 Georges Chehade1†
Georges Chehade1† Nady El Hajj2
Nady El Hajj2 Mohamed Aittaleb2
Mohamed Aittaleb2 Maisa I. Alkailani2
Maisa I. Alkailani2 Yosra Bejaoui2
Yosra Bejaoui2 Asma Mahdi2
Asma Mahdi2 Arwa A. H. Aldaalis2
Arwa A. H. Aldaalis2 Michael Verbiest3Julie Lelotte4
Michael Verbiest3Julie Lelotte4 Nuria Ruiz-Reig1†Irene Durá1Christian Raftopoulos5Nicolas Tajeddine1
Nuria Ruiz-Reig1†Irene Durá1Christian Raftopoulos5Nicolas Tajeddine1 Fadel Tissir1,2*†
Fadel Tissir1,2*†- 1Université Catholique de Louvain, Institute of Neuroscience, Cellular and Molecular Division, Brussels, Belgium
- 2College of Health and Life Sciences, Hamad Bin Khalifa University, Doha, Qatar
- 3Laboratory of Population Genomics, Department of Internal Medicine, Erasmus University Medical Center, Rotterdam, Netherlands
- 4Department of Neuropathology, Saint-Luc University Hospital, Brussels, Belgium
- 5Department of Neurosurgery, Saint-Luc University Hospital, Brussels, Belgium
Background: Glioblastoma is one of the most aggressive primary brain tumors, with a poor outcome despite multimodal treatment. Methylation of the MGMT promoter, which predicts the response to temozolomide, is a well-established prognostic marker for glioblastoma. However, a difference in survival can still be detected within the MGMT methylated group, with some patients exhibiting a shorter survival than others, emphasizing the need for additional predictive factors.
Methods: We analyzed DIAPH3 expression in glioblastoma samples from the cancer genome atlas (TCGA). We also retrospectively analyzed one hundred seventeen histological glioblastomas from patients operated on at Saint-Luc University Hospital between May 2013 and August 2019. We analyzed the DIAPH3 expression, explored the relationship between mRNA levels and Patient’s survival after the surgical resection. Finally, we assessed the methylation pattern of the DIAPH3 promoter using a targeted deep bisulfite sequencing approach.
Results: We found that 36% and 1% of the TCGA glioblastoma samples exhibit copy number alterations and mutations in DIAPH3, respectively. We scrutinized the expression of DIAPH3 at single cell level and detected an overlap with MKI67 expression in glioblastoma proliferating cells, including neural progenitor-like, oligodendrocyte progenitor-like and astrocyte-like states. We quantitatively analyzed DIAPH3 expression in our cohort and uncovered a positive correlation between DIAPH3 mRNA level and patient’s survival. The effect of DIAPH3 was prominent in MGMT-methylated glioblastoma. Finally, we report that the expression of DIAPH3 is at least partially regulated by the methylation of three CpG sites in the promoter region.
Conclusion: We propose that combining the DIAPH3 expression with MGMT methylation could offer a better prediction of survival and more adapted postsurgical treatment for patients with MGMT-methylated glioblastoma.
Introduction
Diaphanous-related formin (DIAPH) 3 (also known as mDia2) belongs to the formins, a family of dimeric multidomain proteins that are conserved in fungi, plants, and animals. Formins are best known for their cardinal functions in actin nucleation, elongation, and organization (1). However, many studies have shown that some formins can bind to microtubules and regulate their dynamics (2). Hence, formins play important roles in remodeling the cytoskeleton and are therefore key regulators of fundamental cellular processes such as division, adhesion, motility, intracellular trafficking, and polarity. In mammals, this family comprises 15 members grouped into seven subfamilies (3). The diaphanous formin subfamily includes DIAPH1 (4), DIAPH2 (5), and DIAPH3 (6).
DIAPH3 is essential for cell division, and several studies have emphasized its role in cytokinesis (7–9). More recently, DIAPH3 has also been shown to be crucial for karyokinesis, specifically for mitotic spindle organization (10) and activation of the spindle assembly checkpoint (11). Consistent with its important role in mitosis, Diaph3 is exclusively expressed in neural progenitors in the developing mouse brain, and its deficiency causes aberrant cell division, chromosomal instability (CIN), and aneuploidy, resulting in the loss of neural progenitor cells and abnormal cortical histogenesis (10, 11). Errors in mitosis often lead to mitotic catastrophe and subsequent cell death or senescence, impeding the proliferation of aneuploid cells (12). Nevertheless, aneuploidy is a hallmark of highly proliferative cancer cells and is generally associated with poor prognosis, disease progression, metastasis, and therapeutic resistance in malignancies (13). Glioblastoma, the most common and aggressive malignant primary brain tumor in adults, is characterized by a very high degree of CIN and aneuploidy (14). This contributes to the intratumoral heterogeneity and is believed to drive therapeutic resistance. However, the underlying mechanisms remain elusive.
The role of DIAPH3 in tumorigenesis was investigated particularly in cancer cell migration and invasion (15–19). Although some of these studies have linked DIAPH3 deficiency to increased amoeboid cell motility through reduced microtubule stability (15, 16), a large body of evidence supports a key role of DIAPH3 in invasion and metastasis. For instance, in breast cancer, DIAPH3 favors the invasion and expansion of macrometastasis by contributing to the actin filament-based formation of invadopodia (17) and filopodium-like protrusions (18), respectively. In patient-derived glioblastoma stem-like cells, indirect evidence from diaphanous formin modulator studies suggests that DIAPH3 contributes to invasion mechanisms (19). This proinvasive role of DIAPH3 does not exclude its role as a genome safeguard since these mechanisms are implicated at different stages of tumor development.
In this work, we explored the relationship between DIAPH3 levels and survival of glioblastoma patients. We show that DIAPH3 is mostly expressed in proliferating malignant cells. Remarkably, high DIAPH3 expression in resection samples, with comparable proliferation rate, predicts a longer survival of patients, especially in the MGMT-methylated group. We also show that the downregulation of DIAPH3 correlates with the methylation of three cytosine-phosphate-guanine (CpG) sites in the promoter.
Methods
The cancer genome atlas and single-cell data mining
The gene expression, mutations and copy number variations of DIAPH3, as well as the clinical data of glioblastoma patients were obtained from The Cancer Genome Atlas (TCGA) portal (https://www.cancer.gov/ccg/research/genome-sequencing/tcga), accessed on 06 December 2023. The single cell level-expression of DIAPH3 and MKI67 in glioblastoma was extracted from the Broad Institute’s single cell portal (https://singlecell.broadinstitute.org/single_cell/study/SCP393/single-cell-rna-seq-of-adult-and-pediatric-glioblastoma), accessed 06 December 2023 (20).
Patient selection and clinical data collection
We retrospectively identified 117 glioblastomas, as defined by histological criteria, operated on at Saint-Luc University Hospital between May 2013 and August 2019. In accordance with the 2021 World Health Organization classification of tumors of the central nervous system (21), we excluded 24 patients from this study, as shown in the data flow diagram. Moreover, we excluded 11 patients for whom only recurrent tumor samples were available and two patients who died within 30 days after the initial surgery. DIAPH3 expression was analyzed in 73 samples. Clinicopathological characteristics and treatment strategies were collected from institutional medical records, as described previously (22). In brief, age was reported at the time of diagnosis, and Karnofsky performance status (KPS) was evaluated before surgery. Tumor location and laterality were determined on preoperative MRI examination. IDH status was determined by immunohistochemistry using an antibody specific to the IDH1 R132H mutation. In addition, sequencing of the IDH1 and IDH2 genes was performed in 21 patients. MGMT promoter methylation status was assessed by quantitative methylation-specific PCR, and the proliferation index was determined by immunohistochemistry using an anti-MKI67 antibody. The extent of resection was expressed as the percentage of residual enhancing tumor volume on early (within 48 hours) postoperative MRI examination compared to the volume on the preoperative scan. The cutoffs for gross total resection (GTR), near-total resection (NTR), subtotal resection (STR) and partial resection (PR) were 100%, 95-99%, 80-94% and <80%, respectively. Radiochemotherapy according to the Stupp protocol was the standard postoperative treatment. However, some patients received hypofractionated radiotherapy (40.05 Gy in 15 fractions) combined with concurrent and adjuvant temozolomide, radiotherapy only, temozolomide only, radiotherapy combined with nivolumab (CheckMate 498) or no adjuvant treatment. Postoperative treatment planning was unavailable in one patient.
Quantitative reverse transcription PCR
Total RNA was extracted from glioblastoma samples using a RNeasy Micro Kit (Qiagen, 74004). The RNA samples were quantified using a Qubit 4.0 Fluorometer (Invitrogen, Carlsbad, CA), and cDNA was produced using a GoScript™ Reverse Transcription Mix, Random Primers (Promega, A2801). Quantitative PCR was performed with iQ™ SYBR® Green Supermix (Bio-Rad, 1708882) using a CFX96 Touch real-time PCR detection system (Bio-Rad, USA). The housekeeping genes GAPDH and RPL13A were used to normalize RNA expression (23). Relative expression was calculated using the Pffafl method. The following primers were used: DIAPH3 forward primer GATGAAACACGGTTGGCAGAGTC, DIAPH3 reverse primer ACTGCTCA-GGTTCACATAAGTTGC; GAPDH forward primer GTCTCCTCTGACTTCAACAGCG, GAPDH reverse primer ACCACCCTGTTGCTGTAGCCAA; RPL13A forward primer CTCA-AGGTGTTTGACGGCATCC, RPL13A reverse primer TACTTCCAGCCAACCTCGTGAG.
Deep bisulfite sequencing
Genomic DNA was extracted from glioblastoma samples using the QIAamp DNA Micro Kit (Qiagen, 56304) and quantified using the Qubit 4.0 Fluorometer (Invitrogen, Carlsbad, CA). In brief, 50 ng of genomic DNA per sample was bisulfite-converted with an EZ-96 DNA Methylation Deep-Well Kit (Zymo Research, Irvine, CA, USA). To determine the accuracy of DNA methylation measurement, a standard curve of 0%, 25%, 50%, 75% and 100% methylated DNA was included. These standards were prepared using human low-methylated and high-methylated genomic DNAs (Epigendx, Hopkinton). To generate amplicons specific to the CpG island in the DIAPH3 promoter, a first PCR was performed with forward and reverse primers including an overhang sequence. The PCR products were cleaned with Ampure XP (Beckman Coulter, Brea) (1.2× beads) and pooled per sample. A second PCR was performed using Nextera XT v2 primers. The samples were pooled and sequenced on an Illumina MiSeq sequencer at 2×300 bp (V3 chemistry). The generated FASTQ files were analyzed using amplikyzer2, a Python-based tool (24). Briefly, the data were demultiplexed and aligned to the reference sequence of each amplicon, and methylation percentage values were calculated per amplicon for each sample at single-CpG resolution. To test the association between DNA methylation levels and DIAPH3 expression, Spearman’s correlation was performed using R software (version 4.0.2). All P values were adjusted for multiple testing (Padj) using Bonferroni correction. The following primers were used: amplicon 1 forward primer TCGTCGGCAGCGTCAGATGTGTATAAGAGA-CAGAAAATAAAACTTAATCCCCAAATTC, amplicon 1 reverse primer GTCTCGTGGGCT-CGGAGATGTGTATAAGAGACAGGTTGGGTTAGGTTGTGTTGATTGT; amplicon 2 forward primer TCGTCGGCAGCGTCAGATGTGTATAAGAGACAGACAATCAACACAACCTAACC-CAAC, amplicon 2 reverse primer GTCTCGTGGGCTCGGAGATGTGTATAAGAGACAGTTT-AGTTTTGTTGGAATTTTATTTG; amplicon 3 forward primer TCGTCGGCAGCGTCAGAT-GTGTATAAGAGACAGAGGGTTTTAGTAGAATTGGAAGGTG, amplicon 3 reverse primer GTCTCGTGGGCTCGGAGATGTGTATAAGAGACAGAAACTCCTAAAAAACTCAACCTAA-CC. The overhanging sequences are italicized.
Statistical analyses
Survival was estimated by Kaplan−Meier analysis and then compared by the log-rank test. Data were censored at the time of last follow-up, and the median follow-up time was calculated using the reverse Kaplan−Meier method. For the comparison of categorical variables, Pearson’s chi-squared test, Fisher’s exact test or the likelihood-ratio test was used, when applicable. For the comparison of continuous variables, the independent-samples t test or Mann−Whitney test was used after verification of the normality of the distribution by the Kolmogorov−Smirnov test and Shapiro−Wilk test. Univariate and multivariate Cox proportional hazards analyses were performed to estimate predictors of overall survival (OS). Hazard ratios (HRs) with 95% confidence intervals (CIs) were calculated. Statistical analyses were performed using IBM SPSS Statistics for Windows, version 27 (IBM Corp., Armonk, N.Y., USA). Graphs were created using GraphPad Prism for Windows, version 9.1.2 (GraphPad Software, La Jolla, California, USA). n.s., not significant; *, P < 0.05; **, P < 0.01; ***, P < 0.001. The center values, 95% CIs, sample sizes, P values and statistical tests used are specified in the legends of figures and tables.
Results
DIAPH3 is frequently altered in glioblastoma and expressed in proliferating malignant cells
According to TCGA, 36% of glioblastoma patients harbor copy number alterations of DIAPH3, most of which (35%) are copy losses (Figure 1A), whereas only 1% of these patients have a mutation in DIAPH3 (Figure 1B). Notably, DIAPH3 copy loss is caused by the focal loss of the locus 13q22.1, which harbors DIAPH3 (25). Single-cell RNA sequencing analyses in glioblastoma (20) showed that DIAPH3 is mainly expressed in malignant cell population (13.73% of malignant cells express DIAPH3, compared to 1.99% of macrophages, 1.06% of T cells and 0% of oligodendrocytes). Interestingly, DIAPH3 grossly overlapped with MKI67 in proliferating subpopulations of cells (Figures 1C, D). Accordingly, cell state-based hierarchical clustering of malignant cell population (20) showed that the expression of DIAPH3 and MKI67 is much higher in neural-progenitor-like, oligodendrocyte-progenitor-like, and astrocyte-like states than in the mesenchymal-like state (Figures 1E, F).
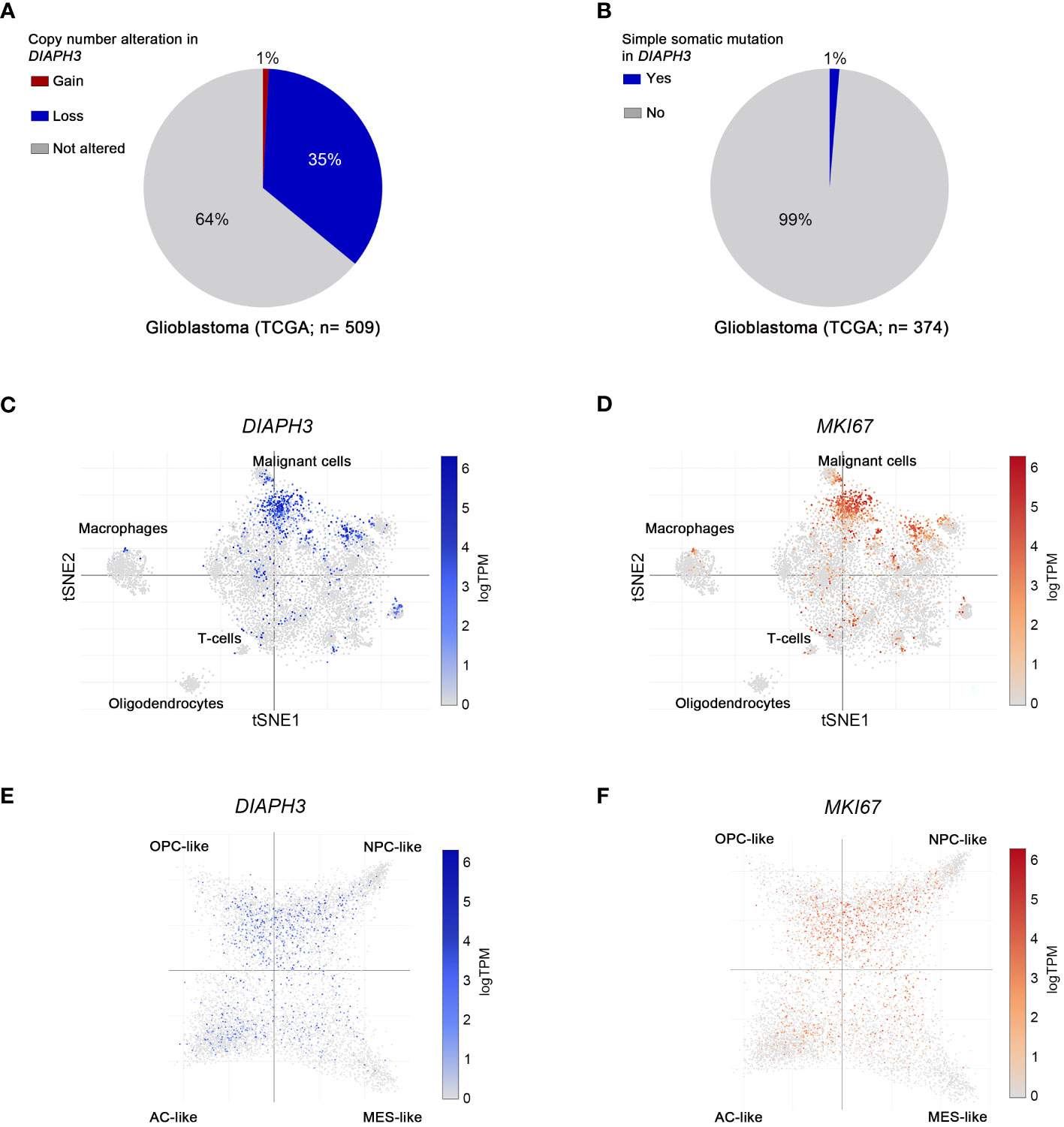
Figure 1 Molecular profiling of DIAPH3 in glioblastoma according to publicly available datasets. (A, B) The frequency of glioblastoma patients harboring gains [(A), red; frequency=1%, n=509], losses [(A), blue; frequency=35%, n=509] or simple somatic mutations [(B); frequency=1%, n=374] in DIAPH3 according to the “The Cancer Genome Atlas” (TCGA) program. (C, D) t-distributed stochastic neighbor embedding (tSNE) plots of single cells isolated from glioblastoma samples. Glioblastoma cell composition, DIAPH3 (C) and MKI67 (D) expression at a single cell resolution are shown. (E, F) Cell state-based hierarchical clustering of glioblastoma cell populations showing the expression of DIAPH3 (E) and MKI67 (F) in neural-progenitor-like (NPC-like), oligodendrocyte-progenitor-like (OPC-like), astrocyte-like (AC-like), and mesenchymal-like (MES-like) states. TCGA, the cancer genome atlas; TPM, transcripts per million; tSNE, t-distributed stochastic neighbor embedding; NPC-like, neural-progenitor-like; OPC-like, oligodendrocyte-progenitor-like; AC-like, astrocyte-like; MES-like, mesenchymal-like.
DIAPH3 expression predicts survival of patients with MGMT-methylated glioblastoma
Given the association of DIAPH3 loss with aneuploidy in murine embryonic neural stem cells (10, 11), and the negative impact of aneuploidy on cancer prognosis (13), we investigated whether DIAPH3 levels could affect the prognosis of human glioblastoma. We evaluated DIAPH3 relative expression by quantitative reverse transcription PCR in 73 IDH-wild-type glioblastomas from patients operated on between May 2013 and August 2019 (Figure 2A). The median patient follow-up period was 51.0 months (95% CI: 24.6-77.5), and the median OS was 15.6 months (95% CI: 13.5-17.7; Figure 2B). This analysis uncovered variable DIAPH3 expression levels with median and mean values of 0.155 and 0.243 arbitrary units (a.u.), respectively. We used the median value as a cutoff and grouped patients into DIAPH3-high (n=36) and DIAPH3-low (n=37) groups (Figure 2C). Kaplan−Meier survival analysis revealed a longer OS in the DIAPH3-high group than in the DIAPH3-low group (P=0.002, log-rank test; HR=0.454, 95% CI: 0.274-0.751, P=0.002, Cox proportional hazards analysis; Figure 2D), despite comparable clinicopathological characteristics (Table 1), comparable postoperative treatment (Table 2), and better microsurgical resections in the DIAPH3-low group (Table 2). For the DIAPH3-high group, the median OS was 20.2 months (95% CI: 14.9-25.5; Figure 2D) versus 11.7 months (95% CI: 7.3-16.2; Figure 2D) in the DIAPH3-low group. Univariate Cox proportional hazards analysis revealed that age (HR=2.178, 95% CI: 1.211-3.919, P=0.009), methylation of the MGMT promoter (HR=0.497, 95% CI: 0.292-0.846, P=0.010), administration of radiochemotherapy according to the Stupp protocol (HR=0.399, 95% CI: 0.214-0.745, P=0.004) and the expression level of DIAPH3 (HR=0.454, 95% CI: 0.274-0.751, P=0.002) were predictors of OS (Figure 2E). Multivariate analysis including these variables confirmed that age (HR=2.298, 95% CI: 1.031-5.122, P=0.042), methylation of the MGMT promoter (HR=0.343, 95% CI: 0.188-0.628, P=0.001), and the expression level of DIAPH3 (HR=0.487, 95% CI: 0.289-0.819, P=0.007) can independently predict the OS of glioblastoma patients (Figure 2F).
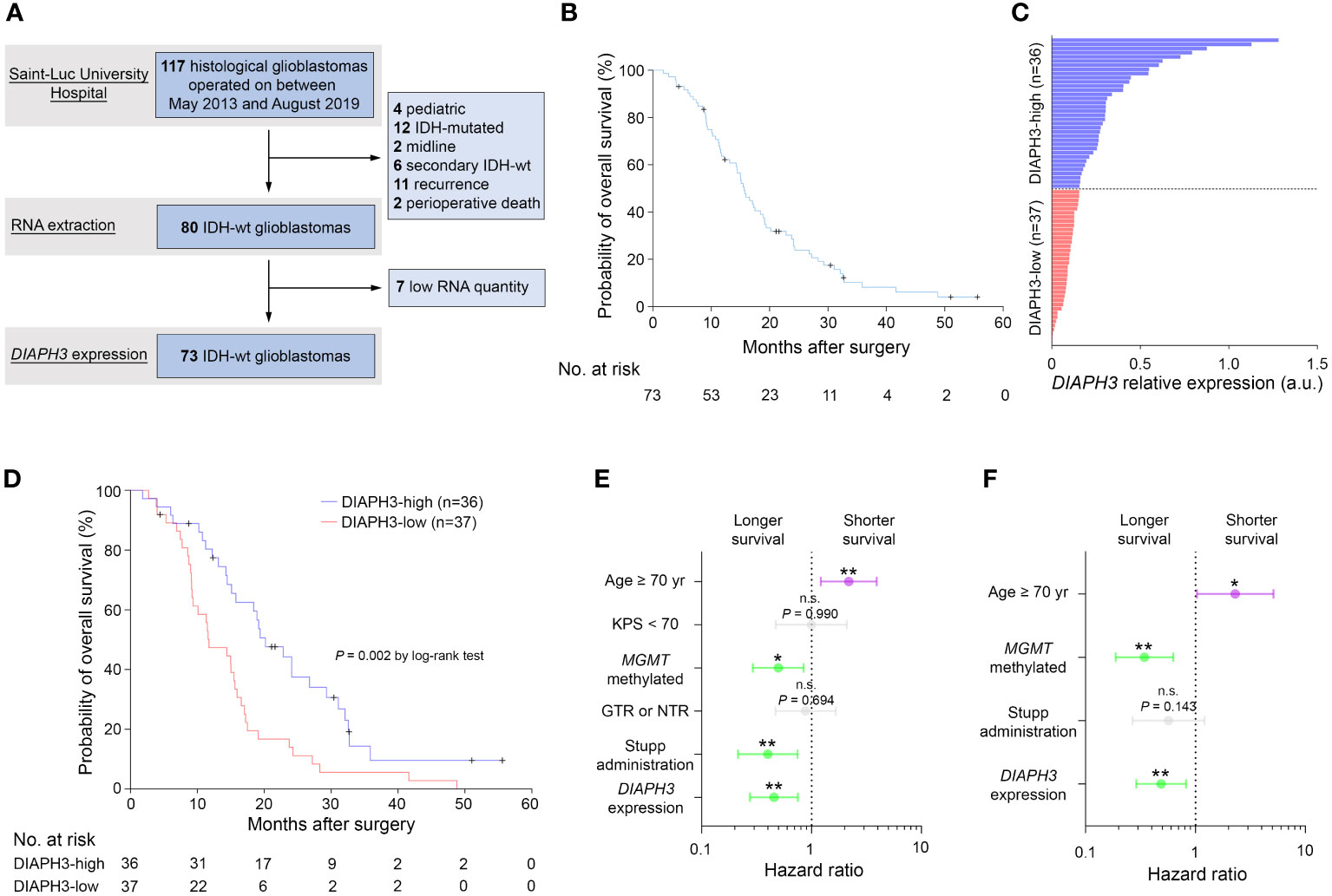
Figure 2 Low expression of DIAPH3 predicts a poor clinical course of glioblastoma. (A) Data flow diagram for the glioblastoma patient cohort. (B) Kaplan−Meier analysis for overall survival (OS) (median OS=15.6 months, 95% CI: 13.5-17.7, n=73). (C) DIAPH3 relative expression in 73 IDH-wild-type glioblastoma patients. DIAPH3-high (n=36) and DIAPH3-low (n=37) groups were formed using the median value of DIAPH3 relative expression as a cutoff. (D) Kaplan−Meier analysis for OS (DIAPH3-high: median OS=20.2 months, 95% CI: 14.9-25.5, n=36; DIAPH3-low: median OS=11.7 months, 95% CI: 7.3-16.2, n=37; P=0.002 by log-rank test). (E) Univariate Cox proportional hazards analysis for OS (Age ≥ 70 yrs: HR=2.178, 95% CI: 1.211-3.919, P=0.009; KPS < 70: HR=0.995, 95% CI: 0.472-2.099, P=0.990; MGMT methylated: HR=0.497, 95% CI: 0.292-0.846, P=0.010; GTR or NTR: HR=0.881, 95% CI: 0.468-1.658, P=0.694; Stupp administration: HR=0.399, 95% CI: 0.214-0.745, P=0.004; DIAPH3 expression: HR=0.454, 95% CI: 0.274-0.751, P=0.002). (F) Multivariate Cox proportional hazards analysis for OS (Age ≥ 70 yrs: HR=2.298, 95% CI: 1.031-5.122, P=0.042; MGMT methylated: HR=0.343, 95% CI: 0.188-0.628, P=0.001; Stupp administration: HR=0.568, 95% CI: 0.267-1.210, P=0.143; DIAPH3 expression: HR=0.487, 95% CI: 0.289-0.819, P=0.007). wt, wild-type; a.u., arbitrary units; KPS, Karnofsky performance status; GTR, gross total resection; NTR, near-total resection. n.s., not significant; *, P < 0.05; **, P < 0.01; ***, P < 0.001.
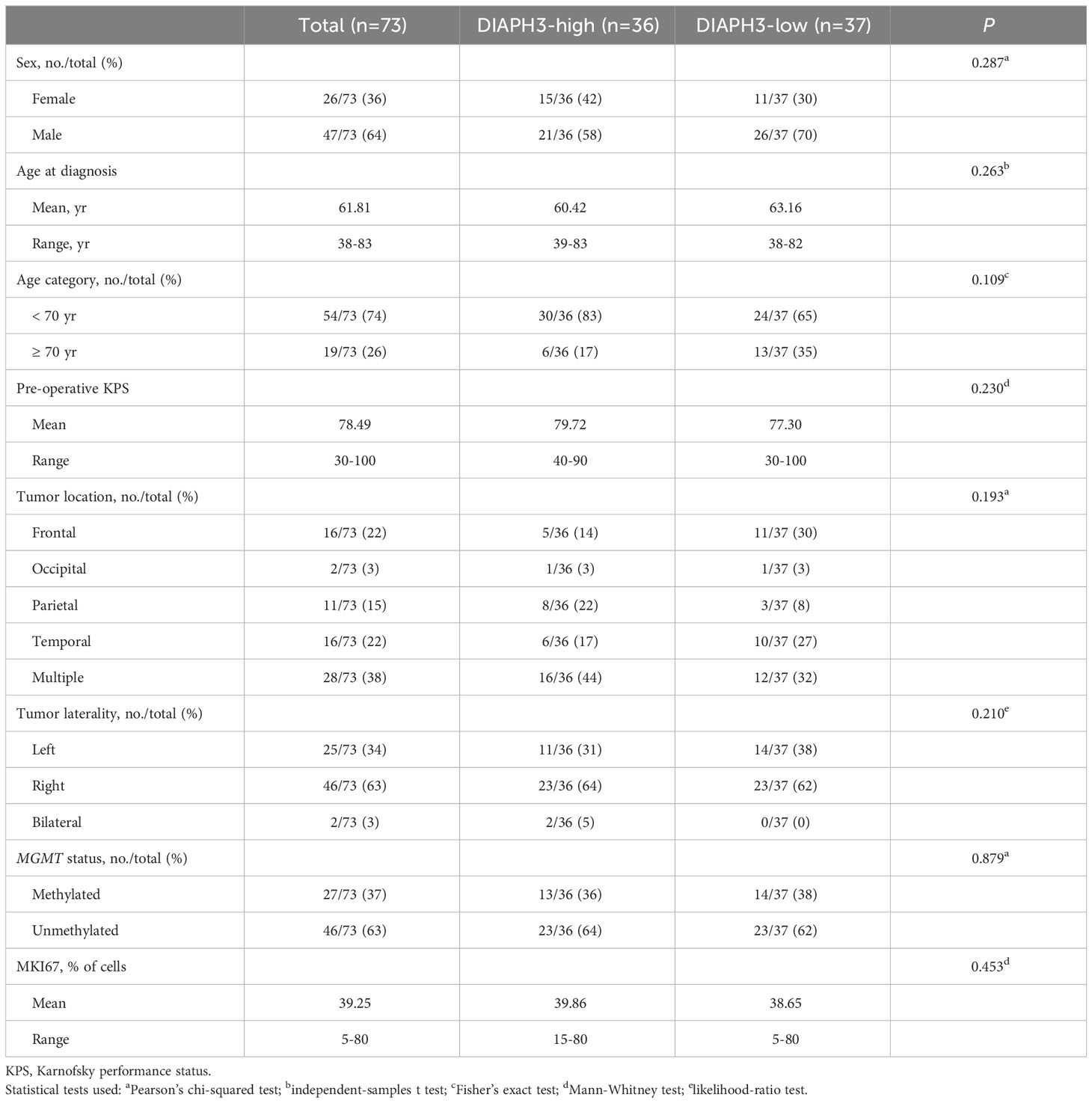
Table 1 Clinicopathological characteristics of patients.
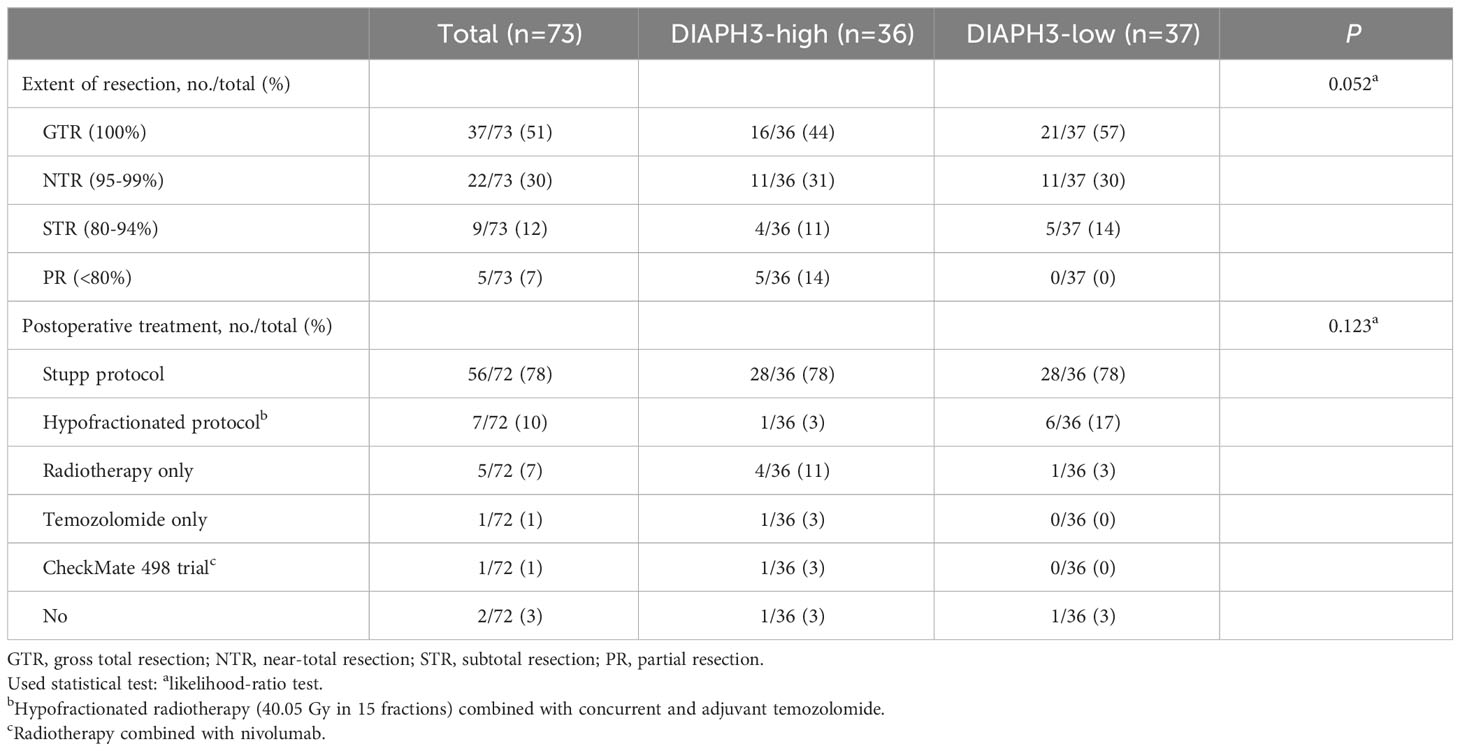
Table 2 Treatment received by patients.
Importantly, when we analyzed separately the effect of DIAPH3 expression on survival in MGMT-methylated and unmethylated glioblastomas, we observed a striking difference between high and low expression of DIAPH3 within the MGMT-methylated group (P=0.018, log-rank test; HR=0.369, 95% CI: 0.156-0.871, P=0.023, Cox proportional hazards analysis; Figure 3A), while the effect of DIAPH3 expression level was not significant within the MGMT-unmethylated group (P=0.086, log-rank test; HR=0.579, 95% CI: 0.307-1.091, P=0.091, Cox proportional hazards analysis; Figure 3B). To corroborate this finding, we analyzed the relationship between DIAPH3 expression and OS in the TCGA IDH-wild-type glioblastoma cohort (n=109; Supplementary Figure 1A). By setting the median value as a cutoff (Supplementary Figure 1B), no difference in OS was observed between DIAPH3-high and DIAPH3-low groups (P=0.190, log-rank test; HR=0.748, 95% CI: 0.484-1.157, P=0.192, Cox proportional hazards analysis; Supplementary Figure 1C). However, when we stratified the samples according to the MGMT methylation status, we detected a positive correlation between DIAPH3 expression and overall survival in the MGMT-methylated group (MGMT-methylated: P=0.035, log-rank test; HR=0.475, 95% CI: 0.234-0.963, P=0.039, Cox proportional hazards analysis; MGMT-unmethylated: P=0.616, log-rank test; HR=1.160, 95% CI: 0.650-2.071, P=0.616, Cox proportional hazards analysis; Figures 3C, D). These results suggest that DIAPH3 expression can predict survival of patients with MGMT-methylated glioblastomas.
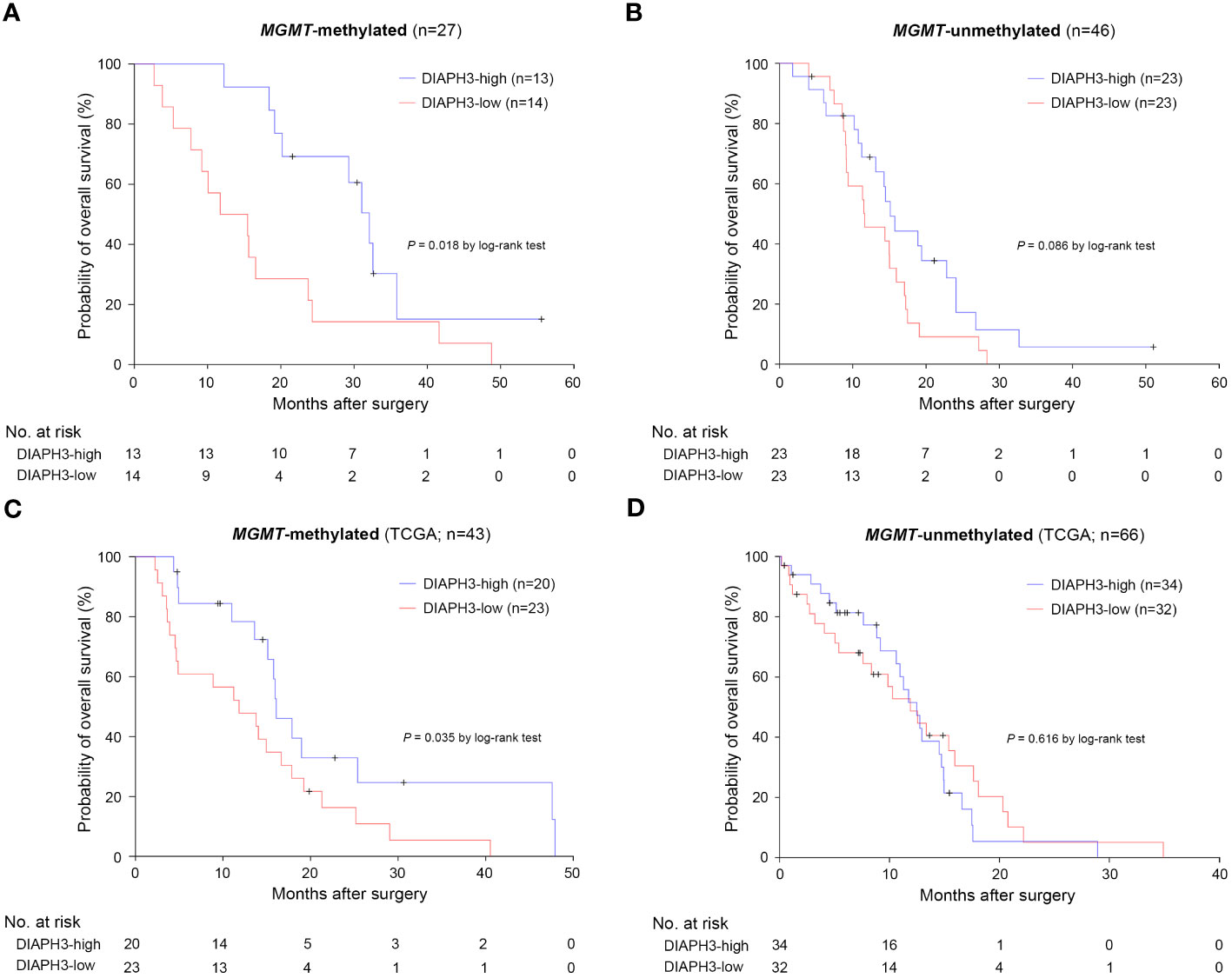
Figure 3 The impact of DIAPH3 expression on survival is more pronounced in MGMT-methylated glioblastoma. (A) Kaplan−Meier analysis for overall survival (OS) in MGMT-methylated glioblastoma (DIAPH3-high: median OS=32.1 months, 95% CI: 28.1-36.1, n=13; DIAPH3-low: median OS=11.7 months, 95% CI: 1.9-21.6, n=14; P=0.018 by log-rank test). (B) Kaplan−Meier analysis for OS in MGMT-unmethylated glioblastoma (DIAPH3-high: median OS=15.1 months, 95% CI: 13.0-17.3, n=23; DIAPH3-low: median OS=11.6 months, 95% CI: 5.9-17.4, n=23; P=0.086 by log-rank test). (C) Kaplan−Meier analysis for OS in the TCGA MGMT-methylated glioblastoma cohort (DIAPH3-high: median OS=16.1 months, 95% CI: 13.5-18.7, n=20; DIAPH3-low: median OS=11.8 months, 95% CI: 4.2-19.5, n=23; P=0.035 by log-rank test). (D) Kaplan−Meier analysis for OS in the cancer genome atlas (TCGA) MGMT-unmethylated glioblastoma cohort (DIAPH3-high: median OS=12.5 months, 95% CI: 10.2-14.8, n=34; DIAPH3-low: median OS=11.9 months, 95% CI: 7.8-16.0, n=32; P=0.616 by log-rank test). TCGA, the cancer genome atlas.
Finally, we comparatively analyzed the overall survival according the MGMT promoter methylation but independently of the DIAPH3 expression level, we detected a higher effect of MGMT methylation in Saint-Luc University Hospital cohort compared to TCGA (Saint-Luc University Hospital Cohort, MGMT-unmethylated: median OS=14.5 months, 95% CI: 10.9-18.0, n=46; MGMT-methylated: median OS=20.2 months, 95% CI: 11.4-29.0, n=27; P=0.009, log-rank test; TCGA cohort, MGMT-unmethylated: median OS=12.5 months, 95% CI: 10.5-14.4, n=66; MGMT-methylated: median OS=15.1 months, 95% CI: 12.6-17.6, n=43; P=0.022, log-rank test; Supplementary Figures 2A, B).
Methylation of three CpG sites in the DIAPH3 promoter are associated with DIAPH3 downregulation
To investigate the mechanisms underlying the regulation of DIAPH3 expression, we screened the DIAPH3 promoter for CpG islands using different in silico tools (e.g., “EMBOSS Cpgplot” and “DataBase of CpG islands and Analytical Tool”) and public databases. All these tools predicted the presence of a CpG island spanning 13:60163334-60164405 (Figures 4A, B). We assessed the methylation of this CpG island in glioblastoma samples using deep bisulfite sequencing. After conversion, we sequenced three amplicons spanning 62 CpG sites in 72 glioblastoma samples for which DIAPH3 expression was available (Figure 4C). Although the DIAPH3 promoter was mostly unmethylated, three CpG sites, namely, CpG 6, CpG 28 and CpG 29, showed a variable level of methylation between samples (0-8%, 0-7% and 0-3% for CpG 6, CpG 28 and CpG 29, respectively; Figure 4D). Importantly, the methylation levels at these three CpG sites were negatively correlated with DIAPH3 expression (Padj=0.008, 0.002, and 0.003 for CpG 6, CpG 28 and CpG 29, respectively; Figure 4E), suggesting that the methylation of these sites contributes to DIAPH3 downregulation in glioblastoma samples.
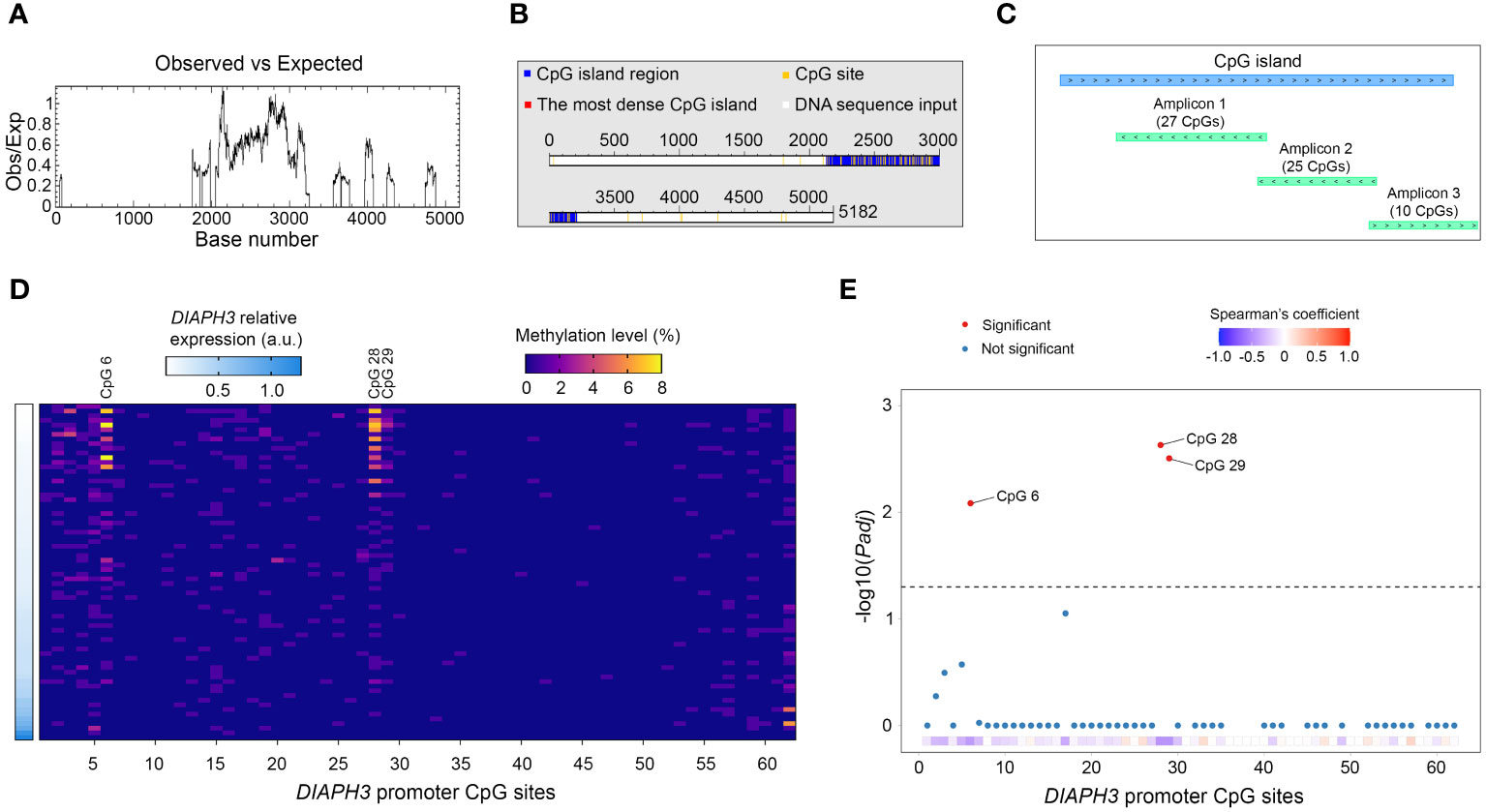
Figure 4 Methylation profiling of DIAPH3 promoter in glioblastoma. (A, B) In silico screening of cytosine-phosphate-guanine (CpG) islands in the promoter region of DIAPH3 (5182 nucleotides) using “EMBOSS Cpgplot” (A) and “DataBase of CpG islands and Analytical Tool” (DBCAT) (B). (C) Schematic diagram illustrating the main CpG island in DIAPH3 promoter region as well as the different amplicons sequenced, after bisulfite conversion. (D) Heatmap showing DIAPH3 relative expression as well as the methylation level of 62 DIAPH3 promoter CpG sites in 72 glioblastoma samples. (E) Spearman’s correlation between CpG methylation and DIAPH3 expression (CpG 6: Spearman’s coefficient=-0.431, Padj=0.008; CpG 28: Spearman’s coefficient=-0.462, Padj=0.002; CpG 29: Spearman’s coefficient=-0.455, Padj=0.003). CpG, cytosine-phosphate-guanine; a.u., arbitrary units.
Discussion
Despite extensive advances in the molecular characterization of glioblastoma, its treatment and prognosis have not improved over the last two decades. Hence, there are still critical gaps in the understanding of this disease’s pathophysiology.
In this study, we investigated the expression of DIAPH3 in glioblastoma and uncovered a positive correlation between DIAPH3 expression level and patients’ survival. Importantly, the impact of DIAPH3 was more prominent in MGMT-methylated glioblastomas. The clinical interest in the methylation status of MGMT promoter in glioblastoma patients stemmed from the implementation of temozolomide as a standard of care treatment in 2005 (26). Methylation of the MGMT promoter increases the sensitivity to temozolomide in glioblastoma patients, extending their survival (27). However, survival curves between MGMT-methylated and MGMT-unmethylated glioblastomas diverge starting from nine months (Supplementary Figure 2A) and (27), suggesting that other factors may contribute to predict survival in the MGMT-methylated glioblastomas. DIAPH3 could be one of these factors, since assessing its expression in MGMT-methylated tumors offers a better prediction of patient survival. Our results suggest that MGMT and DIAPH3 may cooperatively contribute to the repair of temozolomide-induced DNA damage. In the absence of MGMT (MGMT-methylated) DIAPH3 would affect the response to temozolomide whereas in its presence (MGMT-unmethylated) the effect of DIAPH3 would be not significant (our cohort) or masked (TCGA cohort). Mechanistically, we speculate that the low expression of DIAPH3 in proliferating malignant cells could favor aneuploidy, as found in murine embryonic neural stem cells. Aneuploidy in turn, would increase endogenous DNA damage through oxidative stress (increase in reactive oxygen species) (28) and replication stress (stalled replication forks) (29), and activate intrinsic DNA damage response. Further investigations are needed to test this hypothesis.
The impact of DIAPH3 expression on survival in the TCGA cohort is milder than in our cohort. We believe that the TCGA database may not be optimal because it is multicentric and therefore heterogeneous. It includes patients operated on between 1997 and 2011, a long period spanning the pre- and post-temozolomide eras. This has a considerable impact on survival especially in the MGMT-methylated group as evidenced by their rather low median OS in TCGA (15.1 months), compared with our cohort (20.2 months) and the initial report by Hegi and colleagues (21.7 months) (27).
DIAPH3 expression is a tightly regulated process. During embryogenesis in mice, Diaph3 is ubiquitously expressed before the ninth embryonic day. However, as development proceeds, its expression becomes more confined. In the brain, Diaph3 is exclusively expressed by neural stem/progenitor cells and excluded from postmitotic cells. Using targeted deep bisulfite sequencing, a highly sensitive method, we show that methylation of three CpG sites in the DIAPH3 promoter contributes at least partially to its regulation in glioblastoma. The methylation level of the three CpG sites that correlate with low expression of DIAPH3 is mild (maximum 8%). This level is likely underestimated given the significant tumor cell heterogeneity, and the fact that whereas the expression of DIAPH3 is restricted to proliferating cells (Figures 1C–F) (20), the methylation level was calculated as the percentage of methylated CpG in all tumor cells. Other epigenetic (e.g., histone acetylation) or genetic mechanisms may also be implicated. For instance, a point mutation in the 5’ untranslated region of DIAPH3 increases DIAPH3 expression, leading to auditory neuropathy autosomal dominant 1 (AUNA1) (30), suggesting that this mutation may impede the binding of a transcriptional repressor. Moreover, DIAPH3 copy number variations could impact its expression level through a gene dosage effect. A better understanding of the molecular mechanisms underlying DIAPH3 expression should help identify modifiers of DIAPH3 expression with therapeutic potential. Of note, two types of modulators of DIAPH3 activity have been described: small molecule inhibitor of FH2 domain (SMIFH2), which inhibits formins (31), and intramimics 01 and 02 (IMM-01 and IMM-02), which activate them (32). The main weakness of these modulators is their lack of specificity, given that they modify the activity of multiple formins, increasing the probability of potential side effects. Hence, the search for molecules able to specifically target DIAPH3 remains essential.
Conclusions
In this study, we report that DIAPH3 expression is positively correlated with overall survival of patients with MGMT-methylated glioblastoma. We show that DIAPH3 is mostly expressed in proliferating malignant cells in glioblastoma and that the methylation of three CpG sites in the DIAPH3 promoter contributes to its downregulation.
Data availability statement
All the data generated in the study have been included in the main manuscript or as a Supplementary Material.
Ethics statement
The studies involving humans were approved by Comité d’Ethique Hospitalo-Facultaire Saint-Luc –UCL, agreement number 2018/27NOV/450. The studies were conducted in accordance with the local legislation and institutional requirements. The human samples used in this study were acquired from The Saint-Luc University Hospital Biobank. Written informed consent for participation was not required from the participants or the participants’ legal guardians/next of kin in accordance with the national legislation and institutional requirements.
Author contributions
FT: Conceptualization, Funding acquisition, Project administration, Resources, Supervision, Validation, Writing – original draft, Writing – review & editing. GC: Conceptualization, Formal analysis, Investigation, Writing – original draft, Writing – review & editing. NE: Formal analysis, Investigation, Methodology, Writing – review & editing. MA: Investigation, Project administration, Validation, Writing – review & editing. MIA: Formal analysis, Validation, Visualization, Writing – review & editing. YB: Investigation, Methodology, Visualization, Writing – review & editing. AM: Investigation, Visualization, Writing – review & editing. AA: Investigation, Visualization, Writing – review & editing. MV: Formal analysis, Investigation, Methodology, Writing – review & editing. JL: Investigation, Resources, Validation, Writing – review & editing. NR: Investigation, Validation, Writing – review & editing. ID: Investigation, Validation, Visualization, Writing – review & editing. CR: Investigation, Methodology, Resources, Validation, Writing – review & editing. NT: Conceptualization, Formal analysis, Investigation, Validation, Visualization, Writing – review & editing.
Funding
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. This work was supported by the following grants: F/2022/1967 from the Belgian Foundation against Cancer, PDR T0236.20 from the Fund for Scientific Research (FNRS); CDR J.0175.23 from the FNRS; 0913351 from the FNRS/FWO-Excellence of Science Program; and SARA-HBKU-OVPR-TG-HBKU-INT-VPR-TG-02-10 from Hamad Bin Khalifa University. This work was made possible by NPRP-Standard (NPRP-S) 14th Cycle grant # [NPRP14S-0404-210140] from the Qatar National Research Fund (a member of Qatar Foundation). The findings herein reflect the work, and are solely the responsibility, of the authors. Open Access funding provided by the Qatar National Library. GC is a Research Fellow FNRS and the laureate of the 2021 Helaers Research Prize for Neurosurgery, and FT is an Honorary Research Director FNRS.
Acknowledgments
We thank Isabelle Lambermont, Salma El Mere, and Mila Jhamai for technical assistance; and Aurélie Bertrand for helping with the statistical analysis.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The author(s) declared that they were an editorial board member of Frontiers, at the time of submission. This had no impact on the peer review process and the final decision.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fonc.2024.1359652/full#supplementary-material
Supplementary Figure 1 | Relationship between DIAPH3 expression and survival in the cancer genome atlas IDH-wild-type glioblastoma cohort. (A) Data flow diagram for the cancer genome atlas (TCGA) IDH-wild-type glioblastoma patient cohort. (B) DIAPH3 expression in 109 IDH-wild-type glioblastoma patients. DIAPH3-high (n=54) and DIAPH3-low (n=55) groups were formed using the median value of DIAPH3 expression as a cutoff. (C) Kaplan−Meier analysis for overall survival (OS) (DIAPH3-high: median OS=14.7 months, 95% CI: 12.4-17.1, n=54; DIAPH3-low: median OS=11.9 months, 95% CI: 8.1-15.7, n=55; P=0.190 by log-rank test). TCGA, the cancer genome atlas; wt, wild-type; TPM, transcripts per million.
Supplementary Figure 2 | Overall survival according to the MGMT methylation status. (A) Kaplan−Meier analysis for overall survival (OS) in Saint-Luc University Hospital glioblastoma cohort (MGMT-unmethylated: median OS=14.5 months, 95% CI: 10.9-18.0, n=46; MGMT-methylated: median OS=20.2 months, 95% CI: 11.4-29.0, n=27; P=0.009 by log-rank test). (B) Kaplan−Meier analysis for OS in the cancer genome atlas (TCGA) glioblastoma cohort (MGMT-unmethylated: median OS=12.5 months, 95% CI: 10.5-14.4, n=66; MGMT-methylated: median OS=15.1 months, 95% CI: 12.6-17.6, n=43; P=0.022 by log-rank test). TCGA, the cancer genome atlas.
References
1. Goode BL, Eck MJ. Mechanism and function of formins in the control of actin assembly. Annu Rev Biochem. (2007) 76:593–627. doi: 10.1146/annurev.biochem.75.103004.142647
2. Chesarone MA, DuPage AG, Goode BL. Unleashing formins to remodel the actin and microtubule cytoskeletons. Nat Rev Mol Cell Biol. (2010) 11:62–74. doi: 10.1038/nrm2816
3. Higgs HN, Peterson KJ. Phylogenetic analysis of the formin homology 2 domain. Mol Biol Cell. (2005) 16:1–13. doi: 10.1091/mbc.e04-07-0565
4. Watanabe N, Madaule P, Reid T, Ishizaki T, Watanabe G, Kakizuka A, et al. p140mDia, a mammalian homolog of Drosophila diaphanous, is a target protein for Rho small GTPase and is a ligand for profilin. EMBO J. (1997) 16:3044–56. doi: 10.1093/emboj/16.11.3044
5. Bione S, Sala C, Manzini C, Arrigo G, Zuffardi O, Banfi S, et al. A human homologue of the Drosophila melanogaster diaphanous gene is disrupted in a patient with premature ovarian failure: evidence for conserved function in oogenesis and implications for human sterility. Am J Hum Genet. (1998) 62:533–41. doi: 10.1086/301761
6. Alberts AS, Bouquin N, Johnston LH, Treisman R. Analysis of RhoA-binding proteins reveals an interaction domain conserved in heterotrimeric G protein beta subunits and the yeast response regulator protein Skn7. J Biol Chem. (1998) 273:8616–22. doi: 10.1074/jbc.273.15.8616
7. Tominaga T, Sahai E, Chardin P, McCormick F, Courtneidge SA, Alberts AS. Diaphanous-related formins bridge Rho GTPase and Src tyrosine kinase signaling. Mol Cell. (2000) 5:13–25. doi: 10.1016/S1097-2765(00)80399-8
8. Watanabe S, Ando Y, Yasuda S, Hosoya H, Watanabe N, Ishizaki T, et al. mDia2 induces the actin scaffold for the contractile ring and stabilizes its position during cytokinesis in NIH 3T3 cells. Mol Biol Cell. (2008) 19:2328–38. doi: 10.1091/mbc.e07-10-1086
9. Mei Y, Zhao B, Yang J, Gao J, Wickrema A, Wang D, et al. Ineffective erythropoiesis caused by binucleated late-stage erythroblasts in mDia2 hematopoietic specific knockout mice. Haematologica. (2016) 101:e1–5. doi: 10.3324/haematol.2015.134221
10. Lau EO, Damiani D, Chehade G, Ruiz-Reig N, Saade R, Jossin Y, et al. DIAPH3 deficiency links microtubules to mitotic errors, defective neurogenesis, and brain dysfunction. Elife. (2021) 10. doi: 10.7554/eLife.61974
11. Damiani D, Goffinet AM, Alberts A, Tissir F. Lack of Diaph3 relaxes the spindle checkpoint causing the loss of neural progenitors. Nat Commun. (2016) 7:13509. doi: 10.1038/ncomms13509
12. Vitale I, Galluzzi L, Castedo M, Kroemer G. Mitotic catastrophe: a mechanism for avoiding genomic instability. Nat Rev Mol Cell Biol. (2011) 12:385–92. doi: 10.1038/nrm3115
13. Ben-David U, Amon A. Context is everything: aneuploidy in cancer. Nat Rev Genet. (2020) 21:44–62. doi: 10.1038/s41576-019-0171-x
14. Taylor AM, Shih J, Ha G, Gao GF, Zhang X, Berger AC, et al. Genomic and functional approaches to understanding cancer aneuploidy. Cancer Cell. (2018) 33:676–89.e3. doi: 10.1016/j.ccell.2018.03.007
15. Hager MH, Morley S, Bielenberg DR, Gao S, Morello M, Holcomb IN, et al. DIAPH3 governs the cellular transition to the amoeboid tumour phenotype. EMBO Mol Med. (2012) 4:743–60. doi: 10.1002/emmm.201200242
16. Morley S, You S, Pollan S, Choi J, Zhou B, Hager MH, et al. Regulation of microtubule dynamics by DIAPH3 influences amoeboid tumor cell mechanics and sensitivity to taxanes. Sci Rep. (2015) 5:12136. doi: 10.1038/srep12136
17. Lizarraga F, Poincloux R, Romao M, Montagnac G, Le Dez G, Bonne I, et al. Diaphanous-related formins are required for invadopodia formation and invasion of breast tumor cells. Cancer Res. (2009) 69:2792–800. doi: 10.1158/0008-5472.CAN-08-3709
18. Shibue T, Brooks MW, Inan MF, Reinhardt F, Weinberg RA. The outgrowth of micrometastases is enabled by the formation of filopodium-like protrusions. Cancer Discovery. (2012) 2:706–21. doi: 10.1158/2159-8290.CD-11-0239
19. Becker KN, Pettee KM, Sugrue A, Reinard KA, Schroeder JL, Eisenmann KM. The cytoskeleton effectors Rho-kinase (ROCK) and mammalian diaphanous-related (mDia) formin have dynamic roles in tumor microtube formation in invasive glioblastoma cells. Cells. (2022) 11. doi: 10.3390/cells11091559
20. Neftel C, Laffy J, Filbin MG, Hara T, Shore ME, Rahme GJ, et al. An integrative model of cellular states, plasticity, and genetics for glioblastoma. Cell. (2019) 178:835–49.e21. doi: 10.1016/j.cell.2019.06.024
21. Louis DN, Perry A, Wesseling P, Brat DJ, Cree IA, Figarella-Branger D, et al. The 2021 WHO classification of tumors of the central nervous system: a summary. Neuro Oncol. (2021) 23:1231–51. doi: 10.1093/neuonc/noab106
22. Chehade G, Lawson TM, Lelotte J, Daoud L, Di Perri D, Whenham N, et al. Long-term survival in patients with IDH-wildtype glioblastoma: clinical and molecular characteristics. Acta Neurochir (Wien). (2023) 165:1075–85. doi: 10.1007/s00701-023-05544-3
23. Kreth S, Heyn J, Grau S, Kretzschmar HA, Egensperger R, Kreth FW. Identification of valid endogenous control genes for determining gene expression in human glioma. Neuro Oncol. (2010) 12:570–9. doi: 10.1093/neuonc/nop072
24. Rahmann S, Beygo J, Kanber D, Martin M, Horsthemke B, Buiting K. Amplikyzer: Automated methylation analysis of amplicons from bisulfite flowgram sequencing. PeerJ. (2013) 1:e122v2. doi: 10.7287/peerj.preprints.122v2
25. Brennan CW, Verhaak RG, McKenna A, Campos B, Noushmehr H, Salama SR, et al. The somatic genomic landscape of glioblastoma. Cell. (2013) 155:462–77. doi: 10.1016/j.cell.2013.09.034
26. Stupp R, Mason WP, van den Bent MJ, Weller M, Fisher B, Taphoorn MJ, et al. Radiotherapy plus concomitant and adjuvant temozolomide for glioblastoma. N Engl J Med. (2005) 352:987–96. doi: 10.1056/NEJMoa043330
27. Hegi ME, Diserens AC, Gorlia T, Hamou MF, de Tribolet N, Weller M, et al. MGMT gene silencing and benefit from temozolomide in glioblastoma. N Engl J Med. (2005) 352:997–1003. doi: 10.1056/NEJMoa043331
28. Li M, Fang X, Baker DJ, Guo L, Gao X, Wei Z, et al. The ATM-p53 pathway suppresses aneuploidy-induced tumorigenesis. Proc Natl Acad Sci U.S.A. (2010) 107:14188–93. doi: 10.1073/pnas.1005960107
29. Passerini V, Ozeri-Galai E, de Pagter MS, Donnelly N, Schmalbrock S, Kloosterman WP, et al. The presence of extra chromosomes leads to genomic instability. Nat Commun. (2016) 7:10754. doi: 10.1038/ncomms10754
30. Schoen CJ, Emery SB, Thorne MC, Ammana HR, Sliwerska E, Arnett J, et al. Increased activity of Diaphanous homolog 3 (DIAPH3)/diaphanous causes hearing defects in humans with auditory neuropathy and in Drosophila. Proc Natl Acad Sci U S A. (2010) 107:13396–401. doi: 10.1073/pnas.1003027107
31. Rizvi SA, Neidt EM, Cui J, Feiger Z, Skau CT, Gardel ML, et al. Identification and characterization of a small molecule inhibitor of formin-mediated actin assembly. Chem Biol. (2009) 16:1158–68. doi: 10.1016/j.chembiol.2009.10.006
Keywords: diaphanous formin, glioblastoma, mDia2, O(6)-methylguanine-DNA methyltransferase, MGMT methylation, survival, The Cancer Genome Atlas
Citation: Chehade G, El Hajj N, Aittaleb M, Alkailani MI, Bejaoui Y, Mahdi A, Aldaalis AAH, Verbiest M, Lelotte J, Ruiz-Reig N, Durá I, Raftopoulos C, Tajeddine N and Tissir F (2024) DIAPH3 predicts survival of patients with MGMT-methylated glioblastoma. Front. Oncol. 14:1359652. doi: 10.3389/fonc.2024.1359652
Received: 21 December 2023; Accepted: 07 February 2024;
Published: 22 February 2024.
Edited by:
Lincoln A. Edwards, University of Pittsburgh Medical Center, United StatesReviewed by:
Aiguo Li, National Institutes of Health (NIH), United StatesJack M. Shireman, University of Wisconsin-Madison, United States
Copyright © 2024 Chehade, El Hajj, Aittaleb, Alkailani, Bejaoui, Mahdi, Aldaalis, Verbiest, Lelotte, Ruiz-Reig, Durá, Raftopoulos, Tajeddine and Tissir. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Fadel Tissir, ZmFkZWwudGlzc2lyQHVjbG91dmFpbi5iZQ==; ZnRpc3NpckBoYmt1LmVkdS5xYQ==
†ORCID: Fadel Tissir, orcid.org/0000-0002-9292-6622
Georges Chehade, orcid.org/0000-0001-5172-2378
Nuria Ruiz-Reig, orcid.org/0000-0001-7008-7920