 Lizhe Zhu
Lizhe Zhu Wei Wu
Wei Wu Siyuan Jiang1
Siyuan Jiang1 Shibo Yu
Shibo Yu Jianjun He
Jianjun He Yu Ren
Yu Ren Bin Wang
Bin Wang- 1Department of Breast Surgery, The First Affiliated Hospital of Xi’an Jiaotong University, Xi’an, China
- 2Department of Neurosurgery, The First Affiliated Hospital of Xi’an Jiaotong University, Xi’an, China
Introduction: The PINK1 gene encodes a serine/threonine protein kinase that localizes to mitochondria and has usually been considered to protect cells from stress-induced mitochondrial dysfunction. PINK1 mutations have been observed to lead to autosomal recessive Parkinson’s disease. However, the immunological and prognostic roles of PINK1 across cancers remain unclear.
Material and method: In the current study, we used multiple databases, including Oncomine, PrognoScan, Kaplan-Meier Plotter, GEPIA, TIMER, and cBioportal, to investigate the PINK1 expression distribution and its immunological and prognostic role across cancers.
Results and discussion: Bioinformatics data revealed that the mRNA expression of PINK1 was downregulated in most tumors. Although there was a significant prognostic value of PINK1 expression across cancers, PINK1 played a protective or detrimental role in different kinds of cancers. Liver hepatocellular carcinoma and lung squamous cell carcinoma were selected as representative cancer types for further exploration. We found that PINK1 always played a protective role in liver hepatocellular carcinoma patients in the stratified prognostic analyses of clinicopathological characteristics. There were contradictory results between liver hepatocellular carcinoma and lung squamous cell carcinoma in the correlations of PINK1 expression with immune infiltration, including infiltration of B cells, CD8+ T cells, CD4+ T cells, macrophages, neutrophils, and dendritic cells. Furthermore, specific markers of B cells and CD8+ T cells also exhibited different PINK1-related immune infiltration patterns. In addition, there was a significant association between PINK1 copy number variations and immune infiltrates across cancers.
Conclusion: The mitophagy-related protein PINK1 can work as a biomarker for prognosis and the immune response across cancers.
Introduction
Mitochondria are double-membraned and highly dynamic organelles that play an important role in eukaryotic cells and cellular metabolism (1). Mitochondria are eliminated through a kind of autophagy known as mitophagy during certain developmental conditions or damage (2). It has been proven that mitophagy is helpful in eliminating damaged or old mitochondria to maintain cellular integrity and is beneficial for cellular homeostasis (3). In addition, mitophagy is also involved in differentiation and developmental processes, including red blood cell production and muscle differentiation (4, 5). However, the detailed role of mitophagy in tumorigenesis and tumor progression remains unknown.
The PINK1 gene encodes a serine/threonine protein kinase that localizes to mitochondria and has generally been considered to protect cells from stress-induced mitochondrial dysfunction. PINK1 mutations have been found to lead to autosomal recessive Parkinson’s disease. Research has shown that mitophagy might prevent tumorigenesis by eliminating dysfunctional mitochondria (6). Meanwhile, reports have reflected that the PINK1 kinase can regulate glycolysis and the Warburg effect by controlling mitophagy. Furthermore, PINK1 deficiency reprograms glucose metabolism through HIF1α to maintain cell proliferation and even cancer growth (7). Therefore, PINK1 deletion might be associated with carcinogenesis due to mitophagy dysfunction. However, the role of PINK1 across cancers remains unclear. There are no completed studies on the prognostic value of PINK1 across cancers.
In addition, there are also complex interactions between tumors and their microenvironment. Immune cell infiltration is an essential component of the tumor environment. The tumor microenvironment contains innate immune cells and adaptive immune cells, including neutrophils, macrophages, natural killer cells, and dendritic cells. Cancer cells are under the scrutiny of immune cells all their lives, and only if the immune cells fail to eliminate the preneoplastic cells can cancer develop and progress. Currently, different kinds of effective chemotherapies and radiotherapies partially act by activating the immune response, which restores immunosurveillance (8). With the development of immunotherapy, potential targets have been gradually discovered. For example, studies have shown that PD-L1 expression in systemic immune cell populations is a potential predictive biomarker of responses to PD-L1/PD-1 blockade therapy in lung cancer (9). Angiopoietin-2 can also work as a biomarker and target for immune checkpoint therapy (10). However, there are still patients who respond poorly to immunotherapies. Therefore, it is still necessary to further discover more specific or general immune biomarkers in cancer therapy.
In this study, we combined data from different databases, including Oncomine, PrognoScan, Kaplan-Meier Plotter, GEPIA, TIMER and cBioportal, to explore the role of PINK1 expression in prognosis and the immune response. The findings from this study indicated that PINK1 influenced the prognosis of patients with cancers and might probably via its interaction with infiltrating immune cells.
Method
Oncomine Database and the Human Protein Atlas
Oncomine is an online microarray database with large datasets and over eighty thousand samples of different kinds of cancer (11). This database was employed to analyze PINK1 mRNA expression in different kinds of human cancers. The filters included the gene name, “PINK1”, cancer vs. normal analysis, and mRNA data type. The thresholds were set as the following criteria: gene rank: 10%, fold change: 2, and p-value: 0.001. The datasets with statistically significant differences were recorded. The Human Protein Atlas is a Swedish-based program initiated in 2003 with the aim to map all the human protein in cells, tissues and organs using integration of various omics technologies, including antibody-based imaging, mass spectrometry-based proteomics, transcriptomics and systems biology. It consists of six separate parts, and each focusing on a particular aspect of the genome-wide analysis of the human proteins.
PrognoScan, Kaplan-Meier Plotter, and GEPIA
PrognoScan is a new database for the meta-analysis of the genes with prognostic value (12). It analyzes the relationship between specific gene expression and cancer patient outcomes, such as overall survival (OS) and disease-free survival (DFS), across a large collection of publicly available cancer microarray datasets. Forest plots were drawn with GraphPad Prism 8 to summarize the survival analysis. Kaplan-Meier Plotter is an online tool for meta-analyses based on biomarker assessment (13). It is able to assess the survival of patients with different kinds of cancers based on large sample datasets. GEPIA is a newly developed interactive web server for analyzing the RNA sequencing expression data of 9,736 tumors and 8,587 normal samples from the TCGA and GTEx projects by using a standard processing pipeline (14). We used the above three tools to analyze the prognostic value of PINK1 expression in different kinds of human cancers. Hazard ratios (HRs) with 95% confidence intervals (CIs) and p-values were collected. P <0.05 was considered statistically significant.
TIMER and R&D Systems Immune Cell Markers
The TIMER web server is a comprehensive resource for the systematic analysis of immune infiltrates across diverse cancer types (15, 16). The TIMER algorithm can estimate the abundances of six immune infiltrates, including B cells, CD4+ T cells, CD8+ T cells, neutrophils, macrophages, and dendritic cells. We used TIMER to explore the differential PINK1 expression between tumor and adjacent normal tissue across TCGA tumors. Distributions of PINK1 expression levels were adjusted to log2(TPM) data and displayed using box plots, and the statistical significance of differential expression was evaluated using the Wilcoxon test. In addition, we further analyzed the correlation between PINK1 expression and the abundance of the above six immune infiltrates across cancers. The scatterplots show the purity-corrected partial Spearman’s rho value and statistical significance. To investigate the correlation between PINK1 expression and immune cells, we explored the correlation between the specific gene expression of differential immune cells and PINK1 expression. We selected the representative gene makers of immune cells noted on the R&D Systems website (https://www.rndsystems.com/cn/resources/cell-markers/immune-cells). They were CD19, CD20, and CD3 of B cell; CD8A and CD8B of CD8+ T cell; CXCR5, ICOS and BCL-6 of follicular helper T cell; IL12RB2, WSX-1, and T-BET of T helper cell 1; CCR3, STAT6, and GATA-3 of T helper cell 2; TGFBR2, IRF4, and PU.1 of T helper cell 9; IL-21R, IL-23R, and STAT3 of T helper cell 17; CCR10 and AHR of T helper cell 22; FOXP3, CCR8, and CD25 of regulatory T cell; PD-1 and CTLA4 of T cell exhaustion; CD68 and CD11b of macrophage; NOS2 and ROS of M1 macrophage; ARG1 and MRC1 of M2 macrophage; HLA-G, CD80, and CD86 of tumor associated macrophage; CD14 and CD16 of monocyte; XCL1, KIR3DL1, and CD7 of natural killer cell; CD15 and MPO of neutrophil and CD1C and CD141 of dendritic cell. We displayed the PINK1 expression levels on the X-axis and the expression of immune cell-related gene markers on the Y-axis to draw scatterplots. All the gene expression levels in TIMER were adjusted to log2(TPM) data. Meanwhile, we explored the association of immune infiltration levels among cancers with different somatic copy number alterations (SCNAs) affecting PINK1 expression. The SCNAs in TIMER include deep deletions, arm-level deletions, diploid/normal alterations, arm-level gains and high amplifications. Box plots are presented to show the distributions of each immune subset at each copy number status in all cancers. The infiltration level for each SCNA category was compared with that for normal using a two-sided Wilcoxon rank-sum test.
cBioPortal
The cBioPortal for Cancer Genomics contains large-scale cancer genomics data that can be visualized, downloaded and analyzed. We selected the “TCGA pancancer atlas studies” including 32 studies and 10,967 samples to further explore PINK1 alterations across cancers.
Statistical Analysis
Our results from Oncomine database were analyzed by t-tests and showed in P values, gene rank: 10%, fold change: 2, and P value: 0.001 were used as the criteria to set thresholds. In PrognoScan, the univariate Cox regression model was used to calculate the HR and P value. In GEPIA and Kaplan-Meier Plotter, log rank test was used to calculate the HR and its P value in order to compare survival curves. Furthermore, we used Spearman correlation to analyze the correlation among different gene expression. Above all, P <0.05 was set as statistically significant if there was no special annotation.
Result
PINK1 mRNA and Protein Expression Level Across Cancers
Figure 1 shows the transcription levels of PINK1 in different types of human cancers. We identified PINK1 expression across cancers and compared its mRNA expression with that in corresponding normal tissues based on data from the Oncomine database (Figure 1A). The results showed that PINK1 expression was lower in several cancer groups than in normal tissues, including brain, breast, colorectal, esophageal, head and neck, liver and ovarian cancers as well as leukemia and melanoma. However, the mRNA expression of PINK1 was significantly upregulated in lymphoma. The details of PINK1 expression in the above cancers are shown in Table 1. Over 20 unique datasets revealed that PINK1 had lower mRNA expression in different kinds of cancers than in normal tissues. In contrast, only Compagno reported that overexpression of PINK1 was found in diffuse large B cell lymphoma compared with normal tissues, with a P = 1.63E-17 and a fold change = 2.109 (Table 1).
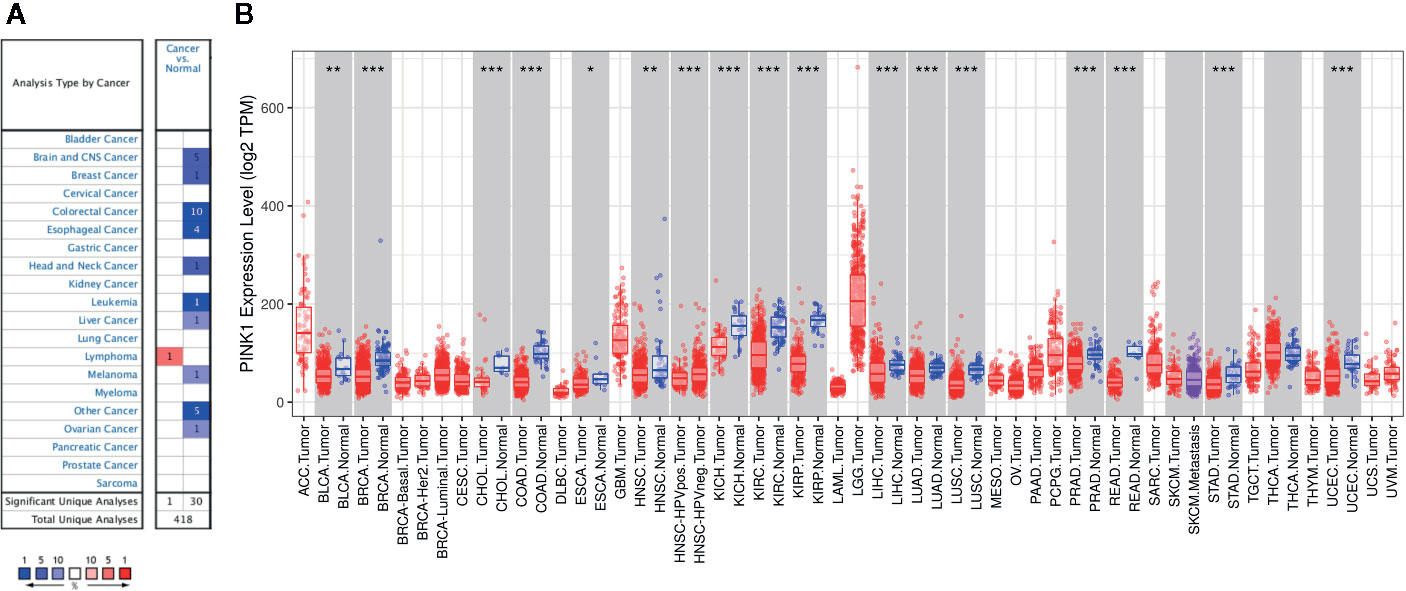
Figure 1 PINK1 expression levels in different types of human cancers. (A) The transcription levels of PINK1 in different types of human cancers. The figure is generated from ONCOMINE with exact thresholds (P-value: 0.001; fold change: 2; gene rank: top 10%). The cell number represents the dataset number that meets all of the thresholds with the color blue for under-expression and color red for over-expression. Cell color is determined by the best gene rank percentile for the analyses within the cell. (B) PINK1 expression levels in different types of human cancers from TCGA data in TIMER. *P < 0.05, **P < 0.01, ***P < 0.001. CNS, central nervous system; ACC, adrenocortical carcinoma; BLCA, bladder urothelial carcinoma; BRCA, breast invasive carcinoma; CESC, cervical and endocervical cancer; CHOL, cholangiocarcinoma; COAD, colon adenocarcinoma; DLBC, diffuse large B-cell lymphoma; ESCA, esophageal carcinoma; GBM, glioblastoma multiforme; HNSC, head and neck cancer; HNSC-HPVpos, head and neck cancer-HPV positive; HNSC-HPVneg, head and neck cancer-HPV negative; KICH, kidney chromophobe; KIRC, kidney renal clear cell carcinoma; KIRP, kidney renal papillary cell carcinoma; LAML, acute myeloid leukemia; LGG, lower grade glioma; LIHC, liver hepatocellular carcinoma; LUAD, lung adenocarcinoma; LUSC, lung squamous cell carcinoma; MESO, mesothelioma; OV, ovarian serous cystadenocarcinoma; PAAD, pancreatic adenocarcinoma; PCPG, pheochromocytoma and paraganglioma; PRAD, prostate adenocarcinoma; READ, rectum adenocarcinoma; SARC, sarcoma; SKCM, skin cutaneous melanoma; STAD, stomach adenocarcinoma; TGCT, testicular germ cell tumors; THCA, thyroid carcinoma; THYM, thymoma; UCEC, uterine corpus endometrial carcinoma; UCS, uterine carcinosarcoma; UVM, uveal melanoma.
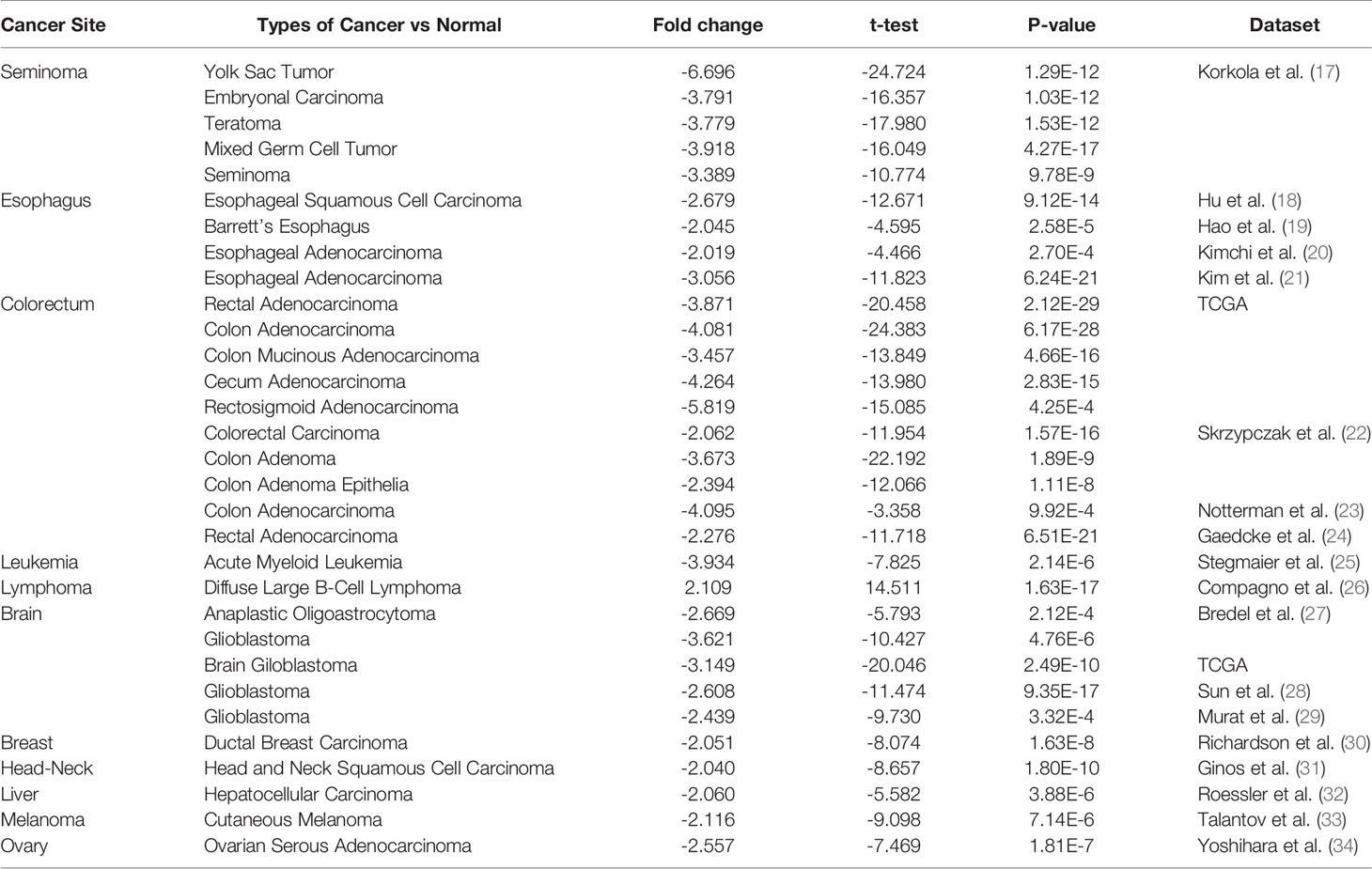
Table 1 Datasets of PINK1 expression in pan-cancers. (ONCOMINE database).
Moreover, we further used TIMER to evaluate the RNA sequencing data of PINK1 in TCGA. Figure 1B shows the details of PINK1 expression across cancers. PINK1 expression was downregulated in many cancer types, including BLCA (bladder urothelial carcinoma), BRCA (breast invasive carcinoma), CHOL (cholangiocarcinoma), COAD (colon adenocarcinoma), ESCA (esophageal carcinoma), HNSC (head and neck cancer), KICH (kidney chromophobe), KIRC (kidney renal clear cell carcinoma), KIRP (kidney renal papillary cell carcinoma), LIHC (liver hepatocellular carcinoma), LUAD (lung adenocarcinoma), LUSC (lung squamous cell carcinoma), PRAD (prostate adenocarcinoma), READ (rectum adenocarcinoma), STAD (stomach adenocarcinoma), and UCEC (uterine corpus endometrial carcinoma). Furthermore, we assessed the PINK1 protein expression level in the human protein atlas datasets (Supplementary Figure 1), most cancer tissues showed moderate granular cytoplasmic positively. Collectively, PINK1 acts as a tumor suppressor in most cancer types.
Prognostic Value of PINK1 Across Cancers
Next, we analyzed the prognostic value of PINK1 expression across cancers in PrognoScan, Kaplan-Meier Plotter, and GEPIA. First, we investigated the prognostic value of PINK1 expression in different cancer types in PrognoScan, and the detailed results are shown in Supplementary Figure 2. We discovered that there was a significant prognostic value of PINK1 expression in seven cancer types: colorectal, ovarian, blood, brain, breast, lung, and soft tissue cancers (Figure 2). The results showed that PINK1 played a protective role in five cancer types, including blood cancer (Figure 2A, OS: Cox P = 0.025802), brain cancer (Figure 2B, OS: Cox P = 0.000357), breast cancer (Figures 2C, D, RFS: Cox P = 0.022997; DMFS: Cox P = 0.002365), lung cancer (Figure 2F, OS: Cox P = 0.004743) and soft tissue cancer (Figure 2I, DRFS: Cox P = 0.037673). However, PINK1 played a detrimental role in colorectal cancer (Figure 2E, DFS: Cox P = 0.001134). Meanwhile, the role of PINK1 in ovarian cancer was controversial (Figures 2G, H, OS of DUKE-OC: Cox P = 0.000154; OS of GSE8841: Cox P = 0.011487).
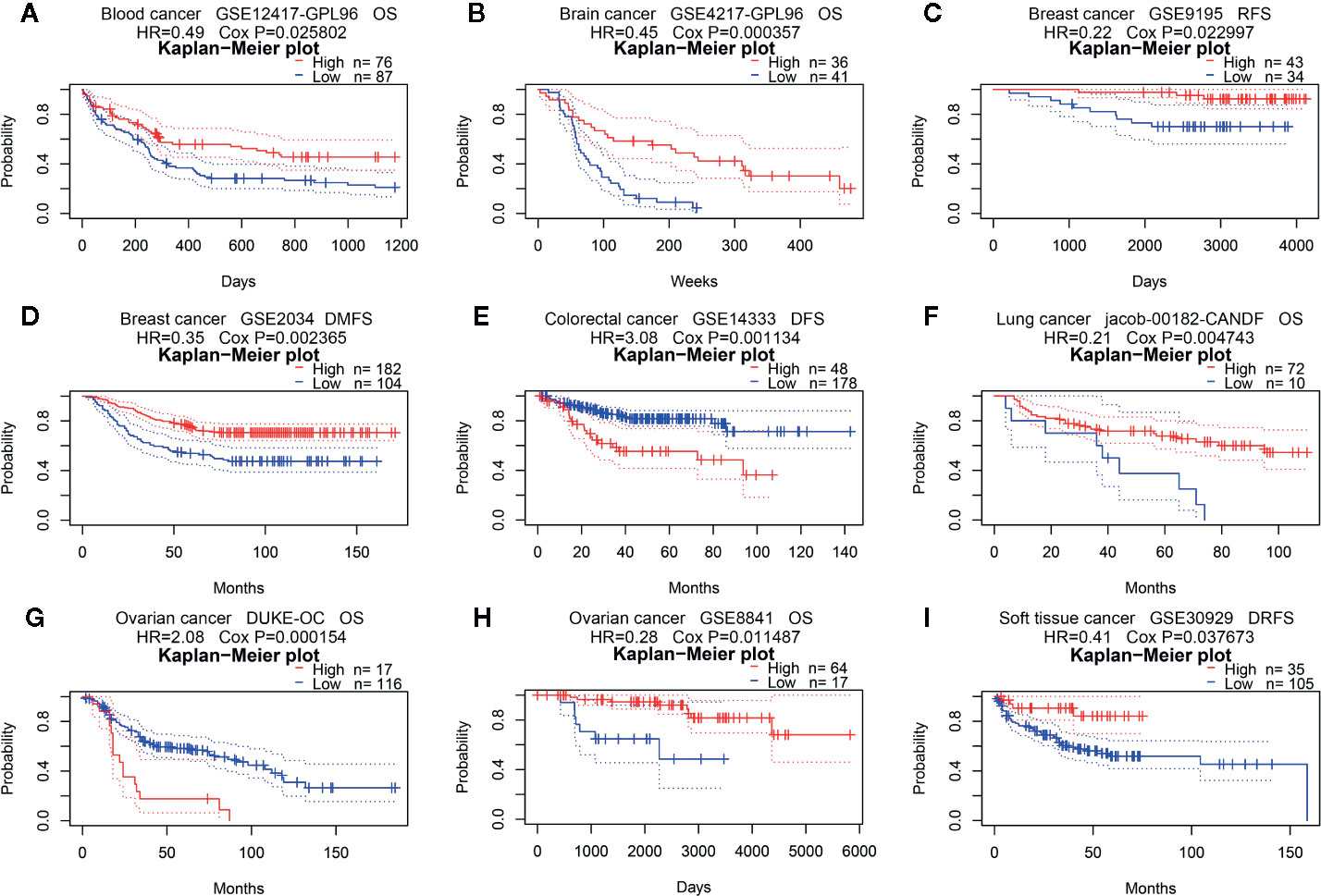
Figure 2 Survival analyses of PINK1 expression in pan-cancers (based on PrognoScan). (A) OS (n = 163) in blood cancer cohort GSE12417-GPL96. (B) OS (n = 77) in brain cancer cohort GSE4217-GPL96. (C) RFS (n = 77) in breast cancer cohort GSE9195. (D) DMFS (n = 286) in breast cancer cohort GSE2034. (E) DFS (n = 226) in colorectal cancer cohort GSE14333. (F) OS (n = 82) in lung cancer cohort jacob-00182-CANDF. (G) OS (n = 133) in ovarian cancer cohort DUKE-OC. (H) OS (n = 81) in ovarian cancer cohort GSE8841. (I) DRFS (n = 140) in soft tissue cancer cohort GSE30929. OS, overall survival; DMFS, distant metastasis free survival; RFS, relapse free survival; DFS, disease free survival; DRFS, distant recurrence free survival.
Second, we used Kaplan-Meier Plotter to further analyze the prognostic value of PINK1 in different types of cancers (Supplementary Table 1). In contrast to PrognoScan, whose data mostly come from the GEO (Gene Expression Omnibus) database, Kaplan-Meier Plotter utilizes Affymetrix microarray data from TCGA. Notably, PINK1 expression was significantly correlated with nine cancer types (Figure 3A). Notably, we newly identified PINK1 as a protective prognostic factor in ESCA (esophageal adenocarcinoma) (OS: log rank P = 0.039) (Figure 3B). This discovery might challenge a previous report stating that high expression of PINK1 is a poor prognostic factor for patients with esophageal squamous cell carcinoma treated with neoadjuvant chemotherapy (35). For HNSC (head and neck squamous cell carcinoma), PINK1 was identified as a protective prognostic factor (OS: log rank P = 0.0072) (Figure 3C). In addition, PINK1 was also found to have a protective effect on OS and RFS in LIHC (liver hepatocellular carcinoma) (OS: log-rank P = 8.7e-05; RFS: log-rank P = 0.0039) (Figures 3D, E). Interestingly, we identified PINK1 as a good prognostic factor in PDAC (pancreatic ductal adenocarcinoma) (OS: log rank P = 0.03) (Figure 3F). This finding may further verify the previous result that PINK1 and PARK2 suppress pancreatic tumorigenesis (7). For THCA (thyroid carcinoma), PINK1 significantly affected overall survival but not relapse-free survival (OS: log-rank P = 0.041) (Figure 3G). Furthermore, PINK1 was newly identified as a detrimental prognostic factor for both OS and RFS in UCEC (uterine corpus endometrial carcinoma) (OS: log rank P = 4.7e-05; RFS: log rank P = 0.046) (Figures 3H, I). In addition, for kidney cancer, PINK1 had a detrimental effect on RFS in only KIRP (kidney renal papillary cell carcinoma) (RFS: log rank P = 0.03) (Figure 3J). For both OV (ovarian cancer) and TGCT (testicular germ cell tumor), PINK1 significantly influenced RFS but not OS (RFS: log-rank P = 0.022; RFS: log-rank P = 0.011) (Figures 3K, L).
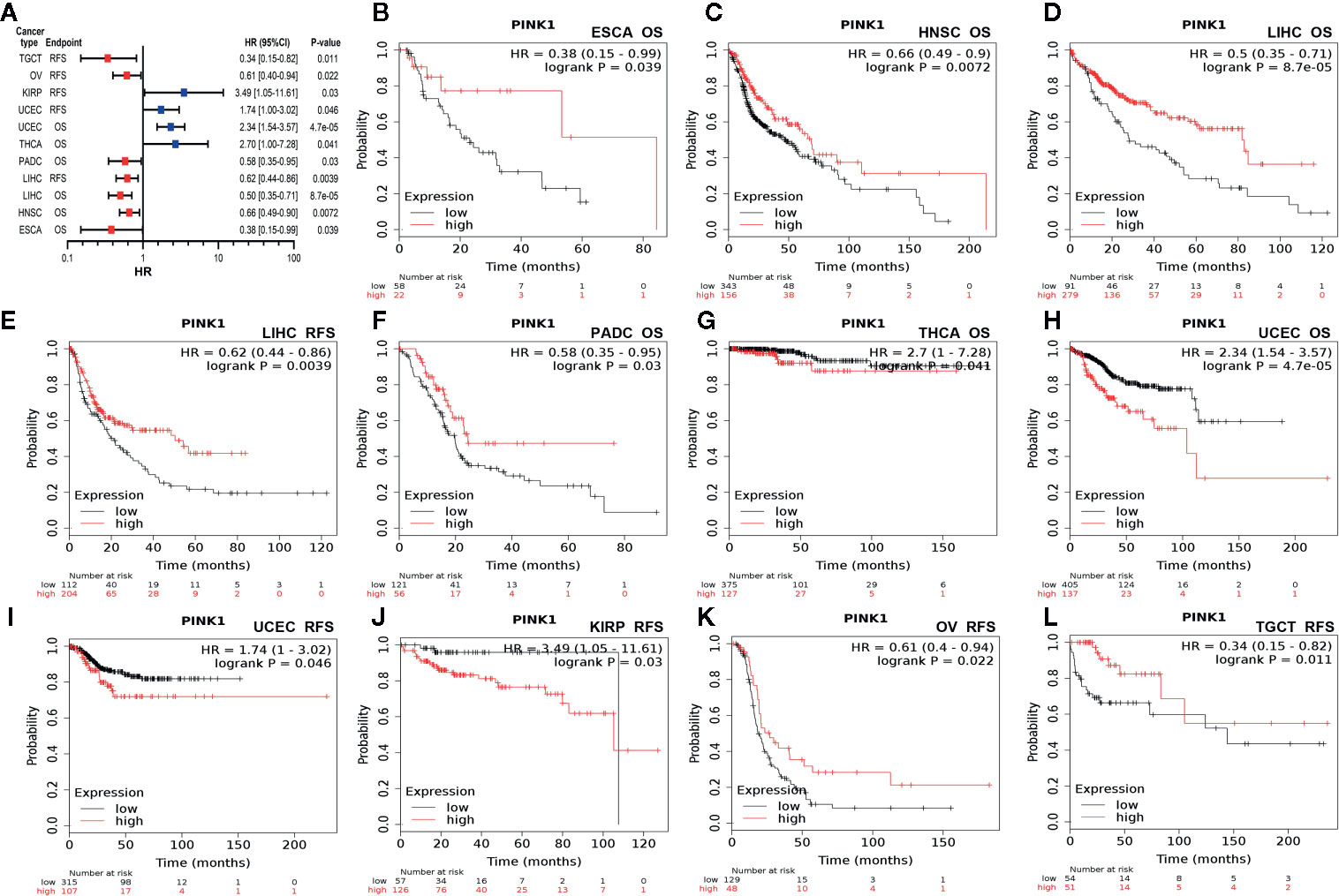
Figure 3 Survival analyses of PINK1 expression in different types of cancer in Kaplan-Meier plotter. (A) Prognostic HR of PIN1K expression in every type pf cancer. (B) OS of ESCA (esophageal adenocarcinoma). (C) OS of HNSC (head-neck squamous cell carcinoma). (D) OS of LIHC (liver hepatocellular carcinoma). (E) RFS of LIHC (liver hepatocellular carcinoma). (F) OS of PADC (pancreatic ductal adenocarcinoma). (G) OS of THCA (thyroid carcinoma). (H) OS of UCEC (uterine corpus endometrial carcinoma). (I) RFS of UCEC (uterine corpus endometrial carcinoma). (J) RFS of KIRP (kidney renal papillary cell carcinoma). (K) RFS of OV (ovarian cancer). (L) RFS of TGCT (testicular germ cell tumor). HR, hazard ratio; CI, confidence interval; OS, overall survival; RFS, relapse free survival.
Third, in addition to our analyses of the microarray data of PINK1 in PrognoScan and Kaplan-Meier Plotter, we used the online tool GEPIA to analyze RNA sequencing data from TCGA. We analyzed the role of PINK1 in 33 kinds of cancer (Supplementary Table 2). Interestingly, PINK1 had a significant overall effect on cancers (OS: total number = 9,500, HR = 0.69, log-rank P = 0; DFS: total number = 9,500, HR = 0.82, log-rank P = 1.4e-07). Compared with low expression of PINK1, high expression of PINK1 was positively correlated with better OS in ACC (adrenocortical carcinoma) and MESO (mesothelioma), DFS in DLBC (lymphoid neoplasm diffuse large B-cell lymphoma), and both OS and DFS in KIRC (kidney renal clear cell carcinoma) and KIRP (kidney renal papillary cell carcinoma). Meanwhile, LIHC (liver hepatocellular carcinoma) outcome was also found to have a positive correlation with PINK1 expression (DFS: HR = 0.75, P-value = 0.057). In contrast, a high expression level of PINK1 was negatively correlated with both OS and DFS in HNSC (head and neck squamous cell carcinoma) and LUSC (lung squamous cell carcinoma). In other cancer types, PINK1 expression had no significant effect on prognosis.
Stratified Prognostic Analyses of PINK1 Expression in LIHC and LUSC
To identify the potential mechanisms of PINK1 expression across cancers, we used LIHC as an example. We used Kaplan-Meier Plotter to explore the correlation between PINK1 expression and several clinicopathological characteristics in LIHC. The results showed that PINK1 played a protective role in LIHC patients with the following clinicopathological characteristics: male sex (OS: P = 0.0056; RFS: P = 0.012), Asian race (OS: P = 8.7e-05; RFS: P = 0.0011), alcohol consumption (OS: P = 0.024; RFS: P = 0.00017), no hepatitis virus infection (OS: P = 0.0068; RFS: P = 0.012), pathology stage 3 (OS: P = 0.00062; RFS: P = 0.0018) and AJCC stage 3 (OS: P = 0.00023; RFS: P = 0.003). Furthermore, for RFS, PINK1 expression also had a significantly positive effect on female (RFS: P = 0.022), white (RFS: P = 0.02), hepatitis virus-infected (RFS: P = 0.024), pathology stage 1 (RFS: P = 0.046) and AJCC stage 1 (RFS: P = 0.025) LIHC patients (Figure 4).
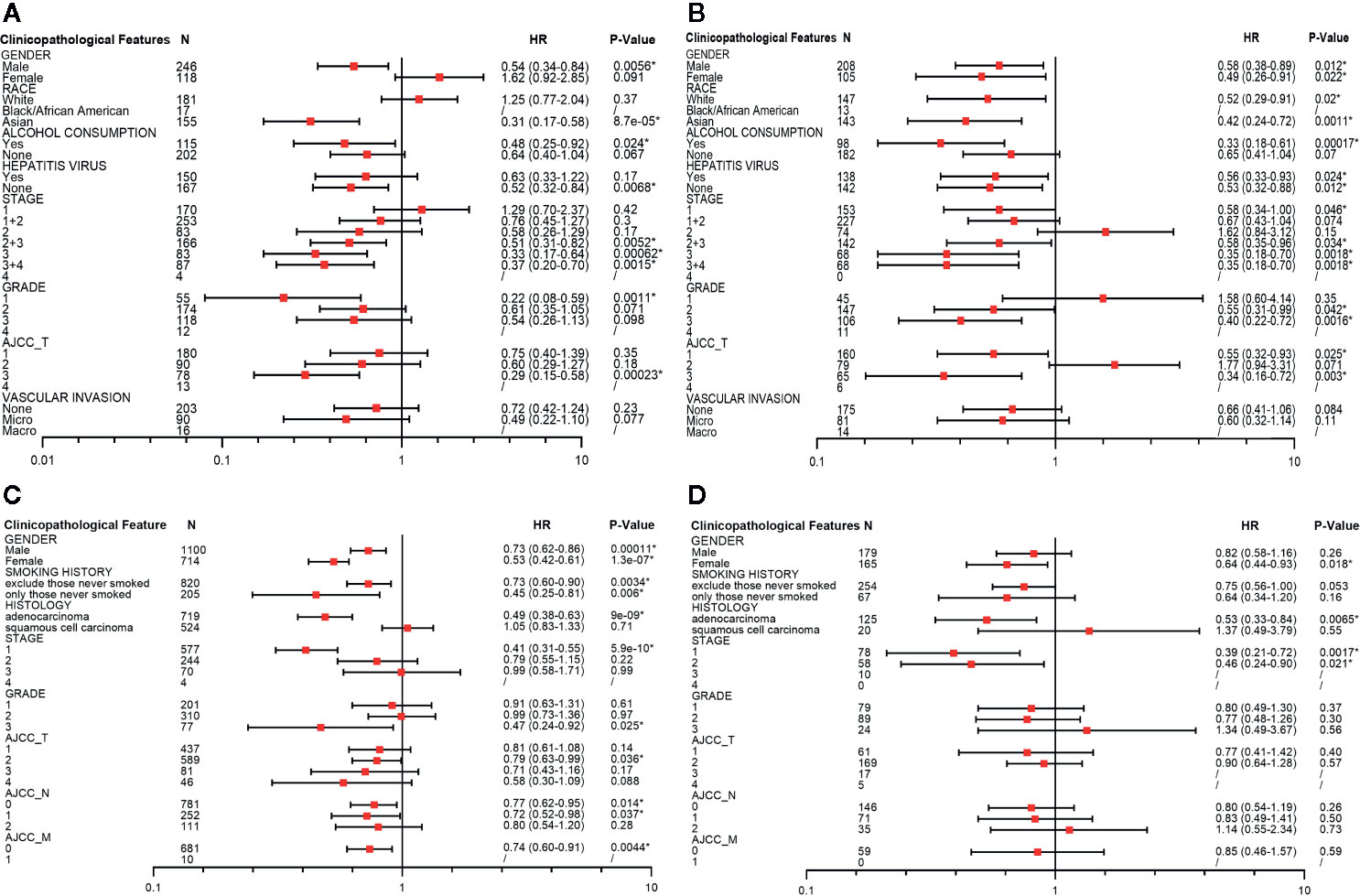
Figure 4 Survival analyses of PINK1 with different clinicopathologic features in liver hepatocellular carcinoma and lung cancer. (A) Correlation of PINK1 mRNA expression with OS in liver hepatocellular carcinoma. (B) Correlation of PINK1 mRNA with RFS in liver hepatocellular carcinoma. (C) Correlation of PINK1 mRNA expression with OS in lung cancer. (D) Correlation of PINK1 mRNA with PPS in lung cancer. *P < 0.05. HR, hazard ratio; OS, overall survival; RFS, relapse free survival; PPS, post progression survival.
We also conducted the detailed evaluation of PINK1 expression and clinicopathological characteristics in lung cancer. Figures 4C, D showed that PINK1 played a protective role in lung cancer patients with the following clinicopathological characteristics: female sex (OS: P = 1.3e-07; PPS: P = 0.018), adenocarcinoma (OS: P = 9e-09; PPS: P = 0.0065) and pathology stage 1(OS: P = 5.9e-10; PPS: P = 0.0017). In addition, for OS, PINK1 expression also had a significantly positive effect on male (OS: P = 0.00011), excluded those never smoked and only those never smoked of smoking history (OS: P = 0.0034; OS: P = 0.006), grade 3 (OS: P = 0.025), AJCC T stage 2 (OS: P = 0.036), AJCC N stage 0 and 1 (OS: P = 0.014; P = 0.037) and AJCC M stage 0 (OS: P = 0.0044). The detailed prognostic values of different clinicopathologic characteristics in LIHC and LUSC are listed in Figure 4.
Conflicting Correlations Between PINK1 Expression and Immune Infiltration in LIHC and LUSC
Determining the interactions between the host immune system and tumor is essential to discover prognostic biomarkers, as well as for reducing drug resistance and developing new cancer therapies (16). It is well known that immune infiltration in the tumor microenvironment can affect patient survival. We also verified that PINK1 might play a prognostic role across cancers. Furthermore, we analyzed the correlation between PINK1 expression and immune infiltration. We used the online TIMER database to explore the correlation between PINK1 expression and the infiltration levels of six immune cell types across cancers. The results showed that PINK1 was significantly correlated with tumor purity in 18 out of 39 cancer types. Meanwhile, the correlations between PINK1 expression and the infiltration levels of B cells, CD8+ T cells, CD4+ T cells, macrophages, neutrophils and dendritic cells were significant in 13, 15, 20, 23, 15, and 21 cancer types, respectively (Supplementary Figure 3).
The GEPIA and TIMER databases are founded on TCGA data. Therefore, based on the findings from GEPIA, LIHC, and LUSC were selected to analyze PINK1 expression and immune infiltration. LIHC represents a cancer in which patients had good survival with high PINK1 expression, while LUSC represents a cancer in which patients had poor survival with high PINK1 expression. Figure 5 shows that PINK1 had significant correlations with tumor purity in both LIHC and LUSC (LIHC: R = -0.109, P = 4.21e-02; LUSC: R = -0.224, P = 7.24e-07). For LIHC, PINK1 expression was significantly negatively correlated with the infiltration levels of four kinds of immune cells: B cells (R = -0.209, P = 9.24e-05), CD8+ T cells (R = -0.11, P = 4.17e-02), macrophages (R = -0.106, P = 4.97e-02) and dendritic cells (R = -0.114, P = 3.60e-02) (Figure 5A). In contrast, the PINK1 expression level was significantly positively correlated with the infiltration level of CD8+ T cells (R = 0.1, P = 2.85e-02), CD4+ T cells (R = 0.452, P = 2.55e-25), macrophages (R = 0.382, P = 5.50e-18), neutrophils (R = 0.223, P = 8.82e-07), and dendritic cells (R = 0.337, P = 5.15e-14) in LUSC (Figure 5B). Taken together, the results above reflect that PINK1 expression might affect cancer patient survival by influencing immune infiltration in the tumor microenvironment.
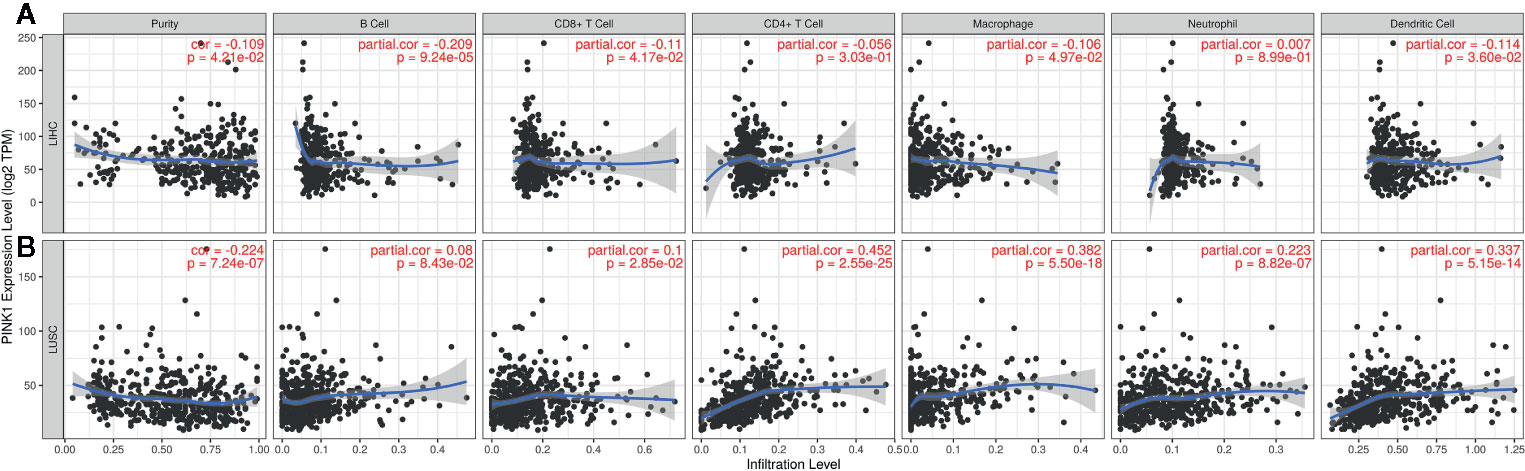
Figure 5 The correlation between PINK1 expression and immune infiltration level in LIHC and LUSC. (A) Correlation of PINK1 expression with tumor purity and infiltrating levels of B cell, CD8+ T cell, CD4+ T cell, macrophage, neutrophil and dendritic cell in LIHC. (B) Correlation of PINK1 expression with tumor purity and infiltrating levels of B cell, CD8+ T cell, CD4+ T cell, macrophage, neutrophil, and dendritic cell in LUSC. P < 0.05 is considered as statistically significant. LIHC, liver hepatocellular carcinoma; LUSC, lung squamous cell carcinoma.
Correlation Between PINK1 Expression and Representative Immune Markers in LIHC and LUSC
The above analyses revealed that PINK1 expression might play an important role in immune infiltration in the tumor microenvironment. We then performed a deeper investigation of the relationship between PINK1 expression and representative immune markers of several immune cells. The immune cells included B cells, CD8+ T cells, follicular helper T cells, T helper 1 cells, T helper 2 cells, T helper 9 cells, T helper 17 cells, T helper 22 cells, regulatory T cells, exhausted T cells, macrophages, M1 macrophages, M2 macrophages, tumor-associated macrophages, monocytes, natural killer cells, neutrophils, and dendritic cells. We selected LIHC and LUSC to explore the relationships in TIMER. Adjustment for tumor purity was performed for accuracy of correlation. PINK1 expression was significantly correlated with 34 of the 45 cell types in LIHC and with 31 of the 45 cell types in LUSC (Table 2). Table 2 shows that the correlation of PINK1 expression with these immune cell types was different in LIHC and LUSC. In LIHC, PINK1 expression was significantly correlated with B cell- and CD8+ T cell-specific markers, while LUSC showed no correlations of PINK1 with these cells. In addition, PINK1 expression was more strongly correlated with tumor-associated macrophage markers, such as HLA-G, CD80, and CD86, in LUSC than in LIHC. These analyses further confirmed that PINK1 expression plays an important role in immune infiltration in cancers such as LIHC and LUSC. These results might contribute to the understanding of the survival differences seen in cancer patients.
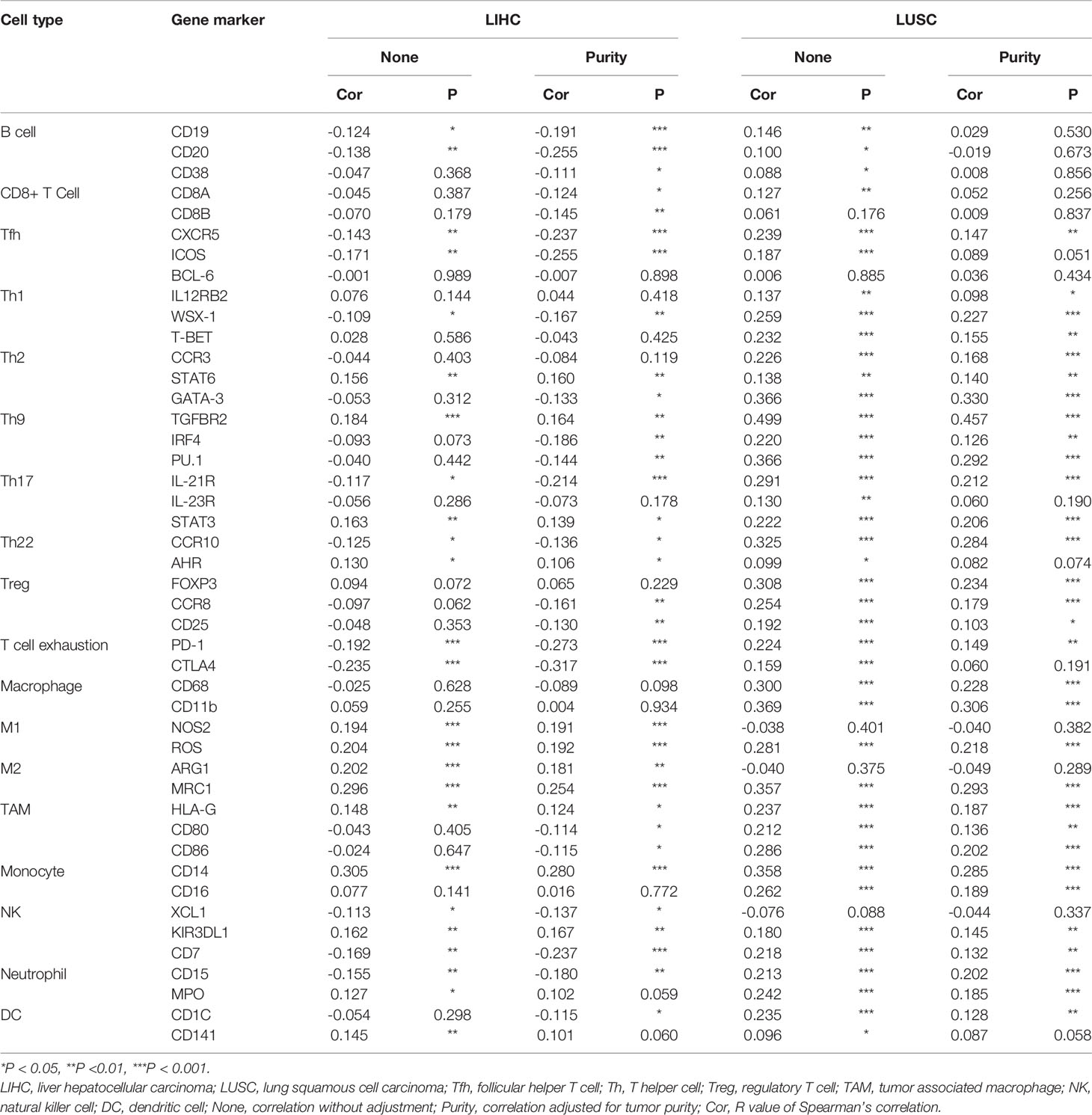
Table 2 The correlation between PINK1 expression and immune cells related gene markers. (TIMER database).
Association Between PINK1 Copy Number Variations and Immune Infiltrates Across Cancers
Supplementary Figure 4 shows that the PINK1 gene is frequently altered in across cancers. The association between PINK1 copy number variations and immune infiltrates in different kinds of cancer was investigated. Figure 6 shows nine of the most significant relationships between the changes in PINK1 copy number variations and six types of immune infiltrates in all cancers. In particular, deletion of PINK1 was associated with substantially lower levels of six immune infiltrates in breast invasive carcinoma, colon adenocarcinoma, head and neck cancer, lower grade glioma, lung adenocarcinoma, lung squamous cell carcinoma, sarcoma, skin cutaneous melanoma, and stomach adenocarcinoma. However, deletion of PINK1 was associated with substantially higher CD4+ T cell, neutrophil and dendritic cell counts in breast invasive carcinoma. These findings might suggest the potential mechanism by which PINK1 alterations predict the response to immune therapy.
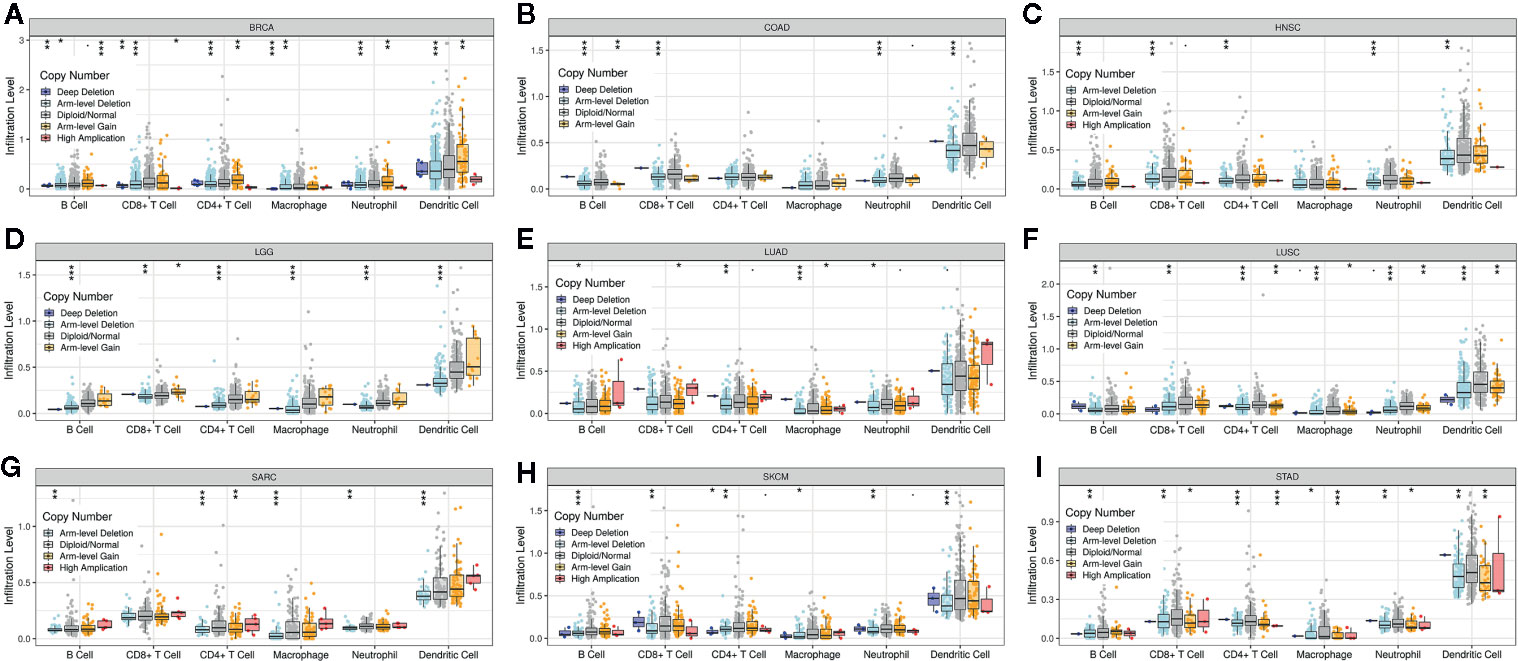
Figure 6 The association between PINK1 copy number variations and immune infiltrates in (A) BRCA (breast invasive carcinoma), (B) COAD (colon adenocarcinoma), (C) HNSC (head and neck cancer), (D) LGG (lower grade glioma), (E) LUAD (lung adenocarcinoma), (F) LUSC (lung squamous cell carcinoma), (G) SARC (sarcoma), (H) SKCM (skin cutaneous melanoma), (I) STAD (stomach adenocarcinoma). *P < 0.05, **P < 0.01, ***P < 0.001.
Discussion
As an evolutionarily conserved cellular process, mitophagy selectively eliminates dysfunctional mitochondria by directing them to the autophagosome and degrading them. However, when mitophagy is dysregulated, mitochondria are damaged and accumulate, causing carcinogenesis and cancer progression (36). PINK1 does not affect the mitophagy process directly (37). Mitochondrial depolarization recedes PINK1 cleavage and causes PINK1 accumulation at the outer membrane of mitochondria by the TOM/TIM complex. The ubiquitin-tagged outer mitochondrial membrane proteins bind to diverse LC3 binding regions that involve different kinds of autophagy adaptors, such as OPTN, NBR1, and TAX1BP1 (38, 39). In addition to the role of ubiquitin-dependent receptors in mitophagy, PINK1 also plays an extensive role in cancer. Research has shown that PINK1 can work as a regulator of the Warburg effect and a negative regulator of glioblastoma growth, and the loss of PINK1 contributes to the Warburg effect through ROS-dependent stabilization of HIF1A (40). Silencing of PINK1 inhibited the proliferation of lung cancer cells and blocked the cell cycle (41). However, the role of PINK1 in metastasis in different cancers has not yet been widely studied. Whether PINK1 expression influences cancer patient survival remains unknown. Our analyses were all based on human datasets. According to our study, we found that PINK1 expression was lower in several cancers, including brain, breast, colorectal, esophageal, head and neck, liver, and ovarian cancers as well as leukemia and melanoma, at the mRNA level. Both the Oncomine and TIMER databases showed low expression levels of PINK1 in most cancer types. This might indicate that PINK1 plays a protective role across cancers. However, this was a preliminary assumption. To confirm the role of PINK1 in cancers, we analyzed the prognosis of patients according to PINK1 expression in different kinds of cancer with data from various databases.
Our analyses showed that PINK1 played a protective role in five cancer types, including blood cancer, brain cancer, breast cancer, lung cancer, and soft tissue cancer. However, PINK1 played a detrimental role in colorectal cancer, and the role of PINK1 in ovarian cancer was controversial. Unlike PrognoScan, which uses data mostly from GEO, the Kaplan-Meier Plotter utilizes Affymetrix microarray data from TCGA. PINK1 expression was significantly correlated with nine cancer types and played a protective role in ESCA, HNSC, LIHC, PADC, OV, and TGCT. The results were not completely consistent. These discrepancies in PINK1 expression in different kinds of cancer might be due to the heterogeneity of data collection as well as a result of the presence of specific biological properties in each cancer type. However, the PINK1 expression in LIHC and LUSC showed consistency in the three databases. We further explored the correlation between PINK1 expression and several clinicopathological characteristics in LIHC and lung cancer. Interestingly, the results showed that PINK1 always played a protective role in LIHC patients with the following clinicopathological characteristics: male sex, Asian race, alcohol consumption, no hepatitis virus infection, pathology stage 3, and AJCC stage 3. The similar results could also be got from lung cancer. Taken together, our results revealed that the role of PINK1 might be context dependent and could vary among different cancers. PINK1 played a protective role in most cancers. However, the discrepant PINK1 expression in different database might reflect the different underlying mechanisms which need to be further verified by experiments. We all know that cancer is a highly heterogeneous disease, and the crosstalk within the tumor is also very diverse. For example, a drug that is effective for a certain gene in a certain cancer is also effective for other cancers that express that gene. In response to this situation, researchers believe that the molecular signals that stem from the same genes in different cancer cells are different, leading to different therapeutic effects. Generally, the gene changes in the same cancer may be different. Similarly, the expression and function of the same gene in different cancers are also different (42, 43). Above all, these findings could provide a view on the utility of the mitophagy-related protein PINK1 as a prognostic factor across cancers.
Scientific observations have exemplified the critical influence of mitochondrial metabolism on malignant transformation and tumor progression (44–46). Defects in immunity also contribute to carcinogenesis, cancer progression, and poor cancer treatment efficacy. Take anti-PD1 immunotherapy as an example, in most advanced cancers, except Hodgkin lymphoma (which has high PD-L1/L2 expression) and melanoma (which has high tumor mutational burden), the objective response rate with anti-PD-1/PD-L1 monotherapy is only ~20%. It suggested that the roles of PD-1/PD-L1 in immune suppression remain to be better defined and important immune regulatory mechanisms within or outside of the PD-1/PD-L1 network need to be discovered (47). This concept prompted us to elucidate the immunological role of the mitophagy-related protein PINK1 across cancers to determine whether the immune system plays any role in the prognostic value of PINK1.
There have been several studies about the role of PINK1 expression in tumor immune function. Sun et al. discovered that a lack of PINK1 alters glial innate immune responses and enhances nitric oxide-mediated neuron death (48). Basit F verified that dendritic cells required PINK1-mediated phosphorylation of BCKDE1α to promote fatty acid oxidation for immune function (48). None of the studies focused on the relationship between PINK1 expression and immune infiltration in cancer patients. We used the online TIMER database to explore the correlation between PINK1 expression and the levels of six kinds of immune infiltrates across cancers. In our study, results showed that PINK1 had significant correlations with tumor purity in both LIHC and LUSC (LIHC: P = 4.21e-02; LUSC: P = 7.24e-07), which indicating their comparative enrichment in tumor cells and may be attributable to the enrichment patterns of PINK1 in the tumor microenvironment. The results showed that PINK1 was significantly correlated with six different immune infiltrates, B cells, CD8+ T cells, CD4+ T cells, macrophages, neutrophils and dendritic cells, across cancers. In addition to the contradictory results of the prognostic value of PINK1 expression in LIHC and LUSC, there were also conflicting results in the correlations between PINK1 expression and immune infiltrates in LIHC and LUSC. For LIHC, PINK1 expression was significantly negatively correlated with immune infiltration levels in four kinds of immune cells: B cells, CD8+ T cells, macrophages and dendritic cells, while PINK1 was positively correlated with these cell types in LUSC. We further explored the correlation between PINK1 expression and representative immune markers in LIHC and LUSC, and PINK1 expression was more strongly correlated with tumor-associated macrophage markers such as HLA-G, CD80, and CD86 in LUSC than in LIHC. Moreover, PINK1 expression was significantly correlated with B cell- and CD8+ T cell-specific markers in LIHC, while no correlation of PINK1 expression with these markers was shown in LUSC. Meanwhile, the increase of CD4+ T cells and neutrophils would result in the poorer prognosis of LUSC than LIHC. Several studies, though not comprehensively elucidated, showed consistency with our analysis. Various CD4+ T cells, including regulatory T (Treg) and T helper 17 (Th17) CD4+ T cell have been observed to mechanistically promote tumorigenesis, cancer progression and metastasis through immunosuppressive and pro-inflammatory functions. Specifically, Tregs contributes to poorer prognosis of LUSC by inducing immunosuppression through contact-dependent mechanisms in LUSC such as the expression of cytotoxic T-lymphocyte-associated protein 4 (CTLA-4), programmed cell death 1 (PD-1), programmed death-ligand 1 (PD-L1) et al. Tregs also influences the tumor microenvironment during the progression of LUSC by inhibiting CD8+ T cell-mediated anti-tumor immunity and resulting in tumor cell death. Th17 contributes to poorer prognosis of LUSC by expressing the transcription factors RORγt/RORC2 (mouse/human) and RORα, which drive Th17 differentiation and produce pro-inflammatory cytokines IL-17, which has been shown to promote tumor growth by increasing angiogenesis, metastasis and macrophage infiltration into tumors. Th17 cells also produces other cytokines in addition to IL-17, including IL-22, which contributes to angiogenesis and metastasis (49). For neutrophils, Marta et al. had reported that in LUSC, anti-angiogenic therapy increased the expression of stem cell maker CD15 which also worked as a neutrophil related gene, which would cause disease progression and also induce tumor proliferation (50). Another neutrophil related gene MPO have been found to be released from neutrophils in lung tissue in response to exposure to various pulmonary insults. MPO had been also shown to activate an intermediate metabolite of B(a)P to the highly reactive and carcinogenic B(a)P diol epoxide and to enhance the binding of B(a)P diol to lung DNA in vitro and finally lead to disease progression (51). These results confirmed our hypothesis that PINK1 expression in LIHC and LUSC correlates with immune cell in different manners, which might help explain the differences in patient survival. With the development of scientific research, the tumor environment including immune cell infiltration could help us elucidate the mechanisms behind tumor development. However, we could only figure out the significant correlations between immune cell infiltration and PINK1 expression in tumors, the cause-effect relationship was hard to established. Recent studies have already showed some possible mechanisms deciphering why PINK1 expression correlated with different prognoses and immune infiltration in pan-cancer. It was acknowledged that the immune system prevents cancer from developing by activating T cells and macrophages to attack tumor cells. However, once the tumor progresses beyond this early stage, the immune TME would transform to support cancer cells, promote tumor progression and suppress immune cell mediated cytotoxicity (52, 53). Our results also find that the immune infiltration levels of antigen presenting cells such as macrophages and B cells are significantly correlated with the PINK1 expression level in cancers. And there were also researchers reported the mitophagy could reduce CD8+ T cell activation. Besides, the mitophagy was also involved in the development and differentiation of immune cells such like NK cells, macrophages and T cells (52). In addition, we also find out that there was a strong association between PINK1 copy number variation and immune infiltrates across cancers. All of the above findings revealed that the correlation between PINK1 expression and immune infiltration exists but various in different types of cancer, it might contribute to patient survival by influencing immune infiltration in the tumor microenvironment.
Although we performed a substantial exploration of the role of PINK1 expression in both prognostic and immunological aspects, there were some limitations of our analysis. First, multiple analyses based on diverse databases could provide a large amount of data as well as data heterogeneity. Therefore, some of the results were conflicting. Second, our analyses were based on pancancer data in patients, and it might be difficult to perform experiments to prove all the ideas at the same time. Further in vivo/in vitro experiments and even clinical trials are needed to verify the highlights from our bioinformatics analysis. Third, although we found that PINK1 expression correlated well with immune infiltration and patient survival, we did not find that PINK1 affects cancer patient prognosis via immune infiltration. This hypothesis remains to be verified.
Conclusion
In conclusion, the mitophagy-related protein PINK1 is significantly correlated with prognosis and immune infiltration across cancers, especially in LIHC and LUSC. PINK1 might work as a new biomarker for prognostic prediction and immune cell infiltration across cancers in the future.
Data Availability Statement
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found in the article/Supplementary Material.
Author Contributions
LZ: Collection and/or assembly of data, data analysis and interpretation, manuscript writing, methodology, and software. WW: Collection and/or assembly of data, data analysis and interpretation, manuscript writing and editing. SJ: Data analysis, manuscript writing and interpretation. SY: Manuscript writing and project administration. YY: Manuscript writing and project administration. KW: Manuscript writing. JH: Conception/design. YR: Conception/design, supervision and editing. BW: Conception/design, supervision, and editing. All authors contributed to the article and approved the submitted version.
Funding
This study was supported by grants from National Natural Science Fund of China (number 81702633).
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We showed our great thanks to Dr. Taiwen Li for his technical support of TIMER. We also showed our special thanks to Oncomine, PrognoScan, Kaplan-Meier Plotter, GEPIA, TIMER, cBioportal for their open access databases.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fonc.2020.569887/full#supplementary-material
References
1. Pickles S, Vigié P, Youle RJ. Mitophagy and Quality Control Mechanisms in Mitochondrial Maintenance. Curr Biol (2018) 28(4):R170–85. doi: 10.1016/j.cub.2018.01.004
2. Wong YC, Kim S, Peng W, Krainc D. Regulation and Function of Mitochondria-Lysosome Membrane Contact Sites in Cellular Homeostasis. Trends Cell Biol (2019) 29(6):500–13. doi: 10.1016/j.tcb.2019.02.004
3. Wai T, Langer T. Mitochondrial Dynamics and Metabolic Regulation. Trends Endocrinol Metab (2016) 27(2):105–17. doi: 10.1016/j.tem.2015.12.001
4. Senft D, Ronai ZA. Regulators of mitochondrial dynamics in cancer. Curr Opin Cell Biol (2016) 39:43–52. doi: 10.1016/j.ceb.2016.02.001
5. Mizushima N, Levine B. Autophagy in mammalian development and differentiation. Nat Cell Biol (2010) 12(9):823–30. doi: 10.1038/ncb0910-823
6. Drake LE, Springer MZ, Poole LP, Kim CJ, Macleod KF. Expanding perspectives on the significance of mitophagy in cancer. Semin Cancer Biol (2017) 47:110–24. doi: 10.1016/j.semcancer.2017.04.008
7. Li C, Zhang Y, Cheng X, Yuan H, Zhu S, Liu J, et al. PINK1 and PARK2 Suppress Pancreatic Tumorigenesis through Control of Mitochondrial Iron-Mediated Immunometabolism. Dev Cell (2018) 46(4):441–55.e8. doi: 10.1016/j.devcel.2018.07.012
8. Zitvogel L, Galluzzi L, Smyth MJ, Kroemer G. Mechanism of action of conventional and targeted anticancer therapies: reinstating immunosurveillance. Immunity (2013) 39(1):74–88. doi: 10.1016/j.immuni.2013.06.014
9. Bocanegra A, Fernandez-Hinojal G, Zuazo-Ibarra M, Arasanz H, Garcia-Granda MJ, Hernandez C, et al. PD-L1 Expression in Systemic Immune Cell Populations as a Potential Predictive Biomarker of Responses to PD-L1/PD-1 Blockade Therapy in Lung Cancer. Int J Mol Sci (2019) 20(7):1631. doi: 10.3390/ijms20071631
10. Wu X, Giobbie-Hurder A, Liao X, Connelly C, Connelly EM, Li J, et al. Angiopoietin-2 as a Biomarker and Target for Immune Checkpoint Therapy. Cancer Immunol Res (2017) 5(1):17–28. doi: 10.1158/2326-6066.CIR-16-0206
11. Rhodes DR, Yu J, Shanker K, Deshpande N, Varambally R, Ghosh D, et al. ONCOMINE: a cancer microarray database and integrated data-mining platform. Neoplasia (2004) 6(1):1–6. doi: 10.1016/s1476-5586(04)80047-2
12. Mizuno H, Kitada K, Nakai K, Sarai A. PrognoScan: a new database for meta-analysis of the prognostic value of genes. BMC Med Genomics (2009) 2:18. doi: 10.1186/1755-8794-2-18
13. Nagy Á, Lánczky A, Menyhárt O, Győrffy B. Validation of miRNA prognostic power in hepatocellular carcinoma using expression data of independent datasets. Sci Rep (2018) 8(1):9227. doi: 10.1038/s41598-018-27521-y
14. Tang Z, Li C, Kang B, Gao G, Li C, Zhang Z. GEPIA: a web server for cancer and normal gene expression profiling and interactive analyses. Nucleic Acids Res (2017) 45(W1):W98–W102. doi: 10.1093/nar/gkx247
15. Li T, Fan J, Wang B, Traugh N, Chen Q, Liu JS, et al. TIMER: A Web Server for Comprehensive Analysis of Tumor-Infiltrating Immune Cells. Cancer Res (2017) 77(21):e108–10. doi: 10.1158/0008-5472.CAN-17-0307
16. Li B, Severson E, Pignon JC, Zhao H, Li T, Novak J, et al. Comprehensive analyses of tumor immunity: implications for cancer immunotherapy. Genome Biol (2016) 17(1):174. doi: 10.1186/s13059-016-1028-7
17. Korkola JE, Houldsworth J, Chadalavada RS, Olshen AB, Dobrzynski D, Reuter VE, et al. Down-regulation of stem cell genes, including those in a 200-kb gene cluster at 12p13.31, is associated with in vivo differentiation of human male germ cell tumors. Cancer Res (2006) 66(2):820–7. doi: 10.1158/0008-5472.CAN-05-2445
18. Hu N, Clifford RJ, Yang HH, Wang C, Goldstein AM, Ding T, et al. Genome wide analysis of DNA copy number neutral loss of heterozygosity (CNNLOH) and its relation to gene expression in esophageal squamous cell carcinoma. BMC Genomics (2010) 11:576. doi: 10.1186/1471-2164-11-576
19. Hao Y, Triadafilopoulos G, Sahbaie P, Young HS, Omary MB, Lowe AW. Gene expression profiling reveals stromal genes expressed in common between Barrett’s esophagus and adenocarcinoma. Gastroenterology (2006) 131(3):925–33. doi: 10.1053/j.gastro.2006.04.026
20. Kimchi ET, Posner MC, Park JO, Darga TE, Kocherginsky M, Karrison T, et al. Progression of Barrett’s metaplasia to adenocarcinoma is associated with the suppression of the transcriptional programs of epidermal differentiation. Cancer Res (2005) 65(8):3146–54. doi: 10.1158/0008-5472.CAN-04-2490
21. Kim SM, Park YY, Park ES, Cho JY, Izzo JG, Zhang D, et al. Prognostic biomarkers for esophageal adenocarcinoma identified by analysis of tumor transcriptome. PloS One (2010) 5(11):e15074. doi: 10.1371/journal.pone.0015074
22. Skrzypczak M, Goryca K, Rubel T, Paziewska A, Mikula M, Jarosz D, et al. Modeling oncogenic signaling in colon tumors by multidirectional analyses of microarray data directed for maximization of analytical reliability. PloS One (2010) 5(10):e13091. doi: 10.1371/journal.pone.0013091
23. Notterman DA, Alon U, Sierk AJ, Levine AJ. Transcriptional gene expression profiles of colorectal adenoma, adenocarcinoma, and normal tissue examined by oligonucleotide arrays. Cancer Res (2001) 61(7):3124–30.
24. Gaedcke J, Grade M, Jung K, Camps J, Jo P, Emons G, et al. Mutated KRAS results in overexpression of DUSP4, a MAP-kinase phosphatase, and SMYD3, a histone methyltransferase, in rectal carcinomas. Genes Chromosomes Cancer (2010) 49(11):1024–34. doi: 10.1002/gcc.20811
25. Stegmaier K, Ross KN, Colavito SA, O’Malley S, Stockwell BR, Golub TR. Gene expression-based high-throughput screening(GE-HTS) and application to leukemia differentiation. Nat Genet (2004) 36(3):257–63. doi: 10.1038/ng1305
26. Compagno M, Lim WK, Grunn A, Nandula SV, Brahmachary M, Shen Q, et al. Mutations of multiple genes cause deregulation of NF-kappaB in diffuse large B-cell lymphoma. Nature (2009) 459(7247):717–21. doi: 10.1038/nature07968
27. Bredel M, Bredel C, Juric D, Harsh GR, Vogel H, Recht LD, et al. Functional network analysis reveals extended gliomagenesis pathway maps and three novel MYC-interacting genes in human gliomas. Cancer Res (2005) 65(19):8679–89. doi: 10.1158/0008-5472.CAN-05-1204
28. Sun L, Hui AM, Su Q, Vortmeyer A, Kotliarov Y, Pastorino S, et al. Neuronal and glioma-derived stem cell factor induces angiogenesis within the brain. Cancer Cell (2006) 9(4):287–300. doi: 10.1016/j.ccr.2006.03.003
29. Murat A, Migliavacca E, Gorlia T, Lambiv WL, Shay T, Hamou MF, et al. Stem cell-related “self-renewal” signature and high epidermal growth factor receptor expression associated with resistance to concomitant chemoradiotherapy in glioblastoma. J Clin Oncol (2008) 26(18):3015–24. doi: 10.1200/JCO.2007.15.7164
30. Richardson AL, Wang ZC, De Nicolo A, Lu X, Brown M, Miron A, et al. X chromosomal abnormalities in basal-like human breast cancer. Cancer Cell (2006) 9(2):121–32. doi: 10.1016/j.ccr.2006.01.013
31. Ginos MA, Page GP, Michalowicz BS, Patel KJ, Volker SE, Pambuccian SE, et al. Identification of a gene expression signature associated with recurrent disease in squamous cell carcinoma of the head and neck. Cancer Res (2004) 64(1):55–63. doi: 10.1158/0008-5472.can-03-2144
32. Roessler S, Jia HL, Budhu A, Forgues M, Ye QH, Lee JS, et al. A unique metastasis gene signature enables prediction of tumor relapse in early-stage hepatocellular carcinoma patients. Cancer Res (2010) 70(24):10202–12. doi: 10.1158/0008-5472.CAN-10-2607
33. Talantov D, Mazumder A, Yu JX, Briggs T, Jiang Y, Backus Y, et al. Novel genes associated with malignant melanoma but not benign melanocytic lesions. Clin Cancer Res (2005) 11(20):7234–42. doi: 10.1158/1078-0432.CCR-05-0683
34. Yoshihara K, Tajima A, Komata D, Yamamoto T, Kodama S, Fujiwara H, et al. Gene expression profiling of advanced-stage serous ovarian cancers distinguishes novel subclasses and implicates ZEB2 in tumor progression and prognosis. Cancer Sci (2009) 100(8):1421–8. doi: 10.1111/j.1349-7006.2009.01204.x
35. Yamashita K, Miyata H, Makino T, Masuike Y, Furukawa H, Tanaka K, et al. High Expression of the Mitophagy-Related Protein Pink1 is Associated with a Poor Response to Chemotherapy and a Poor Prognosis for Patients Treated with Neoadjuvant Chemotherapy for Esophageal Squamous Cell Carcinoma. Ann Surg Oncol (2017) 24(13):4025–32. doi: 10.1245/s10434-017-6096-8
36. Panigrahi DP, Praharaj PP, Bhol CS, Mahapatra KK, Patra S, Behera BP, et al. The emerging, multifaceted role of mitophagy in cancer and cancer therapeutics. Semin Cancer Biol (2019) 66:45–58. doi: 10.1016/j.semcancer.2019.07.015
37. Meßling S, Matthias J, Xiong Q, Fischer S, Eichinger L. The two Dictyostelium discoideum autophagy 8 proteins have distinct autophagic functions. Eur J Cell Biol (2017) 96(4):312–24. doi: 10.1016/j.ejcb.2017.03.014
38. Lazarou M, Sliter DA, Kane LA, Sarraf SA, Wang C, Burman JL, et al. The ubiquitin kinase PINK1 recruits autophagy receptors to induce mitophagy. Nature (2015) 524(7565):309–14. doi: 10.1038/nature14893
39. Heo JM, Ordureau A, Swarup S, Paulo JA, Shen K, Sabatini DM, et al. RAB7A phosphorylation by TBK1 promotes mitophagy via the PINK-PARKIN pathway. Sci Adv (2018) 4(11):eaav0443. doi: 10.1126/sciadv.aav0443
40. Agnihotri S, Golbourn B, Huang X, Remke M, Younger S, Cairns RA, et al. PINK1 Is a Negative Regulator of Growth and the Warburg Effect in Glioblastoma. Cancer Res (2016) 76(16):4708–19. doi: 10.1158/0008-5472.CAN-15-3079
41. Liu L, Zuo Z, Lu S, Wang L, Liu A, Liu X. Silencing of PINK1 represses cell growth, migration and induces apoptosis of lung cancer cells. BioMed Pharmacother (2018) 106:333–41. doi: 10.1016/j.biopha.2018.06.128
42. Stewart A, Coker EA, Pölsterl S, Georgiou A, Minchom AR, Carreira S, et al. Differences in Signaling Patterns on PI3K Inhibition Reveal Context Specificity in KRAS-Mutant Cancers. Mol Cancer Ther (2019) 18(8):1396–404. doi: 10.1158/1535-7163.MCT-18-0727
43. Shimomura A, Yamamoto N, Kondo S, Fujiwara Y, Suzuki S, Yanagitani N, et al. First-in-Human Phase I Study of an Oral HSP90 Inhibitor, TAS-116, in Patients with Advanced Solid Tumors. Mol Cancer Ther (2019) 18(3):531–40. doi: 10.1158/1535-7163.MCT-18-0831
44. Zamzami N, Marchetti P, Castedo M, Decaudin D, Macho A, Hirsch T, et al. Sequential reduction of mitochondrial transmembrane potential and generation of reactive oxygen species in early programmed cell death. J Exp Med (1995) 182(2):367–77. doi: 10.1084/jem.182.2.367
45. Porporato PE, Filigheddu N, Pedro JMB, Kroemer G, Galluzzi L. Mitochondrial metabolism and cancer. Cell Res (2018) 28(3):265–80. doi: 10.1038/cr.2017.155
46. Chen DS, Mellman I. Elements of cancer immunity and the cancer-immune set point. Nature (2017) 541(7637):321–30. doi: 10.1038/nature21349
47. Xu-Monette ZY, Zhang M, Li J, Young KH. PD-1/PD-L1 Blockade: Have We Found the Key to Unleash the Antitumor Immune Response? Front Immunol (2017) 8:1597. doi: 10.1038/s41598-017-18786-w
48. Basit F, de Vries IJM. Dendritic Cells Require PINK1-Mediated Phosphorylation of BCKDE1α to Promote Fatty Acid Oxidation for Immune Function. Front Immunol (2019) 10:2386. doi: 10.3389/fimmu.2019.02386
49. Marshall EA, Ng KW, Kung SH, Conway EM, Martinez VD, Halvorsen EC, et al. Emerging roles of T helper 17 and regulatory T cells in lung cancer progression and metastasis. Mol Cancer (2016) 15(1):67. doi: 10.1186/s12943-016-0551-1
50. Larrayoz M, Pio R, Pajares MJ, Zudaire I, Ajona D, Casanovas O, et al. Contrasting responses of non-small cell lung cancer to antiangiogenic therapies depend on histological subtype. EMBO Mol Med (2014) 6(4):539–50. doi: 10.1002/emmm.201303214
51. Feyler A, Voho A, Bouchardy C, Kuokkanen K, Dayer P, Hirvonen A, et al. Point: myeloperoxidase -463G –> a polymorphism and lung cancer risk. Cancer Epidemiol Biomarkers Prev (2002) 11(12):1550–4.
52. Wu G, Ganley IG, Hill JA, Yin XM, Dong Z, Xu Y, et al. Emerging views of mitophagy in immunity and autoimmune diseases. Autophagy (2019) 1–15. doi: 10.1080/15548627.2019.1603547
Keywords: PINK1, pan-cancer, prognostic, immune infiltration, biomarker
Citation: Zhu L, Wu W, Jiang S, Yu S, Yan Y, Wang K, He J, Ren Y and Wang B (2020) Pan-Cancer Analysis of the Mitophagy-Related Protein PINK1 as a Biomarker for the Immunological and Prognostic Role. Front. Oncol. 10:569887. doi: 10.3389/fonc.2020.569887
Received: 05 June 2020; Accepted: 15 October 2020;
Published: 10 November 2020.
Edited by:
Christian Badr, Massachusetts General Hospital and Harvard Medical School, United StatesReviewed by:
Franz Rödel, University Hospital Frankfurt, GermanyVictor C. Kok, Asia University, Taiwan
Copyright © 2020 Zhu, Wu, Jiang, Yu, Yan, Wang, He, Ren and Wang. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Bin Wang, MDAzOTQ1QHhqdHVmaC5lZHUuY24=; Yu Ren, MDAxMDE0QHhqdHVmaC5lZHUuY24=
†These authors have contributed equally to this work