 Anna Christine Trego1,2
Anna Christine Trego1,2 Evan Galvin1Conor Sweeney1Sinéad Dunning1Cillian Murphy1Simon Mills1Corine Nzeteu2Christopher Quince3
Evan Galvin1Conor Sweeney1Sinéad Dunning1Cillian Murphy1Simon Mills1Corine Nzeteu2Christopher Quince3 Stephanie Connelly4
Stephanie Connelly4 Umer Zeeshan Ijaz4*
Umer Zeeshan Ijaz4* Gavin Collins1,4,5*
Gavin Collins1,4,5*- 1Microbial Communities Laboratory, School of Natural Sciences, National University of Ireland Galway, Galway, Ireland
- 2Microbial Ecology Laboratory, School of Natural Sciences, National University of Ireland Galway, Galway, Ireland
- 3Warwick Medical School, University of Warwick, Warwick, United Kingdom
- 4Infrastructure and Environment, School of Engineering, University of Glasgow, Glasgow, United Kingdom
- 5Ryan Institute, National University of Ireland Galway, Galway, Ireland
Methanogenic sludge granules are densely packed, small, spherical biofilms found in anaerobic digesters used to treat industrial wastewaters, where they underpin efficient organic waste conversion and biogas production. Each granule theoretically houses representative microorganisms from all of the trophic groups implicated in the successive and interdependent reactions of the anaerobic digestion (AD) process. Information on exactly how methanogenic granules develop, and their eventual fate will be important for precision management of environmental biotechnologies. Granules from a full-scale bioreactor were size-separated into small (0.6–1 mm), medium (1–1.4 mm), and large (1.4–1.8 mm) size fractions. Twelve laboratory-scale bioreactors were operated using either small, medium, or large granules, or unfractionated sludge. After >50 days of operation, the granule size distribution in each of the small, medium, and large bioreactor sets had diversified beyond—to both bigger and smaller than—the size fraction used for inoculation. Interestingly, extra-small (XS; <0.6 mm) granules were observed, and retained in all of the bioreactors, suggesting the continuous nature of granulation, and/or the breakage of larger granules into XS bits. Moreover, evidence suggested that even granules with small diameters could break. “New” granules from each emerging size were analyzed by studying community structure based on high-throughput 16S rRNA gene sequencing. Methanobacterium, Aminobacterium, Propionibacteriaceae, and Desulfovibrio represented the majority of the community in new granules. H2-using, and not acetoclastic, methanogens appeared more important, and were associated with abundant syntrophic bacteria. Multivariate integration (MINT) analyses identified distinct discriminant taxa responsible for shaping the microbial communities in different-sized granules.
Introduction
Biofilms form in a wide range of natural and built environments (Sutherland, 2001), and have important significance for biogeochemical cycling in Nature (Battin et al., 2016; Flemming and Wuertz, 2019), and further clinical (Arciola et al., 2018) and industrial implications (Jensen et al., 2016). However, though biofilms are classically found as layers, or films, attached to suitable surfaces—from rocks, to medical devices, to ship hulls—aggregation may also occur due to self-immobilization of cells into discrete structures, such as flocs or granules, without the involvement of a surface (Lettinga et al., 1980; Togashi et al., 2014; Wilbanks et al., 2014). Many such examples can be found in engineered environments, such as in biological wastewater treatment, where prevailing conditions of shear, and hydrodynamic, stresses promote flocculation and granulation. Common types include anaerobic ammonium oxidizing (annamox) granules (Kartal et al., 2010), aerobic granules (Beun et al., 1999), and anaerobic (methanogenic) granules (Lettinga et al., 1980), which underpin the success of several high-rate anaerobic wastewater treatment technologies.
Anaerobic granules are small, with diameters generally ranging 0.1–5.0 mm (Ahn, 2000; Batstone and Keller, 2001; Shin et al., 2019), densely-packed biofilm spheres. They comprise a complex microbial community capable of the complete mineralization of organic pollutants through the anaerobic digestion (AD) pathway (Liu et al., 2002; Batstone et al., 2019). The settleabilty of anaerobic granules accounts for long biomass retention—even in “upflow” bioreactors, such as the upflow anaerobic sludge bed (UASB) and expanded granular sludge bed (EGSB) bioreactors, operated with short hydraulic retention times (HRT), and very high volumetric loading and upflow velocities (van Lier et al., 2015). The size distribution of anaerobic granules varies according to wastewater type (Batstone and Keller, 2001), but has also been linked to the hydrodynamics of the digester system (Arcand et al., 1994). Moreover, granule size has been linked to porosity (Wu et al., 2016) and permeability (Afridi et al., 2017)—having further implications for biofilm structure, mass transfer, gas diffusion, and activity (Bhunia and Ghangrekar, 2007; Jiang et al., 2016; Wu et al., 2016; Afridi et al., 2017).
A single granule contains a diverse and dynamic microbial community, capable of adapting to various changes in environmental conditions (McKeown et al., 2009; Cerrillo et al., 2016; Kuroda et al., 2016; Na et al., 2016; Zhu et al., 2017; Keating et al., 2018). Microbial cells are juxtaposed, and immobilized, within a complex matrix of extracellular polymeric substances (EPS) (MacLeod et al., 1995). Within these highly organized consortia, a collection of microbial trophic groups mediates a cascade of interdependent reactions resulting in complete degradation of complex organic wastewater pollutants (Zehnder and Brock, 1980; Dolfing, 1992; Batstone et al., 2019). Equally, the consortium’s species rely on efficient mass transfer of substrates and complex metabolic interdependencies (Embree et al., 2015).
Granulation is a process whereby suspended particles and planktonic cells accumulate, forming small dense biofilm aggregates (Liu et al., 2002; Show et al., 2020). Unlike conventional biofilm formation, which is a well-documented phenomenon (O’Toole et al., 2000; Hall-Stoodley et al., 2004), the specific mechanisms involved in anaerobic sludge granulation are still being teased-out (Sanjeevi et al., 2013; Kim et al., 2014; Gagliano et al., 2017, 2020; Sudmalis et al., 2018; Faria et al., 2019). The topic has been comprehensively reviewed, and the various theories summarized, which can be categorized as physical, microbial, or thermodynamic (Hulshoff Pol et al., 2004; Show et al., 2020). However, none has been solely accepted as a “unified theory on anaerobic granulation” (Liu et al., 2003). The primary consensus seems to be that the genus Methanothrix (Methanosaeta), a group of acetoclastic methanogens, specifically Methanothrix soehngenii (Tindall, 2014), are key organisms during the process (Hulshoff Pol et al., 2004; Show et al., 2020). These archaea can either (i) aggregate together, (ii) attach to suspended particles, or (iii) potentially form a bridge between existing microflocs—aiding in the critical first step of forming granule precursors (Dubourgier et al., 1987; Weigant, 1987; Jian and Shi-yi, 1993). In fact, recent studies still confirm the importance of Methanosaeta, even at high salinity (Gagliano et al., 2020).
Many studies have focused on granulation (Show et al., 2020), and associated dynamics of physico-chemical properties and microbial community structure. Fewer studies, however, have addressed the ultimate fate of granular biofilms. Moreover, several studies have reported the spontaneous disintegration of granular biofilms, ultimately leading to process failure (McHugh et al., 2006; Baloch et al., 2008; Kobayashi et al., 2015). Studies into development and fate of granules could help prevent or mitigate such phenomena. Therefore, the primary purpose of this study was to monitor granular growth; to determine whether granules grow and develop in a predictable way, from small to medium and, finally, to large. Díaz et al. (2006) proposed that small granules could be considered “young” and larger granules “old,” or more mature. However, the eventual fate of large, old granules remains unclear. This is the first instance, of which we are aware, when granules have been compartmentalized into size-resolved fractions (small, medium, and large), which were then used to separately start up bioreactors to investigate granule development and fate in such a way. Moreover, undisturbed sludge, providing a “meta-community” and full complement of size fractions, was used as a comparator. The extent, nature and ecology of “new” granules emerging in the experiments was monitored.
Materials and Methods
Source and Fractionation of Biomass
Anaerobic sludge was obtained from a full-scale (8256 m3), mesophilic (37°C), EGSB bioreactor in the Netherlands treating potato-processing wastewater. The full-scale bioreactor was operated at an upflow velocity of 1.2 m h–1 and an HRT of 6.86 h.
Granules were grouped (Figure 1) into five distinct size classifications: extra-small (XS; Ø, < 0.6 mm), small (S; Ø, 0.6–1.0 mm), medium (M; Ø, 1.0–1.4 mm), large (L; Ø, 1.4–1.8 mm), and extra-large (XL; Ø, > 1.8 mm). Granules were size-separated by passing the biomass through stainless steel sieves, separating specific size ranges. Triplicate samples from each size were stored at −20°C for subsequent DNA extractions. The remainder of the size-separated biomass was subsequently stored in phosphate-buffered saline (PBS) solution under an N2 headspace at 4°C prior to bioreactor inoculation.
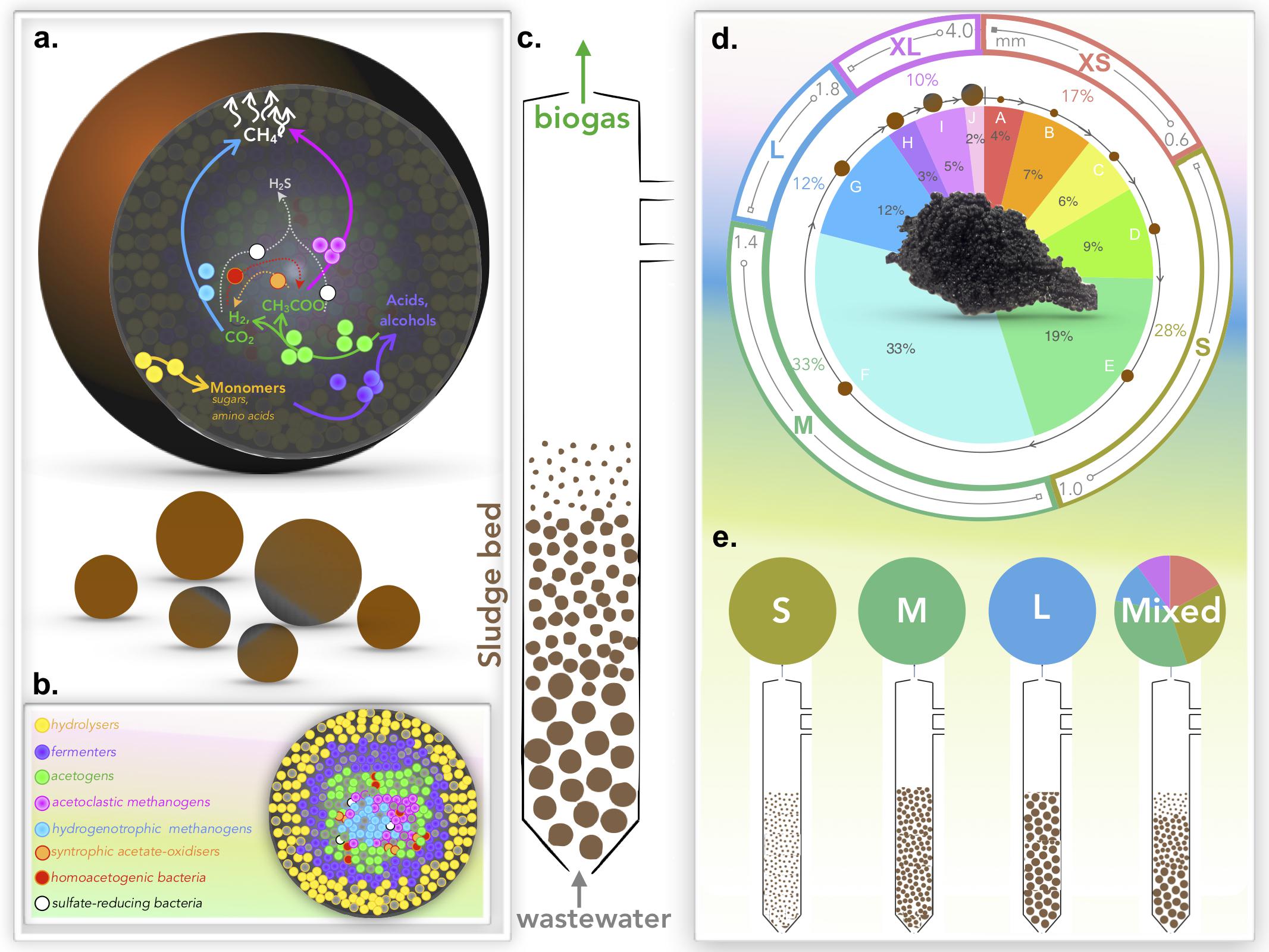
Figure 1. Schematics illustrating: (a) the AD pathway of organic matter degradation in the context of a granule; (b) theoretical distribution of the main trophic groups catalyzing the process; (c) the engineered bioreactor system used to apply granules for wastewater treatment and biogas generation; (d) size distribution of biomass whereby granules were binned for this study into five size groups: extra-small (XS), small (S), medium (M), large (L), and extra-large (XL); and (e) the experimental set-up used to test granular growth where bioreactors were inoculated with either S, M, L, or the naturally distributed (mixed) biomass.
Bioreactor Design and Operation
Twelve, identical laboratory-scale (2 L) glass, EGSB bioreactors (Supplementary Figure S1) were constructed, and operated in four sets of triplicates: the first set (RS1–RS3) containing only S granules; the second set (RM1–RM3) containing only M-sized granules; the third set (RL1–RL3) containing only L granules; and the fourth set (RN1–RN3) started with the unfractionated, naturally distributed (N) biomass (Figure 1).
Apart from granule size in the starter biomass, the 12 bioreactors, each inoculated with 15 gVS Lbioreactor–1, were operated identically at a 24 h HRT for 51 days. The biomass was allowed a 48-h acclimatization period at 37°C, regulated using built-in water jackets and recirculating water baths (Grant Optima, T100-ST12; Grant Instruments Ltd., Shepreth, United Kingdom), before feeding and recirculation were commenced, which were controlled using peristaltic pumps (Watson and Marlow 2058 and 300 series, respectively; Cork, Ireland). Influent was introduced at the base of each bioreactor at an OLR of 15.7 g COD L–1 day–1, and bioreactor liquor was recirculated through the system to achieve the superficial upflow velocity of 1.2 m/h according to the same set-up, and approach, as described previously (Collins et al., 2005; Madden et al., 2014).
The saccharide-rich, synthetic wastewater, based on recommendations and component ratios previously reported (Ahn and Forster, 2000), consisted of glucose (3.75 g/L), fructose (3.75 g/L), sucrose (3.56 g/L), yeast extract (1.45 g/L), and urea (2.15 g/L). The synthetic wastewater was further supplemented with trace elements (Shelton and Tiedje, 1984), and supplied to the 12 bioreactors from a single, thoroughly mixed reservoir to ensure homogeneity. Sodium bicarbonate (10 g/L) was added to the influent on day 6, and for the remainder of the experiment to act as a pH buffer, as the pH of the bioreactor liquor had dropped to 4 during the first week. This modification stabilized the influent pH at 7.8 for the remainder of the trial.
Upon take-down, on day 51, the biomass wet-weight was recorded, to determine total biomass loss or gain, and the entirety was re-fractionated into XS, S, M, L, and XL fractions to determine new size distribution. Samples from each size fraction were stored at -20°C for DNA extractions and sequencing.
Sampling and Analytical Techniques to Monitor Bioreactor Performance
Biogas concentrations of methane, and effluent concentrations of total COD (tCOD), soluble COD (sCOD), volatile fatty acids (VFA), and pH, were monitored three times a week throughout the 51-day trial. Biogas methane concentrations were determined using a VARIAN CP-3800 gas chromatograph (Varian, Inc., Walnut Creek, CA, United States; details available in Supplementary Material). Methane yield efficiency was calculated using the theoretical yield based on COD and reported as an average over three operational phases. pH was measured using a benchtop meter (Hanna Instruments, Woonsocket, RI, United States). COD was measured using pre-prepared COD test kits (Reagacon, Shannon, Ireland) and following the recommendation of the manufacturer. Samples for tCOD assays were each prepared by adding a homogenous sample directly to the test kit, while for sCOD, the sample was first centrifuged for 10 min at 14,000 r/min and the supernatant was added to the test kit. COD tests were incubated for 2 h at 150°C and concentrations were determined using a spectrophotometer (Hach Dr/4000; Hach Company, Loveland, CO, United States) at 435 nm. VFA contents of supernatant from effluent samples were separated, and quantified, using gas chromatography (Varian 450-GC; Varian, Inc., Walnut Creek, CA, United States; details available in Supplementary Material).
DNA Extraction
For each sample investigated, a mass of 0.1 g wet sludge was transferred to respective, sterile tubes in triplicate. Genomic DNA was extracted on ice following the DNA/RNA co-extraction method (Griffiths et al., 2000), which is based on bead beating in 5% (w/v) cetyl trimethylammonium bromide (CTAB) extraction buffer, followed by phenol-chloroform extraction. Quality of nucleic acids was assessed using a NanoDropTM spectrophotometer (Thermo Fisher Scientific, Waltham, MA, United States), and concentrations were determined using a Qubit fluorometer (Invitrogen, Carlsbad, CA, United States) and normalized to 5 ng DNA μL–1 for storage at -80°C.
High-Throughput Gene Sequencing
Partial 16S rRNA gene sequences were amplified using the universal bacterial and archaeal primers, 515F and 806R, and under to the conditions previously applied by Caporaso et al. (2011), but using 2x KAPA HiFi HotStart ReadyMix (Roche; Clarehill, Clare, Ireland). After clean-up using an AMPure XP purification kit (Beckman Coulter, Clare, Ireland), according to the manufacturer’s instructions, amplicons were sequenced, with PhiX (PhiX Control Kit v3) as internal control, on an Illumina MiSeq platform (at FISABIO, Valencia, Spain).
Bioinformatics and Statistical Analysis
Abundance tables were generated by constructing OTUs. An OTU table was generated for this study by matching the original barcoded reads against clean OTUs (a total of 2,793 OTUs for n = 49 samples) at 97% similarity (a proxy for species-level separation). Statistical analyses were performed in R (v. 3.4.4) using the combined data generated from the bioinformatics as well as meta data associated with the study. Alpha diversity analyses included the calculation of Shannon entropies and rarefied richness. Further multivariate integration (MINT)–sparse projection to latent structure (sPLS) algorithms identified study-wise discriminants. Additional details are available in Supplementary Material.
Results
Bioreactor Performance
Each of the four size-constrained sets of bioreactors responded similarly throughout the trial, regardless of being inoculated with differently sized granules. During the 4 days, influent pH decreased to 4.1 in each of the bioreactors. After supplementation of the influent with sodium bicarbonate, the pH stabilized (mean, pH 7.8) over the remainder of the experiment. Biogas methane concentrations were low during the initial acidification, but increased throughout the rest of the trial (Figure 2).
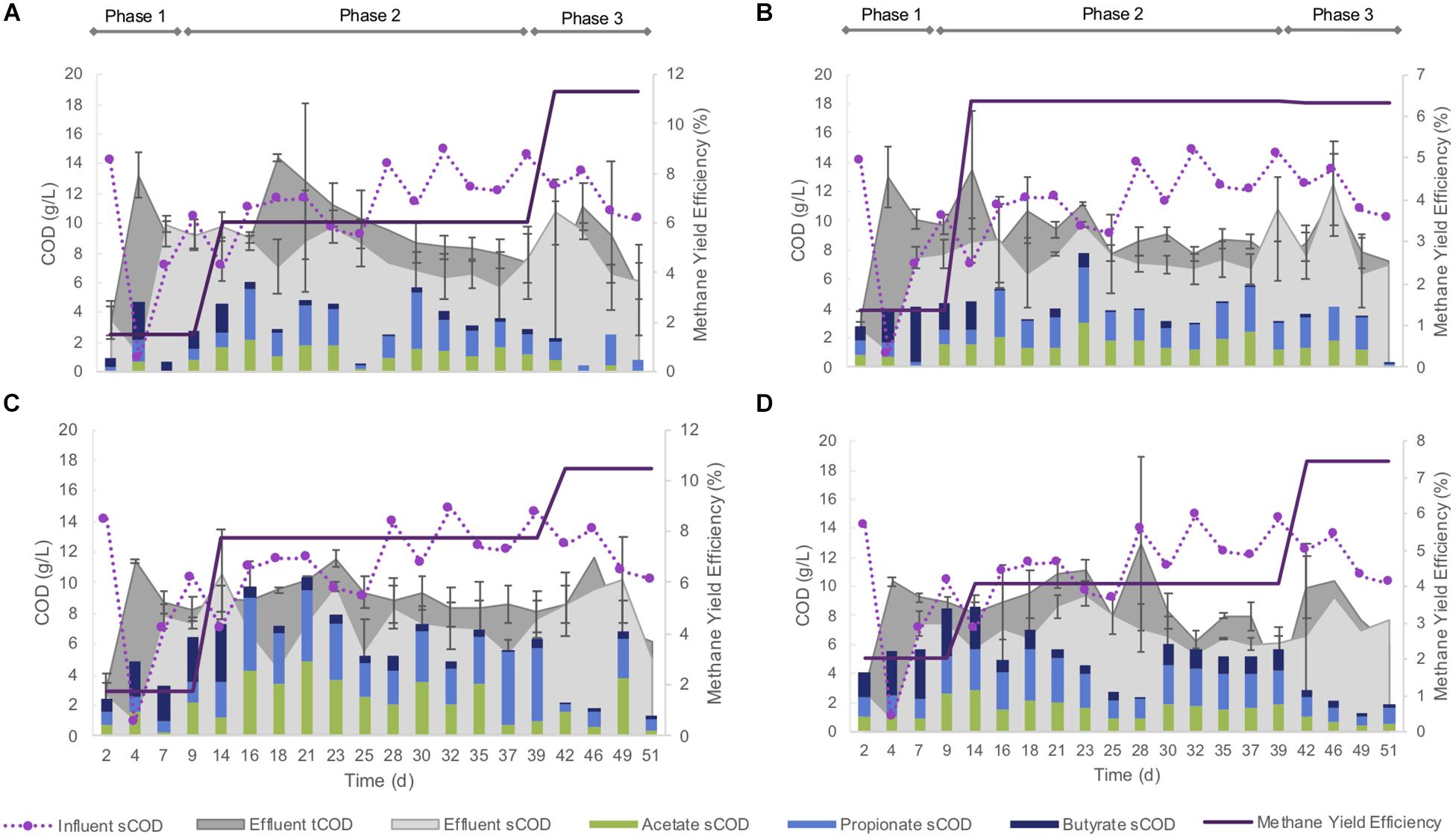
Figure 2. Methane yield efficiency; COD conversions (n = 3); and key VFA (acetate, propionate, and butyrate) contributions to effluent sCOD; in each of the four bioreactor sets: (A) RS1–RS3; (B) RM1–RM3; (C) RL1–RL3; (D) RN1–RN3.
This pH shock also produced a strong effect on the COD removal. During the first 25 days of operation, more COD left the bioreactors than was supplied to them (Figure 2)—indicating biomass washout and a very unstable performance. This was largely reversed over the remainder of the trial, and COD removal, and methane yield efficiency continued to improve over the subsequent weeks, culminating in roughly 50% sCOD removal efficiencies by each of the bioreactors. Nonetheless, COD removal was lower again during the final approximately 2 weeks of the trial (Figure 2). Acetate, propionate, and butyrate contributed to 50–90% of effluent sCOD (Figure 2).
Biomass washout was observed from each bioreactor variously over the course of the 51-day experiment (Supplementary Figure S2), including from the “naturally distributed” condition (RN1–3). Bioreactor RN2 failed—and was stopped—on day 22, due to the loss of 52% of the biomass. The remaining 11 bioreactors experienced losses reaching up to 50%. Washout of biomass was noted particularly during the initial few days during the initial acidification, and again at the end of the trial, evidenced by increased COD in the effluent. A net gain in biomass was observed in only two bioreactors, RL1 and RL3.
Shifts in Granule Size Distribution
Operation of laboratory-scale bioreactors, inoculated with size-constrained granules, allowed the emergence of “new” granule sizes, to be detected and studied. Size fractionation of biomass at the conclusion of the trial showed that the distribution of granule sizes had changed, and new granules—or “emerging sizes”—were apparent in all of the bioreactors (Figure 3). In all three of the RL bioreactors, and in two of the RM bioreactors, a full range of sizes (from the XS, S, M, L, XL classifications) had emerged (Figure 3). In the two surviving RN bioreactors, granules each of the five size classifications were still present, although the proportion of granules in M or above had increased. In fact, with only the exception of L granules in RS2 and RM2, and XL granules in RS2 and RS3, all five sizes emerged from all bioreactors (Figure 3).
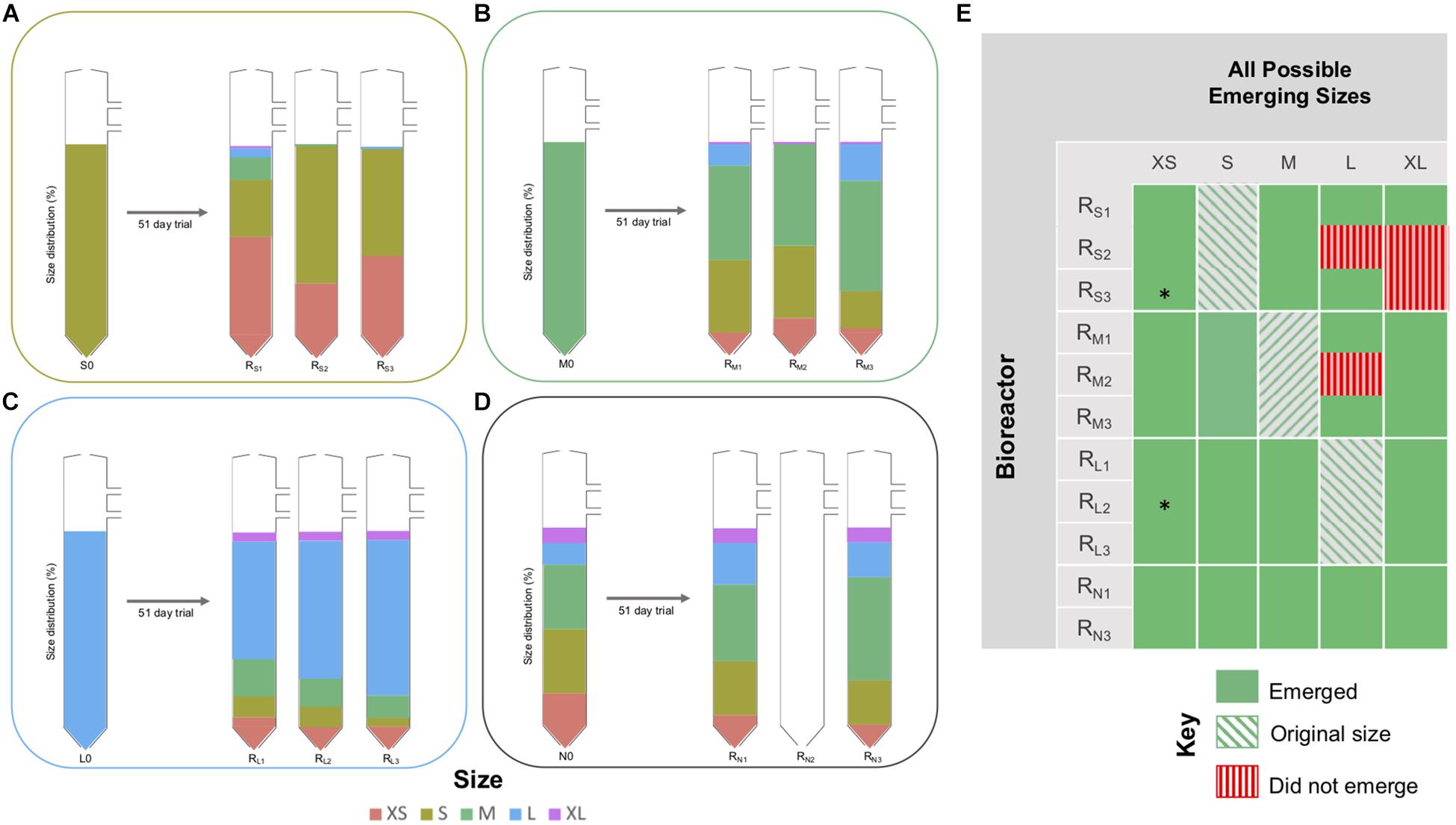
Figure 3. Changes in distribution of granule sizes in the RS, RM, RL, and RN bioreactors during the trial (day 0 and each of the respective bioreactors at day 51), showing: (A) RS1–RS3; (B) RM1–RM3; (C) RL1–RL3; (D) RN1 and RN3 bioreactors. Colors indicate the granule size classification and their proportion of the total biomass present. (E) Map indicating frequency of observations of emerging sizes across the experiment. No sequencing data available for samples marked with (*).
Microbial Community Structure of New Sizes
Alpha diversity measurements, using Shannon entropy, indicated similar trends for granules from the RM and RL bioreactors (Figure 4). A reduction in alpha diversity was apparent from S through to XL granules (i.e., there was more diversity in the microbial communities found in S granules than in bigger ones). Nonetheless, the alpha diversity in XS granules was significantly lower than in S granules. In fact, the diversity found in XS granules was similar to the diversity in XL granules (Figure 4). Size fractions emerging from RS, however, were statistically similar, and did not follow the same diversity trend.
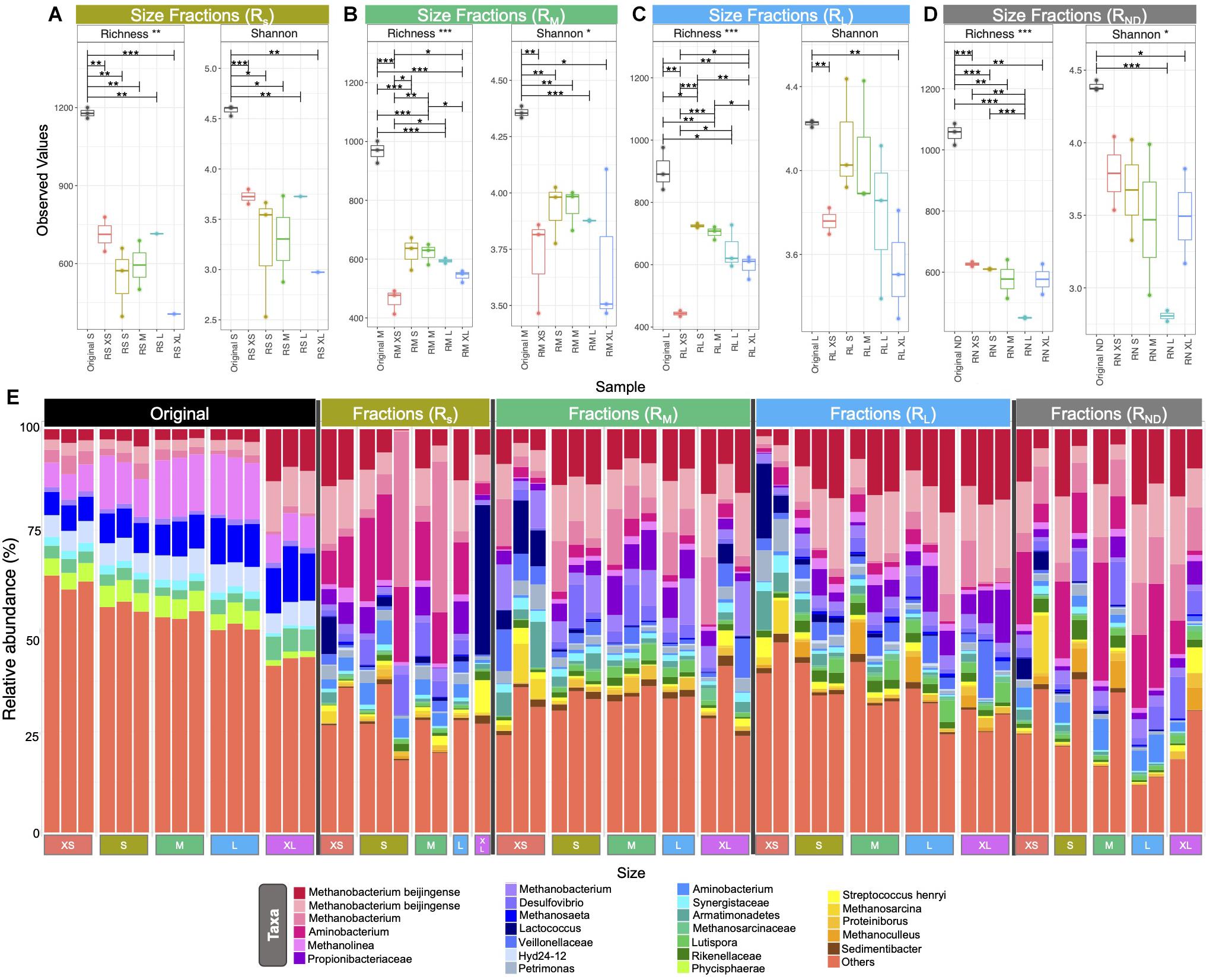
Figure 4. Box plots (A–D) of rarefied richness of the various size classifications from across the four bioreactor sets: (A) RS1–RS3; (B) RM1–RM3; (C) RL1–RL3; (D) RN1 and RN3; and bar chart (E) showing the top 25 relatively most abundant OTUs in original and new granules. Lines for figures (A–D) connect samples where differences were significant (ANOVA) indicated by *p < 0.05, **p < 0.01, ***p < 0.001.
The initial (day 0) community structure comprised of a mix of hydrogenotrophic (Methanobacterium, Methanolinea) and acetoclastic (Methanosaeta) methanogens (archaea). At the same time, the bacteria found to be relatively most abundant were generally all heterotrophic fermenters. Over the course of the trial, the make-up of the most abundant taxa shifted considerably. Across all of the new (or growing) granules—i.e., the emerging sizes from the bioreactors—the community structure was dominated by four operational taxonomic unit (OTU) classifications of Methanobacterium, in many cases accounting for 25–50% of the relative abundance of all taxa (Figure 4). Interestingly, Methanosaeta completely disappeared from among the 25 most abundant OTUs. Other highly abundant taxa included Aminobacterium, Propionibacteiraceae, and Desulfovibrio.
Multivariate integration algorithms used for study-wise discriminant analyses identified a total of 38 “discriminant” OTUs from 11 distinct phyla using two identified “components” (Supplementary Figure S3). Mean relative abundances of these OTUs showed two general groupings: (i) those OTUs more abundant in either, or both, of the XS and XL sized granules, and (ii) those OTUs which were more abundant in the S, M, and L granule sizes.
Discussion
Emerging Sizes: Granules Grow
This study demonstrates that methanogenic granules in anaerobic digesters do, indeed, “grow.” In each of the nine bioreactors started up with granules from a discrete size classification (Figure 1), the final distribution of granule sizes shifted to include new (or “grown”) granules that were either larger or smaller than the original granules (Figure 3), while also still containing granules of the original sizes. The emergence of larger granules almost certainly indicates the growth of granules due to cell replication and the accumulation of formerly planktonic cells from the surrounding environment. The observation of granules smaller than the original biomass might be explained in two ways: that (i) completely new granules formed from planktonic cells in the wastewater and the granulation process was continually initiated inside the digester, or (ii) parts of older, larger granules broke away and provided the foundation for new, small granules (Figure 5). The second explanation offers a potential mechanism of granule development. What is actually likely, we suggest, is that both phenomena proceed simultaneously.
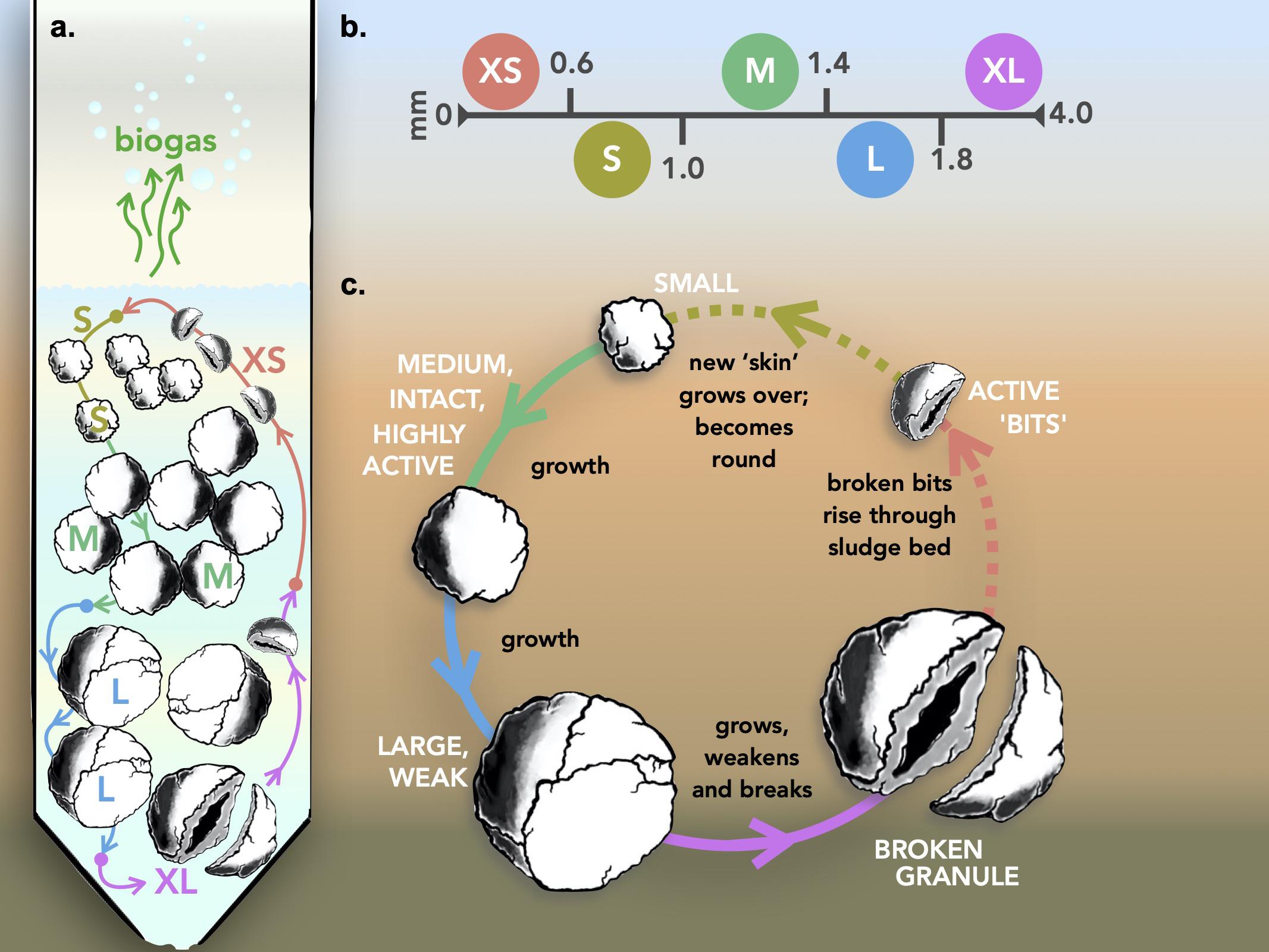
Figure 5. Granular growth and biofilm development model. (a) Operation of the model inside an anaerobic bioreactor; (b) size fraction parameters; and (c) the generalized growth model including the break-up of larger granules to form new, smaller granules.
An important component of the experiment was the set of bioreactors (RN) started up with a full complement of granule sizes, representing a “meta-community” of individual ecosystems (individual granules)—inspired in part by the recent description (Rillig et al., 2017) of soil aggregates as parallel incubators of evolution. In the RN bioreactors, the relative size distribution shifted toward larger granules. This may be due to granule growth. Conversely, it may be that small granules were preferentially lost from RN bioreactors, thus shifting the relative size distribution. However, smaller granules remained abundant in the other (RS, RM, or RL) bioreactors and many XS granules appeared to resist washout, ultimately suggesting that the shift in size distribution toward larger sizes was the result of growth.
Notably, the shift in size, and breaking of granules, while likely a natural process, was potentially further facilitated by the stressful conditions inside the bioreactors, similar to observations made regarding granule disintegration under high VFA conditions (McHugh et al., 2006). Moreover, we may also consider that the hydrodynamic conditions with respect to bioreactor scale (Connelly et al., 2017) and HRT (Arcand et al., 1994) may also have contributed to changes in the size distribution.
Emerging Pathways: Dominance of Hydrogenotrophic Methanogenesis
Early in the trial the reactors were subject to a severe drop in pH, which although not ideal, grants insights into the microbial community of anaerobic granules under stress. The predominant members of the microbiome across all of the samples from the end of the trial included Methanobacterium, Aminobacterium, Propionibacteriaceae, and Desulfovibrio species—none of which were dominant initially. The shift in community was likely a consequence of the pH stress early in the trial. Previous studies (Hori et al., 2006) found that low pH and increasing VFA concentrations in anaerobic digesters resulted in more abundant Methanosarcina (acetoclastic and hydrogenotrophic methanogens) and Methanothermobacter (hydrogenotrophic methanogens) but fewer Methanoculleus (also hydrogenotrophic), concluding that VFA accumulation strongly influences archaeal community structure.
This was also supported by our study. Methanosaeta (Methanothrix)—an acetoclastic methanogen, which was abundant in the granules on day 0—was not detected in the new granules, while Methanobacterium—autotrophic, H2-using methanogens (Jarrell et al., 1982; Pennings et al., 1998; Shlimon et al., 2004; Ma et al., 2005; Lennon and Jones, 2011) also capable of formate reduction (Schauer and Ferry, 1980)—were dominant and likely feeding on increased dissolved hydrogen resulting from the accumulating VFA (Cord-Ruwisch et al., 1997). Propionibacteriaceae—a family of heterotrophic glucose fermenters, producing propionate and acetate as primary products (Akasaka et al., 2003)—were also abundant in new granules, likely as VFA-producing acetogens. It is, of course, interesting to observe that granules grew in this experiment without the apparent dominant involvement of the filamentous Methanosaeta (Methanothrix), which tends to contradict the conventional understanding of granulation microbiology (Weigant, 1987; Hulshoff Pol et al., 2004). Other filamentous bacteria, however, were present such as Streptococcus, which was recently linked to granulation in high-salinity wastewaters (Gagliano et al., 2020).
Emerging Ecology: Supporting Syntrophic Relationships
The dominance of hydrogenotrophic methanogens (Supplementary Figure S3) by the end of the trial appeared to support the abundance of syntrophic bacteria, including Aminobacterium—heterotrophic fermenters of amino acids that grow well with methanogenic, H2-consuming partners, such as Methanobacterium (Baena et al., 1998; Hamdi et al., 2015)—and Desulfovibrio—sulfate-reducing bacteria (SRB) widespread in the environment (Goldstein et al., 2003), where they respire hydrogen or organic acids (Heidelberg et al., 2004) often in syntrophy with methanogens (Meyer et al., 2013). Interspecies metabolite exchange and hydrogen transfer (Stams and Plugge, 2009) between syntrophic partners is critical in AD because the oxidation of organic acids and alcohols by acetogens may be thermodynamically feasible only when hydrogenotrophic methanogens (in this case, likely the Methanobacterium) consume, and maintain sufficiently low concentrations of, H2. It is clear that the microbial community responded to the prevailing environmental stresses within the bioreactors. Indeed, had there not been an accumulation of VFA in the bioreactors and a striking dominance of the H2-oxidizing methanogens, a different community—perhaps characterized more strongly by the acetoclastic methanogens, may have developed.
Emerging Discriminants: Size-Specific OTUs
In general, the communities of all differently sized granules were very similar with some, though few, significant differences in alpha diversity and rarefied richness. Nonetheless, 32 study-wise discriminants could be identified, using MINT-sPLS analysis, which were responsible for minor community shifts across the different sizes from each bioreactor set. Phylogenetically, these discriminants formed two distinct clades—the first made up primarily of the phyla Firmicutes, Synergistetes, and Chloroflexi, and the second clade comprising of Proteobacteria, Spirochaetae, Bacteroidetes, and Euryarchaeota. Many of the discriminant OTUs were generally upregulated in the S, M, or L granules, or were upregulated in either or both XS and XL granules. For example, Lactococcus, a glucose fermenter and primary member of the lactic acid bacteria group, and Stenotrophomonas, a likely nitrate reducer, were both upregulated in XS and XL granules, but rare in S, M, and L granules. Conversely, other taxa, such as the Phycisphaerae, Leptospiraceae, and Bdellovibrio, were upregulated in the S, M, and L granules but infrequent in XS or XL granules.
Granular Growth and Biofilm Fate
Granulation and the subsequent growth of granules, resulting in various size distributions is not a new concept (Arcand et al., 1994; Batstone and Keller, 2001; Hulshoff Pol et al., 2004; Show et al., 2004), nor is it unique to anaerobic granules (Volcke et al., 2012; Zhou et al., 2016). Granular growth, which was clearly evidenced in our study, was also accompanied, and reported here for the first time, by the accumulation of XS granules in each of the initially size-constrained bioreactors, supporting the idea that granules break apart into smaller aggregates at some point during their “lifetime.” These smaller pieces did not appear to be selectively washed away, but were retained, making up a group of “new” smaller granules, which, in turn, will continue to grow. Such a concept is supported by previous evidence, that the larger the granule becomes, the more structurally unstable it is (Díaz et al., 2006), and that it eventually breaks apart. Our idea is that these broken bits, still containing an active microbial community eventually round off (due to shear forces within the digester) and become the basis for new, small granules, so that the process is cyclical (Figure 5).
To accept such a concept regarding granular growth, we would need to see that bioreactors initially containing only small granules, would eventually contain medium, then large and, finally, XL granules. An equivalent scenario would be observed for each bioreactor set. Equally, clear trends in microbial community structure might be observed across the different sizes. For example, an XL granule would have a similar community structure to an XS granule, but may be significantly different to an S or M granule.
Although the experimental design provided an interesting means to uncover the trajectory and fate of granules, each set of bioreactors was started with a different, size-limited, microbial consortium. Thus, granules grew from size-constrained consortia rather than a replete reservoir of granule sizes. Nonetheless, this study does provide evidence for “growing” granules and for the emergence and retention of very small granules, which are either the result of bigger aggregates breaking apart, or continued growth of de novo granules—but likely both. Granule growth was apparent in all nine of the RS, RM, and RL bioreactors. Indeed, most contained granules—albeit, sometimes very few—from each of the five size classifications used. Moreover, this study would suggest, based on emergence of XS granules in the RS bioreactors (Figure 3), that even small granules can break apart.
Conclusion
In summary, granules were demonstrated to be dynamic aggregates inside anaerobic digesters, appearing to follow a progressive growth pattern from small, to medium to large. XS granules emerged in all bioreactors, regardless of the starting size distribution. These either formed de novo, from the aggregation of free cells, or as a result of larger granules breaking apart. Further experiments should be done, under more stable bioreactor conditions, and with more intensive sampling regimes, to provide more evidence. The rate of biomass accumulation, as well as requirements to replace biomass, impinge on bioreactor performance and are important considerations in biomass management in anaerobic digesters. The results of experiments based on innovative approaches to track the fate of growing granules will provide valuable information to environmental engineers running bioreactors and to microbial ecologists studying community assembly phenomena, alike.
Author’s Note
This manuscript has been released as a preprint at BioRxiv (Trego et al., 2019).
Data Availability Statement
The sequencing data from this study are available on the European Nucleotide Archive under the study accession number PRJEB28212 (http://www.ebi.ac.uk/ena/data/view/PRJEB28212).
Author Contributions
AT, SC, UI, and GC designed the study. AT performed all of the physico-chemical characterization with assistance from CM, SM, EG, CS, SD, and CN. AT prepared the sequencing libraries. UI wrote the scripts for data analysis, which was conducted by AT. Results were interpreted by AT, CQ, UI, and GC. AT drafted the manuscript. UI and GC revised the document. All authors approved the manuscript and agreed for accountability of the work therein.
Funding
SC was supported by the Engineering and Physical Sciences Research Council, United Kingdom (EP/J00538X/1). CQ was funded by an MRC fellowship MR/M50161X/1 as part of the CLoud Infrastructure for Microbial Genomics (CLIMB) consortium MR/L015080/1. CM was supported by Erasmus and by the University of Turin and NUI Galway. UI was funded by NERC IRF NE/L011956/1. GC, SM, and AT were supported by a European Research Council Starting Grant (3C-BIOTECH 261330) and by a Science Foundation Ireland Career Development Award (17/CDA/4658) to GC. AT was further supported by a Thomas Crawford Hayes bursary from NUI Galway, and a Short-Term Scientific Mission grant through the EU COST Action 1302.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
The authors thank NVP Energy for providing anaerobic sludge granules.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2020.01126/full#supplementary-material
References
Afridi, Z. U. R., Wu, J., Cao, Z. P., Zhang, Z. L., Li, Z. H., Poncin, S., et al. (2017). Insight into mass transfer by convective diffusion in anaerobic granules to enhance biogas production. Biochem. Eng. J. 127, 154–160. doi: 10.1016/j.bej.2017.07.012
Ahn, J.-H., and Forster, C. F. (2000). Kinetic analyses of the operation of mesophilic and thermophilic anaerobic filters treating a simulated starch wastewater. Process Biochem. 36, 19–23. doi: 10.1016/S0032-9592(00)00166-7
Ahn, Y. (2000). Physicochemical and microbial aspects of anaerobic granular biopellets. J. Environ. Sci. Health A 35, 1617–1635. doi: 10.1080/10934520009377059
Akasaka, H., Ueki, A., Hanada, S., Kamagata, Y., and Ueki, K. (2003). Propionicimonas paludicola gen. nov., sp. nov., a novel facultatively anaerobic, Gram-positive, propionate-producing bacterium isolated from plant residue in irrigated rice-field soil. Int. J. Syst. Evol. Microbiol. 53, 1991–1998. doi: 10.1099/ijs.0.02764-0
Arcand, Y., Guiot, S. R., Desrochers, M., and Chavarie, C. (1994). Impact of the reactor hydrodynamics and organic loading on the size and activity of anaerobic granules. Chem. Eng. J. Biochem. Eng. J. 56, B23–B35. doi: 10.1016/0923-0467(94)87028-4
Arciola, C. R., Campoccia, D., and Montanaro, L. (2018). Implant infections: adhesion, biofilm formation and immune evasion. Nat. Rev. Microbiol. 16, 397–409. doi: 10.1038/s41579-018-0019-y
Baena, S., Fardeau, M. L., Labat, M., Ollivier, B., Thomas, P., Garcia, J. L., et al. (1998). Aminobacterium colombiensegen. nov. sp. nov., an amino acid-degrading anaerobe isolated from anaerobic sludge. Anaerobe 4, 241–250. doi: 10.1006/anae.1998.0170
Baloch, M. I., Akunna, J. C., Kierans, M., and Collier, P. J. (2008). Structural analysis of anaerobic granules in a phase separated reactor by electron microscopy. Bioresour. Technol. 99, 922–929. doi: 10.1016/j.biortech.2007.03.016
Batstone, D. J., Hülsen, T., and Oehmen, A. (2019). Metabolic modelling of mixed culture anaerobic microbial processes. Curr. Opin. Biotechnol. 57, 137–144. doi: 10.1016/j.copbio.2019.03.014
Batstone, D. J., and Keller, J. (2001). Variation of bulk properties of anaerobic granules with wastewater type. Water Res. 35, 1723–1729. doi: 10.1016/S0043-1354(00)00446-2
Battin, T. J., Besemer, K., Bengtsson, M. M., Romani, A. M., and Packmann, A. I. (2016). The ecology and biogeochemistry of stream biofilms. Nat. Rev. Microbiol. 14, 251–263. doi: 10.1038/nrmicro.2016.15
Beun, J. J., Hendriks, A., van Loosdrecht, M. C. M., Morgenroth, E., Wilderer, P. A., and Heijnen, J. J. (1999). Aerobic granulation in a sequencing batch reactor. Water Res. 33, 2283–2290. doi: 10.1016/S0043-1354(98)00463-1
Bhunia, P., and Ghangrekar, M. M. (2007). Required minimum granule size in UASB reactor and characteristics variation with size. Bioresour. Technol. 98, 994–999. doi: 10.1016/j.biortech.2006.04.019
Caporaso, J. G., Lauber, C. L., Walters, W. A., Berg-Lyons, D., Lozupone, C. A., Turnbaugh, P. J., et al. (2011). Global patterns of 16S rRNA diversity at a depth of millions of sequences per sample. Proc. Natl. Acad. Sci. U.S.A. 108, 4516–4522. doi: 10.1073/pnas.1000080107
Cerrillo, M., Morey, L., Viñas, M., and Bonmatí, A. (2016). Assessment of active methanogenic archaea in a methanol-fed upflow anaerobic sludge blanket reactor. Appl. Microbiol. Biotechnol. 100, 10137–10146. doi: 10.1007/s00253-016-7862-4
Collins, G., Foy, C., McHugh, S., and O’ Flaherty, V. (2005). Anaerobic treatment of 2,4,6-trichlorophenol in an expanded granular sludge bed-anaerobic filter (EGSB-AF) bioreactor at 15°C. FEMS Microbiol. Ecol. 53, 167–178. doi: 10.1016/j.femsec.2004.10.008
Connelly, S., Shin, S. G., Dillon, R. J., Ijaz, U. Z., Quince, C., Sloan, W. T., et al. (2017). Bioreactor scalability: laboratory-scale bioreactor design influences performance, ecology, and community physiology in expanded granular sludge bed bioreactors. Front. Microbiol. 8:664. doi: 10.3389/fmicb.2017.00664
Cord-Ruwisch, R., Mercz, T. I., Hoh, C. Y., and Strong, G. E. (1997). Dissolved hydrogen concentration as an on-line control parameter for the automated operation and optimization of anaerobic digesters. Biotechnol. Bioeng. 56, 626–634. doi: 10.1002/(sici)1097-0290(19971220)56:6<626::aid-bit5>3.0.co;2-p
Díaz, E. E., Stams, A. J. M., Amils, R., and Sanz, J. L. (2006). Phenotypic properties and microbial diversity of methanogenic granules from a full-scale upflow anaerobic sludge bed reactor treating brewery wastewater. Appl. Environ. Microbiol. 72, 4942–4949. doi: 10.1128/AEM.02985-05
Dolfing, J. (1992). The energetic consequences of hydrogen gradients in methanogenic ecosystems. FEMS Microbiol. Ecol. 101, 183–187. doi: 10.1111/j.1574-6968.1992.tb05774.x
Dubourgier, H., Prensier, G., and Albagnac, G. (1987). “Structure and microbial activities of granular anaerobic sludge,” in Granular Anaerobic Sludge: Microbiology and Technology, eds G. Lettinga, A. Zehnder, J. Grotenhuis, and L. Hulshoff Pol (Amsterdam: Elsevier), 18–33.
Embree, M., Liu, J. K., Al-Bassam, M. M., and Zengler, K. (2015). Networks of energetic and metabolic interactions define dynamics in microbial communities. Proc. Natl. Acad. Sci. U.S.A. 112, 15450–15455. doi: 10.1073/pnas.1506034112
Faria, C. V., Souza, D. F., Pontes, T. M., Amaral, M. C. S., and Fonseca, F. V. (2019). Strategies of anaerobic sludge granulation in an EGSB reactor. J. Environ. Manage. 244, 69–76. doi: 10.1016/j.jenvman.2019.05.015
Flemming, H.-C., and Wuertz, S. (2019). Bacteria and archaea on Earth and their abundance in biofilms. Nat. Rev. Microbiol. 17, 247–260. doi: 10.1038/s41579-019-0158-9
Gagliano, M. C., Ismail, S. B., Stams, A. J. M., Plugge, C. M., Temmink, H., and Van Lier, J. B. (2017). Biofilm formation and granule properties in anaerobic digestion at high salinity. Water Res. 121, 61–71. doi: 10.1016/j.watres.2017.05.016
Gagliano, M. C., Sudmalis, D., Pei, R., Temmink, H., and Plugge, C. M. (2020). Microbial community drivers in anaerobic granulation at high salinity. Front. Microbiol. 11:235. doi: 10.3389/fmicb.2020.00235
Goldstein, E. J. C., Citron, D. M., Peraino, V. A., and Cross, S. A. (2003). Desulfovibrio desulfuricans bacteremia and review of human Desulfovibrio infections. J. Clin. Microbiol. 41, 2752–2754. doi: 10.1128/JCM.41.6.2752-2754.2003
Griffiths, R. I., Whiteley, A. S., O’Donnell, A. G., and Bailey, M. J. (2000). Rapid Method for coextraction of DNA and RNA from natural environments for analysis of ribosomal DNA- and rRNA-based microbial community composition. Appl. Environ. Microbiol. 66, 5488–5491. doi: 10.1128/aem.66.12.5488-5491.2000
Hall-Stoodley, L., Costerton, J. W., and Stoodley, P. (2004). Bacterial biofilms: from the Natural environment to infectious diseases. Nat. Rev. Microbiol. 2:95. doi: 10.1038/nrmicro821
Hamdi, O., Ben Hania, W., Postec, A., Bouallagui, H., Hamdi, M., Bonin, P., et al. (2015). Aminobacterium thunnarium sp. nov., a mesophilic, amino acid-degrading bacterium isolated from an anaerobic sludge digester, pertaining to the phylum Synergistetes. Int. J. Syst. Evol. Microbiol. 65, 609–614. doi: 10.1099/ijs.0.068965-0
Heidelberg, J. F., Seshadri, R., Haveman, S. A., Hemme, C. L., Paulsen, I. T., Kolonay, J. F., et al. (2004). The genome sequence of the anaerobic, sulfate-reducing bacterium Desulfovibrio vulgaris hildenborough. Nat. Biotechnol. 22:554. doi: 10.1038/nbt959
Hori, T., Haruta, S., Ueno, Y., Ishii, M., and Igarashi, Y. (2006). Dynamic transition of a methanogenic population in response to the concentration of volatile fatty acids in a thermophilic anaerobic digester. Appl. Environ. Microbiol. 72, 1623–1630. doi: 10.1128/aem.72.2.1623-1630.2006
Hulshoff Pol, L. W., de Castro Lopes, S. I., Lettinga, G., and Lens, P. N. L. (2004). Anaerobic sludge granulation. Water Res. 38, 1376–1389. doi: 10.1016/j.watres.2003.12.002
Jarrell, K. F., Colvin, J. R., and Sprott, G. D. (1982). Spontaneous protoplast formation in Methanobacterium bryantii. J. Bacteriol. 149, 346–353. doi: 10.1128/jb.149.1.346-353.1982
Jensen, H., Biggs, C. A., and Karunakaran, E. (2016). The importance of sewer biofilms. Wiley Interdiscip. Rev. Water 3, 487–494. doi: 10.1002/wat2.1144
Jian, C., and Shi-yi, L. (1993). Study on mechanism of anaerobic sludge granulation in UASB reactors. Water Sci. Technol. 28, 171–178. doi: 10.2166/wst.1993.0159
Jiang, J., Wu, J., Zhang, Z., Poncin, S., Falk, V., and Li, H. Z. (2016). Crater formation on anaerobic granular sludge. Chem. Eng. J. 300, 423–428. doi: 10.1016/j.cej.2016.05.053
Kartal, B., Kuenen, J. G., and van Loosdrecht, M. C. M. (2010). Sewage treatment with anammox. Science 328, 702–703. doi: 10.1126/science.1185941
Keating, C., Hughes, D., Mahony, T., Cysneiros, D., Ijaz, U. Z., Smith, C. J., et al. (2018). Cold adaptation and replicable microbial community development during long-term low-temperature anaerobic digestion treatment of synthetic sewage. FEMS Microbiol. Ecol. 94:fiy095. doi: 10.1093/femsec/fiy095
Kim, D.-H., Lee, M.-K., Moon, C., Yun, Y.-M., Lee, W., Oh, S.-E., et al. (2014). Effect of hydraulic retention time on lactic acid production and granulation in an up-flow anaerobic sludge blanket reactor. Bioresour. Technol. 165, 158–161. doi: 10.1016/j.biortech.2014.03.097
Kobayashi, T., Xu, K.-Q., and Chiku, H. (2015). Release of extracellular polymeric substance and disintegration of anaerobic granular sludge under reduced sulfur compounds-rich conditions. Energies 8, 7968–7985. doi: 10.3390/en8087968
Kuroda, K., Nobu, M. K., Mei, R., Narihiro, T., Bocher, B. T. W., Yamaguchi, T., et al. (2016). A single-granule-level approach reveals ecological heterogeneity in an upflow anaerobic sludge blanket reactor. PLoS One 11:e0167788. doi: 10.1371/journal.pone.0167788
Lennon, J. T., and Jones, S. E. (2011). Microbial seed banks: the ecological and evolutionary implications of dormancy. Nat. Rev. Microbiol. 9:119. doi: 10.1038/nrmicro2504
Lettinga, G., van Velsen, A. F. M., Hobma, S. W., de Zeeuw, W., and Klapwijk, A. (1980). Use of the upflow sludge blanket (USB) reactor concept for biological wastewater treatment, especially for anaerobic treatment. Biotechnol. Bioeng. 22, 699–734. doi: 10.1002/bit.260220402
Liu, Y., Xu, H.-L., Show, K.-Y., and Tay, J.-H. (2002). Anaerobic granulation technology for wastewater treatment. World J. Microbiol. Biotechnol. 18, 99–113. doi: 10.1023/A:1014459006210
Liu, Y., Xu, H.-L., Yang, S.-F., and Tay, J.-H. (2003). Mechanisms and models for anaerobic granulation in upflow anaerobic sludge blanket reactor. Water Res. 37, 661–673. doi: 10.1016/S0043-1354(02)00351-2
Ma, K., Liu, X., and Dong, X. (2005). Methanobacterium beijingense sp. nov., a novel methanogen isolated from anaerobic digesters. Int. J. Syst. Evol. Microbiol. 55, 325–329. doi: 10.1099/ijs.0.63254-0
MacLeod, F. A., Guiot, S. R., and Costerton, J. W. (1995). Electron microscopic examination of the extracellular polymeric substances in anaerobic granular biofilms. World J. Microbiol. Biotechnol. 11, 481–485. doi: 10.1007/BF00286356
Madden, P., Al-Raei, A. M., Enright, A. M., Chinalia, F. A., de Beer, D., O’Flaherty, V., et al. (2014). Effect of sulfate on low-temperature anaerobic digestion. Front. Microbiol. 5:376. doi: 10.3389/fmicb.2014.00376
McHugh, S., Collins, G., and O’Flaherty, V. (2006). Long-term, high-rate anaerobic biological treatment of whey wastewaters at psychrophilic temperatures. Bioresour. Technol. 97, 1669–1678. doi: 10.1016/j.biortech.2005.07.020
McKeown, R. M., Scully, C., Enright, A. M., Chinalia, F. A., Lee, C., Mahony, T., et al. (2009). Psychrophilic methanogenic community development during long-term cultivation of anaerobic granular biofilms. ISME J. 3:1231. doi: 10.1038/ismej.2009.67
Meyer, B., Kuehl, J. V., Deutschbauer, A. M., Arkin, A. P., and Stahl, D. A. (2013). Flexibility of syntrophic enzyme systems in Desulfovibrio species ensures their adaptation capability to environmental changes. J. Bacteriol. 195, 4900–4914. doi: 10.1128/jb.00504-13
Na, J.-G., Lee, M.-K., Yun, Y.-M., Moon, C., Kim, M.-S., and Kim, D.-H. (2016). Microbial community analysis of anaerobic granules in phenol-degrading UASB by next generation sequencing. Biochem. Eng. J. 112, 241–248. doi: 10.1016/j.bej.2016.04.030
O’Toole, G., Kaplan, H. B., and Kolter, R. (2000). Biofilm formation as microbial development. Annu. Rev. Microbiol. 54, 49–79. doi: 10.1146/annurev.micro.54.1.49
Pennings, J. L. A., Keltjens, J. T., and Vogels, G. D. (1998). Isolation and characterization of Methanobacterium thermoautotrophicum ΔH mutants unable to grow under hydrogen-deprived conditions. J. Bacteriol. 180, 2676–2681. doi: 10.1128/jb.180.10.2676-2681.1998
Rillig, M. C., Muller, L. A. H., and Lehmann, A. (2017). Soil aggregates as massively concurrent evolutionary incubators. ISME J. 11:1943. doi: 10.1038/ismej.2017.56
Sanjeevi, R., Abbasi, T., and Abbasi, S. A. (2013). Role of Calcium (II) in Anaerobic Sludge Granulation and UASB Reactor Operation: A Method to Develop Calcium-Fortified Sludge Outside the UASB Reactors. New Delhi: NISCAIR-CSIR.
Schauer, N. L., and Ferry, J. G. (1980). Metabolism of formate in Methanobacterium formicicum. J. Bacteriol. 142, 800–807. doi: 10.1128/jb.142.3.800-807.1980
Shelton, D. R., and Tiedje, J. M. (1984). Isolation and partial characterization of bacteria in an anaerobic consortium that mineralizes 3-chlorobenzoic acid. Appl. Environ. Microbiol. 48, 840–848. doi: 10.1128/aem.48.4.840-848.1984
Shin, S. G., Ahn, Y., Park, C., Choi, Y.-K., Cho, H.-M., and Cho, S.-K. (2019). Size and morphological analyses of ultrasonicated hydrogen-producing granules using a simple method. Int. J. Hydrogen Energy 44, 2246–2252. doi: 10.1016/j.ijhydene.2018.07.032
Shlimon, A. G., Friedrich, M. W., Niemann, H., Ramsing, N. B., and Finster, K. (2004). Methanobacterium aarhusense sp. nov., a novel methanogen isolated from a marine sediment (Aarhus Bay, Denmark). Int. J. Syst. Evol. Microbiol. 54, 759–763. doi: 10.1099/ijs.0.02994-0
Show, K.-Y., Wang, Y., Foong, S.-F., and Tay, J.-H. (2004). Accelerated start-up and enhanced granulation in upflow anaerobic sludge blanket reactors. Water Res. 38, 2293–2304. doi: 10.1016/j.watres.2004.01.039
Show, K.-Y., Yan, Y., Yao, H., Guo, H., Li, T., Show, D.-Y., et al. (2020). Anaerobic granulation: a review of granulation hypotheses, bioreactor designs and emerging green applications. Bioresour. Technol. 300:122751. doi: 10.1016/j.biortech.2020.122751
Stams, A. J. M., and Plugge, C. M. (2009). Electron transfer in syntrophic communities of anaerobic bacteria and archaea. Nat. Rev. Microbiol. 7:568. doi: 10.1038/nrmicro2166
Sudmalis, D., Gagliano, M. C., Pei, R., Grolle, K., Plugge, C. M., Rijnaarts, H. H. M., et al. (2018). Fast anaerobic sludge granulation at elevated salinity. Water Res. 128, 293–303. doi: 10.1016/j.watres.2017.10.038
Sutherland, I. W. (2001). The biofilm matrix – an immobilized but dynamic microbial environment. Trends Microbiol. 9, 222–227. doi: 10.1016/S0966-842X(01)02012-1
Tindall, B. J. (2014). The genus name Methanothrix Huser et al. 1983 and the species combination Methanothrix soehngenii Huser et al. 1983 do not contravene Rule 31a and are not to be considered as rejected names, the genus name Methanosaeta Patel and Sprott 1990 refers to the same taxon as Methanothrix soehngenii Huser et al. 1983 and the species combination Methanothrix thermophila Kamagata et al. 1992 is rejected: supplementary information to Opinion 75. Judicial Commission of the International Committee on Systematics of Prokaryotes. Int. J. Syst. Evol. Microbiol. 64, 3597–3598. doi: 10.1099/ijs.0.069252-0
Togashi, T., Sasaki, H., and Yoshimura, J. (2014). A geometrical approach explains Lake Ball (Marimo) formations in the green alga, Aegagropila linnaei. Sci. Rep. 4:3761. doi: 10.1038/srep03761
Trego, A. C., Galvin, E., Sweeney, C., Dunning, S., Murphy, C., Mills, S., et al. (2019). De novo growth of methanogenic granules indicates a biofilm life-cycle with complex ecology. bioRxiv [preprint]. doi: 10.1101/667386
van Lier, J. B., van der Zee, F. P., Frijters, C. T. M. J., and Ersahin, M. E. (2015). Celebrating 40 years anaerobic sludge bed reactors for industrial wastewater treatment. Rev. Environ. Sci. Bio Technol. 14, 681–702. doi: 10.1007/s11157-015-9375-5
Volcke, E. I. P., Picioreanu, C., De Baets, B., and van Loosdrecht, M. C. M. (2012). The granule size distribution in an anammox-based granular sludge reactor affects the conversion—Implications for modeling. Biotechnol. Bioeng. 109, 1629–1636. doi: 10.1002/bit.24443
Weigant, W. (1987). “The ‘spagetti theory’ on anaerobic sludge formation, or the inevitability of granulation,” in Granular Anaerobic Sludge: Microbiology and Technology, eds G. Lettinga, A. Sehnder, J. Grotenhuis, and L. Hulshoff Pol (Netherlands: Pudoc. Wageningen), 146–152.
Wilbanks, E. G., Jaekel, U., Salman, V., Humphrey, P. T., Eisen, J. A., Facciotti, M. T., et al. (2014). Microscale sulfur cycling in the phototrophic pink berry consortia of the Sippewissett Salt Marsh. Environ. Microbiol. 16, 3398–3415. doi: 10.1111/1462-2920.12388
Wu, J., Afridi, Z. U. R., Cao, Z. P., Zhang, Z. L., Poncin, S., Li, H. Z., et al. (2016). Size effect of anaerobic granular sludge on biogas production: a micro scale study. Bioresour. Technol. 202, 165–171. doi: 10.1016/j.biortech.2015.12.006
Zehnder, A. J., and Brock, T. D. (1980). Anaerobic methane oxidation: occurrence and ecology. Appl. Environ. Microbiol. 39, 194–204. doi: 10.1128/aem.39.1.194-204.1980
Zhou, J., Zhang, Z., Zhao, H., Yu, H., Alvarez, P. J. J., Xu, X., et al. (2016). Optimizing granules size distribution for aerobic granular sludge stability: effect of a novel funnel-shaped internals on hydraulic shear stress. Bioresour. Technol. 216, 562–570. doi: 10.1016/j.biortech.2016.05.079
Keywords: anaerobic digestion, biofilms, methanogens, microbial communities, sludge granules, wastewater
Citation: Trego AC, Galvin E, Sweeney C, Dunning S, Murphy C, Mills S, Nzeteu C, Quince C, Connelly S, Ijaz UZ and Collins G (2020) Growth and Break-Up of Methanogenic Granules Suggests Mechanisms for Biofilm and Community Development. Front. Microbiol. 11:1126. doi: 10.3389/fmicb.2020.01126
Received: 28 January 2020; Accepted: 05 May 2020;
Published: 03 June 2020.
Edited by:
Hyung-Sool Lee, University of Waterloo, CanadaReviewed by:
Caroline M. Plugge, Wageningen University and Research, NetherlandsSeung Gu Shin, Pohang University of Science and Technology, South Korea
Changsoo Lee, Ulsan National Institute of Science and Technology, South Korea
Copyright © 2020 Trego, Galvin, Sweeney, Dunning, Murphy, Mills, Nzeteu, Quince, Connelly, Ijaz and Collins. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Umer Zeeshan Ijaz, dW1lci5pamF6QGdsYXNnb3cuYWMudWs=; Gavin Collins, Z2F2aW4uY29sbGluc0BudWlnYWx3YXkuaWU=