 Ana Hickey1,2
Ana Hickey1,2 Panagiota Stamou1
Panagiota Stamou1 Sreeram Udayan1,2
Sreeram Udayan1,2 Ana Ramón-Vázquez1
Ana Ramón-Vázquez1 Maria Esteban-Torres1,3
Maria Esteban-Torres1,3 Francesca Bottacini1,3
Francesca Bottacini1,3 Jerzy Adam Woznicki1
Jerzy Adam Woznicki1 Owen Hughes4
Owen Hughes4 Silvia Melgar1
Silvia Melgar1 Marco Ventura5
Marco Ventura5 Douwe Van Sinderen1,3
Douwe Van Sinderen1,3 Valerio Rossini1*†
Valerio Rossini1*† Ken Nally1,2*†
Ken Nally1,2*†- 1APC Microbiome Ireland, University College Cork, Cork, Ireland
- 2School of Biochemistry and Cell Biology, University College Cork, Cork, Ireland
- 3School of Microbiology, University College Cork, Cork, Ireland
- 4Luminex Corporation, Austin, TX, United States
- 5Life Sciences and Environmental Sustainability, University of Parma, Parma, Italy
Exopolysaccharide (EPS) is a bacterial extracellular carbohydrate moiety which has been associated with immunomodulatory activity and host protective effects of several gut commensal bacteria. Bifidobacterium breve are early colonizers of the human gastrointestinal tract (GIT) but the role of EPS in mediating their effects on the host has not been investigated for many strains. Here, we characterized EPS production by a panel of human B. breve isolates and investigated the effect of EPS status on host immune responses using human and murine cell culture-based assay systems. We report that B. breve EPS production is heterogenous across strains and that immune responses in human THP-1 monocytes are strain-specific, but not EPS status-specific. Using wild type and isogenic EPS deficient mutants of B. breve strains UCC2003 and JCM7017 we show that EPS had strain-specific divergent effects on cytokine responses from murine bone marrow derived macrophages (BMDMs) and dendritic cells (BMDCs). The B. breve UCC2003 EPS negative (EPS–) strain increased expression of cytokine genes (Tnfa, Il6, Il12a, and Il23a) relative to untreated BMDCs and BMDCs treated with wild type strain. B. breve UCC2003 and JCM7017 EPS– strains increased expression of dendritic cell (DC) activation and maturation marker genes (Cd80, Cd83, and Cd86) relative to untreated BMDCs. Consistent with this, BMDCs co-cultured with B. breve UCC2003 and JCM7017 EPS– strains engineered to express OVA antigen activated OVA-specific OT-II CD4+ T-cells in a co-culture antigen-presentation assay while EPS proficient strains did not. Collectively, these data indicate that B. breve EPS proficient strains use EPS to prevent maturation of DCs and activation of antigen specific CD4+ T cells responses to B. breve. This study identifies a new immunomodulatory role for B. breve EPS and suggests it may be important for immune evasion of adaptive immunity by B. breve and contribute to host-microbe mutualism.
Introduction
Bifidobacteria elicit protective immunomodulatory effects on murine and human hosts including reduction of gastrointestinal (GI) diseases and potentially systemic effects (Sivan et al., 2015; Sarkar and Mandal, 2016; Riedel, 2018). However, mechanisms underpinning these effects are still poorly understood (Fischbach, 2018; Round and Palm, 2018). Identification and characterization of specific bifidobacterial microbe-associated molecular patterns (MAMPs), their corresponding host pattern recognition receptors (PRRs) (Round et al., 2011) and associated immune cell responses is needed in order to understand proposed immunomodulatory properties of bifidobacteria. One of these candidate bifidobacterial MAMPs is EPS, a surface-associated glycan which is used by pathogens and other microbiota members to manipulate host responses (Castro-Bravo et al., 2018b). There is large structural diversity in EPS polymers produced by bifidobacteria, even between strains of the same species (Bottacini et al., 2014; Hidalgo-Cantabrana et al., 2014; Ferrario et al., 2016). The genes responsible for EPS biosynthesis are clustered within one or two transcriptional units. Variations in monosaccharide components and quantities of such, in addition to differences in glycosidic linkages and the degree of branching have been observed for different bifidobacteria strains (Leivers et al., 2011; Hidalgo-Cantabrana et al., 2014; Inturri et al., 2017; Castro-Bravo et al., 2018a). This variation could, in principle, lead to a vast number of distinct EPS structures and, theoretically to different immunomodulatory effects on the host. Reports indicate that purified EPS from two different bifidobacterial species can be detected by host innate immune PRRs – toll like receptor (TLR) 2 and TLR4 (Schiavi et al., 2018; Castro-Bravo et al., 2019). Bifidobacterial EPS has also been shown to ameliorate inflammation in murine pulmonary and intestinal models of inflammation (Hidalgo-Cantabrana et al., 2016; Schiavi et al., 2018). Previously, our group showed that EPS deficient B. breve UCC2003 had profound immunomodulatory effects on the host with an increase in pro-inflammatory cytokines interferon (IFN)-γ, tumor necrosis factor (TNF)-α and IL-12 detected from splenocytes cultured with the bacteria. These effects were associated with an increase in neutrophils, macrophages, natural killer cells, B cells and IFN-γ+, TNF-α+, and IL-12+ T cells (Fanning et al., 2012). Furthermore, the administration of EPS deficient B. breve to mice increased susceptibility to Citrobacter rodentium infection and reduced persistence of B. breve in the murine GIT (Fanning et al., 2012). Although many studies have used purified EPS to characterize host responses, this approach is susceptible to contamination with additional MAMPs derived from components of bacterial cell walls, membranes, or even cytoplasms (Alhudhud et al., 2018). An alternative reductionist approach is to investigate the effects of EPS using EPS deficient isogenic mutants of EPS producers and compare immune cell responses to both WT EPS proficient and mutant EPS deficient strains. EPS deficient mutants have been created for several bifidobacterial strains such as Bifidobacterium longum ssp. longum 35624 (Schiavi et al., 2016), B. longum 105-A (Tahoun et al., 2017), Bifidobacterium breve UCC2003 (Fanning et al., 2012), and B. breve JCM7017 (Alhudhud et al., 2018). Another consideration when investigating EPS-mediated effects is to use experimental cell-based systems from different host species as bifidobacteria are found and isolated from various ecological niches (Turroni et al., 2018). In this study, we analyzed a panel of 12 B. breve strains, which were isolated from humans, to assess EPS production. Two of these strains had matching isogenic EPS deficient mutant strains as previously described (B. breve UCC2003 and JCM7017) (Fanning et al., 2012; Alhudhud et al., 2018) and were used to determine the contribution of EPS to host immune cell responses. We found that B. breve EPS production was required to prevent maturation of dendritic cells and dendritic cell-mediated activation of CD4+ T cells responses.
Materials and Methods
Bacterial Culture Conditions and Plasmid Transformation
All bacterial strains used in this study are listed in Table 1. Strains were routinely cultured in reinforced clostridial medium (RCM; Oxoid cat. CM0149), sub-cultured into de Man Rogosa and Sharpe Medium (MRS; Becton Dickonson Difco cat. 288130) supplemented with 0.05% cysteine-HCl (Sigma) and incubated anaerobically at 37°C. Culture of bifidobacterial strains for growth curves was carried out in modified MRS (mMRS) medium, which was prepared from first principles (De Man et al., 1960) and supplemented with 0.05% cysteine-HCl (Sigma) and 1% glucose (Sigma). Cultures containing either B. breve UCC2003 EPS– or B. breve JCM7017 EPS– mutants were supplemented with 0.01 mg/mL tetracycline (Sigma) and cultures containing bacteria transformed with the pMG-mCherry-OVA plasmid were supplemented with 0.01 mg/mL chloramphenicol (Sigma). The pMG-mCherry-OVA plasmid was constructed from the pMG-mCherry plasmid as described by Grimm et al. (2014). Transformation of the pMG-mCherry-OVA plasmid into B. breve strains was achieved by electroporation with subsequent screening for chloramphenicol-resistant colonies on RCM agar plates containing 0.01 mg/mL chloramphenicol (Sigma). Further confirmation of mCherry-OVA expression was carried out by flow cytometry for mCherry expression (excitation at 545 ± 30 nm, emission at 610 ± 75 nm) using the BD FACSAria III Fusion Cell sorter, polymerase chain reaction and western blot. Flow cytometry data was further analyzed using FCS Express version 5 software (De Novo).
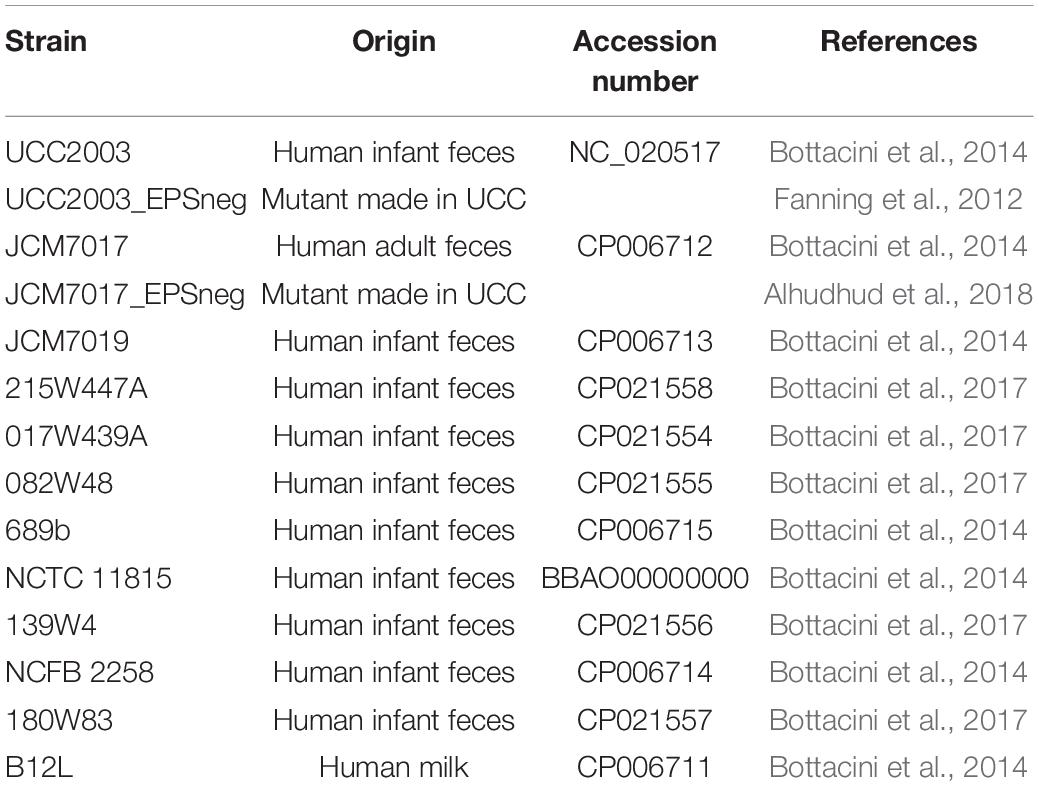
Table 1. List of B. breve strains used in study, their origin of isolation and genome accession number.
Electron Microscopy
Bacteria were prepared as described. Materials were directly adsorbed onto a carbon film membrane on a 300-mesh copper grid, stained with 1% uranyl acetate, dissolved in distilled water, and dried at room temperature. Grids were examined with Hitachi HT7700 electron microscope operated at 80 kV (Elexience – France), and images were acquired with a charge-coupled device camera (AMT). This work was carried out at the facilities and expertise of MIMA2 MET – GABI, INRA, Agroparistech (78352 Jouy-en-Josas, France).
In silico and in vitro Analysis of EPS Production
In silico analysis of B. breve fully sequenced strains was carried out using comparative genome analysis according to a previously described method (Bottacini et al., 2018). Whole genome sequences of 12 B. breve strains were retrieved from the National Centre for Biotechnology Information (NCBI) database. The genomic region previously described as associated to EPS biosynthesis in B. breve UCC2003 (Fanning et al., 2012; Bottacini et al., 2017), encoded between a priming glycosyl transferase (Bbr_0430) and a chain length determinant (Bbr_0474) was used as starting point for predicting homologous EPS-encoding regions in the 12 B. breve strains here analyzed. Presence/absence of homologous genes was retrieved by comparative genome analysis using “all-against-all,” bi-directional BLASTP alignments49 (cut-off: E-value < 0.0001, with at least 50% identity across at least 50% of either protein sequence). Based on previous descriptions for the B. breve spp., each strain was classified as an EPS “producer” or “non-producer” based on the presence of: (a) an EPS locus larger than 30 Kb, (b) at least three glycosyltransferase (GTF)-encoding genes within the predicted EPS locus, (c) a priming glycosyltransferase, (d) a gene encoding a flippase, (e) a chain length determinant. EPS status was assessed in vitro by a sedimentation assay as previously described (Fanning et al., 2012; Tahoun et al., 2017). Briefly, optical density measurements (OD600 nm) of B. breve strains and EPS– mutants were taken over a 6 h time period grown in mMRS medium, which was prepared from first principles (De Man et al., 1960) and supplemented with 0.05% cysteine-HCl and 1% glucose, lactose or maltose (Sigma) without agitation. A drop in OD values due to sedimentation is a characteristic of EPS non-producers.
Mice
Mice used for this study were 8–12 weeks old from a C57BL/6 background. WT C57BL/6 mice for isolation of bone marrow and generation of BMDMs and BMDCs were purchased from Envigo (Oxfordshire, United Kingdom). OT-II mice were acquired from Jackson Laboratories (Bar Harbor, United States) and breeding was maintained in house. All mice were housed in UCC Biological Services Unit under specific pathogen free (SPF) conditions. Standard housing conditions were maintained with 12 h darkness and 12 h light, temperature controlled at 21°C and 50% humidity. Animals were fed standard chow pellets and water was given ad libitum. The animal work was performed in accordance with EU legislation, in accordance with EU Directive 2010/63/EU, for the protection of animals used for scientific purposes and approved by the Animal Experimentation Ethics Committee of University College Cork, Euthanasia Only Application ID 2018/007.
THP-1 Human Monocytic NF-κB Reporter Cells
THP1-XBlueTM and THP1-XBlueTM-defMyD (MyD88 deficient) NF-κB reporter cell lines were purchased from Invivogen. Cells were routinely cultured using Roswell Park Memorial Institute medium (RPMI 1640; Sigma cat. R8758) supplemented with 10% Fetal Bovine Serum (FBS; Sigma), 1% Penicillin-Streptomycin (P/S; Sigma), 100 μg/mL Normocin (Invivogen) and 200 μg/mL Zeocin (Invivogen). For the THP1-XBlueTM-defMyD cells 100 μg/mL Hygromycin B Gold (Invivogen) was added to the medium. Cells were continuously maintained at a density between 7 × 105 cells/mL and 2 × 106 cells/mL. NF-κB activity in THP-1 reporter cell lines was assayed using Quanti-blue (Invivogen cat. rep-qb1), as per manufacturer’s instructions.
GM-CSF-Derived Bone Marrow Derived Macrophages and Dendritic Cells
Bone marrow isolation was carried out using aseptic techniques. This method was based on the work of Helft et al. (2015). Briefly, bone marrow was isolated from the femurs and tibias of selected mice by flushing using RPMI and filtered through a 70 μm strainer. Cells were collected by centrifugation at 200 × g at 4°C for 10 min. The cell pellet was resuspended in differentiation media – RPMI containing 10% FBS, 1% P/S and 20 ng/mL GM-CSF (PeproTech cat. 315-03). Cells were counted and 107 cells were seeded per well in 6 well plates with each well containing 4 mL of differentiation media. On day 2 half of the content was collected from each well, centrifuged and cells were resuspended in RPMI containing 10% FBS, 1% P/S and 40 ng/mL GM-CSF and added back to the 6 well plates. On day 3, all content from each well was harvested, centrifuged and cells were resuspended in RPMI containing 10% FBS, 1% P/S and 20 ng/mL GM-CSF before being added back to the 6 well plates. Both suspension cells (containing the DC fraction) and adherent cells (containing the macrophage fraction) were collected on day 6 and prepared for sorting. Prior to collection, adherent cells were harvested using Stempro Accutase (Thermo Scientific). Cells were stained using anti-mouse CD11c Alexa Fluor 700 (BioLegend cat. 117320), anti-mouse I-A/I-E fluorescein isothiocyanate (BioLegend cat. 107606), anti-mouse CD11b R-phycoerythrin cyanine 7 (BioLegend cat. 101216) and anti-mouse CD115 Allophycocyanin (BioLegend cat. 135510). Cells were then analyzed using the BD FACSAriaIII Fusion Cell sorter and sorted into pure populations of BMDMs and BMDCs based on a gating strategy modified from Helft et al. (2015). Data was further analyzed using FCS Express version 5 software (De Novo).
Co-culture of Mammalian Cells and Bifidobacteria
For all experiments, after an overnight culture of bacteria an OD600 nm of 1 corresponded to 109 CFU of bacteria. Bacteria were washed twice in sterile PBS and resuspended in the appropriate mammalian cell culture medium with no antibiotics and concentration was adjusted as required. For cytokine and NF-κB activity experiments, BMDMs, BMDCs and THP-1 monocytes were seeded in 96 well plates at a density of 5 × 104 cells/well in RPMI with 10% FBS and incubated with bacteria at a multiplicity of infection of 10:1 bacteria:cells. For BMDM and BMDCs experiments 100 ng/mL of LPS-B5 (Invivogen cat. tlrl-b5lps) and 5 μM of ODN 1585 CpG (Invivogen cat. tlrl-1585) were added as a positive controls, respectively. Co-cultures were incubated for 24 h at 37°C in 5% CO2. After 24 h, cells were collected by centrifugation and supernatants analyzed for NF-κB activity (THP-1 cells only). For reverse transcription-quantitative polymerase chain reaction (RT-qPCR) BMDMs or BMDCs were seeded at 2.5 × 105 cells were seeded per well in 12 well plates in RPMI with 10% FBS and incubated with bacteria at a multiplicity of infection of 10:1. 100 ng/mL of LPS-B5 (Invivogen cat. tlrl-b5lps) was used as a positive control in BMDM cultures and 5 μM of ODN 1585 CpG (Invivogen cat. tlrl-1585) was used as a positive control in BMDCs cultures. Co-cultures were incubated for 4 or 8 h at 37°C in 5% CO2. After 4 or 8 h, BMDMs were washed with ice cold PBS once and lysed using RLT lysis buffer (Qiagen). BMDCs were transferred to 1.5 mL tubes and centrifuged for 7 min at 400 × g at 4°C. Supernatants were removed and cells lysed in RLT lysis buffer. Both BMDM and BMDs lysates were immediately stored at −80°C until total RNA was extracted.
Cytokine ELISAs
BMDM and BMDCs co-culture supernatants were analyzed for secreted cytokines TNF-α and IL-10 using mouse TNF-alpha DuoSet ELISA (R&D Systems cat. DY410) and mouse IL-10 DuoSet ELISA (R&D Systems cat. DY417) as per manufacturer’s instructions. Supernatants from T cell and BMDM or BMDCs co-cultures were analyzed for secreted cytokines TNF-α and IL-2 using mouse TNF-alpha DuoSet ELISA (R&D Systems cat. DY410) and mouse IL-2 DuoSet ELISA (R&D Systems cat. DY402) as per manufacturer’s instructions. TNF-α production by THP-1 reporter cell lines was measured in supernatants from these experiments using human TNF-α ELISA Duoset (R&D Systems cat. DY210).
RT-qPCR
RNA was isolated from BMDMs and BMDCs using RNeasy Micro Kit (Qiagen cat. 74004) which included DNase treatment. Total RNA was quantified using Qubit RNA high sensitivity kit (Biosciences cat. Q32852). Complementary DNA (cDNA) was synthesized from 100 ng of total RNA using Transcriptor Reverse Transcriptase (Roche) and random hexamer primers (Roche) as per manufacturer’s instructions. qPCR assays were designed using the Roche Universal Probe Library Assay Design Centre (Table 2). qPCR assays were carried out by using SensiFAST Probe No-ROX Kit (Bioline), 500 nM primers, 250 nM corresponding ProbeLibrary probe from Universal ProbeLibrary (Roche) and 2 ng of cDNA. PCR reactions were run on a LightCycler 480 instrument (Roche) and the 2–ΔΔCT method (Mar et al., 2009; McCall et al., 2014) was used to calculate relative gene changes in gene expression compared to β-actin which served as a housekeeping gene (Livak and Schmittgen, 2001).
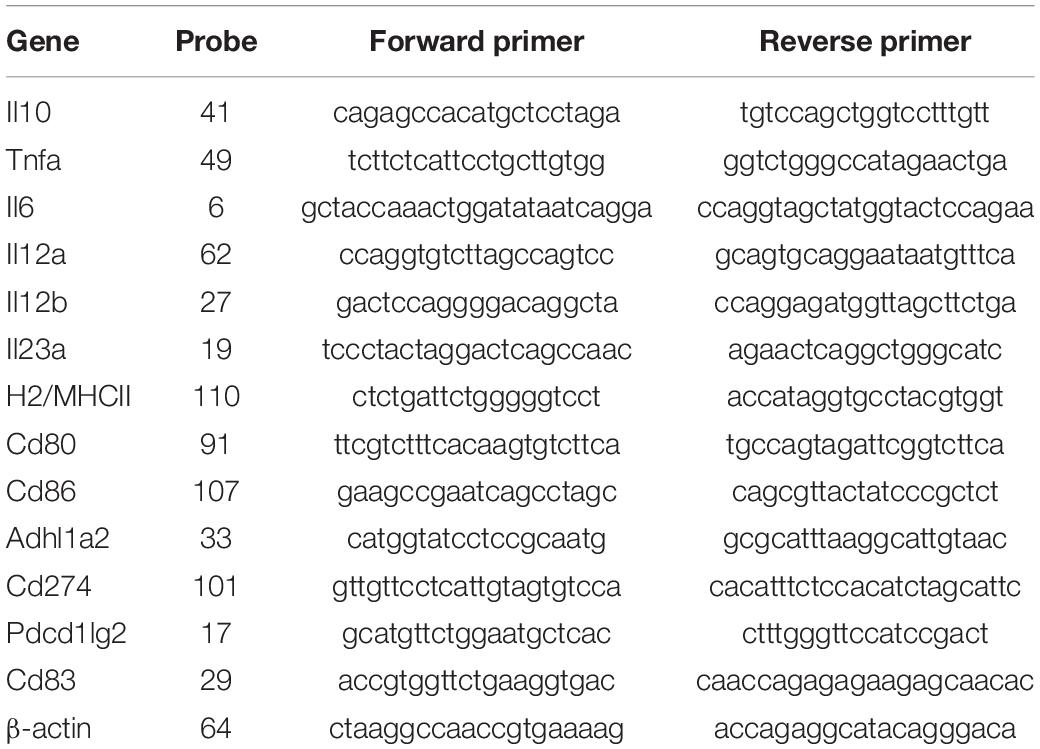
Table 2. List of genes, Universal ProbeLibrary (UPL) probes and primers used for RT-qPCR.
Western Blotting
After an overnight culture, bacterial cells were resuspended in ice-cold 10 mM Tris-HCl pH 8, transferred to tubes containing glass microbeads and bead-beated three times. Lysates were centrifuged at maximum speed for 15 min at 4°C and supernatants were transferred to fresh 1.5 mL tubes. Protein was quantified using Pierce BCA Protein Assay Kit (Thermo Scientific) as per manufacturer’s instructions. Fifty μg of protein for each sample was standardized and the appropriate volume of 4× LDS sample buffer and 10× sample reducing agent (Novex) was added to each sample. Protein was denatured by heating at 70°C for 10 min. Samples were separated on a 4–12% Bis-Tris Plus Gel (Invitrogen) alongside SeeBlue Plus2 Prestained Standard ladder (Invitrogen) and transferred onto a PVDF membrane (Millipore) for antibody detection. Membranes were stained with Ponceau S to confirm protein transfer, blocked for 1 h rocking at room temperature with 5% milk powder (Sigma) dissolved in 0.05% Tween-20 TBS (TBST), washed three times with TBST and incubated rocking overnight at 4°C with anti-OVA (Abbiotec cat. 250803) in 5% milk in TBST. Membranes were washed with TBST and incubated with HRP-conjugated goat anti-rabbit secondary antibody (Sigma) in 5% milk in TBST. Membranes were washed with TBST and WesternBright substrate (Advansta) was added. Images were acquired using Fujifilm LAS-3000 instrument and software.
Co-culture Antigen Presentation Assay With Bacterial – BMDC– CD4+ T Cells
5 × 104 sorted BMDCs were seeded per well in 96 well plates in RPMI with 10% FBS but no antibiotics. Bacteria expressing mCherry-OVA were added at ratio 10:1. OVA protein (Sigma) was added at a final concentration of 150 μg/mL as a positive control. The cells were incubated with OVA protein or bacteria for 8 h at 37°C in a cell culture incubator at 5% CO2. CD4+ T cells were isolated from OT-II mice by spleen homogenization. Red blood cells were lysed using RBC lysis buffer (Invitrogen) and CD4+ T cells separated magnetically using mouse CD4+ T Cell Isolation Kit (Miltenyi cat. 130-104-454) as per manufacturer’s instructions. Isolated T cells were then labeled with carboxyfluorescein succinimidyl ester (CFSE) Cell Division Tracker kit (BioLegend cat. 423801) as per manufacturer’s instructions. After 8 h co-incubation with OVA protein or bacteria, media was removed from the BMDCs cultures and fresh media containing 200 μg/mL gentamicin (Sigma) was added for 30 min at 37°C. The cells were washed three times with PBS and fresh media was added along with CFSE-labeled CD4+ T cells from OT-II mice at a ratio of 1:1 T cells:BMDCs. Plates were incubated at 37°C in 5% CO2 for 5 days. T cells were then collected by centrifugation and stained with anti-mouse CD4 Brilliant Violet421 (BioLegend cat. 135510) and anti-mouse CD25 R-phycoerythrin cyanine 7 (BioLegend cat. 101916). T cell proliferation and activation was analyzed using the BD FACSCelesta Analyzer and data was further analyzed using FCS Express version 5 software (De Novo).
Image Stream Analysis
1 × 106 sorted BMDCs were seed in 6-well plates and co-cultured overnight at 37°C with B. breve EPS+ and B. breve EPS– at MOI 10:1. Cultured BMDCs alone were used as negative controls. Cells were washed and incubated with gentamicin (200 μg/mL) for 30 min at 37°C and then washed 3 times in 1 × PBS and stained with CD45 APC, as previously described. Cells were washed twice in wash buffer, fixed with 1% paraformaldehyde and re-suspended in 50 μl of wash buffer. Fixed and stained cell samples were analyzed with an Amnis ImageStreamX MkII cytometer running INSPIRE software version 200.1.620.0 At least 2000 CD45 positive cells were acquired from each experimental sample. Data was analyzed using IDEAS version 6.2.187.0.
Statistical Analysis
For every experiment at least three biological replicates with three technical replicates per experiment were performed. Statistical analysis was compiled using one way ANOVA with Dunnett’s comparison in GraphPad Prism version 5.03. Statistical comparisons were completed using student’s t-tests and p-values less than 0.05 were considered significant.
Results
Heterogeneous Production of EPS by Human B. breve Strains
The EPS status (Table 3) of twelve B. breve strains (Table 1) was predicted by in silico analyses of B. breve genomes for presence of genes required for EPS production. We used the EPS– mutant of B. breve UCC2003 as a positive control for an EPS sedimentation assay. Because the carbohydrate source can significantly impact EPS production we screened all B. breve strains in media supplemented with different carbohydrates (glucose, lactose, and maltose) (Raftis et al., 2011). B. breve UCC2003 and JCM7017 wild type (WT) strains did not sediment but their EPS– mutants did over a period of 6 h (Figure 1A). B. breve JCM7019 was a consistent EPS producer while B. breve NCTC 11815 sedimented in both glucose and lactose but not in maltose, despite the fact that it was predicted not to produce EPS (Figure 1A and Table 3). B. breve strains 017W439A and 215W447A were EPS producers when cultured in all three carbohydrate sources while the 139W4 and NCFB 2258 strains were not (Figure 1B). B. breve strains 082W48 and 689b were EPS producers when cultured in all three carbohydrate sources while B. breve strains 180W83 and B12L were not (Figure 1C). Comparison between the predicted and actual EPS status of B. breve strains (based on EPS sedimentation assay) demonstrated that all predictions were correct except for strain 139W4 which was predicted to produce EPS but actually sedimented in vitro irrespective of the carbohydrate source used (Table 3).
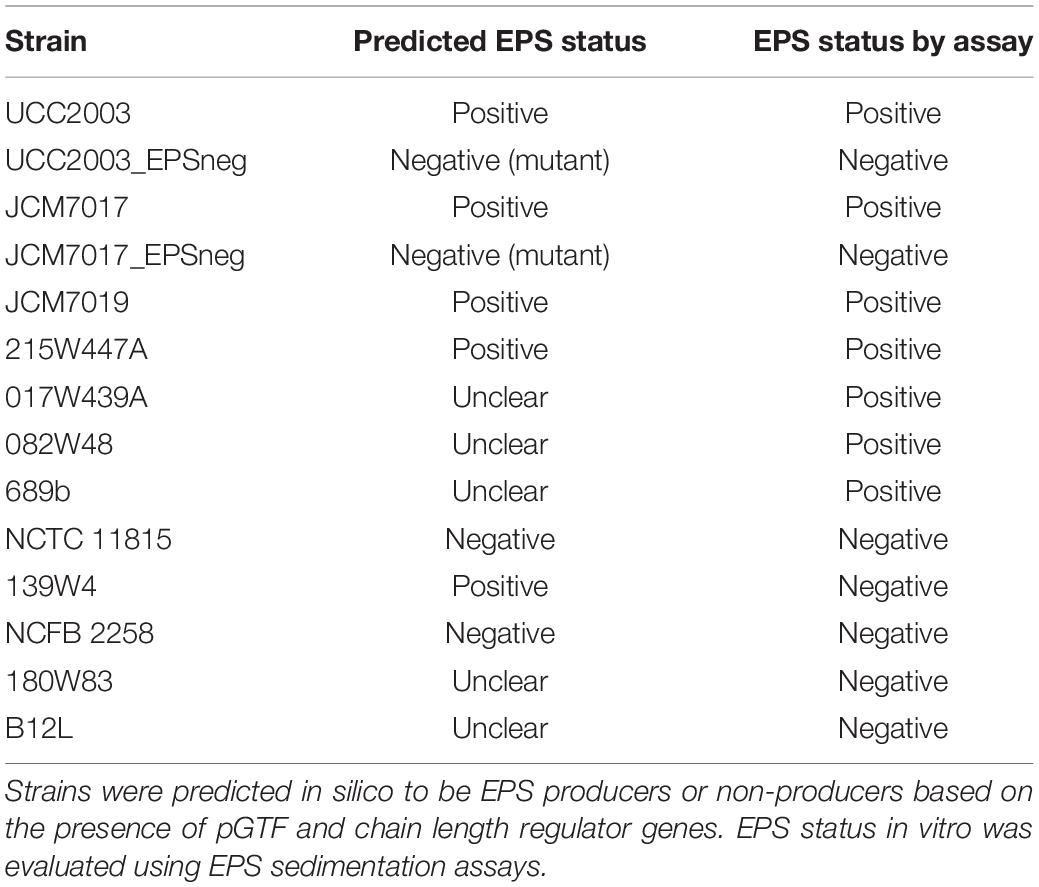
Table 3. EPS status of B. breve strains.
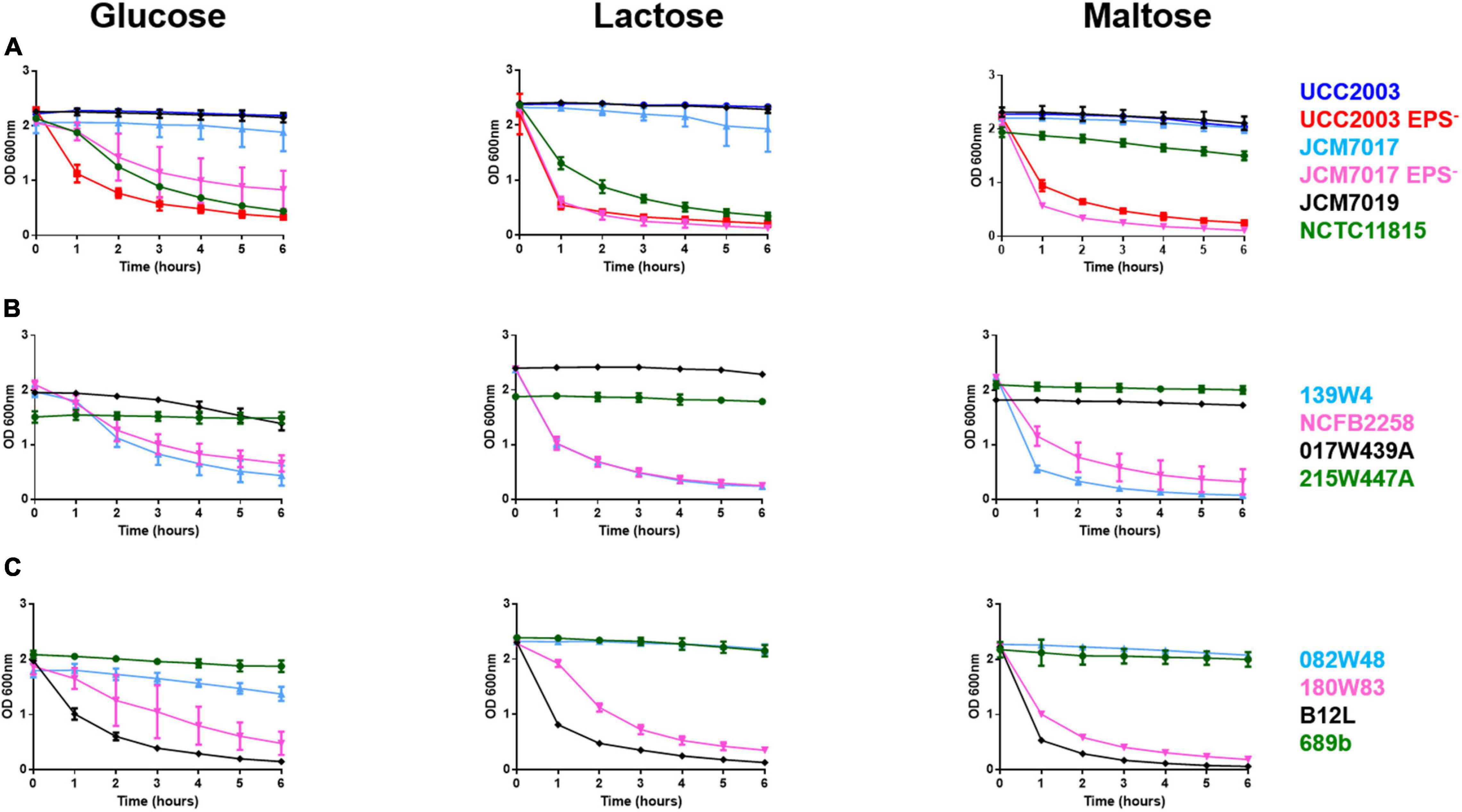
Figure 1. Heterogeneity of EPS production by B. breve strains in different carbon sources. B. breve strains were cultured overnight, homogenized before OD readings were taken from cultures over 6 h with no disturbance to cultures at all. Bacteria were grown in media containing either glucose, lactose, or maltose as a carbon source. This was done for B. breve UCC2003, UCC2003 EPS–, JCM7017, JCM7017 EPS–, JCM7019 and NCTC 11815 (A), 139W4, NCFB 2258, 215W447A and 017W439A (B), 082W48, 180W83, B12L and 689b (C). Data show the average of three independent experiments.
Human THP-1 Monocyte Responses to B. breve Strains Are MyD88-Dependent
To investigate if EPS production by B. breve strains either positively or negatively affected recognition and response to the bacteria by monocytes, the panel of bacteria were co-cultured with WT and MyD88–/– human THP-1 monocyte nuclear factor (NF)-κB reporter cell lines (Figure 2A). NF-κB responses to the panel of B. breve strains showed that there was no clear EPS-dependent associated innate immune response. Although strains (B. breve 215W447A, 689b, and JCM7019) which stimulated highest activation of NF-κB were all EPS positive, the strain (017W439A) which stimulated lowest activation of NF-κB was also EPS positive (Figure 2A). There was decreased activation of NF-κB by B. breve UCC2003 and JCM7017 EPS– isogenic mutant strains compared to their WT parental strains (Figure 2A). Production of TNF-α by THP-1 cells showed a similar trend to NF-κB responses (Figure 2B) for all strains. The lack of response to all strains in the MyD88–/– THP-1 cell line showed that NF-κB activation and TNF-α production was uniformly MyD88-dependent in this cell system (Figure 2). Overall, there was no correlation between EPS status and immune activation of human THP-1 monocytic cells, although all responses to B. breve were MyD88-dependent, regardless of strain.
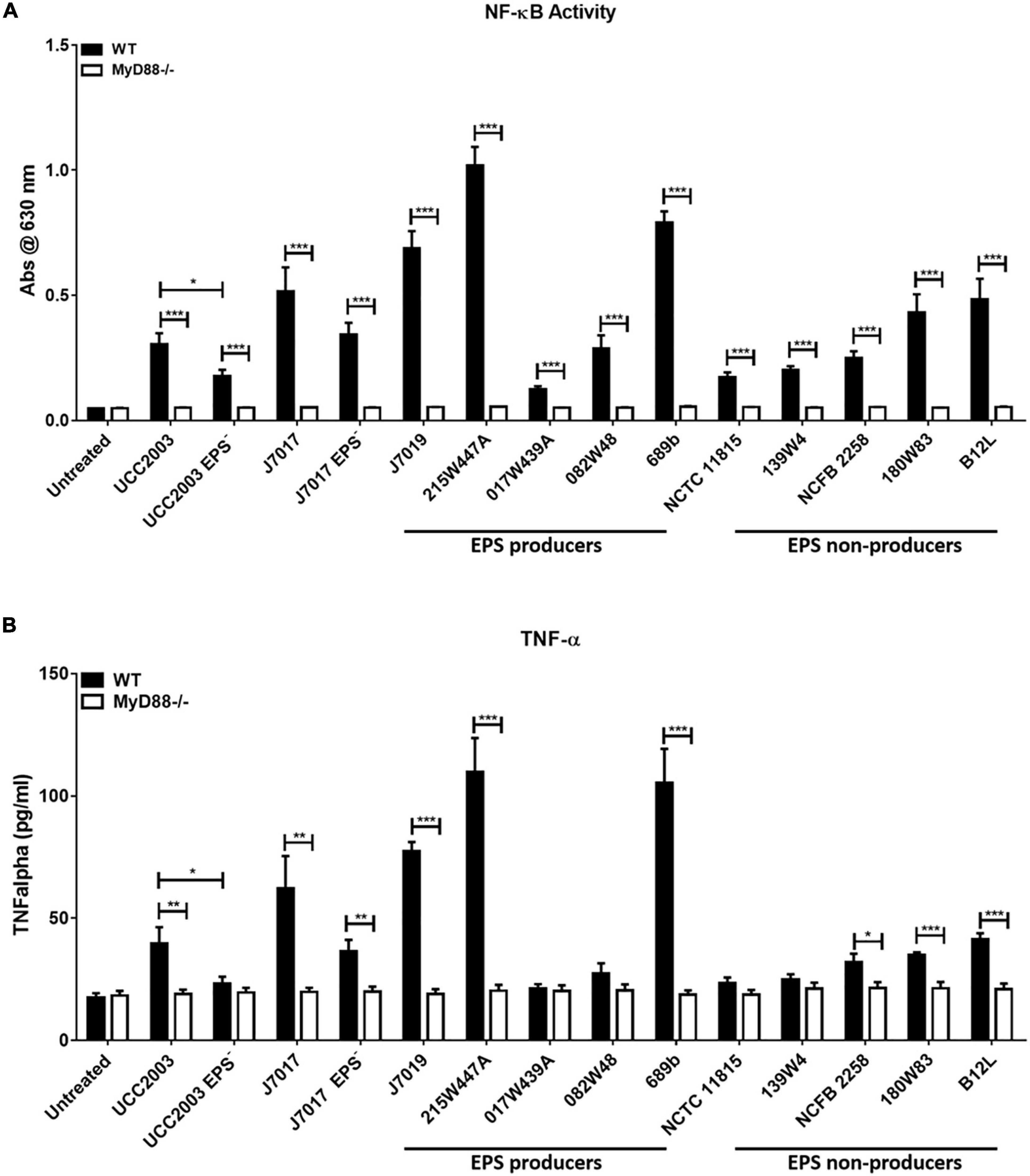
Figure 2. Human THP-1 monocyte responses to B. breve strains are MyD88-dependent but EPS-independent. B. breve strains were incubated with THP1-XBlue and THP1-XBlue-defMyD reporter cell lines for 24 h. (A) NF-κB responses were assessed by measuring reporter gene production, secreted embryonic alkaline phosphatase. (B) TNF-α secretion was assessed by ELISA. Data shown is the average of three independent experiments performed in triplicate wells. Data are presented as mean ± SEM. Statistical analysis was assessed using the student t-test in GraphPad Prism where *p-value < 0.05, **p-value < 0.005, and ***p-value < 0.0005.
EPS Modulates Cytokine Response of BMDMs to B. breve UCC2003 and JCM7017
Because we observed no association between EPS production status and immunomodulation by different B. breve strains in THP-1 cells, we investigated the role of EPS on immunomodulation specifically by B. breve UCC2003 and JCM7017 using the EPS– isogenic mutants of both strains. EPS production by B. breve UCC2003 and JCM7017 parental strains and their respective isogenic EPS– mutants was first confirmed by transmission electron microscopy (TEM) (Figure 3). TEM images showed a noticeably thicker layer of EPS surrounding the WT B. breve UCC2003 strain compared with JCM7017 WT strain (Figures 3A,C). In addition TEM confirmed absence of EPS on the surface of both the B. breve UCC2003 EPS– and JCM7017 EPS– isogenic mutant strains (Figures 3B,D). To understand how EPS affects macrophage and DC cytokine responses to B. breve UCC2003 and JCM7017, primary BMDMs (Figure 4A) and BMDCs (Figure 4B) were co-cultured with the strains and their isogenic EPS– mutants for 24 h. The absence of EPS from B. breve UCC2003 enhanced BMDM TNF-α and IL10 cytokine responses, while the absence of EPS from B. breve JCM7017 reduced cytokine responses (Figures 4C,D). BMDCs did not produce significant TNF-α or IL10 in response to any bacterial strain (Figures 4E,F).
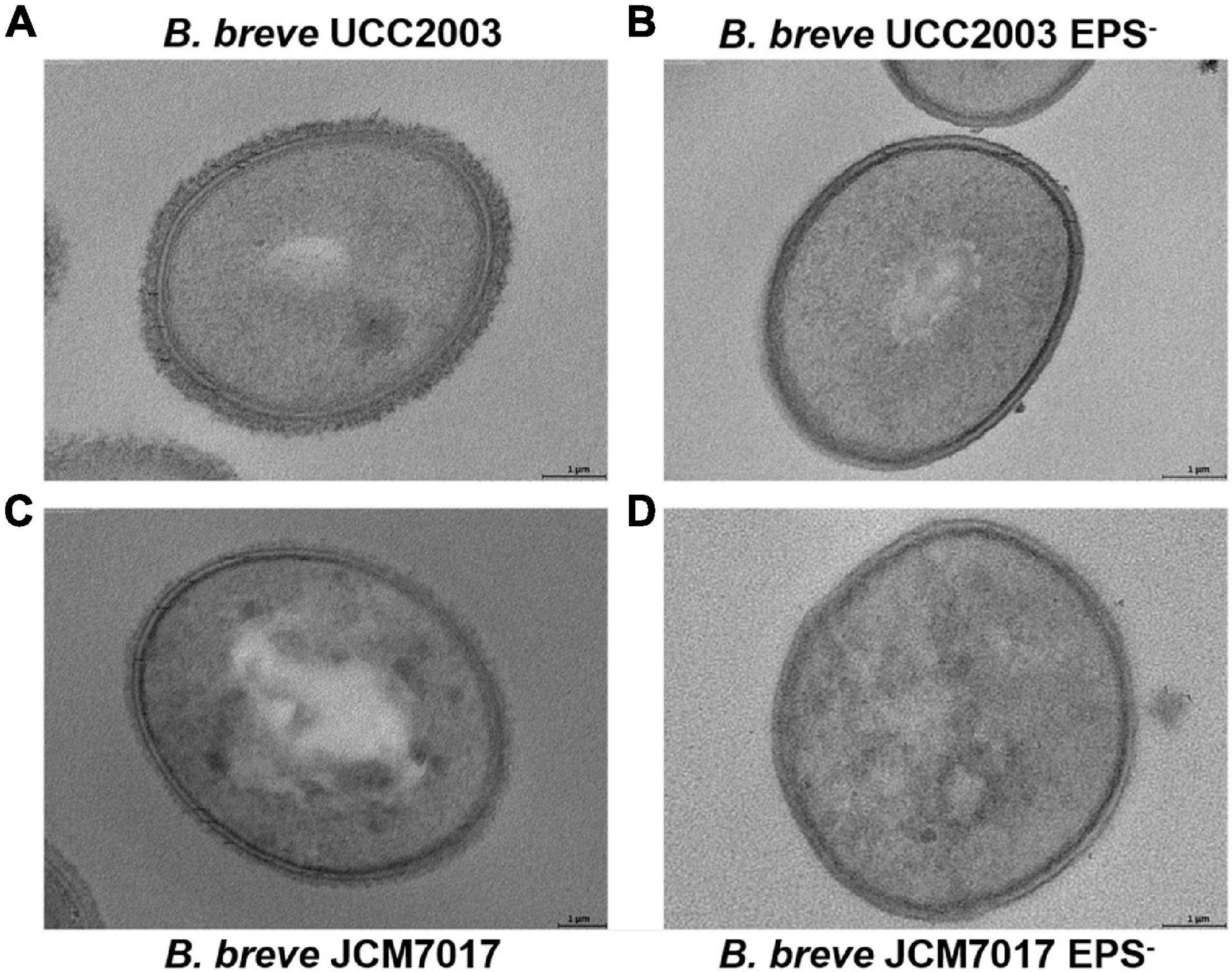
Figure 3. Confirmation of EPS production by B. breve UCC2003, JCM7017 and EPS– mutants. Transmission electron microscopy images of B. breve UCC2003 (A), B. breve UCC2003 EPS– (B), B. breve JCM7017 (C), and B. breve JCM7017 EPS– (D).
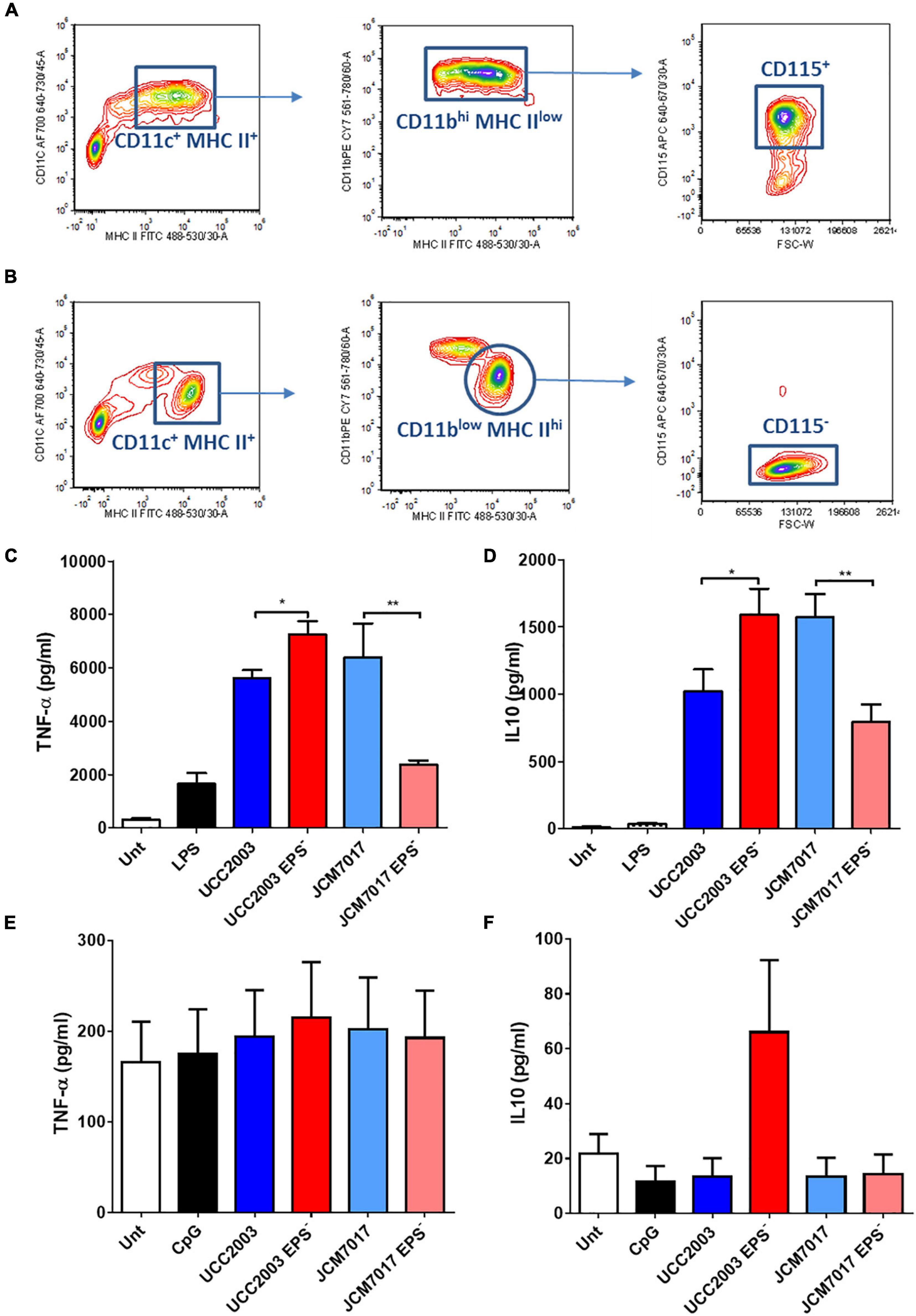
Figure 4. Opposing effects of EPS on murine macrophage cytokine responses to B. breve strains. (A,B) Gating strategy to identify and sort BMDMs. The bone marrow cells were analyzed and sorted by multi-color flow cytometry. Cells were stained for MHC-II, CD11b, CD11c, and CD115. (A) For the adherent cells, MHC-II vs. CD11b contour plots were obtained by gating on MHC-II+CD11c+ cells. Forward size scattered (FSC) vs. CD115 contour plots were obtained by gating on MHC-IIlowCD11bhigh cells. CD115+ cells were sorted and considered as pure macrophages population. (B) For the suspension cells MHC-II vs. CD11b contour plots were obtained by gating on MHC-II+CD11c+ cells. FSC vs. CD115 contour plots were obtained by gating on MHC-IIhighCD11blow cells. CD115– cells were sorted and considered as pure dendritic cells population. (C,D) TNF-α and IL10 secretion from BMDMs co-cultured with B. breve UCC2003, JCM7017 and their isogenic EPS– mutants. (E,F) TNF-α and IL10 secretion from BMDCs co-cultured with B. breve UCC2003, JCM7017 and their isogenic EPS– mutants. Cytokines were analyzed by ELISA. Data is the average of three independent experiments performed in triplicates. Statistical analysis was assessed using the student t-test in GraphPad Prism where *p-value < 0.05 and **p-value < 0.005.
EPS Reduces Activation of BMDCs by B. breve UCC2003 and JCM7017
To further understand the innate immune response of phagocytes to B. breve UCC2003 and JCM7017 and how EPS can affect this, we co-cultured sorted BMDCs with the two WT bacterial strains and their EPS– isogenic mutants for 4 h (Figure 5 and Supplementary Table 1) and 8 h (Supplementary Figure 1). We also co-cultured BMDMs with the strains for 8 h (Supplementary Figure 2). RT-qPCR was performed for cytokine genes (Il10, Tnfa, Il6, Il12a, Il12b, and Il23a), antigen presentation and co-stimulatory genes (H2/MHC-II, Cd80, Cd83, and Cd86), and immune tolerogenic genes (Aldh1a2, Cd274, and Pdcd1lg2). B. breve UCC2003, JCM7017 strains and their EPS– mutants significantly increased the expression of cytokine genes Il10, Tnfa, Il6 (Figures 5A–C) relative to untreated BMDCs at 4 h. B. breve UCC2003 EPS– also increased expression of both Il12a/Il12p35 and Il23a/Il12p19 compared to untreated cells (Figure 5D). However, no significant differences were observed between groups for the expression of Il12b/Il12p40, the gene coding for the other subunit of both IL-12p35/p40 and IL-23p19/p40 cytokines which are associated with Th1 and Th17 responses, respectively. No effects were seen on H2/MHC-II (Figure 5G) or Pdcd1lg2/PDL1 (Figure 5K) expression. Significantly, EPS– mutants of both B. breve UCC2003 and JCM7017 – but not EPS WT strains - increased the expression of the T cell co-stimulatory genes Cd80 (Figure 5H) and Cd83 (Figure 5M), relative to untreated BMDCs while Cd86 (Figure 5I) expression was increased by both B. breve UCC2003 EPS– and WT JCM7017. The absence of EPS from B. breve UCC2003 further enhanced expression of Tnfa, IL6, and Il23a relative to the WT parental strain (Figures 5B,C,F). No significant differences between the parental WT and EPS– mutant strains of B. breve JCM7017 were observed for any of the genes tested. After 8 h co-culture (Supplementary Figure 1 and Supplementary Table 1), all four tested strains continued to significantly enhance Il10 and Il6 expression relative to untreated BMDCs (Supplementary Figures 1A,C). Expression of Tnfa was also significantly increased by all strains except WT B. breve UCC2003 (Supplementary Figure 1B). The absence of EPS from B. breve UCC2003 increased Tnfa transcription (Supplementary Figure 1B) and decreased Il6 transcription relative to its parental WT strain (Supplementary Figure 1C). Both EPS– mutants induced the expression of Il23a compared to untreated (Supplementary Figure 1F) and WT B. breve UCC2003 decreased expression of H2/MHC-II (Supplementary Figure 1G) and Cd83 (Supplementary Figure 1M) relative to untreated cells. Importantly, absence of EPS from B. breve UCC2003 increased transcription of Cd83 (Supplementary Figure 1M) at 8 h relative to its parental WT strain. In BMDM after 8 h of co-culture with the strains (Supplementary Figure 2 and Supplementary Table 1), the absence of EPS from B. breve JCM7017 increased Il10 (Supplementary Figure 2A), Tnfa (Supplementary Figure 2B), and Il12a (Supplementary Figure 2D) transcription. B. breve UCC2003 and its associated EPS– mutant showed a similar, non-significant trend for Tnfa and Il12a gene expression. EPS absence from B. breve UCC2003 increased Cd274 expression (Supplementary Figure 2K). No other significant differences were observed in the genes analyzed (Supplementary Figures 2C,E–M). Overall, this gene expression data indicate that the presence of EPS suppresses the expression of cytokine and co-stimulatory genes required for activation and maturation of dendritic cells in response to B. breve UCC2003 and JCM7017 strains.
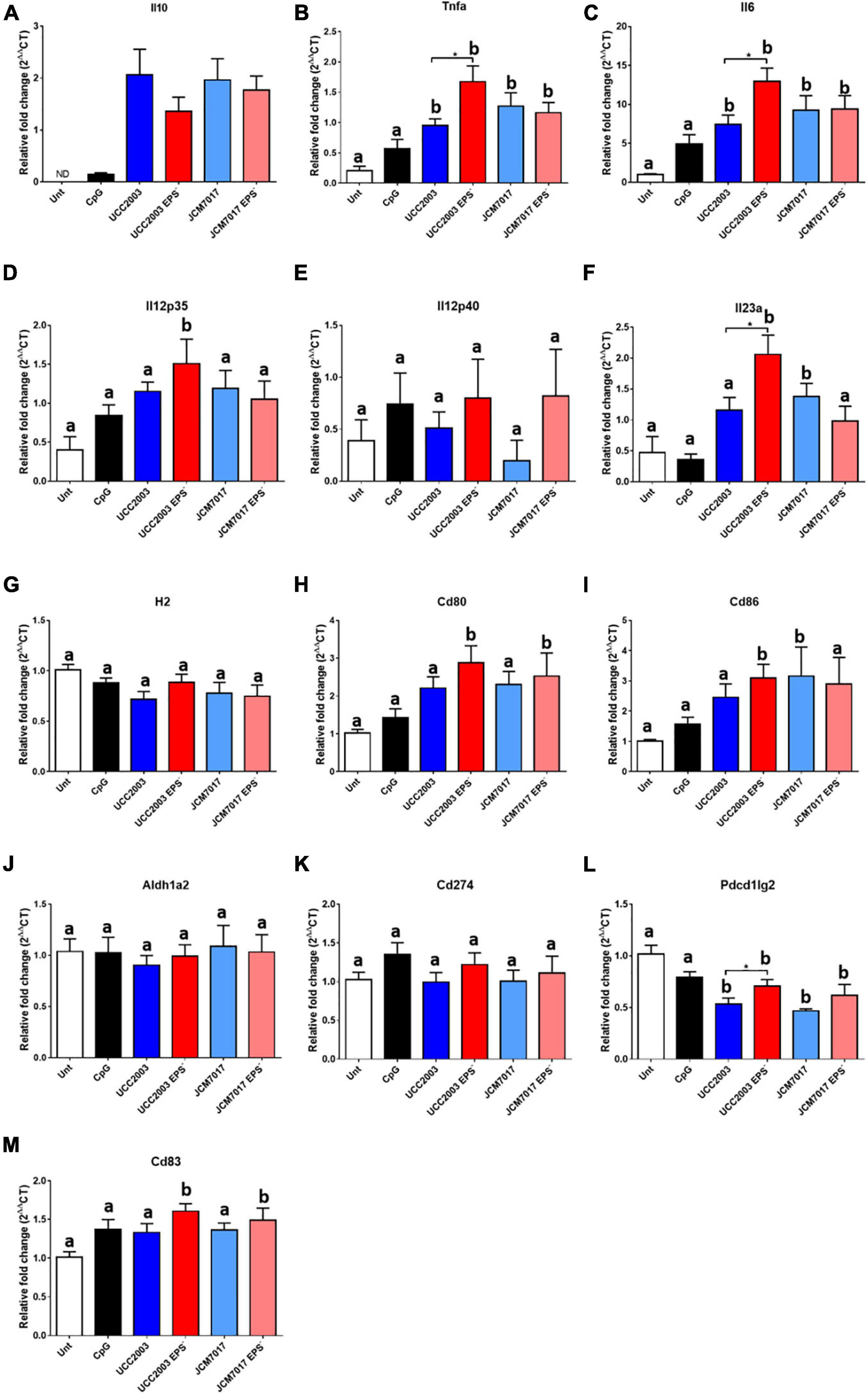
Figure 5. EPS modulates the expression of immunomodulatory genes in BMDCs. Sorted BMDCs were co-cultured or not [untreated (Unt)] with B. breve strains as indicated (CpG was used as a positive control) for 4 h. RT-qPCR was carried out and the expression of (A–F) cytokine genes (G–I) antigen presentation and co-stimulatory genes and (J–M) tolerogenic genes were assessed. Expression changes are relative to β-actin. Data show the average of three independent experiments. Gene expression differences between the groups were compared first with the untreated control. Different letters (a and b) indicate statistical difference for each presented gene expression. Letter a indicates no statistical difference between the indicated group and the untreated control, letter b indicates significant differences between the indicated group and the untreated control (p < 0.05 of Dunnett’s test). Then, the student t-test was used to compare wild type Bifidobacteria (both UCC2003 and JCM7017) with their isogenic EPS mutants (*p-value < 0.05 and **p-value < 0.005).
B. breve EPS Blocks Antigen Specific Activation of CD4+ T Cells by BMDCs
To further investigate how EPS impacts both the recognition and response of phagocytes to B. breve UCC2003 and JCM7017, an antigen presentation experiment was designed (Helft et al., 2015). B. breve UCC2003 and JCM7017 and their corresponding isogenic EPS– mutant strains were transformed with pMG-mCherry or pMG-mCherry OVA plasmid. FACS analysis validated expression of mCherry in the transformant bacteria (Figure 6A and Supplementary Figure 3A). Western blotting analysis confirmed that the bacteria expressed mCherry-OVA (Figure 6B and Supplementary Figure 3B). Bacterial growth profiles showed that strains were not seriously affected in any obvious manner by the presence of the plasmid (Figures 6C,D). Antigen presentation assays were set up as illustrated in Figure 6E. Sorted BMDCs were co-cultured with B. breve strains transformed or not with pMG-mCherry (used as negative control) or pMG-mCherry-OVA for 8 h. BMDCs were treated with OVA protein as a positive control. After this time, potential extracellular bacteria were killed with gentamicin and BMDCs were washed prior to co-culturing with OVA specific OT-II CD4+ T cells for 5 days. T cell activation was determined by examining the numbers of proliferated (CFSElow) CD25+CD4+ T cells. Flow cytometry analysis showed that the absence of EPS from both B. breve UCC2003 and JCM7017 significantly increased T cell activation (Figure 6F and Supplementary Figure 4). To investigate if EPS affected uptake of bacteria, BMDCs were co-cultured with B. breve UCC2003 strains transformed with pMG-mCherry. After this time, BMDCs were analyzed by Image Stream flow cytometry to identify and quantify uptake of bacteria by cells. B. breve UCC2003 EPS+ and EPS– were detected inside CD45+ (Supplementary Figure 4B). cells and no difference in similarity was observed between cells which had taken up these strains (Supplementary Figure 4C). To investigate persistence of viable B. breve UCC2003 and EPS– inside DCs, we co-cultured these bacteria with BMDCs for 2 h. Then, after a gentamicin step to kill extracellular bacteria, we lysed DCs at different time points and plated cell lysates on RCA plates (Supplementary Figure 4D). Colonies of B. breve UCC2003 were recovered at significantly higher levels compared to B. breve UCC2003 EPS– (Supplementary Figure 4D). Collectively, these results indicate that while EPS does not affect the uptake of bacteria by DCs, it does reduce processing and presentation of bacterial derived OVA antigen and activation of antigen specific CD4+ T cells by DCs.
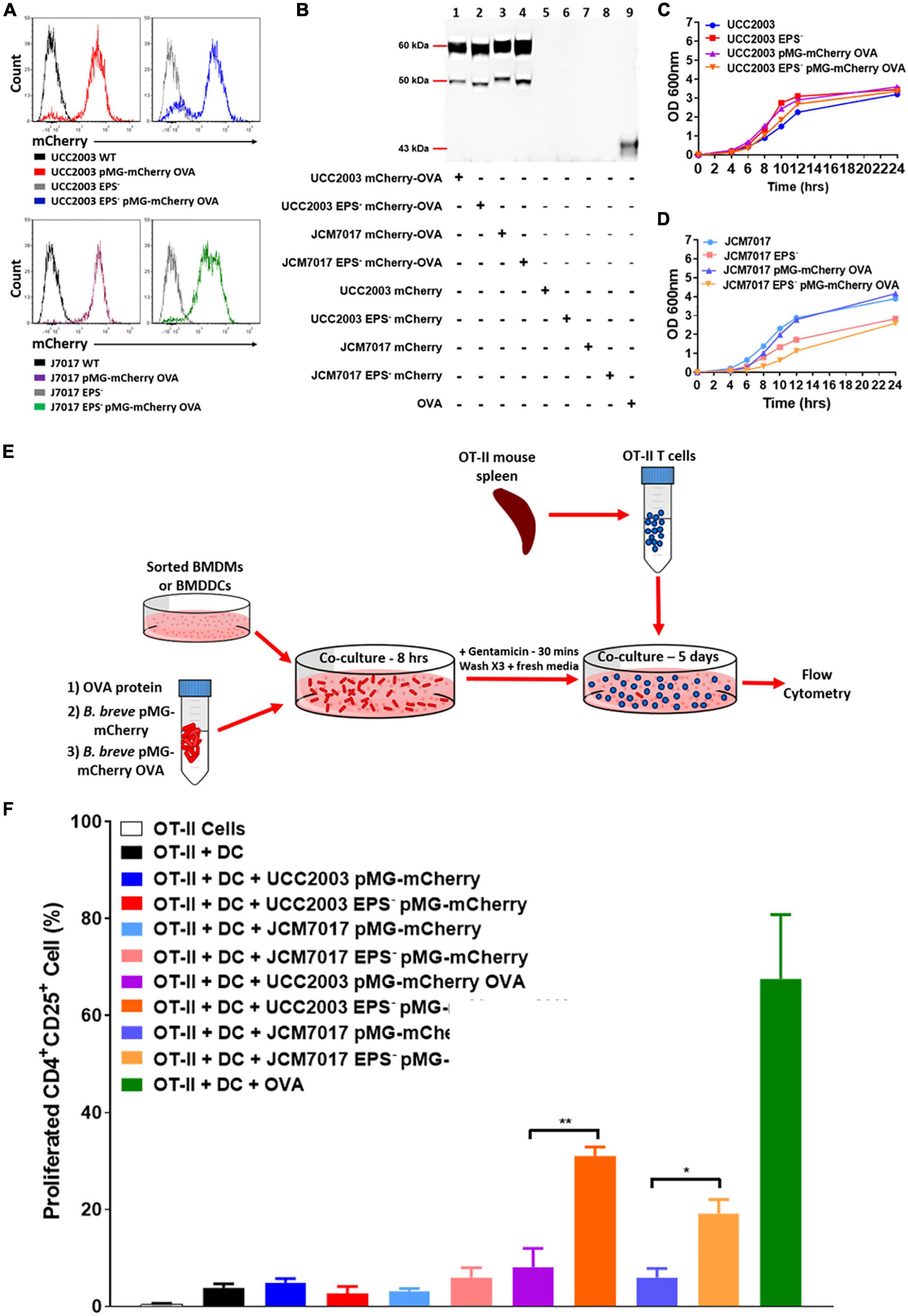
Figure 6. EPS protects from antigen presentation of B. breve to OT-II CD4+ T cells by DCs. (A) mCherry expression (pMG-mCherry OVA) in the indicated B. breve strains by flow cytometry. Histograms were obtained by gating on bacterial cells. Black and gray histograms were obtained from parental B. breve UCC2003 and JCM7017 wild type or EPS–. Red and blue histograms were obtained from B. breve UCC2003 wt and B. breve UCC2003 EPS– transformed with the plasmid pMG-mCherry OVA. Purple and green histograms were obtained from B. breve JCM7017 wt and B. breve JCM7017 EPS– transformed with the plasmid pMG-mCherry OVA. (B) Western blot for OVA protein production by pMG-mCherry-OVA and pMG-mCherry transformed strains of B. breve as indicated. 0.025 μg of ovalbumin was used as a positive control. (C,D) Growth curves of strains transformed or untransformed with the pMG-mCherry-OVA plasmid over 24 h. Growth curves are representative of three experiments. (E) Schematic of the antigen presentation assay setup. (F) Sorted BMDCs were co-cultured or not with different bacterial strains expressing or not OVA (as indicated) for 8 h. OVA protein was used as positive control. mCherry only expressing bacteria were used as negative controls. Cells were then co-cultured with OT-II CD4+ T cells for 5 days and analyzed by multi-color flow cytometry. T cells were stained for CD4, CD25, and CFSE. Percentage of proliferated CD4+CD25+ OT-II T cells was quantitated and presented as mean ± SEM. Data is the average of three independent experiments performed in triplicate wells. Statistical analysis was assessed using the student t-test in GraphPad Prism where *p-value < 0.05 and **p-value < 0.005.
Discussion
In this paper, we report on the production and immunomodulatory effects of EPS from a collection of human-isolated B. breve strains. EPS was produced differentially across a panel of strains and EPS status did not correlate with obvious immunomodulatory effects in a series of immune cell-based assay systems. However, B. breve EPS was required for suppression of gene expression associated with and required for full maturation of DCs. Consistent with this, EPS was necessary to prevent processing and presentation of antigen from the strains and antigen-specific activation of CD4+ T cells by DCs. Overall, we have identified a new immunomodulatory role for EPS which indicates that EPS is utilized by B. breve for immune evasion perhaps as a mechanism to promote host-microbe mutualism.
The panel of B. breve strains screened appeared to represent a near equal number of EPS producers and non-producers. Heterogeneity in EPS production has been noted before for bifidobacteria and, taking into account the fact that the genes of EPS biosynthesis loci have a lower GC content than the average bifidobacterial genome, it is proposed that some species and strains acquired the ability to produce EPS by horizontal gene transfer (Hidalgo-Cantabrana et al., 2014; Ferrario et al., 2016). By assessing the phenotypic production of EPS by a panel of B. breve strains in different carbohydrate sources, we were able to identify consistent EPS producers and non-producers and compare this phenotypic behavior to in silico analysis. The carbohydrate source has previously been shown to impact on EPS production in both lactobacilli (Raftis et al., 2011) and bifidobacteria (Audy et al., 2010). For example lactose induced the highest production of the glycan in B. longum subsp. longum CRC 002 compared to glucose, galactose, and fructose (Audy et al., 2010) and perhaps this is because lactose itself is a source of glucose and galactose, the two major constituents of EPS in the strain. In addition to carbon source, EPS composition and production can be impacted by other environmental factors such as O2, bacterial growth phase, temperature and pH (Degeest et al., 2001). Therefore, it is possible that strains that were phenotypically negative for EPS production in glucose, lactose and maltose-containing medium may be positive under different growth conditions. For example, B. breve 139W4 was predicted to produce EPS but did not do so in any of the prepared media under the experimental conditions used. It is possible that it is an EPS producer but that the growth conditions used were not optimal for EPS biosynthesis in this strain. Similarly, this may explain why B. breve NCTC 11815 sedimented as an EPS non-producer in glucose and lactose but not in maltose. A screen of bifidobacterial type strains showed that of the 48 (sub)species, 56% were EPS producers, 21% were non-producers and 23% displayed an intermediate phenotype (Ferrario et al., 2016). This variability could also be altered by growth in different media but the mechanistic reasons for this have yet to be explored.
When we assayed for immunomodulatory effects of EPS we found that NF-κB activation in THP-1 monocytes did not correlate with EPS status. This may be due to differences between EPS structures and/or composition of the EPS producers or the presence/absence of other MAMPs involved in triggering NF-κB responses. The fact that there was strain-specific modulation rather than EPS-associated modulation is not entirely surprising as strain-specific effects have been characterized for bifidobacteria (López et al., 2010, 2011, 2012). EPS from Lactobacillus plantarum can act in a TLR2-dependent manner to activate signal transducer and activator of transcription 3 (STAT3) independently of NF-κB (Zhou et al., 2018). It is certainly possible that there may be other mononuclear phagocyte responses to these strains which were not investigated here and that are associated with EPS status. In our study the TLR signaling adaptor MyD88 was necessary to activate the NF-κB pathway by all B. breve strains tested. This was not entirely surprising considering previous studies linking bifidobacterial immunomodulation to the TLR family and MyD88 (Jeon et al., 2012; Schiavi et al., 2018; Verma et al., 2018; Castro-Bravo et al., 2019). The absence of EPS from both B. breve UCC2003 and JCM7017 drove different cytokine responses by BMDMs. While the absence of EPS from B. breve UCC2003 increased the production and secretion of IL10 and TNF-α from BMDMs at 24 h the opposite effect was observed for responses to EPS deficient B. breve JCM7017 with decreased production and secretion of IL10 and TNF-α detected. It is possible that EPS acts as a cloaking MAMP, reducing the exposure of other bacterial MAMPs on the same cell to PRRs, or even that it shields other MAMPs completely. Any of these scenarios could explain why EPS from B. breve UCC2003 appears to modulate more anti-inflammatory effects while EPS from B. breve JCM7017 appears to induce more pro-inflammatory effects. It would be useful to understand the composition and structure of B. breve UCC2003 EPS in order to compare to the already characterized EPS of B. breve JCM7017 (Alhudhud et al., 2018), although TEM images did show significant differences in EPS density between the two strains with B. breve UCC2003 having a thicker EPS layer. However, a pure population of sorted BMDCs did not produce IL10 or TNF-α at all to B. breve UCC2003 or JCM7017 after 24 h of exposure. BMDCs generated and FACS purified using the approach by Helft et al. (2015) demonstrate a tolerogenic phenotype (Vander Lugt et al., 2017), and represent a pure population of DCs with no contaminating macrophages. This might explain why they did not show an obvious inflammatory response to our B. breve strains.
Because we detected EPS-dependent differences in cytokine response of BMDMs to B. breve UCC2003 and JCM7017, we analyzed the expression of a panel of immune genes in BMDMs and BMDCs co-cultured with the strains to further understand how EPS can modulate phagocyte immune responses. Our data indicates that both phagocyte populations sensed the presence of the bacteria and responded through PRR-mediated signaling pathways and transcriptional mechanisms to alter gene expression. In BMDCs, the absence of EPS from B. breve UCC2003 but not from JCM7017, significantly increased Tnfa, Il6, and Il23a expression after 4 h co-culture and decreased, H2/MHC-II, Pdcd1lg2/PD-L2, and Cd83 expression after 8 h co-culture. The absence of EPS from B. breve JCM7017, but not from B. breve UCC2003, significantly increased Il10 and Il23a expression after 8 h co-culture. Significantly, in BMDCs the absence of EPS increased the expression of genes for co-stimulatory molecules (Cd80, Cd86 for UCC2003 and Cd83 for both UCC2003 and JCM7017) typically associated with maturation of DCs. This data is consistent with similar suppressive effects of EPS on expression of cytokine genes (Il12a/Il12p35 and Il23a/Il12p19) associated with T cell differentiation and with suppression of T cell activation in our co-culture antigen presentation assay. In BMDMs, the absence of EPS in B. breve UCC2003 increased the expression of Cd274 and the absence of EPS from B. breve JCM7017, but not from B. breve UCC2003, increased Il10, Tnfa, and Il12a expression after 8 h. Indeed, we noted an interesting discrepancy in BMDMs between cytokine (Il10/IL10 and Tnfa/TNF-α) protein data at 24 h and gene expression data at 8 h in response to the EPS deficient JCM7017 strain. While the strain increased Il10 and Tnfa gene expression at 8 h it decreased IL10 and TNF-α cytokine production at 24 h. This curious finding may have an underlying biological mechanism associated with it that is particular to the JCM7017 EPS– strain and the kinetics of BMDM response to this strain. There is evidence in the literature of similar discrepancies and disconnect between cytokine mRNA expression and protein production and secretion. For example in the context of Crohn’s disease it has been reported that despite macrophages showing normal levels and stability of cytokine mRNA intracellular levels of the protein and secretion were abnormally low (Smith et al., 2009). Interestingly, the WT B. breve JCM7017 strain did not induce Il12b/IL12p40 transcription at all in BMDMs. Il12b/IL12p40 encodes a subunit that can be used in conjunction with Il12a/IL12p35 to generate the IL-12 cytokine (an important inducer of TH1 differentiation) or dimerise with Il23a/IL23p19 to create the IL-23 cytokine (an inducer for TH17 differentiation). Il12b/IL12p40 can alternatively homodimerise to induce macrophage chemotaxis or encourage primed DC migration (Cooper and Khader, 2007). The fact that B. breve JCM7017 did not induce expression of the Il12b/IL12p40 subunit in BMDMs implies that the bacteria may not support differentiation of TH1 or TH17 T cells. It also implies that EPS is responsible for controlling this as the EPS– mutant upregulated the expression of Il12b/IL12p40 in BMDMs.
We used mCherry-Ova expressing B. breve UCC2003 and JCM7017 strains in a co-culture antigen presenting assay to investigate if BMDCs were able to present bacterially expressed antigens to T cells, or if they were just unresponsive to the tested bacterial strains. Although T cell activation was increased by both EPS– mutants expressing OVA, the B. breve UCC2003 EPS– mcherry-OVA strain induced stronger T cell activation than the B. breve JCM7017 EPS– mcherry-OVA strain. This suggests that EPS is more important for B. breve UCC2003 to avoid triggering T cell responses than it is for B. breve JCM7017. This observation also ties in with a previous report that the absence of EPS from B. breve UCC2003 increased T cell numbers from splenocytes cultured with the bacteria (Fanning et al., 2012). It appears that EPS in different strains may be involved in different functions, possibly because of differences in EPS composition and production. Indeed, higher molecular weight EPS is implicated in better bacterial survival but also reduces traits such as host cell adhesion and aggregation with other bacteria (Yu et al., 2019) and so there may be a trade-off between bacterial fitness and ability to affect cell–cell interactions with host and neighbors. This is possibly the reason for the heterogeneity in EPS production observed, even between EPS producers such as B. breve UCC2003 and JCM7017. The absence of EPS from these two strains revealed that it modulates host immune cell responses differently by the two strains but that it also elicits similar functions such as prevention of antigen presentation by BMDCs to T cells. While use of isogenic EPS– mutants offers a potential advantage over the use of purified EPS, observed responses to the bacteria did not clarify if the effect is directly because of the EPS binding a PRR, shielding other MAMP/MAMPs, altering binding affinity of other MAMPs to PRRs, reduction of intra-cellular processing of the bacteria through the phagolysosome or a combination of the above. Certainly EPS is able to shield MAMPs as an EPS– mutant of Lactobacillus rhamnosus GG revealed that the bacteria produced sortase-dependent pili on its surface (Lebeer et al., 2009) and a similar observation was made for B. longum 105-A (Tahoun et al., 2017). Additionally, it has been shown that while purified EPS from B. animalis subsp. lactis triggers TLR4 activation, the whole bacteria itself activates TLR2, TLR2/1 and TLR2/6 (Castro-Bravo et al., 2019). Our study shows that while EPS production was heterogeneous amongst human-isolated B. breve strains; EPS from B. breve UCC2003 and JCM7017 had similar effects on maturation and antigen presentation function of BMDCs. It is important to acknowledge that the biochemical composition and density of EPS observed in both WT strains may contribute to the differences observed in BMDM and BMDCs immune responses. Further detailed biochemical, molecular and cell-based investigations are required to understand the biochemistry and immunological effects of different EPS structures and their effects on dendritic cell biology. Our observations suggest that EPS may contribute to an important immunomodulatory effect of B. breve strains by blocking activation and antigen presentation function of DCs. This would allow these strains to manipulate DCs for immunomodulatory effects promoting immune evasion and host-microbe mutualism by these gut symbionts.
Data Availability Statement
The original contributions generated for this study are included in the article/Supplementary Material, further inquiries can be directed to the corresponding authors.
Ethics Statement
Use of animals for this study was reviewed and approved by Animal Experimentation Ethics Committee of University College Cork.
Author Contributions
VR, DV, SM, and KN: conceptualization. AH, VR, PS, SU, AR-V, ME-T, and FB: methodology and writing – original draft. AH, VR, PS, and AR-V: validation. AH, VR, PS, AR-V, JW, and OH: formal analysis. AH, VR, PS, SU, and AR-V: investigation. MV, DV, SM, and KN: resources. AH, VR, PS, and AR-V: data curation. AH, VR, and KN: writing – review and editing. AH, VR, PS, and AR-V: visualization. KN, SM, and DV: supervision. KN: project administration. DV and KN: funding acquisition. All authors contributed to the article and approved the submitted version.
Funding
This work was supported by grants from Science Foundation Ireland – namely a research center grant (SFI-12/RC/2273) to APC Microbiome Ireland.
Conflict of Interest
OH is employed by the company Luminex Corporation which manufactures the Amnis ImageStream flow cytometer used in this study.
The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We would like to acknowledge and thank the Flow Cytometry Platform, APC Microbiome Ireland located at University College Cork for assistance with flow cytometry analysis.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2021.653587/full#supplementary-material
References
Alhudhud, M., Sadiq, S., Ngo, H., Hidalgo-Cantabrana, C., Ruas-Madiedo, P., Van Sinderen, D., et al. (2018). Extraction of the same novel homoglycan mixture from two different strains of Bifidobacterium animalis and three strains of Bifidobacterium breve. Benef. Microbes 9, 663–674. doi: 10.3920/bm2017.0145
Audy, J., Labrie, S., Roy, D., and LaPointe, G. (2010). Sugar source modulates exopolysaccharide biosynthesis in Bifidobacterium longum subsp. longum CRC 002. Microbiology 156, 653–664. doi: 10.1099/mic.0.033720-0
Bottacini, F., Morrissey, R., Esteban-Torres, M., James, K., van Breen, J., Dikareva, E., et al. (2018). Comparative genomics and genotype-phenotype associations in Bifidobacterium breve. Sci. Rep. 8:10633.
Bottacini, F., Morrissey, R., Roberts, R. J., James, K., van Breen, J., Egan, M., et al. (2017). Comparative genome and methylome analysis reveals restriction/modification system diversity in the gut commensal Bifidobacterium breve. Nucleic Acids Res. 46, 1860–1877. doi: 10.1093/nar/gkx1289
Bottacini, F., O’Connell Motherway, M., Kuczynski, J., O’Connell, K. J., Serafini, F., Duranti, S., et al. (2014). Comparative genomics of the Bifidobacterium breve taxon. BMC Genomics 15:170. doi: 10.1186/1471-2164-15-170
Castro-Bravo, N., Margolles, A., Wells, J. M., and Ruas-Madiedo, P. (2019). Exopolysaccharides synthesized by Bifidobacterium animalis subsp. lactis interact with TLR4 in intestinal epithelial cells. Anaerobe 56, 98–101. doi: 10.1016/j.anaerobe.2019.02.014
Castro-Bravo, N., Sánchez, B., Margolles, A., and Ruas-Madiedo, P. (2018a). “Biological activities and applications of Bifidobacterial Exopolysaccharides: from the bacteria and host perspective,” in The Bifidobacteria and Related Organisms, eds P. Mattarelli, B. Biavati, W. H. Holzapfel, and B. J. B. Wood (Amsterdam: Academic Press), 177–193. doi: 10.1016/b978-0-12-805060-6.00010-7
Castro-Bravo, N., Wells, J. M., Margolles, A., and Ruas-Madiedo, P. (2018b). Interactions of Surface Exopolysaccharides From Bifidobacterium and Lactobacillus within the intestinal environment. Front. Microbiol. 9:2426. doi: 10.3389/fmicb.2018.02426
Cooper, A. M., and Khader, S. A. (2007). IL-12p40: an inherently agonistic cytokine. Trends Immunol. 28, 33–38. doi: 10.1016/j.it.2006.11.002
De Man, J. C., Rogosa, M., and Sharpe, M. E. (1960). A medium for the cultivation of lactobacilli. J. Appl. Bacteriol. 23, 130–135. doi: 10.1111/j.1365-2672.1960.tb00188.x
Degeest, B., Vaningelgem, F., and De Vuyst, L. (2001). Microbial physiology, fermentation kinetics, and process engineering of heteropolysaccharide production by lactic acid bacteria. Int. Dairy J 11, 747–757. doi: 10.1016/s0958-6946(01)00118-2
Fanning, S., Hall, L. J., Cronin, M., Zomer, A., MacSharry, J., Goulding, D., et al. (2012). Bifidobacterial surface-exopolysaccharide facilitates commensal-host interaction through immune modulation and pathogen protection. Proc. Natl. Acad. Sci. U.S.A. 109, 2108–2113. doi: 10.1073/pnas.1115621109
Ferrario, C., Milani, C., Mancabelli, L., Lugli, G. A., Duranti, S., Mangifesta, M., et al. (2016). Modulation of the eps-ome transcription of bifidobacteria through simulation of human intestinal environment. FEMS Microbiol. Ecol. 92:fiw056. doi: 10.1093/femsec/fiw056
Fischbach, M. A. (2018). Microbiome: focus on causation and mechanism. Cell 174, 785–790. doi: 10.1016/j.cell.2018.07.038
Grimm, V., Gleinser, M., Neu, C., Zhurina, D., and Riedel, C. U. (2014). Expression of fluorescent proteins in bifidobacteria for analysis of host-microbe interactions. Appl. Environ. Microbiol. 80, 2842–2850. doi: 10.1128/aem.04261-13
Helft, J., Böttcher, J., Chakravarty, P., Zelenay, S., Huotari, J., Schraml, B. U., et al. (2015). GM-CSF mouse bone marrow cultures comprise a heterogeneous population of CD11c+ MHCII+ macrophages and dendritic cells. Immunity 42, 1197–1211. doi: 10.1016/j.immuni.2015.05.018
Hidalgo-Cantabrana, C., Algieri, F., Rodriguez-Nogales, A., Vezza, T., Martínez-Camblor, P., Margolles, A., et al. (2016). Effect of a ropy exopolysaccharide-producing bifidobacterium animalis subsp. lactis strain orally administered on DSS-Induced colitis mice model. Front. Microbiol. 7:868. doi: 10.3389/fmicb.2016.00868
Hidalgo-Cantabrana, C., Sánchez, B., Milani, C., Ventura, M., Margolles, A., and Ruas-Madiedo, P. (2014). Genomic overview and biological functions of exopolysaccharide biosynthesis in Bifidobacterium spp. Appl. Environ. Microbiol. 80, 9–18. doi: 10.1128/aem.02977-13
Inturri, R., Molinaro, A., Di Lorenzo, F., Blandino, G., Tomasello, B., Hidalgo-Cantabrana, C., et al. (2017). Chemical and biological properties of the novel exopolysaccharide produced by a probiotic strain of Bifidobacterium longum. Carbohydr. Polym. 174, 1172–1180. doi: 10.1016/j.carbpol.2017.07.039
Jeon, S. G., Kayama, H., Ueda, Y., Takahashi, T., Asahara, T., Tsuji, H., et al. (2012). Probiotic Bifidobacterium breve Induces IL-10-Producing Tr1 Cells in the Colon. PLoS Pathog. 8:e1002714. doi: 10.1371/journal.ppat.1002714
Lebeer, S., Verhoeven, T. L. A., Francius, G., Schoofs, G., Lambrichts, I., Dufrêne, Y., et al. (2009). Identification of a gene cluster for the biosynthesis of a long, galactose-rich exopolysaccharide in Lactobacillus rhamnosus GG and functional analysis of the priming glycosyltransferase. Appl. Environ. Microbiol. 75, 3554–3563. doi: 10.1128/aem.02919-08
Leivers, S., Hidalgo-Cantabrana, C., Robinson, G., Margolles, A., Ruas-Madiedo, P., and Laws, A. P. (2011). Structure of the high molecular weight exopolysaccharide produced by Bifidobacterium animalis subsp. lactis IPLA-R1 and sequence analysis of its putative eps cluster. Carbohydr. Res. 346, 2710–2717. doi: 10.1016/j.carres.2011.09.010
Livak, K. J., and Schmittgen, T. D. (2001). Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 25, 402–408. doi: 10.1006/meth.2001.1262
López, P., González-Rodríguez, I., Gueimonde, M., Margolles, A., and Suárez, A. (2011). Immune response to Bifidobacterium bifidum strains support Treg/Th17 plasticity. PLoS One 6:e24776. doi: 10.1371/journal.pone.0024776
López, P., Gueimonde, M., Margolles, A., and Suárez, A. (2010). Distinct Bifidobacterium strains drive different immune responses in vitro. Int. J. Food Microbiol. 138, 157–165. doi: 10.1016/j.ijfoodmicro.2009.12.023
López, P., Monteserín, D. C., Gueimonde, M., de los Reyes-Gavilán, C. G., Margolles, A., Suárez, A., et al. (2012). Exopolysaccharide-producing Bifidobacterium strains elicit different in vitro responses upon interaction with human cells. Food Res. Int. 46, 99–107. doi: 10.1016/j.foodres.2011.11.020
Mar, J. C., Kimura, Y., Schroder, K., Irvine, K. M., Hayashizaki, Y., Suzuki, H., et al. (2009). Data-driven normalization strategies for high-throughput quantitative RT-PCR. BMC Bioinformatics 10:110. doi: 10.1186/1471-2105-10-110
McCall, M. N., McMurray, H. R., Land, H., and Almudevar, A. (2014). On non-detects in qPCR data. Bioinformatics 30, 2310–2316. doi: 10.1093/bioinformatics/btu239
Raftis, E. J., Salvetti, E., Torriani, S., Felis, G. E., and O’Toole, P. W. (2011). Genomic diversity of Lactobacillus salivarius. Appl. Environ. Microbiol. 77, 954–965. doi: 10.1128/aem.01687-10
Riedel, C. U. (2018). “Chapter 13 - clinical significance of bifidobacteria,” in The Bifidobacteria and Related Organisms, eds P. Mattarelli, B. Biavati, W. H. Holzapfel, and B. J. B. Wood (Cambridge, MA: Academic Press), 221–234. doi: 10.1016/b978-0-12-805060-6.00013-2
Round, J. L., Lee, S. M., Li, J., Tran, G., Jabri, B., Chatila, T. A., et al. (2011). The toll-like receptor 2 pathway establishes colonization by a commensal of the human microbiota. Science 332, 974–977. doi: 10.1126/science.1206095
Round, J. L., and Palm, N. W. (2018). Causal effects of the microbiota on immune-mediated diseases. Sci. Immunol. 3:eaao1603. doi: 10.1126/sciimmunol.aao1603
Sarkar, A., and Mandal, S. (2016). Bifidobacteria—Insight into clinical outcomes and mechanisms of its probiotic action. Microbiol. Res. 192, 159–171. doi: 10.1016/j.micres.2016.07.001
Schiavi, E., Gleinser, M., Molloy, E., Groeger, D., Frei, R., Ferstl, R., et al. (2016). The surface-associated exopolysaccharide of Bifidobacterium longum 35624 plays an essential role in dampening host proinflammatory responses and repressing local TH17 responses. Appl. environ. Microbiol. 82, 7185–7196. doi: 10.1128/aem.02238-16
Schiavi, E., Plattner, S., Rodriguez-Perez, N., Barcik, W., Frei, R., Ferstl, R., et al. (2018). Exopolysaccharide from Bifidobacterium longum subsp. longum 35624TM modulates murine allergic airway responses. Benef. Microbes 9, 761–773. doi: 10.3920/bm2017.0180
Sivan, A., Corrales, L., Hubert, N., Williams, J. B., Aquino-Michaels, K., Earley, Z. M., et al. (2015). Commensal Bifidobacterium promotes antitumor immunity and facilitates anti–PD-L1 efficacy. Science 350, 1084–1089. doi: 10.1126/science.aac4255
Smith, A. M., Rahman, F. Z., Hayee, B., Graham, S. J., Marks, D. J., Sewell, G. W., et al. (2009). Disordered macrophage cytokine secretion underlies impaired acute inflammation and bacterial clearance in Crohn’s disease. J. Exp. Med. 206, 1883–1897. doi: 10.1084/jem.20091233
Tahoun, A., Masutani, H., El-Sharkawy, H., Gillespie, T., Honda, R. P., Kuwata, K., et al. (2017). Capsular polysaccharide inhibits adhesion of Bifidobacterium longum 105-A to enterocyte-like Caco-2 cells and phagocytosis by macrophages. Gut Pathog. 9:27.
Turroni, F., Milani, C., Van Sinderen, D., and Ventura, M. (2018). “Chapter 12 - bifidobacteria: ecology and coevolution with the host,” in The Bifidobacteria and Related Organisms, eds P. Mattarelli, B. Biavati, W. H. Holzapfel, and B. J. B. Wood (Cambridge, MA: Academic Press), 213–220. doi: 10.1016/b978-0-12-805060-6.00012-0
Vander Lugt, B., Riddell, J., Khan, A. A., Hackney, J. A., Lesch, J., DeVoss, J., et al. (2017). Transcriptional determinants of tolerogenic and immunogenic states during dendritic cell maturation. J. Cell Biol. 216, 779–792. doi: 10.1083/jcb.201512012
Verma, R., Lee, C., Jeun, E. J., Yi, J., Kim, K. S., Ghosh, A., et al. (2018). Cell surface polysaccharides of Bifidobacterium bifidum induce the generation of Foxp3(+) regulatory T cells. Sci. Immunol. 3:eaat6975.
Yu, R., Zuo, F., Ma, H., and Chen, S. (2019). Exopolysaccharide-Producing Bifidobacterium adolescentis strains with similar adhesion property induce differential regulation of inflammatory immune response in Treg/Th17 Axis of DSS-Colitis Mice. Nutrients 11:782. doi: 10.3390/nu11040782
Keywords: Bifidobacterium breve, exopolysaccharides, host immune responses, macrophages, dendritic cells, antigen presentation
Citation: Hickey A, Stamou P, Udayan S, Ramón-Vázquez A, Esteban-Torres M, Bottacini F, Woznicki JA, Hughes O, Melgar S, Ventura M, Van Sinderen D, Rossini V and Nally K (2021) Bifidobacterium breve Exopolysaccharide Blocks Dendritic Cell Maturation and Activation of CD4+ T Cells. Front. Microbiol. 12:653587. doi: 10.3389/fmicb.2021.653587
Received: 14 January 2021; Accepted: 11 May 2021;
Published: 16 June 2021.
Edited by:
Lifeng Zhu, Nanjing Normal University, ChinaReviewed by:
Koji Hosomi, National Institutes of Biomedical Innovation, Health and Nutrition, JapanFabien P. Blanchet, Institut National de la Santé et de la Recherche Médicale (INSERM), France
Aristobolo Mendes Da Silva, Minas Gerais State University, Brazil
Copyright © 2021 Hickey, Stamou, Udayan, Ramón-Vázquez, Esteban-Torres, Bottacini, Woznicki, Hughes, Melgar, Ventura, Van Sinderen, Rossini and Nally. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Valerio Rossini, dmFsZXJpby5yb3NzaW5pQHVjYy5pZQ==; Ken Nally, ay5uYWxseUB1Y2MuaWU=
†These authors share senior authorship