
 Rohit Sachdeva1,2
Rohit Sachdeva1,2
- 1 Laboratory of Molecular Neurogenetics, Department of Experimental Medical Science, Wallenberg Neuroscience Center, Lund University, Lund, Sweden
- 2 Lund Stem Cell Center, Lund University, Lund, Sweden
In adult mammals, neural stem cells (NSCs) are found in two niches of the brain; the subventricular zone by the lateral ventricles and the subgranular zone of the dentate gyrus in the hippocampus. Neurogenesis is a complex process that is tightly controlled on a molecular level. Recently, microRNAs (miRNAs) have been implicated to play a central role in the regulation of NCSs. miRNAs are small, endogenously expressed RNAs that regulate gene expression at the post-transcriptional level. However, functional studies of miRNAs are complicated due to current technical limitations. In this review we describe recent findings about miRNAs in NSCs looking closely at miR-124, miR-9, and let-7. In addition, we highlight technical strategies used to investigate miRNA function, accentuating limitations, and potentials.
Introduction
Neural stem cells (NSCs) are multipotent cells that can give rise to various cell types in the central nervous system (CNS), including neurons, astrocytes, and oligodendrocytes (Alvarez-Buylla et al., 2002). In adult mammals, neurogenesis is limited to two brain regions; the subventricular zone (SVZ) by the lateral ventricles and the subgranular zone (SGZ) of the dentate gyrus in the hippocampus (reviewed in Zhao et al., 2008). NSCs in the SVZ, so called type-B cells, are of astrocytic lineage and generate transient amplifying type-C cells. Those in turn differentiate into migrating neuroblasts, type-A cells. Neuroblasts then migrate along a defined pathway, the rostral migratory stream, to mature and form interneurons in the olfactory bulb (Doetsch et al., 1999). NSCs of the SGZ are radial glia-like type 1 cells. They generate type 2 intermediate proliferating progenitors that subsequently give rise to type 3 neuroblasts migrating into the subgranular layer, to become mature functional glutamatergic granule neurons (Kempermann et al., 2004).
Although the functional role of adult neurogenesis still is debated, it is undoubtedly a complex and highly regulated process. Recently, microRNAs (miRNAs) have been suggested as players of neurogenesis, controlling expression of key regulatory genes (Gao, 2010; Shi et al., 2010; Luikart et al., 2011). miRNAs are one of the central determinants of mRNA abundance, functioning as standby mediators of mRNA regulation. They are small (21–23 nucleotides long), non-coding endogenously expressed RNA that bind to complementary mRNA targets, resulting in a decrease in target mRNA activity (Bartel, 2009).
Since the discovery of the first miRNA in 1993, they have been identified in animals, plants, and viruses and more than 1000 miRNA sequences have so far been found in humans (www.mirbase.org). Many are conserved across species. Some have a general expression pattern, others are specifically expressed in certain tissues or cell types and expression can be spatially and temporally restricted. Several labs have utilized conditional deletion of Dicer, a key enzyme in miRNA biogenesis, as a tool to study the role of miRNAs in neurogenesis. Disruption of the function of Dicer during brain development results in gross anatomical changes and in some cases it is embryonic lethal (Giraldez et al., 2005; Davis et al., 2008; De Pietri Tonelli et al., 2008; Kawase-Koga et al., 2009). In vitro, NSCs can be cultured in the absence of Dicer but are unable to generate neurons or astrocytes upon differentiation (Andersson et al., 2010). Although, Dicer knockout (KO) studies indicate that miRNAs are important regulators of neurogenesis and NSC self-renewal, they are difficult to interpret. Not only because the simultaneous deficiency of all miRNAs, but the long half-life of mature miRNA makes conditional Dicer KO experiments difficult to control from a temporal aspect, moreover there are concerns that non-miRNA related functions of Dicer contribute to the observed phenotypes (Konopka et al., 2010; Kaneko et al., 2011). For example, when Dicer was conditionally deleted in the retinal pigmented epithelium, cell death occurred via a mechanism that is independent on miRNA, rather depending on the accumulation of retrotransposon transcripts (Kaneko et al., 2011). With this in mind, studies of the functional role of individual miRNAs are a necessity.
In 2007 there were four simultaneous reports of miRNA KO mice (Rodriguez et al., 2007; Thai et al., 2007; van Rooij et al., 2007; Zhao et al., 2007), which enabled loss-of-function studies of specific miRNAs. However, for several miRNA, including many of those involved in neurogenesis, a classic KO strategy is complicated to apply due to single miRNA species being located in multiple copies in separate regions of the genome, in clusters or located within other genes. One example of such a miRNA family is miR-124, which is transcribed as three different primary transcripts from three independent loci. miR-124-1, miR-124-2, and miR-124-3 all produce the same mature miRNA (Griffiths-Jones, 2006). Another example is miR-9 which is expressed from three independent loci, miR-9-1, miR-9-2, and miR-9-3. The mature sequence is identical, and conserved in vertebrates and mammals. With let-7 the situation is even more complex with 12 human let-7 genes encoding for nine distinct, but closely related, mature forms of the miRNA (Griffiths-Jones, 2006).
Recently, a KO mouse for miR-124-1 was described showing that reduction of miR-124 has severe consequences for neuronal survival and axonal outgrowth (Sanuki et al., 2011). While this study demonstrates the importance of miR-124, it also highlights the problems of using a classic KO strategy. It is clear from the study by Sanuki et al. (2011) that compensation of miR-124-2 and miR-124-3 influence the phenotype of the miR-124-1 KO mouse. Likewise, a KO mice that have two of the three copies of miR-9 deleted has been reported (Shibata et al., 2011). These mice suffer from severe developmental effects of the brain. These two reports suggest that conditional deletion of all three copies of either miR-9 or miR-124 is necessary in order to fully understand the role of these miRNAs in adult neurogenesis. Although challenging and time consuming, such a strategy may be feasible for miR-9 and miR-124. However, for let-7 such a strategy appears unlikely to be successful given the large size of the let-7-family.
Adding to this, in situ analysis of the expression profile of individual miRNA is difficult due to the small size of the mature miRNA, which leads to poor resolution obtained in the brain with current histological techniques. As such, the study of the functional role of individual miRNAs in vivo is complicated and makes interpretation and comparison between different studies challenging.
Below, we highlight recent advances in the understanding of selected miRNAs in neurogenesis. We describe the current status of the field, existing attempts to study loss of miRNA function, and point out technical limitations that need to be circumvented in order to move the field forward.
Expression Profiling of miRNA in Neural Stem Cells
It is fairly straightforward to profile miRNA-expression patterns from bulk RNA samples, either at single species resolution using for example Northern blot or PCR-techniques, or at a global level using miRNA arrays, PCR-array, deep sequencing of small RNAs or other more specialized platforms. These methods all have their innate differences and parallel analysis of the same samples using different techniques may give significantly different results (see, e.g., Hebert and Nelson, 2011 for a discussion on this matter). Since there is currently no “gold standard” for transcriptional profiling of miRNA, the use of independent techniques to verify results is therefore necessary. Nevertheless, these approaches have revealed the complexity of miRNA-expression patterns among different cell types and have allowed identification of a number of candidate miRNAs that appear to be enriched in cultured NSCs. However, the technical difficulties of purifying populations of NSCs and progenitors from in vivo material, using for example fluorescence activated cell sorting, make these approaches problematic to transfer to the in vivo setting (see Table 1).
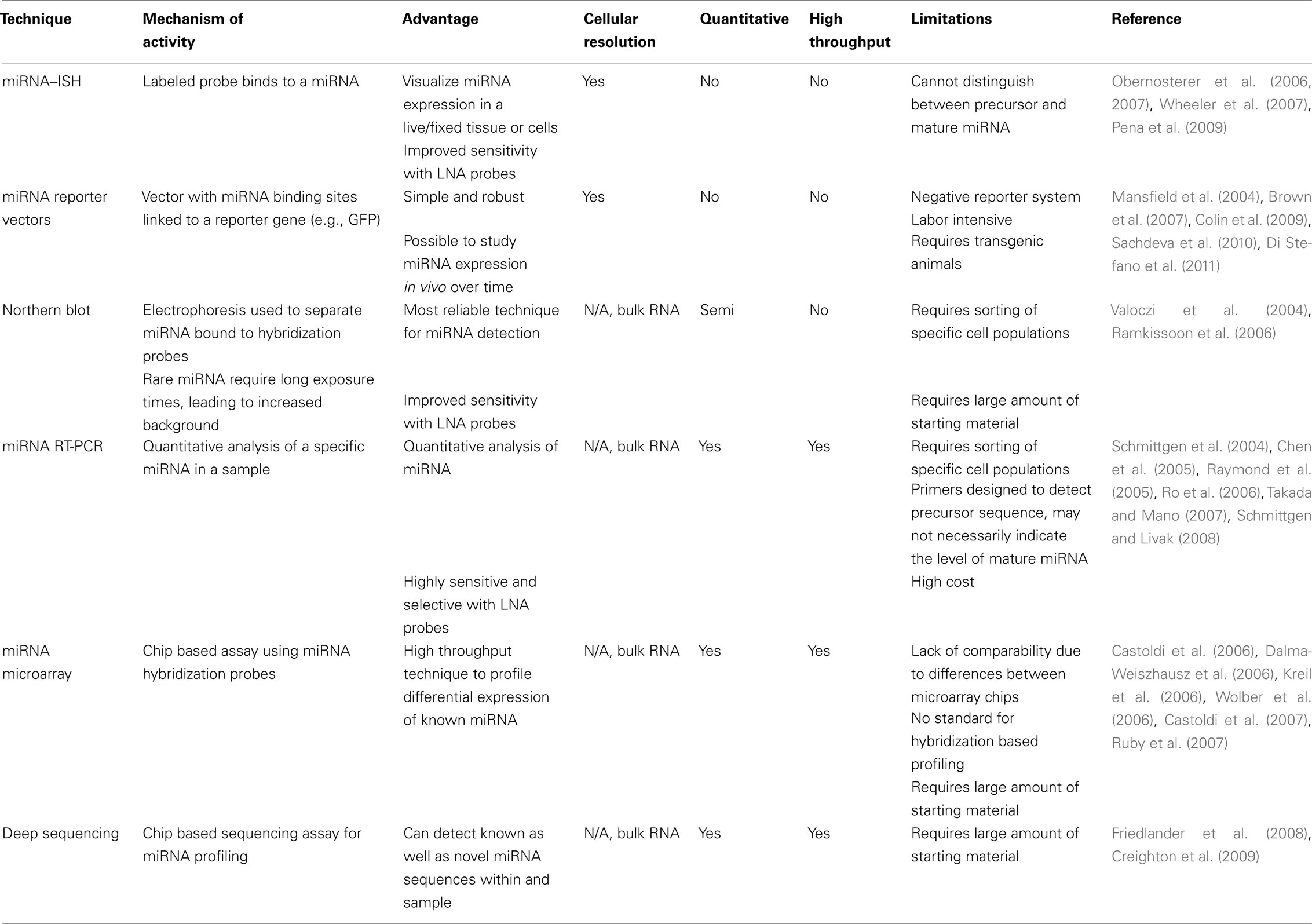
Table 1. Evaluation of different miRNA-visualization techniques.
Histological approaches to study miRNA expression in brain tissue have to a great extent relied on in situ hybridization (ISH) techniques. Due to the small size of the miRNA it is not possible to use standard ISH protocols; an additional fixation step of the miRNA is needed and probe hybridization must be optimal (Pena et al., 2009). Locked nucleic acid (LNA) modified oligonucleotides is preferable to use, since the melting temperature of the LNA probe/miRNA duplex is increased, resulting in stringent hybridization conditions, which in turn increases specificity and sensitivity (reviewed in Obernosterer et al., 2007; Wheeler et al., 2007). There are, however, challenges with ISH. First, discriminating between precursor and mature miRNA is difficult when using ISH. To do so, additional probes that target all the various precursor transcripts need to be used (Obernosterer et al., 2006). However, this can be technically challenging when analyzing miRNAs with multiple precursor transcripts (such as miR-9 or miR-124). Furthermore, the results from this method are of limited resolution, thereby making it difficult to distinguish between two adjacent cells. In addition, ISH is also problematic to use in combination with other labeling techniques that are routinely used in NSC-research. We have in our lab not been able to adopt protocols that allow the use of miRNA–ISH in combination with, for example BrdU-labeling, which is widely used in this field. This is primarily due to the stringent treatment of the tissue that is necessary for ISH, which is incompatible with the tissue treatments for BrdU-labeling. The problem of in situ miRNA-expression analysis is a major concern for the study of miRNA in the nervous system where it is essential to understand the cellular localization with regards to functionality.
More recently, miRNA reporter or sensor vectors have been used to visualize the expression pattern of endogenous miRNA in cells. These are gene transfer vectors that contain a reporter gene (i.e., GFP) along with binding sites for specific miRNA (Mansfield et al., 2004; Brown et al., 2007). In the case that a cell is actively expressing the specific miRNA, the expression of the reporter gene will be suppressed by the binding of the miRNA to the complimentary binding sites. Thus, this system reports the absence of the target miRNA, and cells that do not express it will be GFP-positive. This technique is highly specific, simple, and robust and makes it possible to study miRNA expression as cells differentiate. Injection of a miR-124 reporter vector into the brain will separate reporter gene expression between different cell types such as neurons and astrocytes (Colin et al., 2009). The system has been used to segregate differentiated neural cells in pluripotent cell cultures, based on the expression of miR-292 that is expressed in embryonic stem cells but not in NSCs (Sachdeva et al., 2010), as well as the opposite where a miRNA let-7a reporter was used to select undifferentiated cells from more differentiated cells (Di Stefano et al., 2011). In NSCs in vivo, sensor vectors have been used to track the expression of miR-132 in the DG (Luikart et al., 2011). The use of sensor vectors has the advantage that they measure the activity of the miRNA rather than expression per se allowing easy determination if the mature miRNA is present. Also, the use of a fluorescent reporter gene allows excellent morphological analysis of cells in vivo. Still, the technique is time-consuming including the generation of viral vectors followed by experimentation in cell culture or in vivo models. However, the versatility of the technique opens up the possibility of generating transgenic reporter animals, making it possible to visualize the expression pattern of a specific miRNA throughout an organism over time (see, e.g., Gentner et al., 2010). Although this approach remains to be tested in the CNS, it may be an attractive alternative in order to achieve sensitive, high-quality expression analysis of miRNAs in vivo.
Loss of Function Studies of miRNAs
As mentioned above, generation of KO mice for individual miRNAs is often complicated since many miRNAs are present in several copies or in clusters while some are present within introns of genes. To circumvent this issue several knockdown or inhibition approaches have been developed over the last few years (see Table 2).
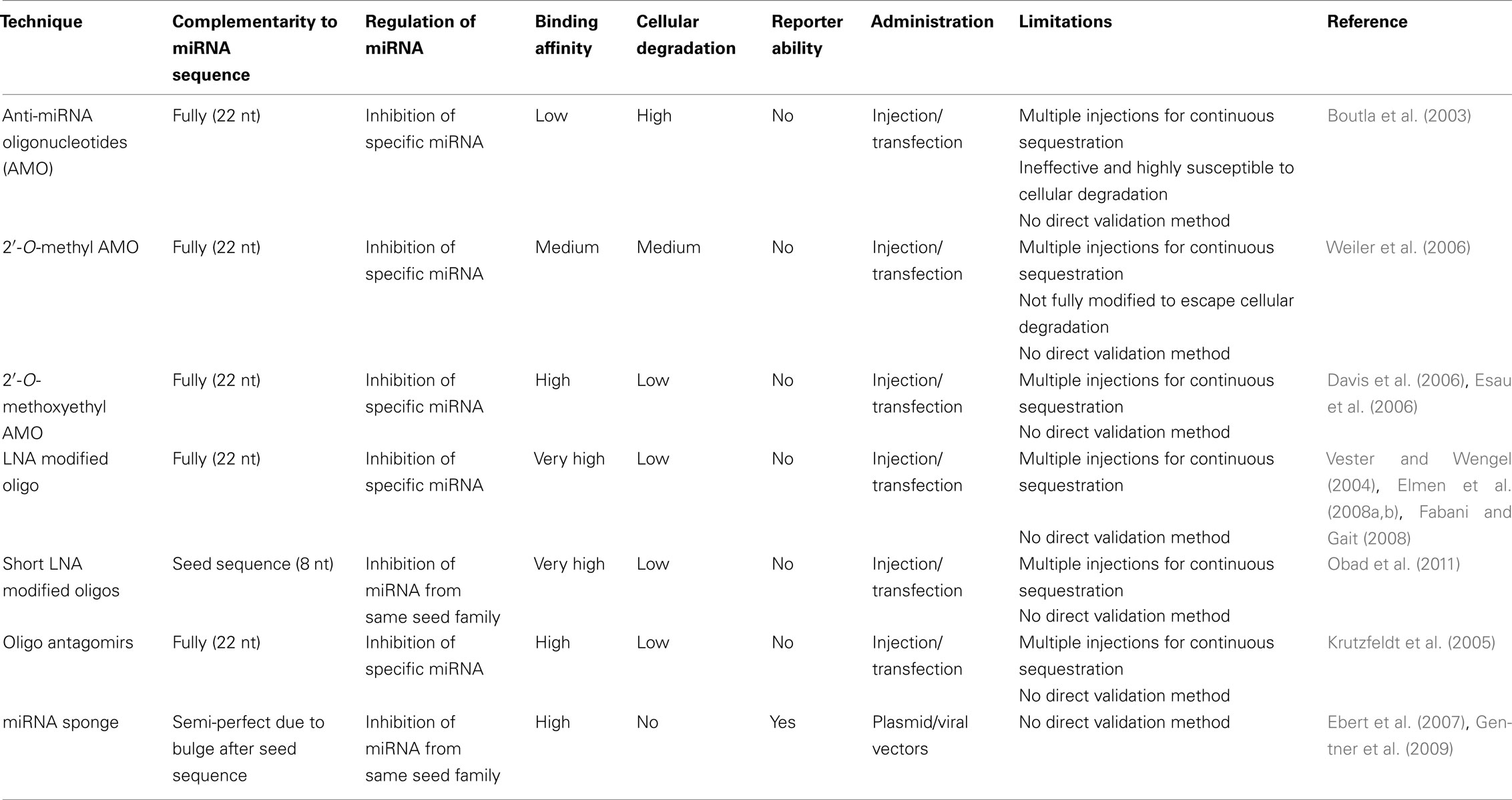
Table 2. Comparison of miRNA loss-of-function techniques.
An early approach to analyze specific miRNAs was by using anti-miRNA oligonucleotide (AMO) which are nucleic acids that are antisense to the miRNA, thus hindering the interaction between the miRNA and target mRNA. This technique was first used to inactivate miR-2 and miR-13 in Drosophila, in the search for miRNA-target genes (Boutla et al., 2003). It was soon found that unmodified oligonucleotides are ineffective since the cellular machinery degrades them. However there are various chemical modified variants that are efficient. 2′-O-methyl AMO is a simple chemical modification where the methyl group reduces the chances of endonucleolytic cleavage, and improves binding affinity to the miRNA (Weiler et al., 2006). Another variant is 2′-O-methoxyethyl AMO, which are fully modified oligonucleotides, with higher affinity and specificity than 2′-O-methyl AMO variants (Davis et al., 2006; Esau et al., 2006). Recently, LNA (locked nucleic acid) modified oligonucleotides allow for even further stabilization of the miRNA/target duplex structure improving silencing and also making it possible to the use small oligonucleotides that enables targeting of entire miRNA-families (Vester and Wengel, 2004; Obad et al., 2011).
Anti-miRNA oligonucleotide targets single miRNAs with high specificity, as they are completely complementary to the mature sequence of the miRNA (Boutla et al., 2003). Despite the successful knockdown of miRNA in vitro and in vivo using this method, it has several limitations. First, a direct measurement of the down regulation of miRNA is difficult, because AMO binds to the miRNA and sequesters it from its target rather than inducing its degradation (Krutzfeldt et al., 2005; Davis et al., 2009). Therefore the only possible way to confirm the decrease of miRNA is to use indirect methods, whereby one can measure the level of expression of a reporter gene containing a target sequence of the miRNA or analyzing upregulation of endogenous target genes. Secondly it is not possible to identify the cells in which the AMOs are active as they do not carry a reporter. On top of that, the level of AMO should preferably be kept constant to allow a continuous sequestration of the miRNA. These limitations make the technique complicated to use when targeting NSCs in vivo.
Oligonucleotide antagomirs are chemically modified; cholesterol conjugated single stranded RNA analogs complementary to a target miRNA (Krutzfeldt et al., 2005). The modification includes a partial phosphorothioate backbone in addition to 2′-O-methoxyethyl to inhibit Ago-2-mediated cleavage. In vivo, antagomirs have been given through intravenous injection where they appear to efficiently target miRNA in various tissues. However, antagomirs do not cross the blood brain barrier, which means that in the brain, antagomirs have the same limitations as AMOs have.
MicroRNA sponges are transcripts expressed from strong promoters, containing multiple, tandem binding sites to a selected member of the miRNA seed family of interest (Ebert et al., 2007). The binding site is imperfect, containing a bulge, for preventing RNA interference cleavage and degradation of the sponge RNA through endonucleolytic cleavage by Ago-2. The main advantage of miRNA sponges is the possibility to achieve stable expression from integrated transgenes in vivo (Gentner et al., 2009). This can be used for studying long-term effects of miRNA loss-of-function and also allows for stably expressing cell lines to be generated. The use of vector-mediated delivery also enables the incorporation of a reporter gene in order to identify the modified cells. Another advantage is that sponges complementarily bind to the seed sequence of the miRNA, which means that one sponge can target an entire family of miRNAs. In summary, these features make sponge vectors an attractive approach to study miRNA function in vivo in NSCs. Recently; this technology was employed to demonstrate that miR-132 affects the integration of new-born neurons in the adult hippocampus (Luikart et al., 2011). A previous report used a retrovirus expressing the cre-recombinase to delete a floxed miR-132 gene allowing a side-by-side comparison of sponge vs. conditional KO (Magill et al., 2010; Luikart et al., 2011). The sponge recapitulates some, but not all, defects detected following a complete deletion of miR-132 suggesting that the use of a sponge reduces rather than eliminates levels of miR-132 (Magill et al., 2010; Luikart et al., 2011). As with the use of AMOs, the sponge vectors also have other limitations, including the difficulty of validating the down regulation of a specific miRNA and the only possible way to confirm the decrease of miRNA is to use indirect methods as described above.
Taken together, it is evident that is technically challenging to perform loss-of-function studies of miRNAs in vivo in NSCs. The problem of validating the inhibition, the use of transient systems together with the appearance of potential “off-target” effects makes the interpretation of several studies challenging. This fact is highlighted by several studies coming to contradictory conclusions. An example of this are two studies performed in the developing chick, one report suggested that miR-124 plays no role in neurogenesis while another found that miR-124 modestly promotes neurogenesis (Cao et al., 2007; Visvanathan et al., 2007).
On the other hand, overexpression studies of miRNAs are quite simple to perform. Direct delivery of miRNA-duplexes or the use of various plasmid based approaches and viral vectors have been effectively used to overexpress miRNA. There are several relatively simple designs of vectors enabling stable expression that all appear to work efficiently (Krutzfeldt et al., 2006; Amendola et al., 2009). The statement that gain of function studies are easier to perform than loss of function studies is reflected in the literature and a great extent of the insight gained in the miRNA field is from overexpression studies.
In the last part of this review we discuss in detail three miRNAs that have been functionally implicated in neurogenesis; miR-9, miR-124, and the let-7 family. We give an overview of the current understanding of how these miRNAs influence neurogenesis and also highlight the technical shortcomings that still prevent a full understanding of the role of these miRNAs in vivo. Moreover we propose technical developments that are necessary for the field to move forward. In addition to the three above-mentioned miRNA, there is a growing literature of other miRNAs, including for example miR-125b, miR-132, miR-137, and miR-184 that influence neurogenesis (Le et al., 2009; Liu et al., 2010; Magill et al., 2010; Szulwach et al., 2010; Luikart et al., 2011; Sun et al., 2011). Several of the technical problems that limit our understanding of miR-9, miR-124, and the let-7 family also hold true for other miRNAs.
miR-124
miR-124 is perhaps the best characterized brain-specific miRNA, and accounts for 25–48% of all brain miRNA (Lagos-Quintana et al., 2002). miR-124 is expressed in neurons and have been proposed to suppress non-neural transcripts to promote neural identity (Visvanathan et al., 2007; De Pietri Tonelli et al., 2008; Maiorano and Mallamaci, 2009; Farrell et al., 2011; Liu et al., 2011a; Yoo et al., 2011). Several studies suggest that miR-124 is not expressed in other cells of the CNS such as astrocytes (see, e.g., Smirnova et al., 2005) while Ponomarev et al. (2010) also found it to be expressed in microglia and downregulated in activated microglia, being a key regulator of microglia quiescence. miR-124 is not expressed in NSCs but is suggested to regulate the temporal progression of neurogenesis in SVZ. It is upregulated in the transition between type-C and type-A cells and further upregulated as the neuroblasts exit the cell cycle (Cheng et al., 2009).
Delivery of miR-124 to mouse or human cells in vitro causes the global expression profile of mRNA to shift toward that of the brain. Lim et al. (2005) transfected miR-124 duplexes, Yu et al. (2008) used a DNA plasmid with primary miR-124 transcript whereas Yoo et al. (2011) delivered miR-124 with a lentiviral vector harboring a miR-124 precursor, all resulting in promoting a neural phenotype. In addition, transfection of miR-124 duplexes into glioblastoma cells inhibits proliferation and induces differentiation (Silber et al., 2008).
In vitro loss of function, by using antisense 2′-O-methyl AMO, results in delayed neurite outgrowth (Yu et al., 2008) and upregulation of non-neural transcripts (Conaco et al., 2006). Blocking miR-124 in SVZ cell populations in vivo by delivering antisense 2′-O-methyl AMO by a micro-osmotic pump into the ventricle, maintains neural progenitors as dividing precursors (Cheng et al., 2009). On the contrary, injecting a retrovirus overexpressing miR-124 into the SVZ promotes precocious neural maturation (Cheng et al., 2009). However, the in vivo role of miR-124 remains unclear and controversial. Although miR-124 has been reported to be an important regulator of neurogenesis both in the developing and the adult brain, contradictory findings have been published suggesting miR-124 to be less important for neurogenesis in the developing spinal cord (Cao et al., 2007; Visvanathan et al., 2007).
Based on in vitro miR-124 suppression or overexpression experiments, numerous miR-124 target genes have been found and validated. miR-124 has been shown to suppress several components of the RE1 silencing transcription factor (REST) pathway (Conaco et al., 2006; Wu and Xie, 2006). REST is a master regulator of neuronal phenotype (Lunyak and Rosenfeld, 2005) and together with co-repressors it recruits histone deacetylases to suppress non-neural genes. miR-124 and REST act reciprocally; miR-124 represses REST in neurons to promote expression of neural genes, whereas REST downregulates miR-124 in non-neural cells to inhibit expression of neural genes (Conaco et al., 2006). Another miR-124 target is Ptbp1 (Makeyev et al., 2007), a repressor of alternative splicing in non-neural cells demonstrating that miR-124 promotes a neuronal transcriptome by altering splicing. These targets suggest that miR-124 expression maintains a neuronal transcriptome by repressing non-neuronal genes at several levels. Other targets include Jagged1 in the Notch signaling pathway (Cheng et al., 2009; Liu et al., 2011b) and Sox9 that controls adult neurogenesis (Cheng et al., 2009; Grandjean et al., 2009; Farrell et al., 2011). These targets suggest that miR-124 also plays a role in regulating the exit of a self-renewing state of NSC. However, these are only a few examples since computational algorithms suggest more than 1000 miR-124 targets (Griffiths-Jones, 2006). Thus, the functional role of miR-124 is likely to be complex.
The large number of miR-124 target genes together with the observation that overexpression of miR-124 can induce a neuronal gene program suggest that miR-124 plays a crucial role in establishing and maintaining a neuronal transcription network. In light of this, it is surprising that the in vivo phenotypes found when blocking miR-124 using antisense technology is subtle, characterized mainly by a delay in differentiation or without detectable malformations (Cao et al., 2007; Cheng et al., 2009). However, a recent report using a classic KO strategy shows that deletion of miR-124-1 leads to major developmental phenotypes including small brain size, defective axonal outgrowth, and cell death confirming a crucial role for miR-124 in neurogenesis (Sanuki et al., 2011). As mentioned above, it is clear that compensation of miR-124-2 and miR-124-3 influence the phenotype of the miR-124-KO mouse, which highlights the need for generation of a conditional triple-miR-124-KO mouse. Although this will be challenging and time consuming, such a strategy is necessary in order to understand the role of miR-124 during brain development and will assist the understanding of miR-124 in adult neurogenesis. However, since the use of retroviral or lentiviral vectors allow targeting of NSCs in vivo, the application of stable inhibition vectors using for example miRNA sponges to reassess the role of miR-124 in adult NSCs is an interesting alternative.
miR-9
Another well-studied brain-specific miRNA involved in neurogenesis is miR-9 that is expressed in NSCs and upregulated upon neural differentiation (Krichevsky et al., 2006; De Pietri Tonelli et al., 2008; Zhao et al., 2008; Laneve et al., 2010; Bonev et al., 2011; Shibata et al., 2011; Yoo et al., 2011). Initial studies, done in zebrafish, showed that miR-9 directs late organizer activity of the midbrain hindbrain boundary (MHB; Leucht et al., 2008). The MHB is an organizing center in the vertebrate neural tube essential for proper development of the midbrain and anterior hindbrain (Wurst and Bally-Cuif, 2001). In human cells, miR-9 was found to have an important role in migration and proliferation of NSCs. miR-9 expression is turned on in NSCs when differentiated from hESCs. Knockdown experiments in neurospheres, using an LNA antisense probe, led to reduced proliferation, and an increased migration (Delaloy et al., 2010). In contrast, Zhao et al. found reduced proliferation accompanied with increased differentiation of mouse NSCs when overexpressing miR-9 by RNA duplexes. When they knocked down miR-9 with 2′-O-methyl AMO, proliferation increased (Zhao et al., 2009). In mouse, miR-9 has been shown to have a regional diversity along the anterior/posterior-axis; knockdown in hindbrain leads to a failure of cell cycle, promoting proliferation of neural progenitor cells, whereas cells lacking miR-9 in the forebrain undergo p53-dependent apoptosis (Bonev et al., 2011).
Just as miR-124, miR-9 has reciprocal actions with REST; REST inhibits miR-9-2 in undifferentiated neuroblastoma cells, REST and CREB inactivation triggers miR-9-2 activation (Laneve et al., 2010). Other miR-9 targets include Stathmin that increases microtubule instability (Delaloy et al., 2010) and Tlx that regulate stem cell fate (Zhao et al., 2009). Tlx is suppressed by miR-9 to negatively regulate stem cell proliferation and accelerate neural differentiation (Zhao et al., 2009). Hairy1 has also been suggested to mediate the effects of miR-9 on proliferation (Bonev et al., 2011).
miR-9 expression pattern during mouse development has been investigated by ISH; it is expressed in the developing medial pallium although it is most abundant in cortex. When a miR-9 AMO was electroporated into E11.5 cerebral cortex, deficiencies in differentiation of Cajal–Retzius cells and early born neurons were seen, suggested to be due to the increased expression of the target gene Foxg1 (Shibata et al., 2008). Generation of a miR-9-2 and miR-9-3 double KO mouse, that is the two most abundant forms during telencephalon development, resulted in major phenotypic brain defects. Cortical layers and VZ were reduced, lateral ventricles expanded, the proliferative zones hyperplasic and differentiated structures reduced. In addition, mice suffered from growth retardation and died within 1 week, demonstrating the importance for miR-9 in neurogenesis (Shibata et al., 2011).
These reports clearly demonstrate that miR-9 influence NSCs, perhaps by regulating self-renewal and migration. Still, much remains unclear regarding the functional role of miR-9 in NSCs in vivo. As with miR-124, it will be interesting to follow the generation of a conditional triple-miR-9-KO mouse or the application of stable miRNA sponges to study the functional role of miR-9.
The Let-7-Family
Let-7 is one of the first miRNA discovered in C. elegans, the first known human miRNA and it is conserved over species. There are 12 human let-7 genes encoding for nine distinct mature forms of the miRNA, let-7a through to let-7i. Increased let-7 expression is seen in early neurogenesis and neural differentiation while it is decreased in the adult brain. ISH shows induction of let-7 already at E9.5 in the developing CNS (Wulczyn et al., 2007). Let-7 expression closely resembles the expression of other brain-enriched miRNAs (Wulczyn et al., 2007).
In utero electroporation of let-7b duplexes injected into the lateral ventricles of E13.5 mice causes a reduction in cell cycle progression in NSCs (Zhao et al., 2010). In vitro, let-7 plays a role in neural differentiation of embryonic neural progenitors. Overexpression of let-7b in NSCs causes reduced proliferation and an increase in neural differentiation (Zhao et al., 2010). Suppression of let-7, using a miRNA-sponge, causes an increase in levels of lin28 protein (Rybak et al., 2008). Lin28 is a protein that specifically binds and blocks processing of let-7, thereby inducing pluripotency (Rybak et al., 2008; Balzer et al., 2010). Lin28 is expressed broadly throughout the neural tube early during development where neural differentiation has not begun (Balzer et al., 2010), at this stage it co-localizes with Sox2, a maker for NSCs. It has also been shown that let-7b suppresses the expression of Tlx (Zhao et al., 2010).
These reports suggest that the let-7-family serves as key regulators of NSC proliferation and accelerated neural differentiation (Wulczyn et al., 2007; Zhao et al., 2010). However, the large size of the let-7 family posses a technical hurdle for the generation of loss-of-function mutants, which limits our understanding of the role of let-7. In the future it appears likely that the use of sponge-vectors allowing stable inhibition of the entire family may be the most feasible choice to study the functional role of let-7 in NSCs in vivo.
Conclusion
It is likely that we have so far only begun to realize the complexity of miRNA-mediated regulation of NSCs. The multitude of miRNA complimentary targets in the genome implicates the complexity of miRNA gene regulation. Therefore studies of miRNA-target regulation in specific cell types at various developmental time points are essential. Other questions that need to be answered are; can several miRNAs act to suppress the same mRNA simultaneously and do they have compensatory, collaborative or competitive effects?
The development of new biotechnological tools such as miRNA sponges and transgenic reporter systems will enable new types of studies that will clarify the functional properties of individual miRNAs. In the coming years it will be extremely interesting to follow this field as it matures and unravels the full role of miRNAs in NSCs.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
Our work is funded by the Swedish research council and the Swedish cancer foundation. We are grateful to Shane Grealish for excellent comments on the manuscript.
References
Alvarez-Buylla, A., Seri, B., and Doetsch, F. (2002). Identification of neural stem cells in the adult vertebrate brain. Brain Res. Bull. 57, 751–758.
Amendola, M., Passerini, L., Pucci, F., Gentner, B., Bacchetta, R., and Naldini, L. (2009). Regulated and multiple miRNA and siRNA delivery into primary cells by a lentiviral platform. Mol. Ther. 17, 1039–1052.
Andersson, T., Rahman, S., Sansom, S. N., Alsio, J. M., Kaneda, M., Smith, J., O’carroll, D., Tarakhovsky, A., and Livesey, F. J. (2010). Reversible block of mouse neural stem cell differentiation in the absence of Dicer and microRNAs. PLoS ONE 5, e13453. doi:10.1371/journal.pone.0013453
Balzer, E., Heine, C., Jiang, Q., Lee, V. M., and Moss, E. G. (2010). LIN28 alters cell fate succession and acts independently of the let-7 microRNA during neurogliogenesis in vitro. Development 137, 891–900.
Bonev, B., Pisco, A., and Papalopulu, N. (2011). MicroRNA-9 reveals regional diversity of neural progenitors along the anterior-posterior axis. Dev. Cell 20, 19–32.
Boutla, A., Delidakis, C., and Tabler, M. (2003). Developmental defects by antisense-mediated inactivation of micro-RNAs 2 and 13 in Drosophila and the identification of putative target genes. Nucleic Acids Res. 31, 4973–4980.
Brown, B. D., Gentner, B., Cantore, A., Colleoni, S., Amendola, M., Zingale, A., Baccarini, A., Lazzari, G., Galli, C., and Naldini, L. (2007). Endogenous microRNA can be broadly exploited to regulate transgene expression according to tissue, lineage and differentiation state. Nat. Biotechnol. 25, 1457–1467.
Cao, X., Pfaff, S. L., and Gage, F. H. (2007). A functional study of miR-124 in the developing neural tube. Genes Dev. 21, 531–536.
Castoldi, M., Benes, V., Hentze, M. W., and Muckenthaler, M. U. (2007). miChip: a microarray platform for expression profiling of microRNAs based on locked nucleic acid (LNA) oligonucleotide capture probes. Methods 43, 146–152.
Castoldi, M., Schmidt, S., Benes, V., Noerholm, M., Kulozik, A. E., Hentze, M. W., and Muckenthaler, M. U. (2006). A sensitive array for microRNA expression profiling (miChip) based on locked nucleic acids (LNA). RNA 12, 913–920.
Chen, C., Ridzon, D. A., Broomer, A. J., Zhou, Z., Lee, D. H., Nguyen, J. T., Barbisin, M., Xu, N. L., Mahuvakar, V. R., Andersen, M. R., Lao, K. Q., Livak, K. J., and Guegler, K. J. (2005). Real-time quantification of microRNAs by stem-loop RT-PCR. Nucleic Acids Res. 33, e179.
Cheng, L. C., Pastrana, E., Tavazoie, M., and Doetsch, F. (2009). miR-124 regulates adult neurogenesis in the subventricular zone stem cell niche. Nat. Neurosci. 12, 399–408.
Colin, A., Faideau, M., Dufour, N., Auregan, G., Hassig, R., Andrieu, T., Brouillet, E., Hantraye, P., Bonvento, G., and Deglon, N. (2009). Engineered lentiviral vector targeting astrocytes in vivo. Glia 57, 667–679.
Conaco, C., Otto, S., Han, J. J., and Mandel, G. (2006). Reciprocal actions of REST and a microRNA promote neuronal identity. Proc. Natl. Acad. Sci. U.S.A. 103, 2422–2427.
Creighton, C. J., Reid, J. G., and Gunaratne, P. H. (2009). Expression profiling of microRNAs by deep sequencing. Brief. Bioinformatics 10, 490–497.
Dalma-Weiszhausz, D. D., Warrington, J., Tanimoto, E. Y., and Miyada, C. G. (2006). The affymetrix genechip platform: an overview. Meth. Enzymol. 410, 3–28.
Davis, S., Lollo, B., Freier, S., and Esau, C. (2006). Improved targeting of miRNA with antisense oligonucleotides. Nucleic Acids Res. 34, 2294–2304.
Davis, S., Propp, S., Freier, S. M., Jones, L. E., Serra, M. J., Kinberger, G., Bhat, B., Swayze, E. E., Bennett, C. F., and Esau, C. (2009). Potent inhibition of microRNA in vivo without degradation. Nucleic Acids Res. 37, 70–77.
Davis, T. H., Cuellar, T. L., Koch, S. M., Barker, A. J., Harfe, B. D., McManus, M. T., and Ullian, E. M. (2008). Conditional loss of Dicer disrupts cellular and tissue morphogenesis in the cortex and hippocampus. J. Neurosci. 28, 4322–4330.
De Pietri Tonelli, D., Pulvers, J. N., Haffner, C., Murchison, E. P., Hannon, G. J., and Huttner, W. B. (2008). miRNAs are essential for survival and differentiation of newborn neurons but not for expansion of neural progenitors during early neurogenesis in the mouse embryonic neocortex. Development 135, 3911–3921.
Delaloy, C., Liu, L., Lee, J. A., Su, H., Shen, F., Yang, G. Y., Young, W. L., Ivey, K. N., and Gao, F. B. (2010). MicroRNA-9 coordinates proliferation and migration of human embryonic stem cell-derived neural progenitors. Cell Stem Cell 6, 323–335.
Di Stefano, B., Maffioletti, S. M., Gentner, B., Ungaro, F., Schira, G., Naldini, L., and Broccoli, V. (2011). A miRNA-based system for selecting and maintaining the pluripotent state in human induced pluripotent stem cells. Stem Cells 29, 1684–1695.
Doetsch, F., Caille, I., Lim, D. A., Garcia-Verdugo, J. M., and Alvarez-Buylla, A. (1999). Subventricular zone astrocytes are neural stem cells in the adult mammalian brain. Cell 97, 703–716.
Ebert, M. S., Neilson, J. R., and Sharp, P. A. (2007). MicroRNA sponges: competitive inhibitors of small RNAs in mammalian cells. Nat. Methods 4, 721–726.
Elmen, J., Lindow, M., Schutz, S., Lawrence, M., Petri, A., Obad, S., Lindholm, M., Hedtjarn, M., Hansen, H. F., Berger, U., Gullans, S., Kearney, P., Sarnow, P., Straarup, E. M., and Kauppinen, S. (2008a). LNA-mediated microRNA silencing in non-human primates. Nature 452, 896–899.
Elmen, J., Lindow, M., Silahtaroglu, A., Bak, M., Christensen, M., Lind-Thomsen, A., Hedtjarn, M., Hansen, J. B., Hansen, H. F., Straarup, E. M., McCullagh, K., Kearney, P., and Kauppinen, S. (2008b). Antagonism of microRNA-122 in mice by systemically administered LNA-antimiR leads to up-regulation of a large set of predicted target mRNAs in the liver. Nucleic Acids Res. 36, 1153–1162.
Esau, C., Davis, S., Murray, S. F., Yu, X. X., Pandey, S. K., Pear, M., Watts, L., Booten, S. L., Graham, M., McKay, R., Subramaniam, A., Propp, S., Lollo, B. A., Freier, S., Bennett, C. F., Bhanot, S., and Monia, B. P. (2006). miR-122 regulation of lipid metabolism revealed by in vivo antisense targeting. Cell Metab. 3, 87–98.
Fabani, M. M., and Gait, M. J. (2008). miR-122 targeting with LNA/2′-O-methyl oligonucleotide mixmers, peptide nucleic acids (PNA), and PNA-peptide conjugates. RNA 14, 336–346.
Farrell, B. C., Power, E. M., and Mc Dermott, K. W. (2011). Developmentally regulated expression of Sox9 and microRNAs 124, 128 and 23 in neuroepithelial stem cells in the developing spinal cord. Int. J. Dev. Neurosci. 29, 31–36.
Friedlander, M. R., Chen, W., Adamidi, C., Maaskola, J., Einspanier, R., Knespel, S., and Rajewsky, N. (2008). Discovering microRNAs from deep sequencing data using miRDeep. Nat. Biotechnol. 26, 407–415.
Gao, F. B. (2010). Context-dependent functions of specific microRNAs in neuronal development. Neural Dev. 5, 25.
Gentner, B., Schira, G., Giustacchini, A., Amendola, M., Brown, B. D., Ponzoni, M., and Naldini, L. (2009). Stable knockdown of microRNA in vivo by lentiviral vectors. Nat. Methods 6, 63–66.
Gentner, B., Visigalli, I., Hiramatsu, H., Lechman, E., Ungari, S., Giustacchini, A., Schira, G., Amendola, M., Quattrini, A., Martino, S., Orlacchio, A., Dick, J. E., Biffi, A., and Naldini, L. (2010). Identification of hematopoietic stem cell-specific miRNAs enables gene therapy of globoid cell leukodystrophy. Sci. Transl. Med. 2, 58ra84.
Giraldez, A. J., Cinalli, R. M., Glasner, M. E., Enright, A. J., Thomson, J. M., Baskerville, S., Hammond, S. M., Bartel, D. P., and Schier, A. F. (2005). MicroRNAs regulate brain morphogenesis in zebrafish. Science 308, 833–838.
Grandjean, V., Gounon, P., Wagner, N., Martin, L., Wagner, K. D., Bernex, F., Cuzin, F., and Rassoulzadegan, M. (2009). The miR-124-Sox9 paramutation: RNA-mediated epigenetic control of embryonic and adult growth. Development 136, 3647–3655.
Griffiths-Jones, S. (2006). miRBase: the microRNA sequence database. Methods Mol. Biol. 342, 129–138.
Hebert, S. S., and Nelson, P. T. (2011). Studying microRNAs in the brain: technical lessons learned from the first ten years. Exp. Neurol. doi: 10.1016/j.expneurol.2011.12.004. [Epub ahead of print].
Kaneko, H., Dridi, S., Tarallo, V., Gelfand, B. D., Fowler, B. J., Cho, W. G., Kleinman, M. E., Ponicsan, S. L., Hauswirth, W. W., Chiodo, V. A., Kariko, K., Yoo, J. W., Lee, D. K., Hadziahmetovic, M., Song, Y., Misra, S., Chaudhuri, G., Buaas, F. W., Braun, R. E., Hinton, D. R., Zhang, Q., Grossniklaus, H. E., Provis, J. M., Madigan, M. C., Milam, A. H., Justice, N. L., Albuquerque, R. J., Blandford, A. D., Bogdanovich, S., Hirano, Y., Witta, J., Fuchs, E., Littman, D. R., Ambati, B. K., Rudin, C. M., Chong, M. M., Provost, P., Kugel, J. F., Goodrich, J. A., Dunaief, J. L., Baffi, J. Z., and Ambati, J. (2011). DICER1 deficit induces Alu RNA toxicity in age-related macular degeneration. Nature 471, 325–330.
Kawase-Koga, Y., Otaegi, G., and Sun, T. (2009). Different timings of Dicer deletion affect neurogenesis and gliogenesis in the developing mouse central nervous system. Dev. Dyn. 238, 2800–2812.
Kempermann, G., Jessberger, S., Steiner, B., and Kronenberg, G. (2004). Milestones of neuronal development in the adult hippocampus. Trends Neurosci. 27, 447–452.
Konopka, W., Kiryk, A., Novak, M., Herwerth, M., Parkitna, J. R., Wawrzyniak, M., Kowarsch, A., Michaluk, P., Dzwonek, J., Arnsperger, T., Wilczynski, G., Merkenschlager, M., Theis, F. J., Kohr, G., Kaczmarek, L., and Schutz, G. (2010). MicroRNA loss enhances learning and memory in mice. J. Neurosci. 30, 14835–14842.
Kreil, D. P., Russell, R. R., and Russell, S. (2006). Microarray oligonucleotide probes. Meth. Enzymol. 410, 73–98.
Krichevsky, A. M., Sonntag, K. C., Isacson, O., and Kosik, K. S. (2006). Specific microRNAs modulate embryonic stem cell-derived neurogenesis. Stem Cells 24, 857–864.
Krutzfeldt, J., Poy, M. N., and Stoffel, M. (2006). Strategies to determine the biological function of microRNAs. Nat. Genet. 38(Suppl.), S14–S19.
Krutzfeldt, J., Rajewsky, N., Braich, R., Rajeev, K. G., Tuschl, T., Manoharan, M., and Stoffel, M. (2005). Silencing of microRNAs in vivo with “antagomirs”. Nature 438, 685–689.
Lagos-Quintana, M., Rauhut, R., Yalcin, A., Meyer, J., Lendeckel, W., and Tuschl, T. (2002). Identification of tissue-specific microRNAs from mouse. Curr. Biol. 12, 735–739.
Laneve, P., Gioia, U., Andriotto, A., Moretti, F., Bozzoni, I., and Caffarelli, E. (2010). A minicircuitry involving REST and CREB controls miR-9-2 expression during human neuronal differentiation. Nucleic Acids Res. 38, 6895–6905.
Le, M. T., Xie, H., Zhou, B., Chia, P. H., Rizk, P., Um, M., Udolph, G., Yang, H., Lim, B., and Lodish, H. F. (2009). MicroRNA-125b promotes neuronal differentiation in human cells by repressing multiple targets. Mol. Cell. Biol. 29, 5290–5305.
Leucht, C., Stigloher, C., Wizenmann, A., Klafke, R., Folchert, A., and Bally-Cuif, L. (2008). MicroRNA-9 directs late organizer activity of the midbrain-hindbrain boundary. Nat. Neurosci. 11, 641–648.
Lim, L. P., Lau, N. C., Garrett-Engele, P., Grimson, A., Schelter, J. M., Castle, J., Bartel, D. P., Linsley, P. S., and Johnson, J. M. (2005). Microarray analysis shows that some microRNAs downregulate large numbers of target mRNAs. Nature 433, 769–773.
Liu, C., Teng, Z. Q., Santistevan, N. J., Szulwach, K. E., Guo, W., Jin, P., and Zhao, X. (2010). Epigenetic regulation of miR-184 by MBD1 governs neural stem cell proliferation and differentiation. Cell Stem Cell 6, 433–444.
Liu, K., Liu, Y., Mo, W., Qiu, R., Wang, X., Wu, J. Y., and He, R. (2011a). MiR-124 regulates early neurogenesis in the optic vesicle and forebrain, targeting NeuroD1. Nucleic Acids Res. 39, 2869–2879.
Liu, X. S., Chopp, M., Zhang, R. L., Tao, T., Wang, X. L., Kassis, H., Hozeska-Solgot, A., Zhang, L., Chen, C., and Zhang, Z. G. (2011b). MicroRNA profiling in subventricular zone after stroke: MiR-124a regulates proliferation of neural progenitor cells through Notch signaling pathway. PLoS ONE 6, e23461. doi:10.1371/journal.pone.0023461
Luikart, B. W., Bensen, A. L., Washburn, E. K., Perederiy, J. V., Su, K. G., Li, Y., Kernie, S. G., Parada, L. F., and Westbrook, G. L. (2011). miR-132 mediates the integration of newborn neurons into the adult dentate gyrus. PLoS ONE 6, e19077. doi:10.1371/journal.pone.0019077
Lunyak, V. V., and Rosenfeld, M. G. (2005). No rest for REST: REST/NRSF regulation of neurogenesis. Cell 121, 499–501.
Magill, S. T., Cambronne, X. A., Luikart, B. W., Lioy, D. T., Leighton, B. H., Westbrook, G. L., Mandel, G., and Goodman, R. H. (2010). MicroRNA-132 regulates dendritic growth and arborization of newborn neurons in the adult hippocampus. Proc. Natl. Acad. Sci. U.S.A. 107, 20382–20387.
Maiorano, N. A., and Mallamaci, A. (2009). Promotion of embryonic cortico-cerebral neuronogenesis by miR-124. Neural Dev. 4, 40.
Makeyev, E. V., Zhang, J., Carrasco, M. A., and Maniatis, T. (2007). The MicroRNA miR-124 promotes neuronal differentiation by triggering brain-specific alternative pre-mRNA splicing. Mol. Cell 27, 435–448.
Mansfield, J. H., Harfe, B. D., Nissen, R., Obenauer, J., Srineel, J., Chaudhuri, A., Farzan-Kashani, R., Zuker, M., Pasquinelli, A. E., Ruvkun, G., Sharp, P. A., Tabin, C. J., and McManus, M. T. (2004). MicroRNA-responsive “sensor” transgenes uncover Hox-like and other developmentally regulated patterns of vertebrate microRNA expression. Nat. Genet. 36, 1079–1083.
Obad, S., Dos Santos, C. O., Petri, A., Heidenblad, M., Broom, O., Ruse, C., Fu, C., Lindow, M., Stenvang, J., Straarup, E. M., Hansen, H. F., Koch, T., Pappin, D., Hannon, G. J., and Kauppinen, S. (2011). Silencing of microRNA families by seed-targeting tiny LNAs. Nat. Genet. 43, 371–378.
Obernosterer, G., Leuschner, P. J., Alenius, M., and Martinez, J. (2006). Post-transcriptional regulation of microRNA expression. RNA 12, 1161–1167.
Obernosterer, G., Martinez, J., and Alenius, M. (2007). Locked nucleic acid-based in situ detection of microRNAs in mouse tissue sections. Nat. Protoc. 2, 1508–1514.
Pena, J. T., Sohn-Lee, C., Rouhanifard, S. H., Ludwig, J., Hafner, M., Mihailovic, A., Lim, C., Holoch, D., Berninger, P., Zavolan, M., and Tuschl, T. (2009). miRNA in situ hybridization in formaldehyde and EDC-fixed tissues. Nat. Methods 6, 139–141.
Ponomarev, E. D., Veremeyko, T., Barteneva, N., Krichevsky, A. M., and Weiner, H. L. (2010). MicroRNA-124 promotes microglia quiescence and suppresses EAE by deactivating macrophages via the C/EBP-alpha-PU.1 pathway. Nat. Med. 17, 64–70.
Ramkissoon, S. H., Mainwaring, L. A., Sloand, E. M., Young, N. S., and Kajigaya, S. (2006). Nonisotopic detection of microRNA using digoxigenin labeled RNA probes. Mol. Cell. Probes 20, 1–4.
Raymond, C. K., Roberts, B. S., Garrett-Engele, P., Lim, L. P., and Johnson, J. M. (2005). Simple, quantitative primer-extension PCR assay for direct monitoring of microRNAs and short-interfering RNAs. RNA 11, 1737–1744.
Ro, S., Park, C., Jin, J., Sanders, K. M., and Yan, W. (2006). A PCR-based method for detection and quantification of small RNAs. Biochem. Biophys. Res. Commun. 351, 756–763.
Rodriguez, A., Vigorito, E., Clare, S., Warren, M. V., Couttet, P., Soond, D. R., Van Dongen, S., Grocock, R. J., Das, P. P., Miska, E. A., Vetrie, D., Okkenhaug, K., Enright, A. J., Dougan, G., Turner, M., and Bradley, A. (2007). Requirement of bic/microRNA-155 for normal immune function. Science 316, 608–611.
Ruby, J. G., Stark, A., Johnston, W. K., Kellis, M., Bartel, D. P., and Lai, E. C. (2007). Evolution, biogenesis, expression, and target predictions of a substantially expanded set of Drosophila microRNAs. Genome Res. 17, 1850–1864.
Rybak, A., Fuchs, H., Smirnova, L., Brandt, C., Pohl, E. E., Nitsch, R., and Wulczyn, F. G. (2008). A feedback loop comprising lin-28 and let-7 controls pre-let-7 maturation during neural stem-cell commitment. Nat. Cell Biol. 10, 987–993.
Sachdeva, R., Jonsson, M. E., Nelander, J., Kirkeby, A., Guibentif, C., Gentner, B., Naldini, L., Bjorklund, A., Parmar, M., and Jakobsson, J. (2010). Tracking differentiating neural progenitors in pluripotent cultures using microRNA-regulated lentiviral vectors. Proc. Natl. Acad. Sci. U.S.A. 107, 11602–11607.
Sanuki, R., Onishi, A., Koike, C., Muramatsu, R., Watanabe, S., Muranishi, Y., Irie, S., Uneo, S., Koyasu, T., Matsui, R., Cherasse, Y., Urade, Y., Watanabe, D., Kondo, M., Yamashita, T., and Furukawa, T. (2011). miR-124a is required for hippocampal axogenesis and retinal cone survival through Lhx2 suppression. Nat. Neurosci. 14, 1125–1134.
Schmittgen, T. D., Jiang, J., Liu, Q., and Yang, L. (2004). A high-throughput method to monitor the expression of microRNA precursors. Nucleic Acids Res. 32, e43.
Schmittgen, T. D., and Livak, K. J. (2008). Analyzing real-time PCR data by the comparative C(T) method. Nat. Protoc. 3, 1101–1108.
Shi, Y., Zhao, X., Hsieh, J., Wichterle, H., Impey, S., Banerjee, S., Neveu, P., and Kosik, K. S. (2010). MicroRNA regulation of neural stem cells and neurogenesis. J. Neurosci. 30, 14931–14936.
Shibata, M., Kurokawa, D., Nakao, H., Ohmura, T., and Aizawa, S. (2008). MicroRNA-9 modulates Cajal-Retzius cell differentiation by suppressing Foxg1 expression in mouse medial pallium. J. Neurosci. 28, 10415–10421.
Shibata, M., Nakao, H., Kiyonari, H., Abe, T., and Aizawa, S. (2011). MicroRNA-9 regulates neurogenesis in mouse telencephalon by targeting multiple transcription factors. J. Neurosci. 31, 3407–3422.
Silber, J., Lim, D. A., Petritsch, C., Persson, A. I., Maunakea, A. K., Yu, M., Vandenberg, S. R., Ginzinger, D. G., James, C. D., Costello, J. F., Bergers, G., Weiss, W. A., Alvarez-Buylla, A., and Hodgson, J. G. (2008). miR-124 and miR-137 inhibit proliferation of glioblastoma multiforme cells and induce differentiation of brain tumor stem cells. BMC Med. 6, 14. doi:10.1186/1741-7015-6-14
Smirnova, L., Grafe, A., Seiler, A., Schumacher, S., Nitsch, R., and Wulczyn, F. G. (2005). Regulation of miRNA expression during neural cell specification. Eur. J. Neurosci. 21, 1469–1477.
Sun, G., Ye, P., Murai, K., Lang, M. F., Li, S., Zhang, H., Li, W., Fu, C., Yin, J., Wang, A., Ma, X., and Shi, Y. (2011). miR-137 forms a regulatory loop with nuclear receptor TLX and LSD1 in neural stem cells. Nat. Commun. 2, 529.
Szulwach, K. E., Li, X., Smrt, R. D., Li, Y., Luo, Y., Lin, L., Santistevan, N. J., Li, W., Zhao, X., and Jin, P. (2010). Cross talk between microRNA and epigenetic regulation in adult neurogenesis. J. Cell Biol. 189, 127–141.
Takada, S., and Mano, H. (2007). Profiling of microRNA expression by mRAP. Nat. Protoc. 2, 3136–3145.
Thai, T. H., Calado, D. P., Casola, S., Ansel, K. M., Xiao, C., Xue, Y., Murphy, A., Frendewey, D., Valenzuela, D., Kutok, J. L., Schmidt-Supprian, M., Rajewsky, N., Yancopoulos, G., Rao, A., and Rajewsky, K. (2007). Regulation of the germinal center response by microRNA-155. Science 316, 604–608.
Valoczi, A., Hornyik, C., Varga, N., Burgyan, J., Kauppinen, S., and Havelda, Z. (2004). Sensitive and specific detection of microRNAs by northern blot analysis using LNA-modified oligonucleotide probes. Nucleic Acids Res. 32, e175.
van Rooij, E., Sutherland, L. B., Qi, X., Richardson, J. A., Hill, J., and Olson, E. N. (2007). Control of stress-dependent cardiac growth and gene expression by a microRNA. Science 316, 575–579.
Vester, B., and Wengel, J. (2004). LNA (locked nucleic acid): high-affinity targeting of complementary RNA and DNA. Biochemistry 43, 13233–13241.
Visvanathan, J., Lee, S., Lee, B., Lee, J. W., and Lee, S. K. (2007). The microRNA miR-124 antagonizes the anti-neural REST/SCP1 pathway during embryonic CNS development. Genes Dev. 21, 744–749.
Weiler, J., Hunziker, J., and Hall, J. (2006). Anti-miRNA oligonucleotides (AMOs): ammunition to target miRNAs implicated in human disease? Gene Ther. 13, 496–502.
Wheeler, G., Valoczi, A., Havelda, Z., and Dalmay, T. (2007). In situ detection of animal and plant microRNAs. DNA Cell Biol. 26, 251–255.
Wolber, P. K., Collins, P. J., Lucas, A. B., De Witte, A., and Shannon, K. W. (2006). The Agilent in situ-synthesized microarray platform. Meth. Enzymol. 410, 28–57.
Wu, J., and Xie, X. (2006). Comparative sequence analysis reveals an intricate network among REST, CREB and miRNA in mediating neuronal gene expression. Genome Biol. 7, R85.
Wulczyn, F. G., Smirnova, L., Rybak, A., Brandt, C., Kwidzinski, E., Ninnemann, O., Strehle, M., Seiler, A., Schumacher, S., and Nitsch, R. (2007). Post-transcriptional regulation of the let-7 microRNA during neural cell specification. FASEB J. 21, 415–426.
Wurst, W., and Bally-Cuif, L. (2001). Neural plate patterning: upstream and downstream of the isthmic organizer. Nat. Rev. Neurosci. 2, 99–108.
Yoo, A. S., Sun, A. X., Li, L., Shcheglovitov, A., Portmann, T., Li, Y., Lee-Messer, C., Dolmetsch, R. E., Tsien, R. W., and Crabtree, G. R. (2011). MicroRNA-mediated conversion of human fibroblasts to neurons. Nature 476, 228–231.
Yu, J. Y., Chung, K. H., Deo, M., Thompson, R. C., and Turner, D. L. (2008). MicroRNA miR-124 regulates neurite outgrowth during neuronal differentiation. Exp. Cell Res. 314, 2618–2633.
Zhao, C., Deng, W., and Gage, F. H. (2008). Mechanisms and functional implications of adult neurogenesis. Cell 132, 645–660.
Zhao, C., Sun, G., Li, S., Lang, M. F., Yang, S., Li, W., and Shi, Y. (2010). MicroRNA let-7b regulates neural stem cell proliferation and differentiation by targeting nuclear receptor TLX signaling. Proc. Natl. Acad. Sci. U.S.A. 107, 1876–1881.
Zhao, C., Sun, G., Li, S., and Shi, Y. (2009). A feedback regulatory loop involving microRNA-9 and nuclear receptor TLX in neural stem cell fate determination. Nat. Struct. Mol. Biol. 16, 365–371.
Keywords: microRNA, neural stem cells, neurogenesis, miR-124, miR-9, let-7
Citation: Åkerblom M, Sachdeva R and Jakobsson J (2012) Functional studies of microRNAs in neural stem cells: problems and perspectives. Front. Neurosci. 6:14. doi: 10.3389/fnins.2012.00014
Received: 03 October 2011;
Accepted: 19 January 2012;
Published online: 07 February 2012.
Edited by:
Yanhong Shi, City of Hope, USAReviewed by:
Carlos P. Fitzsimons, University of Amsterdam, NetherlandsChunnian Zhao, California State University, USA
Copyright: © 2012 Åkerblom, Sachdeva and Jakobsson. This is an open-access article distributed under the terms of the Creative Commons Attribution Non Commercial License, which permits non-commercial use, distribution, and reproduction in other forums, provided the original authors and source are credited.
*Correspondence: Johan Jakobsson, Laboratory of Molecular Neurogenetics, Wallenberg Neuroscience Center, Lund University, BMC A11, 221 84 Lund, Sweden. e-mail:am9oYW4uamFrb2Jzc29uQG1lZC5sdS5zZQ==