing amastigotes was observed (Figures 3E, Supplementary Figure S3), along with respiratory epithelium loss (Supplementary Figure S4B), and parasitized macrophages near cartilage (Supplementary Figure S4E). In the NB group, similar edema and neutrophil infiltration were seen, with many parasitized macrophages (Figure 3F). Respiratory epithelium showed slight morphological changes, including cellular stress and detachment (Supplementary Figures S4C, F). These findings indicate that Leishmania infection impacts the inner nasal mucosa in this model.
Nasal mucosa infection produced higher systemic antibodies titers compared to ear dermis infection
To assess immune responses to different nasal mucosa infections by Leishmania, we measured antibody levels. The NC group showed the highest IgM, IgG, and IgG1 levels, followed by NB and ED (Figures 4A-F). IgG2a and IgG2b were highest in NB, then NC (Figures 4G-J). Overall, NC and NB had stronger antibody responses than ED, while ST had the lowest. These findings suggest mucocutaneous infection triggers a stronger humoral response than ear dermis infection, with antibody profiles varying by inoculation site.
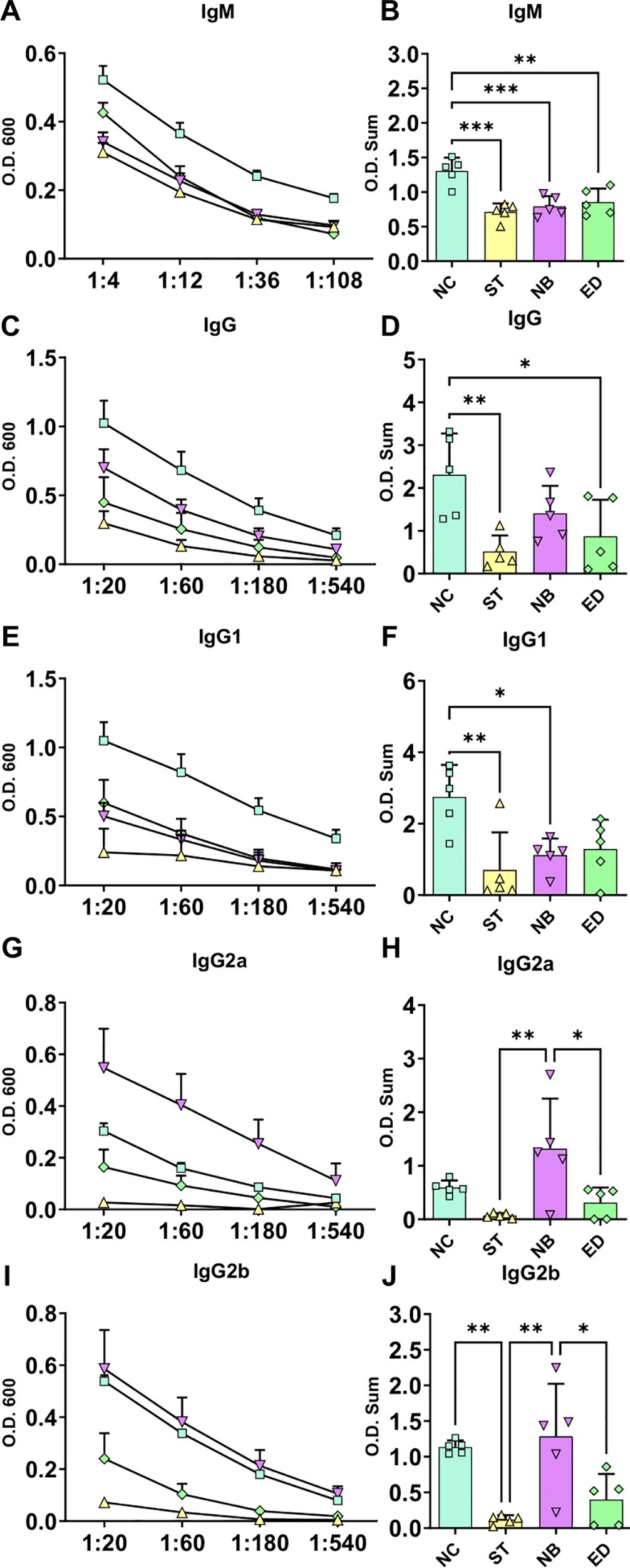
Figure 4. Systemic antibodies infection profile. Systemic antibodies responses in the different infected groups. (A, C, E, G, I) Ig titration using four dilutions, represented by O.D. 600 absorbance measurements (Standard Error of The Mean (SEM)). (B, D, F, H, J) Ig O.D. Sum from each animal and groups. NC groups produce more IgM, total IgG and IgG1, while NB mode of infection have more IgG2a and IgG2b. NC, ST, NB, ED groups represented by square; triangle; inverse triangle and diamond respectively. Statistics: One-way ANOVA; *p<0,05; **p<0,005; ***p<0,0005.
Cytotoxic and effector CD4+ and CD8+ T cells from draining lymph nodes were increased in NC, however, CD4+ T cells producing IL-10 were impaired in nasal mucosa infection
Flow cytometry of draining lymph nodes showed reduced CD4+ T cell frequency in the ED group but not in nasal infections. The NC and NB groups had increased total CD4+ T cells (Figures 5A, E) and higher numbers of cells expressing cytotoxic markers (CD107a, granzyme B, perforin) across NC, NB, and ED groups (Figures 5B-D), despite unchanged frequencies (Supplementary Figures S6-S8).
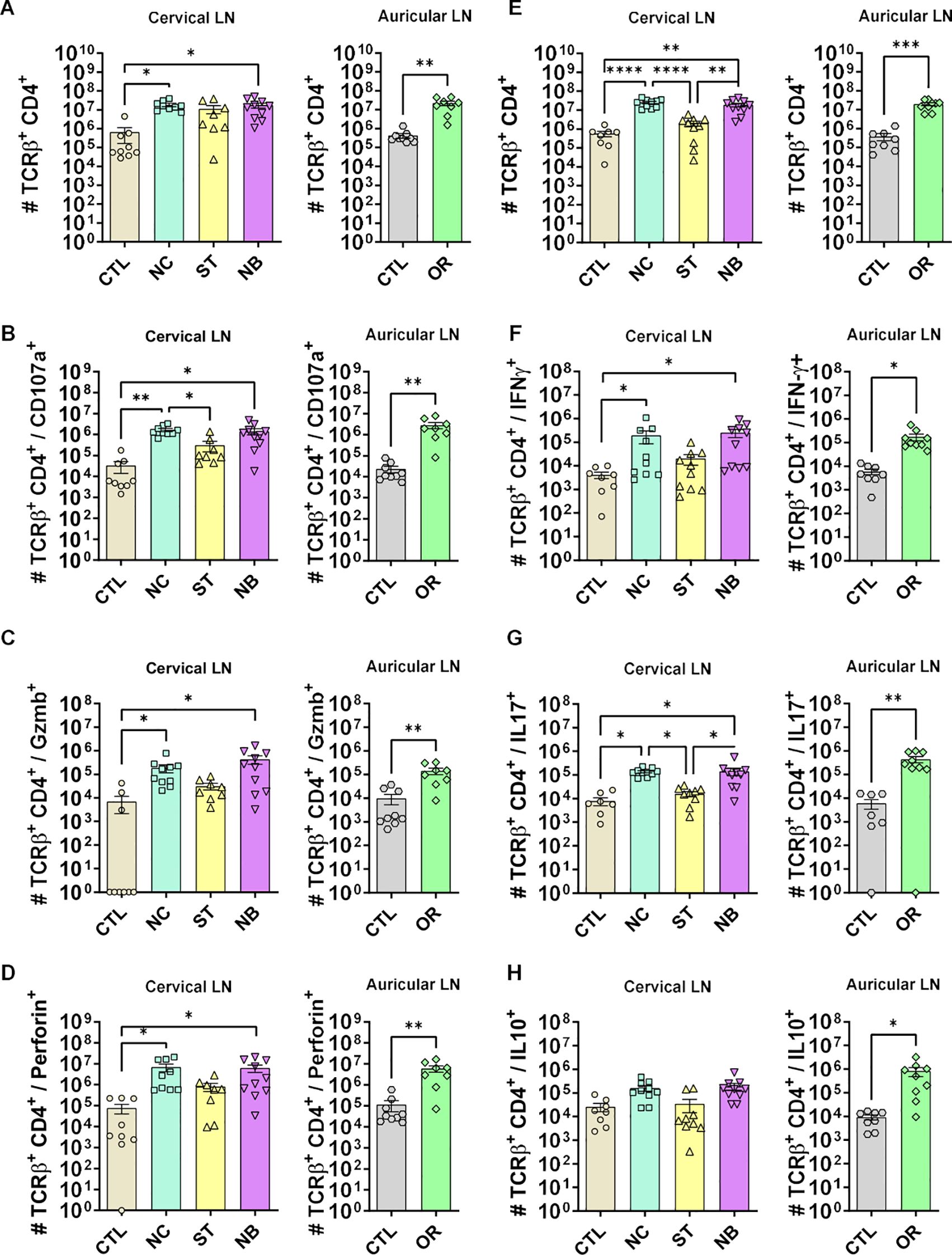
Figure 5. CD4+ T cells cytotoxic and effector profile. (A-D) The cytotoxic profile of T helper lymphocytes was measured by the expression of CD107a, Granzyme B and Perforin. (A) Cervical and auricular lymph nodes total Lymphocytes counts are shown in logarithmic scale; (B) total cells expressing CD107a, (C) Granzyme B; (D) Perforin. (E-H) The effector profile of T helper lymphocytes was measured by the expression of IL-10, IL-17 and IFN-γ. (E) Cervical and auricular lymph nodes total lymphocytes counts are shown in logarithmic scale; (F) total cells expressing IFN-γ, (G) IL-17; (H) IL-10. Data accumulative of two independent experiments (3–6 animals per group). Statistics: plot with Standard Error of The Mean (SEM), t-test was used for all groups and samples *p<0,05, **<0,005, ***p<0,0005 ****p<0,00005.
Although frequencies remained unchanged (Supplementary Figures S9, Supplementary Figures S10), total IFN-γ+ (Figure 5F) and IL-17+ (Figure 5G) cells increased in NC, NB, and ED groups. CD4+ IL-10+ T cells increased only in the ED group (Figure 5H), with lower IL-10-producing CD4+ T cell frequencies in nasal infections compared to ED (Figures 6A, B), highlighting distinct expansion dynamics between nasal and dermal infections.
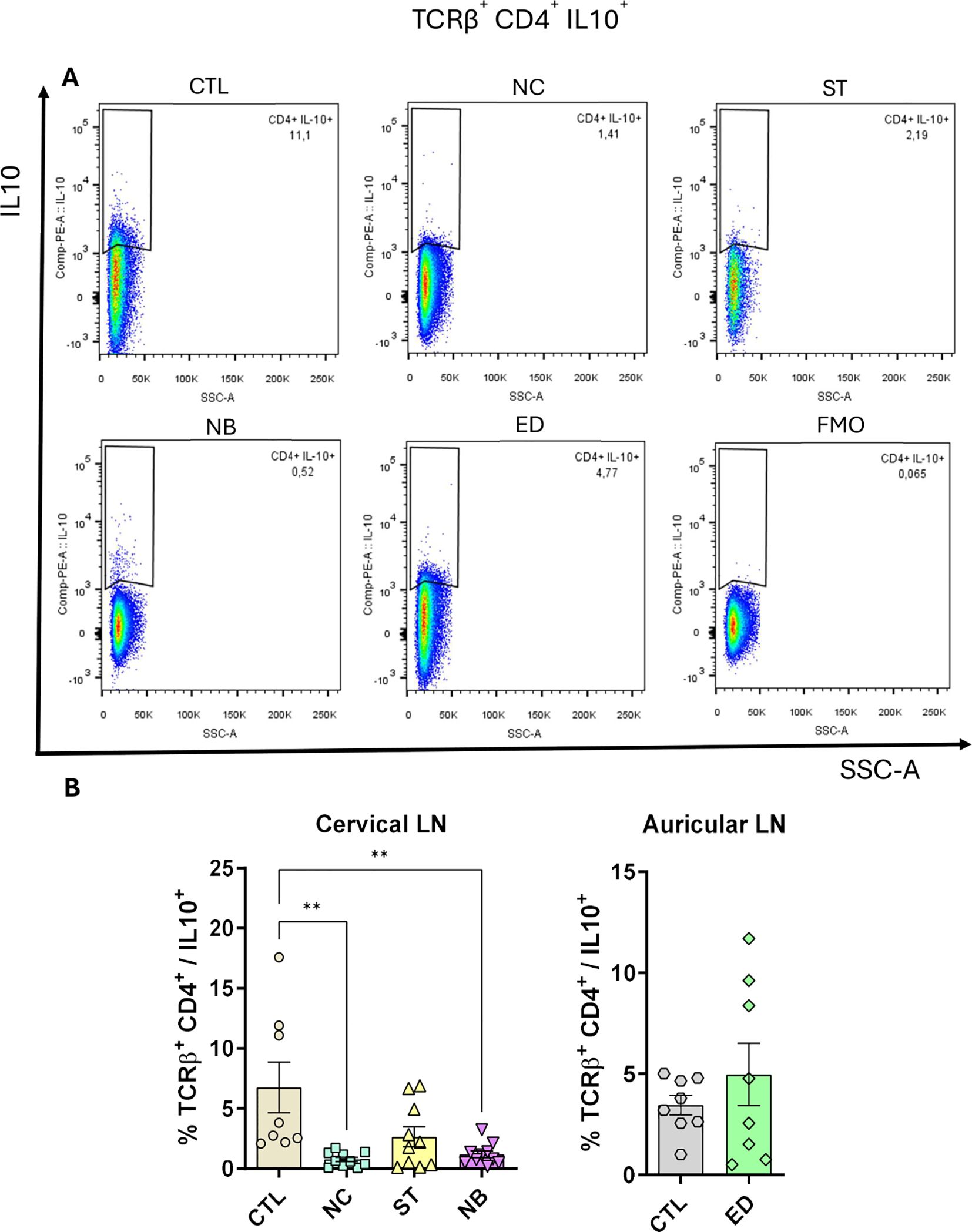
Figure 6. CD4+ T cells IL-10+ is impaired. The IL-10 producing CD4+ T cells was measured and the frequency is shown. (A) Pseudocolor dot plot showing the representative animals of each group. (B) Frequency plot from cervical and auricular lymph nodes from infected animals vs controls. Data accumulative of two independent experiments (3–6 animals per group). Statistics: plot with Standard Error of The Mean (SEM), t-test was used for all groups and samples **<0,005.
We analyzed whether changes in CD4+ IL-10+ T cells were linked to CD4+ CD25+ FoxP3+ Tregs. Treg numbers increased in NC, NB, and ED groups without frequency changes (Supplementary Figure S11). Similar trends were observed for CD25- FoxP3+, CD25+ FoxP3-, and CD25- FoxP3- cells (Supplementary Figures S12-Supplementary Figure S14), indicating the IL-10 reduction is unrelated to regulatory T cell frequency.
We also analyzed CD8+ T lymphocytes during mucosal infections. NC and NB groups showed an expansion in total CD8+ T cells (Figures 7A, E) with frequencies similar to CD4+ T cells (Supplementary Figure S15). Cytotoxic CD8+ T cells markers increased in NC, NB, and ED groups (Figures 7B-D), with NC showing higher CD8+ Perforin+ frequency (Supplementary Figure S18), nut no frequency changes to CD107a and Granzyme B respectively (Supplementary Figure S16, Supplementary Figure S17). There were no IFN-γ+ CD8+ significant changes in frequency (Figure 7F) or number (Supplementary Figure S19). IL-17+ CD8+ T cells increased in NB and ED groups (Figure 7G) without frequency changes (Supplementary Figures S20). CD8+ IL-10+ T cells showed no numerical change (Figure 7H), but frequency decreased in mucosa-infected and ED groups (Supplementary Figure S21).
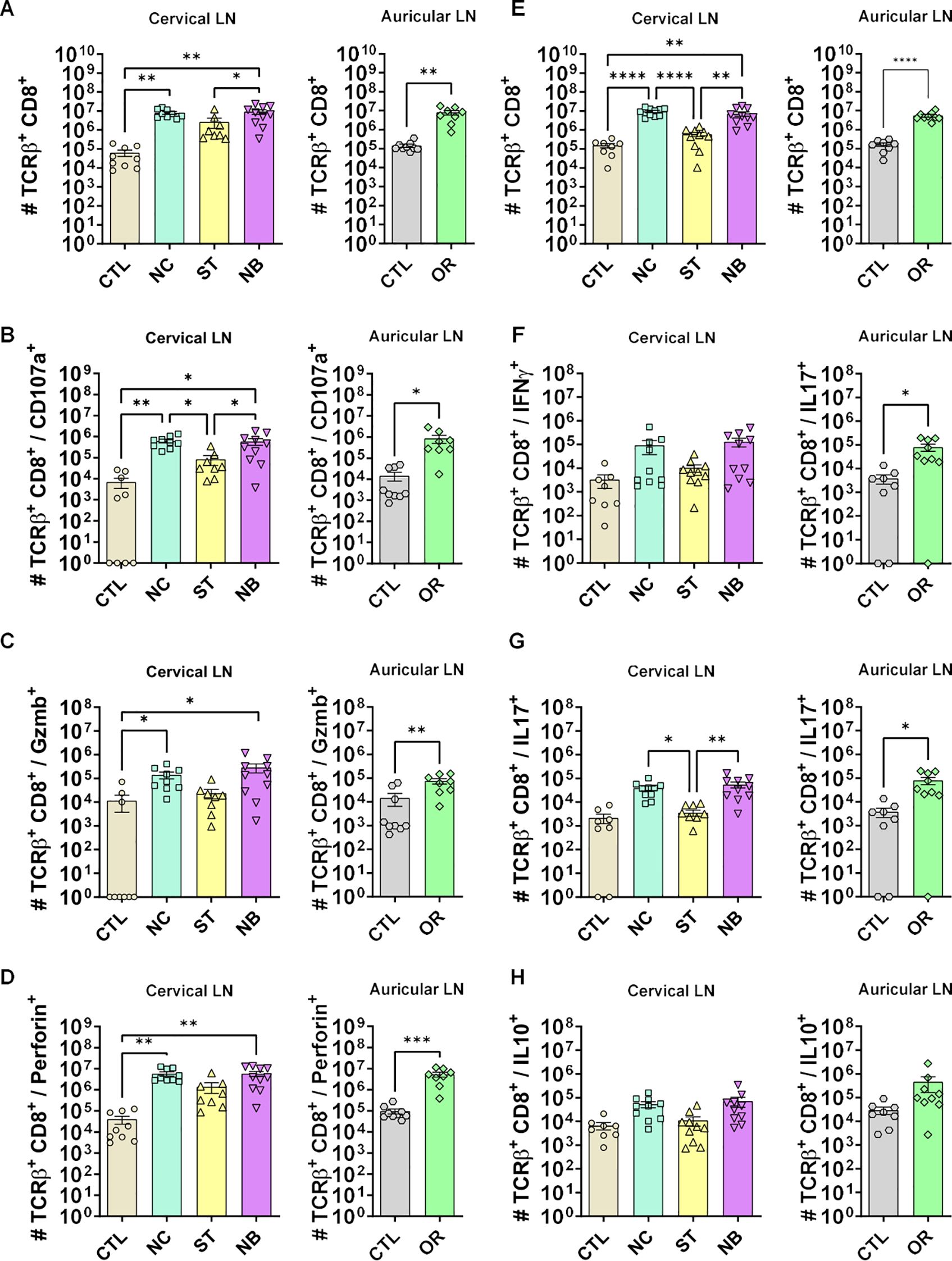
Figure 7. Cytotoxic and effector profile of CD8+ T cells. (A-D) The cytotoxic profile of T helper lymphocytes was measured by the expression of CD107a, Granzyme B and Perforin. (A) Cervical and auricular lymph nodes total Lymphocytes counts are shown in logarithmic scale; (B) total cells expressing CD107a, (C) Granzyme B; (D) Perforin. (E-H) The effector profile of T helper lymphocytes was measured by the expression of IL-10, IL-17 and IFN-γ. (E) Cervical and auricular lymph nodes total lymphocytes counts are shown in logarithmic scale; (F) total cells expressing IFN-γ, (G) IL-17; (H) IL-10. Data accumulative of two independent experiments (3–6 animals per group). Statistics: plot with Standard Error of The Mean (SEM), t-test was used for all groups and samples * p<0,05, **<0,005, ***<0,0005.
Discussion
Mucocutaneous Leishmaniasis (MCL) causes severe tissue damage in nasal, oral, and pharyngeal mucosa, leading to edema and erythema in affected areas (3, 5). In our mouse model, the NC and NB groups developed visible edema and erythema by week 6, progressing over time. Lesions in the NC group formed at the nose tip and extended dorsally, resembling the experimental mucosal leishmaniasis in dogs and hamsters (16, 17). In the NB group, lesions spread to the upper lips and premaxilla, mimicking clinical features of human MCL (18, 19).
Some animals developed necrotic lesions after 12 weeks of infection. While L. amazonensis typically causes non-necrotizing lesions in BALB/c mice resembling diffuse cutaneous leishmaniasis in humans (20). Similar variation in infection sites was observed with Leishmania major (21), possibly linked to site-specific microbiota. Nasal microbiota, including Staphylococcus and Streptococcus species (22, 23), may exacerbate tissue damage by recruiting neutrophils and CD8 IL-17+ cells during infection (24, 25).
Mucosal leishmaniasis in humans often affects septa, cartilage, and nasal cavities, with occasional bone destruction visible via computed tomography (3, 26–28). In this study, microCT revealed edema in the nasal and premaxilla regions of NC and NB groups. Histopathology showed dense cellular infiltrates and infected cells near cartilage and the nasal mucosa, with respiratory epithelium changes, consistent with previous findings (11).
Despite MCL caused by Leishmania braziliensis typically shows fewer parasites at the site than cutaneous leishmaniasis (29), our model NC and NB groups had higher parasite loads, in the draining lymph nodes and showing also infected cells in nasal mucosa. This may reflect L. amazonensis characteristics, which produce higher parasite loads than L. braziliensis in mice (30), and as evidenced by the case of MCL caused by L. amazonensis, where parasite amastigotes were described as easily visualized in H&E sections (31). Unlike previous models focusing on nasal infection (16, 17, 32, 33), this study uniquely examined infections within the nasal mucosa cavity, revealing that the inoculum site significantly influences lesion severity and parasite load across NC, ST, and NB groups. Necrosis was observed in some animals, a feature absents in other models (16, 17, 33). Mucosal infections showed clinical aspects similar to, or more severe than cutaneous ear lesions, aligning with findings in Leishmania panamensis-infected hamsters (17). While other models required approximately 8 months to develop lesions (11), this approach shortened progression to 6–8 weeks. Future studies could compare axenic amastigote infections, given their higher virulence and differing proliferation kinetics from promastigotes (34, 35).
MCL severity correlates with higher anti-Leishmania IgG levels (36). In our model, mucosal infections (NC and NB) produced more IgM and IgGs than dermal infections, the high antibody levels produced by the mucosal infection compared to dermis can be interpreted as an increased inflammatory response that may contribute to the severity of pathology. Interesting, the mucosal infection site is contributing to the type of immune response with NC favoring IgG1 (Th2 response) and NB favoring IgG2a (Th1 response), according to the antibody dichotomy (37).
To compare nasal mucosa infections with human MCL, we performed flow cytometry on lymph node cells. Mucosal infection (NC and NB) increased CD4+ and CD8+ T cells expressing cytotoxic markers (CD107a, Granzyme B, Perforin) and IL-17, with less IFN-γ, but no increase in IL-10-expressing cells, unlike the ED group. In MCL, CD4+ T cells produce high IFN-γ and TNF-α, with elevated Th17 responses that recruit neutrophils and cause tissue damage (12, 38, 39). MCL also shows higher granzyme A-expressing cells, linked to greater damage (34), with a lower IL-10 receptor expression, despite normal IL-10 expression (39). Our data suggests a similar imbalance between cytotoxic and regulatory responses, with reduced IL-10+ T cells and increased IL-17+ T cells potentially contributing to severity, as IL-10 limits Th17-mediated pathology in experimental leishmaniasis (40).
The overall IL-10 production by T cells is important to protect against excessive inflammation and damage during the peak of infection for Leishmania mexicana and L. major (40, 41). Tregs (CD4+ CD25+ FoxP3+ T cells) and Tr1 (CD4+ CD25- FoxP3- IL-10+) are the most important subsets of T cells that provide IL-10 during immune-responses (42). In the L. amazonensis mice infection, the CD4+ CD25+ T cells can contribute to parasite control decreasing the inflammatory response (43). Furthermore, CD4+ CD25+ FoxP3+ T cells are also implicated to reduce inflammation and parasite load in BALB/c mice infection by Leishmania panamensis (44). Moreover, on visceral leishmaniasis mice model, the regulatory T cells had no effect on Leishmania donovani parasite proliferation but were able to reduce tissue damage (45). However, the IL-10 production by T cells is also frequently linked to parasite persistence and in some circumstances an increased pathology. The IL-10 production by CD4+ CD25- FoxP3- IL-10+ T cells in a Th1 environment is important for the lesional persistence of L. major (Seidman strain) and L. mexicana (41, 46), while for the L. major (Friedlin) that causes a self-healing pathology, the IL-10 production by CD4+ CD25+ FoxP3+ Tregs prevents the sterile cure (47). In our model, we can hypothesize that the inflammatory response of mucosal sites is exacerbated by the absence of T cell IL-10 production that is not contributing to the parasite control, instead it is favoring the recruitment of host cells for parasite replication and increasing tissue damage.
Taken together, L. amazonensis infection in BALB/c mice induces an immune response that reflects some aspects of the human MCL, with increased cytotoxic and effector cells but a compromised regulatory response. However, our studies didn’t find some MCL features like cavity formation, septal perforations, and bone destruction, possibly due to parasite species or host lineage differences, or the shortened experimental period compared to human pathology (28). To address this, ongoing research is being performed with C57BL/6 strain for a stronger Th1 response (15, 48). We also are planning to test the L. braziliensis hamster model for its higher susceptibility for this species (49). As NC and NB sites differed in some characteristics, we believe that the combination of these inoculum sites may better represent the full mucosal leishmaniasis profile in mice. More studies are needed to confirm the lymphocyte profile in the mucosal site.
In conclusion, when infected directly at nasal mucosa, L. amazonensis parasites are more proliferative and lead to an increased inflammatory response compared to ear dermis infection, which can be associated with the IL-10 impairment related to the immunopathology of MCL.
Data availability statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Ethics statement
This study was conducted in accordance with the local legislation and institutional requirements being approved by the Ethics Committee on the Animal Use in Experimentation - under the protocol CEUA No. 133/23 of the Health Sciences Center (CCS) from the University of Rio de Janeiro.
Author contributions
AD-R: Conceptualization, Methodology, Writing – original draft, Writing – review & editing, Data curation, Formal analysis, Investigation, Project administration. JD: Conceptualization, Investigation, Writing – review & editing, Supervision. IB: Investigation, Writing – review & editing. DD-A: Investigation, Writing – review & editing. NM: Investigation, Writing – review & editing. HP: Investigation, Writing – review & editing. JR: Investigation, Writing – review & editing. ES-J: Writing – review & editing, Formal analysis. AS-G: Writing – review & editing, Methodology. MO: Methodology, Writing – review & editing. LC: Resources, Writing – review & editing, Validation. DDO: Resources, Validation, Writing – review & editing. AD-C: Resources, Validation, Writing – review & editing. AF-M: Resources, Writing – review & editing, Formal analysis, Investigation, Methodology. HDM: Investigation, Writing – review & editing, Conceptualization, Funding acquisition, Methodology, Resources, Supervision, Writing – original draft.
Funding
The author(s) declare that financial support was received for the research and/ or publication of this article. This work was supported by Fundação de Amparo à Pesquisa Estado do Rio de Janeiro – FAPERJ Process E-26/200.770/2022 and CNE: 200.993/2022. Conselho Nacional de Desenvolvimento Cientifico e Tecnológico (CNPQ - Process PQ1D307632/2022-9).
Acknowledgments
We would like to acknowledge the Centro Nacional de Bioimagem (CENABIO) for the equipment and technical support in tomography analysis. We would also like to acknowledge the Platform of Histotechnology (FIOCRUZ-RJ) for the processing of histology samples.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The author(s) declared that they were an editorial board member of Frontiers, at the time of submission. This had no impact on the peer review process and the final decision.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fimmu.2025.1621781/full#supplementary-material
References
1. Mann S, Frasca K, Scherrer S, Henao-Martínez AF, Newman S, Ramanan P, et al. A review of leishmaniasis: current knowledge and future directions. Curr Trop Med Rep (Internet). (2021) 8:1–12. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC7966913/.
2. World Health Organization. Leishmaniasis. Whoint (2022). Available at: https://www.who.int/news-room/fact-sheets/detail/leishmaniasis (Accessed June 30, 2022).
3. Lessa MM, HA L, Castro TWN, Oliveira A, Scherifer A, Machado P, et al. Mucosal leishmaniasis: epidemiological and clinical aspects. Braz J Otorhinolaryngology. (2007) 73:843–7. doi: 10.1016/s1808-8694(15)31181-2
4. Goto H and Lindoso JAL. Cutaneous and mucocutaneous leishmaniasis. Infect Dis Clinics North America. (2012) 26:293–307. doi: 10.1016/j.idc.2012.03.001
5. Strazzulla A, Cocuzza S, Pinzone MR, Postorino MC, Cosentino S, Serra A, et al. Mucosal leishmaniasis: an underestimated presentation of a neglected disease. BioMed Res Int (Internet). (2013) 2013. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3703408/.
6. Handler MZ, Patel PA, Kapila R, Al-Qubati Y, and Schwartz RA. Cutaneous and mucocutaneous leishmaniasis: Clinical perspectives. J Am Acad Dermatol (Internet). (2015) 73:897–908. doi: 10.1016/j.jaad.2014.08.051.
7. Cincurá C, de Lima CMF, Machado PRL, Oliveira-Filho J, Glesby MJ, Lessa MM, et al. Mucosal leishmaniasis: A Retrospective Study of 327 Cases from an Endemic Area of Leishmania (Viannia) Braziliensis. Am J Trop Med Hygiene (Internet). (2017) 97:761–6. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC5590558/.
8. Martins SS, Barroso DH, Rodrigues BC, da Motta JdOC, Freire GSM, Pereira LIdA, et al. A pilot randomized clinical trial: oral miltefosine and pentavalent antimonials associated with pentoxifylline for the treatment of american tegumentary leishmaniasis. Front Cell Infection Microbiol (Internet). (2021) 11:700323. doi: 10.3389/fcimb.2021.700323
9. Sampaio RNR, Silva JSFe, Paula CDRd, Porto C, Motta JdOCd, Pereira LIdA, et al. A randomized, open-label clinical trial comparing the long-term effects of miltefosine and meglumine antimoniate for mucosal leishmaniasis. Rev da Sociedade Bras Medicina Trop. (2019) 52. doi: 10.1590/0037-8682-0292-2018
10. Calvopina M, Jijon S, Serrano E, and Kato H. Case report: successful treatment with miltefosine of severe new world mucosal leishmaniasis caused by leishmania guyanensis. Am J Trop Med Hygiene. (2020) 103:752–5. doi: 10.4269/ajtmh.19-0867
11. Cupolilo SM, Souza F, Abreu-Silva AL, Calabrese KS, and Goncalves C. Biological behavior of Leishmania (L.) amazonensis isolated from a human diffuse cutaneous leishmaniasis in inbred strains of mice. PubMed. Natl Institutes Health;. (2003) 18:1059–65. doi: 10.14670/hh-18.1059
12. Boaventura V, Santos CS, Ribeiro C, de J, Santos, Clarêncio J, et al. Human mucosal leishmaniasis: Neutrophils infiltrate areas of tissue damage that express high levels of Th17-related cytokines. Eur J Immunol Wiley-Blackwell;. (2010) 40:2830–6. doi: 10.1002/eji.200940115
13. Sacks D and Noben-Trauth N. The immunology of susceptibility and resistance to Leishmania major in mice. Nat Rev Immunol. (2002) 2:845–58. doi: 10.1038/nri933
14. Lopez Kostka S, Dinges S, Griewank K, Iwakura Y, Udey MC, and von Stebut E. IL-17 promotes progression of cutaneous leishmaniasis in susceptible mice. J Immunol. (2009) 182:3039–46. doi: 10.4049/jimmunol.0713598
15. Dos-Santos JS, Firmino-Cruz L, Ramos TD, Fonseca-Martins AMd, Oliveira-Maciel D, De-Medeiros JVR, et al. Characterization of sv129 mice as a susceptible model to leishmania amazonensis. Front Med. (2019) 6. doi: 10.3389/fmed.2019.00100
16. Pirmez C, Marzochi MCA, and Coutinho SG. Experimental canine mucocutaneous leishmaniasis (Leishmania Braziliensis Braziliensis). Memórias do Instituto Oswaldo Cruz. (1988) 83:145–51. doi: 10.1590/s0074-02761988000200001
17. Osorio Y, Melby PC, Pirmez C, Chandrasekar B, Guarín N, and Travi BL. The site of cutaneous infection influences the immunological response and clinical outcome of hamsters infected with Leishmania panamensis. Parasite Immunol Wiley-Blackwell;. (2003) 25:139–48. doi: 10.1046/j.1365-3024.2003.00615.x
18. Marsden PD. Mucosal leishmaniasis due to Leishmania (Viannia) Braziliensis L(V)b in Três Braços, Bahia-Brazil. Rev da Sociedade Bras Medicina Tropical. (1994) 27:93–101. doi: 10.1590/s0037-86821994000200007
19. Lambertucci JR, Coulaud R, and Cristina L. Mucosal leishmaniasis. Revista da sociedade brasileira de medicina tropical. Braz Soc Trop Medicine;. (2003) 36:307–8. doi: 10.1590/S0037-86822003000200017
20. Pérez H, Arredondo B, and González M. Comparative study of American cutaneous leishmaniasis and diffuse cutaneous leishmaniasis in two strains of inbred mice. Infection Immunity. (1978) 22:301–7. doi: 10.1128/iai.22.2.301-307.1978
21. Baldwin TM, Elso C, Curtis J, Buckingham L, and Handman E. The site of leishmania major infection determines disease severity and immune responses. Infection and immunity. Am Soc Microbiology;. (2003) 71:6830–4.
22. Bomar L, Brugger SD, and Lemon KP. Bacterial microbiota of the nasal passages across the span of human life. Curr Opin Microbiol. (2018) 41:8–14. doi: 10.1016/j.mib.2017.10.023
23. Thangaleela S, Sivamaruthi BS, Kesika P, Bharathi M, and Chaiyasut C. Nasal microbiota, olfactory health, neurological disorders and aging—A review. Microorganisms. (2022) 10:1405.
24. Borbón TY, Scorza BM, Clay GM, Lima Nobre de Queiroz F, Sariol AJ, Bowen JL, et al. Coinfection with Leishmania major and Staphylococcus aureus enhances the pathologic responses to both microbes through a pathway involving IL-17A. PloS Negl Trop Diseases. (2019) 13:e0007247. doi: 10.1371/journal.pntd.0007247
25. Singh TP, Carvalho AM, Sacramento LA, Grice EA, Scott P, and Stäger S. Microbiota instruct IL-17A-producing innate lymphoid cells to promote skin inflammation in cutaneous leishmaniasis. PloS Pathogens. (2021) 17:e1009693. doi: 10.1371/journal.ppat.1009693
26. Rojas-Jaimes J, Lescano AG, Frischtak HL, and Arenas J. Case report: mucosal leishmaniasis presenting with nasal septum perforation after almost thirty years. Am J Trop Med Hygiene. (2018) 99:327–30.
27. Bhowate RR, Bhargava PA, and Badki SD. Mrunal meshram. Mucosal leishmaniasis involving the nostril and maxillary sinus: A case report. Cureus. (2022) 14:e30289. doi: 10.7759/cureus.30289
28. Camargo RA, de Nicodemo AC, Sumi DV, Mello Santiago Gebrim EM, Tuon FF, de Camargo LM, et al. Facial structure alterations and abnormalities of the paranasal sinuses on multidetector computed tomography scans of patients with treated mucosal leishmaniasis. PloS Negl Trop Diseases. Public Library Science;. (2014) 8:e3001–1. doi: 10.1371/journal.pntd.0003001
29. Jara M, Adaui V, Valencia BM, Martinez D, Alba M, Castrillon C, et al. Real-time PCR assay for detection and quantification of leishmania (Viannia) organisms in skin and mucosal lesions: exploratory study of parasite load and clinical parameters. J Clin Microbiol. (2013) 51:1826–33. doi: 10.1128/JCM.00208-13
30. Carvalho AK, Silveira FT, Passero LFD, Gomes CMC, Corbett CEP, and Laurente MD. Leishmania (V.) Braziliensis and L. (L.) amazonensis promote differential expression of dendritic cells and cellular immune response in murine model. Parasite Immunol. (2012) 34:395–403. doi: 10.1111/j.1365-3024.2012.01370.x
31. Sampaio RN, Marsden PD, Cuentas EAL, Cuba CAC, and Grimaldi J. Leishmania mexicana amazonensis isolated from a patient with fatal mucosal leishmaniasis. Revista da Sociedade Brasileira de Medicina Tropical. FapUNIFESP (SciELO);. (1985) 18:273–4.
32. Teva A, Porrozzi R, Cupolillo E, Pirmez C, Mp O-N, and Grimaldi G. Leishmania (Viannia) Braziliensis-induced chronic granulomatous cutaneous lesions affecting the nasal mucosa in the rhesus monkey (Macaca mulatta) model. Parasitology. (2003) 127:437–47. doi: 10.1017/s0031182003004037
33. Cortes S, Esteves C, Maurício I, Maia C, Cristovão JM, Miles M, et al. In vitro and in vivo behavior of sympatric Leishmania (V.) Braziliensis, L. (V.) Peruviana and their hybrids. Parasitology. (2011) 139:191–9. doi: 10.1017/s0031182011001909
34. Mendes Costa D, Cecílio P, Santarém N, Cordeiro-da-Silva A, and Tavares J. Murine infection with bioluminescent Leishmania infantum axenic amastigotes applied to drug discovery. Sci Rep. (2019) 9. doi: 10.1038/s41598-019-55474-3
35. Baek K-H, Piel L, Rosazza T, Prina E, Späth GF, and No JH. Infectivity and drug susceptibility profiling of different leishmania-host cell combinations. Pathogens. (2020) 9:393. doi: 10.3390/pathogens9050393
36. Lima CMF, Magalhães AS, Costa R, Barreto CC, Machado PRL, Carvalho EM, et al. High anti-leishmania igG antibody levels are associated with severity of mucosal leishmaniasis. Front Cell Infection Microbiol. (2021) 11.
37. Bretscher PA. On the mechanism determining the th1/th2 phenotype of an immune response, and its pertinence to strategies for the prevention, and treatment, of certain infectious diseases. Scandinavian J Immunol. (2014) 79:361–76. doi: 10.1111/sji.12175
38. Bacellar O, Lessa HA, Schriefer A, Machado P, de Jesus AR, Dutra WO, et al. Up-regulation of th1-type responses in mucosal leishmaniasis patients. Infect Immun. (2002) 70:6734–40. doi: 10.1128/IAI.70.12.6734-6740.2002
39. Faria DR, Gollob KJ, Barbosa J, Schriefer A, Machado PRL, Lessa H, et al. Decreased in situ expression of interleukin-10 receptor is correlated with the exacerbated inflammatory and cytotoxic responses observed in mucosal leishmaniasis. Infection Immunity. (2005) 73:7853–9. doi: 10.1128/IAI.73.12.7853-7859.2005
40. Gonzalez-Lombana C, Gimblet C, Bacellar O, Oliveira WW, Passos S, Carvalho LP, et al. IL-17 mediates immunopathology in the absence of IL-10 following leishmania major infection. Langhorne J editor. PloS Pathogens. (2013) 9:e1003243. doi: 10.1371/journal.ppat.1003243
41. Buxbaum LU. Interleukin-10 from T Cells, but Not Macrophages and Granulocytes, Is Required for Chronic Disease in Leishmania mexicana Infection. Infection and Immunity (Internet). Am Soc Microbiol. (2015) 83:1366–71. https://pmc.ncbi.nlm.nih.gov/articles/PMC4363430/.
42. Ng THS, Britton GJ, Hill EV, Verhagen J, Burton BR, and Wraith DC. Regulation of adaptive immunity; the role of interleukin-10. Front Immunol. (2013) 4:3389/fimmu.2013.00129. doi: 10.3389/fimmu.2013.00129
43. Ji J, Masterson J, Sun J, and Soong L. CD4+CD25+ Regulatory T Cells Restrain Pathogenic Responses during Leishmania amazonensis Infection. J Immunol. (2005) 174:7147–53. doi: 10.4049/jimmunol.174.11.7147
44. Ehrlich A, Castilho TM, Goldsmith-Pestana K, Chae W-J, L M Bothwell A, Sparwasser T, et al. The immunotherapeutic role of regulatory T cells in leishmania (Viannia) panamensis infection. J Immunol. (2014) 193:2961–70. doi: 10.4049/jimmunol.1400728
45. Bunn PT, Montes de Oca M, de Labastida Rivera F, Kumar R, Ng SS, Edwards CL, et al. Distinct Roles for CD4+ Foxp3+ Regulatory T Cells and IL-10–Mediated Immunoregulatory Mechanisms during Experimental Visceral Leishmaniasis Caused by Leishmania donovani. J Immunol. (2018) 201:3362–72. doi: 10.4049/jimmunol.1701582
46. Anderson CF, Oukka M, Kuchroo VJ, and Sacks D. CD4+CD25–Foxp3– Th1 cells are the source of IL-10–mediated immune suppression in chronic cutaneous leishmaniasis. J Exp Med. (2007) 204:285–97. doi: 10.1084/jem.20061886
47. Belkaid Y, Piccirillo CA, Mendez S, Shevach EM, and Sacks DL. CD4+CD25+ regulatory T cells control Leishmania major persistence and immunity. Nature. (2002) 420:502–7.
48. Tomiotto-Pellissier F, Miranda-Sapla MM, Silva TF, Taciane B, Gonçalves MD, Concato V, et al. Murine Susceptibility to Leishmania amazonensis Infection Is Influenced by Arginase-1 and Macrophages at the Lesion Site. Frontiers in Cellular and Infection Microbiology. Front Media;. (2021) 11. doi: 10.3389/fcimb.2021.687633
Keywords: leishmaniasis, mucosa, L. amazonensis, BALB/c, experimental infection
Citation: Da-Rocha AA, Dos-Santos JS, Bittencourt IS, De-Almeida DB, Manhães NCS, Praxedes H, Romano JVP, da Silva-Junior EB, da Silva-Gonçalves AJ, Oliveira MP, Covre LP, Gomes DCO, Da-Cruz AM, da Fonseca-Martins AM and de Matos Guedes HL (2025) Mice mucosal leishmaniasis model shown high parasite load, increased cytotoxicity and impaired IL-10+ T cell response. Front. Immunol. 16:1621781. doi: 10.3389/fimmu.2025.1621781
Received: 01 May 2025; Accepted: 10 June 2025;
Published: 04 August 2025.
Edited by:
Diego Luis Costa, University of São Paulo, BrazilReviewed by:
Lucas P. Carvalho, Gonçalo Moniz Institute (IGM), BrazilLaís Amorim Sacramento, University of Pennsylvania, United States
Copyright © 2025 Da-Rocha, Dos-Santos, Bittencourt, De-Almeida, Manhães, Praxedes, Romano, da Silva-Junior, da Silva-Gonçalves, Oliveira, Covre, Gomes, Da-Cruz, da Fonseca-Martins and de Matos Guedes. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Herbert Leonel de Matos Guedes, aGVyYmVydEBtaWNyby51ZnJqLmJy; aGVyYmVydEBpb2MuZmlvY3J1ei5icg==