 Silvijus Abramavicius1,2
Silvijus Abramavicius1,2 Asbjørn G. Petersen1
Asbjørn G. Petersen1 Nirthika S. Renaltan1
Nirthika S. Renaltan1 Judit Prat-Duran1Roberta Torregrossa3Edgaras Stankevicius4
Judit Prat-Duran1Roberta Torregrossa3Edgaras Stankevicius4 Matthew Whiteman3
Matthew Whiteman3 Ulf Simonsen1*
Ulf Simonsen1*- 1Department of Biomedicine, Pulmonary and Cardiovascular Pharmacology, Aarhus University, Aarhus, Denmark
- 2Institute of Physiology and Pharmacology, Lithuanian University of Health Sciences, Kaunas, Lithuania
- 3Medical School, University of Exeter, Exeter, United Kingdom
- 4Institute of Cardiology, Lithuanian University of Health Sciences, Kaunas, Lithuania
Donors of H2S may be beneficial in treating cardiovascular diseases where the plasma levels of H2S are decreased. Therefore, we investigated the mechanisms involved in relaxation of small arteries induced by GYY4137 [(4-methoxyphenyl)-morpholin-4-yl-sulfanylidene-sulfido-λ5-phosphane;morpholin-4-ium], which is considered a slow-releasing H2S donor. Sulfides were measured by use of 5,5′-dithiobis-(2-nitro benzoic acid), and small rat mesenteric arteries with internal diameters of 200–250 µm were mounted in microvascular myographs for isometric tension recordings. GYY4137 produced similar low levels of sulfides in the absence and the presence of arteries. In U46619-contracted small mesenteric arteries, GYY4137 (10−6–10–3 M) induced concentration-dependent relaxations, while a synthetic, sulfur-free, GYY4137 did not change the vascular tone. L-cysteine (10−6–10–3 M) induced only small relaxations reaching 24 ± 6% at 10–3 M. Premixing L-cysteine (10–3 M) with Na2S and GYY4137 decreased Na2S relaxation and abolished GYY4137 relaxation, an effect prevented by an nitric oxide (NO) synthase inhibitor, L-NAME (Nω-nitro-L-arginine methyl ester). In arteries without endothelium or in the presence of L-NAME, relaxation curves for GYY4137 were rightward shifted. High extracellular K+ concentrations decreased Na2S and abolished GYY4137 relaxation suggesting potassium channel-independent mechanisms are also involved Na2S relaxation while potassium channel activation is pivotal for GYY4137 relaxation in small arteries. Blockers of large-conductance calcium-activated (BKCa) and voltage-gated type 7 (KV7) potassium channels also inhibited GYY4137 relaxations. The present findings suggest that L-cysteine by reaction with Na2S and GYY4137 and formation of sulfides, inhibits relaxations by these compounds. The low rate of release of H2S species from GYY4137 is reflected by the different sensitivity of these relaxations towards high K+ concentration and potassium channel blockers compared with Na2S. The perspective is that the rate of release of sulfides plays an important for the effects of H2S salt vs. donors in small arteries, and hence for a beneficial effect of GYY4137 for treatment of cardiovascular disease.
Introduction
Hydrogen sulfide (H2S) is considered an essential signaling molecule in the cardiovascular and nervous systems (Szabó, 2007; Wallace et al., 2018) and a variety of pathophysiological changes including cancer, glycometabolic disorders, diabetes, sepsis, and human malignt hyperthermia are associated with altered endogenous levels of H2S (Szabó, 2007; Szabo and Papapetropoulos, 2017; Vellecco et al., 2020). In the cardiovascular system, endogenous H2S can lead to both vasodilatation and vasoconstriction (Li et al., 2015; Hedegaard et al., 2016; Gheibi et al., 2018).
Several mechanisms mediate vasodilatation induced by addition of exogenous H2S salts, including lowering of smooth muscle cells calcium by activation of K channels (Skovgaard et al., 2011; Hedegaard et al., 2016), enhancement of nitric oxide (NO) signaling (Szabo, 2017), and changes in intracellular pH by inhibition of an acid-sensitive Cl2/HCO3-exchanger (Lee et al., 2006; Esechie et al., 2008; Malekova et al., 2009; Perniss et al., 2017). The opening of potassium channels leads to hyperpolarization and smooth muscle relaxation. Different types of K channels are involved in H2S vasodilatation, including in large arteries ATP-sensitive K channels (KATP) (Zhao and Wang, 2002; Kubo et al., 2007; Webb et al., 2008; Martelli et al., 2013a), voltage-gated K channels (KV7, KCNQ) (Martelli et al., 2013a; Hedegaard et al., 2014), and 4-aminopyridine-sensitive voltage-gated potassium channels (Cheang et al., 2010). In resistance arteries, H2S vasodilatation involves KATP channels (Tang et al., 2005), large-conductance calcium-dependent potassium channels (BKCa) (Jackson-Weaver et al., 2011; Jackson-Weaver et al., 2013), and KV7 channels (Schleifenbaum et al., 2010; Hedegaard et al., 2016), but also potassium channel-independent vasodilatation (Hedegaard et al., 2016).
In a variety of human diseases, e.g., hypertension and atherosclerotic disease, the plasma levels of H2S are decreased (Wang, 2012). Several series of H2S donors have recently been developed to substitute for the decreased H2S levels (Feng et al., 2015; Steiger et al., 2017; Szabo and Papapetropoulos, 2017). The H2S donating compounds compromises of two major groups: the inorganic salts NaHS and Na2S, which are rapid H2S releasers, and compounds associated with a slow release of H2S, e.g., diallyl disulfide and GYY4137 (4-methoxyphenyl)-morpholin-4-yl-sulfanylidene-sulfido-λ5-phosphane;morpholin-4-ium) (Li et al., 2008; Martelli et al., 2013b) and AP39, AP123, and AP67 (Whiteman et al., 2006). NaSH and Na2S produce an instant pH-dependent dissociation to H2S and at high concentrations, e.g., 1 mM, induce vasorelaxation (Zhao et al., 2001; Li et al., 2008), while 100 µM GYY4137 associated with release of <1 µM H2S is associated with relaxation (Hedegaard et al., 2016).
GYY4137 exhibits vasorelaxant, hypotensive, anti-inflammatory, and anti-cancer activity effects (Li et al., 2008; Martelli et al., 2013a; Lee et al., 2011; Liu et al., 2013), and it is considered as a slow-releasing H2S donor (Li et al., 2008; Feng et al., 2015). Different mechanisms have been reported to be involved in the release of sulfide species from GYY4137, including conversion by cystathionine γ-lyase (CSE) (Chitnis et al., 2013) and interaction with cysteine (Martelli et al., 2013b). In a recent study where changes in H2S gas were detected with microelectrodes, we observed that GYY4137 induced full relaxation of small mesenteric arteries without producing detectable changes in amperometric currents (Hedegaard et al., 2016). Hence, it is unclear whether H2S gas contributing to the pharmacodynamic effects of GYY4137 is below detection level or whether GYY4137 induces vasodilatation by mechanisms independent of H2S gas e.g., commercial GYY4137 has dichlormethane leading to formation of carbon monoxide (CO) (Alexander et al., 2015).
To examine whether sulfides are involved in GYY4137 relaxations, measurements of sulfides were conducted and compared to a hydrolyzed version of GYY4137 (Alexander et al., 2015). Na2S was chosen for comparison as its vasodilating effects previously have been associated with increases in H2S gas (Hedegaard et al., 2016). To investigate whether CSE or L-cysteine contribute to the release of H2S from GYY4137, the effect of an inhibitor of CSE and L-cysteine were examined on GYY4137 relaxation. Release of H2S from Na2S and GYY4137 appear to have different kinetics and that may change the involvement of potassium channels, and therefore relaxations induced by the two compounds were investigated in the presence of blockers of potassium channels. Small mesenteric arteries contribute significantly to vascular resistance and blood pressure in intact animals (Fenger-Gron et al., 1995), and therefore the vasodilatation studies were performed in rat small mesenteric arteries.
Methods and Materials
Animals and Preparation of Samples
The investigation was carried out in accordance to the Guide for the Care and the Use of Laboratory Animals published by the United States National Institutes of Health (NIH Publication No. 85–23, revised 1996) and followed the ARRIVE guidelines (McGrath and Lilley, 2015). Adult male Wistar rats (12–14 weeks) were killed by decapitation and subsequent exsanguination. The protocol was approved by the Danish Animal Experiments Inspectorate (permission 2014-15-2934-0159).
Chemicals and Materials
The following drugs were used: noradrenaline, acetylcholine (ACh), L-NAME (Nω-nitro-L-arginine methyl ester), Na2S, XE991 [10,10-bis(4-pyridinylmethyl)-9(10H)-anthracenone dihydrochloride], glibenclamide from Sigma-Aldrich (St. Louis, MO), 5,5′-dithiobis-(2-nitro benzoic acid) (DTNB) (Sigma), D,L-propargylglycine (PPG; an irreversible inhibitor of the enzyme cystathionine γ-lyase (CSE), an H2S synthase inhibitor), 1,4-Dithiothreitol (DTT). GYY4137 ((4-methoxyphenyl)-morpholin-4-yl-sulfanyl-sulfanylidene- λ5-phosphane sodium salt) was synthesized, as described previously by us (Alexander et al., 2015). Fresh Na2S solution was prepared every day. To neutralize pH of the solution, hydrochloric acid was added until a pH of 7.35–7.45 was obtained. The composition of the physiologic salt solution (PSS) was NaCl 119 mM, NaHCO3 25 mM, glucose 5.5 mM, CaCl2 1.6 mM, KH2PO4 1.18 mM, MgSO4 1.17 mM, and EDTA 0.027 mM. High potassium solution, KPSS, was PSS with NaCl exchanged for KCl on equimolar basis.
Measurements of Release of Sulfide Species From GYY4137
Sulfide species (H2S and HS−) released from GYY4137 were assessed in vitro as described previously (Li et al., 2008). In brief, 100 mM phosphate buffer pH 7.40 was incubated with 1.0 or 0.1 mM GYY4137 at 25 or 37°C. A phosphate buffer with pH of 3.01 was also tested as acidic conditions have been shown to promote H2S release from GYY4137 (Li et al., 2008; Hedegaard et al., 2016). At appropriate times, 20 µl aliquots were removed and added to 96-well microplates containing 50 µl 1 mM DTNB and 50 µl 1 M HEPES buffer pH 8.0, and absorbance was measured at 412 nm on a plate reader. The concentration of H2S formed from GYY4137 was calculated from a standard curve of NaSH (1–500 µM) for each of the respective time points.
Functional Studies in Small Mesenteric Arteries
Third branch mesenteric arteries were dissected from the mesenteric vascular bed and mounted on 40 µm steel wires in microvascular myographs (Danish Myotechnology, Aarhus, Denmark) for isometric tension recording as previously described (Mulvany and Halpern, 1976). The vessels were equilibrated in oxygenated (5% CO2, 20% O2, 75% N2) PSS at 37°C and for 30 min, and by stretching normalized to a lumen diameter (d100) equivalent to 100 mm Hg, after which tension was set to 90% x d100 (Mulvany and Halpern, 1976). At this tension, the internal lumen diameters were 200–250 µm. After normalization, the arterial segments were stimulated with KPSS, washed in PSS, and stimulated with noradrenaline (10 µM). Arteries were only included if they developed an active force corresponding to a transmural pressure of 100 mmHg. The PowerLab data system and Chart 5.5 (ADInstruments, Oxfordshire, United Kingdom) was used to record the data. The mechanical responses of the vessel segments were measured as active wall tension (ΔT), which is the changes in force (ΔF) divided by twice the segment length (2L).
Experimental Protocol
To determine whether the effects of GYY4137 were due to H2S released from it, the parent compound was compared with an analog, which is normally produced by a two-step hydrolytic degradation of GYY4137 over weeks, but in this study synthesized as previously described (Alexander et al., 2015).
The biosynthetic enzymes for H2S production (CBS and CSE) were reported to be involved in the relaxant effects of GYY4137 in bovine ciliary arteries (Chitnis et al., 2013). Therefore, small mesenteric arteries were incubated with D, L-propargylglycine (PPG, 10 mM), which is an irreversible inhibitor of CSE, and concentration-response curves were constructed for Na2S and GYY4137. L-cysteine is considered substrate for formation of endogenous H2S (Wang, 2012), and was also reported as a scavenger of HS− giving rise to formation of sulfides (Koike et al., 2017). Concentration-response curves for L-cysteine were constructed in U46619 (0.3 µM)-contracted preparations. To investigate an eventual scavenger effect, we conducted two set of experiments. In a first set of experiments, solutions of cysteine and, respectively, Na2S and GYY4137 were pre-mixed in airtight containers and then added to the arteries contracted with U46619 at 0 and 10 h after the mixing. At 10 h the amount of sulfides was also measured using the DTNB assay as described above. In a second series of experiments, small mesenteric arteries were incubated with L-cysteine (10–3 M) or the thiol reducing agent, 1,4-dithiothreitol (10−3 M), and concentration-response curves were constructed for GYY4137 and Na2S. The control and examination of drugs were run in parallel, and only one concentration-response curve was constructed for each vasodilator per animal.
To investigate the role of the endothelium in relaxations induced by Na2S and GYY4137, arterial segments with and without endothelium were mounted. The endothelial cells were removed by introducing into to the lumen a human scalp hair and rubbing back and forth several times (Hedegaard et al., 2016). The effectiveness of the procedure was assessed by absence of relaxation to acetylcholine in noradrenaline-contracted arteries, while vessel with endothelium were accepted only if acetylcholine-induced (10–5 M) relaxation on noradrenalin-induced (5 × 10–6 M) contraction was larger than 50%, and exclusion following these criteria explains unequal group numbers are reported. The preparations were contracted with U46619 (0.3 µM) giving a contraction level corresponding to 50–60% of the contractions induced by 125 mM KPSS, and when the contraction was stable cumulative concentration-response curves were constructed for Na2S (10−6–10–3 M) or GYY4137 (10−6–10–3 M). Preparations were incubated with the NO synthase inhibitor, L-NAME (300 µM), the preparations were contracted with U46619 (0.1 µM) to obtain contraction similar to controls levels, concentration-response curves were constructed for increasing concentrations of Na2S, GYY4137, and acetylcholine.
High concentrations of the NO donor, sodium nitroprusside (SNP) with NaSH yield formation of a nitrosothiol and inhibits NO-induced rat aorta relaxation (Ali et al., 2006; Whiteman et al., 2006). To investigate the interaction with NO, concentration-response curves for SNP (10−9–10–4 M) were obtained in the absence and presence of GYY4137 or Na2S.
The involvement of K channels in Na2S and GYY4137 induced relaxation were examined by comparing relaxations in U46619 and high-potassium physiologic saline solution (KPSS)-contracted arteries. To investigate the specific K channels involved, the preparations were incubated for 30 min with a selective blocker of ATP-sensitive K channels, glibenclamide (1 µM) (Mulvany et al., 1990), a selective blocker of large-conductance calcium-activated K channels, iberiotoxin (100 nM) (Giangiacomo et al., 1992), and a blocker of voltage-gated KV7 channels, XE991 (10 µM) (Yeung et al., 2007), and concentration-response curves were obtained for GYY4137 and Na2S.
Data and Statistical Analysis
All data were presented as mean ± S.E.M. with a significance level of p < 0.05, and n representing the number of individual animals (n > 5 for each protocol). Statistical comparisons between H2S release at time 0 and 10 h by GYY4137 and Na2S were performed by Student´s t-test. The two-way analysis of variance (ANOVA) was used to compare the different conditions affecting release of sulfide species from GYY4137 and concentration-response curves obtained in functional studies of isolated mesenteric arteries. The assumptions of the ANOVA approach were verified by inspection of Q-Q plots. The graphs and statistical analyses were performed using GraphPad Prism 7.0 (GraphPad Software, La Jolla, CA).
Results
Role of H2S in Na2S and GYY4137 Relaxation
The release of free H2S from GYY4137 was examined spectrophotometrically by the use of DTNB. Following previous studies (Li et al., 2008), incubation of 0.1 mM GYY4137 at pH 7.4 25°C resulted in a slow-release, which reached an end value of 8.33 µM after 90 min of incubation (Supplementary Figure S1A). This release was augmented by an increased starting concentration of GYY4137 (1 mM) (Supplementary Figure S1A). Sulfide release was also significantly increased under acidic conditions and by increased temperatures (Supplementary Figures S1B,C respectively). L-cysteine by itself increased the spectrophotometrically measured absorbance, but there was a further increase in sulfide release by combining cysteine and GYY4137 (Supplementary Figure S1D). The presence of rat mesenteric artery did not affect sulfide release from GYY4137 (Figure 1A). The results suggest that the release of sulfides from GYY4137 is independent of the presence of mesenteric artery.
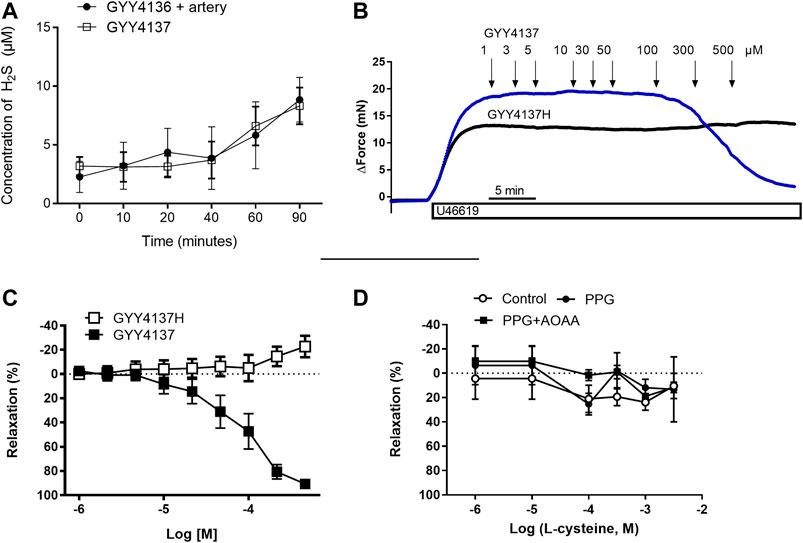
FIGURE 1. Role of H2S for GYY4137 relaxations small mesenteric arteries. (A) Sulfide release from GYY4137 in the absence and the presence of small mesenteric arteries (B) Original recordings in small mesenteric arteries showing contraction to 3 µM U46619 and addition of increasing concentrations of GYY4137 (1–500 µM) and the hydrolyzed form of GYY4137H (1–500 µM) (C) Average relaxations induced by the parent compound and the hydrolyzed form of GYY4137. (D) Effect of the substrate for H2S synthesis, L-cysteine, on vascular tone in small mesenteric arteries. The arteries were incubated with an inhibitors of H2S biosynthesis using the cystathionine-gamma-lyase (CSE) inhibitor, PPG (1 mM), and the cystathionine-beta-synthase (CBS)/cystathionine-gamma-lyase (CSE) inhibitor aminooxyacetic acid (AOAA, 1 mM). L-cysteine was added in increasing concentrations (1 µM-3 mM). Data are means ± SEM (n = 6).
Concentration-response curves were obtained in U46619 (0.3 µM)-contracted preparations for GYY4137 and the hydrolyzed product of GYY4137, GYY4137H to investigate whether the relaxant effects of GYY4137 were due to H2S release. We found that GYY4137 induced concentration-dependent relaxations while there was no change in vascular tone by adding the GYY4137H (Figures1B,C), suggesting that release of sulfides is pivotal for GYY4137 relaxation.
In U46619-contracted arteries with endothelium, the H2S substrate, and thiol-containing amino acid, L-cysteine (10−6–10–2 M) induced small relaxations which were 24 ± 6% at 10–3 M, and these relaxations were not inhibited in the presence of PPG (Figure 1D). Pre-mixing L-cysteine (10–3 M) with either Na2S or GYY4137 and adding it immediately after the mixing or 10 h later showed, that the presence of L-cysteine inhibited Na2S and GYY4137 relaxations (Figures 2A,B). The relaxations induced by Na2S and GYY4137 were comparable 10 h after the mixing of the solutions (Figures 2A,B), and this was also the case for the amount of sulfides measured in the solutions (Figures 2C,D). After preincubation with L-cysteine (10−3 M), instead of relaxations,3 × 10–6 to 10–3 M Na2S and 10−4–10–3 M GYY4137 induced contractions, which at the highest concentrations were contractions followed by relaxations (Figures 3A–D). In the presence of both L-NAME and L-cysteine, Na2S-induced contractions were reduced (Figures 3C,G), while the inhibitory effect of L-cysteine on GYY4137 relaxation was prevented (Figures 3F,H). To examine whether enzymatic conversion by CSE or interaction with thiol-groups play a role for Na2S and GYY4137 relaxations, the preparations were pre-incubated with an inhibitor of CSE, PPG (10–3 M), or the thiol reducing agent DTT (10–3 M), but these treatments did not change concentration-response curves for Na2S and GYY4137 (Figure 4).
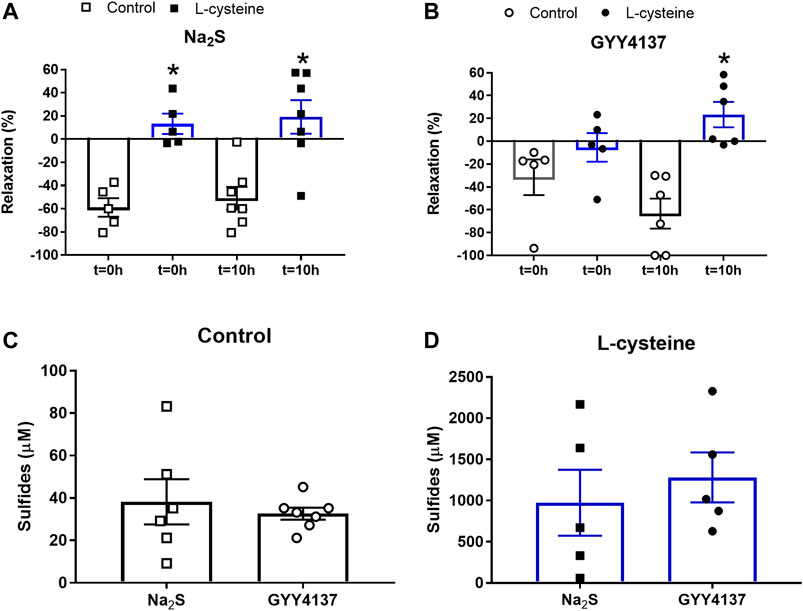
FIGURE 2. Effect of L-cysteine on Na2S and GYY4137 relaxations in rat small mesenteric arteries. Mixture of Na2S (300 µM) or GYY4137 (1 mM) with and without L-cysteine (1 mM) was added to air-tight bottles and relaxations measured to time = 0 and 10 h for (A) Na2S and (B) GYY4137, and the amount of sulfides measured at 10 h for (C) Na2S and GYY4137 in the absence, and for (D) Na2S and GYY4137 in the presence of L-cysteine, where the concentration of sulfides generated by L-cysteine were subtracted. Please note the difference in scale comparing data in (C and D). Data are means ± SEM (n = 6). *p < 0.05, Students t-test.
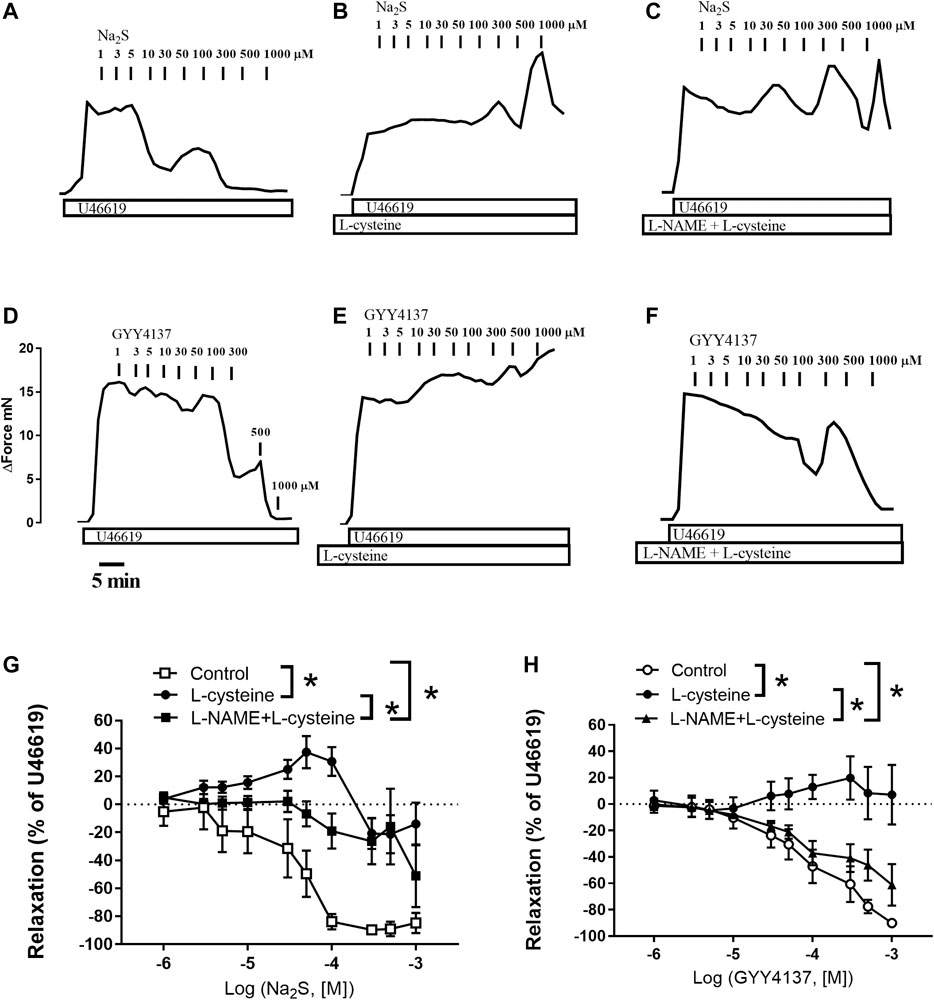
FIGURE 3. Effect of L-cysteine and L-NAME incubation on Na2S and GYY4137 relaxations in rat small mesenteric arteries. Original traces showing the effect of increasing concentrations of Na2S in U46619-contracted preparations in (A) control conditions (B) in the presence of L-cysteine (1 mM), and (C) in the presence of L-cysteine and the nitric oxide synthase inhibitor, L-NAME (100 µM). Original traces showing the effect of increasing concentrations of GYY4137 in U46619-contracted preparations in (D) control conditions (E) in the presence of L-cysteine (1 mM), and (F) in the presence of L-cysteine and the nitric oxide synthase inhibitor, L-NAME (100 µM). The horizontal bar indicates time in min and the vertical bar increase in force in mN (G) Average effect of L-cysteine without and with L-NAME on Na2S-induced vascular relaxation (H) Average effect of L-cysteine without and with L-NAME on GYY4137-induced vascular relaxation. Data are means ± SEM (n = 6). *p < 0.05, two-way ANOVA.
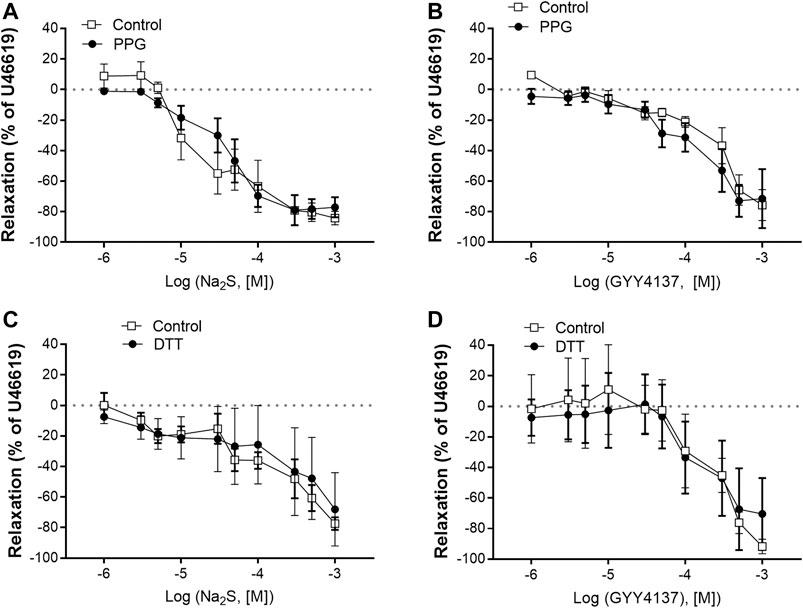
FIGURE 4. Effect of the cystathionine γ-lyase (CSE) inhibitor, PPG, and DTT on Na2S and GYY4137 relaxations in rat mesenteric arteries (A) Effect of PPG (1 mM) on Na2S induced vascular relaxation (B) effect of PPG on GYY4137 induced vascular relaxation (C) effect of DTT (1 mM) on Na2S induced vascular relaxation (D) effect of DTT on GYY4137 induced vascular relaxation. Where no error bars are indicated, error lies within dimensions of the symbol. *p < 0.05, two-way ANOVA. All data is represented as mean ± SEM (n = 5–6).
Taken together our results show that L-cysteine converts Na2S and GYY4137 relaxations to contraction associated with markedly higher sulfide concentrations. This effect of L-cysteine on vascular tone was partly reversed in the presence of the endothelial NO synthase inhibitor, L-NAME.
Effect of Endothelial Cell Removal and NO in Na2S and GYY4137 Relaxation
In contrast to L-cysteine (10–3 M), incubation with L-NAME significantly rightward shifted concentration-response curves for acetylcholine relaxation in small mesenteric arteries (Figure 5A). In U46619-contracted arteries, Na2S induced concentration-dependent relaxations, which were of similar magnitude in vessel segments with and without endothelium (Figure 5B), while concentration-response curves for GYY4137 were significantly rightward shifted in vessels without endothelium (Figure 5C). In the presence of an inhibitor of NO synthase, L-NAME (10–4 M), the concentration-response curves for Na2S were unaltered (Figure 5B), while L-NAME rightward shifted concentration-response curves for GYY4137 (Figure 5C). These results suggest that in rat small mesenteric arteries, endothelium-derived NO is of importance for some of the effects of GYY4137 on vascular tone, while there were no significant differences for Na2S.
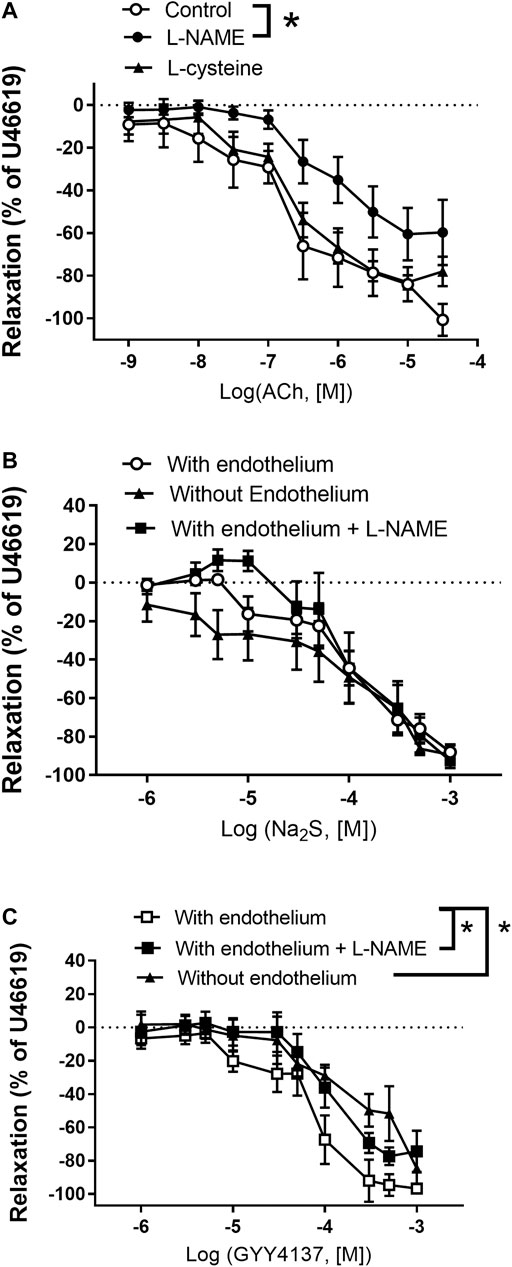
FIGURE 5. Role of the endothelium and NO for Na2S and GYY4137 relaxation (A) Average relaxations induced by acetylcholine (ACh) in the absence (n = 7) and the presence of either L-NAME (100 µM) (n = 6) or L-cysteine (1 mM) (n = 7) in rat mesenteric arteries (B) Na2S induced vascular relaxation in vessels with (n = 11) and without endothelium (n = 5), or with endothelium in the presence of an NO synthase inhibitor, L-NAME (n = 6) (C) GYY4137 induced vascular relaxation in vessels with (n = 12) and without endothelium (n = 6) or with endothelium in the presence L-NAME (n = 6). Where no error bars are indicated, error lies within dimensions of symbol. All data are means ± SEM. *p < 0.05, two-way ANOVA.
To investigate the effect of Na2S and GYY4137 on NO donor-induced relaxations, the small mesenteric arteries were incubated with vehicle, Na2S (3 × 10–4 M), or GYY4137 (10–3 M), then contracted to the same level with U46619 (0.3 µM), and increasing concentrations of SNP was added. We found that in the presence of Na2S, concentration-response curves for SNP were leftward shifted, while the presence of GYY4137 did not change the relaxation responses for SNP in small mesenteric arteries (Figure 6).
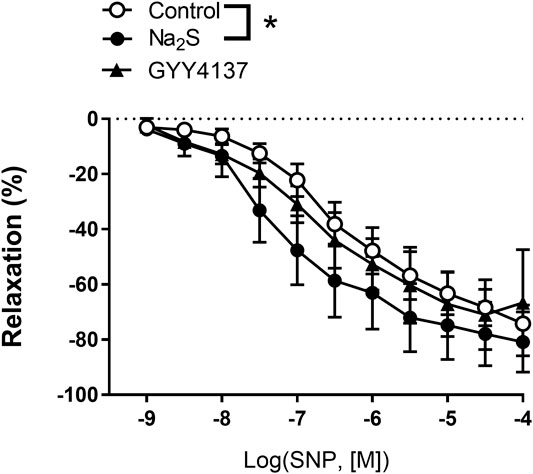
FIGURE 6. Effect of Na2S and GYY4137 on relaxations induced by the NO donor, sodium nitroprusside (SNP) in small mesenteric arteries. The arteries were incubated with and inhibitor of NO synthase, L-NAME (10 µM), and Na2S (300 µM) (n = 6), or GYY4137 (1 mM) (n = 5), contracted with U46619 (0.3 µM) and concentration-response curves constructed for SNP. *p < 0.05, two-way ANOVA. All data are means ± SEM.
Involvement of K Channels in GYY4137 and Na2S-Induced Vascular Relaxation
In preparations contracted with high extracellular potassium (60 mM KPSS), relaxations induced by GYY4137 were abolished, while relaxations induced by Na2S were significantly decreased compared with responses obtained in U46619 (0.3 µM)-contracted preparations (Figures 7A–D). In contrast to GYY4137, Na2S still induced 60% maximum relaxation in 60 mM KPSS-contracted preparations (Figures 7E,F), hence suggesting that K channels are pivotal for GYY4137-induced relaxations, while K channels and also other mechanisms contribute to Na2S relaxation. To investigate the K channel subtypes involved in the relaxations, the preparations were incubated with blockers of ATP-sensitive K channels (glibenclamide), BKCa (iberiotoxin), and of KV7 channels (XE991). Glibenclamide decreased Na2S relaxation, while GYY4137 relaxation was unaltered in U46619-contracted arteries (Figures 8A,B). Iberiotoxin and XE991 significantly decreased relaxations induced by both Na2S and GYY4137 (Figures 8C–F).
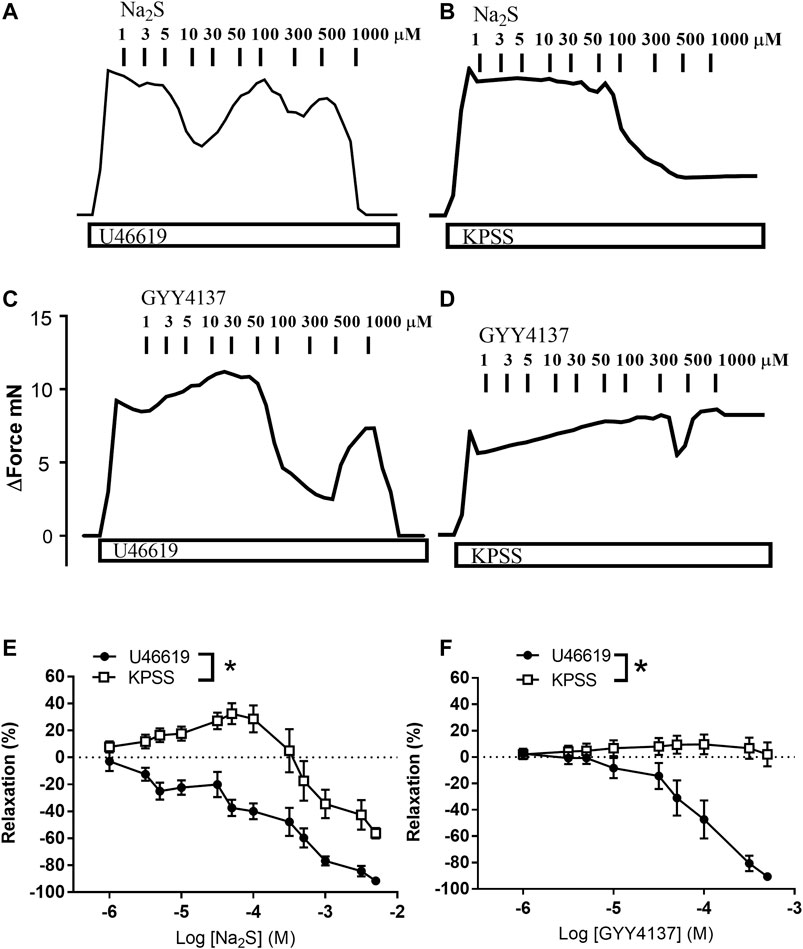
FIGURE 7. Effect of high extracellular potassium (KPSS) on Na2S and GYY4137 relaxation. Original traces showing contractions induced by 3 µM U46619 or 60 mM KPSS and effect of increasing concentrations of (A,B) Na2S (1–1,000 µM) or (C,D) GYY4137 (1–1,000 µM). (E) Average Na2S induced vascular relaxations in vessels contracted with U46619 or KPSS; (F) Average GYY4137 induced vascular relaxations in vessels contracted with U46619 or KPSS. Where no error bars are indicated, error lies within dimensions of symbol. All data are means ± SEM (n = 5–10). *p < 0.05, two-way ANOVA.
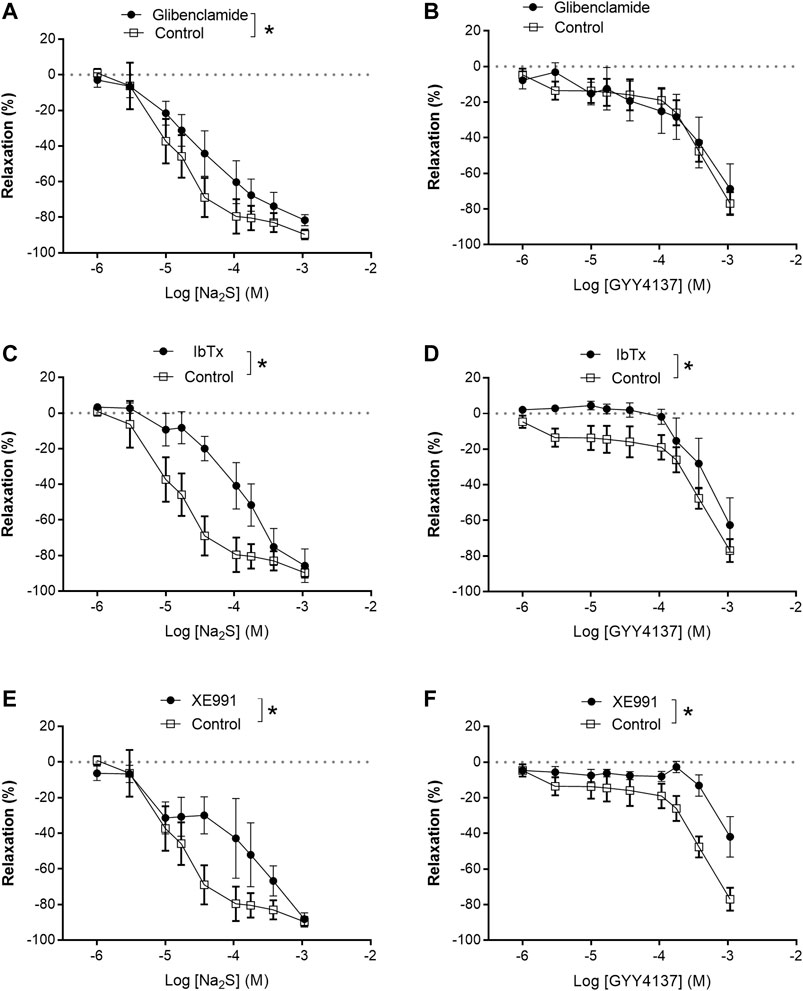
FIGURE 8. Effect of K channels blockers on Na2S and GYY4137 relaxation. (A) Effect of inhibition of KATP channels by glibenclamide (1 µM) on Na2S-induced vascular relaxation; (B) effect of inhibition of KATP channels by glibenclamide on GYY4137-induced vascular relaxation; (C) effect of selective blockade of BKCa channels by iberiotoxin (IBTX, 100 nM) on Na2S-induced vascular relaxation; (D) effect of selective blockade of BKCa channels by IBTX on GYY 4137-induced vascular relaxation; (E) effect of selective blockade of Kv7 channels by XE991 (10 µM) on Na2S-induced vascular relaxation; (F) effect of selective blockade of KV7 by XE991 on GYY4137-induced vascular relaxation. Where no error bars are indicated, errors lies within dimensions of symbol. All data are means ± SEM (n = 8–10). *p < 0.05, two-way ANOVA.
Discussion
The main findings in the present study are that GYY4137 spontaneously releases low amounts of sulfides leading to relaxation, and that L-cysteine by direct chemical interaction inhibits Na2S and GYY4137 relaxations. The observation of sulfide release is supported by the tissue-independent effect on sulfide release measured from GYY4137 and that the hydrolyzed control, GYY4137H, in contrast to the parent molecule, fails to relax small mesenteric arteries. Moreover, Na2S induced comparable relaxations after dissolving at 0 h compared to 10 h storage, while GYY4137 relaxation was markedly increased by storage for 10 h in airtight containers and yielded sulfide accumulation similar to Na2S, and suggesting GYY4137 is associated with slow release of H2S. These findings suggest rate of H2S release plays an essential role for the effect on vascular tone, where high levels of H2S from Na2S interacts leftward shift concentration-response curves for the NO donor SNP, while low levels of H2S from GYY4137 interact with endogenous endothelium-derived NO leading to relaxation. Moreover, blockers of KATP, BKCa, and KV7 channels affected Na2S and GYY4137 concentration-response curves differently.
It has previously been shown that GYY4137 releases H2S (Li et al., 2008; Whiteman et al., 2010; Martelli et al., 2013b), and in agreement with these studies, we found that GYY4137 concentration-dependently releases small amounts of sulfides. This release is markedly enhanced by increasing the temperature and acidifying the solutions. In contrast, at physiological conditions (pH 7.4, 37°C), we found by simultaneous measurements of H2S gas and relaxation that GYY4137 caused relaxation of small rat arteries without releasing detectable amounts of H2S gas (Hedegaard et al., 2016). However, the lack of relaxant effect of the hydrolyzed GYY4137 control compound, GYY4137H (Alexander et al., 2015), suggests that GYY4137-induced vessel relaxation requires H2S.
Several H2S releasing compounds with slow releasing rates, including organic polysulphides of garlic, e.g., diallyl disulfide and arylthiamides require the presence of reduced glutathione or thiols to release H2S (Benavides et al., 2007; Martelli et al., 2013b). However, in the presence of arterial tissue, the amount of sulfides measured from GYY4137 was not increased suggesting the H2S release is tissue-independent. Moreover, Na2S induced comparable relaxations immediately after dissolving the salt compared to solutions stored in airtight containers for 10 h, while GYY4137 stored for 10 h yielded sulfide accumulations similar to Na2S and markedly increased relaxation. These findings suggest rate of H2S release plays an essential role for the effect on vascular tone of GYY4137.
Plasma L-cysteine concentrations are in the range of 3.5–11 μmol/L (Chawla et al., 1984) and L-cysteine is considered one of the primary substrates leading to formation of H2S. It has at high concentrations been found to increase formation of H2S in mammalian tissues such as kidney (Jackson-Weaver et al., 2013), and to cause relaxations in small cerebral (Streeter et al., 2012) and retinal arteries (Takır et al., 2015), although 1–300 μM L-cysteine had no effect (Takır et al., 2015). In agreement with the latter study, we only observed small relaxations by adding increasing concentrations of L-cysteine to U46619-contracted arteries and no effect on acetylcholine relaxation. In rat mesenteric arteries, the expression of CSE is high in perivascular and adventitial tissue and associated with formation of H2S (Jackson-Weaver et al., 2011; Li et al., 2013). In the present study, we carefully removed adhering tissue and cannot exclude L-cysteine will contribute to endogenous H2S formation to a larger degree in mesenteric arteries with adhering perivascular tissue.
L-cysteine is considered a scavenger of nitroxyl (HNO) (Andrews et al., 2009), and studies in cell cultures reported that L-cysteine by direct interaction may scavenge HS− and lead to formation of inactive sulfides (Miyamoto et al., 2017). By mixing L-cysteine with Na2S or GYY4137, we observed increased accumulation of sulfides (Figure 2). Considering that L-cysteine by itself only had small effect on vascular tone and did not change acetylcholine relaxation, the effect of L-cysteine on Na2S and GYY4137 may be ascribed to a direct chemical reaction, and thereby inactivating Na2S and GYY4137 relaxation.
In previous studies, NaSH and Na2S induced contraction followed by relaxation is observed in the perfused mesenteric vascular bed and in isolated arteries (Ali et al., 2006; Di Villa Bianca et al., 2011; Hedegaard et al., 2016). In the presence of L-cysteine, low concentrations of Na2S and GYY4137 induced marked contractions of the isolated rat mesenteric arteries, and when L-cysteine was mixed with Na2S or GYY4137, we observed an increased sulfide accumulation. Polysulfides (H2S2 and H2S3) have been suggested to play a role in the effects of H2S or to produce many of the effects previously attributed to H2S (Kimura, 2019), but the effects of these unstable compounds were reported to be inhibited in the presence of L-cysteine (Miyamoto et al., 2017), and in previous studies we observed that polysulfides (K2Sn) induce relaxations in rat mesenteric arteries (Hedegaard et al., 2016). Therefore, it seems unlikely that L-cysteine by interaction with Na2S and GYY4137 just leads to formation of inactive sulfides. Instead the conversion of the Na2S and GYY4137 relaxations to contractions by L-cysteine treatment may result from inhibition of polysulfides. However, inhibition of formation of polysulfides with DTT (Miyamoto et al., 2017) did not affect Na2S and GYY4137 relaxation (Figures 4C,D). Therefore, a more speculative mechanism is that L-cysteine together with Na2S or GYY4137 result in the formation of a product that may interfere with endothelial NO synthase leading to contraction, e.g., formation of cIMP instead of cGMP by NO as described in large coronary arteries (Chen et al., 2014) (Figure 9A). Indeed in the presence of L-cysteine, L-NAME by inhibition of NO synthase restored the GYY4137 relaxations. In agreement with these findings we in preliminary studies observed in 1H-, 31P-NMR spectra that L-cysteine incubation with GYY4137 formed a product (results not shown) which support the formation of a product that may interfere with vascular tone in small mesenteric arteries, although other experimental approaches will be required to characterize the product formed by L-cysteine and GYY4137.
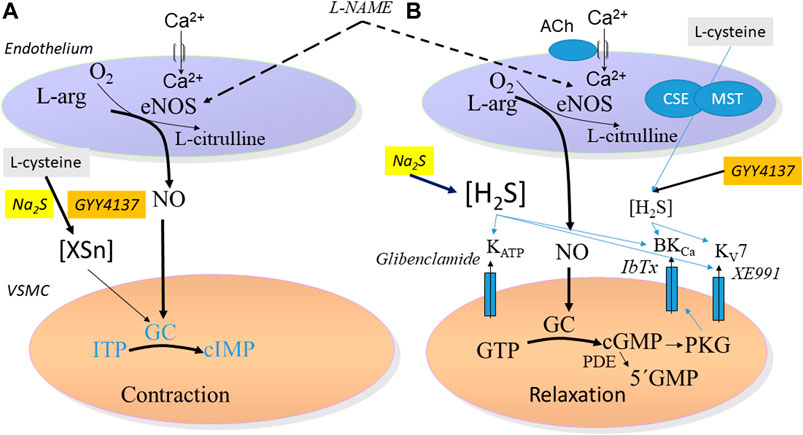
FIGURE 9. Schematic figure proposing the mechanisms involved in (A) contraction in small mesenteric arteries induced by Na2S and GYY4137 in the presence of L-cysteine. This involve a formation of a sulfide product (XSn) that may modulate guanylate cyclase leading to contraction of the vascular smooth muscle cells (VSMC). The effect is partly reversed by the NO synthase inhibitor, L-NAME (B) Relaxation induced by Na2S and GYY4137 leads to, respectively, fast and slow release of H2S and followed by activation of subsets of different potassium channels: ATP-sensitive K (KATP) channels by Na2S, and of large-conductance calcium activated (BKCa) and voltage-gated type 7 (KV7) channels by GYY4137.
While 500 and 1,000 nmol/kg Na2S failed to change blood pressure in normotensive rats (Tomasova et al., 2015), intravenous injection of 39 μmol/kg Na2S decreased mean arterial blood pressure with 45 mmHg in anesthetized mice (Eberhardt et al., 2014). Treatment with 56 μmol/kg/day of NaSH administered by intraperitoneal injection lowered also blood pressure in spontaneously hypertensive rats (Ni et al., 2018). These findings suggest that high doses of H2S salts lowers blood pressure by vasodilatation. As mentioned in the introduction, several mechanisms have been suggested to mediate NaSH and Na2S vasodilatation, depending on the vascular preparations that have been studied. In previous studies in resistance arteries, H2S vasodilatation was found to involve KATP channels (Tang et al., 2005), BKCa channels (Jackson-Weaver et al., 2011; Jackson-Weaver et al., 2013), and KV7 channels (Schleifenbaum et al., 2010; Hedegaard et al., 2016). In patch-clamp studies H2S gas 30 μM to 1 mM caused activation of KATP channels in vascular smooth muscle from mesenteric arteries (Tang et al., 2005), and 10 µM NaSH hyperpolarized mesenteric arteries by iberiotoxin-sensitive mechanism also suggesting the involvement BKCa channels (Jackson-Weaver et al., 2011, 2013), and NaHS (1 mM) hyperpolarized rat aorta and directly activated KV7 channels in CHO cells (Martelli et al., 2013a). Recent studies have also shown that direct activation of KV7 channels by H2S donors protects against neuropathic pain (Di Cesare Mannelli et al., 2017), and that direct persulfidation of KV7 channels by H2S plays an important role in skeletal muscle hypercontractility in human malignant hyperthermia syndrome (Vellecco et al., 2020). In agreement with studies in resistance arteries, in small mesenteric arteries contracted with the U46619, Na2S in the present study induced relaxations sensitive to high extracellular potassium and blockers of both ATP-sensitive, KV7, and BKCa channels suggesting involvement of these channels in Na2S relaxation, although electrophysiological measurements, e.g., membrane potential measurements or patch-clamp will be required to confirm the activation of KV7 channels by Na2S in this preparation.
Interaction of H2S with the NO pathway is thought to be important for the vascular effects of Na2S. Thus, it was proposed that NO and H2S may act co-operatively to generate nitroxyl (HNO), and that this activates transient receptor potential ankyrin 1 (TRPA1) channels on sensory nerves with subsequent calcitonin gene-related peptide release and relaxation in meningeal and mouse mesenteric arteries (Eberhardt et al., 2014). In constrast, based on studies in mice with downregulation of CSE, it was suggested that physiological concentrations of H2S scavenge endothelium-derived NO, and in the absence of NO leads to activation of smooth muscle KATP and KV channels (Szijártó et al., 2018). In the present study, endothelial cell removal or inhibition of NO synthase failed to change relaxations induced by Na2S in mesenteric arteries. These findings agree with our previous studies showing that NaSH relaxation in rat mesenteric arteries is NO and endothelium-independent. However, incubating the preparations with Na2S leftward shifted concentration-response curves for an exogenous NO donor, SNP suggesting that high concentrations of Na2S and NO synergistically cause vasodilatation in rat mesenteric arteries. Moreover, our results support that Na2S causes relaxation by activation of K channels in the smooth muscle layer. High concentrations of Na2S also relax contractions induced by high extracellular potassium suggest that K channel independent mechanisms are involved (Figure 5A), and may similar to NaSH involve inhibition of mitochondrial complex I and III (Hedegaard et al., 2016).
The mechanism of GYY4137 induced vascular relaxation has previously been observed to be endothelium-dependent in rat aorta rings (Li et al., 2008). In small mesenteric arteries, GYY4137 relaxations were reduced in preparations without endothelium, and by an inhibitor of NO synthase suggesting endothelium-derived NO is involved in relaxations induced by GYY4137. Interestingly, incubation with GYY4137 failed to change relaxations induced by SNP suggesting that high H2S concentrations are required to act synergistically with an NO donor, but also implying that the interaction of GYY4137 with endothelium-derived NO is likely at smooth muscle level.
In aorta segments and ciliary arteries KATP channels were found involved in GYY4137 relaxation (Li et al., 2008; Chitnis et al., 2013). Here, we provide evidence that potassium channels may play a pivotal role in the vascular relaxations induced by GYY4137, as high extracellular potassium completely inhibited GYY4137 relaxation. Also, blockers of smooth muscle KV7 and BKCa channels, XE991 and iberiotoxin markedly inhibited relaxation, suggesting these channels are involved in relaxations induced by GYY4137 in rat mesenteric arteries (Figure 9B). Therefore, the mechanisms involved in GYY4137 relaxation regarding both the endothelium and involvement of K channels seems different from the mechanisms involved in Na2S relaxation, reflecting different rate and levels of H2S reaching the vascular preparations when Na2S salt and GYY4137 are added to an organ bath in similar conditions.
Conclusion and Perspectives
The present findings suggest that L-cysteine by reaction with Na2S and GYY4137 and formation of sulfides, inhibits relaxations by these compounds. The low rate of release of H2S species from GYY4137 is reflected by the different sensitivity of these relaxations towards high K+ concentration and K channel blockers compared with Na2S. The perspective is that the rate of release of sulfides plays an important for the effects of H2S salt vs. donors in small arteries.
Data Availability Statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Ethics Statement
The animal study was reviewed and approved by Danish Animal Experiments Inspectorate permission 2014-15-2934-01059.
Author Contributions
Participated in research design: US and MW. Conducted experiments: SA, AP, NR, and JP-D. Contributed new reagents or analytic tools: MW and RT. Performed data analysis: AP, SA, ES, and US. Wrote or contributed to the writing of the manuscript: SA, AP, MW, ES, and US. Manuscript final version approval: All Authors.
Funding
The Danish Medical Research Council (6110-00622B) and the Danish Heart Foundation (grant 17-R116-A7616-22074) supported US. The Medical Research Council, United Kingdom (MR/M022708/1), supported MW, and RT was funded by the Brain Ridge Scholarship, United Kingdom.
Conflict of Interest
MW and the University of Exeter have patents on slow release hydrogen sulfide releasing molecules and their therapeutic use.
The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
The authors thank Heidi Knudsen and Susie Mogensen for technical assistance.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fphar.2021.613989/full#supplementary-material.
References
Alexander, B. E., Coles, S. J., Fox, B. C., Khan, T. F., Maliszewski, J., Perry, A., et al. (2015). Investigating the generation of hydrogen sulfide from the phosphonamidodithioate slow-release donor GYY4137. Med. Chem. Commun. 6, 1649–1655. doi:10.1039/c5md00170f
Ali, M. Y., Ping, C. Y., Mok, Y. Y., Ling, L., Whiteman, M., Bhatia, M., et al. (2006). Regulation of vascular nitric oxide in vitro and in vivo; a new role for endogenous hydrogen sulphide?. Br. J. Pharmacol. 149, 625–634. doi:10.1038/sj.bjp.0706906
Andrews, K. L., Irvine, J. C., Tare, M., Apostolopoulos, J., Favaloro, J. L., Triggle, C. R., et al. (2009). A role for nitroxyl (HNO) as an endothelium-derived relaxing and hyperpolarizing factor in resistance arteries. Br. J. Pharmacol. 157, 540–550. doi:10.1111/j.1476-5381.2009.00150.x
Benavides, G. A., Squadrito, G. L., Mills, R. W., Patel, H. D., Isbell, T. S., Patel, R. P., et al. (2007). Hydrogen sulfide mediates the vasoactivity of garlic. Proc. Natl. Acad. Sci. USA 104, 17977–17982. doi:10.1073/pnas.0705710104
Chawla, R. K., Lewis, F. W., Kutner, M. H., Bate, D. M., Roy, R. G., and Rudman, D. (1984). Plasma cysteine, cystine, and glutathione in cirrhosis. Gastroenterology 87, 770–776. doi:10.1016/0016-5085(84)90069-6
Cheang, W. S., Wong, W. T., Shen, B., Lau, C. W., Tian, X. Y., Tsang, S. Y., et al. (2010). 4-Aminopyridine-sensitive K+ channels contributes to NaHS-induced membrane hyperpolarization and relaxation in the rat coronary artery. Vascul. Pharmacol. 53, 94–98. doi:10.1016/j.vph.2010.04.004
Chen, Z., Zhang, X., Ying, L., Dou, D., Li, Y., Bai, Y., et al. (2014). cIMP synthesized by sGC as a mediator of hypoxic contraction of coronary arteries. Am. J. Physiol. Heart Circ. Physiol. 307, H328–H336. doi:10.1152/ajpheart.00132.2014
Chitnis, M. K., Njie-Mbye, Y. F., Opere, C. A., Wood, M. E., Whiteman, M., and Ohia, S. E. (2013). Pharmacological actions of the slow release hydrogen sulfide donor GYY4137 on phenylephrine-induced tone in isolated bovine ciliary artery. Exp. Eye Res. 116, 350–354. doi:10.1016/j.exer.2013.10.004
Di Villa Bianca, R. E., Sorrentino, R., Coletta, C., Mitidieri, E., Rossi, A., Vellecco, V., et al. (2011). Hydrogen sulfide-induced dual vascular effect involves arachidonic acid cascade in rat mesenteric arterial bed. J. Pharmacol. Exp. Ther. 337, 59–64. doi:10.1124/jpet.110.176016
Di Cesare Mannelli, L., Lucarini, E., Micheli, L., Mosca, I., Ambrosino, P., Soldovieri, M. V., et al. (2017). Effects of natural and synthetic isothiocyanate-based H2S-releasers against chemotherapy-induced neuropathic pain: role of Kv7 potassium channels. Neuropharmacology 121, 49–59. doi:10.1016/j.neuropharm.2017.04.029
Eberhardt, M., Dux, M., Namer, B., Miljkovic, J., Cordasic, N., Will, C., et al. (2014). H2S and NO cooperatively regulate vascular tone by activating a neuroendocrine HNO-TRPA1-CGRP signalling pathway. Nat. Commun. 5, 4381–4417. doi:10.1038/ncomms5381
Esechie, A., Kiss, L., Olah, G., Horváth, E. M., Hawkins, H., Szabo, C., et al. (2008). Protective effect of hydrogen sulfide in a murine model of acute lung injury induced by combined burn and smoke inhalation. Clin. Sci. 115, 91–97. doi:10.1042/CS20080021
Fenger-Gron, J., Mulvany, M. J., and Christensen, K. L. (1995). Mesenteric blood pressure profile of conscious, freely moving rats. J. Physiol. 488, 753. doi:10.1113/jphysiol.1995.sp021006
Feng, W., Teo, X. Y., Novera, W., Ramanujulu, P. M., Liang, D., Huang, D., et al. (2015). Discovery of new H 2 S releasing phosphordithioates and 2,3-Dihydro-2-phenyl-2-sulfanylenebenzo[d][1,3,2]oxazaphospholes with Improved Antiproliferative activity. J. Med. Chem. 58, 6456–6480. doi:10.1021/acs.jmedchem.5b00848
Giangiacomo, K. M., Garcia, M. L., and McManus, O. B. (1992). Mechanism of iberiotoxin block of the large-conductance calcium-activated potassium channel from bovine aortic smooth muscle. Biochemistry 31, 6719. doi:10.1021/bi00144a011
Gheibi, S., Jeddi, S., Kashfi, K., and Ghasemi, A. (2018). Regulation of vascular tone homeostasis by NO and H2S: Implications in hypertension. Biochem. Pharmacol. 149, 42–59. doi:10.1016/j.bcp.2018.01.017
Hedegaard, E. R., Gouliaev, A., Winther, A. K., Arcanjo, D. D., Aalling, M., Renaltan, N. S., et al. (2016). Involvement of potassium channels and calcium-independent mechanisms in hydrogen sulfide-induced relaxation of rat mesenteric small arteries. J. Pharmacol. Exp. Ther. 356, 53–63. doi:10.1124/jpet.115.227017
Hedegaard, E. R., Nielsen, B. D., Kun, A., Hughes, A. D., Krøigaard, C., Mogensen, S., et al. (2014). KV 7 channels are involved in hypoxia-induced vasodilatation of porcine coronary arteries. Br. J. Pharmacol. 171, 69–82. doi:10.1111/bph.12424
Jackson-Weaver, O., Paredes, D. A., Gonzalez Bosc, L. V., Walker, B. R., and Kanagy, N. L. (2011). Intermittent hypoxia in rats increases myogenic tone through loss of hydrogen sulfide activation of large-conductance Ca(2+)-activated potassium channels. Circ. Res. 108, 1439–1447. doi:10.1161/CIRCRESAHA.110.228999
Jackson-Weaver, O., Osmond, J. M., Riddle, M. A., Naik, J. S., Bosc, L. V. G., Walker, B. R., et al. (2013). Hydrogen sulfide dilates rat mesenteric arteries by activating endothelial large-conductance Ca2+-activated K+ channels and smooth muscle Ca2+ sparks. Am. J. Physiology-Heart Circulatory Physiol. 304, H1446–H1454. doi:10.1152/ajpheart.00506.2012
Kimura, H. (2019). Signaling by hydrogen sulfide (H2S) and polysulfides (H2Sn) in the central nervous system. Neurochem. Int. 126, 118–125. doi:10.1016/j.neuint.2019.01.027
Koike, S., Nishimoto, S., and Ogasawara, Y. (2017). Cysteine persulfides and polysulfides produced by exchange reactions with H2S protect SH-SY5Y cells from methylglyoxal-induced toxicity through Nrf2 activation. Redox Biol. 12, 530–539. doi:10.1016/j.redox.2017.03.020
Kubo, S., Doe, I., Kurokawa, Y., Nishikawa, H., and Kawabata, A. (2007). Direct inhibition of endothelial nitric oxide synthase by hydrogen sulfide: contribution to dual modulation of vascular tension. Toxicology 232, 138–146. doi:10.1016/j.tox.2006.12.023
Lee, S. W., Hu, Y. S., Hu, L. F., Lu, Q., Dawe, G. S., and Moore, P. K. (2006). Hydrogen sulphide regulates calcium homeostasis in microglial cells. Glia. 54, 116–124. doi:10.1002/glia.20362
Lee, Z. W., Zhou, J., Chen, C. S., Zhao, Y., Tan, C. H., Li, L., et al. (2011). The slow-releasing hydrogen sulfide donor, GYY4137, exhibits novel anti-cancer effects in vitro and in vivo. PLoS One 6, e21077. doi:10.1371/journal.pone.0021077
Li, L., Whiteman, M., Guan, Y. Y., Neo, K. L., Cheng, Y., Lee, S. W., et al. (2008). Characterization of a novel, water-soluble hydrogen sulfide-releasing molecule (GYY4137): new insights into the biology of hydrogen sulfide. Circulation 117, 2351–2360. doi:10.1161/CIRCULATIONAHA.107.753467
Li, R., Andersen, I., Aleke, J., Golubinskaya, V., Gustafsson, H., and Nilsson, H. (2013). Reduced anti-contractile effect of perivascular adipose tissue on mesenteric small arteries from spontaneously hypertensive rats: role of Kv7 channels. Eur. J. Pharmacol. 698, 310–315. doi:10.1016/j.ejphar.2012.09.026
Li, S., Ping, N. N., Cao, L., Mi, Y. N., and Cao, Y. X. (2015). H2S induces vasoconstriction of rat cerebral arteries via cAMP/adenylyl cyclase pathway. Toxicol. Appl. Pharmacol. 289, 389–396. doi:10.1016/j.taap.2015.10.021
Liu, Z., Han, Y., Li, L., Lu, H., Meng, G., Li, X., et al. (2013). The hydrogen sulfide donor, GYY4137, exhibits anti‐atherosclerotic activity in high fat fed apolipoprotein E‐/‐ mice. Br. J. Pharmacol. 169, 1795–1809. doi:10.1111/bph.12246
Malekova, L., Krizanova, O., and Ondrias, K. (2009). H(2)S and HS(‐) donor NaHS inhibits intracellular chloride channels. Gen. Physiol. Biophys. 28, 190–194. doi:10.4149/gpb_2009_02_190
Martelli, A., Testai, L., Breschi, M. C., Lawson, K., McKay, N. G., Miceli, F., et al. (2013a). Vasorelaxation by hydrogen sulphide involves activation of Kv7 potassium channels. Pharmacol. Res. 70, 27. doi:10.1016/j.phrs.2012.12.005
Martelli, A., Testai, L., Citi, V., Marino, A., Pugliesi, I., Barresi, E., et al. (2013b). Arylthioamides as H2S donors: l-cysteine-activated releasing properties and vascular effects in vitro and in vivo. ACS Med. Chem. Lett. 4, 904–908. doi:10.1021/ml400239a
McGrath, J. C., and Lilley, E. (2015). Implementing guidelines on reporting research using animals (ARRIVE etc.): New requirements for publication in BJP. Br. J. Pharmacol. 172, 3189–3193. doi:10.1111/bph.12955
Miyamoto, R., Koike, S., Takano, Y., Shibuya, N., Kimura, Y., Hanaoka, K., et al. (2017). Polysulfides (H2Sn) produced from the interaction of hydrogen sulfide (H2S) and nitric oxide (NO) activate TRPA1 channels. Sci. Rep. 7. doi:10.1038/srep4599510.1038/srep45995
Mulvany, M. J., and Halpern, W. (1976). Mechanical properties of vascular smooth muscle cells in situ. Nature 260, 617. doi:10.1038/260617a0
Mulvany, M. J., Videbaek, L. M., Hughes, A. D., and Aalkjaer, C. (1990). Mechanism of the vasodilator action of pinacidil. Blood Vessels 27, 314. doi:10.1159/000158824
Ni, X., Zhang, L., Peng, M., Shen, T. W., Yu, X. S., Shan, L. Y., et al. (2018). Hydrogen sulfide attenuates hypertensive inflammation via regulating connexin expression in spontaneously hypertensive rats. Med. Sci. Monit. 24, 1205–1218. doi:10.12659/msm.908761
Perniss, A., Preiss, K., Nier, M., and Althaus, M. (2017). Hydrogen sulfide stimulates CFTR in Xenopus oocytes by activation of the cAMP/PKA signalling axis. Sci. Rep. 7, 3517. doi:10.1038/s41598-017-03742-5
Szabo, C. (2017). Hydrogen sulfide, an enhancer of vascular nitric oxide signaling: mechanisms and implications. Am. J. Physiol. Cell Physiol. 312, C3–C15. doi:10.1152/ajpcell.00282.2016
Schleifenbaum, J., Köhn, C., Voblova, N., Dubrovska, G., Zavarirskaya, O., Gloe, T., et al. (2010). Systemic peripheral artery relaxation by KCNQ channel openers and hydrogen sulfide. J. Hypertens. 28, 1875. doi:10.1097/HJH.0b013e32833c20d5
Skovgaard, N., Gouliaev, A., Aalling, M., and Simonsen, U. (2011). The role of endogenous H2S in cardiovascular physiology. Curr. Pharm. Biotechnol. 12, 1385–1393. doi:10.2174/138920111798280956
Steiger, A. K., Yang, Y., Royzen, M., and Pluth, M. D. (2017). Bio-orthogonal “click-and-release” donation of caged carbonyl sulfide (COS) and hydrogen sulfide (H2S). Chem. Commun. (Camb). 53, 1378–1380. doi:10.1039/c6cc09547j
Streeter, E., Hart, J., and Badoer, E. (2012). An investigation of the mechanisms of hydrogen sulfide-induced vasorelaxation in rat middle cerebral arteries. Naunyn. Schmiedebergs. Arch. Pharmacol. 385, 991–1002. doi:10.1007/s00210-012-0779-2
Szabó, C. (2007). Hydrogen sulphide and its therapeutic potential. Nat. Rev. Drug Discov. 6, 917–935. doi:10.1038/nrd2425
Szabo, C., and Papapetropoulos, A. (2017). International union of basic and clinical Pharmacology. CII: pharmacological modulation of H2S levels: H2S donors and H2S biosynthesis inhibitors. Pharmacol. Rev. 69, 497–564. doi:10.1124/pr.117.014050
Szijártó, I. A., Markó, L., Filipovic, M. R., Miljkovic, J. L., Tabeling, C., Tsvetkov, D., et al. (2018). Cystathionine γ-lyase-produced hydrogen sulfide controls endothelial NO bioavailability and blood pressure. Hypertension 71, 1210–1217. doi:10.1161/HYPERTENSIONAHA.117.10562
Takır, S., Ortaköylü, G. Z., Toprak, A., and Uydeş-Doğan, B. S. (2015). NaHS induces relaxation response in prostaglandin F(2α) precontracted bovine retinal arteries partially via K(v) and K(ir) channels. Exp. Eye Res. 132, 190–197. doi:10.1016/j.exer.2015.02.002
Tang, G., Wu, L., Liang, W., and Wang, R. (2005). Direct stimulation of K(ATP) channels by exogenous and endogenous hydrogen sulfide in vascular smooth muscle cells. Mol. Pharmacol. 68, 1757–1764. doi:10.1124/mol.105.017467
Tomasova, L., Pavlovicova, M., Malekova, L., Misak, A., Kristek, F., Grman, M., et al. (2015). Effects of AP39, a novel triphenylphosphonium derivatised anethole dithiolethione hydrogen sulfide donor, on rat haemodynamic parameters and chloride and calcium Cav3 and RyR2 channels. Nitric Oxide 46, 131–144. doi:10.1016/j.niox.2014.12.012
Vellecco, V., Martelli, A., Bibli, I. S., Vallifuoco, M., Manzo, O. L., Panza, E., et al. (2020). Anomalous Kv 7 channel activity in human malignant hyperthermia syndrome unmasks a key role for H2 S and persulfidation in skeletal muscle. Br. J. Pharmacol. 177, 810–823. doi:10.1111/bph.14700
Wallace, J. L., Vaughan, D., Dicay, M., MacNaughton, W.K, and de Nucci, G. (2018). Hydrogen sulfide-releasing therapeutics: translation to the clinic. Antioxid. Redox Signal. 28, 1533–1540. doi:10.1089/ars.2017.7068
Wang, R. (2012). Physiological implications of hydrogen sulfide: a whiff exploration that blossomed. Physiol. Rev. 92, 791–896. doi:10.1152/physrev.00017.2011
Webb, G. D., Lim, L. H., Oh, V. M., Yeo, S. B., Cheong, Y. P., Ali, M. Y., et al. (2008). Contractile and vasorelaxant effects of hydrogen sulfide and its biosynthesis in the human internal mammary artery. J. Pharmacol. Exp. Ther. 324, 876–882. doi:10.1124/jpet.107.133538
Whiteman, M., Li, L., Kostetski, I., Chu, S. H., Siau, J. L., Bhatia, M., et al. (2006). Evidence for the formation of a novel nitrosothiol from the gaseous mediators nitric oxide and hydrogen sulphide. Biochem. Biophys. Res. Commun. 343, 303. doi:10.1016/j.bbrc.2006.02.154
Whiteman, M., Li, L., Rose, P., Tan, C. H., Parkinson, D. B., and Moore, P. K. (2010). The effect of hydrogen sulfide donors on lipopolysaccharide-induced formation of inflammatory mediators in macrophages. Antioxid. Redox Signal 12, 1147. doi:10.1089/ars.2009.2899
Yeung, S. Y., Pucovský, V., Moffatt, J. D., Saldanha, L., Schwake, M., Ohya, S., et al. (2007). Molecular expression and pharmacological identification of a role for K(v)7 channels in murine vascular reactivity. Br. J. Pharmacol. 151, 758–770. doi:10.1038/sj.bjp.0707284
Zhao, W., and Wang, R. (2002). H(2)S-induced vasorelaxation and underlying cellular and molecular mechanisms. Am. J. Physiol. Heart Circ. Physiol. 283, H474–H480. doi:10.1152/ajpheart.00013.2002
Keywords: GYY4137, sodium sulfide, hydrogen sulfide, potassium channels, small mesenteric arteries
Citation: Abramavicius S, Petersen AG, Renaltan NS, Prat-Duran J, Torregrossa R, Stankevicius E, Whiteman M and Simonsen U (2021) GYY4137 and Sodium Hydrogen Sulfide Relaxations Are Inhibited by L-Cysteine and KV7 Channel Blockers in Rat Small Mesenteric Arteries. Front. Pharmacol. 12:613989. doi: 10.3389/fphar.2021.613989
Received: 04 October 2020; Accepted: 12 February 2021;
Published: 26 March 2021.
Edited by:
Lei Xi, Virginia Commonwealth University, United StatesReviewed by:
Takayuki Matsumoto, Hoshi University, JapanVincenzo Calderone, University of Pisa, Italy
Copyright © 2021 Abramavicius, Petersen, Renaltan, Prat-Duran, Torregrossa, Stankevicius, Whiteman and Simonsen. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Ulf Simonsen, dXNAYmlvbWVkLmF1LmRr