 Lígia A. B. M. Mendonça1*
Lígia A. B. M. Mendonça1* Rosângela dos Santos Ferreira2*Rita de Cássia Avellaneda Guimarães2
Rosângela dos Santos Ferreira2*Rita de Cássia Avellaneda Guimarães2 Alinne P. de Castro1
Alinne P. de Castro1 Octávio L. Franco1,3
Octávio L. Franco1,3 Rosemary Matias4,5Cristiano M. E. Carvalho1,5
Rosemary Matias4,5Cristiano M. E. Carvalho1,5- 1S-Inova Biotech Post Graduate Program in Biotechnology, Catholic University Dom Bosco, Campo Grande, Brazil
- 2Post Graduate Program in Health and Development in the Central-West Region of Brazil, Federal University of Mato Grosso do Sul, Campo Grande, Brazil
- 3Center of Proteomic and Biochemical Analysis, Post Graduate Program in Genomic Sciences and Biotechnology, Catholic University of Brasilia, Brasilia, Brazil
- 4Post Graduate Program in Environmental Sciences and Agricultural Sustainability, Catholic University Dom Bosco, Campo Grande, Brazil
- 5Post Graduate Program in Environment and Regional Development, University Anhanguera Uniderp, Campo Grande, Brazil
Colorectal cancer exerts a strong influence on the epidemiological panorama worldwide, and it is directly correlated to etiologic factors that are substantiated by genetic and environmental elements. This complex mixture of factors also has a relationship involving the structural dependence and composition of the gut microbiome, leading to a dysbacteriosis process that may evolve to serious modifications in the intestinal lining, eventually causing the development of a neoplasm. The gastrointestinal tract presents defense strategies and immunological properties that interfere in intestinal permeability, inhibiting the bacterial translocation, thus maintaining the integrity of intestinal homeostasis. The modulation of the intestinal microbiome and the extinction of risk factors associated with intestinal balance losses, especially of environmental factors, make cell and defense alterations impossible. This modulation may be conducted by means of functional foods in the diet, especially soluble fibers, polyunsaturated fatty acids, antioxidants and prebiotics that signal immunomodulatory effects in the intestinal microbiota, with preventive and therapeutic action for colorectal cancer. In summary, this review focuses on the importance of dietary modulation of the intestinal microbiota as an instrument for dysbacteriosis and, consequently, for the prevention of colorectal cancer, suggesting anticarcinogenic, and antiangiogenic properties. Among the intestinal modulating agents considered here are functional foods, especially flaxseed, oat and soy, composing a Bioactive Food Compound.
Introduction
Colorectal carcinogenesis (CRC) is a neoplastic modality with a wide and varied incidence and geographical distribution (1). CRC dissemination can be observed from the alarming epidemiological panorama, and future estimates demonstrate a considerable increase in the number of new cases of and deaths from CRC (2).
CRC etiology could be based on numerous genetic and environmental changes. Damage to the intestinal environment and mutations can culminate in the development of inflammatory bowel diseases (IBDs). The IBDs are one of the major genetic risk factors and are strongly linked to changes in the composition of the gut microbiota and the intestinal cell microenvironment (3). The environmental factors involved in CRC etiology derived from deleterious effects on gut microbiota (bacterial translocation, increased intestinal permeability and the inflammatory process) are influenced by inappropriate eating habits (4), such as excessive consumption of fast (5) and fried foods, excessive salt, saturated fat, red meat, and sugary beverages, and low fiber intake (6).
Therefore, functional foods and their bioactive substances provide a viable and accessible alternative in minimizing damaged to the intestinal microenvironment. Fibers and polyunsaturated fatty acids (PUFAs), especially those of the n-3 series, are examples of functional foods that act on gut microbiota composition, decreasing the number of harmful bacteria, such as Helicobacter pylori (7–12). These foods also act in minimizing the inflammatory intestine process, preventing tumorigenesis promoted by CRC (13).
These etiological findings connect directly to immune system activities that show numerous and important protective strategies, such as the synthesis and secretion of mucus from goblet cells (14). This mucus released into the intestinal region is constituted by antibacterial factors and immunomodulatory molecules (15).
The gastrointestinal tract has its organization based on the presence of pattern recognition receptors (PRRs) that recognize particularities and characteristics of pathogens and the toll-like receptors that are related to the cascade of inflammatory signaling (9, 16). Therefore, it is essential to maintain the intestinal microbiome homeostasis, from the precise relationship among the quantity and quality of the resident microbial agents. Such a relationship seems to be fundamental for the maintenance of intestinal epithelial cell integrity and for adequate immune response, avoiding the development of dysbacteriotic processes (17). However, the interindividual variability, in face of endogenous and exogenous factors, determine the gut microbiota formation playing an important role in dysbacteriosis, leading or not to diseases development such as CRC (18). Birth types seems to be one of main factor that may influence in gut microbiota formation, being that, the composition of gut bacterial community is different in infants delivered by cesarean section in comparison to infants born by vaginal delivery (19, 20). Besides that, infant's lifestyle also influences the microbiota gut formation (21). In this context, the ethnicity, geography and socioeconomic variation seems to strongly influence the occurrence of a particular microorganism or not (22–24). The host genetics also could be an important influence factor of gut microbiota, as well as, xenobiotic exposure (pesticides, environmental pollution and chemical substances, improper eating habits and use of medicament). In the case of xenobiotic exposure, the authors mention that microorganisms that compound the gut microbiota can alter the chemical structure of poluents and minimize the harmful effects on host physiology, being a reliable gut microbiota for human health (25). Thus, this scenario shows that research in this line is challenging and yet to be explored and further investigated (18). Another challenge of gut microbiota study is to move beyond classification of the bacteria agents, but also to elucidate the mechanisms underlying their influence on host health (25).
In summary, this review aims to describe the dysbacteriotic process and its consequences in the composition and conditions of the gastrointestinal tract. It also aims to describe how the immune system and its agents are involved in the repair of dysbiosis and the development of CRC and how the modulation of the gut microbiota takes place in response to functional foods and their bioactive constituents.
Colorectal Carcinogenesis
Cancer is a multifaceted disease that presents abnormal and disordered cell growth as the main pathological characteristic (26). Another important feature of cancer is the alarming epidemiological framework that indicates a substantial increase in the number of deaths (13 million) and new cases (21.7 million) by the year 2030 (2).The IARC (2) and Cancer Research UK (27) emphasize that the epidemiological context of cancer by the year 2030 may be even greater, reaching 23.6 million new cases. This increase could be strongly correlated to risk factors including smoking, alcoholism, and a poor junk food diet (5) normally composed of saturated and trans fat, salt and sugars, constituting low nutritional quality (28). Other risk factors are also cited, including the high consumption of red meat and sugary drinks and a low fiber intake and sedentary lifestyle (6). In this scenario, CRC is highlighted because it is a silent disease, especially in the early stages, not presenting clear symptoms, thus substantially impairing the diagnostic process, treatment and cure. In advanced cases, signs and symptoms can be perceived, related to changes in the gastrointestinal tract that including: diarrhea or constipation, incomplete intestinal emptying, hemorrhoids, and occult blood in the eliminations, besides associated symptoms such as cramps and stomach pain, fatigue, weight loss and low red blood cell count (29–31).
Such symptoms, if diagnosed and identified early, contribute to a brief and assertive treatment, which includes local and systemic treatment. The former is characterized by a surgical process, radiation therapy and ablation, while the latter is targeted chemotherapy (29, 30, 32). CRC presents high frequency (1, 30, 33). For Arnold et al. (34), the number of new cases of this neoplasm will increase by 60% by the year 2030, representing 2.2 million affected and 1.1 million deaths. This type of cancer is reported as the third most common form in males and the second in females (26, 35), and it is considered the second cause of death worldwide. CRC ranks first in incidence and is responsible for 47,100 new cases per year, with an average mortality of 28.2/100,000 inhabitants in the countries of the West, South, North and West Central Europe (36, 37). In these countries, over the last 40 years, CRC has presented different profiles for the level of mortality related to gender, age group, advancement of health conditions, greater access to screening methods, with consequent early detection of the disease and specialized care (38).
CRC in Asian countries occupies first place in terms of mortality and incidence (39, 40). The disease has grown significantly in recent years, although to a lesser extent than in western countries (40), and it presents wide geographic variation, especially in developed countries, such as Japan, South Korea, Singapore and Malaysia (41, 42). The epidemiological history of CRC in the USA presents similar characteristics to the global picture regarding incidence and mortality. For the year 2017, 95.520 new cases of colon cancer and 93.910 new cases of rectal cancer were estimated, distributed evenly among men and women. A similar framework can be observed in relation to the number of deaths, with 27.150 among men and 23.110 among women (43). The men CRC incidence may occur due to risk factors exposure such as smoking and alcohol, for example (43, 44). In women older than 65 years, a lower CRC survival rate predominates (45). Such information, according to some authors, may be associated with oestrogens that plays an essential role in modulating the developing CRC risk. This is due to a differential gut hormone receptors expression, although this role has not being well elucidated (46, 47). In Brazil, CRC presents heterogeneity, distributed unevenly among regions. In 2018, 17.380 new cases were estimated among men and 18.980 among women, due to the differences associated with biological and pathophysiological factors (48), like age, since women have a longer life expectancy, a condition that makes them more vulnerable due to hormonal changes, a condition similar to the global framework (43, 46, 47). In addition to gender, there are other etiologic factors that may be related to CRC, including advanced age, genetic alterations, erroneous eating habits, sedentary lifestyle, obesity, alcoholism, smoking, contact with microorganisms and diseases such as: ulcerative colitis (UK) (49), Crohn's disease (CD), familial polyposis (FP), and Lynch syndrome (LS) (35). These factors, if not detected and corrected in advance, guarantee the development of initial lesions in the lining of the colon. These lesions are also commons in IBDs that are secondary CRC causes of and are characterized by long-term intestinal inflammation (50) and genetic alteration, culminated in oxidative stress, cellular proliferation, and activation of immune system (51).
These lesions, if not treated, can evolve in the deeper muscular layers and throughout the intestinal wall, culminating in the development of CRC. These lesions, if not treated, can evolve in the deeper muscular layers and throughout the intestinal wall, culminating in the development of CRC (26).
The Relationship Between Gut Microbiota and Colorectal Carcinogenesis
Dysbacteriosis leads to the development of several diseases, especially those of an inflammatory order, such as IBD and CRC (52). The etiology of CRC can be attributed to the proliferation of H. pylori, Streptococcus bovis Orla-Jensen, Enterococcus faecalis Schleifer and Kilpper-Bälz (1984) and Orla-Jensen (1919), Clostridium septicum, Escherichia coli T. Escherich (1885), Fusobacterium spp. Knorr (1922), Bacteroides fragilis (3, 8) and Streptococcus gallolyticus (3) (Table S1).
These microorganisms (Table S1) are etiologically related to CRC through innumerable effects. H. pylori have been associated with CRC, especially in hypergastrinemia relapses cases (8, 53). Already the relation of the bacterium s. bovis with CRC occurs due to the installation of a pro-inflammatory process that can lead to the recruitment of leukocytes and the development of pre-neoplastic lesions (54).
The carcinogenic property of E. faecalis occurs in bacteremia cases, characterized by frequent bacterial translocations with consequent increase in intestinal permeability, in addition to reactive oxygen species (ROS) formation (55). The intestinal permeability increase could also be a consequence of E. coli translocation to the intestinal region, causing serious epithelium lesions. Such deleterious effect may be related to toxins release and chronic inflammatory process development (8, 56). In such case C. septicum virulence and its ability to translocate to intestinal tissue may justify a strictly relation to CRC development (57). Fusobacterium and B. fragilis, from production and release of enterotoxins stimulate the immune response, causing alterations in intestinal epithelial cells (58, 59). These pre-neoplastic CRC effects caused by these microorganisms may occur in association. Besides that, Gagnière et al. (8) cite and establish an interconnection between genotoxins and bacterial metabolism, in addition to oxidative stress and inflammatory process, demonstrating the bacterial etiological multidirectionality of CRC.
Exploring Interactions Among Immune Responses, Intestinal Microbiota and Colorectal Carcinogenesis
The gastrointestinal tract of the host is composed of thick layers of mucus. These layers are composed of mucin glycoproteins and other antibacterial factors, such as C-α-defensins lectins, lysozyme and phospholipase A2, which are excreted by Paneth cells. When in homeostasis, these substances confer perfect regulation on microbial abundance and the immune response. Otherwise, the development of intestinal dysbiosis is established (60).
Complementing the intestinal immune defense, there are receptors whose function is to mediate the recognition process. These are the so-called PRRs that initiate the step of recognizing pathogen-associated molecular patterns (PAMPs). There are also toll-like receptors, of glycoprotein nature, known as the interface between the intestinal epithelial barrier and the immune system, which synthesize and release cytokines and chemokines and are responsible for the transcription of genes related to the triggering of the immunologic response. These receptors compose a singular superfamily called interleukin-1 receptors/toll-like receptors and are located in innumerable cells, in the intestinal epithelial barrier and in intracellular compartments (61, 62) (Figure 1).
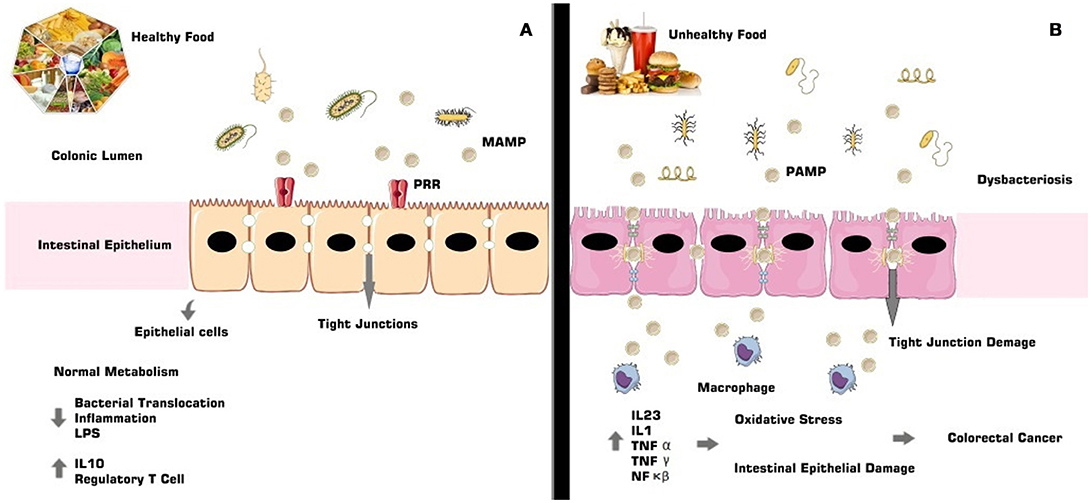
Figure 1. The gut microbiota composition is directly involved with immunologic and intestinal homeostasis (A). In dysbacteriosis the signaling cascade results in the activity of specific receptors and their ligands, leading to the release of pro-inflammatory agents, such as cytokines and interleukins, allowing oxidative damage, associated with non-intestinal epithelial lesions, and consequent development of CRC (B).
Dysbacteriosis promotes the development of an inflammatory process characterized by increased expression of pro-inflammatory genes and specific receptors such as TLR3 and TLR4, activating NF-κB and IRF3 (interferon 3 regulating factor) (16, 61), besides the immunological agents MAPK (mitogen-activated protein kinase), AP1 (activator protein 1), STAT (signal transducer and activator of transcription), and IRF3 (interferon IFN-regulatory factor 3) (63).
Metabolic pathways, such as TLR4, are sensitized and activated by endotoxins found in Gram-negative bacteria, called lipopolysaccharides (LPS), which when released by bacterial lysis are mediated by acute inflammatory factors such as NF-κB (64). This inflammatory response to LPS occurs in the presence of cell and platelet adhesion molecules, pro-inflammatory cytokines [TNF-α, interleukin 6 (IL6), interleukin (IL1)], enzymes iNOS (nitric oxide synthase) and COX-2 (cyclooxygenase 2) receptors, bradykinin B1 receptors and the IkB-α repressor protein (63, 65).
The inflammatory process involved in dysbacteriosis can be the etiological cause of CRC and is associated with hyperplastic damage in the intestinal epithelium and with lamina propria fragmentation. These characteristics confer an environment that is conducive to bacterial translocation and to the development of pro-neoplastic lesions (3, 66, 67).
Inflammation of the intestinal tissue, due to a dysbacteriotic process, for example, is accompanied by the extinction of tumor suppressor genes and of genes that aid in the repair of genetic material. This also produces inflammatory-immunological conditions that will lead to the development of an environment conducive to the cancerous installation (68). In the inflammatory-immunological process that is characteristic of dysbacteriosis and that can cause CRC, activation of the NF-κB pathway occurs, and this then causes conformational changes in the neoplastic environment, stimulates action of the pro-inflammatory cytokines, such as IL6, which has an important pathogenic role in the progression of CRC and regulates genes related to the tumor necrosis factor (TNF) and COX-2 enzyme, both highly expressed in neoplastic conditions (66). Regulatory T cells are also expressed in inflammatory conditions that can lead to CRC (3).
Therefore, it is verified that the gut microbiota and its composition (specific microorganisms) are important promoters of the inflammatory-immunological response, conditioning or not the development of a pro-mutagenic environment for CRC (69, 70).
Functional Foods as Gut Microbiota Modulators and Chemopreventive Action
The modulation of the intestinal microbiota and its consequent beneficial effects, which include protection against pathogenic invaders, immune system stimulation, and selective beneficial bacteria growth, occurs through the use of prebiotics, probiotics (71) and the association of the two in varying amounts forming the symbiotic (71–74).
In examining this association, it is important to consider the most common phyla identified in most individuals (Firmicutes and Bacteroides and, to a lesser extent, Actinobacteria and Proteobacteria) (4, 75) and the constant adaptation and quantitative and qualitative modification of the intestinal microbial composition. These modifications interfere in physiological, metabolic and immunological effects, considering the close relationship between the healthy process and disease (76).
Therefore, intestinal microbial community modifications that culminate in the extinction of the community's balance and the characterization of an intestinal dysbiosis status, as previously reviewed, can be related to the onset of chronic non-communicable diseases (NCDs), such as obesity, insulin resistance and cardiovascular diseases (77, 78), and particularly to CRC (3). Considering the benefits and effects of modulation of the gut microbiota, especially in the prevention and treatment of CRC, there are functional foods that motivate countless studies, with the objective of identifying their properties and applications. Functional foods are composed of bioactive ingredients with beneficial properties for metabolic and physiological well-being (79, 80).
Future prospects indicate that the constitution of the intestinal microbiota can be enhanced by the insertion of functional foods into the diet, with special emphasis on chemopreventive, biologically active compounds present in foods of plant origin and that can prevent and/or attenuate the development of CRC (81, 82).
In this case, flexseed (Linum usitatissimum L.) (83, 84), oat bran (Avena sativa L.) (85) and soybean Glycine max L.) (86), foods that are sources of PUFAs, especially the n-3 series (11), soluble food fibers (11, 12) and phytochemicals and antioxidants (87) (Table S2).
The anticarcinogenic property of soluble fibers like lignan and β-glucan could be justified by short chain fatty acids (SCFAs) formation, in special acetate, propionate and butyrate, from its gut microbiota fermentation (Figure 2). The SCFAs are responsible for the intracellular and colonic pH decrease, making the intestinal environment more acidic. Such condition inhibits pathogenic organism proliferation, toxic degradation products formation (88), DNA damage induction (89), the cell proliferation and increase apoptosis of tumor cells (90, 91). These factors could be reduced by dietary fibers that also have the capacity to act in the pre-neoplastic process, with a reduction in the inflammatory and oxidative process (73, 92) (Figures 1B, 3). However, such favorable CRC effects could be perceived in a dose-dependent manner (93). The ability to block neoplastic cell proliferation and induce apoptosis, as well as pre-neoplastic CRC conditions, is also a feature of n-3 series PUFAs. These lipids also could alter the cell cycle components and acts on the immune system markers and gene expression modulation by regulating CRC related genes expression (94, 95) (Figures 1B, 3). These effects according to D'Eliseo and Velotti (96) are perceived at different periods and exposure doses. Antioxidants, especially the isoflavones, genistein, and daidzein, block the DNA bases attack, avoiding the lesions formation and cellular integrity losses, due to free radicals' neutralization generated by cellular metabolism or exogenous factors (97, 98). In contrast, high food consumption when submitted to the fermentation process may be related to the increased risk of developing CRC (99). The other protective effects could be related to damaged cell membranes reconstitution and further DNA molecule damage removal, since oxidative stress may be directly related to the pre-neoplastic conditions in phases of initiation and promotion of cancer (100). Besides of the inflammatory process with increased activity of inflammatory cytokines and intestinal permeability (101) (Figures 1B, 3).
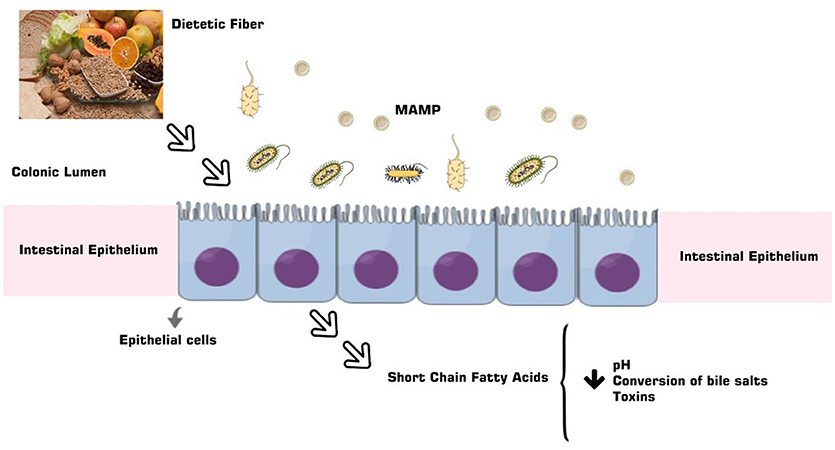
Figure 2. The PUFAs derive from a complex relationship between the gut microbiota and the diet, besides the characteristics of the microbiome, and are a product of the fermentation of dietary fiber and lignan absorbed by the small intestine.
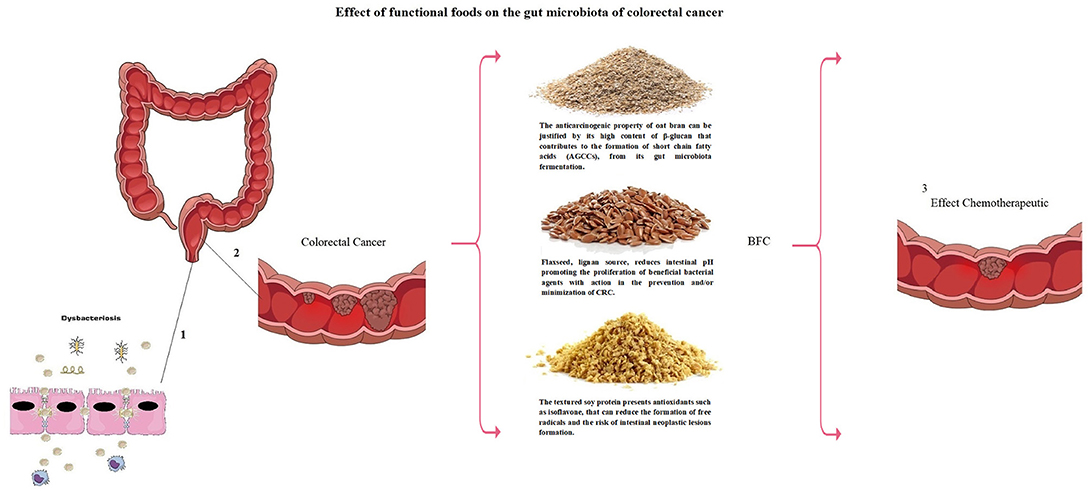
Figure 3. Dysbacteriosis process (1) causing colorectal cancer (2). Modulation of gut microbiota may be conducted by means of functional foods (BFC: Bioactive Food Compound—National Institute of Industrial Property – n. BR 10 2013 018002 5) with preventive and therapeutic action for colorectal cancer (3).
The immunomodulatory effects of flaxseed are perceived from its prebiotic properties. Because it is loaded with mucilage, this food maintains the integrity of the intestinal epithelial barrier, minimizing inflammatory processes, guaranteeing the proliferation of beneficial phyla to the detriment of putrefying and harmful species. This modification guarantees an improvement in the immune defense of the host, reducing the risk of developing NCDs, especially IBDs and CRC (102). The prebiotic effect of flaxseed is also presented by oats and soybeans. These two functional foods are filled with soluble fiber, contributing to the balance of the gut microbiota from its modulation. Gut microbiota modulation promotes the proliferation of probiotic phyla, guaranteeing the absorption of nutrients, protection against pathogens, and modulation of the immune system (103, 104).
The modulation of the gut microbiota composition from these functional foods also occurs in the presence of phytochemicals. Maintaining the balance of the gut microbiota ensures perfect immune defense and disease prevention, specifically CRC, decreasing cell proliferation, stimulating the induction of apoptosis, inhibition of angiogenesis and delay of the metastatic process (105, 106).
Finally, these functional foods (flaxseed, oat bran, and soybean) form a bioactive food compound (BFC) that is patented by National Institute of Industrial Property (number BR 10 2013 018002 5), published by Ministry of Development and Foreign Trade, Brazil in august 2015. BFC is rich in proteins, in PUFAs (60.41%) (n-3 series (30%) and n-6 series (27%)) and also in dietary fibers. Among such fiber it is possible to find soluble fibers including lignan and β-glucans, both with high viscosity and fermentable potential (107). Moreover BFC has been studied as adjuvant therapy in the NCDs directly related to metabolic syndrome (MS) including atherosclerosis. Such food composition exhibit low atherogenicity, thrombogenicity, and hypocholesterolemia/hypercholesterolemia, also acting at anthropometric measurements reduction (108, 109). Additionally studies with BFC demonstrate direct chemopreventive and/or chemotherapeutic relation with CRC (85, 86, 94, 110).
Therefore, a diet rich in vegetables, fruits, oilseeds, low glycemic complex carbohydrates, mono, and polyunsaturated fats and proteins presents a good balance in quality and quantity of phytochemicals, especially phenolic compounds such as flavonoids. These substances have considerable antioxidant activity, a property associated with a lower incidence of NCDs and lower resulting mortality, specifically CRC (111, 112).
The chemopreventive agents found in functional foods can be used with a powerful preventive effect, as their multiple mechanisms of action aid in the treatment and in the reduction of the risk of CRC recurrence, besides contributing to the improvement of the quality of life of the carriers of CRC in Figure 3.
However, it is noted with this review that there is a potential number of studies demonstrating the role of the intestinal microbiota and its imbalance at CRC pathogenesis. Associated with this problem are the functional foods that may act as anticarcinogenic and chemotherapeutic in animal models, being the model of choice for most research in this emerging field. This review proposes the continuity of potent studies that demonstrate causality in various experimental and human models, since some observations in animal models may not be applicable in people, although both present some significant similarities, genetic, anatomical, and physiological factors differ (113–115).
Conclusion and Future Prospects
Dietary modulation of the intestinal microbiota, avoiding the intake of foods that stimulate dysbiosis and intestinal inflammation, is an efficient strategy to prevent the development of CRC. In this review, several studies have demonstrated the functional chemopreventive and/or chemotherapeutic value of the foods that compose the BFC in CRC. These effects may stimulate the use of flaxseed, oat bran and soya in the diet, as part of sweet and savory preparations, as well as in the industrial production of cereal bars and added to bakery products.
Future prospects point to the recommendation of a specific dietary plan, from the ingestion of functional foods that compose the BFC, highlighting the importance of its bioactive compounds, versatility of use and ease of access. The functional properties and the economic character of these functional foods demonstrate a participation in reducing health costs and improving the quality of life of the general population, especially in the prevention and treatment of NCDs, with an emphasis on CRC.
Taking advantage of the socio-demographic aspects and economic opportunities that favor the use of BFC in CRC, it is proposed that studies on this subject should continue, considering the need for clarification on the synergism and/or antagonism between bioactive compounds, as well as the daily dose of use and the possible mechanisms of action of these food substances.
It is concluded with this review that modifiable lifestyle factors such as western standard diet with high glycaemic load, unfavorable energy balance (obese or physically inactive) and smoking may have an important role in the formation of the intestinal microbiota, which is determinant for a favorable prognosis in CRC. Related to this, it is important to consider intraindividual differences in diet response as well as the great difficulty of nutrition and food professionals in stimulating compliance with these dietary recommendations.
Author Contributions
LM and RdSF drafted the manuscript, composed the figures, and critically revised the manuscript. LM, RdSF, and RdCAG conceived the manuscript and finalized the draft. RM have written some part. RdCAG, AdC, OF, RM, and CC revised the manuscript. All authors read and approved the final manuscript. RM wrote the bioactive compounds and that revised the final version of the manuscript.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
This work was supported by National Council for Scientific and Technological Development (CNPq), Coordination of Improvement of Higher Level Personnel (CAPES), Foundation for Support to the Development of Teaching, Science and Technology of the State of Mato Grosso do Sul (FUNDECT) and Foundation for Research Support of the Distrito Federal (FAPDF), Don Bosco Catholic University (UCDB).
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fonc.2018.00325/full#supplementary-material
References
1. Bordini HP, Kremer JL, Fagundes TR, Melo GP, Conchon-Costa I, Silva SS, et al. Protective effect of metformin in an aberrant crypt foci model induced by 1,2-dimethylhydrazine: modulation of oxidative stress and inflammatory process. Mol Carcinog. (2017) 53:913–22. doi: 10.1002/mc.22545
2. International Agency for Research on Cancer (IARC). Global Cancer Facts Figures 2017. (2017). Available online at: https://www.cancer.org/research/cancer-facts-statistics/all-cancer-facts-figures/cancer-facts-figures-2017.html (Accessed April 15, 2017).
3. Brennan AC and Garrett WS. Gut microbiota, inflammation, and colorectal cancer. Annu Rev Microbiol. (2016) 70:395–411. doi: 10.1146/annurev-micro-102215-095513
4. Calças N, Mendonça LABM, Peres L, Santos RF, Elisei C, Castro A. Diet as a therapy for gut dysbacteriosis. JSM Biochem Mole Biol. (2016) 4:1021. Available online at: https://www.jscimedcentral.com/Biochemistry/vol4issue1.php (Accessed 11, August 2018).
5. Adedeji OA, John UI. Food environment and unhealthy eating habits among adolescents in tertiary institutions. Am Int J Res Humanit Arts Soc Sci. (2015) 9:72–77. Available online at: http://iasir.net/AIJRHASSpapers/AIJRHASS15-115.pdf (Accessed 11, August 2018).
6. Riccio P, Rossano R. Nutrition facts in multiple sclerosis. ASN Neuro. (2015) 2015:1–20. doi: 10.1177/1759091414568185
7. Gul K, Singh AK, Jabeen R. Avenanthramides inhibit proliferation of human colon cancer cell lines in vitro. Crit Rev Food Sci Nutr. (2015) 8:1007–16. doi: 10.1080/01635581.2010.492090
8. Gagnière J, Raisch J, Veziant J, Barnich N, Bonnet R, Buc E, et al. Gut microbiota imbalance and colorectal cancer. World J Gastroenterol. (2016) 22:501–8. doi: 10.3748/wjg.v22.i2.501
9. Gao R, Gao Z, Huang L. Qin, H. Gut microbiota and colorectal cancer. Eur J Clin Microbiol Infect Dis. (2017) 36:757–69. doi: 10.1007/s10096-016-2881-8
10. Papandreou D, Tul Noor Z, Rashed M. The role of soluble, insoluble fibers and their bioactive compounds in cancer: a mini review. Int J Food Sci Nutr. (2015) 6:1–11. doi: 10.4236/fns.2015.61001
11. Singh RK, Chang HW, Yan D, Lee KM, Ucmak D, Wong K. et al. Influence of diet on the gut microbiome and implications for human health. J Transl Med Epidemiol. (2017) 15:1–17. doi: 10.1186/s12967-017-1175-y
12. Wypych TP, Marsland BJ. Diet hypotheses in light of the microbiota revolution: new perspectives. Nutrients (2017) 9:E537. doi: 10.3390/nu9060537
13. Yang K, Li H, Dong J, Dong Y, Wang CZ. Expression profile of polyunsaturated fatty acids in colorectal cancer. World J Gastroenterol. (2015) 21:2405–12. doi: 10.3748/wjg.v21.i8.2405
14. Birchenough GMH, Johansson MEV, Gustafsson JK, Bergstrom JH, Hansson GC. New developments in goblet cell mucus secretion and function. Mucosal Immunol. (2015) 8:712–19. doi: 10.1038/mi.2015.32
15. Johansson MEV and Hansson GC. Immunological aspects of intestinal mucus and mucins. Nat Rev Immunol. (2015) 16:1–11. doi: 10.1038/nri.2016.88
16. Frosali S, Pagliari D, Gambassi G, Landolfi R, Pandolfi F, Cianci R. How the intricate interaction among toll-like receptors, microbiota, and intestinal immunity can influence gastrointestinal pathology. J Immunol. (2015) 2015:489821. doi: 10.1155/2015/489821
17. Nakatsu G, Li X, Zhou H, Sheng J, Wong SH, Wu WK. Gut mucosal microbiome across stages of colorectal carcinogenesis. Nat Commun. (2015) 6:8727. doi: 10.1038/ncomms9727
18. Martin R, Makino H, Yavuz AC, Ben-Amor K, Roelofs M, Ishikawa E. et al. Early-life events, including mode of delivery and type of feeding, siblings and gender, shape the developing gut microbiota. PLoS ONE (2016) 11:e0158498. doi: 10.1371/journal.pone.0158498
19. Mueller NT, Bakacs E, Combellick J, Grigoryan Z, Dominguez-Bello MG. The infant microbiome development: mom matters. Trends Mol Med. (2015) 21:109–17. doi: 10.1016/j.molmed.2014.12.002
20. Dominguez-Bello MG, Jesus-Laboy KM, Shen N, Cox LM, Amir A, Gonzalez A. et al. Partial restoration of the microbiota of cesarean-born infants via vaginal microbial transfer. Nat Med. (2016) 22:250–3. doi: 10.1038/nm.4039
21. Matamoros S, Gras-Leguen C, Le Vacon F, Potel G, La Cochetiere MF. Development of intestinal microbiota in infants and its impact on health. Cell Press (2013) 21:167–73. doi: 10.1016/j.tim.2012.12.001
22. Prideaux L, Kang S, Wagner J, Kamm MA. Impact of ethnicity geography and disease on the microbiota in health and infammatory bowel disease. Inflamm Bowel Dis. (2013) 19:2906–18. doi: 10.1097/01.MIB.0000435759.05577.12
23. Chong CW, Ahmad AF, Lim YAL, Teh CSJ, Yap IKS, Lee SC. Effect of ethnicity and socioeconomic variation to the gut microbiota composition among pre-adolescent in Malaysia. Sci Rep. (2015) 5:13338. doi: 10.1038/srep13338
24. Gupta VK, Paul S, Dutta C. Geography ethnicity or subsistence-specific variations in human microbiome composition and diversity. Front Microbiol. (2017) 8:1162. doi: 10.3389/fmicb.2017.01162
25. Koppel N, Rekdal VM, Balskus EP, Chemical transformation of xenobiotics by the human gut microbiota. Science (2017). 356:6344. doi: 10.1126/science.aag2770
26. Zaafar DK, Zaitone SA, Moustafa YM. Role of metformin in suppressing 1,2-dimethylhydrazine-induced colon cancer in diabetic and non-diabetic mice: effect on tumor angiogenesis and cell proliferation. PLoS ONE (2014) 9:e100562. doi: 10.1371/journal.pone.0100562
27. Cancer Research UK. Available online at: https://www.cancerresearchuk.org. (Accessed April 16, 2017).
28. Al-Saad E. Causes and effects of fast food. Biomed J Sci Tech Res. (2016) 5:279–80. Available online at: http://www.ijstr.org/final-print/apr2016/Causes-And-Effects-Of-Fast-Food.pdf (Accessed August 11, 2018).
29. Ekholm K, Gronberg C, Borjeson S, Bertero C. The next of kin experiences of symptoms and distress among patients with colorectal cancer: diagnosis and treatment affecting the life situation. Eur J Oncol Nurs. (2013) 17:125–30. doi: 10.1016/j.ejon.2012.05.003
30. Rohrl K, Guren MG, Miaskowski C, Cooper BA, Diep LM, Rustoen T. No differences in symptom burden between colorectal cancer patients receiving curative versus palliative chemotherapy. J Pain Symptom Manage. (2016) 52:539–47. doi: 10.1016/j.jpainsymman.2016.04.008
31. International Agency for Research on Cancer (IARC). Colorectal Cancer Overview. (2017). Available online at: https://old.cancer.org/acs/groups/cid/documents/webcontent/003047-pdf.pdf. (Accessed April 15, 2017).
32. International Agency for Research on Cancer (IARC). Treating Colorectal Cancer. (2017). Available online at: https://www.cancer.org/cancer/colon-rectal-cancer/treating.html. (Accessed April 15, 2017).
33. Toes-Zoutendijk E, Van Leerdam ME, Dekker E, Van Hees F, Penning C, Nagtegaal I. Real-time monitoring of results during first year of dutch colorectal cancer screening program and optimization by altering fecal immunochemical test cut off levels. Gastroenterology (2017) 152:767.e2–75. doi: 10.1053/j.gastro.2016.11.022
34. Arnold M, Sierra MS, Laversanne M, Soerjomataram I, Jemal A, Bray F. Global patterns and trends in colorectal cancer incidence and mortality. Gut (2017) 4:683–91. doi: 10.1136/gutjnl-2015-310912
35. Belcheva A, Martin A. Gut microbiota and colon cancer: the carbohydrate link. Mol Cell Oncol. (2015) 1:e96630. doi: 10.4161/23723548.2014.969630
36. Altobelli E, D'Aloisio F, Angeletti PM. Colorectal cancer screening in countries of European Council outside of the EU-28. World J Gastroenterol. (2016) 22:4946–57. doi: 10.3748/wjg.v22.i20.4946
37. Europa Colon. (2017). Colorectal Cancer. Health check barometer 2017. Available in: http://www.europacolon.com/Documents/Uploaded/305-Document-barometermaps2017.pdf. (Accessed September 7, 2017).
38. Ouakrim DA, Pizor C, Boniol M, Malvezzi M, Boniol M, Negri E. et al. Trends in colorectal cancer mortality in Europe: retrospective analysis of the WHO mortality database. BMJ (2015) 351:h4970. doi: 10.1136/bmj.h4970
39. Yeoh GK. In What Asia-Pacific Populations is CRC Screening Justified? (2015). Available online at: http://www.worldendo.org/wp-content/uploads/2016/04/02_CRC_Justified_Yeoh.pdf. (Accessed September 7, 2017).
40. Kato T, Alonso S, Muto Y, Noda H, Miyakura Y, Suzuki K. et al. Clinical characteristics of synchronous colorectal cancers in Japan. World J Surg Oncol. (2016) 14:272. doi: 10.1186/s12957-016-1027-x
41. Sano Y, Byeon JS, Li XB, Wong MCS, Chiu HM, Rerknimitr R. et al. Colorectal cancer screening of the general population in East Asia. Dig Endosc. (2016) 28:243–9. doi: 10.1111/den.12579
42. Veettil SK, Lim KG, Chaiyakunapruk N, Ching SM, Hassan MRA. Colorectal cancer in Malaysia: Its burden and implications for a multiethnic country. Asian J Surg. (2016) 40:1–9. doi: 10.1016/j.asjsur.2016.07.005
43. American Cancer Society. Available online at: https://www.cancer.org/ (Accessed September 7, 2017).
44. Murphy G, Devesa SS, Cross AJ, Inskip PD, McGlynn KA, Cook MB. Sex disparities in colorectal cancer incidence by anatomic subsite, race and age. Int J Cancer (2011) 128:1668–75. doi: 10.1002/ijc.25481
45. Kim SE, Paik HY, Yoon H, Lee JE, Kim N, Sung MK. Sex and gender specific disparities in colorectal cancer risk. World J Gastroenterol. (2015) 21:5167–75. doi: 10.3748/wjg.v21.i17.5167
46. Barone M, Tanzi S, Lofano K, Scavo MP, Guido R, Demarinis L, et al. Estrogens, phytoestrogens and colorectal neoproliferative lesions. Genes Nutr. (2008) 3:7–13. doi: 10.1007/s12263-008-0081-6
47. Nussler NC, Reinbacher K, Shanny N, Schirmeier A, Glanemann M, Neuhaus P. et al. Sex-specific differences in the expression levels of estrogen receptor subtypes in colorectal cancer. Gend Med. (2008) 5:209–17. doi: 10.1016/j.genm.2008.07.005
48. National Cancer Institute (INCA),. Estimativa 2016 – Síntese de Resultados e Comentários. (2016). Available online at: http://santacasadermatoazulay.com.br/wp-content/uploads/2017/06/estimativa-2016-v11.pdf (Accessed April 15, 2017).
49. Desai D, Shah S, Deshmukh A, Abraham P, Joshi A, Gupta T. et al. Colorectal cancers in ulcerative colitis from a low-prevalence area for colon cancer. World J Gastroenterol. (2015) 21:3644–49. doi: 10.3748/wjg.v21.i12.3644
50. Axelrad JE, Lichtiger S, Yajnik V. Inflammatory bowel disease and cancer: the role of inflammation, immunosuppression, and cancer treatment. World J Gastroenterol. (2016) 22:4794–801. doi: 10.3748/wjg.v22.i20.4794
51. Kim ER, Chang DK. Colorectal cancer in inflammatory bowel disease: the risk, pathogenesis, prevention and diagnosis. World J Gastroenterol. (2014) 20:9872–881. doi: 10.3748/wjg.v20.i29.9872
52. Yamamoto M and Matsumoto S. Gut microbiota and colorectal cancer. Genes Environ. (2016) 38:11. doi: 10.1186/s41021-016-0038-8
53. Papastergiou V, Karatapanis S, Georgopoulos SD. Helicobacter pyloriand colorectal neoplasia: Is there a causal link? World J Gastroenterol. (2016) 22:649–58. doi: 10.3748/wjg.v22.i2.649
54. Paritsky M, Pastukh N, Brodsky D, Isakovich N, Peretz A. Association of Streptococcus bovispresence in colonic content with advanced colonic lesion World J Gastroenterol. (2015) 21:5663–67. doi: 10.3748/wjg.v21.i18.5663
55. Amarnani R, Rapose A. Colon cancer and enterococcus bacteremia co-affection: a dangerous alliance. J Infect Public Health (2017) 16:30148. doi: 10.1016/j.jiph.2016.09.009
56. Khan S. Potential role of Escherichia coli DNA mismatch repair proteins in colon cancer. Crit Rev Oncol Hematol. (2015) 96:475–82. doi: 10.1016/j.critrevonc.2015.05.002
57. Jessamy K, Ojevwe FO, Ubagharaji E, Sharma A, Anozie O, Gilman CA, et al. Clostridium septicum: an unusual link to a lower gastrointestinal bleed. Case Rep Gastroenterol. (2016) 10:489–93. doi: 10.1159/000448881
58. Purcell RV, Pearson J, Aitchison A, Dixon L, Frizelle FA, Keenan JI. Colonization with enterotoxigenic Bacteroides fragilisis associated with early-stage colorectal Neoplasia. PLoS ONE (2017) 12:e0171602. doi: 10.1371/journal.pone.0171602
59. Boleij A, Hechenbleikner EM, Goodwin AC, Badani R. Stein EM, Lazarev MG. et al. The Bacteroides fragilis toxin gene is prevalent in the colon mucosa of colorectal cancer patients. Clin Inf Dis. (2015) 60:208–15. doi: 10.1093/cid/ciu787
60. Wehkamp J, Chu H, Shen B, Feathers RW, Kays RJ, Lee SK. et al. Paneth cell antimicrobial peptides: topographical distribution and quantification in human gastrointestinal tissues. FEBS Lett. (2006) 580:5344–350. doi: 10.1016/j.febslet.2006.08.083
61. Dowling JK, Mansell A. Toll-like receptors: the Swiss army knife of immunity and vaccine development. Clin Transl Immunol. (2016) 5:e85. doi: 10.1038/cti.2016.22
62. Narayanan KB, Park HH. Toll/interleukin-1 receptor (TIR) domain-mediated cellular signaling pathways. Apoptosis (2015) 20:169–209. doi: 10.1007/s10495-014-1073-1
64. Jiang Y, Lu X, Man C, Han L, Shan Y, Qu X, et al. Lactobacillus acidophilus induces cytokine and chemokine production via NF-κB and p38 mitogen-activated protein kinase signaling pathways in intestinal epithelial cells. Clin Vaccine Immunol. (2012) 19:603–8. doi: 10.1128/CVI.05617-11
65. Yiu JHC, Dorweiler B, Woo CW. Interaction between gut microbiota and toll-like receptor: from immunity to metabolism. J Mol Med Clin Appl. (2017) 95:13–20. doi: 10.1007/s00109-016-1474-4
66. Park MH and Hong JT. Roles of NF-κB in cancer and inflammatory diseases and their therapeutic approaches. Cells (2016) 5:5020015. doi: 10.3390/cells5020015
67. Ijssennagger N, Belzer C, Hooiveld GJ, Dekker J, van Mil SWC, Müller M. et al. Gut microbiota facilitates dietary heme-induced epithelial hyperproliferation by opening the mucus barrier in colon. Proc. Natl. Acad. Sci. U.S.A. (2015) 112:10038–10043. doi: 10.1073/pnas.1507645112
68. Bultman SJ. Interplay between diet, gut microbiota, epigenetic events, and colorectal cancer. Mol Nutr Food Res. (2017) doi: 10.1002/mnfr.201500902. [Epub ahead of print].
69. Shaw KA, Bertha M, Hofmekler T, Chopra P, Vatanen T, Srivatsa A. Dysbiosis, inflammation, and response to treatment: a longitudinal study of pediatric subjects with newly diagnosed inflammatory bowel disease. NPJ Genom Med. (2016) 8:75. doi: 10.1186/s13073-016-0331-y
70. Sun J and Kato I. Gut microbiota, inflammation and colorectal cancer. Genes Dis. (2016) 3:130–43. doi: 10.1016/j.gendis.2016.03.004
71. Hu S, Wang L, Jiang Z. Dietary additive probiotics modulation of the intestinal microbiota. Protein Pept Lett. (2017) 24:382–87. doi: 10.2174/0929866524666170223143615
72. Roberfroid M, Gibsin GR, Hoyles L, McCartney AL, Rastall R, Rowland I. Prebiotic effects: metabolic and health benefits. Br J Nutr. (2010) 104:S1–63. doi: 10.1017/S0007114510003363
73. Liu X, Cau S, Zhang Z. Modulation of gut microbiota–brain axis by probiotics, prebiotics, and diet. J Agric Food Chem. (2015) 63:7885–95. doi: 10.1021/acs.jafc.5b02404
74. Dinesh KD, Renuka, Puniya M, Shandiya UK, Dhewa T, Kumar N. Gut microbiota modulation and its relationship with obesity using prebiotic fibers and probiotics: a review. Front Microbiol. (2017) 8:563. doi: 10.3389/fmicb.2017.00563
75. Qin J, Li R, Raes J, Arumugam M, Burgdorf KS, Manichanh C. A human gut microbial gene catalogue established by metagenomic sequencing. Nature (2010) 464:59–65. doi: 10.1038/nature08821
76. Daliria EBM, Lee BH. New perspectives on probiotics in health and disease. J Adv Nutr Hum Metab. (2015) 4:56–65. doi: 10.1016/j.fshw.2015.06.002
77. Ettinger G, MacDonald K, Reid G, Burton JP. The influence of the human microbiome and probiotics on cardiovascular health. Gut Microb. (2014) 5:719–28. doi: 10.4161/19490976.2014.983775
78. Harsa ST. Functional food products and ingredients for gut health. J Nutr Ther. (2016) 5:27–8. Available online at: http://www.lifescienceglobal.com/pms/index.php/jnt/article/download/3974/2287. (Access 11 August 2018).
79. Khalid G, Singh AK, Jabeen R. Nutraceuticals and functional foods: the foods for future world. Crit Rev Food Sci Nutr. (2016) 56:2617–27. doi: 10.1080/10408398.2014.903384
80. Agriculture and Agri-Food Canada. Functional foods and natural health products sector. (2017). Available online at: <http://www.agr.gc.ca/eng/industry-markets-and-trade/market-information-by-sector/functional-foods-and-natural-health-products/?id=1170856376710> (Accessed September 06, 2017).
81. Tsoukas MA, Ko BJ, Witte TR, Dincer F, Hardman WE, Mantzoros CS. Dietary walnut suppression of colorectal cancer in mice: mediation by miRNA patterns and fatty acid incorporation. J Nutr Biochem. (2015) 26:776–83. doi: 10.1016/j.jnutbio.2015.02.009
82. Aghajanpour M, Nazer MR, Obeidavi Z, Akbari M, Ezati P, Kor NM. Functional foods and their role in cancer prevention and health promotion: a comprehensive review. Am J Cancer Res. (2017) 7:740–769.
83. Brahe LK, Le Chatelier E, Prifti E, Pons N, Kennedy S, Blaedel T. Dietary modulation if the gut microbiota – a randomised controlled trial in obese postmenopausal women. Br J Nutr. (2015) 114:406–17. doi: 10.1017/S0007114515001786
84. Chishty S, Bissu M. Health benefits and nutritional value of flaxseed - a review. Indian J Appl Res. (2016) 6:243–5.
85. Shen RL, Wang Z, Dong JL, Xiang QS, Liu YQ. Effects of oat soluble and insoluble β-glucan on 1,2-dimethylhydrazine-induced early colon carcinogenesis in mice. Food Agric Immunol. (2016) 19: 657–66. doi: 10.1080/09540105.2016.1148664
86. Ko KP, Yeo Y, Yoon JH, Kim CS, Tokudome S, Ngoan LT. Plasma phytoestrogens concentration and risk of colorectal cancer in two different Asian populations. Clin Nutr. (2017) doi: 10.1016/j.clnu.2017.07.014
87. Marín L, Miguélez EM, Villar CJ, Lombó F. Bioavailability of dietary polyphenols and gut microbiota metabolism: antimicrobial properties. Biomed Res Int. (2015) 2015:905215. doi: 10.1155/2015/905215
88. Li YH, Niu YB, Sun Y, Zhang F, Liu CX, Fan L, et al. Role of phytochemicals in colorectal cancer prevention. World J Gastroenterol. (2015) 21:9262–72. doi: 10.3748/wjg.v21.i31.9262
89. Lu K, Mahbub R, Fox JG. Xenobiotics: interaction with the intestinal microflora. ILAR J. (2015) 56:218–27. doi: 10.1093/ilar/ilv018
90. Bommaredd U, Zhang XY, Kaushik RS, Dwivedi C. Effects of components present in flaxseed on human colon adenocarcinoma Caco-2 cells: possible mechanisms of flaxseed on colon cancer development in animals. Drug Discov Ther. (2010) 4:184–9. Available online at: http://www.ddtjournal.com/getabstract.php?id=321 (Accessed August 11, 2018).
91. Singh KK, Mridula D, Rehal J, Barnwal P. Flaxseed: a potential source of food, feed and fiber. Crit. Rev. Food Sci. Nutr. (2011) 51:210–22. doi: 10.1080/10408390903537241
92. Xu J, Tian G, Ma C, Gao H, Chen C, Yang W, et al. Flaxseed lignan secoisolariciresinol diglucoside ameliorates experimental colitis induced by dextran sulphate sodium in mice. J Func Foods (2016) 26:187–95. doi: 10.1016/j.jff.2016.07.013
93. Busstra MC, Siezen CLE, Grubben MJAL, Van Kranen HJ, Nagengast FM, et al. Tissue levels of fish fatty acids and risk of colorectal adenomas: a case-control study (Netherlands). Cancer Causes and Control (2003) 14:269–276. Available online at: http://www.jstor.org/stable/3553976 (Accessed August 11, 2018).
94. Lee JY, Sim TB, Lee JE, Na HK. Chemopreventive and chemotherapeutic effects of fish oil derived omega-3 polyunsaturated fatty acids on colon carcinogenesis. Clin Nutr. (2017) 6:147–60. doi: 10.7762/cnr.2017.6.3.147
95. Tedelind S, Westberg F, Kjerrulf M, Vidal A. Anti-inflammatory properties of the short-chain fatty acids acetate and propionate: a study with relevance to inflammatory bowel disease. World J Gastroenterol. (2007) 13:2826–32. doi: 10.3748/wjg.v13.i20.2826
96. D'Eliseo D, Velotti F. Omega-3 fatty acids and cancer cell cytotoxicity: implications for multi-targeted cancer therapy. J Clin Med. (2016) 5:E15. doi: 10.3390/jcm5020015
97. Shahidi F. Ho, CT. Antioxidant measurement and applications: an overview. ACS Symp Ser. (2007) 956:2–7. doi: 10.1021/bk-2007-0956.ch001
98. Tangvarasittichai S. Oxidative stress, insulin resistance, dyslipidemia and type 2 diabetes mellitus. World J Diabetes (2015) 6:456–80. doi: 10.4239/wjd.v6.i3.456
99. Shin A, Lee J, Lee J, Park MS, Park JW, et al. Isoflavone and soyfood intake and colorectal cancer risk: a case-control study in Korea. PLoS ONE (2015) 10:e0143228. doi: 10.1371/journal.pone.0143228
100. Pituch-Zdanowska A, Banaszkiewicz A, Albrecht P. The role of dietary fibre in inflammatory bowel disease. Prz Gastroenterol. (2015) 10:135–141. doi: 10.5114/pg.2015.52753
101. Moussa L, Bézirard V, Salvador-Cartier C, Bacquié V, Lencina C, Lévêque M, et al. A Low Dose of fermented soy germ alleviates gut barrier injury, hyperalgesia and faecal protease activity in a rat model of inflammatory bowel disease. PLoS ONE (2012) 7:e49547. doi: 10.1371/journal.pone.0049547
102. Power KA, Leep D, Zarepoor L, Monk JM, Wu W, Tsao R. et al. Dietary flaxseed modulates the colonic microenvironment in healthy C57Bl/6 male mice which may alter susceptibility to gut-associated diseases J Nutr Biochem. (2015) 28:61–9. doi: 10.1016/j.jnutbio.2015.09.028
103. Cavallini DCU, Suzuki JY, Abdalla DS, Vendramini RC, Pauly-Silveira ND, Roselino MN, Pinto RA. et al. Influence of a probiotic soy product on fecal microbiota and its association with cardiovascular risk factors in an animal model. Lipids Health Dis. (2011) 10:126. doi: 10.1186/1476-511X-10-126
104. Walugembe M, Hsieh JC, Koszewski NJ, Lamont SJ, Persia ME, Rothschild MF. Effects of dietary fiber on cecal short-chain fatty acid and cecal microbiota of broiler and laying-hen chicks. Genet Genomics (2015) 94, 2351–59. doi: 10.3382/ps/pev242
105. Hosseini A and Ghorbani A. Cancer therapy with phytochemicals: evidence from clinical studies. Avicenna J Phytomed. (2015) 5:84–97.
106. Kotecha R, Takami A, Espinoza JL. Dietary phytochemicals and cancer chemoprevention: a review of the clinical evidence. Oncotarget. (2016) 7:52517–29. doi: 10.18632/oncotarget.9593
107. Ferreira RS, Hiane PA, Guimarães RCA, Ramos MIL, Demarque DP, Meira JEC. Physicochemical, microbiological and sensory evaluation of a bioactive food blend. Food Sci Technol. (2014) 34:609–15. doi: 10.1590/1678-457x.6406
108. Ferreira RS, Mendonça LABM, Guimarães RCA, David MM, Hiane PA. Nutritional intervention with a bioactive food compound in HIV/AIDS patients with hypertriglyceridemia: case reports. Int J Dev Res. (2017) 07:17440–44. Available online at: https://www.journalijdr.com/nutritional-intervention-bioactive-food-compound-hiv-aids-patients-hypertriglyceridemia-case-reports (Accessed August 11, 2018).
109. Ferreira RS, Guimarães RCA, Pontes ERJC, Mendonça LABM, Freitas KC, Hiane PA. Effectiveness of a bioactive food compound in anthropometric measures of individuals with HIV/AIDS: a nonrandomized trial. PLoS ONE (2018) 13:e0191259. doi: 10.1371/jornal.pone.0191259
110. Jandacek RJ. Linoleic Acid: a nutritional quandary. Healthcare (2017) 5:25. doi: 10.3390/healthcare5020025
111. Dahham SS, Majid AMA. The impact of life style and nutritional components in primary prevention of colorectal cancer. J Appl Pharm Sci. (2016) 6:237–44. doi: 10.7324/JAPS.2016.60935
112. Rosa LS, Silva NJA, Soares NCP, Monteiro MC, Teodoro AJ. Anticancer properties of phenolic acids in colon cancer – a review. J Nutr Food Sci. (2016) 6:468. doi: 10.4172/2155-9600.1000468
113. Nguyen TLA, Vieira-Silva S, Liston A, Raes J. How informative is the mouse for human gut microbiota research? Dis Models Mech. (2015) 8:1–16. doi: 10.1242/dmm.017400
114. Scheithauer TPM, Dallinga-Thie GM, de Vos WM, Nieuwdorp M, van Raalte DH. Causality of small and large intestinal microbiota in weight regulation and insulin resistance. Mole Metabol. (2016) 5:759–70. doi: 10.1016/j.molmet.2016.06.002
115. Meijnikman AS, Gerdes VE, Nieuwdorp M, Herrema H. Evaluating causality of gut microbiota in obesity and diabetes in humans. Endocr. Rev. (2018) 39:133–153. doi: 10.1210/er.2017-00192
116. Irman M, Ahmad N, Anjum FM, Khan MK, Mushtag Z, Nadeem M, et al. Potential protective properties of flax lignan secossolariciresinol diglucósido. Nutr J. (2015) 14:71. doi: 10.1186/s12937-015-0059-3
117. Nagel JM, Brinkoetter M, Magkos F, Liu X, Chamberland JP. Dietary walnuts inhibit colorectal cancer growth in mice by suppressing angiogenesis. Nutrition (2012) 28:67–75. doi: 10.1016/j.nut.2011.03.004
118. Lima RK, Cardoso MG, Andrade MA, Guimarães PL, Batista LR, Nelson DL. Bactericidal and antioxidant activity of essential oils from Myristica fragrans Houtt and Salvia microphylla H.B.K. J. Am. Oil Chemists' Soc. (2012) 89:523–8. doi: 10.1007/s11746-011-1938-1
119. Dastjerdi MN, Kavoosi F, Valiani A, Esfandiari E, Sanaei M, Sobhanian S, et al. Inhibitory effect of genistein on PLC/PRF5 hepatocellular carcinoma cell line. Int J Prev Med. (2015) 6:54. doi: 10.4103/2008-7802.158914
120. Fu M, Song Y, Wen Z, Lu X, Cui L. Inositol hexaphosphate and inositol inhibit colorectal cancer metastasis to the liver in BALB/c mice. Nutrients (2016) 8:E286. doi: 10.3390/nu8050286
121. Yang G, Shu XO, Li H, Chow WH, Cai H, Zhang X. Prospective cohort study of soy food intake and colorectal cancer risk in women. Am J Clin Nutr. (2009) 89:577–83. doi: 10.3945/ajcn.2008.26742
122. Gutte KB, Sahoo AK, Ranveer RC. Bioactive components of flaxseed and its health benefits. Int J Pharm Sci Rev Res. (2015) 31:42–51.
123. Andres-Franch M, Aliana AG, Hellin VS, Ochoa E, Illan EH, Garcia PL. et al. Streptococcus gallolyticus infection in colorectal cancer and association with biological and clinical factors. PLoS ONE (2017) 12:e0174305. doi: 10.1371/journal.pone.0174305
124. Kumar N, Pruthi V. Potential applications of ferulic acid from natural sources. Biotechnol. Rep. (2014) 4:86–93. doi: 10.1016/j.btre.2014.09.002
125. Viljoen KS, Dakshinamurthy A, Goldberg P, Blackburn JM. Quantitative profiling of colorectal cancer associated bacteria reveals associations between Fusobacterium spp., enterotoxigenic Bacteroides fragilis (ETBF) and clinicopathological features of colorectal cancer. PLoS ONE (2015) 10:e0119462. doi: 10.1371/journal.pone.0119462
126. Cheshmehkani A, Senatorov IS, Kandi P, Singh M, Britt A, Hayslett R. Fish oil and flax seed oil supplemented diets increase FFAR4 expression in the rat colon. Inflamm Res. (2015) 64:809–15. doi: 10.1007/s00011-015-0864-3
127. Wang P, Chen H, Zhu Y, McBrinde J, Fu J, Sang S. Oat Avenanthramide-C (2c) is biotransformed by mice and the human microbiota into bioactive metabolites. J Nutr. (2015) 239–45. doi: 10.3945/jn.114.206508
128. Alphonse PAS, Aloko RE, Anti-carcinogenic and anti -metastatic effects of flaxseed lignan secoisolariciresinol diglucoside (SDG). Discov Phytomed. (2015) 2:12–7. doi: 10.15562/phytomedicine.2015.24
129. Ajdarkosh H, Tameshkel FS, Sohrabi MR, Niya MHK, Hemmasi G, Amirkalali B. et al. Association of Helicobacter pylori infection with colon polyp and colorectal cancer. Br J Med Med Res. (2016) 16:1–6. doi: 10.4021/jocmr880w
130. Fantini M, Benvenuto M, Masuelli L, Frajese GV, Tresoldi I, Modesti A, et al. in vitro and in vivo antitumoral effects of combinations of polyphenols, or polyphenols and anticancer drugs: perspectives on cancer treatment. Int J Mol Sci. (2015) 16:9263–82. doi: 10.3390/ijms16059236
131. Tsai CE, Chiu CT, Rayner CK, Wu KL, Chiu YC, Hu ML. Associated factors in Streptococcus bovis bacteremia and colorectal cancer. Kaohsiung J Med Sci. (2016) 32:196–200. doi: 10.1016/j.kjms.2016.03.003
132. Mima K, Cao Y, Chan AT, Qian ZR, Nowak JA, Masugi Y. Fusobacterium nucleatum in colorectal carcinoma tissue according to tumor location. Clin Transl Gastroenterol. (2016) 7:e200. doi: 10.1038/ctg.2016.53
133. Bonnet M, Buc E, Sauvanet P, Darcha C, Dubois D, Pereira B, et al. Colonization of the human gut by E. coli and colorectal cancer risk. Clin Cancer Res. (2014) 20:859–67. doi: 10.1158/1078-0432.CCR-13-1343
134. Magdy A, Elhadidy M, Ellatif MA, Nakeeb AE, Abdallah E, Thabet W, et al. Enteropathogenic Escherichia coli (EPEC): does it have a role in colorectal tumourigenesis? a prospective cohort study. Int J Surg. (2015) 18:169–73. doi: 10.1016/j.ijsu.2015.04.077
135. Raju J, Bielecki A, Caldwell D, Lok E, Taylor M, Kapal K, et al. Soy isoflavones modulate azoxymethane-induced rat colon carcinogenesis exposed pre- and postnatally and inhibit growth of DLD-1 human colon adenocarcinoma cells by increasing the expression of estrogen Receptor-b. J Nutr. (2009) 139:474–81. doi: 10.3945/jn.108.099200
Keywords: functional foods, intestinal neoplasm, gut microbiota, bioactive compounds, dysbacteriosis
Citation: Mendonça LABM, dos Santos Ferreira R, de Cássia Avellaneda Guimarães R, de Castro AP, Franco OL, Matias R and Carvalho CME (2018) The Complex Puzzle of Interactions Among Functional Food, Gut Microbiota, and Colorectal Cancer. Front. Oncol. 8:325. doi: 10.3389/fonc.2018.00325
Received: 21 April 2018; Accepted: 30 July 2018;
Published: 05 September 2018.
Edited by:
Michael J. Wargovich, The University of Texas Health Science Center at San Antonio, United StatesReviewed by:
Maria Marino, Università degli Studi Roma Tre, ItalySudipto Das, Royal College of Surgeons in Ireland, Ireland
Copyright © 2018 Mendonça, dos Santos Ferreira, de Cássia Avellaneda Guimarães, de Castro, Franco, Matias and Carvalho. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Lígia A. B. M. Mendonça, bG1lbmRvbmNhbnV0cmlAZ21haWwuY29t
Rosângela dos Santos Ferreira, cm9zYW5nZWxhLmZlcnJlaXJhQHVmbXMuYnI=