 Rewaida Abdel-Gaber1*
Rewaida Abdel-Gaber1* Mohammed Albeshr1
Mohammed Albeshr1 Mohamed A. Dkhil2Nada Almohawis3
Mohamed A. Dkhil2Nada Almohawis3 Kareem A. Abdelmeguid4
Kareem A. Abdelmeguid4 Denis Delic5Saleh Al Quraishy1
Denis Delic5Saleh Al Quraishy1 Esam M. Al-Shaebi1
Esam M. Al-Shaebi1- 1Department of Zoology, College of Science, King Saud University, Riyadh, Saudi Arabia
- 2Department of Zoology and Entomology, Faculty of Science, Helwan University, Cairo, Egypt
- 3Department of Chemistry, College of Science, King Saud University, Riyadh, Saudi Arabia
- 4Botany and Microbiology Department, Faculty of Science, Helwan University, Cairo, Egypt
- 5Translational Medicine and Clinical Pharmacology, Boehringer Ingelheim Pharma GmbH & Co. KG, Biberach, Germany
Introduction: The rise of antibiotic-resistant microbes has diminished antibiotic effectiveness, leading to the exploration of alternatives. Krameria lappacea has been used traditionally for various ailments. This study evaluates the in vitro antibacterial and cytotoxic potential of its roots and identifies its active constituents.
Methods: Roots of K. lappacea were acquired from a Riyadh market. They were extracted using methanol and the maceration method, followed by phytochemical screening via Liquid Chromatography-Electrospray Ionization-Mass Spectrometry. Antibacterial properties were assessed using agar well diffusion and broth microdilution methods, while cytotoxicity was tested on human lung A549 and MCF7 breast cancer cell lines via MTT assay.
Results: Phytochemical analysis of the extract identified alkaloids, phenolics (including anthraquinones and chlorogenic acids), and flavonoids (such as dihydrokaempferol, epicatechin, and fisetin). The most susceptible bacteria were Staphylococcus aureus and Listeria monocytogenes, both Gram-positive, while Gram-negative Escherichia coli and Salmonella enterica were resistant. Extract effectively suppressed bacterial growth, particularly against L. monocytogenes and S. aureus, showing moderate activity against E. coli and S. enterica, with MBC values higher than MICs. Cytotoxicity testing yielded IC50 values of 142.27 ± 1.02 μg/ml and 64.81 ± 0.26 μg/ml, respectively, compared to doxorubicin.
Conclusion: Our findings suggest that K. lappacea possesses notable antibacterial activity against Gram-positive bacteria and demonstrates cytotoxic effects against human cancer cell lines, indicating its potential as a natural source of bioactive compounds for antimicrobial and anticancer applications.
1 Introduction
Herbal medicine has been a cornerstone in the treatment of various ailments for centuries (Maciel et al., 2002). The World Health Organization (WHO) defines herbal medicine as a practice encompassing herbs, herbal materials, herbal preparations, and finished herbal products that contain active ingredients derived from plants or other plant materials, or their combinations (Robinson and Zhang, 2011; Chaachouay and Zidane, 2024). The therapeutic value of these plants is attributed to secondary metabolites, which are organic compounds not directly involved in the plant’s growth, development, or reproduction but play crucial roles in the plant’s defense against pathogens and herbivores (Andrade et al., 2007; Ansari et al., 2023). These metabolites include alkaloids, flavonoids, terpenoids, and phenolics, which have been shown to possess anti-inflammatory, antimicrobial, anticancer, and antioxidant properties (Chaachouay and Zidane, 2024). In recent years, there has been a growing trend toward seeking herbal alternatives due to their accessibility, safety, and affordability. Studies have highlighted the neuroprotective and metabolic regulatory functions of plant-based secondary metabolites, underscoring their potential for therapeutic use (El-Shouny et al., 2018; Ilaghi et al., 2021; Abdel-Gaber et al., 2024; Khanam et al., 2025).
Krameria lappacea, commonly known as rhatany, is a slow-growing shrub belonging to the Krameriaceae family. The roots are rich in diverse bioactive compounds, including phenolics, flavonoids, lignan derivatives, tannins, benzofuran derivatives, and oligomeric proanthocyanidins, which contribute to its pharmacological benefits (Aldakheel et al., 2021; Safta et al., 2023; Abdel-Gaber et al., 2024). Earlier phytochemical investigations have reported the presence of flavonoids and oligomeric proanthocyanidins as the dominant constituents (Scholz and Rimpler, 1989; Genovese et al., 2021; Zabka, 2022), which provide a strong basis for subsequent pharmacological exploration. Traditionally, K. lappacea has been used to treat gastrointestinal disorders (Simpson, 1989) and is valued for its anti-inflammatory and anti-hemorrhagic properties, making it useful in conditions such as stomach ailments, oropharyngeal inflammation, and excessive blood loss (Simpson, 1991; Carini et al., 2002; Safta et al., 2023; Abdel-Gaber et al., 2024). More recently, a wide range of pharmacological activities have been reported, including photoprotective (Carini et al., 2002), antidiabetic (Heiss et al., 2012), vasoprotective (Ladurner et al., 2012), antimicrobial (Bombardelli et al., 2002; Ortiz et al., 2019; Genovese et al., 2021; Zabka, 2022), antibacterial (Scholz and Rimpler, 1989; Ortiz et al., 2019; Safta et al., 2023), anticancer (Al-Oqail, 2021), anticoccidial (Alamari et al., 2024), and insecticidal (Al-Fuhaid, 2018) activities.
This study aims to comprehensively assess the effectiveness of the methanolic extract derived from the roots of K. lappacea. The research will focus on three primary objectives: (i) Identification and characterization of active constituents present in the methanolic extract. (ii) Evaluation of the antibacterial activity of the extract against both Gram-positive and Gram-negative bacterial strains. (iii) Assessment of the cytotoxic potential of the K. lappacea extract against human lung cancer cell lines, specifically A549, and breast cancer cell lines, namely MCF7. Through these comprehensive evaluations, the study aims to contribute valuable knowledge regarding the pharmacological benefits of K. lappacea and its potential use in medicinal applications.
2 Materials and methods
2.1 Plant collection and extract preparation
The roots of K. lappacea were obtained from a local market in Riyadh, Saudi Arabia. A qualified taxonomist, Prof. Mohamed A. El-Sheikh, authenticated the plant specimen, which was assigned the voucher number KSU-22958, at the herbarium of the Botany Department, College of Science, King Saud University (Saudi Arabia). Authentication was based on a detailed examination of the plant’s morphological traits (e.g., root structure, color, and texture) in comparison with standard taxonomic references and databases. While the present identification relied on morphological features, we acknowledge that anatomical analysis of root tissues (e.g., transverse sections, xylem and phloem structure, and histological features) would provide an additional confirmatory layer, and we intend to incorporate such analysis in future work.
The extract of the K. lappacea roots was carried out using maceration following the method described by Manikandan et al. (2008). Briefly, the roots were broken into smaller pieces and then coarsely powdered using a Hummer grinder (ED-CG1400). The dried and powdered roots (100 g) were soaked in 1,000 mL of 70% methanol at room temperature for 24 h with occasional stirring. The mixture was then filtered through Whatman No. 1 filter paper, and the residue was re-macerated twice under the same conditions to ensure maximum extraction of phytochemicals. The combined filtrates were concentrated under reduced pressure using a rotary vacuum evaporator, specifically a Buchi model from Switzerland, at 55 °C and subsequently dried to yield the crude extract. The extract was stored at −20 °C until further analysis.
2.2 Liquid chromatography-electrospray ionization-mass spectrometry (LC-ESI-MS)
LC-ESI-MS was employed to identify the phytochemicals in the extract, in triplicate, to provide a comprehensive profile relevant to its pharmacological properties. The analysis was performed using an XEVO TQD triple quadrupole mass spectrometer equipped with an electrospray ionization (ESI) source (Waters Corporation, Milford, MA, USA). Data were acquired in both positive and negative ionization modes. Chromatographic separation was carried out on an ACQUITY UPLC BEH C18 column (2.1 × 50 mm, 1.7 μm particle size) maintained at 40 °C, with a flow rate of 0.2 mL/min. The mobile phase consisted of (A) water containing 0.1% formic acid and (B) acetonitrile containing 0.1% formic acid, using a linear gradient program optimized for phytochemical separation. For mass spectrometric detection, the capillary voltage was set at 3.0 kV, cone voltage at 30 V, desolvation temperature at 350 °C, source temperature at 120 °C, and desolvation gas (nitrogen) flow at 800 L/h. Mass spectra were collected over an m/z range of 100–1,200 with a scan time of 0.2 s. Compound identification was achieved by comparing experimental mass spectra and fragmentation patterns with reference spectra from the Waters UNIFI Scientific Information System and cross-verified using the MassBank and METLIN databases. A matching score threshold of ≥85% similarity was applied for tentative identifications, while compounds with ≥95% match were considered high-confidence identifications. Retention times and fragmentation consistency were additionally considered to confirm compound identity and minimize false positives. This combined approach—utilizing spectral matching, retention time verification, and fragmentation pattern analysis—ensured a robust and reproducible identification of the phytochemicals present in the extract.
2.3 Microorganisms
The test organisms were sourced from the American Type Culture Collection (ATCC, Manassas, USA) and included two Gram-positive bacteria: Listeria monocytogenes (ATCC 7644) and Staphylococcus aureus (ATCC 25923), as well as two Gram-negative bacteria: Escherichia coli (ATCC 8739) and Salmonella enterica (ATCC 14028).
2.4 Evaluation of the antibacterial potential of the methanolic extract
The experiment was conducted using the agar well-diffusion method, following the guidelines set by the Clinical and Laboratory Standards Institute (CLSI, 2012, 2019), Gholizadeh et al. (2013), and Balouiri et al. (2016). In summary, 100 μL of each reference strain (1 × 105 colony-forming unit (CFU)/mL) was separately spread over the Mueller-Hinton Agar (MHA) medium. After the medium had fully solidified, we utilized a sterile cork borer to carefully create wells with a precise diameter of 0.6 cm. These wells were designed to accommodate 50 μL of different concentrations of the methanolic extract, which varied from 0 to 133 mg/mL. The plates were first placed in the refrigerator for 30 min to enhance the diffusion of the sample extracts into the agar (Gonelimali et al., 2018; Eloff, 2019). After this period, the plates were incubated at 37 °C for 24 h. The antibacterial potential of each sample was assessed by measuring the diameter of the zones of inhibition (ZOI) in millimeters (mm), using three replicates. This activity was compared to that of several conventional antibiotics with varying modes of action: amikacin (AK, 30 μg/mL) as a protein synthesis inhibitor; amoxicillin (AX, 25 μg/mL) and ampicillin/sulbactam (SAM, 20 μg/mL) as cell wall synthesis inhibitors; norfloxacin (NOR, 10 μg/mL) and ofloxacin (OFX, 5 μg/mL) as DNA replication inhibitors. Sterile dimethyl sulfoxide (DMSO) of 50 μL was utilized as a negative control.
2.5 Determination of minimum inhibitory concentrations (MICs)
The broth microdilution assay was used to determine the minimum inhibitory concentrations (MICs) of each tested sample, following the protocols outlined by EUCAST (2003), Gholizadeh et al. (2013), and CLSI (2012, 2019). A stock solution of the extract was prepared by carefully dissolving 133 mg of the extract in 1 mL of 1% dimethyl sulfoxide (DMSO). This process involved gradually adding the extract to the DMSO while stirring gently to ensure complete dissolution. The resulting solution was then thoroughly mixed to achieve a homogeneous stock solution suitable for further experimental use. Following this, 100 μL of Mueller-Hinton Broth (MHB) was added to each well from wells 2–12. Then, 150 μL of the stock solution was added to the first column of a microtiter plate. A two-fold dilution was performed by transferring 100 μL from the first well to the next, continuing this process until the 11th well. In the final step, 100 μL of each microbial culture (1 × 105 CFU/mL) was added to each well, except the last one, which served as the blank control. All microtiter plates were incubated at 37 °C for 24 h for the inoculated bacterial strains. Absorbances were measured at a wavelength of 620 nm using an automated microplate reader (ChroMate 4,300, United States), and the results were graphically represented using Microsoft Excel version 2019. Statistical significance was determined with a p-value of < 0.05 compared to the control group. Chloramphenicol (1 mg/mL) and ciprofloxacin (1 μg/mL) were used as positive controls for antibacterial activity.
2.6 Assessment of the minimum bactericidal concentration (MBC)
The minimum bactericidal concentration (MBC) for the extract was determined using the dilution in broth method, following the guidelines set by CLSI (2019). Aliquots of 50 μL were taken from all wells that showed no visible growth after 24 h of incubation at 37 °C and were spread onto Mueller-Hinton agar (MHA) plates. These plates were then incubated for an additional 24 h at 37 °C. The limit of detection (LOD) for this technique is 10 CFU/mL; therefore, the absence of any growth on the MHA plates indicated that the concentration was below this threshold. This implies that the initial concentration of 105 CFU/mL had been significantly reduced to a level below 10 CFU/mL. Subsequently, the MBC was assessed as the minimum concentration of the extract capable of killing more than 99.99% of the bacteria present. Three replicates of each trial were conducted.
2.7 Cytotoxicity assay
The cytotoxicity of the extract was evaluated, in triplicate, using the MTT assay, specifically with 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide, according to Mosmann (1983). The assay was applied to human lung (A549) and breast (MCF7) cancer cell lines obtained from ATCC (USA). The cells were cultured in Dulbecco’s Modified Eagle Medium (DMEM) (Gibco, USA) supplemented with 1% penicillin/streptomycin and 10% fetal bovine serum (FBS) (Gibco, USA). They were incubated at 37 °C in a humidified atmosphere containing 5% CO2 for 24 h in a CO2 incubator (LCO-065AI, Daihan LabTech Co., LTD). In summary, cells were plated in a 96-well culture plate at a density of 2 × 105 cells/ml and allowed to proliferate for 24 h. The cells were treated with various extract concentrations (500, 250, 125, 62.5, 31.25, and 15.625 μg/mL) and doxorubicin. After treatment, 20 μL of MTT solution (5 μg/mL in PBS) was added to each well, and the plate was incubated for 4 h at 37 °C. The yellow tetrazolium salt was reduced to purple formazan crystals, which were then dissolved in 100 μL of DMSO. The absorbance of the resulting colored solution was measured spectrophotometrically at 570 nm on a microplate reader (BioTek, USA). Cell viability (%) was calculated following the method described by Shirley and Lillehoj (2012). The doxorubicin-treated cells served as a positive control. IC50 values (the concentration of extract that causes 50% inhibition) were calculated from the dose–response curve of cell viability percentages using OriginPro software.
3 Results and discussion
The phytochemical profile obtained in this study revealed several compounds, including flavonoids and oligomeric proanthocyanidins, which are consistent with previous reports on K. lappacea (Scholz and Rimpler, 1989; Genovese et al., 2021; Zabka, 2022). However, some detected compounds, such as sanguinarine, an alkaloid, which have not been previously reported in this species and were identified tentatively based on MS/MS fragmentation patterns. Its identification in the present study was based on MS/MS fragmentation and database matching; however, confirmation requires further characterization using authentic standards, multiple reaction monitoring (MRM), and complementary structural analyses (e.g., NMR). Therefore, sanguinarine is reported here as a preliminary finding that warrants validation in future investigations. Previous studies by Yan et al. (2021) and Ardhany et al. (2022) have shown that sanguinarine exhibits potent antibacterial activity through multiple mechanisms, including disruption of bacterial membranes, DNA intercalation, induction of reactive oxygen species (ROS), and inhibition of essential bacterial enzymes, ultimately leading to bacterial cell death. Recent research by Gu et al. (2023) further corroborates these findings, demonstrating that sanguinarine chloride hydrate (SGCH) effectively compromises the cell wall and membrane integrity of Staphylococcus aureus, inducing oxidative stress and bacterial lysis.
In addition to sanguinarine, six flavonoids were detected, each associated with specific biological activities. Eriodictyol possesses antioxidant, anti-inflammatory, neuroprotective, and anticancer properties (Deng et al., 2020). Dihydrokaempferol demonstrates in vitro antimicrobial activity against diverse microorganisms (Elmongy et al., 2022). Isobavachin has broad-spectrum pharmacological effects, including antimicrobial and cytotoxic actions (Chung et al., 2024). Epicatechin exerts antioxidant, antimicrobial, anti-inflammatory, and anticancer effects by scavenging ROS, disrupting bacterial membranes, and inducing apoptosis in cancer cells (Prakash et al., 2019). Fisetin has been shown to inhibit cancer cell proliferation, induce apoptosis, suppress angiogenesis, and protect against oxidative stress (Zhou et al., 2023). Naringenin exhibits significant antioxidant, antimicrobial, and anti-inflammatory activity, contributing to both antibacterial and anticancer effects (Moon et al., 2011; Wang et al., 2016). Recent reviews by Zhang et al. (2025) and Liu et al. (2025) have highlighted the antibacterial potential of flavonoids, emphasizing their mechanisms, including membrane disruption, inhibition of nucleic acid and protein synthesis, and suppression of efflux pumps.
Two phenolic compounds—chlorogenic acid and anthraquinones—were also identified. Chlorogenic acid is known for antibacterial, antioxidant, anti-inflammatory, and anticancer activities, functioning through mechanisms such as ROS scavenging, disruption of bacterial cell walls, and induction of apoptosis in tumor cells (Miao and Xiang, 2020). Anthraquinones are recognized for antimicrobial and anti-inflammatory properties and can compromise bacterial membrane integrity (Malik and Müller, 2016). These findings are consistent with those of Safta et al. (2023), who identified similar compounds in K. lappacea root extracts and attributed their bioactivity to these constituents. Each compound exhibited distinct peak areas and retention times, which were carefully documented in Table 1.
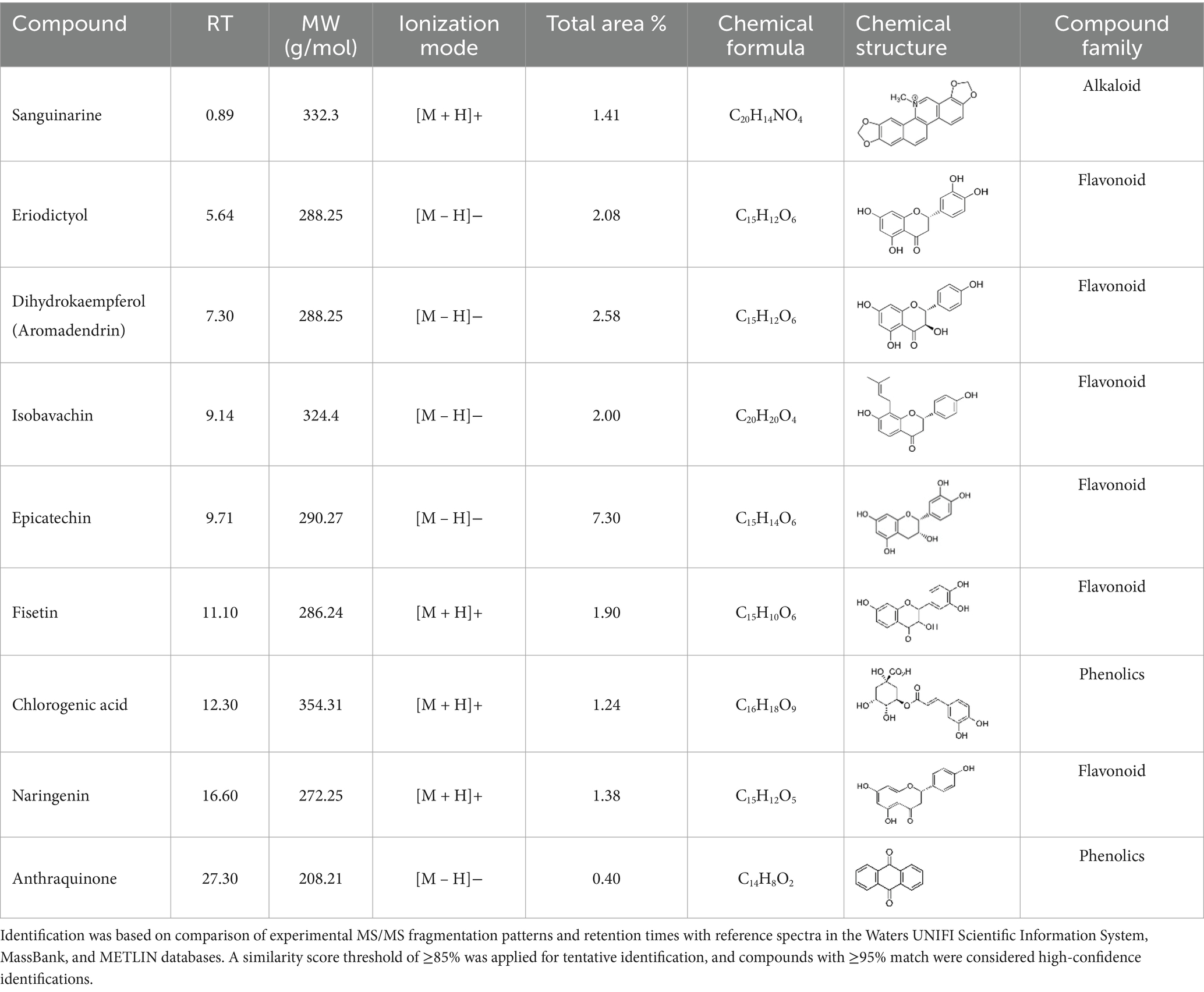
Table 1. Phytochemical compounds were identified from the K. lappacea roots extract using LC-ESI-MS in both positive and negative ionization modes.
The antibacterial activity of K. lappacea extract is likely mediated through several complementary pathways. Flavonoids and related phytochemicals can disrupt bacterial cell walls and membranes, causing leakage of intracellular contents and compromising structural integrity (Genovese et al., 2021). Some compounds intercalate with bacterial DNA, hindering replication and transcription (Abdel-Gaber et al., 2024). Others bind to bacterial enzymes and proteins, impairing energy metabolism, protein synthesis, and cell wall biosynthesis. Certain constituents may also interfere with quorum-sensing pathways, reducing biofilm formation and inducing ROS-mediated oxidative stress, collectively leading to bacterial lysis (Genovese et al., 2021; Abdel-Gaber et al., 2024). Collectively, these mechanisms act synergistically to inhibit bacterial growth and ultimately cause bacteriolysis.
In agar diffusion assays, the extract produced a ZOI of 14 mm against S. aureus and 19 mm against L. monocytogenes (Figure 1; Table 2). This aligns partially with Babiker et al. (2020), who reported a 19 mm ZOI for Krameria triandra against S. aureus, highlighting species-specific differences in bacterial sensitivity potentially linked to variations in the phytochemical profile and structure–activity relationships of the extracts. The stronger effect against L. monocytogenes may reflect differences in cell wall composition and permeability among Gram-positive bacteria (Abdel-Gaber et al., 2024). In contrast, Gram-negative bacteria (E. coli and S. enterica) were resistant, likely due to the protective outer membrane and lipopolysaccharide layer, which restricts entry of hydrophobic phytochemicals (Inouye et al., 2001; Horne et al., 2020; Salem et al., 2023). This resistance was consistent with Nayf and Salman (2021), who observed larger inhibition zones with standard antibiotics (such as Amikacin, Amoxicillin, Ampicillin/sulbactam, Norfloxacin, and Ofloxacin) compared to plant extracts. Overcoming Gram-negative resistance may require combining phytochemicals with agents that disrupt the outer membrane or inhibit efflux pumps, thereby improving intracellular accumulation of active compounds. Several studies have reported such effects: Wesseling and Martin (2022) highlighted peptide-based permeabilizers that sensitize bacteria to antimicrobials, Duffey et al. (2024) catalogued numerous efflux pump inhibitors in E. coli and Pseudomonas, and Al-Sallami et al. (2023) demonstrated that plant extracts like rosemary, clove, and cumin can potentiate antibiotic activity via efflux pump inhibition. Similarly, Minnelli et al. (2024) showed that amphiphilic α-hydrazido acids permeabilize bacterial membranes and act synergistically with antibiotics. Building on these precedents, combining K. lappacea extract with safe permeabilizers or efflux pump inhibitors could broaden its spectrum against Gram-negative pathogens, a hypothesis that warrants validation through synergistic assays, intracellular accumulation studies, and toxicity assessments.
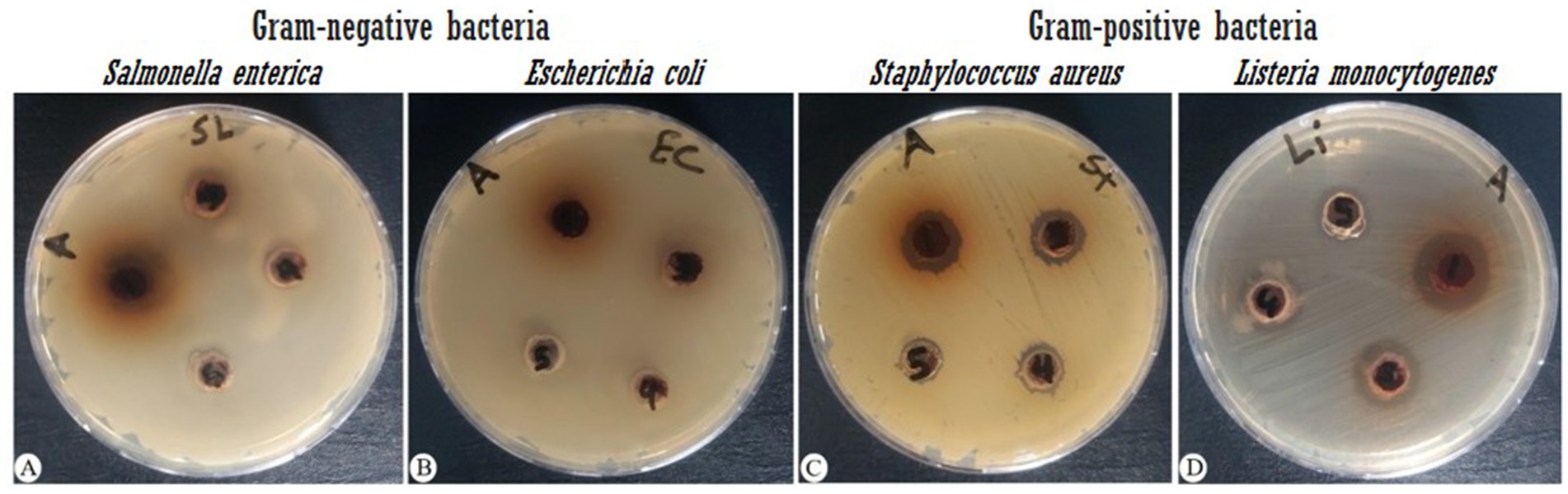
Figure 1. Antibacterial activity of K. lappacea methanolic extract against four bacterial strains, measured as the zone of inhibition (mm). Extract concentrations tested were 133, 66.7, 33.3, 16.7, and 8.3 mg/mL. (A) Salmonella enterica (ATCC 14028), (B) Escherichia coli (ATCC 8739), (C) Staphylococcus aureus (ATCC 25923), and (D) Listeria monocytogenes (ATCC 7644).
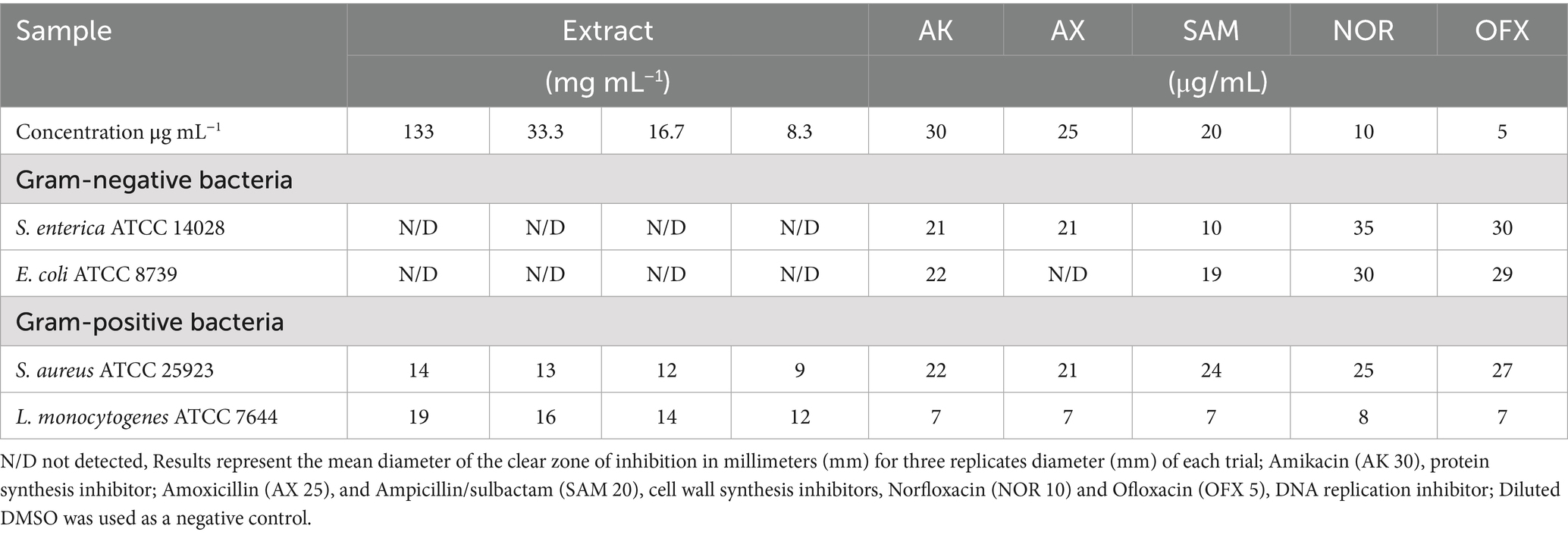
Table 2. Antibacterial susceptibility test results of the extract.
MIC and MBC analyses revealed effective inhibition of Gram-positive strains, particularly L. monocytogenes (MIC = 67 mg/mL) and S. aureus (MIC = 133 mg/mL), with MBCs ranging from 34 to 67 mg/mL (Figure 2; Table 3). These results are supported by Bouarab-Chibane et al. (2019) and Hochma et al. (2021), highlighting the susceptibility of Gram-positive bacteria to phenolic compounds due to their simpler cell wall structure. In line with our results, Al-Oqail (2021) investigated the antibacterial and cytotoxic activities of K. lappacea root extracts. Their findings revealed that the ethanolic extract exhibited inhibitory effects against both Gram-positive and Gram-negative bacteria, with MIC values ranging from 250 to 1,000 μg/mL. Mendes et al. (2022) highlighted that phenolic compounds, such as anthraquinones and chlorogenic acids, act by disrupting peptidoglycan cross-links, leading to leakage of proteins and phospholipids, thereby compromising membrane integrity and causing bacterial death. In contrast, certain flavonoids, such as dihydrokaempferol-3-O-α-L-rhamnoside, have been reported to overcome Gram-negative barriers depending on chemical structure (Elmongy et al., 2022). Alkaloids, including sanguinarine, further enhance antibacterial efficacy by compromising membranes, inhibiting DNA and protein synthesis, and inducing ROS-mediated stress (Yan et al., 2021; Ardhany et al., 2022). Taken together, the present results reinforce the antimicrobial potential of K. lappacea, while comparative studies highlight the importance of both bacterial cell wall composition and specific phytochemical structures in determining susceptibility.
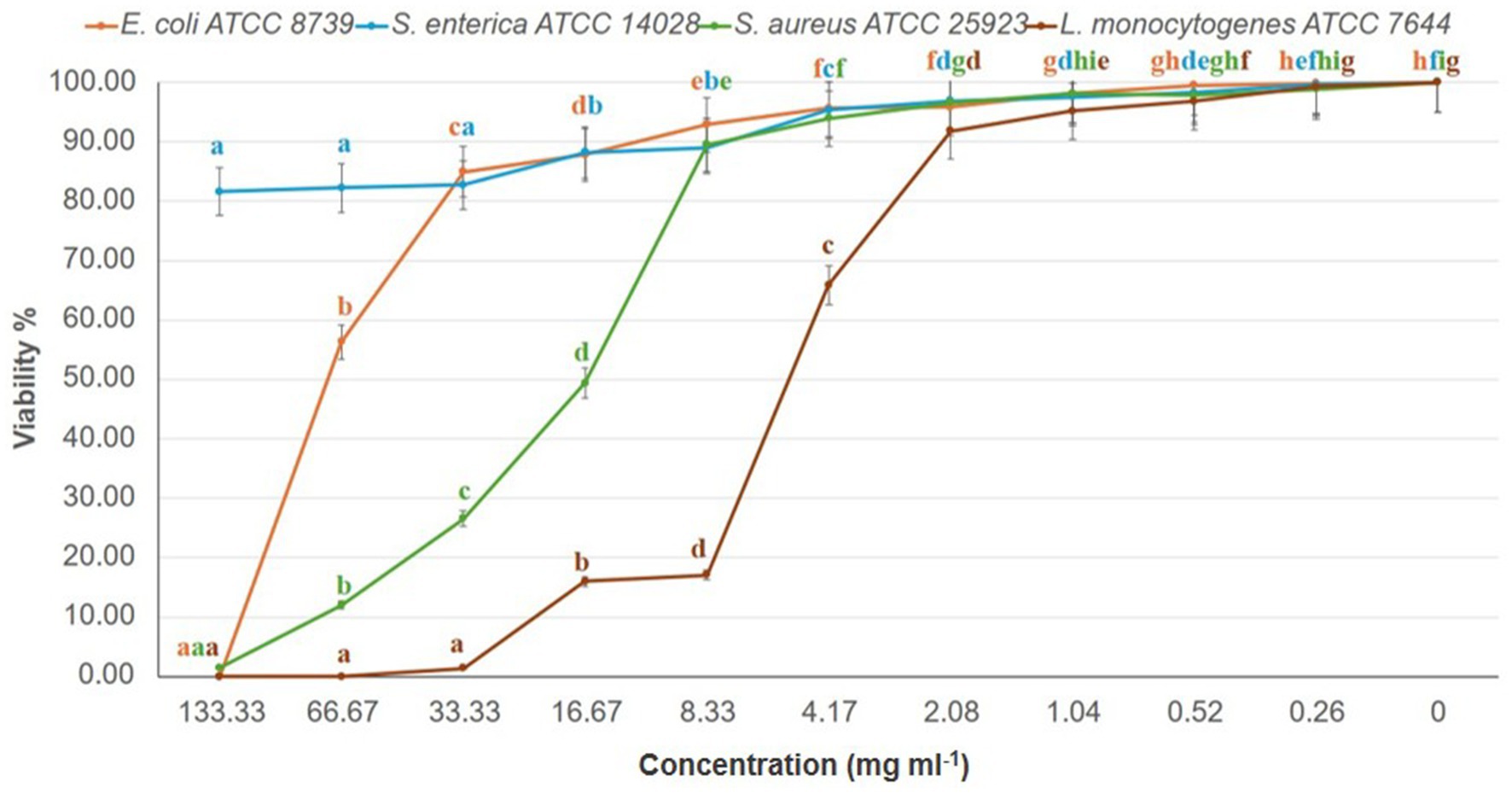
Figure 2. Dose–response effect of K. lappacea methanolic extract on the viability (%) of four bacterial reference strains—Salmonella enterica (ATCC 14028), Escherichia coli (ATCC 8739), Staphylococcus aureus (ATCC 25923), and Listeria monocytogenes (ATCC 7644)—assessed using the broth dilution assay. Extract concentrations tested were 133, 66.7, 33.3, 16.7, and 8.3 mg/mL. Error bars represent standard deviations from three independent experiments (n = 3). Means with different letters are significantly different (p < 0.0001) according to one-way ANOVA followed by Duncan’s post hoc test.
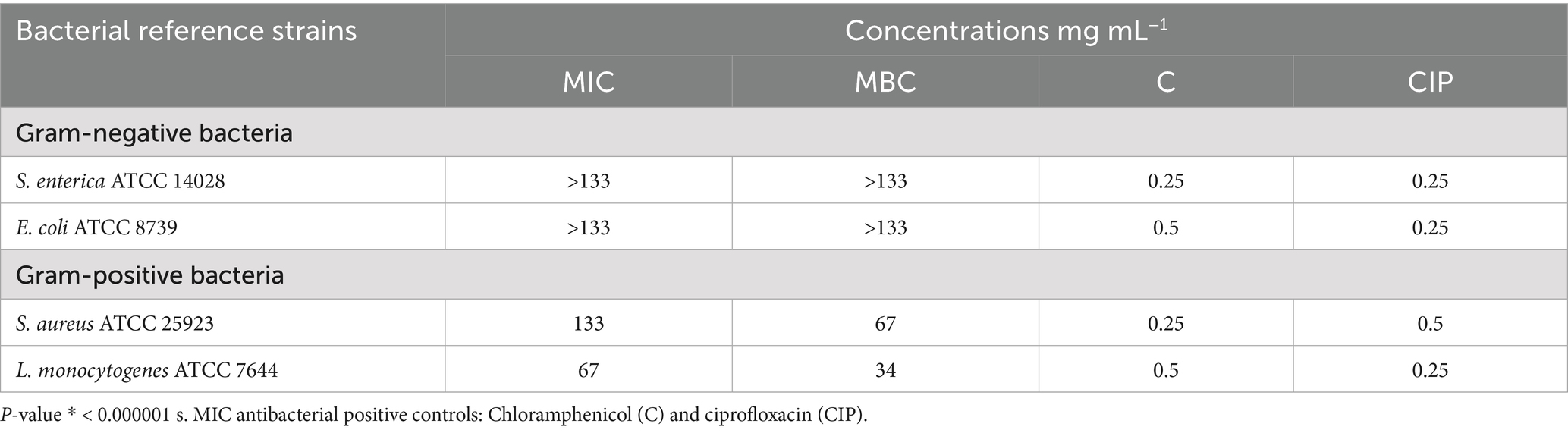
Table 3. Minimum inhibitory concentrations (MICs) and minimum bactericidal concentrations (MBCs) of the extract.
The cytotoxic activity of the K. lappacea extract was evaluated against A549 (lung carcinoma) and MCF7 (breast carcinoma) cell lines using the MTT assay (Figure S1; Table 4), showing dose-dependent inhibition of cell viability. The extract demonstrated higher cytotoxicity against MCF7 (IC₅₀ = 64.81 ± 0.26 μg/mL) than A549 (IC₅₀ = 142.27 ± 1.02 μg/mL), whereas doxorubicin—a widely used chemotherapeutic—exhibited substantially greater potency at much lower concentrations. This highlights the relatively moderate activity of the extract compared to established anticancer agents. In line with our results, Al-Oqail (2021) reported that the extract demonstrated cytotoxicity against MCF-7 breast cancer cells, with IC₅₀ values between 8.883 and 3.17 μM. These results align with our study’s observations and provide a broader context for understanding the therapeutic potential of K. lappacea extracts. The cytotoxic effects of the extract are attributed to flavonoids (epicatechin, fisetin) and phenolics (chlorogenic acid), which exert their activity through multiple mechanistic pathways: induction of apoptosis via caspase activation, mitochondrial dysfunction with loss of membrane potential and cytochrome c release, generation of reactive oxygen species (ROS) causing oxidative stress, interference with cell cycle progression, and inhibition of oncogenic signaling pathways such as PI3K/Akt and MAPK (Govarthanan et al., 2016; Prakash et al., 2019; Miao and Xiang, 2020; Zhou et al., 2023). These mechanisms collectively reduce proliferation and promote selective cancer cell death. Importantly, while the extract demonstrates only moderate potency compared with doxorubicin and reference antibiotics (e.g., ampicillin and vancomycin), such findings underscore the potential of exploring synergistic strategies. Previous studies have shown that combining phytochemicals with conventional chemotherapeutics or antibiotics can enhance therapeutic efficacy, reduce required drug dosages, and overcome resistance mechanisms (Wesseling and Martin, 2022). Future investigations should therefore assess whether K. lappacea root extract, or its bioactive constituents, can act synergistically with standard anticancer agents like doxorubicin or with frontline antibiotics. Such studies would not only clarify the clinical relevance of our findings but also open new avenues for the rational development of combination therapies.
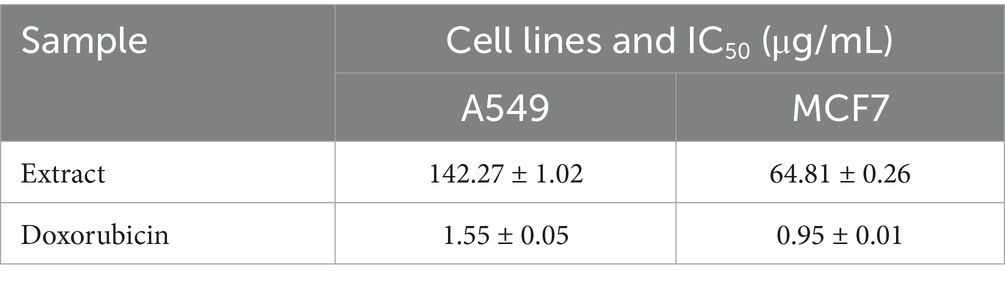
Table 4. IC50 of the studied extract.
Overall, LC-ESI-MS analysis links specific alkaloids and flavonoids to their reported antibacterial and cytotoxic mechanisms, providing a comprehensive rationale for the extract’s multifactorial biological activity. These findings reinforce K. lappacea’s potential as a source of bioactive compounds for antimicrobial and anticancer applications, while highlighting that efficacy is influenced by phytochemical composition, target cell type, and the inherent sensitivity of bacterial or cancer cells relative to established reference agents like doxorubicin and standard antibiotics.
4 Conclusion
Preliminary in vitro findings indicate that K. lappacea may possess bacteriostatic activity against Gram-positive bacteria and exhibit cytotoxic effects on human lung and breast (MCF7) cancer cell lines. However, subsequent studies will focus on identifying precise molecular targets (e.g., bacterial cell wall biosynthesis, efflux pump inhibition, apoptosis, or cell cycle arrest in cancer cells) and delineating signaling pathways through proteomic, transcriptomic, and molecular docking approaches.
Impact statement
Krameria lappacea demonstrates promising bacteriostatic activity against Gram-positive bacteria, suggesting potential as a complementary or alternative strategy to conventional antibiotics. It also exhibits cytotoxic effects against human lung (A549) and breast (MCF7) cancer cells, supporting its relevance as a source of bioactive compounds with anticancer potential. While further studies are required to elucidate its mechanisms of action and validate therapeutic applications, the consistency between its traditional use and the present findings highlights its potential role in developing novel antimicrobial and anticancer agents.
Data availability statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Author contributions
RA-G: Investigation, Writing – review & editing, Funding acquisition, Validation, Formal analysis, Supervision, Methodology, Data curation, Resources, Visualization, Software, Conceptualization, Project administration, Writing – original draft. MA: Investigation, Writing – original draft, Resources, Funding acquisition, Visualization, Formal analysis, Software, Validation, Conceptualization, Data curation, Project administration, Supervision, Writing – review & editing, Methodology. MD: Resources, Project administration, Formal analysis, Writing – original draft, Funding acquisition, Validation, Visualization, Data curation, Writing – review & editing, Supervision, Investigation, Conceptualization, Software, Methodology. NA: Software, Investigation, Supervision, Methodology, Funding acquisition, Writing – review & editing, Writing – original draft, Conceptualization, Project administration, Formal analysis, Visualization, Resources, Data curation, Validation. KA: Project administration, Writing – review & editing, Validation, Visualization, Supervision, Data curation, Methodology, Writing – original draft, Formal analysis, Conceptualization, Software, Investigation, Resources, Funding acquisition. DD: Project administration, Conceptualization, Funding acquisition, Validation, Writing – review & editing, Software, Resources, Formal analysis, Supervision, Writing – original draft, Methodology, Data curation, Visualization, Investigation. SQ: Conceptualization, Methodology, Supervision, Validation, Software, Data curation, Investigation, Visualization, Resources, Writing – review & editing, Funding acquisition, Project administration, Writing – original draft, Formal analysis. EA-S: Writing – review & editing, Writing – original draft, Software, Data curation, Methodology, Investigation, Visualization, Resources, Project administration, Funding acquisition, Validation, Conceptualization, Formal analysis, Supervision.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. The work was funded Ongoing Research Funding Program (ORF-2025-1079), King Saud University, Riyadh, Saudi Arabia.
Acknowledgments
This study was supported by the Ongoing Research Funding Program (ORF-2025-1079), King Saud University, Riyadh, Saudi Arabia.
Conflict of interest
DD was employed by company Boehringer Ingelheim Pharma GmbH & Co. KG.
The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The handling editor FH declared a shared affiliation with the authors RA-G, MA, NA, EA-S, SQ at the time of review.
Generative AI statement
The authors declare that no Gen AI was used in the creation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2025.1662564/full#supplementary-material
References
Abdel-Gaber, R., Alamari, G., Dkhil, M. A., Meryk, A., Al-Shaebi, E. M., and Al-Quraishy, S. (2024). Krameria lappacea root extract’s anticoccidial properties and coordinated control of CD4 T cells for IL-10 production and antioxidant monitoring. Front. Immunol. 15:1404297. doi: 10.3389/fimmu.2024.1404297
Alamari, G., Abdel-Gaber, R., Al-Shaebi, E. M., and Al-Quraishy, S. (2024). Anticoccidial and jejunum-protective effects of Krameria lappacea roots extract on experimental Eimeria papillata infection. Microsc. Res. Tech. 87, 1467–1478. doi: 10.1002/jemt.24531
Aldakheel, R. K., Gondal, M. A., Nasr, M. M., Dastageer, M. A., and Almessiere, M. A. (2021). Quantitative elemental analysis of nutritional, hazardous and pharmacologically active elements in medicinal Rhatany root using laser induced breakdown spectroscopy. Arab. J. Chem. 14:102919. doi: 10.1016/j.arabjc.2020.102919
Al-Fuhaid, N. (2018). Insecticidal activities of Phyllanthus emblica, Prunus mahaleb, Cerasus mahaleb, Piper nigrum, Krameria lappacea and Phoenix dactylifera on larvae Trogoderma granarium everts. Int. J. Agric. For. 8, 53–59. doi: 10.5923/j.ijaf.20180802.01
Al-Oqail, M. M. (2021). Anticancer efficacies of Krameria lappacea extracts against human breast cancer cell line (MCF-7): role of oxidative stress and ROS generation. Saudi Pharm. J. 29, 244–251. doi: 10.1016/j.jsps.2021.01.008
Al-Sallami, D., Alsultan, A., Abbas, K. H., and Clarke, S. R. (2023). Evaluation of efflux pump inhibitory activity of some plant extracts and using them as adjuvants to potentiate the inhibitory activity of some antibiotics against Staphylococcus aureus. Open Vet. J. 13, 42–47. doi: 10.5455/OVJ.2023.v13.i1.5
Andrade, S. F., Cardoso, L. G., and Bastos, J. K. (2007). Anti-inflammatory and antinociceptive activities of extract, fractions and populonic acid from bark wood of Austroplenckia populnea. J. Ethnopharmacol. 109, 464–471. doi: 10.1016/j.jep.2006.08.023
Ansari, N., Yadav, D. S., Singh, P., Agrawal, M., and Agrawal, S. B. (2023). Ozone exposure response on physiological and biochemical parameters Vis-à-Vis secondary metabolites in a traditional medicinal plant Sida cordifolia L. Ind. Crop. Prod. 194:116267. doi: 10.1016/j.indcrop.2023.116267
Ardhany, S. D., Fansuri, M. A., and Novaryatiin, S. (2022). Pharmacognostic study of Bawang Dayak (Eleutherine bulbosa (mill.) Urb.) and its clay mask against acne-causing bacteria. Trop. J. Nat. Prod. Res. 6, 1614–1621. doi: 10.26538/tjnpr/v6i10.9
Babiker, A. A. E., Mohammed, S. O., Al-Asiri, H. A., Al-Humaid, F. K., and Al-Subaihi, W. M. (2020). Antimicrobial activity of some plant extracts against MRSA. Glob. Sci. J. 8, 231–238.
Balouiri, M., Sadiki, M., and Ibnsouda, S. K. (2016). Methods for in vitro evaluating antimicrobial activity: a review. J. Pharm. Anal. 6, 71–79. doi: 10.1016/j.jpha.2015.11.005
Bombardelli, E., Morazzoni, P., Cristoni, A., Seghizzi, R., and Indena, S. A. (2002). Pharmaceutical and cosmetic formulations with antimicrobial activity (U.S. patent no. 6,475,536)
Bouarab-Chibane, L., Forquet, V., Lantéri, P., Clément, Y., Léonard-Akkari, L., Oulahal, N., et al. (2019). Antibacterial properties of polyphenols: characterization and QSAR (quantitative structure-activity relationship) models. Front. Microbiol. 10:829. doi: 10.3389/fmicb.2019.00829
Carini, M., Aldini, G., Orioli, M., and Facino, R. M. (2002). Antioxidant and photoprotective activity of a lipophilic extract containing neolignans from Krameria triandra roots. Planta Med. 68, 193–197. doi: 10.1055/s-2002-23167
Chaachouay, N., and Zidane, L. (2024). Plant-derived natural products: a source for drug discovery and development. Drugs Drug Cands. 3, 184–207. doi: 10.3390/ddc3010011
Chung, Y. C., Song, S. J., Lee, A., Jang, C. H., Kim, C. S., and Hwang, Y. H. (2024). Isobavachin, a main bioavailable compound in Psoralea corylifolia, alleviates lipopolysaccharide-induced inflammatory responses in macrophages and zebrafish by suppressing the MAPK and NF-κB signaling pathways. J. Ethnopharmacol. 321:117501. doi: 10.1016/j.jep.2023.117501
CLSI (2012). Methods for dilution antimicrobial susceptibility tests for bacteria that grow aerobically, approved standard. 9th Edn. Wayne, PA: Clinical and Laboratory Standards Institute CLSI document M07-A9.
CLSI (2019). Performance standards for antimicrobial susceptibility testing. 29th Edn. Wayne, PA: Clinical and Laboratory Standards Institute.
Deng, Z., Hassan, S., Rafiq, M., Li, H., He, Y., Cai, Y., et al. (2020). Pharmacological activity of eriodictyol: the major natural polyphenolic flavanone. Evid. Based Complement. Alternat. Med. 2020:6681352. doi: 10.1155/2020/6681352
Duffey, M., Jumde, R. P., Costa, R. M. A., Ropponen, H. K., Blasco, B., and Piddock, L. J. V. (2024). Extending the potency and lifespan of antibiotics: inhibitors of gram-negative bacterial efflux pumps. ACS Infect. Dis. 10, 1458–1482. doi: 10.1021/acsinfecdis.4c00091
Elmongy, E. I., Negm, W. A., Elekhnawy, E., El-Masry, T. A., Attallah, N. G. M., Altwaijry, N., et al. (2022). Antidiarrheal and antibacterial activities of Monterey cypress phytochemicals: in vivo and in vitro approach. Molecules 27:346. doi: 10.3390/molecules27020346
Eloff, J. N. (2019). Avoiding pitfalls in determining the antimicrobial activity of plant extracts and publishing the results. BMC Complement. Altern. Med. 19, 1–8. doi: 10.1186/s12906-019-2519-3
El-Shouny, W. A., Ali, S. S., Sun, J., Samy, S. M., and Ali, A. (2018). Drug resistance profile and molecular characterization of extended spectrum beta-lactamase (Escherichia coli)-producing Pseudomonas aeruginosa isolated from burn wound infections. Microb. Pathog. 116, 301–312. doi: 10.1016/j.micpath.2018.02.005
European Committee on Antimicrobial Susceptibility Testing (EUCAST) (2003). Determination of minimum inhibitory concentrations (MICs) of antibacterial agents by broth microdilution. EUCAST Discussion Document E. Def 5.1. Clin. Microbiol. Infect. 9, 1–7.
Genovese, C., D’Angeli, F., Bellia, F., Distefano, A., Spampinato, M., Attanasio, F., et al. (2021). In vitro antibacterial, anti-adhesive and anti-biofilm activities of Krameria lappacea (Dombey) Burdet & B.B. Simpson root extract against methicillin-resistant Staphylococcus aureus strains. Antibiotics (Basel) 10:428. doi: 10.3390/antibiotics10040428
Gholizadeh, S. S., Baserisalehi, M., and Bahador, N. (2013). Study on bioactive compounds produced by soil origin Brevibacillus spp. Nat. Environ. Pollut. Technol. 12, 209–214.
Gonelimali, F. D., Lin, J., Miao, W., Xuan, J., Charles, F., and Chen, M. (2018). Antimicrobial properties and mechanism of action of some plant extracts against food pathogens and spoilage microorganisms. Front. Microbiol. 9:1639. doi: 10.3389/fmicb.2018.01639
Govarthanan, M., Mythili, R., Selvankumar, T., Kamala-Kannan, S., Rajasekar, A., and Chang, Y. C. (2016). Bioremediation of heavy metals using an endophytic bacterium Paenibacillus sp. RM isolated from the roots of Tridax procumbens. 3 Biotech 6:242. doi: 10.1007/s13205-016-0560-1
Gu, Y., Dong, J., Li, J., Luo, Q., Dong, X., Tang, G., et al. (2023). Antibacterial activity and mechanism of sanguinarine against Staphylococcus aureus by interfering with the permeability of the cell wall and membrane and inducing bacterial ROS production. Front. Vet. Sci. 10:1121082. doi: 10.3389/fvets.2023.1121082
Heiss, E. H., Baumgartner, L., Schwaiger, S., Heredia, R. J., Atanasov, A. G., Rollinger, J. M., et al. (2012). Ratanhiaphenol III from Ratanhiae radix is a PTP1B inhibitor. Planta Med. 78, 678–681. doi: 10.1055/s-0031-1298242
Hochma, E., Yarmolinsky, L., Khalfin, B., Nisnevitch, M., Ben-Shabat, S., and Nakonechny, F. (2021). Antimicrobial effect of phytochemicals from edible plants. PRO 9:2089. doi: 10.3390/pr9112089
Horne, J. E., Brockwell, D. J., and Radford, S. E. (2020). Role of the lipid bilayer in outer membrane protein folding in gram-negative bacteria. J. Biol. Chem. 295, 10340–10367. doi: 10.1074/jbc.REV120.011473
Ilaghi, M., Sharifi, I., Sharififar, F., Sharifi, F., Oliaee, R. T., Babaei, Z., et al. (2021). The potential role and apoptotic profile of three medicinal plant extracts on Leishmania tropica by MTT assay, macrophage model and flow cytometry analysis. Parasite Epidemiol. Control 12, e00201–e00208. doi: 10.1016/j.parepi.2021.e00201
Inouye, S., Yamaguchi, H., and Takizawa, T. (2001). Screening of the antibacterial effects of a variety of essential oils on respiratory tract pathogens, using a modified dilution assay method. J. Infect. Chemother. 7, 251–254. doi: 10.1007/s101560170022
Khanam, S., Mishra, P., Faruqui, T., Alam, P., Albalawi, T., Siddiqui, F., et al. (2025). Plant-based secondary metabolites as natural remedies: a comprehensive review on terpenes and their therapeutic applications. Front. Pharmacol. 16:1587215. doi: 10.3389/fphar.2025.1587215
Ladurner, A., Atanasov, A. G., Heiss, E. H., Baumgartner, L., Schwaiger, S., Rollinger, J. M., et al. (2012). 2-(2, 4-dihydroxyphenyl)-5-(E)-propenylbenzofuran promotes endothelial nitric oxide synthase activity in human endothelial cells. Biochem. Pharmacol. 84, 804–812. doi: 10.1016/j.bcp.2012.06.029
Liu, Y., Zhu, J., Liu, Z., Zhi, Y., Mei, C., and Wang, H. (2025). Flavonoids as promising natural compounds for combating bacterial infections. Int. J. Mol. Sci. 26:2455. doi: 10.3390/ijms26062455
Maciel, M. A. M., Pinto, A. C., Veiga, V. F., Grynberg, N. F., and Echevarria, A. (2002). Plantas medicinais: a necessidade de estudos multidisciplinares. Quim Nova 25, 429–438. doi: 10.1590/S0100-40422002000300016
Malik, E. M., and Müller, C. E. (2016). Anthraquinones as pharmacological tools and drugs. Med. Res. Rev. 36, 705–748. doi: 10.1002/med.21391
Manikandan, P., Letchoumy, P. V., Gopalakrishnan, M., and Nagini, S. (2008). Evaluation of Azadirachta indica leaf fractions for in vitro antioxidant potential and in vivo modulation of biomarkers of chemoprevention in the hamster buccal pouch carcinogenesis model. Food Chem. Toxicol. 46, 2332–2343. doi: 10.1016/j.fct.2008.03.013
Mendes, C. R., Dilarri, G., Forsan, C. F., Sapata, V. M. R., Lopes, P. R. M., de Moraes, P. B., et al. (2022). Antibacterial action and target mechanisms of zinc oxide nanoparticles against bacterial pathogens. Sci. Rep. 12:2658. doi: 10.1038/s41598-022-06657-y
Miao, M., and Xiang, L. (2020). Pharmacological action and potential targets of chlorogenic acid. Adv. Pharmacol. 87, 7–88. doi: 10.1016/bs.apha.2019.12.002
Minnelli, C., Mangiaterra, G., Laudadio, E., Citterio, B., and Rinaldi, S. (2024). Investigation on the synergy between membrane permeabilizing amphiphilic α-hydrazido acids and commonly used antibiotics against drug-resistant bacteria. Molecules 29:4078. doi: 10.3390/molecules29174078
Moon, P. D., Choi, I. H., and Kim, H. M. (2011). Naringenin suppresses the production of thymic stromal lymphopoietin through the blockade of RIP2 and caspase-1 signal cascade in mast cells. Eur. J. Pharmacol. 671, 128–132. doi: 10.1016/j.ejphar.2011.09.163
Mosmann, T. (1983). Rapid colorimetric assay for cellular growth and survival: application to proliferation and cytotoxicity assays. J. Immunol. Methods 65, 55–63. doi: 10.1016/0022-1759(83)90303-4
Nayf, E. M., and Salman, H. A. (2021). Antibacterial activity of aquatic extract of Myrtus communis leaves against periodontitis isolated bacteria. IOP Conf. Ser. Earth Environ. Sci. 880:012047. doi: 10.1088/1755-1315/880/1/012047
Ortiz, S., Lecsö-Bornet, M., Bonnal, C., Houze, S., Michel, S., Grougnet, R., et al. (2019). Bioguided identification of triterpenoids and neolignans as bioactive compounds from anti-infectious medicinal plants of the Taira Atacama’s community (Calama, Chile). J. Ethnopharmacol. 231, 217–229. doi: 10.1016/j.jep.2018.10.029
Prakash, M., Basavaraj, B. V., and Murthy, K. N. C. (2019). Biological functions of epicatechin: plant cell to human cell health. J. Funct. Foods 52, 14–24. doi: 10.1016/j.jff.2018.10.021
Robinson, M. M., and Zhang, X. (2011). “Traditional medicines: global situation, issues and challenges” in The world medicines situation. 3rd ed (Geneva: WHO), 1–14.
Safta, D. A., Ielciu, I., Șuștic, R., Hanganu, D., Niculae, M., Cenariu, M., et al. (2023). Chemical profile and biological effects of an herbal mixture for the development of an oil-in-water cream. Plants 12:248. doi: 10.3390/plants12020248
Salem, S. S., Elsayed, H. E., Shabana, S., Khazaal, M. T., and Moharram, F. A. (2023). Phytochemical profile and antimicrobial activity of essential oils from two Syzygium species against selected oral pathogens. BMC Complement Med. Ther. 23:448. doi: 10.1186/s12906-023-04277-1
Scholz, E., and Rimpler, H. (1989). Proanthocyanidins from Krameria triandra root. Planta Med. 55, 379–384. doi: 10.1055/s-2006-962032
Shirley, M. W., and Lillehoj, H. (2012). The long view: a selective review of 40 years of coccidiosis research. Avian Pathol. 41, 111–121. doi: 10.1080/03079457.2012.666338
Simpson, B. B. (1991). The past and present uses of Rhatany (Krameria, Krameriaceae). Econ. Bot. 45, 397–409. doi: 10.1007/BF02887080
Simpson, B. B.Krameriaceae (1989). Flora neotropica, vol. 49. New York: New York Botanical Garden Press, 1–108.
Wang, C., Fan, R. Q., Zhang, Y. X., Nie, H., and Li, K. (2016). Naringenin protects against isoniazid- and rifampicin-induced apoptosis in hepatic injury. World J. Gastroenterol. 22, 9775–9783. doi: 10.3748/wjg.v22.i44.9775
Wesseling, C. M., and Martin, N. I. (2022). Synergy by perturbing the gram-negative outer membrane: opening the door for gram-positive specific antibiotics. ACS Infect. Dis. 8, 1731–1757. doi: 10.1021/acsinfecdis.2c00193
Yan, Y., Li, X., Zhang, C., Lv, L., Gao, B., and Li, M. (2021). Research progress on antibacterial activities and mechanisms of natural alkaloids: a review. Antibiotics 10:318. doi: 10.3390/antibiotics10030318
Zabka, M. (2022). Antifungal efficacy and convenience of Krameria lappacea for the development of botanical fungicides and new alternatives of antifungal treatment. Agronomy 12:2599. doi: 10.3390/agronomy12112599
Zhang, Z., Cao, M., Shang, Z., Xu, J., Chen, X., Zhu, Z., et al. (2025). Research progress on the antibacterial activity of natural flavonoids. Antibiotics 14:334. doi: 10.3390/antibiotics14040334
Keywords: Krameria lappacea, phytochemical screening, antibacterial activity, gram-bacteria, cytotoxicity
Citation: Abdel-Gaber R, Albeshr M, Dkhil MA, Almohawis N, Abdelmeguid KA, Delic D, Al Quraishy S and Al-Shaebi EM (2025) Antibacterial activity of Krameria lappacea root extract against gram-positive and gram-negative bacteria and its cytotoxicity on lung and breast cancer cell lines. Front. Microbiol. 16:1662564. doi: 10.3389/fmicb.2025.1662564
Edited by:
Fohad Mabood Husain, Department of Food Science and Nutrition, King Saud University, Saudi ArabiaReviewed by:
Ekhtear Hossain, Southern University and A&M College, United StatesFatimah Salim, Atta-ur-Rahman Institute for Natural Product Discovery (AuRIns), Malaysia
Rohit Ruhal, VIT University, India
Copyright © 2025 Abdel-Gaber, Albeshr, Dkhil, Almohawis, Abdelmeguid, Delic, Al Quraishy and Al-Shaebi. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Rewaida Abdel-Gaber, cmFiZGVsZ2FiZXIuY0Brc3UuZWR1LnNh