Abstract
Ion channels play an important role in mediating pain through signal transduction, regulation, and control of responses, particularly in neuropathic pain. Transient receptor potential channel superfamily plays an important role in cation permeability and cellular signaling. Transient receptor potential channel Melastatin 2 (TRPM2) subfamily regulates Ca2+ concentration in response to various chemicals and signals from the surrounding environment. TRPM2 has a role in several physiological functions such as cellular osmosis, temperature sensing, cellular proliferation, as well as the manifestation of many disease processes such as pain process, cancer, apoptosis, endothelial dysfunction, angiogenesis, renal and lung fibrosis, and cerebral ischemic stroke. Toll-like Receptor 4 (TLR4) is a critical initiator of the immune response to inflammatory stimuli, particularly those triggered by Lipopolysaccharide (LPS). It activates downstream pathways leading to the production of oxidative molecules and inflammatory cytokines, which are modulated by basal and store-operated calcium ion signaling. The cytokine production and release cause an imbalance of antioxidant enzymes and redox potential in the Endoplasmic Reticulum and mitochondria due to oxidative stress, which results from TLR-4 activation and consequently induces the production of inflammatory cytokines in neuronal cells, exacerbating the pain process. Very few studies have reported the role of TRPM2 and its association with Toll-like receptors in the context of neuropathic pain. However, the molecular mechanism underlying the interaction between TRPM2 and TLR-4 and the quantum of impact in acute and chronic neuropathic pain remains unclear. Understanding the link between TLR-4 and TRPM2 will provide more insights into pain regulation mechanisms for the development of new therapeutic molecules to address neuropathic pain.
1 Introduction
Pain is defined as a characteristic phenomenon caused by noxious stimuli from pathogenic microorganisms such as bacteria, viruses, and fungi and from injury caused by various forms of mechanical and thermal stimuli. The physiological response to these conditions results in pain. Relief from pain is crucial for an organism’s survival and protection because pain serves as a warning signal for potential harm or injury. It triggers reflexes and behaviors that help avoid or minimize damage, promoting healing and preventing further injury. Without effective pain relief mechanisms, an organism could suffer prolonged stress, which can lead to immune suppression, impaired function, and reduced chances of survival. Therefore, managing pain is not only important for comfort but also vital for the overall wellbeing and preservation of life. Normal pain symptoms involve allodynia, which is non-painful stimuli, such as a light touch or mild temperature change, whereas hyperalgesia refers to an increased sensitivity to painful stimuli generated during minor injury. The symptoms of hypersensitivity involve both allodynia and hyperalgesia, which are intense responses to sensory stimuli, resulting in enhanced sensitivity and discomfort. Persistent and extreme sensitivity are also referred to as chronic pain (Loeser and Melzack, 1999; Świeboda et al., 2013). Acute pain condition is a protective mechanism in which tissue stimulates the healing process. Beyond any alteration in the pain pathway and duration leads to hypersensitivity and intolerable or chronic pain.
Depending on the duration of symptoms, pain is classified as acute pain (<3 months) and chronic pain (>3 months). Pain is also classified into many subtypes such as anatomical pain, physiological pain, pathological pain, deep pain, vascular pain, bone and joint pain, myalgia, organ pain, wired pain, neuralgia, causalgia, convolutional pain, and phantom pain, depending upon the stimuli, location, physiological and social factors (Świeboda et al., 2013; Sela et al., 2002).
According to the International Association for the Study of Pain defines, neuropathic pain is a pain caused by a lesion or disease affecting the somatosensory nervous system. As per the International classification of diseases, conditions for peripheral neuropathic pain include trigeminal neuralgia, polyneuropathy, peripheral nerve injury, and painful radiculopathy. Conditions for central neuropathy include pain caused by brain or spinal cord injury, post-stroke pain, and pain associated with multiple sclerosis (Scholz et al., 2019). Neuropathic pain is associated with damage to the nerves, including central, peripheral, and somatosensory nervous systems. It involves both positive and negative symptoms. Positive symptoms include paresthesia, dysesthesia, superficial pain, allodynia, and hyperalgesia. Negative symptoms include tactile hypoesthesia, thermal hypoesthesia, and pinprick hypoalgesia (Baron, 2006). Neuropathic pain is considered chronic pain that affects the quality of normal life of an individual. Trigeminal neuralgia, diabetic neuropathic pain, cancer pain, and sciatica are some examples of chronic pain. Many of the neuronal complex structures are involved in the activation, transmission, regulation, and healing processes (Backonja, 2003). Nerve endings, dorsal root ganglia, microglia, Schwann cells, and thalamus are the important structures that are involved in chronic pain. Structural changes, numerous cellular interactions, and signaling processes contribute to the nociceptive pathway, including ion channels, receptors, immune cells, and microglia (Finnerup et al., 2020; Kerstman et al., 2013). The transition from acute neuropathic pain to chronic pain is a complex process and involves multiple signaling pathways, central and peripheral processes, and inherited and environmental risks.
Neuroinflammation is an immune-mediated inflammatory response elicited by various stimuli. Cytokines released in response to the inflammation regulate oxidative stress and mediate the injury and/or repair process (Myers et al., 2006). Sensitization of nerve endings and other inflammatory molecules mediate the pain process in which ion channels play an important role in mediating pain, inflammation, and repair depending on Ca2+ ion influx (Kuffler, 2020). Many of the ion channels are regulated either by a change in the voltage (cation/anion exchange) or receptor-mediated responses. These ion channels respond to changes in the cellular external and internal environments, thereby eliciting a physiological response (Bouali-Benazzouz et al., 2021; Vicario et al., 2020). Ion channels exhibit regulatory properties of immune cells, metabolism, and inflammation pathways (Ruck et al., 2022; Bittner et al., 2014). Apart from the homeostatic functions, ion channels may also be involved in the pathological mechanisms of neuronal inflammation that leads to pain. Membrane ion channels that govern sodium, potassium, and calcium ions and their modulation by various inflammatory mediators are involved in neuropathic pain.
Inflammatory responses and oxidative stress are regulated by molecular inhibitors and activators, Toll-like-receptor (TLR-4) is a transmembrane pattern recognition receptor (Fehder et al., 2007), Nicotinamide Adenine Dinucleotide phosphate (NADPH) oxidase (NOX), and Transient receptor potential (TRP), an ion channel and a well-known receptor for immune responses. TLR-4 is associated with many pathological conditions with chronic neuropathic pain. TRPM2 is a calcium-permeable cation channel in sensory neurons. NOX is part of a family of enzymes that includes NOX1, NOX2, NOX3, NOX4, NOX5, DUOX1, and DUOX2. NOX enzymes are known for immune defenses in which Reactive Oxygen Species (ROS) production by phagocytic cells to kill pathogens (Vermot et al., 2021). TRPM2 channels are activated by ROS produced by NOX, which leads to calcium influx. This calcium signaling boosts NOX activity, which amplifies ROS production and oxidative stress. This pathway is associated with several diseases, including neurodegenerative diseases and chronic pain (Görlach et al., 2015).
The present review aims to address pathways in which oxidative stress and inflammatory reactions by TLR-4, NOX, and TRPM2 implicate pain (Figure 1). These findings highlight the biological importance of TRPM2 and TLR-4 for elucidating pain pathways due to producing ROS and inflammatory cytokines, which are important in regulating oxidative stress and inflammation in cells. TRPM2 inhibitors could decrease oxidative stress-induced injuries and neuropathic pain in a wide variety of ways. TLR-4 antagonists may decrease inflammatory responses and alleviate chronic inflammation. The discovery of the mechanisms by which TRPM2 activation drives nerve damage and inflammation, and its association with TLR-4 can show new pathways; such understanding may become apparent as an innovative therapy for disorders assumed to cause or intensify neuropathic pain.
FIGURE 1
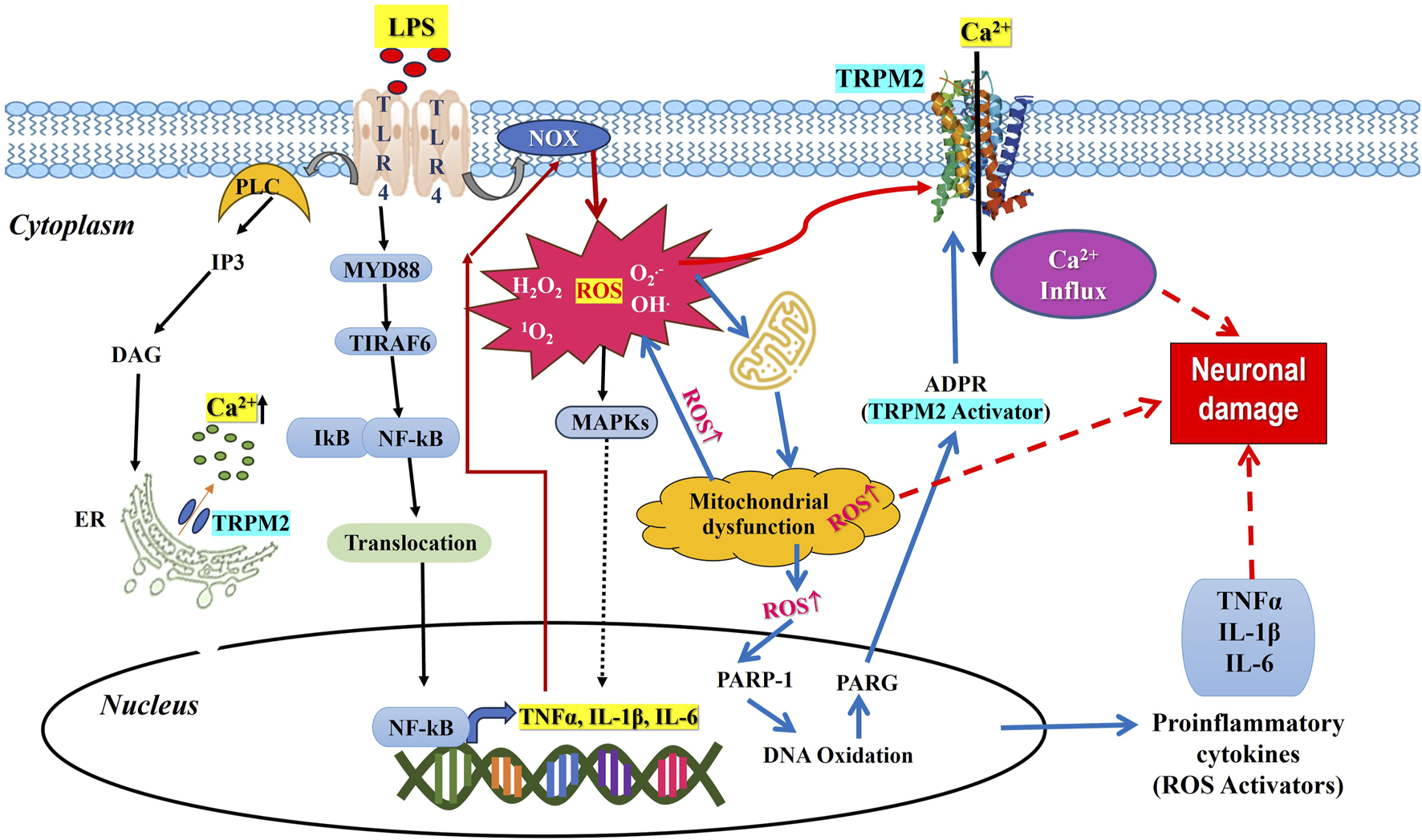
Mechanism of TLR4 induced TRPM2 mediated neuropathic pain.
2 Ion channels and their role in diseases
The process of recognizing pain involves the recognition of pain signals by sensory neurons, initiation of nerve impulses, and transforming the noxious stimuli to pain response. These nociceptive neurons are specialized in the recognition of noxious stimuli, of which ion channels on cell membrane play an important role in intermembrane transporters of various ions, thereby regulating various physiological functions ranging from excitation of tissue transportation, cell migration tumorigenesis, atherosclerosis, and apoptosis (Kew and Davies, 2010; Schwab et al., 2012; Jentsch, 2016). Voltage-gated ion channels are widely expressed in all tissues, including neurons, such as sodium Nav1.8 and Nav1.9 channels (Giniatullin, 2020). Other ion channels such as purinergic receptor, TRPV1(vanilloid)/A1 (ankyrin), TRPC (canonical), TRPM2, and voltage-gated (sodium, calcium, and potassium) channels are also involved in nociception. Many of the neuronal diseases, including neuropathic pain, are linked to ion channel dysfunction and neuroinflammation (Eren-Koçak and Dalkara, 2021).
3 TRP channel
Transient receptor potential (TRP) channels are widely distributed throughout the tissues. In 1969, TRP was discovered in drosophila and subsequently in humans (Montell and Rubin, 1989; Samanta et al., 2018). The TRP channels superfamily were extended up to 48 members and their major role was found in cation permeability and cellular sensory signaling. These channel proteins span the transmembrane with a C-terminal intracellular domain and N-terminal intracellular domain (Kassmann et al., 2013), which are non-selective ion channels that regulate Ca2+, Mg2+ influx, and other monovalent cations. These TRP channels can be activated in cellular organelles, such as endoplasmic reticulum, lysosomes, Golgi apparatus, and synaptic vesicles by Ca2+ influx. TRP channels are also involved in the regulation of osmotic pressure, temperature, pH oxidation-reduction vascular tone, proliferation, mechanosensing, proliferation, apoptosis, and angiogenesis (Gees et al., 2010). Therefore, TRP channels respond to various stimuli such as stress, mediators, hormones, chemicals, and environmental factors (Lolignier et al., 2016; Kaneko and Szallasi, 2014). Changes in the channel function result in many disorders, such as retinal, skin, cardiac, neuronal, and vascular, forming the basis of mutation in the genes responsible for coding for TRP channel (Tsagareli and Nozadze, 2020). TRP Channels have been implicated in various neurodegenerative diseases such as Alzheimer’s, Parkinson’s, and Huntington’s diseases (Rather et al., 2023). TRP channels are proven to play as central elements of nociception, thereby acting as potential targets for alleviating neuropathic pain. Modulating the TRP channel function offers a wide scope for altering cellular functions, thereby increasing the research interest. Modulation of TRP channels has emerged as a novel field of therapeutic strategy for the treatment of pain, especially neuropathic pain. Many small molecule compounds that modulate TRP channels have entered clinical trials aiming to treat a variety of neuronal diseases (Table 1).
TABLE 1
| Agent | Antagonists/Activators | Indications | Status | Notes | Ref. |
|---|---|---|---|---|---|
| Gabapentin | TRPV1 antagonist (Indirect) | Neuropathic pain, epilepsy | Approved | Gabapentin combined with Capsaicin enters the neurons through TRPV1 to access Voltage-gated channels | Biggs et al. (2015) |
| Pregabalin | TRPV1 antagonist (Indirect) | Neuropathic pain, fibromyalgia, epilepsy | Approved | Pregabalin is used for its antagonistic activity at Voltage-gated Ca2+ channels and binds to alpha-2-delta | Onakpoya et al. (2019) |
| Capsaicin | TRPV1 activator | Neuropathic pain | Approved | Capsaicin desensitizes, calcium-permeable TRPV1 channels and relieves pain | Pasierski and Szulczyk (2022) |
| SB-705498 | TRPV1 antagonist | Rhinitis Pain relief | Clinical Phase II |
SB-705498 relieved pain by suppressing response to skin stimulation by TRPV1 receptor | Lambert et al. (2009) |
| Resiniferatoxin | TRPV1 activator | osteoarthritic pain | Clinical Phase II |
Resiniferatoxin through injection desensitizes TRPV1 | Szallasi (2023) |
| Tramadol | TRPV1 antagonist (Indirect) | neuropathic pain | Approved | Tramadol relieves Neuropathic Pain by regulating mediators in pain signaling (TRPV1) | Quan et al. (2022) |
| XEN-D0501 | TRPV1 receptor antagonist | Chronic pain | Clinical Phase II |
XEN-D0501 is used as a temperature regulator by blocking the activation of TRPV1 receptor | Round et al. (2011) |
| Topiramate | TRPV1 antagonist (Indirect) | Migraine | Approved | Topiramate is used as anti-migraine, activates Na+ channels and Ca2+ channels | Fan et al. (2018) |
| Zonisamide | TRPV1 antagonist (Indirect) | Migraine | Clinical Phase II |
Zonisamide is used a anti-migraine, activates Na+ channels and Ca2+ channel | Asiri and Hassan (2023) |
| GRC 6211 | TRPV1 antagonists | Osteoarthritis pain, chronic pain | Clinical Phase II |
GRC 6211 suppresses neurogenic detrusor overactivity | Santos-Silva et al. (2012) |
| PF-05105679 | TRPM8 antagonists | Cold Pain Sensitivity | Clinical Phase II |
PF-05105679 is used to regulate body temperature | Gosset et al. (2017) |
| AMG-333 | TRPM8 antagonists | Migraine | Clinical Phase II |
TRPM8 is activated. By cold and AMG-333 is a highly selective antagonist | Horne et al. (2018) |
| KRP-2529 | TRPM8 antagonists | hypersensitive bladder disorders | Preclinical | TRPM8 antagonist reduces hypersensitivity by blocking TRPM8 activity | Aizawa et al. (2018) |
| RQ-00434739 | TRPM8 antagonists | hypersensitive bladder disorders | Preclinical | TRPM8 antagonist reduces hypersensitivity by blocking TRPM8 activity | Aizawa and Fujita (2022) |
| HC-030031 | TRPA1 antagonists | Chronic Pain | Preclinical | HC-030031 reduces pain and inflammation by blocking TRPA1 channels | de Almeida et al. (2021) |
| GRC-17536 | TRPA1 antagonists | Hypersensitivity | Clinical Phase II |
TRPA1 antagonists inhibit the activation of the channel that detects pain | Klionsky et al. (2007) |
| AMG-9090 | TRPA1 antagonists | Chronic Pain | Preclinical | TRPA1 antagonist reduces hypersensitivity by blocking TRPA1 activity | Klionsky et al. (2007) |
| LY3526318 | TRPA1 antagonists | Neuropathic Pain | Clinical Phase II |
LY3526318 alleviates pain by inhibiting TRPA1 channel activity | Bamps et al. (2023) |
Present and ongoing TRP ion channels clinical drugs used for pain pathology.
4 TRPM channel structure and functions
TRPM channels are ubiquitous in nature and widely expressed and distributed in tissues and contribute to health and disease. The three domains of these ion channels are N, C, and channel domain, where the N-terminal is associated with four similar melastatin subunits and a pre-S1 domain to play a role in sensing and channel assembly. The channel domain S4 is specific to the TRPM family and participates in a voltage-sensing-like domain, whereas S5-S6 forms the P-loop, which mainly acts as an ion-conducting pore. The C-Domain is composed of a TRP box, which is highly conserved and maintains channel stability, whereas the coiled-coil domain contains motifs that modulate pore gating. The differences in the homology sequence of the C-terminus divide the TRPM subfamily into four groups: TRPM3/TRPM6/TRPM7, highly permeable to Ca2+, TRPM2/TRPM8 nonselective for monovalent and divalent cations, whereas TRPM4/TRPM5 is permeable to monovalent cations. TRPM2/TRPM6/TRPM7 present enzymatic properties at the C terminus, and TRPM2 has an additional domain for nucleoside diphosphate pyrophosphatase, which shares a similar homology with Nudix hydrolase NUDT (-H domain), which exhibit a wide range of functions such as metabolism of nicotinamide adenine dinucleotide (NAD), ADP-ribose, and their derivatives and is characterized by different substrate specificity and intracellular localization. (Samanta et al., 2018; McLennan, 2006). TRPM8 is temperature sensitive and activated by innocuous cold to noxious cold. It is also activated by menthol and icilin, synthetic super-agonist. Both TRPM8 agonists and antagonists, e.g., AMTB, proved to be beneficial in pain pathways (Fernández-Peña and Viana, 2013). TRPM3 is a polymodal nociceptor thought to be involved in the detection of noxious heat (Vriens et al., 2011). TRPM3, together with TRPV1 and TRPA1, mediate acute noxious heat detection in mice (Vandewauw et al., 2018). Given the importance of TRPM channels, there is an increase in research interest in TRPM channels and their role in triggering various diseases, including pain process, cellular osmosis, temperature sensing, cellular proliferation, cancer, apoptosis, endothelial dysfunction, angioparagraphgenesis, renal, and lung fibrosis (Duitama et al., 2020).
5 TRPM2 distribution and its role in various pathologies
TRPM2 is particularly gaining importance as it is permeable to both monovalent and divalent cations, as well as its activation/inhibition in response to various stimuli from both exogenous and endogenous sources, leading to changes in cellular responses that vary from picoseconds to several seconds. This is the only ion channel of the TRPM family associated with enzyme activity at NUDT9-H domain of the C-terminus sensitive to adenosine diphosphate ribose (ADPR) (Sumoza-Toledo and Penner, 2011; Perraud et al., 2001) thereby involving multiple functional roles like Ca2+ homeostasis (Hasan and Zhang, 2018), redox potential (Song et al., 2016), osmotic regulation, temperature sensitive gating (Vilar et al., 2020; Tan and McNaughton, 2018), production of inflammatory mediators (Yamamoto and Shimizu, 2016; Akyuva and Nazıroğlu, 2020), pain modulation (Di et al., 2012; Yüksel et al., 2017), cellular migration, cytoskeleton remodeling (Schwab et al., 2012), and immunity (Knowles et al., 2013; Syed Mortadza et al., 2015). Functional role of TRPM2 have been reported in many diseases such as Alzheimer’s and Parkinson’s disease, ischemic stroke, neuronal cell death, neurovascular functional injury, myocardial ischemia/reperfusion injury, vascular dysfunction, pancreatic β-cell death associated with pancreatitis, acute and chronic diseases, and liver toxicity (Malko and Jiang, 2020). TRPM2 channel is widely expressed in immune cells, microglia, neutrophils, T-lymphocytes. It also acts as a sensor for ROS and plays a key role in the inflammatory response in both normal and pathological states. Since TRPM2 has multifunctional role in oxidative stress and inflammation it would be worth exploring its mechanisms in attenuating pain-related diseases.
5.1 TRPM2 in cerebral ischemic stroke
TRPM2 is widely expressed in neurons and its role in ischemic stroke was evaluated in several in vitro and in vivo studies. TRPM2 have shown to increase the cell survival and inhibit apoptosis by various mechanisms (Ali et al., 2023; Shi et al., 2021). The following few studies represented the role of TRPM2 in ischemic stroke models.TRPM2 in primary cortical cultures of rat subjected to H2O2 induces apoptosis, and this is significantly reversed by using TRPM2 siRNA by inhibiting Ca2+ influx and cell death (Kaneko et al., 2006). The role of TRPM2 in oxidative stress induced by hypoxia has been studied in different animal models. TRPM2 KO mice, when subjected to transient ischemia by carotid artery occlusion, showed a 40% reduction in infarct volume. Some of the molecular pathways that contribute to cell survival are activation of protein kinase B (Akt) pathway and inhibition of glycogen synthase kinase-3β (GSK3β) pathway in TRPM2 KO mice (Alim et al., 2013). Genetic deletion of TRPM2 shows neuroprotective effects and improved sensory and motor outcomes in neonatal mice when compared to the wild type. TRPM2 KO mice showed an increased pro-survival signaling pathway and produced neuroprotective effects through Akt/GSK3β pathway (Huang et al., 2017). The use of TRPM2 inhibitors such as flufenamic acid (Nazıroğlu et al., 2007), clotrimazole (Hill et al., 2004), aminoethoxy diphenyl borate (APB) (Togashi et al., 2008), N-(p-amylcinnamoyl)anthranilic acid (ACA) (Kraft et al., 2006), and TRPM2 shRNA significantly reduced neuronal cell death following oxygen-glucose deprivation in males (Jia et al., 2011). Pharmacological inhibition of TRPM2 inhibits the adhesion of neutrophils in ischemic conditions (Gelderblom et al., 2014). There is a strong correlation between TRPM2 oxidative stress, inflammation, and ischemia followed by reperfusion injury (Huang et al., 2024).
5.2 TRPM2 and microglia
Apart from neuronal cells, TRPM2 is widely expressed in non-neuronal cells such as microglia and astrocytes (Turlova et al., 2018). Hypoxia-induced generation of H2O2 activated microglia increased TRPM2 mediated Ca2+ conductance in middle cerebral artery occlusion model injury (Patel et al., 2013). H2O2-induced oxidative stress revealed upregulation of TRPM2-like conductance and was reversed by flufenamic acid (Fonfria et al., 2006) in rat microglia. An increase in TRPM2 activity is associated with the generation of ROS, leading to the activation of Poly adenosine-diphosphate ribose polymerase 1 and TRPM2 activity were suppressed by inhibiting PKC and Nicotinamide Adenine Dinucleotide Phosphate (NADPH) oxidase (NOX) and downstream Mitogen-Activated Protein Kinase (MAPK)/extracellular signal-regulated kinase (MEK/ERK) pathway (Alawieyah Syed Mortadza et al., 2018). Release of cytokine interleukin-1 beta (IL-1β) from microglia is also mediated through TRPM2-dependent activation of Nucleotide-binding Oligomerization Domain-like receptor pyrin domain 3 inflammasome (Aminzadeh et al., 2018). Selenium, an important essential element through the GSH (Glutathione) peroxidase pathway, prevents interferon-gamma (IFNγ) induced activation of TRPM2 channel-mediated apoptosis in microglia (Akyuva et al., 2021). TRPM2 (knock out) KO in microglia suppressed the kainic acid-induced glial activation, cytokine production, and hippocampus paroxysmal discharges, affirming the role of TRPM2 via AMPK and mTOR pathway in epileptogenesis (Yıldızhan and Nazıroğlu, 2020). When stimulated with interferon-γ, microglia in the brain of C57bl/6 mice displayed excessive currents of TRPM2 (Akyuva et al., 2021). Macrophage exposure to LPS triggers the activation of TRPM2, thereby promoting NO production by regulating MAPK and JNK signaling pathways (Miyake et al., 2014).
5.3 TRPM2 in renal injury and fibrosis
Reactive oxygen species formed by renal ischemia-induced hypoxia and reperfusion activate TRPM2 ion channels. TRPM2-KO mice have shown resistance to renal injury (Khanahmad et al., 2022). Administration of TRPM2 antagonist, 8-bromo cADPR in Wistar rats shown to inhibit renal ischemia-reperfusion injury via caspase 3 inhibition (Eraslan et al., 2019). Cisplatin induced nephrotoxicity is also reduced by the activation of TRPM2 mediated autophagy (Yu et al., 2023). TRPM2-KO mice markedly improved renal dysfunction and its ablation remarkedly suppressed TGFβ mediated JNK activation renal fibrosis (Wang Y. et al., 2019). Thus, TRPM2 highlights its role in renal ischemic injury and fibrosis which is elevated during renal ischemic/reperfusion injury (Gao et al., 2014). Ischemia reperfusion-activated NOX and RAC1 were activated in WT mice but not in TRPM2-KO mice (Gao et al., 2014). Curcumin also reduces the albumin-evoked Ca2+-induced oxidative stress through the TRPM2 channel in renal tubules (Nazıroğlu et al., 2019).
5.4 TRPM2-mediated apoptosis
ROS-dependent neuronal inflammation and death involve multiple mechanisms, including pyro-apoptosis, autophagy, and apoptosis. ROS-mediated TRPM2 inhibits autophagy through downregulation of AMPK-induced Mammalian rapamycin target protein (mTOR), leading to ischemic/reperfusion-induced neuronal cell death (Hu et al., 2021). TRPM2 channels regulate cation permeability across the cell membrane that includes Zn2+ and the accumulation of the Zn2+ intracellularly induces ROS production, triggering lysosomal dysfunction and an increase in neuronal cell death (Ye et al., 2014). In addition, TRPM2 activates Calmodulin-dependent kinase II -mediated phosphorylation of Beclin1 that inhibits autophagy and induces neuronal death (Wang et al., 2016). TRPM2 mediated Ca2+ induced neuronal death was modulated by duloxetine preventing from apoptosis in hippocampus and dorsal root ganglion (DRG) of rats (Demirdaş et al., 2017).
5.5 TRPM2 in autoimmune disorders
The role of TRPM2 in inflammation has been extensively evaluated for Ca2+ mediated oxidative stress (ROS) that, in turn, activates innate immunity, thus delineating its role in autoimmune disorders such as rheumatoid arthritis, type 1 diabetes, multiple sclerosis, inflammatory bowel disease through multiple mechanisms. In an animal model of multiple sclerosis, TRPM2-KO or pharmacological inhibition of TRPM2 inhibits the progression of the disease. Moreover, decreased neutrophil infiltration in the central nervous system was observed in KO mice than in WT (Tsutsui et al., 2018). A decrease in neuronal antioxidant glutathione triggers the increased current of the TRPM2 channel (Belrose et al., 2012). Additional mechanisms such as ROS-ADPR- Poly adenosine diphosphate ribose polymerase mediated TRPM2 current in microglia thereby increase intracellular Ca2+ and thereby induce apoptosis of hippocampal pyramidal neurons in Alzheimer’s disease (Övey and Naziroğlu, 2015). TRPM2 also plays a key role in Amyloid beta (Aβ)/ROS-induced microglial-mediated neuroinflammation and neuronal death (Jiang et al., 2018). Glutathione depletion linked to oxidative stress induces apoptosis mediated through TRPM2 channels in microglial cells with Alzheimer’s disease model.
5.6 TRPM2 in hepatotoxicity
The liver is known as an important metabolic organ that regulates various functions. Acetaminophen is the commonly prescribed drug for pyrexia and is known to cause unwanted effects such as hepatotoxicity. ROS is the key mediator of acetaminophen-induced toxicity. Acetaminophen induced hepatocyte death by increasing Ca2+ influx in cultured rat and mouse hepatocytes which were blocked by ACA, clotrimazole or TRPM2-siRNA, and TRPM2-KO (Wang et al., 2016; Kheradpezhouh et al., 2014). TRPM2-mediated Ca2+ signalling activates Ca2+/CaMKII to inhibit autophagy (Ni et al., 2012). Also, acetaminophen-induced liver injury in WT mice was mitigated by treatment with CaMKII inhibitor KN-93 (Wang et al., 2016). In the hepatic ischemia-reperfusion injury model, TRPM2-mediated Ca2+ influx causes mitochondrial lipid peroxidation due to increasing arachidonate 12-lipoxygenase (Zhong et al., 2023). In addition, pretreatment with antioxidants such as N-acetyl cysteine and thymoquinone inhibits the Ca2+ entry by reducing the TRPM2 gene expression in the hepatocyte model of ischemia-reperfusion injury (Atilgan et al., 2022; Caglar et al., 2024).
6 TRPM2 expression and function in neuronal and non-neuronal pain
TRPM2 ion channels play an important role in generating, transmitting, and transforming nerve signals. Calcium ions play an important role in interneuronal communication, triggering an action potential and the release of certain neurotransmitters and mediators (Malko et al., 2019; Wang H. et al., 2019). Primary afferent neurons such as DRG and trigeminal ganglia also express TRPM2 channels (Vilar et al., 2020). The C-fibers of afferent neurons are mainly responsible for pain perception leading to depolarization. Substance P and calcitonin gene-related peptide (CGRP) of the peptidergic neurons, upon sustained stimulation, exacerbate tissue inflammation by infiltrating immune cells. TRPM2 is known to express abundantly in both peptidergic and non-peptidergic C-fibres (Matsumoto et al., 2016). TRPM2 is known to play a crucial role in afferent ganglia, where it responds to oxidative stress, particularly that caused by inflammatory injuries. This channel contributes to the sensory signaling processes by detecting and reacting to oxidative damage, which can occur during inflammation. The H2O2-induced inward currents in a whole cell patch clamp experiment with DRG neurons H2O2-induced inward currents were reversibly abolished by TRPM2 inhibitors 2-APB and ACA (Nazıroğlu et al., 2011; Naziroğlu et al., 2013). In addition, H2O2-mediated TRPM2 currents were inhibited by NADPH oxidase inhibitors such as apocynin and N-acetyl cysteine, which regulate the H2O2-ADPR-TRPM2 axis (Nazıroğlu, 2017). In cultured trigeminal ganglia upon H2O2 exposure, there is a significant increase in cytokine and chemokine levels which were prevented by treatment with TRPM2 inhibitors 2-APB (Chung et al., 2015). Therefore, TRPM2 plays an important role in the mechanisms of neuronal excitability and in the proinflammatory conditions under oxidative stress.
TRPM2 is widely expressed in immune cells. LPS stimulation of immune cells increased the expression of TRPM2, contributing to the production of cytokines (Wehrhahn et al., 2010). Also, neutrophil infiltration is linked with TRPM2 deficiency in carrageenan-induced inflammation (Haraguchi et al., 2012). IL- 1β secretion is also increased in macrophages by ROS-mediated TRPM2 activation (Zhong et al., 2013). The factors that regulate the calcium gating through TRPM2 include adenosine dinucleotide, cytokines, ROS, and intracellular calcium ions (Wang H. et al., 2019). Increased expression of TRPM2 in macrophages is associated with acceleration of inflammatory signals that regulate the pathophysiology of pain (Jang et al., 2018). Neuronal cell response to LPS is dependent on Ca2+, but the mechanisms involved are poorly elucidated. LPS is known to activate TLR-4, which initiates to activate of the downstream pathways of inflammation, such as phosphorylation of MAPKs, NF-kB translocation to the nucleus, and upregulation of inflammatory genes for cytokines such as TNFα, IL-1β, and IL-6 as shown in Figure 1. There is also evidence that LPS-mediated TLR-4 activates PLC, thereby mobilizing intracellular Ca2+. Thus, the role of Ca2+ and the production of cytokines are interlinked. Then, it comes to the question that controlling Ca2+ entry into the cell decreases inflammation. To understand this, the cytokine production generates ROS from mitochondria via modulating NADPH redox system is to be studied. The cytokine production generates ROS by activating NADPH oxidase, disrupting mitochondrial function, and activating MAPK pathways, including inducible nitric acid synthase and activating inflammasomes, contributing to the inflammatory response and cellular signaling (Bordt and Polster, 2014). The generation of free radicals takes place by intracellular production of H2O2 (Checa and Aran, 2020). Cellular activity sensors such as ion channels, particularly TRPM2, regulate the cation influx, including Ca2+, which further increases the inflammation process. Since regulation of inflammation is Ca2+ dependent which is also regulated by TRPM2 ion channel. Given the importance of TRPM2 ion channel in current scenario, modulating the function of TRPM2 is warranted. TRPM2 modulation with antioxidant compounds will alleviate the inflammation, thereby reducing nociception. Table 2 provides a summary of some research studies that use oxidative stress and inflammation as a model to modulate neuronal pain via TRPM2.
TABLE 2
| Cell Type | Inducer/Animal Model | Associated Pathology | Key observations | Ref. |
|---|---|---|---|---|
| Dorsal Root Ganglia (DRG) of rats | Spinal cord injury and Sciatic nerve injury | Neuronal death and apoptosis | Hypericum perforatum attenuates oxidative stress and apoptosis induced by SCI and SNI and, thereby, reducing, Ca2+ uptake through TRPM2 | Özdemir et al. (2016) |
| TRPM2-KO mice | LPS | bladder inflammation | Lacking TRPM2 attenuates LPS-induced inflammation and hypersensitivity | Kamei et al. (2021) |
| Hippocampal neuron/TRPM2-KO mice | Morphine | Inflammation and apoptosis | Morphine Induces Apoptosis, Inflammation, and Mitochondrial Oxidative Stress via Activation of TRPM2 Channel and Nitric Oxide Signaling Pathways | Osmanlıoğlu et al. (2020) |
| C57/BL6 mice | LPS | Inflammation-induced cognitive impairment | IL-1β or TRPM2 level knockdown helped counter the cognitive impairment caused by significant inflammation | Yang et al. (2024) |
| Sciatic nerve of rats | streptozotocin | Diabetic neuropathic pain | Hesperidin treatment attenuated diabetes-induced neuropathic pain by reducing TRPM2 channel activation | Bayir et al. (2023) |
| Microglia from WT and TRPM2KO | LPS/IFNγ | Inflammation | TRPM2-mediated Ca2+ signaling is necessary to activate p38 MAPK and JNK signaling and results in increased NO production in microglia | Miyake et al. (2014) |
| TRPM2-KO | Capsaicin/H2O2/acetic acid | Inflammation | Acetic acid-induced writhing was significantly attenuated in TRPM2-KO | So et al. (2015) |
| Cultured DRG | Rotenone | Neuropathy | ADPR and rotenone-induced inward electrical currents were reversibly abolished by the TRPM2 inhibitors, APB, and ACA | Nazıroğlu et al. (2011) |
| Hippocampal and DRG of Wistar rats | Middle cerebral artery occlusion/reperfusion | Apoptosis | Dexamethasone shows remarkable neuroprotective impairment effects in the hippocampus and DRG of ischemia-induced rats by reducing Ca2+ via TRPM2 | Akpınar et al. (2016) |
| Cultured rat Trigeminal Ganglia (TG) | H2O2 | Inflammation | TRPM2 role in H2O2-induced expression of inflammatory cytokines was studied | Chung et al. (2015) |
| Rat DRG | ADP-ribose and H2O2 | Neuropathic pain | NADPH oxidase-dependent activation of TRPM2 | Nazıroğlu and Braidy (2017) |
| Human Monocytes | LPS | Cytokine production | TRPM2 is required for LPS-induced cytokine production | Wehrhahn et al. (2010) |
| Mice macrophages | Carrageenan | Inflammatory and neuropathic pain | TRPM2-KO mice develop less severe allodynia | Haraguchi et al. (2012) |
Evidence of TRPM2 mediated neuropathic pain in response to inflammation and oxidative stress.
The role of TRPM2 in acute inflammatory mechanisms is acknowledged not only in neuronal cells but also in nonneuronal cells. TRPM2 channels are widely expressed in various cell types, including microglia (Mortadza et al., 2017; Kraft et al., 2004), macrophages (Zou et al., 2013), neurons (Kaneko et al., 2006; Alim et al., 2013), endothelial cells (Hecquet and Malik, 2009; Mittal et al., 2017; Yang et al., 2006), cardiomyocytes (Yang et al., 2006), dendritic cells (Sumoza-Toledo et al., 2011), and pancreatic beta cells (Ishii et al., 2006; Togashi et al., 2006). This indicates that TRPM2 plays multiple functional roles in many physiological and pathological processes. TRPM2 has been shown to regulate intracellular calcium levels through lysosomal, endoplasmic reticulum, and mitochondrial compartments through various pathways that mediate through both exogenous and endogenous mediators (Zhang et al., 2018).
TRPM2 is known to be regulated by other receptor mechanisms in inflammation and pain, such as the N-methyl-D-aspartate (NMDA) receptor (NMDAR), which activates to promote TRPM2-mediated Ca2+ influx via ERK1/2-dependent PARP-1 recruitment of microglia-dependent neuroinflammation (Raghunatha et al., 2020). TRPM2 channels can also be activated through intracellular signaling cascades initiated by pruritogen receptors and, thereby, neuronal activation. TRPM2 via Protein Kinase C gamma (PKC-γ), a neuro-specific PKC phosphorylates serine/threonine residues, activates and increases the expression of NMDAR, thereby increasing the excitotoxicity of NMDARS, and the interaction between them is thought to be mediated by oxidative stress (Sun and Alkon, 2014). Antioxidants such as Selenium reduced the fibromyalgia induced increase in TRPM2 and TRPV1 currents, pain intensity, ROS, and intracellular free Ca2+ in sciatic and DRG neurons (Yüksel et al., 2017).
7 Toll-like receptors
Toll-like receptors (TLRs) are pattern recognition receptors embedded in the phospholipid membrane as well as endosomes and lysosomes (Sahoo, 2020). In humans, there are ten TLRs (TLR1-TLR10), but overall, there are 12 TLRs. Different TLRs recognize and activate different pathogen-associated molecular patterns (PAMPs) (Kawai and Akira, 2007). TLR-1 and TLR-2 link together and recognize bacteria lipoproteins which activate immune responses. TLR-2 not only forms a heterodimer with TLR-1 but also with TLR-6 to recognize microbial patterns. TLR-3 is found embedded in endosomes, senses viral infections, and activates the production of type-I interferons. TLR-4 is activated by bacterial LPS, which is the main cell wall component of Gram-negative bacteria (Kuzmich et al., 2017). This activation can contribute to inflammation and pain pathways, leading to the release of pro-inflammatory cytokines and other mediators that contribute to the development or exacerbation of neuropathic pain. Some Gram-negative bacteria associated with this process include Escherichia coli (E. coli), Pseudomonas aeruginosa, Klebsiella pneumoniae, Salmonella spp., Neisseria meningitidis, Haemophilus influenzae, Helicobacter pylori (Deng and Chiu, 2021; Staurengo-Ferrari et al., 2022). TLR4 can also be activated by a variety of non-LPS ligands. These ligands include endogenous danger-associated molecular patterns (DAMPs) and exogenous pathogen-associated molecular patterns (PAMPs) other than LPS (Facchini, 2018; Alexzander Asea et al., 2002; Erridge, 2010). TLR-7, TLR-8, and TLR-9 are located in endosomes and lysosomes, while TLR-5, found on the cell surface, recognizes bacterial flagellins involved in motility, leading to the activation of inflammatory cytokines (Tegtmeyer et al., 2020). They are mainly expressed in various immune cells and initiate inflammatory responses. TLR-10, although found on both the cell surface and endosomes, the actual functions are not well-defined (Noreen and Arshad, 2015).
However, Toll-like receptors are a group of receptors that recognize and initiate innate and adaptive immune responses. Like other receptors, they span cellular transmembrane with an external domain of leucine repeats and cytosolic Toll interleukin-1 receptor (TIR) domains to activate downstream pathways. The external domain tends to recognize endotoxins, heat shock proteins, cellular damage products, and High mobility group box 1 (HMGB-1) proteins (Bettoni et al., 2008). The receptors are widely distributed not only on antigen-presenting cells but also on myocytes, adipocytes, thyroid cells, mesangial cells, and fibroblasts. The receptors are also expressed in sensory neurons, microglial cells, and astrocytes (Vaure and Liu, 2014). TLR-4 receptor undergoes dimerization with myeloid differentiation protein-2 upon ligand binding to microdomain lipid rafts and initiates TIR adoption molecules, thereby initiating a signaling cascade.
TLR-4, through the TIR domain-containing adapter-inducing Inhibitor of the kappa B (IkB) pathway, induces NF-kB and expression of proinflammatory cytokines such as IL-1, IL-6, and TNF-α. Activation and inhibition of these pathways create a balance between the production of these cytokines and Interferon Type-1 (IFN-1) (Roy et al., 2016; Kawasaki and Kawai, 2014).
8 Role of TLR-4 in the regulation of neuronal pain and inflammation
TLR4 plays an important role in adaptive immune response, induction, and maintenance of acute and chronic pain states. Recognition of pathogens and products from damaged neurons by microglia leads to activation of TLR-4 and increased expression of proinflammatory mediators. Sustained activation or dysregulation of TLR-4 contributes to inflammatory and autoimmune diseases such as Crohn’s Disease, atherosclerosis, rheumatoid arthritis, type 1 diabetes, and other neurodegenerative diseases (Takeda et al., 2003; Booth et al., 2011; Hosseini et al., 2015).
Microglial activation of TLR-4 after spinal cord injury is pivotal for the induction of pain by modulating proinflammatory cascade and expression of TNFα, IFN-γ, IL-6, IL-1β, and NF-kB (Kuang et al., 2012; Stokes et al., 2013). In rat animal model of chronic constriction injury TLR-4 antagonists and siRNA-mediated suppression of spinal cord TLR-4 signaling prevent the activation of the NF-kB pathway and production of proinflammatory cytokines, thereby attenuating allodynia and thermal hyperalgesia, emphasizing its preventive role in neuropathic pain (Eidson and Murphy, 2013; Wu et al., 2010). TLR-4 also mediates the conversion of acute to chronic pain. Intrathecal administration of TLR-4 antagonists reversed the chronic constriction injury-induced thermal hyperalgesia and mechanical allodynia in wild-type mice, whereas in knockout and point mutant mice, attenuation of thermal hypersensitivity along with spinal microglial activation and lower proinflammatory cytokines suggesting its role in the maintenance of chronic pain (Hutchinson et al., 2008; Christianson et al., 2011; Nicotra et al., 2012; Woller et al., 2017). Administration of TLR-4 antisense oligonucleotide prevents thermal hypersensitivity with reduced expression of mRNA for microglial markers and spinal proinflammatory cytokines (Tanga et al., 2005).
Hydrogen peroxide, a ROS, oxidatively modifies TLR-4 proteins, and it mimics LPS binding (Powers et al., 2006). It activates the release of damaged-associated molecular proteins (DAMPs) from dying cells, which trigger TLR-4 proteins (Shim et al., 2017). H2O2 also modulates the activity of kinases and transcription factors such as NF-κB and MAPKs, which are critical for TLR-4 signaling, which can amplify inflammatory responses (Burgueno et al., 2021). When H2O2 production is increased, it acts as a secondary signal to aid TLR-4-mediated defenses guaranteeing an immune response (Zhou et al., 2013). The interaction between ROS and TLR-4 can intensify inflammation indicating tissue damage and disease development (Gunawardena et al., 2019).
9 Relationship between TRPM2 and TLR-4 in neuropathic pain
The generation of neuropathic pain involves complex pathways within the nervous system, both peripheral and central. The process is initiated by damage or dysfunction in the somatosensory nervous system, which leads to abnormal pain signaling. Neuropathic pain often involves nerve injury or chronic inflammation, leading to increased oxidative stress and inflammatory mediators such as proinflammatory cytokines, chemokine, and ROS, which can activate TRPM2 channels. Understanding the mechanisms by which TRPM2 contributes to neuropathic pain can provide insights into potential therapeutic targets for managing chronic pain conditions.
On the other hand, TLR-4 is a key player in the immune system, primarily recognized for its role in detecting pathogens and initiating inflammatory responses. However, TLR-4 plays a critical role in the pathogenesis of neuropathic pain by driving neuroinflammation, central sensitization, and the maintenance of a chronic pain state. Targeting TLR-4 or its downstream signaling pathways is a potential therapeutic strategy for managing neuropathic pain.
TLR-4 is expressed on the cell membrane and senses pathogen-associated molecular patterns such as LPS, peptidoglycans, chitin, and glucans (Brown et al., 2011). Upon recognition and activation, TLR-4 recruits adaptor molecules such as MyD88 and intracellular Toll-interleukin (IL)-1 receptor domains and promotes downstream pathways leading to the secretion of proinflammatory mediators, including cytokines (Blasius and Beutler, 2010; Santoni et al., 2015). These proinflammatory responses by the immunocompetent cells in the brain, such as microglia, contribute to neuroinflammation leading to pain. In chronic states such as neuropathic pain, microglial activation results in deleterious consequences (Block et al., 2007). Microglia express a wide variety of ion channels, including TRP channels, that are necessary for cytokine production, proliferation, and migration of microglia (Echeverry et al., 2016). Studies indicate that H2O2-induced TRPM2 activation mediates Ca2+ influx, which modulates physiological and pathological cellular functions (Shirakawa and Kaneko, 2018). TRPM2 is expressed in both neurons and glia, and oxidative stress-induced TRPM2 activation is implicated in neuronal diseases (So et al., 2015). Miyake et al. (2014) demonstrated that LPS and IFN-γ can stimulate TRPM2 mediated Ca2+ in microglia. They also showed that activation of TRPM2 results in increased NO production. Interaction between nociceptive neurons and glial cells plays an important role in neuropathic pain (Ren and Dubner, 2010).
TRPM2, a calcium-permeable cation channel present in sensory neurons, is capable of recognizing mechanical, chemical, and thermal stimuli (Santoni et al., 2015). TRPM2 activation leads to increased calcium levels, which also results in a pain-signaling response in the sensory neurons. TLR-4 has been linked to neuroinflammation (Table 3) and the sensitization of nociceptors (Malko et al., 2019). When TLR-4 interacts with ROS, TLR-4 proteins can intensify inflammation and hypersensitivity to pain through the opening of TRPM2 channels. Thus, the pathways of both TRPM2 and TLR4 converge on neuroinflammation (Yamamoto and Shimizu, 2016). Neuropathic Pain is a chronic pain caused by a disease of the somatosensory nervous system (Finnerup et al., 2021). TRPM2 contributes to the release of pro-inflammatory cytokines (Haraguchi et al., 2012), and the production of cytokines, in turn, activates TLR-4, which plays an important role in neuropathic pain and induces inflammatory and immune responses (Kuzmich et al., 2017).
TABLE 3
| Pathological condition/disorders | Experimental paradigm | Treatment | Mechanism/pathways | Principal Effects/Remarks | Ref. |
|---|---|---|---|---|---|
| Neuropathic pain | Thermal and Mechanical Hypersensitivities in a Rat Model of Paclitaxel-Induced Peripheral Neuropathic Pain | Electro-acupuncture (EA) | NF-kappaB inhibitor alpha (NF-κB) and Mitogen-activated protein (MAP) kinase pathways | EA suppresses TLR4 and its downstream signaling molecule MyD88 overexpression in DRGs of paclitaxel-treated rats | Li et al. (2019) |
| Chronic Neuropathic Pain | The therapeutic effect of β-sitosterol on chronic neuropathic pain by performing behavioral tests on Sciatica models (chronic constriction injury on SD rats) | β-Sitosterol | β-sitosterol can affect microglial polarization by inhibiting the TLR4/NF-κB signaling pathway | β-Sitosterol Reduces Pain Sensitivity in the Right Hind Limb of Sprague‒Dawley Rats | Zheng et al. (2023) |
| Neuropathic pain | Therapeutic effect of ferulic acid on the chronic constriction injury induced pain rat model via the von Frey test and acetone experiment | Ferulic acid | The levels of IBA‐1, IL‐1β, iNOS, TLR4, Myd88, p‐NF‐κB, and p‐p38MAPK increased signaling pathway | Ferulic acid can promote injured sciatic nerve repair by reducing neuronal cell apoptosis and inflammatory infiltration though the TLR4/NF‐κB pathway | Zhang et al. (2023) |
| Migraine and Functional Gastrointestinal Disorders | Using a murine model of light aversion produced by compound 48/80. TLR4 in migraine-like behavior and neuronal activation | Compound 48/80 | TLR4 utilizes both MyD88 and TRIF adapter protein pathways for downstream signaling | TLR4 signaling in migraine is the report that naloxone was reported to be effective in treating acute migraine attacks | Ramachandran et al. (2019) |
| Cancer-induced Neuropathic Pain | Cancer-induced neuropathy model and determined the thresholds of cold allodynia and thermal and mechanical hyperalgesia | Ruthenium Red | TRPV1 over TRPV4 antagonism is attributed to its multiple mechanisms of action and different pathways and target receptors other than the TRP channels | The increased expression of TLR4 and ERK1/2 reveals immune response and tumor progression, respectively, and their ultimate decrease is an indicator of nerve damage | Maqboul and Elsadek (2018) |
| Acute Inflammatory Visceral Pain (AIVP) | AIVP was used to examine the antinociceptive efficiency of DEX and assess its effects on the activation of the ERK and TLR4 signaling pathway and release of CGRP during visceral hypersensitivity | Dexmedetomidine (DEX) | TLR4 receptor and its downstream NF-κB and IRF3 were upregulated, and the phosphorylation of P65 and IRF3 also increased upon acetic acid treatment | Antinociceptive effects of DEX might be partially mediated via suppression of the inflammatory responses associated with AIVP | Liu et al. (2018) |
| Chronic Sciatica | The therapeutic effect of α-asarone on CCI of the sciatic nerve via behavioral tests, pathology, and immunohistochemistry | α-Asarone | α-Asarone decreased the mRNA levels of IL1β, TNF-α, TRPV1-4, TRPA1, and TRPM8 | α-Asarone relieves chronic sciatica by decreasing the levels of inflammatory factors, inhibiting peripheral sensitization, and favoring the repair of damaged nerves | Zhang et al. (2023) |
| Multiple Sclerosis | Spinal cord alterations induced by this novel SD rat model of MOG-induced EAE, optimized to avoid motor impairments/disabilities | myelin-oligo dendrocyte-glycoprotein (MOG) | Ligation of TLR4 induces NF-kB activation that primes the NLRP3 inflammasome and can ultimately lead to the production of the proinflammatory cytokine IL-1β t | Doses of 8 and 16 μg MOG could produce long-lasting mechanical allodynia in the absence of motor impairments/disabilities in male SD rats | Kwilasz et al. (2022) |
| Rheumatoid Arthritis) | Molecular mechanisms of Baihu-Guizhi decoction (BHGZD), against active RA were validated by a series of experiments based on the adjuvant-induced arthritis-modified rat model | (BHGZD) | BHGZD suppress TLR4/PI3K/AKT/NFκB signaling-related protein activation, and subsequently inhibit NLRP3 inflammasome-induced pyroptosis | BHGZD effectively improved disease severity of active RA rats, elevating pain thresholds, relieving joint inflammation and bone erosion via inhibiting TLR4/PI3K/AKT/NFκB signaling to suppress the activation of the NLRP3 inflammasome | Li et al. (2022) |
| Interstitial cystitis/bladder pain syndrome (IC/BPS) | IC rat model by intraperitoneal injection of cyclophosphamide. MSC-EVs were isolated from the culture supernatants of human umbilical cord-derived MSCs | Mesenchymal stem cell-derived extracellular vesicles (MSC-EVs) | the expression levels of NLRP3, Caspase-1, IL‐1β and IL‐18 in the SDH were reduced | Injection of MSC-EVs can alleviate neuroinflammation and mechanical allodynia in IC rats by inhibiting NLRP3 inflammasome activation | Zhang et al. (2022) |
| Post-epidural fibrosis | murine laminectomy model to observe the effect of metformin on epidural scars | Metformin | Metformin inhibited the hyper-proliferation of epidural scars after laminectomy via the reduction in fibronectin and collagen deposition by inhibiting the HMGB1/TLR4 and TGF‐β1/Smad3 signaling pathway | metformin may be a potential therapeutic option to mitigate epidural fibrosis after laminectomy | Song et al. (2021) |
Pain signaling pathways through TLR4 and TRP channels.
TRPM2 channel is a newly cited player for neuroinflammation mediated in response to LPS, a classical target for TLR-4 activation where the interaction between TLR-4 and TRPM2 in neuronal cells was least explored. TLR-4-mediated responses required Ca2+ influx through TRPM2 (Schappe et al., 2018; Tauseef et al., 2012), and the binding of LPS to TLR-4 activation increases the production of diacylglycerol (DAG) and the subsequent increase in Ca2+ concentration, resulting in MyD88-NF-kB activation (Mohammadi, 2019). TRPM2-mediated Ca2+ entry is implicated in inflammatory responses in neuronal and non-neuronal cells (Tauseef et al., 2012; Mehta et al., 2003). LPS-TLR-4 activation triggers NF-kB-mediated cytokine production resulting in TRP-mediated Ca2+ influx. Cytokine increases intracellular Ca2+ concentration, thereby inducing early ROS production preceded by TRPM2-mediated Ca2+ influx (Simon et al., 2017; López-Requena et al., 2017). Consequently, TRPM2 regulates total ROS production and Ca2+ influx. The existing studies suggest TRPM2 is not only involved in physiological nociceptive pain, but also in inflammatory and neuropathic pain (So et al., 2015).
The role of TLR-4-induced inflammation via calcium channels has been reported. Activation of TLR-4 through LPS-induced cytokine production results in an imbalance of basal calcium and store-operated calcium channels, particularly TRPM2, in microglia through Orai-1, a downstream protein of TLR4-MAPK (Birla et al., 2022). Various proinflammatory cytokines are produced during the immune response to an infection. The absence of TRPM2 channels improved neuroethology and pathological changes and decreased inflammatory cytokines and apoptosis proteins in TRPM2 knock-out mice in the LPS-induced sepsis model associated with encephalopathy infection (Zhu et al., 2019). In a study, where the LPS-induced production of IL-6, IL-8, and TNF-α in THP1 monocytic cells was significantly attenuated by using shRNA to reduce TRPM2 expression (Wehrhahn et al., 2010). Also, increased cell-free heme activates TLR4-mediated ER stress and ROS leading to spinal microglial activation and neuroinflammation, contributing to acute and chronic pain (Lei et al., 2021). Ketamine, an anesthetic agent, attenuated hypoxia-induced TRPM2-mediated Ca2+ influx, ROS, NMDAR, and ROS in neuronal cell (SH-SY5Y) death (Osmanlioğlu, 2022).
There is a reciprocal relationship between neuroinflammation and oxidative stress. These two processes are not independent but rather complex reciprocal interactions (Teixeira-Santos et al., 2020). NOX activity upregulates the expression of proinflammatory cytokines and proinflammatory cytokines increase ROS through NOX (Kallenborn-Gerhardt et al., 2013). Initiation of TLR4 downstream pathway requires NOX activity (Haslund‐Vinding et al., 2017) and TLR-4 receptors may be required for NOX expression (Lim et al., 2013), thus both are dependent on each other. These interdependent complex mechanisms that regulate neuropathic pain require Ca2+ ion, which is well controlled by the TRPM2 ion channel sensitive to cellular redox potential (Ogawa et al., 2016), and hence modulated TRPM2 can alleviate neuropathic pain.
NOX 2 and NOX 4 have been implicated in chronic pain mechanisms such as neuropathic pain (Grace et al., 2016). Inhibition of NOX and its isoforms 2 and 4 have shown to be beneficial in neuropathic pain models (Kallenborn-Gerhardt et al., 2013). Macrophages produce ROS during immune response, and it is well regulated by NOX. This enzyme activity can be regulated by Ca2+, which is controlled by protein kinase C α and membrane potential (Bedard and Krause, 2007). However, TRPM2KO mice showed greater mortality than WT in LPS-induced inflammation (Di et al., 2012). In addition, TRPM2 deficiency reduces oxidized low-density lipoprotein uptake by macrophages, thereby minimizing macrophage infiltration and inflammatory response by modulation of NOX (Zong et al., 2022).
However, the molecular mechanism underlying the interaction between TRPM2 and TLR4s and the quantum of impact in acute and chronic neuropathic pain remains unclear. It would be worth focusing on TRPM2 interaction with TLR4 in the context of neuropathic pain. Mechanistic studies that focus on TLR4-TRPM2 interaction will open new pathway interactions for treating inflammation-associated diseases such as neuropathic pain.
10 Conclusion
This review summarizes the functional role and mechanisms of TRPM2 that mediate Ca2+ influx under oxidative stress and inflammation linked with TLR-4. The reported literature in this review demonstrates the role of the TRPM2 ion channel in various pathological states and, more importantly, neuropathic pain. Many research findings support the evidence that oxidative stress and LPS-induced TLR4-mediated inflammation exert their effects mediated by the TRPM2 channel in neuronal and non-neuronal tissues. The existing and growing scientific evidence of TRPM2 in various pain pathologies has made researchers and the pharmaceutical industry focus on the development of novel targets for diagnostic and therapeutic approaches for pain treatment. This review also demonstrates the role of various inhibitors/drugs that modulate neuropathic pain in various disease states. However, the role of TRPM2 ion channels and their association with Toll-like receptors in pain management needs to be studied in more detail for the development of effective strategies to treat neuropathic pain.
Statements
Author contributions
VM: Writing–original draft, Investigation. AR: Writing–review and editing, Investigation. ZK: Investigation, Writing–review and editing. SQ: Data curation, Formal Analysis, Writing–review and editing, Investigation, Visualization. FD: Conceptualization, Data curation, Formal Analysis, Funding acquisition, Investigation, Methodology, Resources, Software, Supervision, Validation, Visualization, Writing–review and editing.
Funding
The author(s) declare that financial support was received for the research, authorship, and/or publication of this article.
Acknowledgments
The authors of this review were in part supported by the Fisch College of Pharmacy, at UT-Tyler, Endowment Fellowship funds, and UT-Tyler Office of Research and Scholarship funds.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Glossary
| ACA | N-(p-amylcinnamomyl) Anthranilic acid |
| ADPR | Adenosine diphosphate ribose |
| AIVP | Acute inflammatory visceral pain |
| AKT | Protein kinase B |
| AMPK | Adenosine 5′-monophosphate (AMP)-activated protein kinase |
| APB | Aminoethoxy diphenyl borate |
| Aβ | Amyloid beta |
| BHGZD | Baihu-Guizhi decoction |
| BPS | Bladder pain syndrome |
| cADPR | cyclic ADP-ribose |
| CGRP | Calcitonin gene related peptide |
| DAG | Diacyl glycerol |
| DAMPS | Damage-associated molecular patterns |
| DEX | Dexmeditomidine |
| DRG | Dorsal root ganglia |
| ER | Endoplasmic reticulum |
| ERK | Extracellular signal-regulated kinase |
| GSH | Glutathione |
| GSK3β | Glycogen synthase kinase-3β |
| H2O2 | Hydrogen peroxide |
| HMGB-1 | High mobility group box 1 |
| IFN-1 | Interferon Type 1 |
| IkB | Inhibitor of kappa B |
| IL-1β | Interleukin-1 beta |
| IL-6 | Interleukin-6 |
| IP3 | Inositol triphosphate |
| JNK | c-Jun N-terminal kinase |
| KO | Knock Out |
| LPS | Lipopolysaccharide |
| MAPKs | Mitogen-Activated Protein Kinase |
| MOG | Myeloid oligodendrocyte glycoprotein |
| MSC | Mesenchymal stem cell |
| mTOR | Mammalian rapamycin target protein |
| Myd88 | Myeloid differentiation Primary Response |
| NADPH | Nicotinamide Adenine Dinucleotide phosphate |
| NF-kB | Nuclear factor kappa B |
| NMDA | N-methyl D-aspartate |
| NOX | NADPH oxidase |
| NUDT9-H | Nudix hydroxylase |
| PAMPS | Pathogen-associated molecular patterns |
| PARP1 | Poly adenosine diphosphate ribose polymerase |
| PI | Phosphatidyl inositol |
| PKC | Protein Kinase C |
| PLC | Phospholipase C |
| ROS | Reactive Oxygen Species |
| SCI | Spinal Cord Injury |
| SD | Sprague Dawley |
| TGFβ | Tumor growth factor |
| TIR | Toll interleukin −1 receptor |
| TLR | Toll-like-receptor |
| TNFα | Tumor necrosis factor α |
| TRAF6 | Tumor necrosis factor receptor (TNFR)-associated factor 6 |
| TRIF | TIR (Toll/interleukin-1 receptor) domain-containing adaptor protein |
| TRP | Transient receptor potential |
| TRPA | Transient receptor potential Ankyrin |
| TRPC | Transient receptor potential Canonical |
| TRPM | Transient receptor potential Melastatin |
| TRPV | Transient receptor potential Vanilloid |
| WT | Wild Type |
References
1
Aizawa N. Fujimori Y. Kobayashi J. I. Nakanishi O. Hirasawa H. Kume H. et al (2018). KPR-2579, a novel TRPM8 antagonist, inhibits acetic acid-induced bladder afferent hyperactivity in rats. Neurourol. Urodyn.37 (5), 1633–1640. 10.1002/nau.23532
2
Aizawa N. Fujita T. (2022). The TRPM8 channel as a potential therapeutic target for bladder hypersensitive disorders. J. Smooth Muscle Res.58 (0), 11–21. 10.1540/jsmr.58.11
3
Akpınar H. Nazıroğlu M. Övey İ. S. Çiğ B. Akpınar O. (2016). The neuroprotective action of dexmedetomidine on apoptosis, calcium entry and oxidative stress in cerebral ischemia-induced rats: contribution of TRPM2 and TRPV1 channels. Sci. Rep.6 (1), 37196. 10.1038/srep37196
4
Akyuva Y. Nazıroğlu M. (2020). Resveratrol attenuates hypoxia-induced neuronal cell death, inflammation and mitochondrial oxidative stress by modulation of TRPM2 channel. Sci. Rep.10 (1), 6449. 10.1038/s41598-020-63577-5
5
Akyuva Y. Nazıroğlu M. Yıldızhan K. (2021). Selenium prevents interferon-gamma induced activation of TRPM2 channel and inhibits inflammation, mitochondrial oxidative stress, and apoptosis in microglia. Metab. Brain Dis.36, 285–298. 10.1007/s11011-020-00624-0
6
Alawieyah Syed Mortadza S. Sim J. A. Neubrand V. E. Jiang L. H. (2018). A critical role of TRPM2 channel in Aβ42 -induced microglial activation and generation of tumor necrosis factor-α. Glia66 (3), 562–575. 10.1002/glia.23265
7
Alexzander Asea M. R. Kabingu E. Boch J. A. Baré O. Auron P. E. Stevenson M. A. et al (2002). Novel signal transduction pathway utilized by extracellular HSP70: ROLE of toll-LIKE receptor (TLR) 2 and TLR4. J. Biol. Chem.277 (17), 15028–15034. 10.1074/jbc.M200497200
8
Ali E. S. Chakrabarty B. Ramproshad S. Mondal B. Kundu N. Sarkar C. et al (2023). TRPM2-mediated Ca2+ signaling as a potential therapeutic target in cancer treatment: an updated review of its role in survival and proliferation of cancer cells. Cell Commun. Signal.21 (1), 145. 10.1186/s12964-023-01149-6
9
Alim I. Teves L. Li R. Mori Y. Tymianski M. (2013). Modulation of NMDAR subunit expression by TRPM2 channels regulates neuronal vulnerability to ischemic cell death. J. Neurosci.33 (44), 17264–17277. 10.1523/JNEUROSCI.1729-13.2013
10
Aminzadeh M. Roghani M. Sarfallah A. Riazi G. H. (2018). TRPM2 dependence of ROS-induced NLRP3 activation in Alzheimer's disease. Int. Immunopharmacol.54, 78–85. 10.1016/j.intimp.2017.10.024
11
Asiri Y. I. Hassan M. Z. (2023). An overview of ion channels therapeutics in the treatment of pain. Arabian J. Chem.16 (10), 105180. 10.1016/j.arabjc.2023.105180
12
Atilgan F. A. Atescelik M. Yilmaz M. Turk A. Gurger M. Goktekin M. C. et al (2022). Effects of N-acetyl cysteine on TRPM2 expression in kidney and liver tissues following malathion intoxication. Biotech. and Histochem.97 (5), 340–346. 10.1080/10520295.2021.1986639
13
Backonja M.-M. (2003). Defining neuropathic pain. Anesth. and Analgesia97 (3), 785–790. 10.1213/01.ANE.0000062826.70846.8D
14
Bamps D. Blockeel A. J. Dreesen E. Marynissen H. Laenen J. Van Hecken A. et al (2023). TRPA1 antagonist LY3526318 inhibits the cinnamaldehyde-evoked dermal blood flow increase: translational proof of pharmacology. Clin. Pharmacol. Ther.114 (5), 1093–1103. 10.1002/cpt.3024
15
Baron R. (2006). Mechanisms of disease: neuropathic pain—a clinical perspective. Nat. Clin. Pract. Neurol.2 (2), 95–106. 10.1038/ncpneuro0113
16
Bayir M. H. Yıldızhan K. Altındağ F. (2023). Effect of hesperidin on sciatic nerve damage in STZ-induced diabetic neuropathy: modulation of TRPM2 channel. Neurotox. Res.41 (6), 638–647. 10.1007/s12640-023-00657-0
17
Bedard K. Krause K.-H. (2007). The NOX family of ROS-generating NADPH oxidases: physiology and pathophysiology. Physiol. Rev.87 (1), 245–313. 10.1152/physrev.00044.2005
18
Belrose J. C. Xie Y. F. Gierszewski L. J. MacDonald J. F. Jackson M. F. (2012). Loss of glutathione homeostasis associated with neuronal senescence facilitates TRPM2 channel activation in cultured hippocampal pyramidal neurons. Mol. Brain5 (1), 11. 10.1186/1756-6606-5-11
19
Bettoni I. Comelli F. Rossini C. Granucci F. Giagnoni G. Peri F. et al (2008). Glial TLR4 receptor as new target to treat neuropathic pain: efficacy of a new receptor antagonist in a model of peripheral nerve injury in mice. Glia56 (12), 1312–1319. 10.1002/glia.20699
20
Biggs J. E. Stemkowski P. L. Knaus E. E. Chowdhury M. A. Ballanyi K. Smith P. A. (2015). Suppression of network activity in dorsal horn by gabapentin permeation of TRPV1 channels: implications for drug access to cytoplasmic targets. Neurosci. Lett.584, 397–402. 10.1016/j.neulet.2014.07.033
21
Birla H. Xia J. Gao X. Zhao H. Wang F. Patel S. et al (2022). Toll-like receptor 4 activation enhances Orai1-mediated calcium signal promoting cytokine production in spinal astrocytes. Cell calcium105, 102619. 10.1016/j.ceca.2022.102619
22
Bittner S. Ruck T. Fernández-Orth J. Meuth S. G. (2014). TREK-king the blood–brain-barrier. J. Neuroimmune Pharmacol.9, 293–301. 10.1007/s11481-014-9530-8
23
Blasius A. L. Beutler B. (2010). Intracellular toll-like receptors. Immunity32 (3), 305–315. 10.1016/j.immuni.2010.03.012
24
Block M. L. Zecca L. Hong J.-S. (2007). Microglia-mediated neurotoxicity: uncovering the molecular mechanisms. Nat. Rev. Neurosci.8 (1), 57–69. 10.1038/nrn2038
25
Booth J. Wilson H. Jimbo S. Mutwiri G. (2011). Modulation of B cell responses by Toll-like receptors. Cell tissue Res.343, 131–140. 10.1007/s00441-010-1031-3
26
Bordt E. A. Polster B. M. (2014). NADPH oxidase-and mitochondria-derived reactive oxygen species in proinflammatory microglial activation: a bipartisan affair?Free Radic. Biol. Med.76, 34–46. 10.1016/j.freeradbiomed.2014.07.033
27
Bouali-Benazzouz R. Landry M. Benazzouz A. Fossat P. (2021). Neuropathic pain modeling: focus on synaptic and ion channel mechanisms. Prog. Neurobiol.201, 102030. 10.1016/j.pneurobio.2021.102030
28
Brown J. Wang H. Hajishengallis G. N. Martin M. (2011). TLR-signaling networks: an integration of adaptor molecules, kinases, and cross-talk. J. Dent. Res.90 (4), 417–427. 10.1177/0022034510381264
29
Burgueno J. F. Fritsch J. González E. E. Landau K. S. Santander A. M. Fernández I. et al (2021). Epithelial TLR4 signaling activates DUOX2 to induce microbiota-driven tumorigenesis. Gastroenterology160 (3), 797–808 e6. 10.1053/j.gastro.2020.10.031
30
Caglar K. Dokuyucu R. Agturk G. Tumer C. Tutuk O. Gocmen H. D. et al (2024). Effect of thymoquinone on transient receptor potential melastatin (TRPM) channels in rats with liver ischemia reperfusion model in rats. Iran. J. Basic Med. Sci.27, 319–325. 10.22038/IJBMS.2023.71990
31
Checa J. Aran J. M. (2020). Reactive oxygen species: drivers of physiological and pathological processes. J. Inflamm. Res.13, 1057–1073. 10.2147/JIR.S275595
32
Christianson C. A. Dumlao D. S. Stokes J. A. Dennis E. A. Svensson C. I. Corr M. et al (2011). Spinal TLR4 mediates the transition to a persistent mechanical hypersensitivity after the resolution of inflammation in serum-transferred arthritis. Pain152 (12), 2881–2891. 10.1016/j.pain.2011.09.020
33
Chung M.-K. Asgar J. Lee J. Shim M. S. Dumler C. Ro J. Y. (2015). The role of TRPM2 in hydrogen peroxide-induced expression of inflammatory cytokine and chemokine in rat trigeminal ganglia. Neuroscience297, 160–169. 10.1016/j.neuroscience.2015.03.067
34
de Almeida A. S. Pereira G. C. Brum E. d. S. Silva C. R. Antoniazzi C. T. d. D. Ardisson-Araújo D. et al (2021). Role of TRPA1 expressed in bone tissue and the antinociceptive effect of the TRPA1 antagonist repeated administration in a breast cancer pain model. Life Sci.276, 119469. 10.1016/j.lfs.2021.119469
35
Demirdaş A. Nazıroğlu M. Övey İ. S. (2017). Duloxetine reduces oxidative stress, apoptosis, and Ca 2+ entry through modulation of TRPM2 and TRPV1 channels in the hippocampus and dorsal root ganglion of rats. Mol. Neurobiol.54, 4683–4695. 10.1007/s12035-016-9992-1
36
Deng L. Chiu I. M. (2021). Microbes and pain. PLoS Pathog.17 (4), e1009398. 10.1371/journal.ppat.1009398
37
Di A. Gao X. P. Qian F. Kawamura T. Han J. Hecquet C. et al (2012). The redox-sensitive cation channel TRPM2 modulates phagocyte ROS production and inflammation. Nat. Immunol.13 (1), 29–34. 10.1038/ni.2171
38
Duitama M. Vargas-López V. Casas Z. Albarracin S. L. Sutachan J. J. Torres Y. P. (2020). TRP channels role in pain associated with neurodegenerative diseases. Front. Neurosci.14, 782. 10.3389/fnins.2020.00782
39
Echeverry S. Rodriguez M. J. Torres Y. P. (2016). Transient receptor potential channels in microglia: roles in physiology and disease. Neurotox. Res.30, 467–478. 10.1007/s12640-016-9632-6
40
Eidson L. N. Murphy A. Z. (2013). Blockade of Toll-like receptor 4 attenuates morphine tolerance and facilitates the pain relieving properties of morphine. J. Neurosci.33 (40), 15952–15963. 10.1523/JNEUROSCI.1609-13.2013
41
Eraslan E. Tanyeli A. Polat E. Polat E. (2019). 8‐Br‐cADPR, a TRPM2 ion channel antagonist, inhibits renal ischemia–reperfusion injury. J. Cell. physiology234 (4), 4572–4581. 10.1002/jcp.27236
42
Eren-Koçak E. Dalkara T. (2021). Ion channel dysfunction and neuroinflammation in migraine and depression. Front. Pharmacol.12, 777607. 10.3389/fphar.2021.777607
43
Erridge C. (2010). Endogenous ligands of TLR2 and TLR4: agonists or assistants?J. Leukoc. Biol.87 (6), 989–999. 10.1189/jlb.1209775
44
Facchini F. (2018). Toll-like Receptor 4 (TLR4) therapeutic modulation: a chemical biology approach.
45
Fan P. C. Lai T. H. Hor C. C. Lee M. T. Huang P. Sieghart W. et al (2018). The α6 subunit-containing GABA(A) receptor: a novel drug target for inhibition of trigeminal activation. Neuropharmacology140, 1–13. 10.1016/j.neuropharm.2018.07.017
46
Fehder W. P. Tuluc F. Ho W.-Z. Douglas S. D. (2007). “Macrophages,” in Encyclopedia of stress. Editor FinkG.Second Edition (New York: Academic Press), 634–639.
47
Fernández-Peña C. Viana F. (2013). Targeting TRPM8 for pain relief.
48
Finnerup N. B. Kuner R. Jensen T. S. (2020). Neuropathic pain: from mechanisms to treatment. Physiol. Rev.101, 259–301. 10.1152/physrev.00045.2019
49
Finnerup N. B. Kuner R. Jensen T. S. (2021). Neuropathic pain: from mechanisms to treatment. Physiol. Rev.101 (1), 259–301. 10.1152/physrev.00045.2019
50
Fonfria E. Mattei C. Hill K. Brown J. T. Randall A. Benham C. D. et al (2006). TRPM2 is elevated in the tMCAO stroke model, transcriptionally regulated, and functionally expressed in C13 microglia. J. Recept. Signal Transduct.26 (3), 179–198. 10.1080/10799890600637522
51
Gao G. Wang W. Tadagavadi R. K. Briley N. E. Love M. I. Miller B. A. et al (2014). TRPM2 mediates ischemic kidney injury and oxidant stress through RAC1. J. Clin. investigation124 (11), 4989–5001. 10.1172/JCI76042
52
Gees M. Colsoul B. Nilius B. (2010). The role of transient receptor potential cation channels in Ca2+ signaling. Cold Spring Harb. Perspect. Biol.2 (10), a003962. 10.1101/cshperspect.a003962
53
Gelderblom M. Melzer N. Schattling B. Göb E. Hicking G. Arunachalam P. et al (2014). Transient receptor potential melastatin subfamily member 2 cation channel regulates detrimental immune cell invasion in ischemic stroke. Stroke45 (11), 3395–3402. 10.1161/STROKEAHA.114.005836
54
Giniatullin R. (2020). Ion channels of nociception. Int. J. Mol. Sci.21 (10), 3553.
55
Görlach A. Bertram K. Hudecova S. Krizanova O. (2015). Calcium and ROS: a mutual interplay. Redox Biol.6, 260–271. 10.1016/j.redox.2015.08.010
56
Gosset J. R. Beaumont K. Matsuura T. Winchester W. Attkins N. Glatt S. et al (2017). A cross-species translational pharmacokinetic-pharmacodynamic evaluation of core body temperature reduction by the TRPM8 blocker PF-05105679. Eur. J. Pharm. Sci.109s, S161–s167. 10.1016/j.ejps.2017.06.009
57
Grace P. M. Gaudet A. D. Staikopoulos V. Maier S. F. Hutchinson M. R. Salvemini D. et al (2016). Nitroxidative signaling mechanisms in pathological pain. Trends Neurosci.39 (12), 862–879. 10.1016/j.tins.2016.10.003
58
Gunawardena D. Raju R. Münch G. (2019). Hydrogen peroxide mediates pro-inflammatory cell-to-cell signaling: a new therapeutic target for inflammation?Neural Regen. Res.14 (8), 1430–1437. 10.4103/1673-5374.253529
59
Haraguchi K. Kawamoto A. Isami K. Maeda S. Kusano A. Asakura K. et al (2012). TRPM2 contributes to inflammatory and neuropathic pain through the aggravation of pronociceptive inflammatory responses in mice. J. Neurosci.32 (11), 3931–3941. 10.1523/JNEUROSCI.4703-11.2012
60
Hasan R. Zhang X. (2018). Ca2+ regulation of TRP ion channels. Int. J. Mol. Sci.19 (4), 1256. 10.3390/ijms19041256
61
Haslund-Vinding J. McBean G. Jaquet V. Vilhardt F. (2017). NADPH oxidases in oxidant production by microglia: activating receptors, pharmacology and association with disease. Br. J. Pharmacol.174 (12), 1733–1749. 10.1111/bph.13425
62
Hecquet C. M. Malik A. B. (2009). Role of H2O2-activated TRPM2 calcium channel in oxidant-induced endothelial injury. Thrombosis haemostasis101 (04), 619–625. 10.1160/th08-10-0641
63
Hill K. McNulty S. Randall A. (2004). Inhibition of TRPM2 channels by the antifungal agents clotrimazole and econazole. Naunyn-Schmiedeberg's archives Pharmacol.370, 227–237. 10.1007/s00210-004-0981-y
64
Horne D. B. Biswas K. Brown J. Bartberger M. D. Clarine J. Davis C. D. et al (2018). Discovery of TRPM8 antagonist (S)-6-(((3-Fluoro-4-(trifluoromethoxy)phenyl)(3-fluoropyridin-2-yl)methyl)carbamoyl)nicotinic acid (AMG 333), a clinical candidate for the treatment of migraine. J. Med. Chem.61 (18), 8186–8201. 10.1021/acs.jmedchem.8b00518
65
Hosseini A. M. Majidi J. Baradaran B. Yousefi M. (2015). Toll-like receptors in the pathogenesis of autoimmune diseases. Adv. Pharm. Bull.5 (Suppl. 1), 605–614. 10.15171/apb.2015.082
66
Hu X. Wu L. Liu X. Zhang Y. Xu M. Fang Q. et al (2021). Deficiency of ROS-activated TRPM2 channel protects neurons from cerebral ischemia-reperfusion injury through upregulating autophagy. Oxidative Med. Cell. Longev.2021, 7356266–7356312. 10.1155/2021/7356266
67
Huang P. Qu C. Rao Z. Wu D. Zhao J. (2024). Bidirectional regulation mechanism of TRPM2 channel: role in oxidative stress, inflammation and ischemia-reperfusion injury. Front. Immunol.15, 1391355. 10.3389/fimmu.2024.1391355
68
Huang S. Wang H. Turlova E. Abussaud A. Ji X. Britto L. R. et al (2017). GSK-3β inhibitor TDZD-8 reduces neonatal hypoxic-ischemic brain injury in mice. CNS Neurosci. and Ther.23 (5), 405–415. 10.1111/cns.12683
69
Hutchinson M. R. Zhang Y. Brown K. Coats B. D. Shridhar M. Sholar P. W. et al (2008). Non‐stereoselective reversal of neuropathic pain by naloxone and naltrexone: involvement of toll‐like receptor 4 (TLR4). Eur. J. Neurosci.28 (1), 20–29. 10.1111/j.1460-9568.2008.06321.x
70
Ishii M. Shimizu S. Hara Y. Hagiwara T. Miyazaki A. Mori Y. et al (2006). Intracellular-produced hydroxyl radical mediates H2O2-induced Ca2+ influx and cell death in rat beta-cell line RIN-5F. Cell calcium39 (6), 487–494. 10.1016/j.ceca.2006.01.013
71
Jang Y. Cho P. S. Yang Y. D. Hwang S. W. (2018). Nociceptive roles of TRPM2 ion channel in pathologic pain. Mol. Neurobiol.55, 6589–6600. 10.1007/s12035-017-0862-2
72
Jentsch T. J. (2016). VRACs and other ion channels and transporters in the regulation of cell volume and beyond. Nat. Rev. Mol. Cell Biol.17 (5), 293–307. 10.1038/nrm.2016.29
73
Jia J. Verma S. Nakayama S. Quillinan N. Grafe M. R. Hurn P. D. et al (2011). Sex differences in neuroprotection provided by inhibition of TRPM2 channels following experimental stroke. J. Cereb. Blood Flow and Metabolism31 (11), 2160–2168. 10.1038/jcbfm.2011.77
74
Jiang L.-H. Li X. Syed Mortadza S. A. Lovatt M. Yang W. (2018). The TRPM2 channel nexus from oxidative damage to Alzheimer’s pathologies: an emerging novel intervention target for age-related dementia. Ageing Res. Rev.47, 67–79. 10.1016/j.arr.2018.07.002
75
Kallenborn-Gerhardt W. Schröder K. Geisslinger G. Schmidtko A. (2013). NOXious signaling in pain processing. Pharmacol. and Ther.137 (3), 309–317. 10.1016/j.pharmthera.2012.11.001
76
Kamei J. Aizawa N. Nakagawa T. Kaneko S. Fujimura T. Homma Y. et al (2021). Lacking transient receptor potential melastatin 2 attenuates lipopolysaccharide‐induced bladder inflammation and its associated hypersensitivity in mice. Int. J. Urology28 (1), 107–114. 10.1111/iju.14389
77
Kaneko S. Kawakami S. Hara Y. Wakamori M. Itoh E. Minami T. et al (2006). A critical role of TRPM2 in neuronal cell death by hydrogen peroxide. J. Pharmacol. Sci.101 (1), 66–76. 10.1254/jphs.fp0060128
78
Kaneko Y. Szallasi A. (2014). Transient receptor potential (TRP) channels: a clinical perspective. Br. J. Pharmacol.171 (10), 2474–2507. 10.1111/bph.12414
79
Kassmann M. Harteneck C. Zhu Z. Nürnberg B. Tepel M. Gollasch M. (2013). Transient receptor potential vanilloid 1 (TRPV 1), TRPV 4, and the kidney. Acta physiol.207 (3), 546–564. 10.1111/apha.12051
80
Kawai T. Akira S. (2007). TLR signaling. Semin. Immunol.19 (1), 24–32. 10.1016/j.smim.2006.12.004
81
Kawasaki T. Kawai T. (2014). Toll-like receptor signaling pathways. Front. Immunol.5, 461. 10.3389/fimmu.2014.00461
82
Kerstman E. Ahn S. Battu S. Tariq S. Grabois M. (2013). Neuropathic pain. Handb. Clin. neurology110, 175–187. 10.1016/B978-0-444-52901-5.00015-0
83
Kew J. N. Davies C. H. (2010). Ion channels: from structure to function. Oxford University Press.
84
Khanahmad H. Mirbod S. M. Karimi F. Kharazinejad E. Owjfard M. Najaflu M. et al (2022). Pathological mechanisms induced by TRPM2 ion channels activation in renal ischemia-reperfusion injury. Mol. Biol. Rep.49 (11), 11071–11079. 10.1007/s11033-022-07836-w
85
Kheradpezhouh E. Ma L. Morphett A. Barritt G. J. Rychkov G. Y. (2014). TRPM2 channels mediate acetaminophen-induced liver damage. Proc. Natl. Acad. Sci.111 (8), 3176–3181. 10.1073/pnas.1322657111
86
Klionsky L. Tamir R. Gao B. Wang W. Immke D. C. Nishimura N. et al (2007). Species-specific pharmacology of Trichloro(sulfanyl)ethyl benzamides as transient receptor potential ankyrin 1 (TRPA1) antagonists. Mol. Pain3, 39. 10.1186/1744-8069-3-39
87
Knowles H. Li Y. Perraud A.-L. (2013). The TRPM2 ion channel, an oxidative stress and metabolic sensor regulating innate immunity and inflammation. Immunol. Res.55, 241–248. 10.1007/s12026-012-8373-8
88
Kraft R. Grimm C. Frenzel H. Harteneck C. (2006). Inhibition of TRPM2 cation channels by N‐(p‐amylcinnamoyl) anthranilic acid. Br. J. Pharmacol.148 (3), 264–273. 10.1038/sj.bjp.0706739
89
Kraft R. Grimm C. Grosse K. Hoffmann A. Sauerbruch S. Kettenmann H. et al (2004). Hydrogen peroxide and ADP-ribose induce TRPM2-mediated calcium influx and cation currents in microglia. Am. J. Physiology-Cell Physiology286 (1), C129–C137. 10.1152/ajpcell.00331.2003
90
Kuang X. Huang Y. Gu H. F. Zu X. Y. Zou W. Y. Song Z. B. et al (2012). Effects of intrathecal epigallocatechin gallate, an inhibitor of Toll-like receptor 4, on chronic neuropathic pain in rats. Eur. J. Pharmacol.676 (1-3), 51–56. 10.1016/j.ejphar.2011.11.037
91
Kuffler D. P. (2020). Mechanisms for reducing neuropathic pain. Mol. Neurobiol.57 (1), 67–87. 10.1007/s12035-019-01757-9
92
Kuzmich N. N. Sivak K. V. Chubarev V. N. Porozov Y. B. Savateeva-Lyubimova T. N. Peri F. (2017). TLR4 signaling pathway modulators as potential therapeutics in inflammation and sepsis. Vaccines (Basel)5 (4), 34. 10.3390/vaccines5040034
93
Kwilasz A. J. Clements M. A. Larson T. A. Harris K. M. Litwiler S. T. Woodall B. J. et al (2022). Involvement of TLR2-TLR4, NLRP3, and IL-17 in pain induced by a novel Sprague-Dawley rat model of experimental autoimmune encephalomyelitis. Front. Pain Res. (Lausanne)3, 932530. 10.3389/fpain.2022.932530
94
Lambert G. A. Davis J. B. Appleby J. M. Chizh B. A. Hoskin K. L. Zagami A. S. (2009). The effects of the TRPV1 receptor antagonist SB-705498 on trigeminovascular sensitisation and neurotransmission. Naunyn Schmiedeb. Arch. Pharmacol.380 (4), 311–325. 10.1007/s00210-009-0437-5
95
Lei J. Paul J. Wang Y. Gupta M. Vang D. Thompson S. et al (2021). Heme causes pain in sickle mice via toll-like receptor 4-mediated reactive oxygen species-and endoplasmic reticulum stress-induced glial activation. Antioxidants and Redox Signal.34 (4), 279–293. 10.1089/ars.2019.7913
96
Li W. Wang K. Liu Y. Wu H. He Y. Li C. et al (2022). A novel drug combination of mangiferin and cinnamic acid alleviates rheumatoid arthritis by inhibiting TLR4/nfκb/NLRP3 activation-induced pyroptosis. Front. Immunol.13, 912933. 10.3389/fimmu.2022.912933
97
Li Y. Yin C. Li X. Liu B. Wang J. Zheng X. et al (2019). Electroacupuncture alleviates paclitaxel-induced peripheral neuropathic pain in rats via suppressing TLR4 signaling and TRPV1 upregulation in sensory neurons. Int. J. Mol. Sci.20 (23), 5917. 10.3390/ijms20235917
98
Lim H. Kim D. Lee S. J. (2013). Toll-like receptor 2 mediates peripheral nerve injury-induced NADPH oxidase 2 expression in spinal cord microglia. J. Biol. Chem.288 (11), 7572–7579. 10.1074/jbc.M112.414904
99
Liu Y. Liu W. Wang X. Wan Z. Leng Y. (2018). Dexmedetomidine relieves acute inflammatory visceral pain in rats through the ERK pathway, toll-like receptor signaling, and TRPV1 channel. J. Mol. Neurosci.66 (2), 279–290. 10.1007/s12031-018-1172-5
100
Loeser J. D. Melzack R. (1999). Pain: an overview. lancet353 (9164), 1607–1609. 10.1016/S0140-6736(99)01311-2
101
Lolignier S. Gkika D. Andersson D. Leipold E. Vetter I. Viana F. et al (2016). New insight in cold pain: role of ion channels, modulation, and clinical perspectives. J. Neurosci.36 (45), 11435–11439. 10.1523/JNEUROSCI.2327-16.2016
102
López-Requena A. Boonen B. Van Gerven L. Hellings P. W. Alpizar Y. A. Talavera K. (2017). “Roles of Neuronal TRP Channels in Neuroimmune Interactions,” in Neurobiology of TRP Channels. Editor EmirT. L. R. (CRC Press/Taylor & Francis), 277–294. 10.4324/9781315152837-15
103
Malko P. Jiang L.-H. (2020). TRPM2 channel-mediated cell death: an important mechanism linking oxidative stress-inducing pathological factors to associated pathological conditions. Redox Biol.37, 101755. 10.1016/j.redox.2020.101755
104
Malko P. Syed Mortadza S. A. McWilliam J. Jiang L. H. (2019). TRPM2 channel in microglia as a new player in neuroinflammation associated with a spectrum of central nervous system pathologies. Front. Pharmacol.10, 239. 10.3389/fphar.2019.00239
105
Maqboul A. Elsadek B. (2018). Expression profiles of TRPV1, TRPV4, TLR4 and ERK1/2 in the dorsal root ganglionic neurons of a cancer-induced neuropathy rat model. PeerJ6, e4622. 10.7717/peerj.4622
106
Matsumoto K. Takagi K. Kato A. Ishibashi T. Mori Y. Tashima K. et al (2016). Role of transient receptor potential melastatin 2 (TRPM2) channels in visceral nociception and hypersensitivity. Exp. Neurol.285, 41–50. 10.1016/j.expneurol.2016.09.001
107
McLennan A. (2006). The Nudix hydrolase superfamily. Cell. Mol. Life Sci. CMLS63, 123–143. 10.1007/s00018-005-5386-7
108
Mehta D. Ahmmed G. U. Paria B. C. Holinstat M. Voyno-Yasenetskaya T. Tiruppathi C. et al (2003). RhoA interaction with inositol 1, 4, 5-trisphosphate receptor and transient receptor potential channel-1 regulates Ca2+ entry: role in signaling increased endothelial permeability. J. Biol. Chem.278 (35), 33492–33500. 10.1074/jbc.M302401200
109
Mittal M. Nepal S. Tsukasaki Y. Hecquet C. M. Soni D. Rehman J. et al (2017). Neutrophil activation of endothelial cell-expressed TRPM2 mediates transendothelial neutrophil migration and vascular injury. Circulation Res.121 (9), 1081–1091. 10.1161/CIRCRESAHA.117.311747
110
Miyake T. Shirakawa H. Kusano A. Sakimoto S. Konno M. Nakagawa T. et al (2014). TRPM2 contributes to LPS/IFNγ-induced production of nitric oxide via the p38/JNK pathway in microglia. Biochem. biophysical Res. Commun.444 (2), 212–217. 10.1016/j.bbrc.2014.01.022
111
Mohammadi M. (2019). Role of oxidized phospholipids in inflammatory pain. Universität Würzburg[Doctoral dissertation].
112
Montell C. Rubin G. M. (1989). Molecular characterization of the Drosophila trp locus: a putative integral membrane protein required for phototransduction. Neuron2 (4), 1313–1323. 10.1016/0896-6273(89)90069-x
113
Mortadza S. S. Sim J. A. Stacey M. Jiang L. H. (2017). Signalling mechanisms mediating Zn2+-induced TRPM2 channel activation and cell death in microglial cells. Sci. Rep.7 (1), 45032. 10.1038/srep45032
114
Myers R. R. Campana W. M. Shubayev V. I. (2006). The role of neuroinflammation in neuropathic pain: mechanisms and therapeutic targets. Drug Discov. today11 (1-2), 8–20. 10.1016/S1359-6446(05)03637-8
115
Nazıroğlu M. (2017). Activation of TRPM2 and TRPV1 channels in dorsal root ganglion by NADPH oxidase and protein kinase C molecular pathways: a patch clamp study. J. Mol. Neurosci.61, 425–435. 10.1007/s12031-017-0882-4
116
Nazıroğlu M. Braidy N. (2017). Thermo-sensitive TRP channels: novel targets for treating chemotherapy-induced peripheral pain. Front. Physiology8, 1040. 10.3389/fphys.2017.01040
117
Nazıroğlu M. Çiğ B. Yazğan Y. Schwaerzer G. K. Theilig F. Pecze L. (2019). Albumin evokes Ca2+-induced cell oxidative stress and apoptosis through TRPM2 channel in renal collecting duct cells reduced by curcumin. Sci. Rep.9 (1), 12403. 10.1038/s41598-019-48716-x
118
Nazıroğlu M. Lückhoff A. Jüngling E. (2007). Antagonist effect of flufenamic acid on TRPM2 cation channels activated by hydrogen peroxide. Cell Biochem. Funct.25 (4), 383–387. 10.1002/cbf.1310
119
Nazıroğlu M. Özgül C. Çiğ B. Doğan S. Uğuz A. C. (2011). Glutathione modulates Ca 2+ influx and oxidative toxicity through TRPM2 channel in rat dorsal root ganglion neurons. J. Membr. Biol.242, 109–118. 10.1007/s00232-011-9382-6
120
Naziroğlu M. Uğuz A. C. Ismailoğlu Ö. Çiğ B. Özgül C. Borcak M. (2013). Role of TRPM2 cation channels in dorsal root ganglion of rats after experimental spinal cord injury. Muscle and Nerve48 (6), 945–950. 10.1002/mus.23844
121
Ni H. M. Bockus A. Boggess N. Jaeschke H. Ding W. X. (2012). Activation of autophagy protects against acetaminophen‐induced hepatotoxicity. Hepatology55 (1), 222–232. 10.1002/hep.24690
122
Nicotra L. Loram L. C. Watkins L. R. Hutchinson M. R. (2012). Toll-like receptors in chronic pain. Exp. Neurol.234 (2), 316–329. 10.1016/j.expneurol.2011.09.038
123
Noreen M. Arshad M. (2015). Association of TLR1, TLR2, TLR4, TLR6, and TIRAP polymorphisms with disease susceptibility. Immunol. Res.62 (2), 234–252. 10.1007/s12026-015-8640-6
124
Ogawa N. Kurokawa T. Mori Y. (2016). Sensing of redox status by TRP channels. Cell calcium60 (2), 115–122. 10.1016/j.ceca.2016.02.009
125
Onakpoya I. J. Thomas E. T. Lee J. J. Goldacre B. Heneghan C. J. (2019). Benefits and harms of pregabalin in the management of neuropathic pain: a rapid review and meta-analysis of randomised clinical trials. BMJ Open9 (1), e023600. 10.1136/bmjopen-2018-023600
126
Osmanlioğlu H. Ö. (2022). Ketamine attenuates hypoxia-induced cell death and oxidative toxicity via inhibition of the TRPM2 channel in neuronal cells. J. Cell. Neurosci. Oxidative Stress14 (3), 1095–1104. 10.37212/jcnos.1325007
127
Osmanlıoğlu H. Ö. Yıldırım M. K. Akyuva Y. Yıldızhan K. Nazıroğlu M. (2020). Morphine induces apoptosis, inflammation, and mitochondrial oxidative stress via activation of TRPM2 channel and nitric oxide signaling pathways in the hippocampus. Mol. Neurobiol.57 (8), 3376–3389. 10.1007/s12035-020-01975-6
128
Övey I. Naziroğlu M. (2015). Homocysteine and cytosolic GSH depletion induce apoptosis and oxidative toxicity through cytosolic calcium overload in the hippocampus of aged mice: involvement of TRPM2 and TRPV1 channels. Neuroscience284, 225–233. 10.1016/j.neuroscience.2014.09.078
129
Özdemir Ü. S. Nazıroğlu M. Şenol N. Ghazizadeh V. (2016). Hypericum perforatum attenuates spinal cord injury-induced oxidative stress and apoptosis in the dorsal root ganglion of rats: involvement of TRPM2 and TRPV1 channels. Mol. Neurobiol.53, 3540–3551. 10.1007/s12035-015-9292-1
130
Pasierski M. Szulczyk B. (2022). Beneficial effects of capsaicin in disorders of the central nervous system. Molecules27 (8), 2484. 10.3390/molecules27082484
131
Patel A. R. Ritzel R. McCullough L. D. Liu F. (2013). Microglia and ischemic stroke: a double-edged sword. Int. J. physiology5 (2), 73–90. pathophysiology and pharmacology.
132
Perraud A.-L. Fleig A. Dunn C. A. Bagley L. A. Launay P. Schmitz C. et al (2001). ADP-ribose gating of the calcium-permeable LTRPC2 channel revealed by Nudix motif homology. Nature411 (6837), 595–599. 10.1038/35079100
133
Powers K. A. Szászi K. Khadaroo R. G. Tawadros P. S. Marshall J. C. Kapus A. et al (2006). Oxidative stress generated by hemorrhagic shock recruits Toll-like receptor 4 to the plasma membrane in macrophages. J. Exp. Med.203 (8), 1951–1961. 10.1084/jem.20060943
134
Quan J. Lee J. Y. Choi H. Kim Y. C. Yang S. Jeong J. et al (2022). Effect of pregabalin combined with duloxetine and tramadol on allodynia in chronic postischemic pain and spinal nerve ligation mouse models. Pharmaceutics14 (3), 670. 10.3390/pharmaceutics14030670
135
Raghunatha P. Vosoughi A. Kauppinen T. M. Jackson M. F. (2020). Microglial NMDA receptors drive pro‐inflammatory responses via PARP‐1/TRMP2 signaling. Glia68 (7), 1421–1434. 10.1002/glia.23790
136
Ramachandran R. Wang Z. Saavedra C. DiNardo A. Corr M. Powell S. B. et al (2019). Role of Toll-like receptor 4 signaling in mast cell-mediated migraine pain pathway. Mol. Pain15, 1744806919867842. 10.1177/1744806919867842
137
Rather M. A. Khan A. Wang L. Jahan S. Rehman M. U. Makeen H. A. et al (2023). TRP channels: role in neurodegenerative diseases and therapeutic targets. Heliyon9 (6), e16910. 10.1016/j.heliyon.2023.e16910
138
Ren K. Dubner R. (2010). Interactions between the immune and nervous systems in pain. Nat. Med.16 (11), 1267–1276. 10.1038/nm.2234
139
Round P. Priestley A. Robinson J. (2011). An investigation of the safety and pharmacokinetics of the novel TRPV1 antagonist XEN-D0501 in healthy subjects. Br. J. Clin. Pharmacol.72 (6), 921–931. 10.1111/j.1365-2125.2011.04040.x
140
Roy A. Srivastava M. Saqib U. Liu D. Faisal S. M. Sugathan S. et al (2016). Potential therapeutic targets for inflammation in toll-like receptor 4 (TLR4)-mediated signaling pathways. Int. Immunopharmacol.40, 79–89. 10.1016/j.intimp.2016.08.026
141
Ruck T. Bock S. Pfeuffer S. Schroeter C. B. Cengiz D. Marciniak P. et al (2022). K2P18. 1 translates T cell receptor signals into thymic regulatory T cell development. Cell Res.32 (1), 72–88. 10.1038/s41422-021-00580-z
142
Sahoo B. R. (2020). Structure of fish Toll-like receptors (TLR) and NOD-like receptors (NLR). Int. J. Biol. Macromol.161, 1602–1617. 10.1016/j.ijbiomac.2020.07.293
143
Samanta A. Hughes T. E. Moiseenkova-Bell V. Y. (2018). Transient receptor potential (TRP) channels. Membr. protein complexes Struct. Funct.87, 141–165. 10.1007/978-981-10-7757-9_6
144
Santoni G. Cardinali C. Morelli M. B. Santoni M. Nabissi M. Amantini C. (2015). Danger-and pathogen-associated molecular patterns recognition by pattern-recognition receptors and ion channels of the transient receptor potential family triggers the inflammasome activation in immune cells and sensory neurons. J. neuroinflammation12, 21–10. 10.1186/s12974-015-0239-2
145
Santos-Silva A. Charrua A. Cruz C. D. Gharat L. Avelino A. Cruz F. (2012). Rat detrusor overactivity induced by chronic spinalization can be abolished by a transient receptor potential vanilloid 1 (TRPV1) antagonist. Auton. Neurosci.166 (1-2), 35–38. 10.1016/j.autneu.2011.09.005
146
Schappe M. S. Szteyn K. Stremska M. E. Mendu S. K. Downs T. K. Seegren P. V. et al (2018). Chanzyme TRPM7 mediates the Ca2+ influx essential for lipopolysaccharide-induced toll-like receptor 4 endocytosis and macrophage activation. Immunity48 (1), 59–74. 10.1016/j.immuni.2017.11.026
147
Scholz J. Finnerup N. B. Attal N. Aziz Q. Baron R. Bennett M. I. et al (2019). The IASP classification of chronic pain for ICD-11: chronic neuropathic pain. Pain160 (1), 53–59. 10.1097/j.pain.0000000000001365
148
Schwab A. Fabian A. Hanley P. J. Stock C. (2012). Role of ion channels and transporters in cell migration. Physiol. Rev.92, 1865–1913. 10.1152/physrev.00018.2011
149
Sela R. A. Bruera E. Conner-spady B. Cumming C. Walker C. (2002). Sensory and affective dimensions of advanced cancer pain. Psycho‐Oncology J. Psychol. Soc. Behav. Dimensions Cancer11 (1), 23–34. 10.1002/pon.551
150
Shi R. Fu Y. Zhao D. Boczek T. Wang W. Guo F. (2021). Cell death modulation by transient receptor potential melastatin channels TRPM2 and TRPM7 and their underlying molecular mechanisms. Biochem. Pharmacol.190, 114664. 10.1016/j.bcp.2021.114664
151
Shim Y. J. Tae Y. K. Kang B. H. Park J. S. Jeon S. Y. Min B. H. (2017). Toll-like receptor 4 signaling is required for clusterin-induced tumor necrosis factor-α secretion in macrophage. Biochem. Biophys. Res. Commun.482 (4), 1407–1412. 10.1016/j.bbrc.2016.12.049
152
Shirakawa H. Kaneko S. (2018). Physiological and pathophysiological roles of transient receptor potential channels in microglia-related CNS inflammatory diseases. Biol. Pharm. Bull.41 (8), 1152–1157. 10.1248/bpb.b18-00319
153
Simon F. Tapia P. Armisen R. Echeverria C. Gatica S. Vallejos A. et al (2017). Human peritoneal mesothelial cell death induced by high-glucose hypertonic solution involves Ca2+ and Na+ ions and oxidative stress with the participation of PKC/NOX2 and PI3K/Akt pathways. Front. physiology8, 379. 10.3389/fphys.2017.00379
154
So K. Haraguchi K. Asakura K. Isami K. Sakimoto S. Shirakawa H. et al (2015). Involvement of TRPM2 in a wide range of inflammatory and neuropathic pain mouse models. J. Pharmacol. Sci.127 (3), 237–243. 10.1016/j.jphs.2014.10.003
155
Song K. Wang H. Kamm G. B. Pohle J. Reis F. d. C. Heppenstall P. et al (2016). The TRPM2 channel is a hypothalamic heat sensor that limits fever and can drive hypothermia. Science353 (6306), 1393–1398. 10.1126/science.aaf7537
156
Song Z. Wu T. Sun J. Wang H. Hua F. Nicolas Y. S. M. et al (2021). Metformin attenuates post-epidural fibrosis by inhibiting the TGF-β1/Smad3 and HMGB1/TLR4 signaling pathways. J. Cell Mol. Med.25 (7), 3272–3283. 10.1111/jcmm.16398
157
Staurengo-Ferrari L. Deng L. Chiu I. M. (2022). Interactions between nociceptor sensory neurons and microbial pathogens in pain. Pain163 (Suppl. 1), S57–s68. 10.1097/j.pain.0000000000002721
158
Stokes J. A. Corr M. Yaksh T. L. (2013). Spinal toll-like receptor signaling and nociceptive processing: regulatory balance between TIRAP and TRIF cascades mediated by TNF and IFNβ. PAIN®154 (5), 733–742. 10.1016/j.pain.2013.01.012
159
Sumoza-Toledo A. Lange I. Cortado H. Bhagat H. Mori Y. Fleig A. et al (2011). Dendritic cell maturation and chemotaxis is regulated by TRPM2-mediated lysosomal Ca2+ release. FASEB J.25 (10), 3529–3542. 10.1096/fj.10-178483
160
Sumoza-Toledo A. Penner R. (2011). TRPM2: a multifunctional ion channel for calcium signalling. J. physiology589 (7), 1515–1525. 10.1113/jphysiol.2010.201855
161
Sun M.-K. Alkon D. L. (2014). The “memory kinases”: roles of PKC isoforms in signal processing and memory formation. Prog. Mol. Biol. Transl. Sci.122, 31–59. 10.1016/B978-0-12-420170-5.00002-7
162
Świeboda P. Filip R. Prystupa A. Drozd M. (2013). Assessment of pain: types, mechanism and treatment. Pain2 (7), 2–7.
163
Syed Mortadza S. A. Wang L. Li D. Jiang L. H. (2015). TRPM2 channel-mediated ROS-sensitive Ca2+ signaling mechanisms in immune cells. Front. Immunol.6, 407. 10.3389/fimmu.2015.00407
164
Szallasi A. (2023). Resiniferatoxin: nature's precision medicine to silence TRPV1-positive afferents. Int. J. Mol. Sci.24 (20), 15042. 10.3390/ijms242015042
165
Takeda K. Kaisho T. Akira S. (2003). Toll-like receptors. Annu. Rev. Immunol.21 (1), 335–376. 10.1146/annurev.immunol.21.120601.141126
166
Tan C.-H. McNaughton P. A. (2018). TRPM2 and warmth sensation. Pflügers Archiv-European J. Physiology470 (5), 787–798. 10.1007/s00424-018-2139-7
167
Tanga F. Y. Nutile-McMenemy N. DeLeo J. A. (2005). The CNS role of Toll-like receptor 4 in innate neuroimmunity and painful neuropathy. Proc. Natl. Acad. Sci.102 (16), 5856–5861. 10.1073/pnas.0501634102
168
Tauseef M. Knezevic N. Chava K. R. Smith M. Sukriti S. Gianaris N. et al (2012). TLR4 activation of TRPC6-dependent calcium signaling mediates endotoxin-induced lung vascular permeability and inflammation. J. Exp. Med.209 (11), 1953–1968. 10.1084/jem.20111355
169
Tegtmeyer N. Neddermann M. Lind J. Pachathundikandi S. K. Sharafutdinov I. Gutiérrez-Escobar A. J. et al (2020). Toll-like receptor 5 activation by the CagY repeat domains of Helicobacter pylori. Cell Rep.32 (11), 108159. 10.1016/j.celrep.2020.108159
170
Teixeira-Santos L. Albino-Teixeira A. Pinho D. (2020). Neuroinflammation, oxidative stress and their interplay in neuropathic pain: focus on specialized pro-resolving mediators and NADPH oxidase inhibitors as potential therapeutic strategies. Pharmacol. Res.162, 105280. 10.1016/j.phrs.2020.105280
171
Togashi K. Hara Y. Tominaga T. Higashi T. Konishi Y. Mori Y. et al (2006). TRPM2 activation by cyclic ADP‐ribose at body temperature is involved in insulin secretion. EMBO J.25 (9), 1804–1815. 10.1038/sj.emboj.7601083
172
Togashi K. Inada H. Tominaga M. (2008). Inhibition of the transient receptor potential cation channel TRPM2 by 2‐aminoethoxydiphenyl borate (2‐APB). Br. J. Pharmacol.153 (6), 1324–1330. 10.1038/sj.bjp.0707675
173
Tsagareli M. G. Nozadze I. (2020). An overview on transient receptor potential channels superfamily. Behav. Pharmacol.31 (5), 413–434. 10.1097/FBP.0000000000000524
174
Tsutsui M. Hirase R. Miyamura S. Nagayasu K. Nakagawa T. Mori Y. et al (2018). TRPM2 exacerbates central nervous system inflammation in experimental autoimmune encephalomyelitis by increasing production of CXCL2 chemokines. J. Neurosci.38 (39), 8484–8495. 10.1523/JNEUROSCI.2203-17.2018
175
Turlova E. Feng Z.-p. Sun H.-s. (2018). The role of TRPM2 channels in neurons, glial cells and the blood-brain barrier in cerebral ischemia and hypoxia. Acta Pharmacol. Sin.39 (5), 713–721. 10.1038/aps.2017.194
176
Vandewauw I. De Clercq K. Mulier M. Held K. Pinto S. Van Ranst N. et al (2018). A TRP channel trio mediates acute noxious heat sensing. Nature555 (7698), 662–666. 10.1038/nature26137
177
Vaure C. Liu Y. (2014). A comparative review of toll-like receptor 4 expression and functionality in different animal species. Front. Immunol.5, 316. 10.3389/fimmu.2014.00316
178
Vermot A. Petit-Härtlein I. Smith S. M. E. Fieschi F. (2021). NADPH oxidases (NOX): an overview from discovery, molecular mechanisms to physiology and pathology. Antioxidants (Basel)10 (6), 890. 10.3390/antiox10060890
179
Vicario N. Turnaturi R. Spitale F. M. Torrisi F. Zappalà A. Gulino R. et al (2020). Intercellular communication and ion channels in neuropathic pain chronicization. Inflamm. Res.69, 841–850. 10.1007/s00011-020-01363-9
180
Vilar B. Tan C.-H. McNaughton P. A. (2020). Heat detection by the TRPM2 ion channel. Nature584 (7820), E5–E12. 10.1038/s41586-020-2510-7
181
Vriens J. Owsianik G. Hofmann T. Philipp S. E. Stab J. Chen X. et al (2011). TRPM3 is a nociceptor channel involved in the detection of noxious heat. Neuron70 (3), 482–494. 10.1016/j.neuron.2011.02.051
182
Wang H. Song T. Wang W. Zhang Z. (2019b). TRPM2 participates the transformation of acute pain to chronic pain during injury‐induced neuropathic pain. Synapse73 (10), e22117. 10.1002/syn.22117
183
Wang Q. Guo W. Hao B. Shi X. Lu Y. Wong C. W. M. et al (2016). Mechanistic study of TRPM2-Ca2+-CAMK2-BECN1 signaling in oxidative stress-induced autophagy inhibition. Autophagy12 (8), 1340–1354. 10.1080/15548627.2016.1187365
184
Wang Y. Chen L. Wang K. Da Y. Zhou M. Yan H. et al (2019a). Suppression of TRPM2 reduces renal fibrosis and inflammation through blocking TGF-β1-regulated JNK activation. Biomed. and Pharmacother.120, 109556. 10.1016/j.biopha.2019.109556
185
Wehrhahn J. Kraft R. Harteneck C. Hauschildt S. (2010). Transient receptor potential melastatin 2 is required for lipopolysaccharide-induced cytokine production in human monocytes. J. Immunol.184 (5), 2386–2393. 10.4049/jimmunol.0902474
186
Woller S. A. Eddinger K. A. Corr M. Yaksh T. L. (2017). An overview of pathways encoding nociception. Clin. Exp. rheumatology35 (Suppl. 107), 40–46.
187
Wu F.-x. Bian J. j. Miao X. r. Huang S. d. Xu X. w. Gong D. j. et al (2010). Intrathecal siRNA against Toll-like receptor 4 reduces nociception in a rat model of neuropathic pain. Int. J. Med. Sci.7 (5), 251–259. 10.7150/ijms.7.251
188
Yamamoto S. Shimizu S. (2016). Targeting TRPM2 in ROS-coupled diseases. Pharmaceuticals9 (3), 57. 10.3390/ph9030057
189
Yang K. Chang W. L. Yang P. C. Chien C. L. Lai M. S. Su M. J. et al (2006). Activation of the transient receptor potential M2 channel and poly (ADP-ribose) polymerase is involved in oxidative stress-induced cardiomyocyte death. Cell Death and Differ.13 (10), 1815–1826. 10.1038/sj.cdd.4401813
190
Yang Y. Yao Z. Wang H. Jia S. Wang M. Wang S. et al (2024). Severe inflammation in C57/BL6 mice leads to prolonged cognitive impairment by initiating the IL-1β/TRPM2 pathway. Int. Immunopharmacol.128, 111380. 10.1016/j.intimp.2023.111380
191
Ye M. Yang W. Ainscough J. F. Hu X. P. Li X. Sedo A. et al (2014). TRPM2 channel deficiency prevents delayed cytosolic Zn2+ accumulation and CA1 pyramidal neuronal death after transient global ischemia. Cell death and Dis.5 (11), e1541. 10.1038/cddis.2014.494
192
Yıldızhan K. Nazıroğlu M. (2020). Glutathione depletion and parkinsonian neurotoxin MPP+-induced TRPM2 channel activation play central roles in oxidative cytotoxicity and inflammation in microglia. Mol. Neurobiol.57 (8), 3508–3525. 10.1007/s12035-020-01974-7
193
Yu B. Jin L. Yao X. Zhang Y. Zhang G. Wang F. et al (2023). TRPM2 protects against cisplatin-induced acute kidney injury and mitochondrial dysfunction via modulating autophagy. Theranostics13 (13), 4356–4375. 10.7150/thno.84655
194
Yüksel E. Nazıroğlu M. Şahin M. Çiğ B. (2017). Involvement of TRPM2 and TRPV1 channels on hyperalgesia, apoptosis and oxidative stress in rat fibromyalgia model: protective role of selenium. Sci. Rep.7 (1), 17543. 10.1038/s41598-017-17715-1
195
Zhang C. Huang Y. Ouyang F. Su M. Li W. Chen J. et al (2022). Extracellular vesicles derived from mesenchymal stem cells alleviate neuroinflammation and mechanical allodynia in interstitial cystitis rats by inhibiting NLRP3 inflammasome activation. J. Neuroinflammation19 (1), 80. 10.1186/s12974-022-02445-7
196
Zhang D. Li X. Jing B. Chen Z. Shi H. Zheng Y. et al (2023). α-Asarone attenuates chronic sciatica by inhibiting peripheral sensitization and promoting neural repair. Phytother. Res.37 (1), 151–162. 10.1002/ptr.7603
197
Zhang X. Hu M. Yang Y. Xu H. (2018). Organellar TRP channels. Nat. Struct. and Mol. Biol.25 (11), 1009–1018. 10.1038/s41594-018-0148-z
198
Zheng Y. Zhao J. Chang S. Zhuang Z. Waimei S. Li X. et al (2023). β-Sitosterol alleviates neuropathic pain by affect microglia polarization through inhibiting TLR4/NF-κB signaling pathway. J. Neuroimmune Pharmacol.18 (4), 690–703. 10.1007/s11481-023-10091-w
199
Zhong C. Yang J. Zhang Y. Fan X. Fan Y. Hua N. et al (2023). TRPM2 mediates hepatic ischemia–reperfusion injury via Ca2+-induced mitochondrial lipid peroxidation through increasing ALOX12 expression. Research6, 0159. 10.34133/research.0159
200
Zhong Z. Zhai Y. Liang S. Mori Y. Han R. Sutterwala F. S. et al (2013). TRPM2 links oxidative stress to NLRP3 inflammasome activation. Nat. Commun.4 (1), 1611. 10.1038/ncomms2608
201
Zhou Y. H. Liao S. J. Li D. Luo J. Wei J. J. Yan B. et al (2013). TLR4 ligand/H₂O₂ enhances TGF-β1 signaling to induce metastatic potential of non-invasive breast cancer cells by activating non-Smad pathways. PLoS One8 (5), e65906. 10.1371/journal.pone.0065906
202
Zhu T. Zhao Y. Hu H. Zheng Q. Luo X. Ling Y. et al (2019). TRPM2 channel regulates cytokines production in astrocytes and aggravates brain disorder during lipopolysaccharide-induced endotoxin sepsis. Int. Immunopharmacol.75, 105836. 10.1016/j.intimp.2019.105836
203
Zong P. Feng J. Yue Z. Yu A. S. Vacher J. Jellison E. R. et al (2022). TRPM2 deficiency in mice protects against atherosclerosis by inhibiting TRPM2–CD36 inflammatory axis in macrophages. Nat. Cardiovasc. Res.1 (4), 344–360. 10.1038/s44161-022-00027-7
204
Zou J. Ainscough J. F. Yang W. Sedo A. Yu S. P. Mei Z. Z. et al (2013). A differential role of macrophage TRPM2 channels in Ca²⁺ signaling and cell death in early responses to H₂O₂. Am. J. Physiology-Cell Physiology305 (1), C61–C69. 10.1152/ajpcell.00390.2012
Summary
Keywords
inflammation, neuropathic pain, lipopolysaccharide reactive oxygen species (ROS), nicotinamide adenine dinucleotide phosphate (NADPH) oxidase (NOX), TRPM2 and TLR4
Citation
Mandlem VKK, Rivera A, Khan Z, Quazi SH and Deba F (2024) TLR4 induced TRPM2 mediated neuropathic pain. Front. Pharmacol. 15:1472771. doi: 10.3389/fphar.2024.1472771
Received
30 July 2024
Accepted
02 September 2024
Published
12 September 2024
Volume
15 - 2024
Edited by
Sara Marchesan Oliveira, Federal University of Santa Maria, Brazil
Reviewed by
Cassia Regina Silva, Federal University of Uberlandia, Brazil
Evelyne Silva Brum, Federal University of Rio Grande do Sul, Brazil
Updates
Copyright
© 2024 Mandlem, Rivera, Khan, Quazi and Deba.
This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Farah Deba, fdeba@uttyler.edu
Disclaimer
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.