 Abel Ramos-Vega1*
Abel Ramos-Vega1* Elizabeth Monreal-Escalante2,3
Elizabeth Monreal-Escalante2,3 Bernardo Bañuelos-Hernández4
Bernardo Bañuelos-Hernández4 Miriam Angulo2
Miriam Angulo2 Edgar Trujillo2
Edgar Trujillo2 Carlos Angulo2*
Carlos Angulo2*- 1Centro de Investigación en Ciencia Aplicada y Tecnología Avanzada (CICATA) Unidad Morelos del Instituto Politécnico Nacional (IPN), Xochitepec, Morelos, Mexico
- 2Immunology & Vaccinology Group and Laboratorio Nacional Consejo Nacional de Humanidades, Ciencias y Tecnología (CONAHCYT) de Generación de Vacunas Veterinarias y Servicios de Diagnóstico (LNC-GVD, Centro de Investigaciones Biológicas del Noroeste, La Paz, Mexico
- 3Secretaria de Ciencias, Humanidades, Tecnología e Innovación (SECIHTI)-Centro de Investigaciones Biológicas del Noroeste, La Paz, Mexico
- 4Facultad de Agronomía y Veterinaria, Universidad De La Salle Bajio, León Guanajuato, Mexico
Yeasts have contributed to human and animal health through functional antigen production for vaccine formulations. Some yeast-made vaccines have become a reality for humankind because they have reached commercialization (hepatitis B, HPV, and tick parasitosis). Many other vaccine prototypes are under preclinical and clinical evaluations, hoping for their usage soon. Currently, genomes, genetic modification techniques, and industrial vaccine manufacturing have been successfully developed for Saccharomyces cerevisiae, Komagataella phaffii (formerly Pichia pastoris), and Hansenula polymorpha. Moreover, several yeast species are under research as prospects for vaccine production systems, such as Kluyveromyces lactis, Yarrowia lipolytica, Schizosaccharomyces pombe, Saccharomyces boulardii, and Komagataella phaffii. This review was mainly focused on commercial human and animal vaccines, describing and discussing genetic engineering tools, downstream antigen purification processes, GMP according to regulatory issues, and identifying challenges and future directions on the use of yeast as a vaccine production platform to fight against infectious diseases.
Introduction
Infectious diseases continue to pose a major global health burden, affecting both humans and animals (1). Moreover, zoonotic pathogens are a significant concern due to their ability to cross species barriers and cause widespread outbreaks, as exemplified by the recent COVID-19 pandemic (2). In this sense, the use of antibiotics and vaccination remains the most effective strategy for controlling and preventing infectious [zoonotic] diseases. Nonetheless, the excessive and indiscriminate application of antibiotics leads to several problems, including antimicrobial resistance, environmental contamination, and disruption of the microbial diversity balance (3). In such scenarios, vaccines continue to be the most effective and safest strategy for controlling infectious diseases. Conventional vaccines are based on whole inactivated, attenuated pathogens or purified antigenic components from pathogens, such as subunit proteins or toxoids, all of which are rationally designed to elicit a protective immune response. These platforms have demonstrated substantial efficacy in decreasing disease incidence and mortality caused by infectious agents (4). However, conventional vaccines present several limitations, including safety risks associated with live or attenuated organisms, the need for highly trained personnel, cold-chain dependency, longer development timelines, and high production costs, among others (5). Consequently, novel vaccines are investigated to surpass or mitigate those challenges, which ideally should be cost-effective, suitable for alternative delivery routes, and tailored to resource-limited settings.
The arrival of recombinant DNA technology marked a significant turning point in vaccine development, enabling the precise design and production of specific antigens, for instance, without the need to cultivate or inactivate whole pathogens (6). This advancement has facilitated the development of recombinant protein-based vaccines using expression platforms such as bacteria, yeast, plants, mammalian cells, and insect cells, offering enhanced safety features compared to conventional approaches (7). Vaccines produced using recombinant expression platforms offer potential advantages, including reduced biosafety risks, the possibility of alternative routes of administration, lower production costs, and suitability for resource-limited settings (8). In this regard, yeasts have been used as a host system for recombinant protein production. In line with recombinant vaccine development, Valenzuela et al. (1982) reported for the first time the production of hepatitis B virus surface antigen in yeast, which was later evaluated in animal models (9, 10). To date, only yeast-based vaccines for humans against hepatitis B and human papillomavirus (HPV) have been approved, using Saccharomyces cerevisiae and Hansenula polymorpha as expression platforms (11, 12). In animals, certain commercial yeast-based vaccines targeting cattle ticks have been developed using Komagataella phaffii (formerly Pichia pastoris) as the expression system (13–15). Currently, yeasts offer practical advantages such as ease of genetic engineering, rapid growth, high biomass yield, and the absence of endotoxins. These features, combined with their capacity to express structurally complex recombinant proteins, make them highly suitable for large-scale vaccine production (16). This review outlines the current landscape of commercial vaccines produced in yeast. Although only a few yeast-based vaccines have been approved to date, they demonstrate the potential of yeast as a safe and versatile platform for producing recombinant vaccines. Reviewing these vaccines, their applications, and limitations provides valuable insight and highlights directions to improve their effectiveness, scalability, and applicability for future development in both human and animal health.
Yeast as a vaccine production platform: genetic engineering tools
Successful yeast heterologous protein expression depends on having different and efficient genetic engineering tools for cell transformation. The first genetic yeast transformation through DNA recombinant technology was reported by Hinnen et al. and Beggs in 1978 (17, 18). The initial attempt consisted of producing spheroplasts (cells without a wall) to introduce foreign DNA. Later in 1983, Ito et al. standardized a genetic transformation protocol for whole intact cells, opening the path for an easier method (19). Other methods have been developed over the decades to improve efficient yeast transformation for recombinant protein production (20, 21). Improving protein yield has been an achievement through 1) the plasmid design and 2) the evaluation of different promoters, both constitutive and inducible, to manage gene expression (22).
Since the 90s, different plasmids have been developed to regulate the expression of foreign genes. Yeast vectors could be classified into two groups: plasmids that contain the yeast centromere sequence (CEN) and plasmids with a 2 μ origin of replication (23). Plasmids with CEN (centromere) sequences are mitotically stable yeast replicates, with only a single copy present per cell, whereas the 2µ plasmids are multicopy (about 20 per cell). Christianson et al. (24) developed four episomal plasmids with a high copy number based on endogenous sequences of yeast replication origin 2µ from S. cerevisiae and the bacterial plasmid pBluescript, making a shuttle vector (24). Plasmids for protein expression have been mainly developed for S. cerevisiae, K. phaffii, Yarrowia lipolytica, and emerging yeasts used to express recombinant proteins (25–28).
Mumberg et al. (29) developed a series of plasmids to clone genes under the control of constitutive promoters, allowing different levels of protein yield (29). The different promoters used encompassed weak (CYC1) and strong (TEF and GPD) promoters. Interestingly, Drew and Kim (27) designed an expression plasmid using an inducible GAL1 promoter, the recombinant protein of interest, and the GFP-octa-histidine sequence, which is a multi-copy plasmid integrating by homologous recombination in S. cerevisiae (23).
There are many plasmids based on these basic genetic elements. An extensive review of promoters and terminators for the expression of recombinant protein in non-common yeast, including K. phaffii and Y. lipolytica, has recently been reviewed (30). Given the industrial importance of K. phaffii and its potential for vaccine production, we provide a brief description of the commonly employed promoter. Representative constitutive promoters include GAP, fsLovA, cTRDL, and TEF1; while inducible promoters, which use methanol as the inducer, comprise SNT5, iTRDL, and AOX1. Among these, the methanol-inducible AOX1 promoter is the most widely used, enabling high-level expression of heterologous proteins (31). This system is advantageous for producing vaccines, as it can yield proteins constituting up to 30% of total cell protein (32, 33). While the methanol-inducible system is effective, concerns regarding methanol’s toxicity and flammability have led to the exploration of safer alternatives to induction methods. For instance, some orthologous promoters from yeasts can outperform the expression levels of AOX1 promoter (34).
Downstream antigen purification and additional bioprocesses
Downstream processing of yeast-made antigens involves a series of purification techniques essential for recovering and refining bioproducts from fermentation broths. This process is critical for ensuring product purity and reducing production costs, particularly in the pharmaceutical industry. The key aspects of downstream yeast-made antigen purification and additional bioprocesses will be outlined.
The initial steps often involve removing microbial cells through centrifugation and filtration, which are crucial for clarifying the fermentation broth (35). Techniques such as protein A chromatography are effective for purifying antibodies and removing contaminants like β-glucan, which can pose immunogenic risks (36, 37). Other methods include ion exchange and affinity chromatography, which are tailored to the specific properties of the target biomolecule (37).
The liquid-liquid extraction method is commonly used to extract fermentation products, allowing for the separation of desired compounds from complex mixtures (38). The next step is the scale-up of the fermentation process, which involves optimizing the bioreactor (pH, dissolved oxygen, and medium composition). The fermentation processes used by biopharmaceutical manufacturers have demonstrated increased quantities of therapeutic proteins. However, this increase subsequently leads to capacity bottlenecks in the purification process (known as downstream processing) and is associated with high costs. Downstream processing comprises up to 80 percent of the total production costs (39, 40). Producers are increasingly recognizing the urgent need for improvement and have shifted their focus from enhancing the production process (upstream) to refining the downstream process (41, 42). This includes examining operating modes (batch, fed-batch, and continuous) and their impact on productivity. For instance, Martinez-Hernández et al. (43) developed a fed-batch bioprocess to produce a recombinant vaccine against Entamoeba histolytica in K. phaffii under operational conditions suitable for large-scale bioprocesses (43). Following the scale-up, the production of the recombinant protein reached 0.43 mg/mL, marking a 12-fold increase in production, despite the presence of methanol and oxygen-limited conditions. A maximum volumetric productivity of 3.75 mg/L h was achieved in a bioreactor, compared to 0.26 mg/L h attained in a shake flask. The next step involves assessing post-purification antigen stability, estimating the effects of freeze-thaw and lyophilization cycles, and analyzing protein integrity by SDS-PAGE and Western blot (44). Finally, in process validation, reproducibility must be verified in large-scale production batches (e.g., 10 L to 60 L) (45).
The comparison of purification yields and efficiencies between yeast and E. coli for recombinant antigen production reveals distinct edges and challenges associated with each host. Yeast, particularly species such as S. cerevisiae and K. phaffii, has shown promising results in producing functional recombinant proteins, while E. coli remains a widely used system due to its established protocols and high yields. Yeast can produce highly immunogenic proteins, as seen in the manufacture of vaccines against hepatitis B and HPV (46). Also, advances in understanding yeast metabolism have led to improved yields, especially for complex proteins (47). Companies employ various techniques to achieve effective purification in commercial yeast-based vaccines, focusing on optimizing processes that enhance both the purity and recovery of recombinant proteins. For instance, the purification of the SARS-CoV-2 receptor-binding domain utilized mixed-mode chromatography followed by hydrophobic interaction chromatography, achieving over 99% purity with a yield of approximately 21 mg/L (48). Tam et al. (49) have successfully applied a two-step purification strategy involving ion exchange chromatography (IEC) and size exclusion chromatography (SEC) to purify hepatitis B surface antigen, yielding a purity of 95.48% and a recovery rate of 78.07% (49). Additionally, the characteristics of yeast-based vaccines and the assembly of virus-like particles (VLPs) significantly influence purification efficiency. Particularly, K. phaffii is widely utilized for producing VLPs due to its ability to generate high levels of antigens like the Hepatitis B surface antigen (HBsAg) and poliovirus VLPs (50). However, the assembly process and the intracellular localization of these antigens can complicate the purification process.
On the other hand, E. coli can achieve considerable yields up to 23 mg of purified protein per liter of culture (51), and secretory production methods simplify the purification and improve protein folding crucial for functional proteins (52). Moreover, E. coli strains engineered for better disulfide bond formation can produce biologically active proteins more effectively than yeast in some cases (53). In summary, E. coli remains a robust choice for high-yield production and efficient purification, particularly for simpler proteins, while yeast offers advantages in producing complex proteins and vaccines. However, the choice of host ultimately depends on the specific requirements of the recombinant protein being produced. Finally, all processes must comply strictly with Good Manufacturing Practice (GMP) regulations for clinical trials, including long-term stability evaluations for regulatory approval. The GMP regulations are compulsory for ensuring the safety and efficacy of vaccines produced from yeast for clinical trials (54). These regulations encompass a range of processes, from initial production to quality control, guaranteeing that vaccines meet stringent safety standards before human administration (55).
Good manufacturing practices for compliance with regulatory issues
One of the key advantages of employing yeast systems for vaccine production lies in their well-established use in the biopharmaceutical industry. For several decades, yeasts have served as hosts to produce recombinant vaccines and other biotherapeutics approved for human use (8, 10). This historical precedent facilitates regulatory approval and streamlines compliance with GMP, ensuring that medicinal products are consistently produced and controlled according to rigorous quality standards appropriate for their intended use. GMP compliance is critical throughout the vaccine development pipeline, including preclinical stages, where assessments of immunogenicity, toxicity, and allergenicity are conducted using carefully selected and justified in vitro and in vivo models (56). These evaluations form the foundation for subsequent clinical trials, which are essential for regulatory approval (57).
The standards of GMP demand precise characterization and traceability of production strains, underscoring the advantages of yeast-based systems over mammalian cells in recombinant protein manufacturing. Yeast cultures are inherently less susceptible to contamination by bacteria, fungi, mycoplasma, and viruses, a resilience largely attributed to their ability to proliferate in acidic environments that suppress the growth of many microbial contaminants (58, 59). Notably, yeast-based production also circumvents the risk of endotoxin contamination, a common concern in bacterial systems due to lipopolysaccharide residues (60). Furthermore, VLPs generated in yeast offer a compelling alternative to conventional vaccines derived from attenuated or inactivated viruses. In addition to the economic benefits of yeast cultivation, VLPs lack genetic material, thereby enhancing their safety profile—particularly for immunocompromised individuals (61).
In general terms for vaccine production, adherence to local regulatory frameworks is necessary to avoid delays or complications; however, the primary GMP guidelines are issued by major international agencies such as the European Medicines Agency (EMA), the U.S. Food and Drug Administration (FDA), and the World Health Organization (WHO). To further harmonize regulatory requirements and reduce redundancy in clinical and non-clinical studies, the International Council for Harmonisation of Technical Requirements for Pharmaceuticals for Human Use (ICH) was established (http://ich.org/). The ICH framework represents a significant advancement in aligning global standards, minimizing unnecessary duplication of human trials, and reducing animal use without compromising the integrity of safety and efficacy data. Within the GMP framework, the ICH Quality Guidelines are particularly relevant to vaccine development. These guidelines span 14 core areas, covering topics from stability testing (Q1A–Q1F) to analytical procedure development (Q14), offering a robust foundation for ensuring product quality and regulatory compliance (62). In the United States, the FDA regulates vaccine development and manufacturing under Subchapter F—Biologics, while GMP requirements for drug manufacturing, processing, packaging, and storage are detailed in Subchapter C—Drugs: General. Additional relevant provisions are found in Subchapters D—Drugs for Human Use and A—General (63). The EMA complements these regulations with its Good Pharmacovigilance Practices (GVP) guidelines, which encompass risk management plans, including detailed product profiles (e.g., vaccine composition, adjuvants, preservatives, and residuals), safety specifications, pharmacovigilance strategies, post-authorization efficacy studies, and risk minimization measures (64). Similarly, the WHO provides comprehensive guidance, such as the Guidelines on Clinical Evaluation of Vaccines (65). Regulatory aspects have been fundamental to improving the safety and efficacy of vaccines against infectious diseases. In this context, yeast-based systems offer several advantages for vaccine development: enhanced safety due to the absence of lipopolysaccharides (LPS), and their recombinant protein-based nature enables safe administration in immunocompromised individuals, positioning them as a preferable alternative to attenuated vaccines; scalability enabled by the use of low-cost culture media with minimal risk of contamination; and strong immunogenicity through the potential to produce VLPs-based vaccines.
Human vaccines produced in yeast
To date, at least several recombinant vaccines expressed in yeast have been approved by the U.S. Food and Drug Administration (FDA), European Medicines Agency (EMA), and WHO for human use (see Table 1 and Supplementary Figure S1). Leading manufacturers in this field include Merck & Co., Inc., GlaxoSmithKline Biologicals, Dynavax Technologies Corporation, and Sanofi Pasteur Inc. While the vaccines from these pharmaceutical companies are widely used across many countries, it is worth noting that China has developed a significant portfolio of domestically produced vaccines. These include several HPV vaccines manufactured using yeast-based expression systems—some of which have already been approved, such as the Shanghai Zerun vaccine (Walvax)—as well as others currently in various clinical trial stages (66, 67). Notably, two yeast-derived vaccines—Mosquirix™ (GlaxoSmithKline) and R21/Matrix-M™ (Oxford University, Novavax, Inc., and Serum Institute of India)—have been recommended by the World Health Organization for the prevention of malaria in children residing in endemic regions (68).
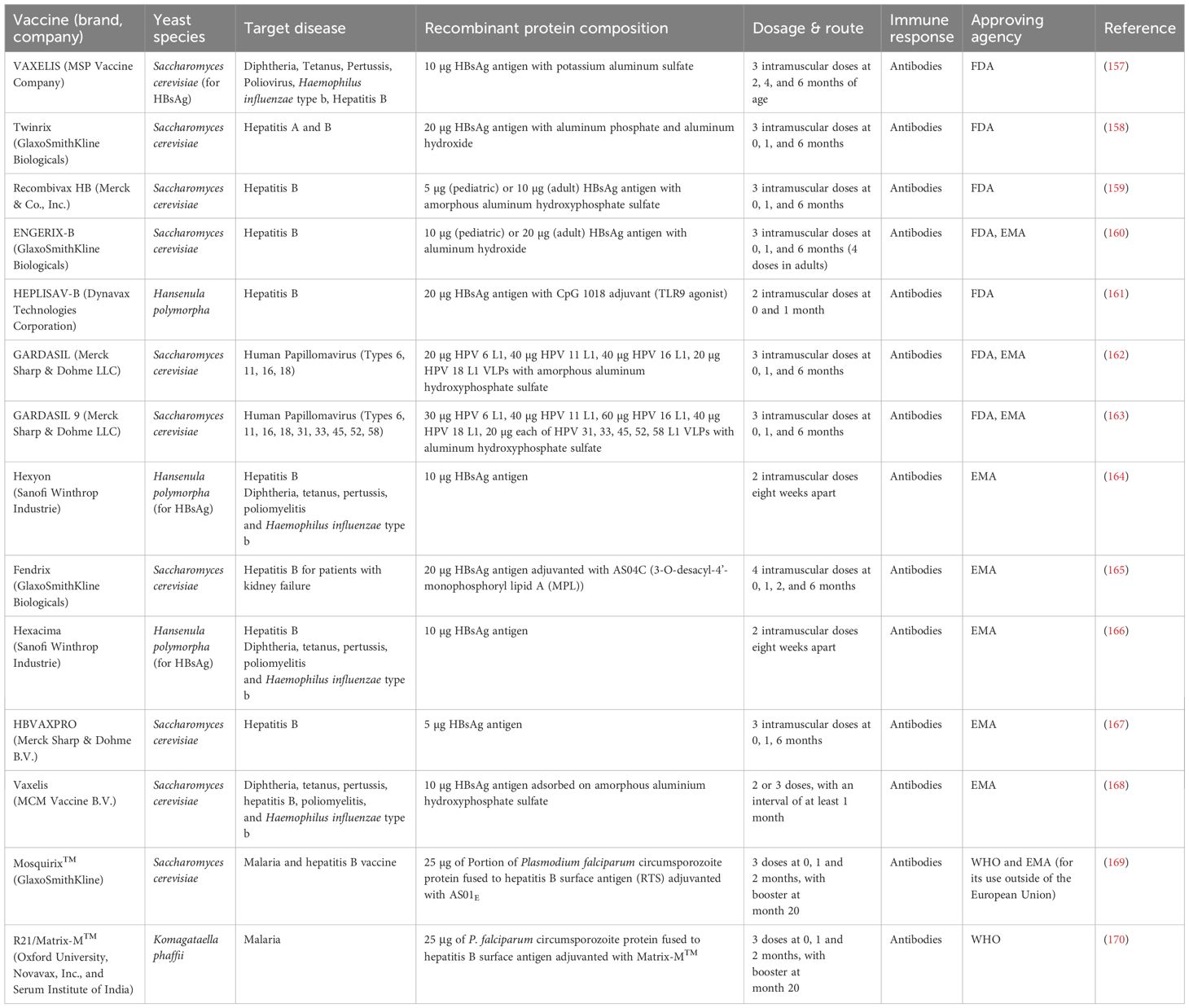
Table 1. Recombinant yeast-expressed vaccines approved by the FDA, EMA, and WHO for human use.
Saccharomyces cerevisiae remains the preferred host for recombinant antigen expression; however, the hepatitis B vaccine HEPLISAV-B, Hexyon, and Hexacima are produced in Hansenula polymorpha, whereas the R21/Matrix-M™ vaccine is produced in K. phaffii. These yeasts are classified as Generally Recognized As Safe (GRAS), capable of growing in low-cost media and yielding high levels of recombinant protein (11, 12). Despite the use of similar genetic transformation tools, these yeasts differ in their regulatory protein expression elements (69, 70). Currently, yeast-expressed vaccines target hepatitis B, malaria and various types of HPV, all formulated as virus-like particles (VLPs). VLP-based strategies offer enhanced immunogenicity compared to non-assembled recombinant proteins (71). Despite more complex proteins requiring mammalian systems, such as glycoprotein E in the SHINGRIX vaccine (produced in Chinese hamster ovary cells), many yeast-derived recombinant vaccines without multiple post-translational modifications and capable of forming VLPs have demonstrated efficacy rates exceeding 95% (72). The main immune response involved in these vaccines is through the production of highly neutralizing antibodies, where the adjuvants play a critical role in the immunogenicity of yeast-based vaccines. Most adjuvanted formulations rely on aluminum salts (see Table 1). An exception is Dynavax’s HEPLISAV-B, which incorporates CpG 1018, a Toll-like receptor 9 (TLR9) agonist. Additional examples include malaria vaccines. Mosquirix™ employs the AS01E adjuvant, which consists of Quillaja saponaria Molina fraction 21 (QS-21) and 3-O-desacyl-4’-monophosphoryl lipid A (MPL). In contrast, R21/Matrix-M™ utilizes the Matrix-M™ adjuvant—a saponin-based formulation developed by Novavax AB (Uppsala, Sweden)—which elicits both Th1 and Th2 immune responses.
Notably, several yeast-derived vaccine candidates are currently undergoing evaluation (see Table 2). One example is the same malaria vaccine R21/Matrix-M™ expressed in H. polymorpha, forming VLPs, which is being evaluated in adult populations (ClinicalTrials.gov ID: NCT05252845). On the other hand, several vaccine prototypes have been evaluated in preclinical studies, including those targeting complex antigens such as the SARS-CoV-2 spike protein and influenza hemagglutinin. These proteins require specific glycosylation patterns, which have driven the development of innovative strategies to optimize yeast expression systems for industrial-scale production (73–75).
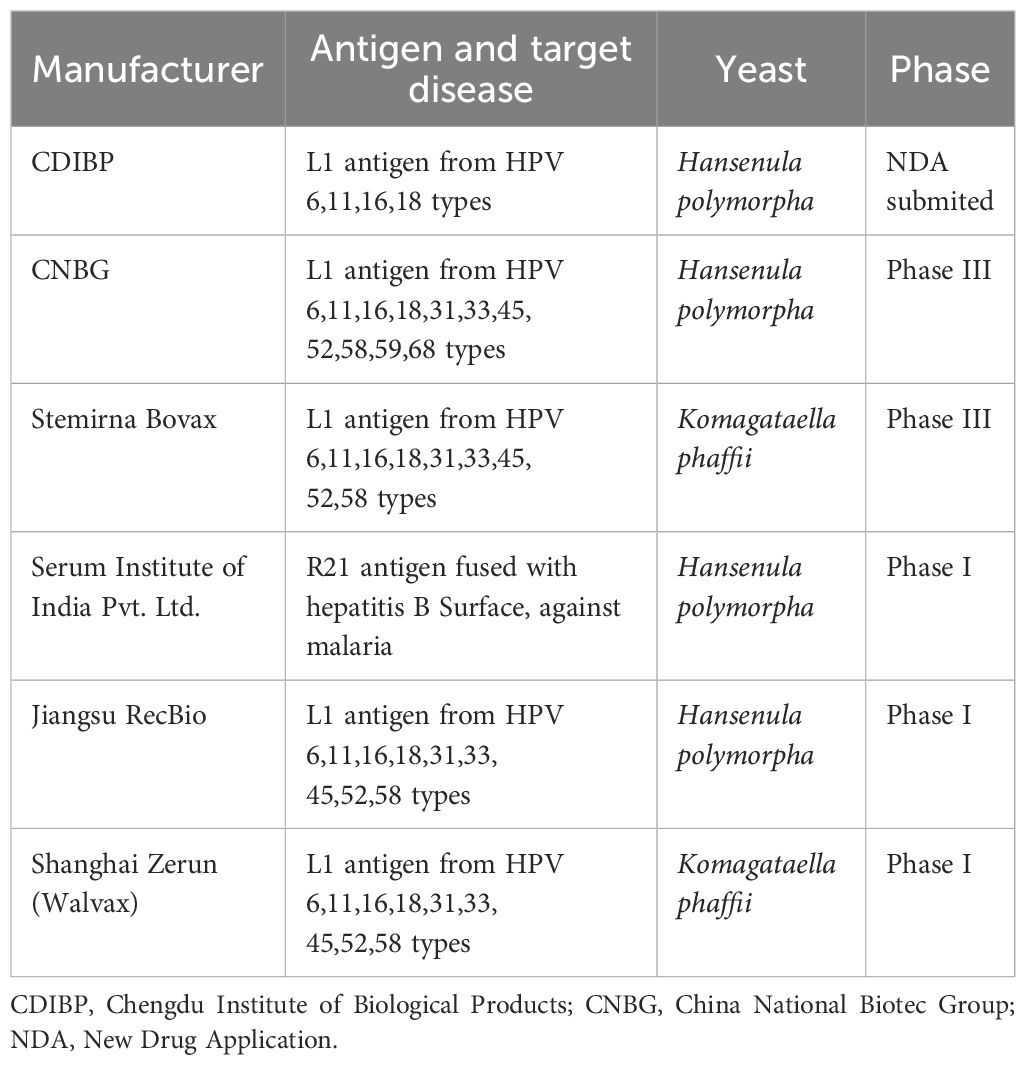
Table 2. Examples of recombinant yeast-expressed vaccines in clinical trials.
Animal vaccines produced in yeast
There are many recombinant yeast-based vaccines at preclinical levels, particularly useful in livestock, poultry, aquaculture, and companion animals, providing targeted protection against pathogenic viruses, bacteria, and parasites (see Supplementary Figure S1) (8, 76, 77). In birds, yeast-produced vaccines include those targeting fowl adenovirus (FAdV) (78) and parasites such as Eimeria tenella (79), where S. cerevisiae-expressed antigenic proteins serve as effective subunit vaccines. For fish, yeast-based vaccines expressing antigens from Cyprinid herpesvirus 2 and 3 have been developed to protect species such as Gibel carp (Carassius auratus gibelio), common carp (Cyprinus carpio) against these viral infections (80–82). In cattle, recombinant yeast vaccines expressing proteins from Babesia bovis (83) (and Theileria parva (84) are being investigated to reduce the impact of tick-borne diseases. For pigs, yeast-expressed vaccines against porcine circovirus type 2 (PCV2) and African swine fever virus (ASFV) have shown promising immune protection (85, 86). In pets, experimental yeast-based vaccines have demonstrated strong immunogenicity against hookworm, Ancylostoma caninum, and canine distemper virus (CDV) in dogs and feline infectious peritonitis virus (FIPV) in cats (75, 87, 88). However, in terms of commercial availability for veterinary purposes, there are just a few recombinant vaccines expressed in yeast, precisely in K. phaffii, and only against ticks (13, 15, 89–91). Although there are several commercial subunit recombinant veterinary vaccines on the market, most of them have been produced in Escherichia coli or baculovirus systems (92, 93). The E. coli expression system is the most widely used due to its simplicity, rapid growth, and low production cost. It allows high yields of recombinant proteins in a short time, making it ideal for large-scale vaccine manufacturing (94, 95). However, E. coli lacks the machinery for post-translational modifications such as glycosylation, which can limit its use for complex eukaryotic antigens (96). On the other hand, the baculovirus system offers a eukaryotic environment that enables proper protein folding, disulfide bond formation, and post-translational modifications, producing more structurally and functionally authentic antigens. This system is especially valuable for glycoproteins and complex multimeric proteins used in vaccines (97, 98). The baculovirus system is more expensive and slower than E. coli but often improves vaccine production and efficacy of immunologically relevant proteins (99). In contrast, the yeast system is a cheap platform that can perform functional post-translational modifications, and its disadvantage is over-glycosylation of proteins or the production of non-mammalian glycan patterns, which could affect vaccine immunogenicity (100, 101). Here, the recombinant yeast-based vaccines commercially available for animals will be reviewed by analyzing doses, efficacy, and immune responses.
Regarding commercial vaccines, the yeast-made vaccine Gavac® (Heber Biotec S.A., Havana, Cuba) and TickGARD® (Biotech Australia Pty. Ldt.) are composed of recombinant Rhipicephalus microplus Bm86 gut antigen expressed in K. phaffii that allows protection against cattle ticks (13, 14, 90, 102). These vaccines are administered in the neck in multiple doses in a recommended schedule of two initial doses on days 0 and 28, followed by a booster dose every 6 months (103). The effectiveness of Bm86-based vaccines like Gavac® and TickGARD® has been assessed through parameters such as tick survival, egg production, and fertility. Studies show variable efficacy (>50% in some cases), influenced by tick strain and cattle breed. Interestingly, vaccines reduced acaricide use by up to 87% and saved $23.4 per animal/year (15). However, efficacy varies due to tick genetic diversity, particularly polymorphisms in the Bm86 gene (104). Additionally, in 2018, Mexico developed Bovimune Ixovac® using the “Media Joya” strain and expressed it in K. phaffii (105). It reduced tick infestations to <10 per animal and tick baths from 14 to 3 per year, with an 86% drop in tick fertility. In Colombia, Tick-Vac®, developed by LIMOR and marketed by TECNOQUÍMICAS, uses Bm86 from tick larvae. It follows a three-dose initial schedule with biannual boosters. Field trials showed 80% protection, with clinical studies reporting 64–96% efficacy and reduced parasite loads across various agroecosystems (15).
The immunological outcomes behind vaccination rely on antigen-presenting cells like dendritic cells and macrophages that process and present Bm86 via bovine leukocyte antigen. This activates naive T cells, which help activate B cells in germinal centers, leading to the formation of plasma and memory B cells (106). Plasma cells produce specific antibodies, while memory cells remain in lymphoid organs awaiting booster doses. Antibodies (IgM and IgG) circulate in the blood and are ingested by ticks during feeding. They bind to Bm86 in the tick gut and activate the classical complement pathway, causing enterocyte lysis and impairing digestion and overall tick health (107, 108). In general, yeast-made veterinary vaccines are an option to fight against animal diseases that remains to be fully exploited.
Limitations and future directions
Yeast as eukaryotic cells have many advantages to produce recombinant vaccines over mammalian cells and bacterial counterparts. However, several disadvantages have led to limited success in reaching commercial vaccines (See Figure 1). The main issues revolve around proper glycosylation, accurate formation of VLPs, and high yields. Although glycosylation is essential for the structural and functional integrity of many antigens—and yeast is capable of performing this post-translational modification—certain glycosylation patterns, such as high-mannose N-glycans, pose challenges to protein stability. These modifications can alter the expected glycosylation profile and critically impact both the immunogenicity and allergenicity of antigens (107). To address these issues, advances in glycoengineering have partially mitigated such limitations (109). In the context of yeast genetic engineering, efforts have focused on generating knockout strains of S. cerevisiae targeting genes involved in mannosylphosphate biosynthesis (110). Particularly, genes such as Mnn2p, Mnn11p, and α-1, 6-mannosyltransferase Och1p have been disrupted to reach this goal (29). Parallel strategies have explored the overexpression of endoglycosidase genes to enhance glycosylation efficiency (111). Both approaches hold promise for the production of complex antigens with more human-like glycosylation profiles.
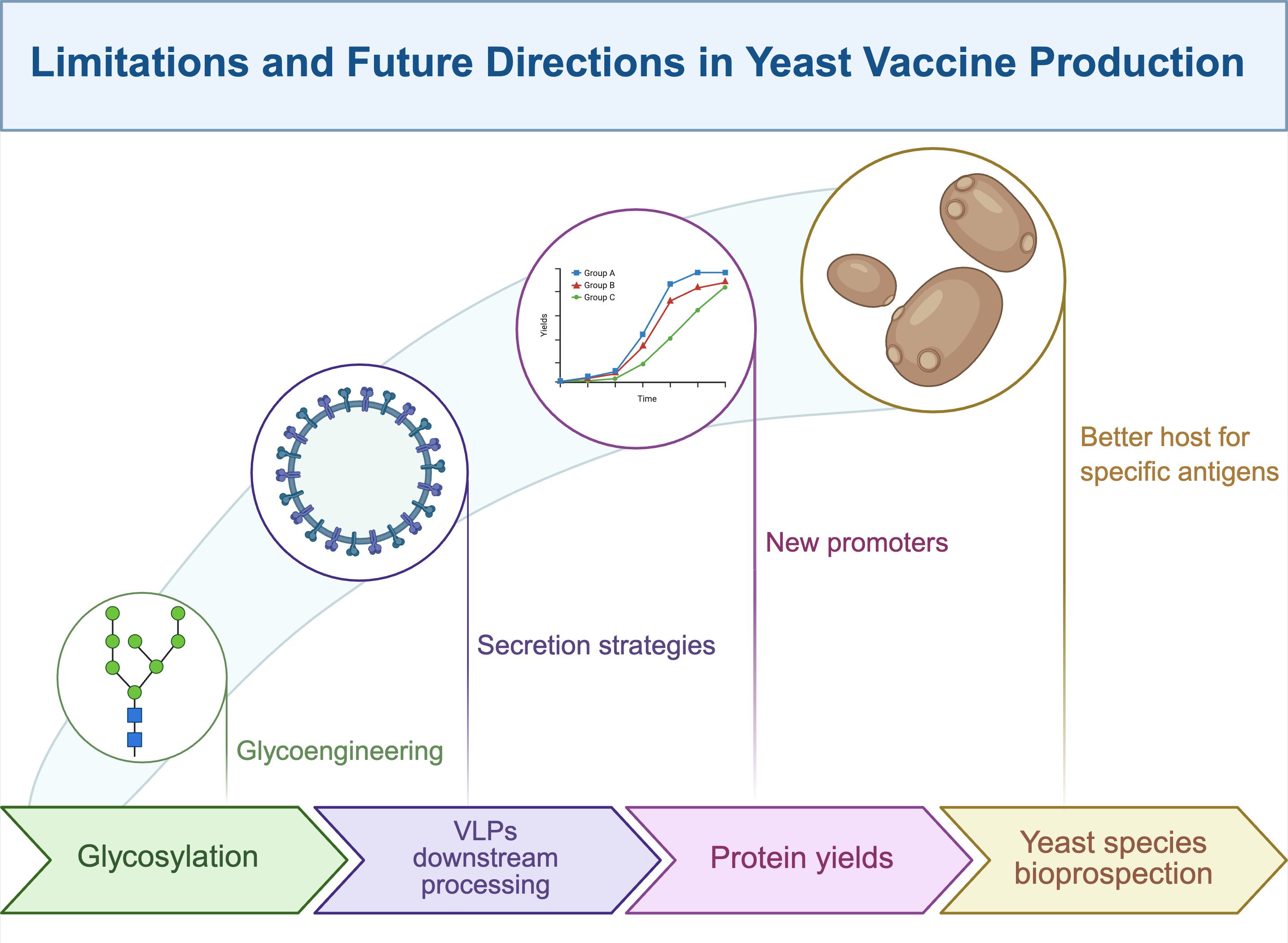
Figure 1. Limitations and future directions in yeast vaccine production.
An alternative strategy involves modifying antigen sequences to eliminate glycosylation sites. For instance, research groups developing yeast-based vaccines against SARS-CoV and SARS-CoV-2 have evaluated the expression in K. phaffii of deglycosylated receptor-binding domain (RBD) fragments. In these constructs, N-linked glycosylated asparagine residues were removed (112). The resulting proteins exhibited high yields without compromising antigenicity and elicited even stronger neutralizing antibody responses than their glycosylated antigen. Building on this approach, subsequent work during the SARS-CoV-2 pandemic targeted Asn331 and Cys538, again demonstrating robust induction of neutralizing antibodies (74). Interestingly, the CRISPR-Cas technology has emerged as a novel tool for genome modification applied in yeast, achieving successful glycoengineering (113). Collectively, these efforts underscore the potential of yeast as a cost-effective platform to produce complex antigens from pathogens of epidemic and pandemic relevance.
As previously discussed, licensed yeast-derived vaccines in humans predominantly rely on VLPs as antigenic platforms. Despite their success, several challenges remain in the production and optimization of VLP-based vaccines. These include efficient VLP assembly and secretion, the formation of enveloped and non-enveloped VLPs, and the tendency of VLPs to aggregate—often necessitating the use of anti-aggregation agents (76, 114). In most cases, VLPs expressed in yeast accumulate intracellularly, which poses significant limitations for large-scale production due to the need for cell lysis and complex downstream processing (115, 116). To circumvent these issues and reduce purification costs, secretion strategies have been explored, notably through the use of signal peptides such as the α-mating factor secretion signal (117, 118). However, unlike other eukaryotic systems capable of producing enveloped VLPs via budding from the plasma membrane (119), yeast lacks this capability under normal conditions. Enveloped VLP production in yeast has only been achieved through the generation of spheroplasts by enzymatic removal of the cell wall (120). A promising strategy involves the co-assembly of VLPs incorporating multiple antigens. These multilayered or mosaic VLPs may enhance vaccine efficacy or confer multivalent protection against diverse viral strains or species (121, 122).
A relevant issue is the improvement of recombinant protein yields. This aspect has claimed attention for research focused on strong AOX1 and GAP promoters (123–125), along with new promoters recently reported (126, 127). Additionally, it is well-known that yeast-preferred codons enhance recombinant antigen production (128). Of interest to improve yeast-made vaccines is that higher copy gene number through repeated sequences can also proportionally increase antigen transcript and recombinant protein (129). Even if high recombinant protein yields can be accurately reached, the downstream protein recovery and purification process faces additional challenges. To surpass this issue, for instance, cell disruption methods have been proposed for better recovery of recombinant antigens, especially those based on response surface methodology (130, 131). Innovations in protein purification have also been pursued, such as non-affinity chromatographic methods (132, 133). Moreover, improvements in classical ion (134, 135) and anion (136) exchange chromatography are being investigated to remove aggregates and optimize the purification process. Notably, current research on optimized buffers during down-processing is relevant for VLP-based recombinant vaccines (137). Lastly, partial purification as a downstream process using nano-colloidal silica adsorbent (Aerosil-380) can help in the separation of antigens (138), serving as a clarification process before final purification through non-affinity and affinity methods. In this regard, the use of nanoparticles is a future direction that promises high-efficiency nano-systems for the purification of antigens produced in yeast (139).
It is convenient to mention that the production of commercial recombinant vaccines in yeast has been limited to S. cerevisiae, K. phaffii, and H. polymorpha. However, many yeast species should be explored to find a better host to produce specific antigens on a case-by-case basis. It means a battery of available yeasts to select the best host in terms of yields, functionality, and safety. Prospects in this direction include the use of Kluyveromyces lactis (140–142), Yarrowia lipolytica (140), Schizosaccharomyces pombe (143–145), and Saccharomyces boulardii (146), in which antigens have been recombinantly produced. In addition to commercial vaccines, efforts in research & development have led to experimental vaccines with promising outcomes to prevent human and animal infectious diseases. Among many others, the most recent studies include experimental vaccines against rabies (147), herpesvirus hematopoietic necrosis (148, 149), Covid-19 (150), polio (151), bovine mastitis (152), tuberculosis (153), ETEC (154), human papillomavirus type 52 (155), and cancer (156).
Conclusion
Yeasts are an antigen production platform for which genetic engineering tools, downstream processes, and industrial manufacturing have led to the benefit of vaccine commercialization to prevent hepatitis B, HPV, malaria, and tick parasitosis (Rhipicephalus microplus). In addition, efforts in R&D of experimental vaccines in preclinical and clinical studies hope to reach commercialization soon to fight against human and animal infectious diseases.
Author contributions
AR-V: Writing – original draft, Funding acquisition, Visualization, Writing – review & editing, Conceptualization. EM-E: Writing – original draft. BB-H: Writing – original draft. MA: Writing – original draft. ET: Writing – original draft. CA: Writing – review & editing, Conceptualization, Writing – original draft.
Funding
The author(s) declare financial support was received for the research and/or publication of this article. This article is part of the funding provided by the Instituto Politécnico Nacional under the SAD-DPP-1883-2025 “Proyecto de Consorcio IPN-UAM-UAEMex” and the SIP 20251094 “Proyecto Multidisciplinario”.
Acknowledgments
The authors wish to thank IBQ. Hassian Javier León Montoya for providing formatting writing support.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The author(s) declared that they were an editorial board member of Frontiers, at the time of submission. This had no impact on the peer review process and the final decision.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fimmu.2025.1697177/full#supplementary-material
Supplementary Figure 1 | Most representative vaccines produced in yeast.
References
1. Baral B, Mamale K, Gairola S, Chauhan C, Dey A, and Kaundal RK. Infectious diseases and its global epidemiology. In: Nanostructured Drug Delivery Systems in Infectious Disease Treatment. Academic Press (2024). p. 1–24. doi: 10.1016/B978-0-443-13337-4.00017-3
2. Weiss RA and Sankaran N. Emergence of epidemic diseases: zoonoses and other origins. Fac Rev. (2022) 11:2. doi: 10.12703/r/11-2
3. Bungau S, Tit DM, Behl T, Aleya L, and Zaha DC. Aspects of excessive antibiotic consumption and environmental influences correlated with the occurrence of resistance to antimicrobial agents. Curr Opin Environ Sci Health. (2021) 19:100224. doi: 10.1016/j.coesh.2020.10.012
4. Rappuoli R, Pizza M, Del Giudice G, and De Gregorio E. Vaccines, new opportunities for a new society. Proc Natl Acad Scie. (2014) 111:12288–93. doi: 10.1073/pnas.1402981111
5. Ghattas M, Dwivedi G, Lavertu M, and Alameh MG. Vaccine technologies and platforms for infectious diseases: current progress, challenges, and opportunities. Vaccines (Basel). (2021) 9:1490. doi: 10.3390/vaccines9121490
6. de Pinho Favaro MT, Atienza-Garriga J, Martiníez-Torró C, Parladé E, Vázquez E, and Corchero JL. Recombinant vaccines in 2022: a perspective from the cell factory. Microb Cell Fact. (2022) 21:203. doi: 10.1186/s12934-022-01929-8
7. Kumar V, Verma A, Singh R, Garg P, Sharma SK, and Singh HN. Recombinant vaccines: Current updates and future prospects. Asian Pac J Trop Med. (2024) 17:338–50. doi: 10.4103/apjtm.apjtm_854_23
8. Kumar R and Kumar P. Yeast-based vaccines: New perspective in vaccine development and application. FEMS Yeast Res. (2019) 19:foz007. doi: 10.1093/femsyr/foz007
9. Valenzuela P, Medina A, Rutter WJ, Ammerer G, and Hall BD. Synthesis and assembly of hepatitis B virus surface antigen particles in yeast. Nature. (1982) 298:347–50. doi: 10.1038/298347a0
10. McAleer WJ, Buynak EB, Maigetter RZ, Wampler DE, Miller WJ, and Hilleman MR. Human hepatitis B vaccine from recombinant yeast. Nature. (1984) 307:178–80. doi: 10.1038/307178a0
11. Da Silva NA and Srikrishnan S. Introduction and expression of genes for metabolic engineering applications in Saccharomyces cerevisiae. FEMS Yeast Res. (2012) 12:197–214. doi: 10.1111/j.1567-1364.2011.00769.x
12. Manfrão-Netto JHC, Gomes AMV, and Parachin NS. Advances in using hansenula polymorpha as chassis for recombinant protein production. Front Bioeng Biotechnol. (2019) 7:94. doi: 10.3389/fbioe.2019.00094
13. Canales M, Enríquez A, Ramos E, Cabrera D, Dandie H, and Soto A. Large-scale production in Pichia pastoris of the recombinant vaccine Gavac™ against cattle tick. Vaccine. (1997) 15:414–22. doi: 10.1016/S0264-410X(96)00192-2
14. Jonsson NN, Matschoss AL, Pepper P, Green PE, Albrecht MS, and Hungerford J. Evaluation of TickGARDPLUS, a novel vaccine against Boophilus microplus, in lactating Holstein–Friesian cows. Vet Parasitol. (2000) 88:275–85. doi: 10.1016/S0304-4017(99)00213-7
15. Galván Arellano DM, Vieyra Reyes P, Montes de Oca Jiménez R, Ortega Lopez J, Martínez Arzate SG, Rivas Santiago B, et al. Proteína Bm86 y su potencial uso como vacuna contra garrapatas en el ganado bovino. Revisión. Rev Mex Cienc Pecu. (2023) 14:672–95. doi: 10.22319/rmcp.v14i3.6255
16. Lestari CSW and Novientri G. Advantages of yeast-based recombinant protein technology as vaccine products against infectious diseases. IOP Conf Ser Earth Environ Sci. (2021) 913:12099. doi: 10.1088/1755-1315/913/1/012099
17. Hinnen A, Hicks JB, and Fink GR. Transformation of yeast. Proc Natl Acad Sci. (1978) 75:1929–33. doi: 10.1073/pnas.75.4.1929
18. Beggs JD. Transformation of yeast by a replicating hybrid plasmid. Nature. (1978) 275:104–9. doi: 10.1038/275104a0
19. Ito H, Fukuda Y, Murata K, and Kimura A. Transformation of intact yeast cells treated with alkali cations. J Bacteriol. (1983) 153:163–8. doi: 10.1128/jb.153.1.163-168.1983
20. Tuite MF. Strategies for the genetic manipulation of saccharomyces cerevisiae. Crit Rev Biotechnol. (1992) 12:157–88. doi: 10.3109/07388559209069191
21. Fraczek MG, Naseeb S, and Delneri D. History of genome editing in yeast. Yeast. (2018) 35:361–8. doi: 10.1002/yea.3308
22. Baghban R, Farajnia S, Rajabibazl M, Ghasemi Y, Mafi A, and Hoseinpoor R. Yeast expression systems: overview and recent advances. Mol Biotechnol. (2019) 61:365–84. doi: 10.1007/s12033-019-00164-8
23. Drew D and Kim H. Preparation of Saccharomyces cerevisiae Expression Plasmids. Humana Press (2012). pp. 41–6. pp. 41–6. doi: 10.1007/978-1-61779-770-5_4.
24. Christianson TW, Sikorski RS, Dante M, Shero JH, and Hieter P. Multifunctional yeast high-copy-number shuttle vectors. Gene. (1992) 110:119–22. doi: 10.1016/0378-1119(92)90454-W
25. Ling H, Teo W, Chen B, Leong SSJ, and Chang MW. Microbial tolerance engineering toward biochemical production: from lignocellulose to products. Curr Opin Biotechnol. (2014) 29:99–106. doi: 10.1016/j.copbio.2014.03.005
26. Peralta-Yahya PP, Zhang F, del Cardayre SB, and Keasling JD. Microbial engineering for the production of advanced biofuels. Nature. (2012) 488:320–8. doi: 10.1038/nature11478
27. Parker W, Taylor A, Razdan A, Escarce J, and Crook N. Enabling technologies for in situ biomanufacturing using probiotic yeast. Adv Drug Delivery Rev. (2025) 223:115605. doi: 10.1016/j.addr.2025.115605
28. Yook S, Deewan A, Ziolkowski L, Lane S, Tohidifar P, and Cheng M. Engineering and evolution of Yarrowia lipolytica for producing lipids from lignocellulosic hydrolysates. Bioresour Technol. (2025) 416:131806. doi: 10.1016/j.biortech.2024.131806
29. Mumberg D, Müller R, and Funk M. Yeast vectors for the controlled expression of heterologous proteins in different genetic backgrounds. Gene. (1995) 156:119–22. doi: 10.1016/0378-1119(95)00037-7
30. Moon SY, An NY, and Lee JY. Transforming non-conventional yeasts into key players in biotechnology: advances in synthetic biology applications. Front Microbiol. (2025) 16:1600187. doi: 10.3389/fmicb.2025.1600187
31. Cregg JM, Barringer KJ, Hessler AY, and Madden KR. Pichia pastoris as a host system for transformations. Mol Cell Biol. (1985) 5:3376–85. doi: 10.1128/mcb.5.12.3376-3385.1985
32. Kielkopf CL, Bauer W, and Urbatsch IL. Expression of cloned genes in pichia pastoris using the methanol-inducible promoter AOX1. Cold Spring Harb Protoc. (2021) 2021:pdb.prot102160. doi: 10.1101/pdb.prot102160
33. Mastropietro G, Aw R, and Polizzi K. Expression of proteins in Pichia pastoris. Methods Enzymology. (2021) 660:53–80. doi: 10.1016/bs.mie.2021.07.004
34. Vogl T, Fischer JE, Hyden P, Wasmayer R, Sturmberger L, and Glieder A. Orthologous promoters from related methylotrophic yeasts surpass expression of endogenous promoters of Pichia pastoris. AMB Express. (2020) 10:38. doi: 10.1186/s13568-020-00972-131
35. Stanbury PF, Whitaker A, and Hall SJ. The recovery and purification of fermentation products. Princ Ferment Technol. (2017), 619–86. doi: 10.1016/B978-0-08-099953-1.00010-7
36. Wang F. Demonstrating β-glucan clearance in CHO- and yeast-produced monoclonal antibodies during downstream purification processes. J Bioprocess Biotech. (2014) 4:2. doi: 10.4172/2155-9821.1000185
37. Bedade D and Pawar S. Downstream processing of biotechnology products. In: Basic Biotech Bioproc Bioentrepreneursh. Academic Press (2023). p. 377–90. doi: 10.1016/B978-0-12-816109-8.00026-X
38. Najafpour GD. Downstream processing. In: Biochemical Engineering and Biotechnology. Elsevier (2015). p. 227–56. doi: 10.1016/B978-0-444-63357-6.00007-9
39. Gottschalk U. Downstream processing. Filtr Purif Biopharm Ind. (2019), 349–82. doi: 10.1201/9781315164953-15
40. Sinha R, Goel N, Fatima H, and Khare SK. Advances in downstream processing techniques. Curr Dev Biotechnol Bioeng. (2022), 353–71. doi: 10.1016/B978-0-323-91167-2.00007-1
41. Prashad M and Tarrach K. Depth filtration: Cell clarification of bioreactor offloads. Filtr Sep. (2006) 43:28–30. doi: 10.1016/S0015-1882(06)70950-8
42. Singh N and Herzer S. Downstream processing technologies/capturing and final purification. In Adv Biochem Eng Biotechnol. (2017) 165:115–78. doi: 10.1007/10_2017_12
43. Martínez-Hernández SL, Marin-Muñoz MA, Ventura-Juarez J, and Jauregui J. Fed-batch cultivation and operational conditions for the production of a recombinant anti-amoebic vaccine in Pichia pastoris system. Rev Mex Ing Quim. (2019) 19:691–705. doi: 10.24275/rmiq/Bio725
44. Liu WC, Inwood S, Gong T, Sharma A, Yu LY, and Zhu P. Fed-batch high-cell-density fermentation strategies for Pichia pastoris growth and production. Crit Rev Biotechnol. (2019) 39:258–71. doi: 10.1080/07388551.2018.1554620
45. De S, Mattanovich D, Ferrer P, and Gasser B. Established tools and emerging trends for the production of recombinant proteins and metabolites in Pichia pastoris. Essays Biochem. (2021) 65:293–307. doi: 10.1042/EBC20200138
46. Bill RM. Playing catch-up with Escherichia coli: using yeast to increase success rates in recombinant protein production experiments. Front Microbiol. (2014) 5:85. doi: 10.3389/fmicb.2014.00085
47. Bill RM. Recombinant protein subunit vaccine synthesis in microbes: a role for yeast? Jou Pha Pha. (2015) 67:319–28. doi: 10.1111/jphp.12353
48. Nagar G, Jain S, Rajurkar M, Lothe R, Rao H, and Majumdar S. Large-scale purification and characterization of recombinant receptor-binding domain (RBD) of SARS-coV-2 spike protein expressed in yeast. Vaccines (Basel). (2023) 11:1602. doi: 10.3390/vaccines11101602
49. Tam YJ, Zeenathul NA, Rezaei MA, Mohd A, Bahaman AR, and Lo SC. Two-step purification strategy for enhanced recovery of recombinant hepatitis B surface antigen from Pichia pastoris. Sep Sci Technol. (2016) 51:816–29. doi: 10.1080/01496395.2015.1135949
50. Lünsdorf H, Gurramkonda C, Adnan A, Khanna N, and Rinas U. Virus-like particle production with yeast: ultrastructural and immunocytochemical insights into Pichia pastoris producing high levels of the Hepatitis B surface antigen. Microb Cell Fact. (2011) 10:48. doi: 10.1186/1475-2859-10-48
51. Maltoni G, Scutteri L, Mensitieri F, Piaz FD, and Hochkoeppler A. High-yield production in Escherichia coli and convenient purification of a candidate vaccine against SARS-CoV-2. Biotechnol Lett. (2022) 44:1313–22. doi: 10.1007/s10529-022-03298-z
52. Choi JH and Lee SY. Secretory and extracellular production of recombinant proteins using Escherichia coli. Appl Microbiol Biotechnol. (2004) 64:625–35. doi: 10.1007/s00253-004-1559-9
53. Wang Z, Böttcher D, Bornscheuer UT, and Müller C. Expression of recombinant hirudin in bacteria and yeast: A comparative approach. Methods Protoc. (2025) 8:89. doi: 10.3390/mps8040089
54. Gomez PL, Robinson JM, and Rogalewicz JA. Vaccine manufacturing. Vaccines. (2013), 44–57. doi: 10.1016/B978-1-4557-0090-5.00019-7
55. Ashfield R. GMP manufacture and quality control. Vaccinol Methods Vaccine Res. (2022), 281–94. doi: 10.1016/B978-0-323-91146-7.00004-4
56. Covarrubias CE, Rivera TA, Soto CA, Deeks T, and Kalergis AM. Current GMP standards for the production of vaccines and antibodies: An overview. Front Public Health. (2022) 10:1021905. doi: 10.3389/fpubh.2022.1021905
57. Gräf DD, Westphal L, and Hallgreen CE. The life cycle of vaccines evaluated by the European Medicines Agency. Vaccine. (2024) 42:126186. doi: 10.1016/j.vaccine.2024.126186
58. Liu X, Jia B, Sun X, Ai J, Wang L, and Wang C. Effect of initial PH on growth characteristics and fermentation properties of saccharomyces cerevisiae. J Food Sci. (2015) 80:M800–M808. doi: 10.1111/1750-3841.12813
59. Barone PW, Wiebe ME, Leung JC, Hussein LITM, Keumurian FJ, and Bouressa J. Viral contamination in biologic manufacture and implications for emerging therapies. Nat Biotechnol. (2020) 38:563–72. doi: 10.1038/s41587-020-0507-2
60. Svraka L, Abdallah HB, and Johansen C. When recombinant proteins go wrong: The hidden pitfall of recombinant protein contamination. Cytokine. (2025) 186:156830. doi: 10.1016/j.cyto.2024.156830
61. Mohsen MO, Zha L, Cabral-Miranda G, and Bachmann MF. Major findings and recent advances in virus–like particle (VLP)-based vaccines. Semin Immunol. (2017) 34:123–32. doi: 10.1016/j.smim.2017.08.014
62. Teasdale A, Elder D, and Nims R. ICH quality guidelines: An implementation guide. Hoboken, NJ: John Wiley & Sons (2017), ISBN: 978-1-118-97111-6.
63. Gruber MF and Marshall VB. Regulation and testing of vaccines. Plotkin’s Vaccines. (2018), 1547–1565.e2. doi: 10.1016/B978-0-323-35761-6.00079-1
64. Ehmann F, Kurz X, Cavaleri M, and Arlett P. Regulation of vaccines in europe. In: Plotkin’s Vaccines. Elsevier (2018). p. 1566–1572.e1. doi: 10.1016/B978-0-323-35761-6.00080-8
65. WHO. Guidelines on clinical evaluation of vaccines: regulatory expectations (2017). World Healt Organization Technical Report. Available online at: https://www.who.int/publications/m/item/WHO-TRS-1004-web-annex-9 (Accessed August 17, 2025).
66. Chen S, Yao L, Wang W, and Tang S. Developing an effective and sustainable national immunisation programme in China: issues and challenges. Lancet Public Health. (2022) 7:e1064–72. doi: 10.1016/S2468-2667(22)00171
67. Zhao XL, Hu SY, Hu JW, Wang HH, Wen TM, and Feng YS. Tackling barriers to scale up human papillomavirus vaccination in China: progress and the way forward. Infect Dis Poverty. (2023) 12:86. doi: 10.1186/s40249-023-01136-6
68. WHO. Malaria vaccines (RTS,S and R21) (2025). Available online at: https://www.who.int/news-room/questions-and-answers/item/q-a-on-rts-s-malaria-vaccine (Accessed October 11, 2025).
69. Suppi S, Michelson T, Viigand K, and Alamäe T. Repression vs. activation of MOX, FMD, MPP1 and MAL1 promoters by sugars in Hansenula polymorpha : the outcome depends on cell’s ability to phosphorylate sugar. FEMS Yeast Res. (2013) 13:219–32. doi: 10.1111/1567-1364.12023
70. Dai Z, Deng S, Culley DE, Bruno KS, and Magnuson JK. Agrobacterium tumefaciens-mediated transformation of oleaginous yeast Lipomyces species. Appl Microbiol Biotechnol. (2017) 101:6099–110. doi: 10.1007/s00253-017-8357-7
71. Nooraei S, Bahrulolum H, Hoseini ZS, Katalani C, Hajizade A, and Easton AJ. Virus-like particles: preparation, immunogenicity and their roles as nanovaccines and drug nanocarriers. J Nanobiotechnology. (2021) 19:59. doi: 10.1186/s12951-021-00806-7
72. Ardiani A, Higgins JP, and Hodge JW. Vaccines based on whole recombinant Saccharomyces cerevisiae cells. FEMS Yeast Res. (2010) 10:1060–9. doi: 10.1111/j.1567-1364.2010.00665.x
73. Lei H, Jin S, Karlsson E, Schultz-Cherry S, and Ye K. Yeast surface-displayed H5N1 avian influenza vaccines. J Immunol Res. (2016) 2016:1–12. doi: 10.1155/2016/4131324
74. Pollet J, Chen WH, Versteeg L, Keegan B, Zhan B, and Wei J. SARS-CoV-2 RBD219-N1C1: A yeast-expressed SARS-CoV-2 recombinant receptor-binding domain candidate vaccine stimulates virus neutralizing antibodies and T-cell immunity in mice. Hum Vaccin Immunother. (2021) 17:2356–66. doi: 10.1080/21645515.2021.1901545
75. Azevedo-Silva J, Amorim M, Tavares-Valente D, Sousa P, Mohamath R, and Voigt EA. Exploring yeast glucans for vaccine enhancement: Sustainable strategies for overcoming adjuvant challenges in a SARS-CoV-2 model. Eur J Pharm Biopharm. (2024) 205:114538. doi: 10.1016/j.ejpb.(2024).114538
76. Srivastava V, Nand KN, Ahmad A, and Kumar R. Yeast-based virus-like particles as an emerging platform for vaccine development and delivery. Vaccines (Basel). (2023) 11:479. doi: 10.3390/vaccines11020479
77. Austriaco N. Yeast oral vaccines against infectious diseases. Front Microbiol. (2023) 14:1150412. doi: 10.3389/fmicb.2023.1150412
78. Cao H, Gu H, Kang H, and Jia H. Development of a rapid reverse genetics system for feline coronavirus based on TAR cloning in yeast. Front Microbiol. (2023) 14:1141101. doi: 10.3389/fmicb.2023.1141101
79. Soutter F, Werling D, Nolan M, Küster T, Attree E, and Marugán-Hernández V. A novel whole yeast-based subunit oral vaccine against eimeria tenella in chickens. Front Immunol. (2022) 13:809711. doi: 10.3389/fimmu.2022.809711
80. Ma Y, Liu Z, Hao L, Wu J, Qin B, and Liang Z. Oral vaccination using Artemia coated with recombinant Saccharomyces cerevisiae expressing cyprinid herpesvirus-3 envelope antigen induces protective immunity in common carp (Cyprinus carpio var. Jian) larvae. Res Vet Sci. (2020) 130:184–92. doi: 10.1016/j.rvsc.2020.03.013
81. Liu Z, Wu J, Ma Y, Hao L, Liang Z, and Ma J. Protective immunity against CyHV-3 infection via different prime-boost vaccination regimens using CyHV-3 ORF131-based DNA/protein subunit vaccines in carp Cyprinus carpio var. Jian. Fish Shellfish Immunol. (2020) 98:342–53. doi: 10.1016/j.fsi.2020.01.034
82. Dong ZR, Mu QJ, Kong WG, Qin DC, Zhou Y, and Wang XY. Gut mucosal immune responses and protective efficacy of oral yeast Cyprinid herpesvirus 2 (CyHV-2) vaccine in Carassius auratus gibelio. Front Immunol. (2022) 13:932722. doi: 10.3389/fimmu.2022.932722
83. Gimenez AM, Francoso KS, Ersching J, Icimoto MY, Oliveira V, and Rodriguez AE. A recombinant multi-antigen vaccine formulation containing Babesia bovis merozoite surface antigens MSA-2a1, MSA-2b and MSA-2c elicits invasion-inhibitory antibodies and IFN-γ producing cells. Parasit Vectors. (2016) 9:577. doi: 10.1186/s13071-016-1862-1
84. Goh S, Kolakowski J, Holder A, Pfuhl M, Ngugi D, and Ballingall K. Development of a potential yeast-based vaccine platform for theileria parva infection in cattle. Front Immunol. (2021) 12:674484. doi: 10.3389/fimmu.2021.674484
85. Patterson R, Eley T, Browne C, Martineau HM, and Werling D. Oral application of freeze-dried yeast particles expressing the PCV2b Cap protein on their surface induce protection to subsequent PCV2b challenge in vivo. Vaccine. (2015) 33:6199–205. doi: 10.1016/j.vaccine.2015.10.003
86. Chen C, Hua D, Shi J, Tan Z, Zhu M, and Tan K. Porcine immunoglobulin fc fused P30/P54 protein of african swine fever virus displaying on surface of S. cerevisiae elicit strong antibody production in swine. Virol Sin. (2021) 36:207–19. doi: 10.1007/s12250-020-00278-3
87. Loukas A, Bethony JM, Mendez S, Fujiwara RT, Goud GN, and Ranjit N. Vaccination with recombinant aspartic hemoglobinase reduces parasite load and blood loss after hookworm infection in dogs. PLoS Med. (2005) 2:e295. doi: 10.1371/journal.pmed.0020295
88. Tizzano MA, Sguazza GH, Picotto LD, Echeverría MG, and Pecoraro MR. The cloning of the virus envelope glycoprotein F of canine distemper virus expressed in Pichia pastoris. Microb Pathog. (2020) 142:104094. doi: 10.1016/j.micpath.2020.104094
89. Willadsen P, Bird P, Cobon GS, and Hungerford J. Commercialisation of a recombinant vaccine against Boophilus microplus. Parasitology. (1995) 110:S43–50. doi: 10.1017/S0031182000001487
90. Suarez M, Rubi J, Pérez D, Cordova V, Salazar Y, and Vielma A. High impact and effectiveness of Gavac™ vaccine in the national program for control of bovine ticks Rhipicephalus microplus in Venezuela. Livest Sci. (2016) 187:48–52. doi: 10.1016/j.livsci.2016.02.005
91. Tabor AE. A review of Australian tick vaccine research. Vaccines (Basel). (2021) 9:1030. doi: 10.3390/vaccines9091030
92. Dellagostin OA, Felix SR, and Jorge S. Recombinant veterinary vaccines. Curr Dev Biotechnol Bioeng. (2017), 439–58. doi: 10.1016/B978-0-444-63660-7.00017-6
93. Aida V, Pliasas VC, Neasham PJ, North JF, McWhorter KL, and Glover SR. Novel vaccine technologies in veterinary medicine: A herald to human medicine vaccines. Front Vet Sci. (2021) 8:654289. doi: 10.3389/fvets.2021.654289
94. Rosano GL and Ceccarelli EA. Recombinant protein expression in Escherichia coli: advances and challenges. Front Microbiol. (2014) 5:172. doi: 10.3389/fmicb.2014.00172
95. Zhang ZX, Nong FT, Wang YZ, Yan CX, Gu Y, and Song P. Strategies for efficient production of recombinant proteins in Escherichia coli: alleviating the host burden and enhancing protein activity. Microb Cell Fact. (2022) 21:191. doi: 10.1186/s12934-022-01917-y
96. Walsh G. Post-translational modifications of protein biopharmaceuticals. Drug Discov Today. (2010) 15:773–80. doi: 10.1016/j.drudis.2010.06.009
97. Chambers AC, Aksular M, Graves LP, Irons SL, Possee RD, and King LA. Overview of the baculovirus expression system. Curr Protoc Protein Sci. (2018) 91:5–4. doi: 10.1002/cpps.47
98. Hong M, Li T, Xue W, Zhang S, Cui L, and Wang H. Genetic engineering of baculovirus-insect cell system to improve protein production. Front Bioeng Biotechnol. (2022) 10:994743. doi: 10.3389/fbioe.2022.994743
99. Trombetta CM, Marchi S, and Montomoli E. The baculovirus expression vector system: a modern technology for the future of influenza vaccine manufacturing. Expert Rev Vaccines. (2022) 21:1233–42. doi: 10.1080/14760584.2022.2085565
100. Laukens B, Visscher CD, and Callewaert N. Engineering yeast for producing human glycoproteins: where are we now? Future Microbiol. (2015) 10:21–34. doi: 10.2217/fmb.14.104
101. Garvey M. Non-mammalian eukaryotic expression systems yeast and fungi in the production of biologics. J Fungi. (2022) 8:1179. doi: 10.3390/jof8111179
102. de la Fuente J, Almazán C, Canales M, Pérez de la Lastra JM, Kocan KM, and Willadsen P. A ten-year review of commercial vaccine performance for control of tick infestations on cattle. Anim Health Res Rev. (2007) 8:23–8. doi: 10.1017/S1466252307001193
103. Vargas-Hernandez M, Santana-Rodríguez E, Montero-Espinosa C, Sordo-Puga Y, Acosta-Hernández A, and Fuentes-Rodríguez Y. Stability, safety and protective immunity of Gavac® vaccine subjected to heat stress. Biotecnol Apl. (2018) 35:35.
104. Martínez-Arzate SG, Sánchez-Bermúdez JC, Sotelo-Gómez S, Diaz-Albiter HM, Hegazy-Hassan W, and Tenorio-Borroto E. Genetic diversity of Bm86 sequences in Rhipicephalus (Boophilus) microplus ticks from Mexico: analysis of haplotype distribution patterns. BMC Genet. (2019) 20:56. doi: 10.1186/s12863-019-0754-8
105. de la Fuente J and Ghosh S. Evolution of tick vaccinology. Parasitology. (2024) 151:1045–52. doi: 10.1017/S003118202400043X
106. Ndawula C. From bench to field: A guide to formulating and evaluating anti-tick vaccines delving beyond efficacy to effectiveness. Vaccines (Basel). (2021) 9:1185. doi: 10.3390/vaccines9101185
107. Hamilton SR and Gerngross TU. Glycosylation engineering in yeast: the advent of fully humanized yeast. Curr Opin Biotechnol. (2007) 18:387–92. doi: 10.1016/j.copbio.2007.09.001
108. Ruiz LM, Orduz S, López ED, Guzmán F, Patarroyo ME, and Armengol G. Immune response in mice and cattle after immunization with a Boophilus microplus DNA vaccine containing bm86 gene. Vet Parasitol. (2007) 144:138–45. doi: 10.1016/j.vetpar.2006.09.033
109. Wildt S and Gerngross TU. The humanization of N-glycosylation pathways in yeast. Nat Rev Microbiol. (2005) 3:119–28. doi: 10.1038/nrmicro1087
110. Kim YH, Kang JY, Gil JY, Kim SY, Shin KK, and Kang HA. Abolishment of N-glycan mannosylphosphorylation in glyco-engineered Saccharomyces cerevisiae by double disruption of MNN4 and MNN14 genes. Appl Microbiol Biotechnol. (2017) 101:2979–89. doi: 10.1007/s00253-017-8101-3
111. Parsaie Nasab F, Aebi M, and Bernhard G. Frey AD. A combined system for engineering glycosylation efficiency and glycan structure in saccharomyces cerevisiae. Appl Environ Microbiol. (2013) 79:997–1007. doi: 10.1128/AEM.02817-12
112. Chen WH, Du L, Chag SM, Ma C, Tricoche N, and Tao X. Yeast-expressed recombinant protein of the receptor-binding domain in SARS-CoV spike protein with deglycosylated forms as a SARS vaccine candidate. Hum Vaccin Immunother. (2014) 10:648–58. doi: 10.4161/hv.27464
113. Dalvie NC, Leal J, Whittaker CA, Yang Y, Brady JR, and Love KR. Host-informed expression of CRISPR guide RNA for genomic engineering in komagataella phaffii. ACS Synth Biol. (2020) 9:26–35. doi: 10.1021/acssynbio.9b00372
114. Kim HJ and Kim HJ. Yeast as an expression system for producing virus-like particles: what factors do we need to consider? Lett Appl Microbiol. (2017) 64:111–23. doi: 10.1111/lam.12695
115. Zhang C, Zhang X, Zhang W, Dai W, Xie J, and Ye L. Enterovirus D68 virus-like particles expressed in Pichia pastoris potently induce neutralizing antibody responses and confer protection against lethal viral infection in mice. Emerg Microbes Infect. (2018) 7:1–22. doi: 10.1038/s41426-017-0005-x
116. Mani S, Tripathi L, Raut R, Tyagi P, Arora U, and Barman T. Pichia pastoris-Expressed Dengue 2 Envelope Forms Virus-Like Particles without Pre-Membrane Protein and Induces High Titer Neutralizing Antibodies. PLoS One. (2013) 8:e64595. doi: 10.1371/journal.pone.0064595
117. Zhou Y, Fan Y, LaPatra SE, Ma J, Xu J, and Meng Y. Protective immunity of a Pichia pastoris expressed recombinant iridovirus major capsid protein in the Chinese giant salamander, Andrias davidianus. Vaccine. (2015) 33:5662–9. doi: 10.1016/j.vaccine.2015.08.054
118. Gupta J, Kaul S, Srivastava A, Kaushik N, Ghosh S, and Sharma C. Expression, purification and characterization of the hepatitis E virus like-particles in the pichia pastoris. Front Microbiol. (2020) 11:141. doi: 10.3389/fmicb.2020.00141
119. Pijlman GP. Enveloped virus-like particles as vaccines against pathogenic arboviruses. Biotechnol J. (2015) 10:659–70. doi: 10.1002/biot.201400427
120. Sakuragi S, Goto T, Sano K, and Morikawa Y. HIV type 1 Gag virus-like particle budding from spheroplasts of Saccharomyces cerevisiae. Proc Natl Acad Sci. (2002) 99:7956–61. doi: 10.1073/pnas.082281199
121. Chandramouli S, Medina-Selby A, Coit D, Schaefer M, Spencer T, and Brito LA. Generation of a parvovirus B19 vaccine candidate. Vaccine. (2013) 31:3872–8. doi: 10.1016/j.vaccine.2013.06.062
122. Liu C, Yao Y, Yang X, Bai H, Huang W, and Xia Y. Production of recombinant human papillomavirus type 52 L1 protein in hansenula polymorpha formed virus-like particles. J Microbiol Biotechnol. (2015) 25:936–40. doi: 10.4014/jmb.1412.12027
123. Kouichi S, Ken-ichi T, Shin-ichiro N, Hiroshi M, and Kenichi M. Production of hepatitis B virion-like particles in yeast. Gene. (1991) 106:143–9. doi: 10.1016/0378-1119(91)90193-F
124. Vassileva A, Chugh DA, Swaminathan S, and Khanna N. Expression of hepatitis B surface antigen in the methylotrophic yeast Pichia pastoris using the GAP promoter. J Biotechnol. (2001) 88:21–35. doi: 10.1016/S0168-1656(01)00254-1
125. Vellanki RN, Komaravelli N, Tatineni R, and Mangamoori LN. Expression of hepatitis B surface antigen in Saccharomyces cerevisiae utilizing glyceraldeyhyde-3-phosphate dehydrogenase promoter of Pichia pastoris. Biotechnol Lett. (2007) 29:313–8. doi: 10.1007/s10529-006-9242-0
126. Rajamanickam V, Metzger K, Schmid C, and Spadiut O. A novel bi-directional promoter system allows tunable recombinant protein production in Pichia pastoris. Microb Cell Fact. (2017) 16:152. doi: 10.1186/s12934-017-0768-8
127. Türkanoğlu Özçelik A, Yılmaz S, and Inan M. Pichia pastoris promoters. Methods Mol Biol. (2019) 1923:97–112. doi: 10.1007/978-1-4939-9024-5_3
128. Gozar MMG, Price VL, and Kaslow DC. Saccharomyces cerevisiae -Secreted Fusion Proteins Pfs25 and Pfs28 Elicit Potent Plasmodium falciparum Transmission-Blocking Antibodies in Mice. Infect Immun. (1998) 66:59–64. doi: 10.1128/IAI.66.1.59-64.1998
129. Tam YJ, Allaudin ZN, Lila MAM, Bahaman AR, Tan JS, and Rezaei MA. Enhanced cell disruption strategy in the release of recombinant hepatitis B surface antigen from Pichia pastoris using response surface methodology. BMC Biotechnol. (2012) 12:70. doi: 10.1186/1472-6750-12-70
130. Gazor M, Ashraf Talesh SS, Hosseini SN, Javidanbardan A, and Khatami M. High recovery of intracellular recombinant HBsAg from Pichia pastoris via continuous pulsed laser cell disruption system optimized by response surface methodology. Biotechnol Appl Biochem. (2019) 66:91–100. doi: 10.1002/bab.1701
131. Hosseini SN, Ghaisari P, Sharifnia S, Khatami M, and Javidanbardan A. Improving the recovery of clarification process of recombinant hepatitis B surface antigen in large-scale by optimizing adsorption-desorption parameters on Aerosil-380. Prep Biochem Biotechnol. (2018) 48:490–7. doi: 10.1080/10826068.2018.1466153
132. Yousefipour M, Khatami M, Javidanbardan A, Hosseini SN, and Mehrnia M. Integration of size-exclusion chromatography and ultracentrifugation for purification of recombinant hepatitis B surface antigen: An alternative method for immunoaffinity chromatography. Prep Biochem Biotechnol. (2019) 49:158–66. doi: 10.1080/10826068.2018.1550658
133. Kimia Z, Hosseini SN, Ashraf Talesh SS, Khatami M, Kavianpour A, and Javidanbardan A. A novel application of ion exchange chromatography in recombinant hepatitis B vaccine downstream processing: Improving recombinant HBsAg homogeneity by removing associated aggregates. J Chromatogr B. (2019) 1113:20–9. doi: 10.1016/j.jchromb.2019.03.009
134. Moazami Goodarzi M, Jalalirad R, Doroud D, Hozouri H, and Aghasadeghi MR. DOE-based process optimization for development of efficient methods for purification of recombinant hepatitis B surface antigen from Pichia pastoris feedstock using Capto adhere resin. Heliyon. (2024) 10:e35124. doi: 10.1016/j.heliyon.2024.e35124
135. Mojarrad Moghanloo GM, Khatami M, Javidanbardan A, and Hosseini SN. Enhancing recovery of recombinant hepatitis B surface antigen in lab-scale and large-scale anion-exchange chromatography by optimizing the conductivity of buffers. Protein Expr Purif. (2018) 141:25–31. doi: 10.1016/j.pep.2017.08.011
136. Goodarzi MM, Jalalirad R, Doroud D, Hozouri H, Aghasadeghi M, and Paryan M. Determining buffer conditions for downstream processing of VLP-based recombinant hepatitis B surface antigen using multimodal resins in bind-elute and flow-through purification modes. Sci Rep. (2023) 13:10745. doi: 10.1038/s41598-023-37614-y
137. Hosseini SN, Javidanbardan A, Alizadeh Salim BS, and Khatami M. Large-scale purification of recombinant hepatitis B surface antigen from Pichia pastoris with non-affinity chromatographic methods as a substitute to immunoaffinity chromatography. Prep Biochem Biotechnol. (2018) 48:683–92. doi: 10.1080/10826068.2018.1487854
138. Hatami Giklou Jajan L, Hosseini SN, Abolhassani M, and Ghorbani M. Progress in affinity ligand-functionalized bacterial magnetosome nanoparticles for bio-immunomagnetic separation of HBsAg protein. PLoS One. (2022) 17:e0267206. doi: 10.1371/journal.pone.0267206
139. Zhao H, Wang Y, Ma Z, Wang Y, and Feng WH. Recombinant Kluyveromyces lactis expressing highly pathogenic porcine reproductive and respiratory syndrome virus GP5 elicits mucosal and cell-mediated immune responses in mice. J Vet Sci. (2014) 15:199. doi: 10.4142/jvs.2014.15.2.199
140. Gebauer M, Hürlimann HC, Behrens M, Wolff T, and Behrens SE. Subunit vaccines based on recombinant yeast protect against influenza A virus in a one-shot vaccination scheme. Vaccine. (2019) 37:5578–87. doi: 10.1016/j.vaccine.2019.07.094
141. Parthasarathi BC, Kumar B, Bhure SK, Sharma AK, Manisha, and Nagar G. Co-Immunization Efficacy of Recombinant Antigens against Rhipicephalus microplus and Hyalomma anatolicumTick Infestations. Pathogens. (2023) 12:433. doi: 10.3390/pathogens12030433
142. Luu VT, Moon HY, Hwang JY, Kang BK, and Kang HA. Development of recombinant Yarrowia lipolytica producing virus-like particles of a fish nervous necrosis virus. J Microbiol. (2017) 55:655–64. doi: 10.1007/s12275-017-7218-5
143. Sasagawa T, Tani M, Basha W, Rose RC, Tohda H, and Giga-Hama Y. A human papillomavirus type 16 vaccine by oral delivery of L1 protein. Virus Res. (2005) 110:81–90. doi: 10.1016/j.virusres.2005.02.001
144. Wu H, Williams K, Singh SR, Scissum-Gunn K, Singh NK, and Teresa D. Integrity of a recombinant hemagglutinin protein of an avian influenza virus. Biotechnol Lett. (2009) 31:1511–7. doi: 10.1007/s10529-009-0047-9
145. Bazan SB, Geginat G, Breinig T, Schmitt MJ, and Breinig F. Uptake of various yeast genera by antigen-presenting cells and influence of subcellular antigen localization on the activation of ovalbumin-specific CD8 T lymphocytes. Vaccine. (2011) 29:8165–73. doi: 10.1016/j.vaccine.2011.07.141
146. Bagherpour G, Ghasemi H, Zand B, Zarei N, Roohvand, and Ardakani EM. Oral administration of recombinant saccharomyces boulardii expressing ovalbumin-CPE fusion protein induces antibody response in mice. Front Microbiol. (2018) 9:723. doi: 10.3389/fmicb.2018.00723
147. Askri H, Kallèl H, Rourou S, Snoussi MA, and Lachheb J. Analytical methods for evaluating the immunogenicity of recombinant rabies virus glycoprotein expressed in the yeast komagataella phaffii. Immunol Invest. (2024) 53:1308–29. doi: 10.1080/08820139.2024.2399589
148. Yang M, Luo S, Zhou Q, Lu J, and Chen J. Immersion immunization with recombinant Saccharomyces cerevisiae displaying ORF25 induced protective immunity against cyprinid herpesvirus 2. J Fish Dis. (2024) 47:e13996. doi: 10.1111/jfd.13996
149. Liang J, Yang M, Li X, Zhou Q, Yang G, and Lu J. A vaccine combining ORF132 and ORF25 expressed by Saccharomyces cerevisiae induces protective immunity in Carassius auratus gibelio against CyHV-2. Fish Shellfish Immunol. (2025) 157:110099. doi: 10.1016/j.fsi.2024.110099
150. Majidi S, Aghaiypour Kolyani K, Akrami M, and Dadar M. An engineered SARS-coV-2 S1 glycoprotein produced in pichia pastoris as a candidate vaccine antigen. Mol Biotechnol. (2025). doi: 10.1007/s12033-025-01409-5
151. Hong Q, Wang S, Wang X, Han W, Chen T, and Liu Y. Vaccine potency and structure of yeast-produced polio type 2 stabilized virus-like particles. Vaccines (Basel). (2024) 12:1077. doi: 10.3390/vaccines12091077
152. Danzelle C, Cunha P, Noleto PG, Gilbert FB, Santos KR, and Staub C. Saccharomyces cerevisiae as a platform for vaccination against bovine mastitis. Vaccine. (2024) 42:126385. doi: 10.1016/j.vaccine.2024.126385
153. Bahrami H, Mosavat A, Soleimanpour S, Farsiani H, Valizadeh N, Rezaee SA, et al. Enhancing Mycobacterium tuberculosis-Ag85B immunogenicity by fusing with human Fcγ1 (Ag85B:hFcγ1). Microb Pathog. (2025) 207:107894. doi: 10.1016/j.micpath.2025.107894
154. Hu D, Li X, Duan X, Yang L, Luo B, Wang L, et al. Recombinant Saccharomyces cerevisiae EBY100/pYD1-FaeG: a candidate for an oral subunit vaccine against F4+ ETEC infection. Appl Environ Microbiol. (2025) 91:e01817–24. doi: 10.1128/aem.01817-24
155. Chairunnisa S, Mustopa AZ, Bela B, Arifah RK, Umami RN, and Firdaus MER. Expression, purification, and immunogenicity study of human papillomavirus type 52 virus-like particles produced in Hansenula polymorpha. Biologicals. (2025) 90:101831. doi: 10.1016/j.biologicals.2025.101831
156. Liang C, Geng L, Dong Y, and Zhang H. VEGF165b mutant can be used as a protein carrier to form a chimeric tumor vaccine with Mucin1 peptide to elicit an anti-tumor response. Mol Immunol. (2024) 175:31–9. doi: 10.1016/j.molimm.2024.09.009
157. FDA. VAXELIS (2023). Available online at: https://www.fda.gov/vaccines-blood-biologics/vaxelis (Accessed August 28, 2025).
158. FDA. TWINRIX (2023). Available online at: https://www.fda.gov/vaccines-blood-biologics/vaccines/twinrix (Accessed August 28, 2025).
159. FDA. Recombivax HB (2019). Available online at: https://www.fda.gov/vaccines-blood-biologics/vaccines/recombivax-hb (Accessed August 28, 2025).
160. FDA. ENGERIX-B (2023). Available online at: https://www.fda.gov/vaccines-blood-biologics/vaccines/engerix-b (Accessed August 28, 2025).
161. FDA. HEPLISAV-B (2024). Available online at: https://www.fda.gov/vaccines-blood-biologics/vaccines/heplisav-b (Accessed August 28, 2025).
162. FDA. GARDASIL (2025). Available online at: https://www.fda.gov/vaccines-blood-biologics/vaccines/gardasil (Accessed August 28, 2025).
163. FDA. GARDASIL 9 (2025). Available online at: https://www.fda.gov/vaccines-blood-biologics/vaccines/gardasil-9 (Accessed August 28, 2025).
164. EMA. Hexyon (2025). Available online at: https://www.ema.europa.eu/en/medicines/human/EPAR/hexyon (Accessed October 11, 2025).
165. EMA. Fendrix (2023). European Medicines Agency. Available online at: https://www.ema.europa.eu/en/medicines/human/EPAR/fendrix (Accessed October 11, 2025).
166. EMA. Hexacima (2025). European Medicines Agency. Available online at: http://ema.europa.eu/en/medicines/human/EPAR/hexacima (Accessed October 11, 2025).
167. EMA. HBVaxPro (2025). European Medicines Agency. Available online at: https://www.ema.europa.eu/en/medicines/human/EPAR/hbvaxpro (Accessed October 11, 2025).
168. EMA. Vaxelis (2024). European Medicines Agency. Available online at: https://www.ema.europa.eu/en/medicines/human/EPAR/vaxelis (Accessed October 11, 2025).
169. EMA. Mosquirix - opinion on medicine for use outside EU (2025). European Medicines Agency. Available online at: https://www.ema.europa.eu/en/opinion-medicine-use-outside-EU/human/mosquirix (Accessed October 11, 2025).
Keywords: recombinant protein expression systems, biopharmaceuticals, eukaryotic cell factories, immune responses, human and animal health
Citation: Ramos-Vega A, Monreal-Escalante E, Bañuelos-Hernández B, Angulo M, Trujillo E and Angulo C (2025) Yeast vaccine production platform for human and animal infectious diseases. Front. Immunol. 16:1697177. doi: 10.3389/fimmu.2025.1697177
Received: 01 September 2025; Accepted: 29 October 2025;
Published: 19 November 2025.
Edited by:
Evan Skowronski, TMG Biosciences, LLC, United StatesReviewed by:
Azam Bolhassani, Pasteur Institute of Iran (PII), IranTiziana Cervelli, National Research Council (CNR), Italy
Copyright © 2025 Ramos-Vega, Monreal-Escalante, Bañuelos-Hernández, Angulo, Trujillo and Angulo. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Abel Ramos-Vega, YXJhbW9zdmVAaXBuLm14; Carlos Angulo, ZWFuZ3Vsb0BjaWJub3IubXg=