Abstract
Background:
Multiple myeloma (MM) is a malignant disease characterized by the clonal proliferation of plasma cells in the bone marrow. Currently incurable, relapse and drug resistance remain significant challenges, necessitating the exploration of novel anti-MM agents. Ferroptosis, a form of cell death driven by iron-dependent lipid peroxidation, has emerged as a critical player in MM pathology and treatment. With advancing research, emerging evidence links ferroptosis to MM pathogenesis and therapeutic strategies. Natural products (NPs) and certain antitumor agents, owing to their broad bioactivities, demonstrate unique pharmacological advantages in MM therapy by targeting ferroptosis-related pathways.
Purpose:
This review systematically examines ferroptosis-related pathways in MM pathogenesis, focusing on pharmacological and toxicological mechanisms of natural products (NPs) and antitumor compounds in modulating ferroptosis-related pathways. It aims to provide novel insights and strategies for MM research and clinical therapy.
Methods:
We systematically retrieved data from PubMed, Web of Science, ScienceDirect, SciFinder, Scopus, and the China National Knowledge Infrastructure (CNKI) spanning database inception to March 2025, followed by taxonomic integrative analysis of NPs’ and antitumor compounds’ echanistic classifications.
Results:
NPs and antitumor compounds exert anti-MM effects via ferroptosis modulation, mechanistically mediated through: 1) lipid metabolism reprogramming; 2) ferritinophagy-driven iron homeostasis regulation; 3) Reactive oxygen species (ROS)-mediated oxidative stress potentiation; 4) autophagic activation; 5) Genes and proteins regulation.
Conclusion:
NPs and antitumor compounds demonstrate therapeutic potential against MM through multi-dimensional ferroptosis modulation, yet clinical translation faces two critical hurdles: 1) predominant focus on single-target mechanisms lacking systems pharmacology-level network analysis; 2) overreliance on in vitro models with insufficient clinical validation. Prioritize developing biomarkers and ferroptosis inducers to advance novel ferroptosis-targeting anticancer compounds.
1 Introduction
Multiple myeloma (MM), a hematologic malignancy characterized by clonal proliferation of bone marrow plasma cells, manifests clinically through CRAB criteria: hypercalcemia, renal dysfunction, anemia, and osteolytic lesions (van de Donk et al., 2021; Malard et al., 2024). Despite significant improvements in patient prognosis over recent decades through therapeutic advances—including proteasome inhibitors (PIs), immunomodulatory drugs (IMiDs), monoclonal antibodies, bispecific antibodies, and chimeric antigen receptor T-cell immunotherapy (CAR-T) (Rajkumar and Kumar, 2020), the disease remains incurable due to inevitable relapse and drug resistance during maintenance therapy. This underscores an urgent need for novel therapeutic targets and modalities.
Ferroptosis, an iron-dependent non-apoptotic cell death driven by excessive lipid peroxidation, was first characterized by Dixon et al., in 2012 through studies on erastin and RAS-selective lethal compounds (Dixon et al., 2012). Emerging as a pivotal research focus, ferroptosis has been implicated in diverse pathologies including hepatic disorders, breast cancer, cardiovascular diseases, and neurodegenerative conditions (Mahoney-Sánchez et al., 2021; Zhang et al., 2022; Cui et al., 2023; Yang F. et al., 2023), while demonstrating therapeutic potential in oncology (Liang et al., 2019; Lei et al., 2022). In MM pathogenesis, accumulating evidence links disease progression to dysregulated ferroptosis signaling pathways, suggesting opportunities for targeted drug development (Zhong et al., 2020; Li W. et al., 2022; Zhang et al., 2023). Furthermore, ferroptosis-related genes (FRGs) show promise as prognostic biomarkers for MM patients (Fu et al., 2022; Qin et al., 2022; Gao et al., 2023; Wang Q. et al., 2023).
This review systematically elucidates the molecular mechanisms of ferroptosis, its interplay with MM pathogenesis, and pharmacologic strategies leveraging ferroptosis modulation for MM intervention. Our synthesis aims to inform mechanistic investigations and advance clinical translation of ferroptosis-targeted therapies (Figure 1).
FIGURE 1
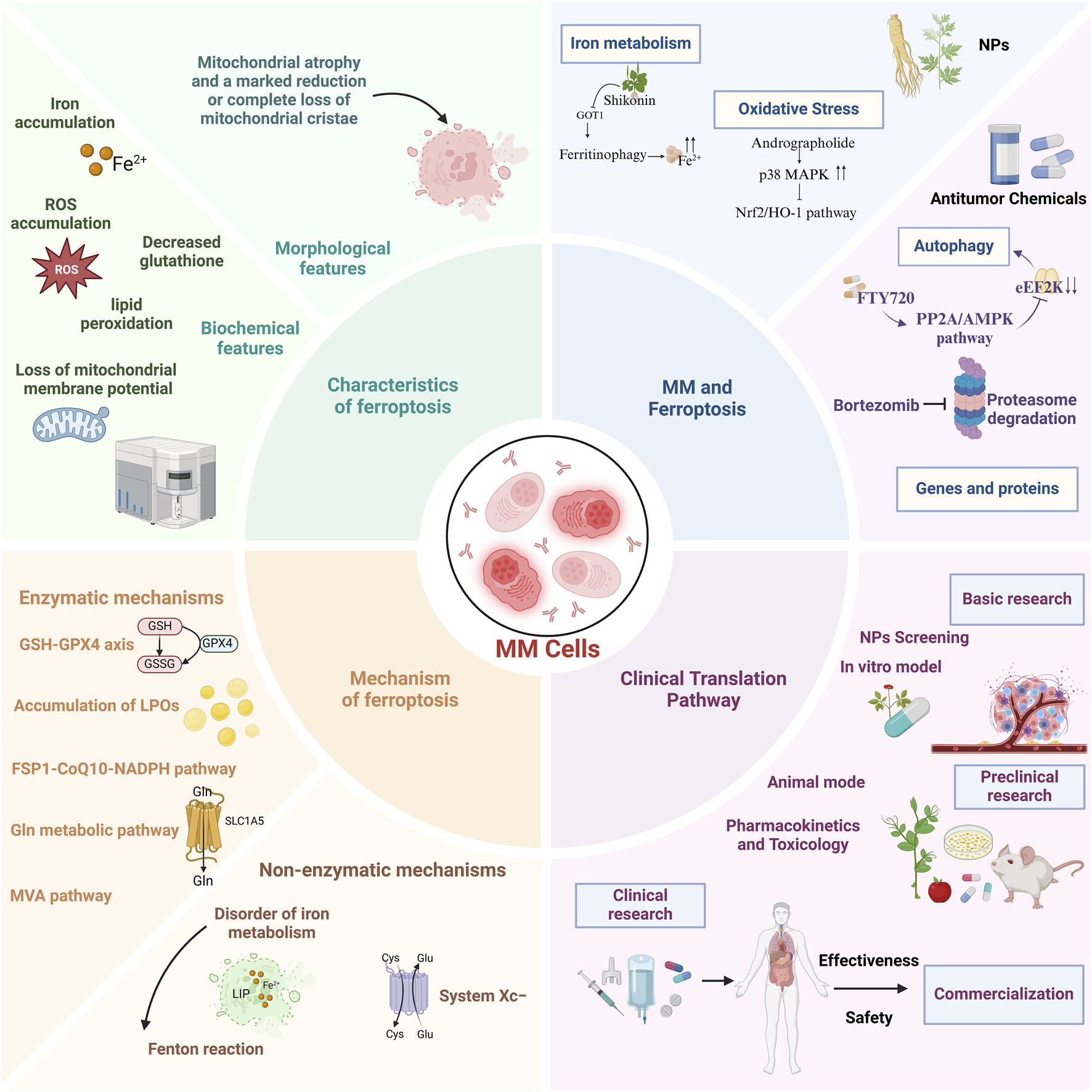
A comprehensive overview of the characteristics, mechanisms, role of ferroptosis in multiple myeloma, and potential clinical translational pathways.
2 Review methodology
In accordance with the Preferred Reporting Items for Systematic Reviews and Meta - Analyses (PRISMA) guidelines, we conducted a systematic review to explore the regulatory role of ferroptosis in MM. Data were collected from research articles published up to March 2025, sourced from eight well - known databases: PubMed, Web of Science, ScienceDirect, Google Scholar, SciFinder, ResearchGate, Scopus, and the China National Knowledge Infrastructure (CNKI). The search terms used were “Multiple myeloma,” “Ferroptosis,” “Natural products,” “Molecular mechanisms,” and “Antitumor effects.” English - language publications, including scientific research, clinical trials, reviews, and meta - analyses, were considered for inclusion. Since this analysis did not involve human or animal subjects, ethical committee approval was not required. After excluding case reports and letters, and subsequent review of abstracts and full - texts, a total of 219 articles were included for evaluation in this manuscript.
3 Overview of ferroptosis
3.1 Evolutionary insights into ferroptosis
The conceptual foundation of ferroptosis was laid in 2003 when Dolma et al. identified erastin as a novel compound selectively lethal to RAS-mutated BJeLR fibroblasts through rapid, irreversible cytotoxicity (Dolma et al., 2003). Subsequent studies revealed the iron-dependent non-apoptotic death mechanism of RAS-mutated cancer cells induced by small molecules RSL3 and RSL5 (Yang and Stockwell, 2008). The term “ferroptosis” was formally proposed in 2012, with mechanistic studies demonstrating erastin’s dual action in HT-1080 fibrosarcoma models: mitochondrial voltage-dependent anion channel (VDAC) modulation and inhibition of the Xc system subunit solute carrier family 7 member 11 (SLC7A11), leading to glutathione depletion and iron-dependent lipid ROS accumulation (Dixon et al., 2012).
A pivotal 2014 study established glutathione peroxidase 4 (GPX4) as the central regulator of ferroptosis, with therapeutic implications validated in tumor xenografts (Yang et al., 2014). Concurrently, p53 was shown to enhance ferroptosis sensitivity by suppressing SLC7A11-mediated cysteine (Cys) uptake, while its acetylation-deficient mutant (p53 3 KR) retained regulatory capacity over ferroptosis under oxidative stress (Jiang et al., 2015). In 2017, acyl-CoA synthetase long-chain family member 4 (ACSL4) was identified as essential for ferroptosis execution by enriching ω6 polyunsaturated fatty acid (PUFA)-containing membranes (Doll et al., 2017). Subsequent work in 2019 uncovered an independent glutathione-bypassing ferroptosis suppression axis mediated by ferroptosis suppressor protein 1 (FSP1) via Coenzyme Q10 (CoQ10) (Doll et al., 2019).
Recent advances have uncovered novel regulatory networks in ferroptosis. The nuclear factor erythroid-2 related factor 2 (Nrf2)-HECT and RLD domain containing E3 ubiquitin protein ligase 2 (HERC2) axis has been shown to modulate cellular iron homeostasis and ferroptosis sensitivity through vesicle associated membrane protein 8 (VAMP8) regulation (Anandhan et al., 2023). Pharmacological targeting of emopamil binding protein (EBP) to block 7-dehydrocholesterol (7-DHC) biosynthesis demonstrates potent ferroptosis induction and antitumor efficacy, with pathway components including cytochrome p450 family 51 subfamily A member 1 (CYP51A1), EBP, and sterol C5-desaturase (SC5D) emerging as potential ferroptosis inhibitors (Freitas et al., 2024; Li Y. et al., 2024). These discoveries have propelled ferroptosis research from basic biological characterization to therapeutic exploration in diverse pathologies including oncology, neurodegenerative disorders, and ischemia-reperfusion injury, offering innovative perspectives for disease mechanism elucidation and intervention strategies (Chen X. et al., 2021; Ju et al., 2023; Wang Y. et al., 2023).
3.2 Morphological characteristics of ferroptotic cells
Ferroptosis manifests distinct morphological alterations that differentiate it from other cell death modalities. Morphologically, ferroptotic cells exhibit cytoplasmic shrinkage, contrasting with apoptotic cell contraction through its association with intracellular lipid peroxide accumulation rather than membrane blebbing. Crucially, these cells preserve plasma membrane integrity and nuclear architecture, lacking characteristic apoptotic features such as chromatin margination, plasma membrane blebbing, or apoptotic body formation (Yagoda et al., 2007; Dixon et al., 2012; Yan H. et al., 2021), unlike autophagic cell death, they show neither double-membrane autophagic vesicles nor compromised membrane integrity (Liang et al., 2019). The pathognomonic ultrastructural features include mitochondrial atrophy with reduced cristae density and increased membrane condensation (Dixon et al., 2012; Stockwell et al., 2017) (Table 1; Figure 2).
TABLE 1
| RCD Types | Morphological features | Biochemical features | Inducing factors |
|---|---|---|---|
| Ferroptosis | Mitochondrial atrophy, accompanied by enhanced double-membrane density, disruption of the outer membrane, and a marked reduction or complete loss of mitochondrial cristaes (Yan et al., 2021b) | Iron accumulation, lipid peroxidation, ROS accumulation, glutathione depletion, increased NADPH oxidation, and loss of mitochondrial membrane potential (Li et al., 2020a; Liu et al., 2022b) | Disruptions in iron metabolism, the accumulation of lipid peroxides, and associated factors |
| Apoptosis | Vacuolization of the plasma membrane, accompanied by nuclear rupture and folding, chromatin condensation, cellular shrinkage, the formation of apoptotic bodies, and pronounced chromatin compaction (VanHook, 2023) | Caspase activation and DNA fragmentation (Chen et al., 2018; Li et al., 2020a) | Regulation of relevant genes under non-pathological conditions |
| Autophagy | The appearance of autophagic vacuoles, which progressively enclose to form autophagosomes and ultimately develop into autolysosomes (Zhao et al., 2024) | LC3-I is converted into LC3-II, and autophagic substrates are degraded (Chen et al., 2018; Li et al., 2020a) | Organelle damage, nutrient deficiency, metabolic disorders, microbial infections, and others |
| Necrosis | The nucleus undergoes condensation, fragmentation, and dissolution; the cell membrane ruptures, accompanied by swelling of the cytoplasm and organelles, as well as chromatin condensation (Kuroe et al., 2021) | ATP levels decline, accompanied by the activation of RIP1, RIP3, and MLKL (Chen et al., 2018; Li et al., 2020a) | Severe pathological damage |
Hallmarks of ferroptosis and other regulated cell death (RCD) modalities: morphological, biochemical, and inducing factors.
NADPH, Nicotinamide adenine dinucleotide phosphate; LC3-I, Microtubule-associated proteins light chain 3-I; LC3-II, Microtubule-associated proteins light chain 3-II; RIP1, Receptor-interacting protein 1; RIP3, Receptor-interacting protein 3; MLKL, Mixed-lineage kinase domain-like.
FIGURE 2
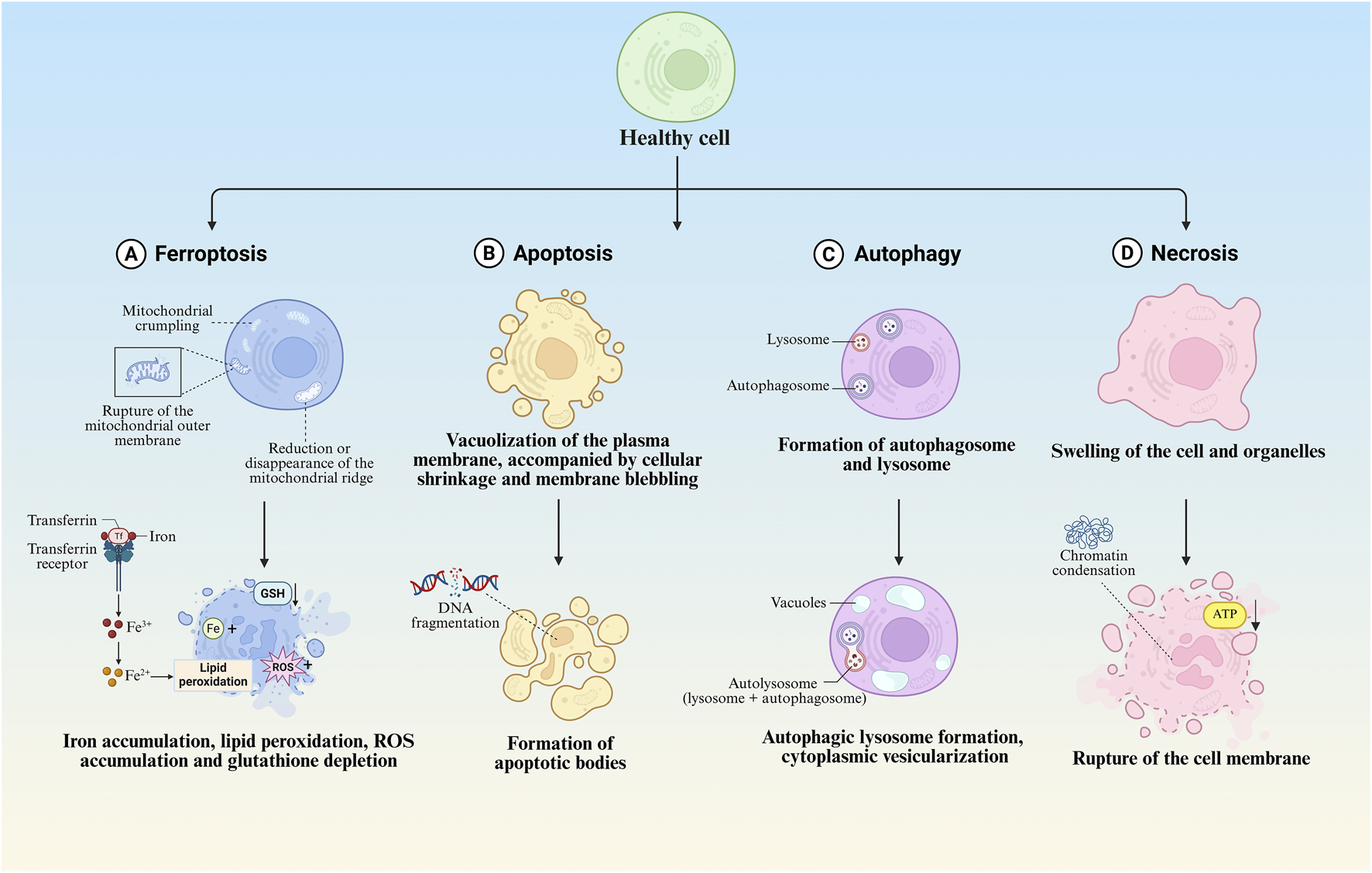
Morphological features of different regulated cell death modalities. (A) Morphological and biochemical characteristics of ferroptosis. (B) Morphological and biochemical characteristics of apoptosis. (C) Morphological and biochemical characteristics of autophagy. (D) Morphological and biochemical characteristics of necrosis.
4 Mechanism of ferroptosis
As an emerging cell death modality, ferroptosis has evolved from the convergence of research in amino acid/lipid metabolism, iron homeostasis, redox biology, selenium regulation, and programmed cell death. Its execution involves a dual-pathway mechanism: enzymatic peroxidation mediated by arachidonate lipoxygenases (ALOXs) and non-enzymatic Fenton reactions. These processes generate cytotoxic phospholipid hydroperoxides that induce membrane damage, ultimately driving ferroptotic cell death.
4.1 Enzymatic mechanisms
4.1.1 The glutathione (GSH)-GPX4 axis
Ferroptosis is characterized by the hallmark feature of intracellular lipid peroxide accumulation, with the glutathione-glutathione peroxidase 4 (GSH-GPX4) axis playing a pivotal role in suppressing lipid peroxidation. GPX4 represents the sole enzymatic system capable of reducing lipid hydroperoxides in mammalian cells, requiring reduced GSH as an essential cofactor for neutralizing lipid peroxides (LPOs). Depletion of GSH or inactivation of GPX4 disrupts this equilibrium, leading to lipid peroxide accumulation and subsequent ferroptosis induction.
Under physiological conditions, cellular homeostasis is maintained through a delicate balance between pro-oxidative systems (comprising iron ions and reactive oxygen species) and antioxidant defenses mediated by the System Xc-/GSH/GPX4 axis. This equilibrium becomes disrupted under pathological states, triggering ferroptotic cell death. As the fourth selenoprotein in the glutathione peroxidase family, GPX4 (originally termed PHGPX for its phospholipid hydroperoxide-reducing capacity) serves as the unique enzymatic regulator of lipid peroxidation through its distinctive structural and functional properties (Xie et al., 2023). The enzyme exerts its protective effects by catalyzing the conversion of phospholipid hydroperoxides (PLOOH) into non-reactive fatty alcohols (PLOH), thereby preventing oxidative membrane damage (Nishida Xavier da Silva et al., 2022), this selenium-dependent peroxidase specifically utilizes reduced GSH to maintain membrane integrity through two critical reactions: reduction of lipid hydroperoxides to corresponding alcohols, and detoxification of hydrogen peroxide to water (Ma et al., 2022; Zhang W. et al., 2024). The catalytic cycle of GPX4 involves dynamic selenocysteine redox transitions. The active site selenol becomes oxidized to selenenic acid during peroxide reduction, subsequently regenerated through GSH-mediated reduction that produces oxidative glutathione (GSSG). This mechanism establishes GSH as an indispensable factor for sustaining GPX4 enzymatic activity (Forcina and Dixon, 2019; Yan H. et al., 2021; Tuncer and Hacioglu, 2024).
4.1.2 Accumulation of LPOs
Ferroptosis represents a distinct form of cell death mechanistically linked to dysregulated lipid metabolism, where the pathological accumulation of LPOs serves as a critical determinant. Lipoxygenases (LOXs), non-heme iron-containing enzymes, catalyze the stereospecific oxygenation of free polyunsaturated fatty acids (PUFAs) into lipid hydroperoxides through molecular oxygen incorporation. This enzymatic activity plays a pivotal role in driving ferroptosis progression (Liu et al., 2023; Mortensen et al., 2023). Membrane-incorporated PUFAs serve as primary targets for ROS-mediated oxidation, with their oxidative susceptibility increasing proportionally to the number of conjugated double bonds (Rouzer and Marnett, 2003), following the initiation of oxidation, free radicals exhibit dual propagation mechanisms: intramolecular migration within lipid bilayers or intermolecular chain reactions amplifying oxidative damage (Samovich et al., 2024). Notably, arachidonic acid (AA) and adrenal acid (AdA), the most oxidation-prone PUFA species, demonstrate particular vulnerability to this peroxidation cascade.
Emerging evidence identifies AA/AdA-derived phosphatidylethanolamine (PE) as a pivotal substrate for ferroptotic lipid peroxidation (Kagan et al., 2017; Samovich et al., 2024). The enzymatic cascade begins with ACSL4 converting free AA/AdA into AA-CoA/AdA-CoA, followed by lysophosphatidylcholine acyltransferase 3 (LPCAT3)-mediated esterification to generate AA-PE/AdA-PE. These phospholipid conjugates undergo iron/LOX-dependent oxidation to PLOOH, thereby initiating membrane peroxidation and ferroptotic execution (Yang et al., 2016; Dierge et al., 2021). Contrastingly, monounsaturated fatty acids (MUFAs) are preferentially incorporated into membrane phospholipids via ACSL3-mediated acyl-CoA synthesis. Exogenous MUFAs suppress ferroptosis through ACSL3 activation, effectively attenuating lipid ROS accumulation (Ru et al., 2024). Notably, Huang et al. identified AS-252424 as a competitive inhibitor targeting the Gln464 residue of ACSL4, effectively blocking enzymatic activity and ferroptotic progression (Huang et al., 2024). Furthermore, redox enzymes contribute to lipid peroxidation by transferring electrons from NADPH to oxygen, generating hydrogen peroxide. This process facilitates Fenton reactions with labile iron pools, thereby amplifying oxidative chain reactions that drive ferroptosis (Yan B. et al., 2021).
4.1.3 The FSP1-CoQ10-NADPH pathway
Researchers identified genes in tumor cells capable of compensating for the deficiency of GPX4, a key enzyme in ferroptosis, through expression cloning. Apoptosis - inducing factor mitochondria - associated 2 (AIFM2), a lipid - binding protein, was recognized as a novel anti - ferroptosis gene, Bersuker et al. renamed AIFM2 as FSP1 (Bersuker et al., 2019; Doll et al., 2019). The inhibition of ferroptosis by FSP1 is mediated by CoQ10. Specifically, the reduced form of CoQ10 can scavenge lipid peroxyl radicals, which mediate lipid peroxidation. FSP1 can regenerate CoQ10 by catalyzing NADPH (Doll et al., 2019). In detail, in the FSP1 - CoQ10 - NADPH pathway, FSP1, as an effective ferroptosis inhibitor, reduces CoQ10 on the plasma membrane to ubiquinol. This process either directly inhibits lipid peroxidation or indirectly promotes the regeneration of tocopherol radicals (vitamin E), a natural chain - breaking antioxidant.
Recent studies have shown that FSP1 possesses a unique carboxyl - terminal domain. This domain can mediate the functional dimerization of FSP1 and the formation of two active sites, which are crucial for the catalytic activity and ferroptosis - inhibitory property of FSP1. In vitro, in the presence of oxygen and NADPH, FSP1 can catalyze the generation of hydrogen peroxide and convert FAD into 6 - hydroxy - FAD. 6 - hydroxy - FAD is not only an active co - factor of FSP1 but also an effective radical - trapping antioxidant that can directly inhibit ferroptosis in cells (Lv et al., 2023). As a glutathione - independent ferroptosis suppressor, FSP1, by regulating the CoQ10 and NADPH pathways, collaborates with GPX4 to inhibit phospholipid peroxidation and ferroptosis, providing new targets and strategies for cancer treatment.
4.1.4 Glutamine metabolic pathway
The glutamine (Gln) metabolic pathway plays a critical role in regulating ferroptosis. Gln is primarily taken up by cells through solute carrier family 1 member 5 (SLC1A5) and subsequently catabolized to glutamate (Glu) by glutaminase (GLS) (Bodineau et al., 2022; Li S. et al., 2023). Glu is then converted to α-ketoglutarate (α-KG) via glutamate dehydrogenase (GLUD1), which enters the tricarboxylic acid (TCA) cycle (Yao et al., 2021), α-KG accumulation promotes oxidative stress by enhancing ROS production and lipid peroxidation, thereby inducing ferroptosis (Gao et al., 2015). Notably, Glu triggers mitochondrial structural and functional alterations, including membrane potential depolarization and reduced electron transport chain activity, further contributing to lipid ROS accumulation. Furthermore, Glu serves as a precursor for glutathione (GSH) synthesis. As the primary intracellular antioxidant, GSH scavenges excessive ROS to mitigate lipid peroxidation, a mechanism crucial for ferroptosis prevention (Kanaan et al., 2024).
Gln participates in the TCA cycle and energy metabolism, providing biosynthetic energy substrates. Under oxidative stress, metabolic reprogramming may influence cellular survival and ferroptosis susceptibility (Suzuki et al., 2022), Gln also modulates iron homeostasis by regulating its uptake and storage, thereby indirectly controlling iron overload risks. Excessive iron accumulation potentiates ROS generation, ultimately triggering ferroptosis (Chen et al., 2024). A recent study demonstrated that Gln protects nucleus pulposus (NP) cells from tert-butyl hydroperoxide (TBHP)-induced ferroptosis and extracellular matrix (ECM) degradation. This protective effect occurs through enhanced Nrf2 stabilization via inhibition of ubiquitin-proteasome degradation, coupled with suppressed lipid oxidation (Fu et al., 2024; Wu J. et al., 2024).
4.1.5 Mevalonate pathway
The mevalonate (MVA) pathway plays a pivotal role in ferroptosis regulation by modulating selenocysteine-tRNA maturation, thereby influencing GPX4 biosynthesis. Selenocysteine (Sec), an essential catalytic residue in GPX4’s active site, is indispensable for its antioxidant function (Chen et al., 2020). The MVA pathway initiates from acetyl-CoA and sequentially generates isopentenyl pyrophosphate (IPP) through reductase-mediated steps, with critical enzymatic involvement from mevalonate kinase (MVK). Crucially, MVA-derived metabolites including IPP and CoQ10 regulate Sec-tRNA maturation and subsequent GPX4 synthesis. The incorporation of Sec during GPX4 translation proves vital for maintaining its redox-protective capacity (Wu et al., 2022). Pharmacological inhibition of the MVA pathway impairs Sec-tRNA biogenesis, leading to GPX4 inactivation, diminished cellular antioxidant defenses, lipid peroxide accumulation, and ultimately ferroptosis induction (Chen et al., 2020).
Recent studies demonstrate that the bisphosphonate alendronate synergistically enhances paclitaxel efficacy, suppresses tumor metastasis, and modifies cytotoxic mechanisms in cancer therapy. Mechanistically, alendronate inhibits mevalonate metabolism, induces mitochondrial morphological alterations, disrupts redox homeostasis, and promotes accumulation of mitochondrial ROS and LPOs - ultimately triggering tumor cell ferroptosis. Notably, hepatocellular carcinoma (HCC) cells overexpressing miR-612 exhibit heightened ferroptosis sensitivity, with miR-612 potentiating lipid ROS accumulation (Song G. et al., 2024). Xing et al. further revealed that miR-612 promotes HCC ferroptosis through HADHA-mediated regulation of the mevalonate pathway, downregulating CoQ10 while elevating intracellular PUFA levels and lipid peroxidation, thereby inhibiting HCC proliferation and metastasis (Xing et al., 2023). Complementary findings by Tang et al. demonstrate that pitavastatin induces ferroptosis in malondialdehyde (MDA)-MB-231 triple-negative breast cancer cells via mevalonate pathway inhibition, with exogenous mevalonate supplementation partially reversing pitavastatin-induced cytotoxicity and the reduced expression of GPx4/FSP1 (Tang W.-J. et al., 2024).
4.2 Non-enzymatic mechanisms
4.2.1 Disorder of iron metabolism
Iron metabolism dysregulation is intrinsically linked to ferroptosis pathogenesis. Under physiological conditions, systemic iron homeostasis is dynamically regulated through coordinated expression of transferrin (Tf), transferrin receptor 1 (TfR1), ferroportin 1 (FPN1), and hepatic iron regulatory proteins (Galy et al., 2024). Tf, a single-chain polypeptide serving as the principal iron-transport protein, serves as a biochemical indicator of systemic iron metabolism (Zhang D. et al., 2024). TfR1, a transmembrane protein mediating cellular iron uptake through Tf-bound iron internalization, has been identified as a specific ferroptosis biomarker (Feng et al., 2020). The iron transport cascade initiates with extracellular Fe3+ binding to Tf, followed by TfR1-mediated endocytosis. Within endosomal vesicles, Fe3+ undergoes six-transmembrane epithelial antigen of prostate 3 (STEAP3)-catalyzed reduction to Fe2+, subsequently exported to cytosol via divalent metal transporter 1 (DMT1) (Wang H. et al., 2023). When iron-binding capacity approaches saturation, excess Fe2+ complexes are sequestered in the labile iron pool (LIP) through ferritin binding. LIP-derived Fe2+ participates in Fenton reactions, generating hydroxyl radical species that drive membrane lipid peroxidation and subsequent ferroptosis (Morales and Xue, 2021), furthermore, ferritin-bound iron undergoes Nuclear receptor coactivator 4 (NCOA4)-mediated autophagic degradation in lysosomes, releasing free Fe2+ into the intracellular compartment (Chen et al., 2022b).
The Fenton reaction, mediated by Fe2+ and hydrogen peroxide, generates hydroxyl radicals that initiate non-enzymatic lipid peroxidation by abstracting hydrogen atoms from lipids, forming lipid radicals (Sun et al., 2022). Notably, extracellular conversion of FPN1-exported Fe2+ into less redox-active Fe3+ is facilitated by ceruloplasmin (CP). CP deficiency disrupts this process, leading to pathological Fe2+ accumulation and subsequent ferroptosis induction (Shang et al., 2020).
4.2.2 System Xc−
System Xc−, a critical intracellular antioxidant component, is an amino acid antiporter composed of the light chain SLC7A11 (xCT) and heavy chain SLC3A2 (4F2hc), which are interconnected by disulfide bonds (Li F.-J. et al., 2022; Aboushousha et al., n.d.). This transporter operates via the Glu and Cys concentration gradient across cellular membranes, primarily facilitating extracellular Cys import and intracellular Glu export in a 1:1 stoichiometric exchange. This mechanism is indispensable for GSH synthesis (Wu et al., 2021; Li F.-J. et al., 2022). Within this pathway, ingested Cys is catalyzed by thioredoxin reductase 1 (TrxR1) to convert it to Cys, followed by conjugation with Glu via glutamate-cysteine ligase (GCL) to yield γ-glutamyl-L-cysteine. This intermediate subsequently combines with glycine to complete GSH biosynthesis, thereby inhibiting ferroptosis (Li F.-J. et al., 2022; Liu J. et al., 2022; Saini et al., 2023; Zhang G. et al., 2024).
System Xc− serves as the pivotal upstream regulator of the System Xc−/GSH/GPX4 axis, with its activity primarily governed by SLC7A11 expression. SLC7A11 downregulation diminishes System Xc− activity, promoting oxidative stress-mediated ferroptosis. Conversely, SLC7A11 upregulation enhances cellular resistance to ferroptosis, a mechanism implicated in tumor chemoresistance (Wang et al., 2020; Xue et al., 2024).
4.2.3 Fenton reaction
The Fenton reaction, a redox interaction between iron and hydrogen peroxide, generates hydroxyl radicals—highly reactive and cytotoxic species that drive ferroptosis by abstracting hydrogen atoms from lipids to initiate non-enzymatic lipid peroxidation (He et al., 2020; Abe et al., 2022). Henning et al. demonstrated that sodium iodate (SI) and dimethyloxalylglycine (DMOG) treatment reduced superoxide dismutase activity in retinal pigment epithelial (RPE) cells, revealing hypoxia-enhanced Fenton reactions as a mechanism exacerbating ferroptosis in RPE cells (Henning et al., 2022). Recent studies indicate phenolic compounds induce ferroptosis in multiple cell types by forming iron complexes that potentiate Fenton reactivity, leading to hydroxyl radical overaccumulation and ferroptosis-like death (Sui et al., 2024). Furthermore, Fenton reaction-based nanocatalytic therapies show promise for targeting ferroptosis-related pathways in cancer treatment (Xing et al., 2021; Zhu et al., 2022; Zhao et al., 2023; Song Q. et al., 2024).
Given that tumor cells exhibit elevated ROS levels due to accelerated metabolic and proliferative activity, ferroptosis—a process intimately linked to redox imbalance—may offer a targeted therapeutic approach for MM and other cancers (Chiang et al., 2018; Du et al., 2023). Rajkumar et al. demonstrated the anti-proliferative effects of ferrotoxic interventions in MM murine models (Rajkumar et al., 2014). Subsequent studies revealed heightened sensitivity of MM cell lines to ferroptosis, with combined bortezomib and iron chelation exhibiting superior efficacy in suppressing disease progression compared to monotherapy, these findings collectively underscore the critical role of ferroptosis in MM pathogenesis (Campanella et al., 2013; Bordini et al., 2017) (Figure 3).
FIGURE 3
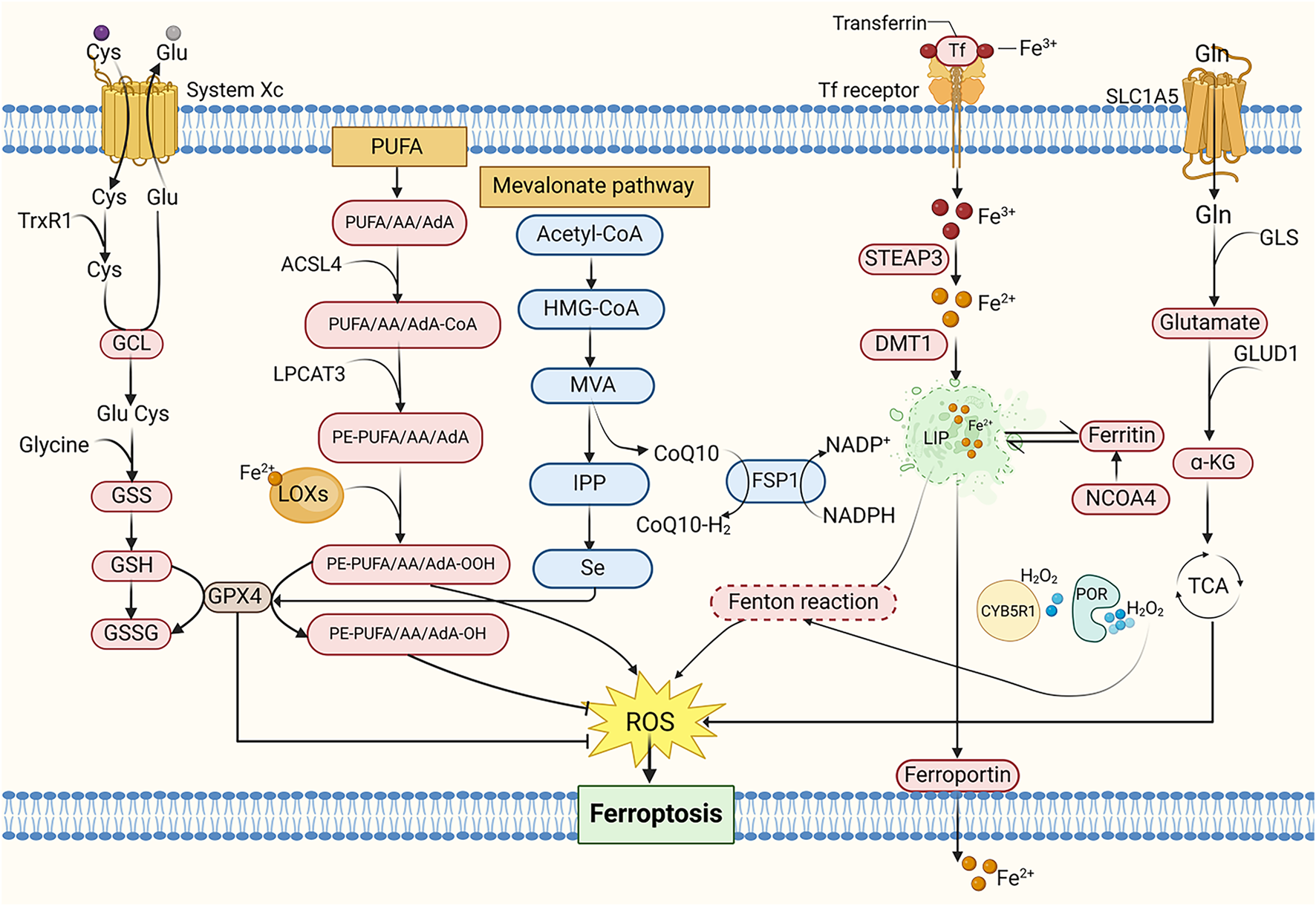
The mechanism of ferroptosis. Cys, Cysteine; Glu, Glutamate; TrxR1, Thioredoxin reductase 1; GCL, Glutamate-cysteine ligase; GSS, Glutathione synthetase; GSH, Glutathione; GSSG, Oxidative glutathione; PUFA, Polyunsaturated fatty acid; AA, Arachidonic acid; AdA, Adrenal acid; ACSL4, Acyl-CoA synthetase long-chain family member 4; LPCAT3, Lysophosphatidylcholine acyltransferase 3; PE, Phosphatidylethanolamine; LOXs, Lipoxygenases; MVA, Mevalonate; IPP, Isopentenyl pyrophosphate; HMG-CoA, 3-Hydroxy-3-Methyl-Glutaryl-CoA; CoQ10, Coenzyme Q10; FSP1, Ferroptosis suppressor protein 1; NADPH, Nicotinamide adenine dinucleotide phosphate; Tf, Transferrin; STEAP3, Six-transmembrane epithelial antigen of prostate 3; DMT1, Divalent metal transporter 1; LIP, Labile iron pool; NCOA4, Nuclear receptor coactivator 4; CYB5R1, NADH-cytochrome b5 reductase; POR, Cytochrome P450 reductase; Gln, Glutamine; SLC1A5, Solute carrier family 1 member 5; GLS, Glutaminase; GLUD1, Glutamate dehydrogenase; α-KG, α-ketoglutarate; TCA cycle, Tricarboxylic acid cycle.
5 Mechanistic differences of ferroptosis in normal and cancerous tissues
Ferroptosis, an iron-dependent lipid peroxidation-driven form of cell death, exhibits significant differences in its mechanisms between normal and cancerous tissues (Jiang X. et al., 2021). These differences are closely related to cell type characteristics, redox status, and the local microenvironment, which directly determine the contrasting roles of inducers and inhibitors in disease therapy.
In normal tissues, ferroptosis is typically associated with cellular stress responses, antioxidant defense mechanisms, and immune regulation. For example, in neurodegenerative diseases such as Alzheimer’s and Parkinson’s diseases, excessive oxidative damage in neuronal cells may accelerate disease progression by inducing ferroptosis, making ferroptosis inhibitors potential therapeutic agents. Ferroptosis inhibitors can slow down neuronal cell death and protect nerve function (Yang et al., 2022; Ryan et al., 2023; Zhang T.-C. et al., 2024).
In contrast, cancerous tissues exhibit a distinct scenario. Tumor cells often possess a strong antioxidant capacity to counteract redox imbalance. Due to the generally higher metabolic activity in cancer cells and the increased levels of ROS associated with cancer progression, they may be more susceptible to ferroptosis (Lei et al., 2022). Furthermore, studies have shown that tumor cells typically require higher iron availability, which may further increase their sensitivity to ferroptosis (Torti and Torti, 2013; Manz et al., 2016). Ferroptosis inducers can promote the accumulation of intracellular iron and lipid peroxides, activating the ferroptosis pathway and leading to tumor cell death. Therefore, ferroptosis inducers are considered a promising therapeutic strategy, especially for treating refractory cancers. By modulating ferroptosis induction or inhibition, different therapeutic effects can be achieved in various disease microenvironments, further highlighting the complexity of ferroptosis in different tissue types.
6 MM and ferroptosis
6.1 Lipid metabolism pathway
6.1.1 AA pathway
Studies have demonstrated significant reductions in key PUFAs, including ferroptosis-inducing AA, within bone marrow aspirates of MM patients (Chen et al., 2023). Elevated expression of ferroptosis-suppressing genes correlates with reduced progression-free survival (PFS) and overall survival (OS) in MM patients (Su et al., 2022). Co-culture experiments of murine MM cells with bone marrow adipocytes revealed that MM cells upregulate fatty acid transport proteins 1 and 4 (FATP1, FATP4) to acquire PUFAs from neighboring adipocytes. PUFA exposure exerts dose-dependent effects on MM cells, with high concentrations triggering lipotoxicity via ferroptosis activation (Panaroni et al., 2022). These findings collectively implicate ferroptosis-mediated lipid metabolism dysregulation in MM pathogenesis. Free fatty acids (FFAs) exhibit dual regulatory roles: low concentrations promote MM proliferation, whereas elevated levels induce ferroptosis-dependent cytotoxicity. Notably, bone marrow lipid content is markedly increased in precursor MM states (MGUS and SMM) compared to healthy controls (Panaroni et al., 2022). AA administration dose-dependently suppresses proliferation and viability in human MM cell lines (MM1S, H929, U266) and enhances apoptosis rates in SCID mouse xenograft models. Therapeutic targeting of ferroptosis may yield superior efficacy during early disease stages (MGUS and SMM) (Panaroni et al., 2018). Paradoxically, while high AA concentrations induce MM cell death, subtoxic levels stimulate tumor growth through FATP-mediated FA uptake and adipocyte lipolysis (Panaroni et al., 2022).
Intriguingly, MM plasma cells exhibit elevated GPX4 and xCT levels compared to normal counterparts. Given their critical roles in ferroptosis suppression, these antioxidant regulators represent promising therapeutic targets for counteracting ROS-mediated cytotoxicity (Mynott et al., 2023). Precise determination of AA’s physiological thresholds is essential to maintain anti-tumor efficacy without triggering pro-survival adaptations.
6.1.2 ACSL4 pathway
ACSL4 is aberrantly overexpressed in MM cell lines and patient-derived samples compared to healthy donors. This dysregulation exerts dual biological effects: ACSL4 enhances fatty acid accumulation by modulating lipid metabolism regulators (e.g., c-Myc and sterol regulatory element-binding proteins, SREBPs) to drive MM proliferation (Liang et al., 2023). However, as a critical ferroptosis driver, ACSL4 governs MM cell sensitivity to ferroptosis inducers like RSL3, with elevated ACSL4 levels potentiating ferroptotic vulnerability (Zhang et al., 2023). ACSL4 knockdown suppresses MM proliferation and attenuates intracellular fatty acid accumulation, potentially via modulation of lipid metabolic regulators. Crucially, ACSL4 silencing confers resistance to ferroptosis, directly linking its expression levels to cellular ferroptotic sensitivity (Zhang et al., 2023). These mechanistic insights position ferroptosis induction as a promising therapeutic strategy for MM, where pharmacological modulation of ACSL4 activity/expression could augment therapeutic efficacy.
6.2 Iron metabolism pathway
Iron overload has been shown to impair MM cell proliferation, an effect reversible by the ferroptosis inhibitor ferrostatin-1. High-dose ferric ammonium citrate (FeAC) triggers MM cell death with concurrent elevation of MDA - a lipid peroxidation byproduct and hallmark of ferroptosis, establishing iron metabolism as a critical modulator in MM pathophysiology (Bordini et al., 2020; Camiolo et al., 2020). Mechanistically, free iron accumulation induces dose-dependent aggregation of polyubiquitinated proteins in iron-sensitive MM H929 cells, while suppressing chymotrypsin-like proteasomal activity in cell lysates. This dual mechanism suggests that iron overload may potentiate proteasome inhibitor efficacy through lipid oxidation and proteostasis disruption, providing a rationale for combinatorial therapeutic strategies (Bordini et al., 2020; Camiolo et al., 2020). Iron homeostasis dysregulation in MM is further linked to altered hepcidin expression patterns. Although hepcidin (a hepatic iron-regulatory peptide) typically restricts systemic iron release (Nemeth and Ganz, 2023), MM patients exhibit paradoxical hepcidin downregulation or functional impairment during iron overload, creating a self-reinforcing cycle of iron accumulation that exacerbates disease progression (Maes et al., 2010; Banaszkiewicz et al., 2022).
6.3 Oxidative stress pathway
Emerging evidence demonstrates that andrographolide, a natural bioactive compound, induces ferroptosis in MM cells via P38-mediated suppression of the Nrf2/heme oxygenase-1 (HO-1) signaling axis, thereby triggering oxidative stress (Li W. et al., 2023). Mechanistically, Nrf2 serves as the master transcriptional regulator of antioxidant responses, orchestrating cellular iron homeostasis, redox balance, and mitochondrial function through transcriptional regulation of FPN1, GPX4, and HO-1. Crucially, Nrf2 knockout models exhibit diminished SLC7A11 and HO-1 protein expression, which potentiates ferroptotic susceptibility (Dong et al., 2020).
6.4 Autophagy pathway
Beclin-1, a core autophagy-related protein, drives autophagosome formation, while UNC-51 like kinase 1 (ULK1) coordinates autophagic initiation through its kinase complex (Kim et al., 2011). Notably, protein phosphatase 2A (PP2A) and AMP-activated protein kinase (AMPK) modulate the phosphorylation status of Beclin-1 to regulate autophagic flux (Peris et al., 2023). PP2A specifically dephosphorylates Beclin-1 at Ser90, whereas AMPK–activated under energy stress–phosphorylates ULK1 to initiate autophagy, and targets Beclin-1 at Ser91/Ser94 residues (Kim et al., 2013; Park et al., 2023). Functionally, the PP2A-AMPK axis orchestrates autophagic-ferroptotic crosstalk in MM cells. PP2A activation via dephosphorylation of its catalytic subunit C at Tyr307 induces AMPK inactivation through Thr172 dephosphorylation. This cascade attenuates eukaryotic elongation factor 2 kinase (eEF2k) activity, thereby relieving its inhibition of eEF2 to enhance energy expenditure and drive dual autophagic-ferroptotic cell death (Zhong et al., 2020). CRISPR/Cas9-mediated knockout of autophagy regulators ATG5/ATG7 in RPMI-8226 MM cells significantly increased cell viability compared to wild-type controls upon RSL3-induced ferroptosis, confirming autophagy’s synergistic role in ferroptotic execution (Li and Lyu, 2024).
6.5 Other regulatory mechanisms
The interplay between ferroptosis and epigenetic regulation has garnered increasing attention due to their shared associations with oxidative stress and dysregulated iron metabolism. Induction of ferroptosis in MM cells upregulates key genes implicated in cellular stress, death, inflammation, and fatty acid metabolism, including ferritin heavy chain 1 (FTH1), HO-1, and SLC7A11. Studies have demonstrated that ferroptosis induction in MM cells triggers DNA methylation and histone modification changes associated with cellular senescence. Logie et al. revealed that MM1 cells exhibit sensitivity to both the ferroptosis inducer RSL3 and epigenetic reprogramming. Liquid chromatography-tandem mass spectrometry (LC-MS/MS) analysis identified the formation of non-heme iron-histone complexes and altered histone modifications linked to DNA repair and senescence. Using EPIC BeadChip profiling, the authors observed significant DNA methylation alterations in ferroptotic MM cells, enriched in genes regulating cell cycle progression and senescence, such as nuclear receptor subfamily 4 group A member 2 (NR4A2). These findings highlight the connection between ferroptosis and epigenomic stress responses, underscoring the therapeutic potential of ferroptosis-inducing agents (Logie et al., 2021).
Emerging evidence indicates that the crosstalk between bone marrow mesenchymal stromal cells (BMSCs) and MM cells enhances ferroptosis susceptibility in the latter. BMSCs elevate intracellular iron levels in MM cells, thereby activating steroid biosynthesis pathways such as lanosterol production. Lanosterol, a major source of ROS in MM cells, is regulated by the CD40 ligand (CD40L)-CD40 receptor interaction, which serves as a critical signaling axis. BMSCs modulate the CD40/CD40L pathway to influence iron homeostasis and lanosterol synthesis in MM cells, driving ROS accumulation and ferroptosis (Jiang et al., 2024). A recent study further revealed that leukocyte immunoglobulin-like receptor B1 (LILRB1), through complex formation with other proteins, promotes low-density lipoprotein and cholesterol uptake. LILRB1 deficiency disrupts cholesterol uptake but activates compensatory cholesterol biosynthesis to maintain cellular cholesterol homeostasis. This shift reduces levels of the anti-ferroptotic metabolite squalene, rendering MM cells more prone to ferroptosis. Given its role in safeguarding MM cells against ferroptosis via cholesterol regulation, LILRB1 represents a promising therapeutic target (Xian et al., 2024).
A recent study revealed that mitochondrial metabolic kinase PCK2 phosphorylates and activates ACSL4, driving phospholipid remodeling linked to ferroptosis. Tumor-reprogrammed cells (TRCs), characterized by stem-like properties, downregulate PCK2 expression to adopt a ferroptosis-resistant state, suggesting that TRCs evade radio- and chemotherapy by reducing ferroptosis susceptibility. These findings highlight novel insights into ferroptosis in malignant tumors (Li Z. et al., 2024). Fu et al. investigated the clinical and biological relevance of ferroptosis-related genes in MM. Their prognostic model underscored the critical role of ferroptosis in MM patient outcomes. In vitro experiments demonstrated synergistic antitumor efficacy of erastin and doxorubicin, mediated by GPX4 degradation and excessive ROS production, which collectively reduced viability of NCI-H929 and RPMI-8226 cells (Fu et al., 2022). Furthermore, glutamine blockade combined with radiotherapy (RT) induces immunogenic ferroptosis in tumor cells. Mechanistically, RT activates the interferon signaling pathway to upregulate interferon regulatory factor 1 (IRF1), an effect amplified by glutamine inhibition. IRF1 drives transferrin receptor expression, elevates intracellular Fe2+ levels, and disrupts iron homeostasis, thereby promoting ferroptosis. The therapeutic potential of this axis in hematologic malignancies warrants further exploration (Song A. et al., 2024) (Figure 4).
FIGURE 4
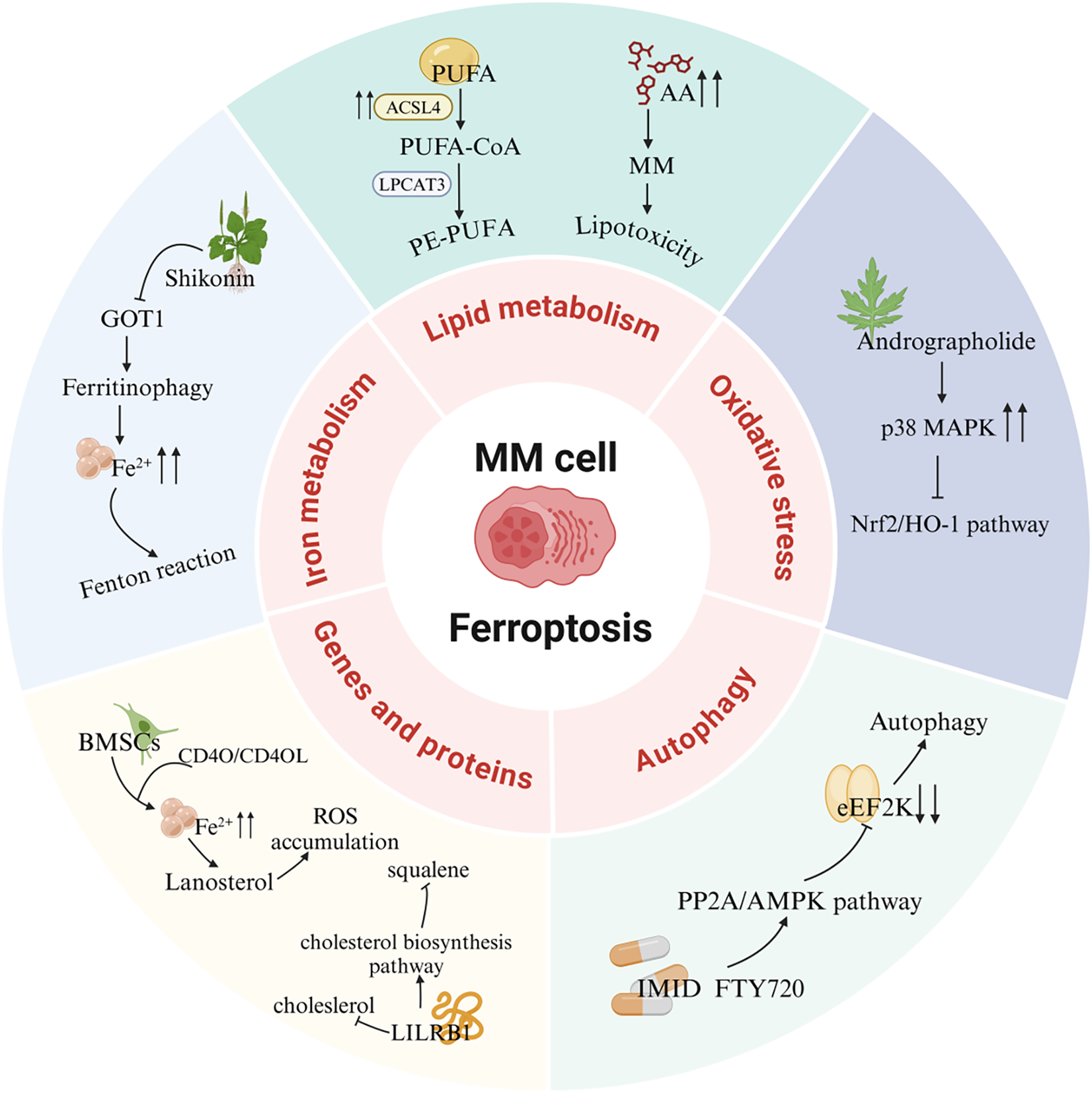
Ferroptosis signaling pathways in MM. PUFA, Polyunsaturated fatty acid; ACSL4, Acyl-CoA synthetase long-chain family member 4; LPCAT3, Lysophosphatidylcholine acyltransferase 3; AA, Arachidonic acid; MAPK, Mitogen-activated protein kinase; Nrf2, Nuclear factor E2-related factor 2; HO-1, Heme oxygenase-1; eEF2K, eukaryotic elongation factor 2 kinase; PP2A, Protein phosphatase 2A; AMPK, AMP-activated protein kinase; IMID, Immunomodulatory drug; FTY720, Fingolimod; BMSCs, Bone marrow mesenchymal stromal cells; LILRB1, Leukocyte immunoglobulin-like receptor B1; GOT1, Glutamic-oxaloacetic transaminase 1.
7 Ferroptosis-based treatment strategies for MM
7.1 Active ingredients of natural products
The partial inhibition of cell death by Ferrostatin-1, Liproxstatin-1, or Deferoxamine, alongside evidence that natural product extracts induce MM cell death, implicates ferroptosis in MM cell demise. This further highlights plant-derived extracts as promising tools for triggering ferroptosis in MM.
Shikonin (SHK), a natural compound isolated from Lithospermum erythrorhizon roots, exhibits diverse bioactivities, including antioxidant, anti-inflammatory, and antitumor effects, and functions as a proteasome inhibitor and necroptosis inducer (Guo et al., 2019; Guo et al., 2023 Y.; Yan et al., 2023). Shikonin exerts anti-angiogenic effects by downregulating the PI3K/AKT and MAPK signaling pathways, and also displays anticancer activity by targeting ROS (Anti-angiogenic effect of Shikonin, 2020). Studies demonstrate that SHK treatment promotes lactate dehydrogenase release, triggers cell death, induces oxidative stress, and elevates ferrous iron and lipid peroxidation levels. Notably, ferroptosis inhibitors reverse SHK-induced cytotoxicity, confirming ferroptosis involvement. Further mechanistic studies revealed that glutamic-oxaloacetic transaminase 1 (GOT1) serves as a key mediator of SHK-driven ferroptosis by promoting ferritinophagy. These findings underscore SHK’s potential as a therapeutic agent for MM (Li W. et al., 2022).
Apigenin (Api), an edible plant-derived flavonoid, has been recognized as an antitumor agent in numerous experimental and biological studies. It demonstrates cytostatic and pro-apoptotic effects across various malignancies through modulation of multiple signaling pathways (Imran et al., 2020; Jiang Z.-B. et al., 2021; Shi et al., 2023; Sudhakaran et al., 2023; Naponelli et al., 2024). A recent study demonstrated that Api significantly inhibits the progression of bladder cancer by targeting VEGF-β (Apigenin inhibits recurrent bladder cancer, 2025). In addition, it has shown potential therapeutic effects against viral infections (Lee et al., 2023). Notably, our investigation reveals that Api induces myeloma cell apoptosis via the STAT1/COX-2/iNOS signaling axis, demonstrating marked growth inhibition in MM cells with the most pronounced concentration-dependent effect observed in NCI-H929 cells. Importantly, pretreatment with ferrostatin-1 and deferoxamine attenuated Api’s efficacy on NCI-H929 cells by over threefold, strongly suggesting the potential involvement of ferroptosis-related pathways in Api-mediated MM cell death. Further mechanistic investigations are warranted to fully elucidate this regulatory network (Adham et al., 2021a).
Artesunate (ART), a water-soluble hemisuccinate derivative of dihydroartemisinin (DHA), originates from artemisinin–an active compound isolated from Artemisia annua with extensive traditional Chinese medicinal applications and current use as a frontline antimalarial agent (Qian et al., 2021; Strik et al., 2024). ART exhibits both anticancer activity and cytotoxicity against solid tumors and leukemias, having been applied as an adjuvant therapy for diverse malignancies (Vakhrusheva et al., 2022; Hill et al., 2023; Fan et al., 2024). ART exerts its antitumor effects through multiple pathways, including PPARγ-SCD and PI3K/AKT/FKHR signaling (Xu et al., 2022; FABP5, 2024). Liang et al. investigated ART’s effects on cellular growth, ROS generation, Fe2+ levels, lipid peroxidation, and ferroptosis-related gene expression, while further exploring ferroptosis mechanisms in MM cells and their association with sterol regulatory element-binding protein 2 (SREBP2) nuclear localization. ART treatment upregulated ROS, Fe2+, and lipid peroxidation levels while suppressing MM cell growth, concomitant with increased ACSL4 expression and decreased GPX4 levels. Notably, ART-induced cell death was reversed by ferroptosis inhibitors ferrostatin-1 (Fer-1) and deferoxamine (DFO). Mechanistically, ART suppressed nuclear localization of SREBP2 in MM cells, accompanied by downregulation of IPP and GPX4, suggesting its capacity to trigger ferroptosis through SREBP2 nuclear localization inhibition and subsequent IPP/GPX4 suppression (Liang et al., 2023).
Eclipta prostrata is a herbaceous plant, and its aerial parts are used as traditional medicine. In traditional Chinese medicine, Eclipta prostrata is employed to treat bleeding, liver diseases, kidney injuries, and snake bites (Fang et al., 2015; Tian et al., 2023). Modern pharmacological studies have shown that Eclipta prostrata exhibits various biological activities, such as anti - tumor, anti - snake venom, anti - inflammatory, antioxidant, and hypolipidemic effects (Timalsina and Devkota, 2021; Yang J. et al., 2023). Its extracts have been found to promote osteogenic differentiation of bone marrow mesenchymal stem cells by targeting METTL3-mediated m6A RNA methylation (Tian et al., 2023). Li et al. found that the ethanol extract of Eclipta prostrata (EEEP) could inhibit the growth of MM cells and induce cell death both in vitro and in vivo. EEEP triggers ferroptosis in MM cells by promoting the accumulation of MDA and Fe2+, lipid peroxidation, and inhibiting GSH. Mechanistically, EEEP - induced lipid peroxidation and MDA accumulation were blocked by the Nrf2 activator NK - 252. EEEP regulates the kelch-like ECH-associated protein 1 (Keap1)/Nrf2/HO - 1 axis and stimulates ferroptosis in MM cells (Li W. et al., 2024).
Thymus vulgaris L. is an important plant that can be used both medicinally and as food. It has demonstrated various pharmacological effects in both traditional medicine and modern medical research. Its main active components include thymol, carvacrol, flavonoids, and volatile oils (Bolatli et al., 2023; Zhou et al., 2023). Arctium lappa L. belongs to the Asteraceae family and has been widely used in traditional Chinese medicine (TCM). It is rich in highly - recognized bioactive metabolites with antioxidant, anticancer, and neuroprotective activities (Yang et al., 2021). Additionally, Arctium lappa L. also shows significant value in nanomedicine applications (Yosri et al., 2023). A study by Adham et al. found that the extracts of Thymus vulgaris (TCF) and Arctium lappa (ACF) can induce cell - cycle arrest and ferroptosis in MM cells and leukemia cells. The researchers observed that myeloma NCI - H929 cells were significantly sensitive to the chloroform fractions of TCF and ACF, and ferroptosis inhibitors eliminated the cytotoxicity of TCF and ACF extracts (Adham et al., 2020).
Andrographolide (AP), a diterpenoid lactone isolated from Andrographis paniculata, is the main component of this plant and exhibits a wide range of pharmacological effects. Multiple studies have demonstrated its potent antitumor activity (Li et al., 2020b; Li et al., 2024 J.; Jiaqi et al., 2023). Andrographolide promotes ferroptosis by inducing mitochondrial dysfunction, thereby inhibiting the development and progression of non-small cell lung cancer (Jiaqi et al., 2023). Research has found that AP can induce the death of MM cells, cause G0/G1 cell - cycle arrest, and trigger oxidative stress responses. These phenomena are accompanied by an increase in the levels of Fe2+ and lipid peroxidation in both the cytoplasm and mitochondria. Further research shows that AP may block the Nrf2/HO - 1 signaling pathway by activating P38, thereby inducing ferroptosis. Additionally, the use of ferroptosis inhibitors can rescue the MM cell death induced by AP, indicating a close relationship between AP - induced MM cell death and the ferroptosis pathway (Li W. et al., 2023).
Ginsenoside Rh4 is an active compound extracted from ginseng. It belongs to protopanaxatriol - type ginsenoside saponins and exhibits a variety of pharmacological activities (Wang et al., 2024). It has been proven to improve antibiotic - induced gut microbiota dysbiosis and intestinal inflammation and inhibit the growth of various tumors (Wu et al., 2018; Dong et al., 2023; Jiang et al., 2023; Bai et al., 2024). Ginsenoside Rh4 suppresses the progression of colorectal cancer by modulating gut microbiota-mediated bile acid metabolism. It also inhibits tumor metastasis by targeting signaling pathways such as Wnt/β-catenin and TGF-β/Smad2/3 (Chen et al., 2022a; Bai et al., 2024). A study by Ying et al. found that ginsenoside Rh4 can inhibit the proliferation of MM cells, induce cell apoptosis, promote cell - cycle arrest, etc., and play a key role in inducing ferroptosis in MM cells (Ying et al., 2023). This process may be related to the inhibition of Sirtuin 2 (SIRT2) expression in MM cells by ginsenoside Rh4. SIRT2 regulates the activity of lipid - peroxidation - related enzymes to reduce the occurrence of ferroptosis and may also participate in regulating the ferroptosis sensitivity of myeloma cells by affecting lipid metabolism and antioxidant regulation (Feng et al., 2024).
Fumaria officinalis is a traditional herbal medicine containing various active components and exhibits multiple pharmacological effects such as antioxidant, antibacterial, anti - inflammatory and antispasmodic, as well as immunomodulatory activities (Raafat and El-Zahaby, 2020; Shah et al., 2024). The chloroform and ethyl acetate (EF) fractions of the Fumaria officinalis extract show significant cytotoxic effects on human acute lymphoblastic leukemia cells and MM cells. Among them, EF induces autophagic cell death, while chloroform can stimulate iron - dependent cell death (Adham et al., 2021b).
Nitidine chloride (NC) is an alkaloid extracted from the dried roots of Zanthoxylum nitidum, a plant of the Rutaceae family. It exhibits various pharmacological activities, including antitumor, anti-inflammatory, antimalarial, and antioxidant effects (Lu et al., 2022). NC has been reported to induce caspase-3/GSDME-dependent pyroptosis in lung cancer by inhibiting the PI3K/Akt pathway (Yu et al., 2022). In multiple myeloma (MM), ATP-binding cassette sub-family B member 6 (ABCB6) has been identified as a key protein mediating ferroptosis resistance by maintaining glutathione (GSH) homeostasis and activating the PI3K/AKT signaling pathway. NC targets ABCB6, suppresses the PI3K/AKT pathway, and induces ferroptosis in MM cells (Yin et al., 2023).
Huachansu Injection is derived from the dried skin of the Bufo gargarizans and has traditionally been used for its detoxifying, anti-inflammatory, and analgesic properties. It is commonly applied in the treatment of advanced-stage cancers and chronic hepatitis B. Huachansu has been shown to inhibit liver cancer cell growth by reducing the activity and expression of glucose-6-phosphate dehydrogenase (G6PD), thereby disrupting the pentose phosphate pathway (Wu Q. et al., 2024). It can also induce apoptosis in gastric cancer cells by increasing ROS levels and suppressing proteasome activity (Deng et al., 2024). Yang et al. were the first to demonstrate that Huachansu Injection induces ferroptosis in multiple myeloma (MM) via the NRF2/HO-1 signaling pathway. Specifically, Huachansu promotes nuclear translocation of NRF2, which activates the transcription of antioxidant genes such as HO-1. HO-1 catalyzes the degradation of heme, leading to the release of free iron, which triggers the Fenton reaction and the accumulation of lipid peroxides such as MDA, ultimately initiating ferroptosis. Moreover, Huachansu downregulates the expression of SLC7A11 and GPX4, weakening the cellular capacity to eliminate lipid peroxides and thereby preventing the suppression of ferroptosis (Yang et al., 2025) (Table 2).
TABLE 2
| Active compounds of TCM | Chemical structure | Molecular formula | CAS number | Animal or cell type | Dosage of drugs used | anti-tumor mechanism | References |
|---|---|---|---|---|---|---|---|
| Shikonin |  | C16H16O5 | 517–89–5 | The human MM cell lines RPMI 8226 and U266; Nod-SCID mice of SPF grade | 4 mg/kg | Ferroptosis and immunogenic cell death are induced in MM cells via GOT1-mediated ferritinophagy | Li et al. (2022b) |
| Apigenin |  | C15H10O5 | 520–36–5 | MM cell line NCI-H929 | 5, 10, 20 or 40 μM | Apigenin can induce ferroptosis in the MM NCI-H929 cell line, but the specific mechanism requires further investigation | Adham et al. (2021a) |
| Artesunate |  | C19H28O8 | 88495–63–0 | The human MM cell lines MM1S (Cat. No. iCell-h291), RPMI8226 (Cat. No. iCell-h183) | 40 μM | Artesunate can trigger ferroptosis in MM by inhibiting the nuclear localization of SREBP2 and downregulating IPP and GPX4 | Liang et al. (2023) |
| Ethanol extract of Eclipta prostrata | -- | -- | -- | The human MM cell lines RPMI-8226 and U266, Nod-SCID mice of SPF grade | 50 mg/kg、100 mg/kg | Eclipta prostrata can induce ferroptosis in MM cells through the Keap1/Nrf2/HO-1 axis | Li et al. (2024c) |
| Thymus serpyllum extract | -- | -- | -- | MM cell line NCI-H929 | 0.001–100 μg/mL | The ferroptosis inhibitors abrogated cytotoxicity of the extracts, indicating that ferroptosis played a role in the process of cell death | Adham et al. (2020) |
| Arctium lappa extract | -- | -- | -- | MM cell line NCI-H929 | |||
| Andrographolide |  | C20H30O5 | 5508–58–7 | The human MM cell lines RPMI-8226 and U266 | 20 or 40 μM | Andrographolide induced ferroptosis in multiple myeloma cells by regulating the P38/Nrf2/HO-1 pathway | Li et al. (2023b) |
| Ginsenoside Rh4 |  | C36H60O8 | 174721–08–5 | MM cell line NCI-H929 | 25, 50, 100 and 200 μM | Ginsenoside Rh4 induces ferroptosis in MM and inhibits its malignant progression by regulating SIRT2 | Ying et al. (2023) |
| Nitidine chloride |  | C21H18ClNO4 | 13063–04–2 | The human MM cell lines RPMI-8226 and U266, mouse MM SP2/0 cells and mouse models | 2, 4, 6 and 8 μM; 4.5 mg/kg, 6 mg/kg | Nitidine chloride induces ferroptosis in MM cells by targeting ABCB6 and suppressing the PI3K/AKT pathway | Yin et al. (2023) |
| Huachansu |  | C26H34O7 | 1108–68–5 | The human MM cell lines ARP-1, CAG and JJN3 | 0.09 μg/mL, 0.18 μg/mL, 0.36 μg/mL | Huachansu Injection induces ferroptosis in multiple myeloma through NRF2/HO-1 signaling pathway | Yang et al. (2025) |
Summary of the mechanisms by which natural products intervene in MM through regulating ferroptosis.
GOT1, Glutamic-oxaloacetic transaminase 1; SREBP2, Sterol regulatory element binding protein 2; IPP, Isopentenyl pyrophosphate; Keap1, kelch-like ECH-associated protein 1; Nrf2, Nuclear factor erythroid 2-related factor 2; HO-1, Heme oxygenase-1; SIRT2, Sirtuin 2; ABCB6, ATP-binding cassette sub-family B member 6.
7.2 Proteasome inhibitors
In MM cells, high proteasome activity is a crucial factor for degrading misfolded immunoglobulins to ensure cell survival. As a proteasome inhibitor, bortezomib has been clinically used to treat patients with MM. Bortezomib exerts its therapeutic effects in MM by binding to the β5 subunit of the 26S proteasome to inhibit the ubiquitin–proteasome pathway, blocking activation of the Nuclear factor kappa-light-chain-enhancer (NF-κB) of activated B cells signaling pathway, inducing mitochondrial apoptosis, and suppressing angiogenesis and bone destruction within the tumor microenvironment. However, the autophagic process activated by the accumulation of intracellular misfolded immunoglobulins leads to bortezomib resistance in multiple myeloma (Cengiz Seval and Beksac, 2018).
MM cells exhibit higher intracellular iron and ferritin levels compared to normal cells. Bortezomib can trigger ferritin degradation, increase intracellular free Fe2+, and promote ferroptosis in MM cells. Further mechanistic studies have shown that bortezomib effectively increases the level of NCOA4 by preventing proteasomal degradation, confirming its role in enhancing ferritinophagy. Moreover, the combined use of bortezomib and the ferroptosis inducer RSL - 3 can synergistically promote ferroptosis in MM cell lines. The synergistic effect of bortezomib and RSL - 3 in MM cells can be successfully counteracted by using the ferroptosis inhibitor liproxstatin - 1 (Zhang et al., 2024f).
Treatment with docosahexaenoic acid or eicosapentaenoic acid (DHA/EPA) can enhance the sensitivity of MM cells to bortezomib. Compared with the combined treatment of DHA/EPA and bortezomib, the sole application of DHA/EPA can significantly reduce the intracellular GSH level and alter the expression of metabolites and key enzymes related to GSH metabolism. RNA - seq indicates that DHA/EPA may enhance the efficacy of bortezomib by activating ferroptosis - related signaling pathways, suggesting that ferroptosis may play a positive role in improving drug sensitivity and overcoming drug resistance in the treatment of MM (Chen J. et al., 2021).
In conclusion, ferroptosis - related genes may become important factors for MM prognosis assessment, and more relevant genes still need to be explored through experiments. Although the above evidence proves the important role of ferroptosis in MM, further research is still needed to determine the ferroptosis inducers applicable in the human body, as well as their concentrations and application methods, in order to exploit the potential of ferroptosis as a new and generalizable clinical strategy.
7.3 Immunomodulatory drugs
Fingolimod (FTY720) was initially used as an immunosuppressant for the treatment of multiple sclerosis (Cengiz Seval and Beksac, 2018). Fingolimod exerts its therapeutic effects in multiple sclerosis by activating sphingosine-1-phosphate receptors (S1PRs) and inducing their internalization, thereby blocking lymphocyte egress from lymphoid tissues to the periphery. This reduces the infiltration of autoreactive T cells into the central nervous system, ultimately attenuating inflammatory damage and slowing disease progression (Kappos et al., 2010). However, subsequent studies have revealed its extensive anti - tumor effects (Hasan Ali et al., 2020). Research has shown that FTY720 can reduce the mRNA and protein levels of GPX4 and SLC7A11 in MM cells. As key regulators of ferroptosis, GPX4 and SLC7A11 are highly expressed in MM cells, indicating that the induction of MM cell death by FTY720 is likely related to the ferroptosis pathway. Further studies have demonstrated that FTY720 activates PP2A to dephosphorylate the AMP - activated protein kinase subunit α (AMPKα) and reduces the expression of phosphorylated eukaryotic elongation factor 2 (eEF2), ultimately leading to MM cell death. FTY720 induces ferroptosis and autophagy through the PP2A/AMPK signaling pathway, and these two modes of cell death can promote each other, providing a new strategy for the treatment of MM (Zhong et al., 2020).
7.4 Other antitumor chemicals
Adapalene (ADA), a third-generation retinoid, is commonly used for the treatment of acne vulgaris. ADA selectively activates retinoic acid receptors (RAR-β/γ), thereby inhibiting excessive proliferation and differentiation of keratinocytes, reducing follicular hyperkeratinization, and downregulating inflammatory cytokines such as IL-6, ultimately contributing to its therapeutic effects in acne (Waugh et al., 2004). Recent studies have shown that ADA induces dose-dependent cell death in MM cells (Cao et al., 2024). ADA induces ferroptosis in MM cells by downregulating the protein expression of GPX4 and SLC7A11, which are key ferroptosis marker proteins. Moreover, studies have found that ADA is beneficial in restoring the sensitivity of cells from patients with bortezomib - resistant relapsed or refractory MM to bortezomib. ADA can disrupt the activation of the NF-κB pathway triggered by bortezomib and promote cell death in the bortezomib - resistant MM subpopulation (Cao et al., 2024).
Zhang et al. investigated the effects of the novel methyltransferase G9a inhibitor DCG066 on MM cells. They found that after DCG066 intervention, the levels of ROS, iron, and MDA in MM cells increased significantly, while the level of GSH decreased. The protein expression levels of SLC7A11, GPX4, Nrf2, and HO - 1 were significantly reduced. These phenomena could be reversed by the ferroptosis inhibitor Ferrostatin-1 and the Nrf2 activator tert - butylhydroquinone, which confirmed that DCG066 can inhibit the proliferation of MM cells and induce ferroptosis through the Nrf2/HO - 1 signaling pathway (Zhang et al., 2024g).
Disulfiram (DSF) is an FDA-approved drug for the treatment of alcohol dependence. It exerts its effect by inhibiting aldehyde dehydrogenase (ALDH), thereby blocking the conversion of ethanol to acetic acid. This leads to the accumulation of acetaldehyde in the body, causing adverse reactions such as nausea, vomiting, and palpitations, which help establish an aversive response to alcohol consumption (Heather, 1989). In recent years, DSF has shown emerging therapeutic potential in hematological malignancies due to its low toxicity and multi-target properties (Weiser Drozdkova and Smesny Trtkova, 2021). Arkan et al. found that treatment with disulfiram (DSF) increased the production of cytoplasmic and mitochondrial ROS in MM cells, led to the loss of mitochondrial membrane potential and an increase in the level of lipid peroxidation, and caused a significant downregulation of ferroptosis - related genes including GPX4 (Arkan and Akcora-Yildiz, 2025). The ferroptosis inhibitor liproxstatin - 1 could alleviate DSF - induced ferroptosis in MM cells by promoting the upregulation of GPX4. DSF can also overcome carfilzomib resistance by increasing the level of lipid peroxidation and exert a synergistic effect with carfilzomib in carfilzomib - resistant MM cell lines, indicating the potential of DSF in the treatment of MM.
T-5224 is an activator protein-1 (AP-1) inhibitor that specifically blocks the DNA-binding activity of the c-Fos/c-Jun heterodimer, thereby suppressing the expression of downstream pro-inflammatory cytokines such as TNF-α and matrix metalloproteinases like MMP-3. Through this mechanism, T-5224 exerts anti-inflammatory, antioxidant, and anti-degenerative effects (Sasakura et al., 2024). Previous studies have shown that T - 5224 can inhibit the proliferation of MM cells and induce their apoptosis (Tang et al., 2023). Tang et al. found that T - 5224 - induced MM cell death could be reversed by the ferroptosis - specific inhibitor ferropstatin-1, and T - 5224 reduced the protein levels of GPX4 and SLC7A11, the key regulators of ferroptosis in MM cells. Further research revealed that T - 5224 reduced the phosphorylation of components in the PI3K and AKT signaling pathways, suggesting that T - 5224 may induce ferroptosis in MM cells by regulating the PI3K/AKT signaling pathway (Tang S. et al., 2024) (Table 3).
TABLE 3
| Anti-tumor chemical | Chemical structure | Molecular formula | CAS number | Animal or cell type | Dosage of drugs used | Anti-tumor mechanism | References |
|---|---|---|---|---|---|---|---|
| Bortezomib |  | C19H25BN4O4 | 179324–69–7 | Human MM cell lines ARP1 and OCI-My5 | 1.5 nM、3 nM | Bortezomib may enhance NCOA4-mediated ferritinophagy by inhibiting the proteasome, thereby increasing free Fe2+ levels and promoting ferroptosis in MM cells | Zhang et al. (2024f) |
| Fingolimod (FTY720) |  | C19H33NO2 | 162359–55–9 | Human MM cell lines U266 and RPMI8226 | 5、10、15 μM | FTY720 induces ferroptosis and autophagy via PP2A/AMPK pathway in MM cells | Zhong et al. (2020) |
| Adapalene |  | C28H28O3 | 106685–40–9 | Human H929 and LP-1 MM cell lines | 20、40、60、80、100 μmol/L | Adapalene induces ferroptosis in MM cells by downregulating the protein expression of GPX4 and SLC7A11 | Cao et al. (2024) |
| DCG066 |  | C30H31F6N3O2 | 494786–13–9 | Human MM cell lines ARH-77 and RPMI-8226 | 5 µM | DCG066 inhibits MM proliferation and induces ferroptosis via the Nrf2/HO-1 pathway | Zhang et al. (2024g) |
| Disulfiram |  | C10H20N2S4 | 97–77–8 | Human MM cell lines NCI H929, U266, and RPMI 8226 | 5、10、25、50 µM | DSF induces ferroptosis in MM cells by downregulating GPX4, increasing cytosolic and mitochondrial ROS, and elevating lipid peroxidation | Arkan and Akcora-Yildiz (2025) |
| T-5224 |  | C29H27NO8 | 530141–72–1 | Human MM cell lines ARP1 and RPMI 8226; Nod-SCID mice of SPF grade | 10、20 µM (in vitro) 20 mg/kg (in vivo) | T-5224 reduces the protein levels of key ferroptosis regulators GPX4 and SLC7A11 in MM cells and induces ferroptosis in MM cells through the PI3K/AKT pathway | Tang et al. (2024a) |
Mechanisms of antitumor chemicals in treating MM through regulation of ferroptosis-related pathways.
NCOA4, Nuclear receptor coactivator 4; PP2A, Protein phosphatase 2A; AMPK, AMP activated protein kinase; DSF, Disulfiram.
In another study, MM cells harboring the t (4; 14) translocation exhibited marked sensitivity to Class II ferroptosis inducers (Class II FINs), whereas t (4; 14)-negative cells were resistant. This selective cytotoxicity is attributed to the overexpression of MMSET, a histone methyltransferase upregulated due to the t (4; 14) translocation. MMSET enhances transcriptional activation of ACSL4, facilitating the incorporation of polyunsaturated fatty acids (PUFAs) into membrane phospholipids. PUFA enrichment increases oxidative damage susceptibility, thereby heightening ferroptosis sensitivity in t (4; 14)-positive MM cells.In vitro assays confirmed that combining Class II FINs with bortezomib results in enhanced ferroptosis induction and significantly reduced viability of t (4; 14)-positive MM cells. As a genetically guided therapeutic strategy, this approach holds great clinical promise. Moreover, MMSET and ACSL4 expression levels may serve as predictive biomarkers for Class II FIN responsiveness, providing a basis for patient stratification (Zhang J. et al., 2024).
8 Regulatory effects of existing MM therapies on the ferroptosis pathway
Currently, no drugs directly targeting the ferroptosis pathway have been approved for the treatment of MM. This review summarizes investigational agents targeting ferroptosis in MM and highlights recent progress in understanding how existing MM therapies modulate ferroptosis (Table 4).
TABLE 4
| Drug Name | Drug Type | Development Stage | Mechanism of Action in Ferroptosis | Target Disease | Study Type | Reference |
|---|---|---|---|---|---|---|
| Bortezomib (combined with RSL3) | Proteasome Inhibitors | Approved Drug | Inhibition of proteasome activity, activates NCOA4-mediated ferritinophagy, leading to the release of free Fe2+ and enhanced accumulation of lipid peroxides | RRMM | Clinical trial, in vitro experiment | Zhang et al. (2024f) |
| DHA/EPA | Omega-3 Fatty Acids | Drug in preclinical study | Significantly reduces intracellular GSH levels, leading to GPX4 inactivation and accumulation of lipid peroxides, thereby promoting ferroptosis; reverses bortezomib resistance | MM resistant to bortezomib | In vitro experiment, in vivo experiment | Chen et al. (2021a) |
| Fingolimod | Immunomodulatory Drugs | Approved Drug | Reduces the expression of GPX4 and SLC7A11 in MM cells and induces ferroptosis and autophagy via the PP2A/AMPK pathway | MM | In vitro experiment | Zhong et al. (2020) |
| Adapalene | Third-generation retinoid | Approved Drug | Adapalene induces ferroptosis by downregulating GPX4 and SLC7A11 expression, leading to glutathione depletion and lipid peroxide accumulation | MM resistant to bortezomib | In vitro experiment | Cao et al. (2024) |
| DCG066 | Methyltransferase G9a inhibitor | Drug in preclinical study | DCG066 activates the Nrf2/HO-1 pathway, reduces the levels of GSH, SLC7A11, and GPX4, increases ROS and MDA accumulation, and induces ferroptosis | MM | In vitro experiment | Zhang et al. (2024g) |
| Disulfiram | Aldehyde dehydrogenase inhibitor | FDA-approved; preclinical in MM | Disulfiram induces ferroptosis by inhibiting GPX4 activity and promoting ROS and lipid peroxide accumulation, primarily through disruption of redox homeostasis | MM BCR-ABL + leukemia Myelodysplastic syndromes Nasopharyngeal Cancer Triple-Negative Breast Cancer Hepatocellular carcinoma Non-small cell lung cancer Glioblastoma Bladder Cancer | Clinical trial, in vitro experiment, in vivo experiment | Li et al. (2020c),2024a; Qiu et al. (2020),Ren et al. (2021),Chu et al. (2023),Liu et al. (2024),Sharma et al. (2024),Zeng et al. (2024) |
| T-5224 | Activator protein-1 inhibitor | Drug in Clinical Trial | T-5224 reduces the protein levels of GPX4 and SLC7A11 in MM cells and induces ferroptosis through the PI3K/AKT pathway | MM | In vitro experiment, in vivo experiment | Tang et al. (2024a) |
| Class II FINs | Ferroptosis inducers | Drug in preclinical study | Class II FINs target the t (4; 14) translocation–driven MMSET–ACSL4 axis to promote the synthesis of PUFA-PLs, thereby enhancing lipid peroxidation sensitivity. In combination with bortezomib, they further induce ferroptosis in t (4; 14)-positive MM cells by increasing Fe2+ release and suppressing GPX4-mediated antioxidant defense | t (4; 14)-positive MM | Clinical trial, in vitro experiment, in vivo experiment | Zhang et al. (2024c) |
| Carfilzomib | Proteasome Inhibitors | Approved Drug | The combination of carfilzomib and125I seed radiation promotes ferroptosis by enhancing intracellular Fe2+ accumulation and downregulating GPX4 expression | Esophageal squamous cell carcinoma | In vitro experiment, in vivo experiment | Wang et al. (2025) |
| PROTAC-PD-Q2 | -- | Treatment in preclinical study | The PROTAC-PD-Q2 probe induces ferroptosis by targeting GPX4 for degradation via the ubiquitin–proteasome system, thereby impairing its ability to repair lipid peroxides, leading to ROS accumulation and triggering a Fenton reaction cascade | -- | In vitro experiment | Zhu et al. (2023) |
| Selinexor | Selective Inhibitor of Nuclear Export | Approved Drug | In CML, selinexor inhibits the nuclear export protein XPO1 to block the NF-κB signaling pathway, downregulate GPX4 expression, and activate NCOA4-mediated ferritinophagy. In combination with ferroptosis inducers, it enhances lipid peroxidation, thereby overcoming drug resistance and inducing ferroptosis | Chronic myeloid leukemia | In vitro experiment | Cui et al. (2024) |
| Venetoclax | BCL-2 inhibitor | Approved Drug | Venetoclax induces apoptosis by inhibiting BCL-2 and indirectly enhances the sensitivity of AML cells to ferroptosis. When combined with ferroptosis inducers, it synergistically promotes lipid peroxide accumulation, overcoming the antioxidant defense threshold of resistant cells | Acute myeloid leukemia | In vitro experiment, in vivo experiment | Shi et al. (2025) |
Development of MM therapeutic agents targeting ferroptosis-related pathways and summary of studies on ferroptosis regulation by approved MM drugs in other diseases.
RRMM, Relapsed/refractory multiple myeloma; DHA/EPA, Docosahexaenoic acid or eicosapentaenoic acid; MDA, Malondialdehyde; FDA, Food and Drug Administration; MMSET, MM SET domain-containing protein; PROTAC, PROteolysis targeting chimera; CML, Chronic myeloid leukemia; XPO1, Exportin 1; NF-κB, Nuclear factor kappa-light-chain-enhancer of activated B cells; BCL-2, B-cell lymphoma-2; AML, Acute myeloid leukemia.
Studies have shown that the combination of the proteasome inhibitor carfilzomib and iodine-125 (125I) seed radiation exhibits potent antitumor effects. Mechanistically, 125I seed radiation induces intracellular accumulation of Fe2+ and lipid peroxides, but also upregulates ferroptosis suppressors SLC7A11 and GPX4. The combination therapy enhances intracellular Fe2+ accumulation and downregulates GPX4 expression, thereby promoting ferroptosis. However, current evidence is limited to solid tumors, and its efficacy in MM remains to be further investigated (Wang et al., 2025). In addition, in vitro experiments suggest that ferroptosis inducers RSL3 and ML162 may synergistically enhance the cytotoxicity of bortezomib and lenalidomide in the MM cell line RPMI-8226, underscoring the therapeutic relevance of ferroptosis-associated pathways in MM (Gao et al., 2023). Pomalidomide, the third-generation IMiD following thalidomide and lenalidomide, has recently been employed in a PROteolysis TArgeting Chimera (PROTAC) strategy. A PROTAC probe named PD-Q2, composed of pomalidomide and the GPX4 inhibitor ML162, effectively induces degradation of GPX4 via the ubiquitin–proteasome system, leading to ROS accumulation and significant suppression of MM cell growth. These findings suggest that GPX4-targeting PROTACs hold promise as novel agents for ferroptosis-based therapy in MM (Zhu et al., 2023).
Selinexor is a small-molecule inhibitor that targets the nuclear export protein Exportin 1 (XPO1). Selinexor-based combination regimens have been recommended by multiple clinical guidelines for the treatment of relapsed/refractory multiple myeloma. In addition, selinexor can block nucleocytoplasmic transport signals in leukemia stem cells and when combined with imatinib, induces cell death in chronic myeloid leukemia (CML) cells. However, drug resistance frequently leads to disease progression. Cui et al. identified a subpopulation within selinexor-resistant cells that exhibits cancer stem cell–like properties, characterized by reduced GPX4 activity and decreased SLC7A11 expression, resulting in impaired GSH synthesis. These cells are resistant to apoptosis-inducing agents but highly sensitive to ferroptosis inducers. Further investigation revealed that co-treatment with selinexor and the ferroptosis inducer RSL3 synergistically eliminates resistant cells through dual mechanisms: inhibition of nuclear export and induction of lipid peroxidation. This study, using single-cell analysis, was the first to reveal a connection between selinexor resistance and ferroptosis evasion in CML, suggesting a novel strategy to reverse drug resistance through ferroptosis induction in hematological malignancies such as MM (Cui et al., 2024).
Venetoclax, a BCL-2 inhibitor, promotes apoptosis in tumor cells. Preclinical data have shown its promising efficacy in both newly diagnosed MM and t (11; 14)-positive MM subtypes (Kaufman et al., 2021). Scavenger receptor class B type 1 (SR-B1), a key receptor in cholesterol metabolism, is significantly upregulated in acute myeloid leukemia (AML) and correlates with poor prognosis. SR-B1 deletion reduces cellular cholesterol uptake and inhibits GPX4 activity. Moreover, SR-B1 deficiency disrupts cholesterol homeostasis, enhances PUFA oxidation, increases lipid ROS, and triggers ferroptosis, while also downregulating BCL-2 expression. Further studies revealed that SR-B1 inhibition can reverse venetoclax resistance and promote AML cell death through ferroptosis. Targeting SR-B1 offers a novel therapeutic strategy in hematologic malignancies. The combination of the SR-B1 inhibitor Block Lipid Transport-1 (BLT-1) and venetoclax has entered a Phase I clinical trial (Shi et al., 2025).
Collectively, these findings highlight the emerging significance of ferroptosis as a therapeutic vulnerability in hematologic malignancies. The integration of ferroptosis-targeted strategies—either through small-molecule inducers, pathway modulators, or combination therapies—may offer novel avenues to overcome drug resistance and improve treatment outcomes in MM and related disorders.
9 Toxicology and side effects
Current evidence reveals an intrinsic paradox between therapeutic efficacy and safety profiles of bioactive compounds - no identified pharmacologically active agent completely avoids non-specific off-target effects on healthy tissues (Guo C. et al., 2023). While natural products demonstrate multi-component synergism with multi-target and multi-pathway regulatory effects on ferroptosis in MM intervention, their complex phytochemical matrices inherently harbor potential toxicological risks and adverse reaction profiles. SHK, as an inhibitor of uridine 5′-diphosphateglucuronosyltransferase, exerts toxic effects on liver tissues (Cheng et al., 2019). An in vitro study demonstrated that shikonin has potent cytotoxicity against 15 types of cancer cells through mitochondrial dysfunction and can induce apoptosis. However, in the same study, shikonin was also found to be toxic to normal cell lines (Wiench et al., 2012). Another study revealed that shikonin induces erythrocyte death by influencing Ca2+ influx and ceramide formation (Lupescu et al., 2014). SHK has also been identified as a mixed and competitive inhibitor of cytochrome P450 enzymes, which may lead to an increased risk of drug interactions and toxicity since these enzymes play a crucial role in drug metabolism (Lupescu et al., 2014). In addition, when administered orally, the LD50 value of shikonin is greater than 1 g/kg, indicating relatively low toxicity. In contrast, the LD50 values for intraperitoneal and intravenous injections are 20 mg/kg and 16 mg/kg respectively, suggesting higher toxicity (Yadav et al., 2022). These results indicate that the administration route of shikonin has a significant impact on its toxicity level. The oral route results in lower absorption rate and less toxicity due to first-pass metabolism, while the injection routes are more toxic as the drug directly enters the circulatory system. Other factors such as metabolism, toxicokinetics, and experimental uncertainties can also affect the drug’s toxicity level.
AP demonstrates potential therapeutic efficacy in MM management, though its toxicity profile and adverse effects warrant critical consideration, particularly regarding cellular, reproductive, and renal impacts. Acute toxicity studies identify 500 mg/kg oral AP as the maximum safe dose (Al Batran et al., 2013), while contrasting evidence shows non-toxicity at 5 mg/kg but exacerbated CCl4-induced hepatotoxicity and weight loss at 50 mg/kg in healthy mice (Lin et al., 2018). A meta-analysis documents 26 cases of AP-induced acute kidney injury manifesting as abdominal pain, nausea/vomiting, and oliguria, with histopathology suggesting acute tubular necrosis (Zhang et al., 2014). Reproductive toxicology reveals that prolonged 25–50 mg/kg AP administration in male rats induces sperm count reduction, motility impairment, and teratozoospermia - though this observation suggests potential male contraceptive applications (Akbarsha and Murugaian, 2000). Synergistic pharmacological interactions emerge with Api enhancing antitumor drug efficacy while reducing toxicity: gemcitabine-Api coadministration exerts superior tumor growth inhibition compared to monotherapy in neoplastic cell lines (Johnson and Gonzalez de Mejia, 2013), yet paradoxically, kaempferol-Api combination only suppresses tumor growth in vitro while increasing tumor burden in vivo (Tang et al., 2017). Adapalene exhibits oral LD50 > 10 mL/kg in rodents, demonstrating retinoid-like side effects during chronic use. Dose-dependent correlations emerge for ovarian gonadomas/thyroid carcinomas in female rats and benign/malignant adrenal medullary pheochromocytomas in males during oral trials (Piérard et al., 2009).
In conclusion, while AP, SHK, ADA, and related compounds exhibit multi-faceted pharmacological potential, their toxicological profiles and adverse effect manifestations remain clinically significant. Elucidating both therapeutic mechanisms and toxicity pathways proves essential for novel therapeutic development and enhancing clinical safety parameters in pharmacological applications.
10 Conclusion and outlook
MM, a B-cell-derived malignant clonal proliferation of plasma cells, ranks as the second most prevalent hematologic malignancy, accounting for 15%–20% of hematological cancer-related mortality (Nooka et al., 2024). This clinically heterogeneous entity is characterized by clonal plasma cell expansion and monoclonal immunoglobulin overproduction, with frequent relapse patterns. Ferroptosis, an emerging regulated cell death modality defined by iron-dependent lipid ROS accumulation, has shown growing therapeutic potential in oncology. This review delineates ferroptosis’s multifaceted roles in MM progression through iron metabolism, oxidative stress, and lipid peroxidation pathways, while systematically summarizing experimental evidence of natural product extracts and antineoplastic compounds targeting MM ferroptosis. However, the intricate pathogenesis of MM and its inherent heterogeneity result in complex interplay between ferroptosis signaling pathways, necessitating further mechanistic elucidation of their crosstalk and regulatory networks. Current ferroptosis research in MM remains confined to preclinical investigations, with clinical translation requiring rigorous validation through human trials. Bridging this translational gap between ferroptosis biology and clinical MM therapeutics represents a critical future research frontier.
While NPs demonstrate potential to modulate ferroptosis through multidimensional mechanisms influencing MM progression, critical challenges hinder clinical translation. Firstly, current investigations remain predominantly confined to single-target/pathway mechanistic elucidation, lacking both comprehensive systems pharmacology analyses of network regulation and high-level evidence-based medical validation. The incomplete clinical evidence chain, heavily reliant on in vitro studies, necessitates prioritized development of ferroptosis-specific biomarkers and innovation in spatiotemporal controlled-release technologies for ferroptosis inducers. These advancements could accelerate discovery of novel ferroptosis pathway-targeting antitumor compounds. Concurrently, substantial research efforts must elucidate the ferroptosis-modulating mechanisms of existing antineoplastic agents through integrated preclinical-clinical investigations, which is crucial for developing novel effective anti-myeloma therapeutics. Secondly, implementing high-quality multicenter, double-blind randomized controlled trials remains imperative to rigorously evaluate the therapeutic efficacy and safety profile of NPs in modulating ferroptosis-related pathways for MM management, thereby ensuring scientifically robust and clinically translatable outcomes. Thirdly, comprehensive characterization of NPs dose-response relationships is essential to establish optimal therapeutic thresholds while mitigating potential long-term toxicity and safety risks. Enhanced investigation into NP toxicological profiles, coupled with innovations in targeted delivery systems to prolong circulatory residence time, could significantly improve treatment precision. Concurrent standardization of NP quality control protocols must be prioritized to ensure reproducibility across preclinical and clinical studies.
In this review, we summarize investigational agents targeting ferroptosis-related pathways in MM and highlight studies in which approved MM therapies modulate ferroptosis in other diseases. Current evidence suggests that MM cells, particularly those resistant to standard therapies or harboring genetic alterations such as the t (4; 14) translocation, are highly sensitive to ferroptosis. However, clinical translation of this mechanism remains in its infancy.
Future research should focus on developing highly selective and potent ferroptosis inducers suitable for hematologic malignancies. These may include small molecules targeting GPX4, SLC7A11, or ferritinophagy regulators like NCOA4, as well as compounds modulating iron metabolism or lipid peroxidation. PROTAC-based degraders and dual-target agents represent promising next-generation strategies. ACSL4 plays a dual role in MM: it promotes tumor growth by activating lipid synthesis, but also initiates ferroptosis by catalyzing the production of PUFA-CoA, a key substrate for lipid peroxidation (Zhang et al., 2023). Notably, ACSL4-deficient MM cells exhibit resistance to RSL3, while ACSL4-high cells are highly sensitive, supporting the rationale for targeting ferroptosis in ACSL4-overexpressing MM subtypes to overcome chemoresistance. Combination therapies also hold great potential. Ferroptosis inducers may synergize with proteasome inhibitors, immunomodulatory drugs, or BCL-2 inhibitors to enhance cytotoxicity and overcome resistance. Molecular stratification based on ferroptosis regulators such as MMSET or ACSL4 could further guide personalized treatment approaches. Finally, emerging technologies—including single-cell sequencing, spatial transcriptomics, and CRISPR-based functional screening—are expected to reveal the ferroptosis regulatory network within the MM tumor microenvironment. Advances in nanomedicine and targeted delivery systems will also be essential to improve therapeutic specificity and reduce toxicity.
However, several critical gaps and challenges remain in current research. The dynamic regulatory mechanisms of the tumor microenvironment (TME) are still unclear. The MM bone marrow microenvironment comprises diverse cell types and soluble factors. Recent studies have shown that GDF-15 secreted by M2-polarized tumor-associated macrophages promotes ferroptosis resistance in AML cells by modulating the SLC7A11/GPX4 axis (Lu and Liao, 2025); however, this mechanism has not yet been validated in MM, and the roles of other immune cells within the TME remain unexplored. Moreover, how intercellular communication in the TME dynamically regulates the stability of key ferroptosis molecules such as SLC7A11 and GPX4 remains poorly understood. Another major challenge lies in the genetic heterogeneity of MM. Current studies have only preliminarily revealed that in t (4; 14) translocation-positive patients, MMSET upregulation enhances ACSL4 transcription and ferroptosis sensitivity (Zhang J. et al., 2024). In contrast, ferroptosis regulatory mechanisms in other high-risk subtypes—such as del (17p), t (14; 16), and 1q21 amplification—remain largely unknown. In particular, how epigenetic modifications, such as DNA methylation, influence the expression of ferroptosis-related genes has yet to be elucidated.
Most current studies remain at the in vitro stage, which presents inherent limitations. The efficacy, safety, and pharmacokinetic profiles of ferroptosis inducers have not been systematically validated in humans. In vitro models fail to recapitulate the complexity of the bone marrow microenvironment and overlook key pharmacokinetic factors such as renal excretion, potentially leading to inaccurate dose prediction. Additionally, drug-resistant cell lines generated in vitro do not fully mimic the clonal evolution of tumors during relapse, which may compromise experimental accuracy. In clinical settings, ferroptosis inducers may pose challenges such as hepatotoxicity, immune interference, and off-target cell damage, all of which require optimization through structural refinement and targeted delivery strategies. Moreover, the high heterogeneity of MM remains a major obstacle, and currently, there is a lack of validated biomarkers to predict ferroptosis sensitivity, limiting precise therapeutic stratification.
In summary, targeting ferroptosis offers a novel pharmacological approach for MM treatment. Future research should focus on drug development, combination strategies, mechanistic studies, and biomarker discovery. The synergistic application of natural products (NPs) with chemotherapeutic agents and other treatment modalities may contribute to a more integrated therapeutic framework. Developing safer and more effective ferroptosis-based strategies holds great promise for patients with relapsed or refractory MM. However, the standardization and clinical translation of ferroptosis inducers remain challenging, highlighting the need for continued multidisciplinary efforts to advance precision medicine in MM.
Statements
Author contributions
JS: Conceptualization, Formal Analysis, Investigation, Writing – original draft. YL: Conceptualization, Data curation, Investigation, Writing – review and editing. WW: Data curation, Investigation, Writing – review and editing. GM: Investigation, Methodology, Writing – review and editing. CL: Investigation, Methodology, Writing – review and editing. LL: Investigation, Methodology, Writing – review and editing. YrW: Investigation, Methodology, Writing – review and editing. YaW: Conceptualization, Formal Analysis, Methodology, Resources, Supervision, Writing – review and editing. RX: Conceptualization, Formal Analysis, Methodology, Resources, Supervision, Writing – review and editing. SC: Conceptualization, Funding acquisition, Methodology, Project administration, Resources, Supervision, Writing – review and editing.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. This work was supported by a project at the departmental level in Shandong Province: Research on Epigenetic Mechanisms of Relapsed and Refractory Multiple Myeloma Patients Based on High - Throughput Methylation Chip Analysis, No. 202303041540.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Glossary
- AA
Arachidonic acid
- ACF
Arctium lappa
- ACSL4
Acyl-CoA synthetase long-chain family member 4
- ADA
Adapalene
- AdA
Adrenic acid
- AIFM2
Apoptosis - inducing factor mitochondria - associated 2
- AMPK
AMP-activated protein kinase
- Api
Apigenin
- ART
Artesunate
- BMSCs
Bone marrow mesenchymal stromal cells
- BTZ
Bortezomib
- CoQ10
Coenzyme Q10
- Cys
cysteine
- CP
Ceruloplasmin
- DHA
Dihydroartemisinin
- DHA/EPA
Docosahexaenoic acid or eicosapentaenoic acid
- DSF
Disulfiram
- EBP
emopamil binding protein
- EEEP
ethanol extract of Eclipta prostrata
- eEF2
eukaryotic elongation factor 2
- eEF2k
eukaryotic elongation factor 2 kinase
- EF
Ethyl acetate
- FPN1
Ferroportin 1
- FRGs
Ferroptosis-related genes
- FSP1
Ferroptosis suppressor protein 1
- FTY720
Fingolimod
- GCL
Glutamate-cysteine ligase
- Gln
Glutamine
- GLS
Glutaminase
- Glu
Glutamate
- GLUD1
Glutamate dehydrogenase
- GOT1
Glutamic-oxaloacetic transaminase 1
- GPX4
Glutathione peroxidase 4
- GSH
Glutathione
- GSSG
Oxidative glutathione
- HCC
Hepatocellular carcinoma
- HO-1
heme oxygenase-1
- IMiDs
immunomodulatory drugs
- IPP
Isopentenyl pyrophosphate
- IRF1
Interferon regulatory factor 1
- LILRB1
Leukocyte immunoglobulin-like receptor B1
- LIP
Labile iron pool
- LOXs
Lipoxygenases
- LPCAT3
Lysophosphatidylcholine acyltransferase 3
- LPOs
lipid peroxides
- MDA
Malondialdehyde
- MM
Multiple myeloma
- MUFAs
Monounsaturated fatty acids
- MVA
Mevalonate
- NADPH
Nicotinamide adenine dinucleotide phosphate
- NPs
Natural products
- Nrf2
Nuclear factor erythroid-2 related factor 2
- PE
Phosphatidylethanolamine
- PIs
Proteasome inhibitors
- PLOH
fatty alcohols
- PLOOH
Phospholipid hydroperoxides
- PP2A
Protein phosphatase 2A
- PUFA
Polyunsaturated fatty acid
- ROS
Reactive oxygen species
- RPE
Retinal pigment epithelial
- RT
Radiotherapy
- Sec
Selenocysteine
- SHK
Shikonin
- SLC1A5
Solute carrier family 1 member 5
- SLC7A11
Solute carrier family 7 member 11
- TCA cycle
Tricarboxylic acid cycle
- TCF
Thymus vulgaris
- Tf
Transferrin
- TfR1
Transferrin receptor 1
- TRCs
Tumor-reprogrammed cells
- ULK1
UNC-51 like kinase 1
- YAP1
Yes-associated protein 1.
References
1
AbeC.MiyazawaT.MiyazawaT. (2022). Current use of fenton reaction in drugs and food. Molecules27, 5451. 10.3390/molecules27175451
2
AboushoushaR.van der VeldenJ.HamiltonN.PengZ.MacPhersonM.EricksonC.et al (n.d.). Glutaredoxin attenuates glutathione levels via deglutathionylation of Otub1 and subsequent destabilization of system xC. Sci. Adv.9, eadi5192. 10.1126/sciadv.adi5192
3
AdhamA. N.AbdelfatahS.NaqishbandiA. M.MahmoudN.EfferthT. (2021a). Cytotoxicity of apigenin toward multiple myeloma cell lines and suppression of iNOS and COX-2 expression in STAT1-transfected HEK293 cells. Phytomedicine80, 153371. 10.1016/j.phymed.2020.153371
4
AdhamA. N.NaqishbandiA. M.EfferthT. (2021b). Cytotoxicity and apoptosis induction by Fumaria officinalis extracts in leukemia and multiple myeloma cell lines. J. Ethnopharmacol.266, 113458. 10.1016/j.jep.2020.113458
5
AdhamN.HegazyM. E.NaqishbandiA. M.EfferthT. (2020). Induction of apoptosis, autophagy and ferroptosis by Thymus vulgaris and Arctium lappa extract in leukemia and multiple myeloma cell lines. Molecules25, 5016. 10.3390/molecules25215016
6
AkbarshaM. A.MurugaianP. (2000). Aspects of the Male reproductive toxicity/male antifertility property of andrographolide in albino rats: effect on the testis and the cauda epididymidal spermatozoa. Phytotherapy Res.14, 432–435. 10.1002/1099-1573(200009)14:6<432::AID-PTR622>3.0.CO;2-I
7
Al BatranR.Al-BayatyF.Al-ObaidiM. M. J.AbdullaM. A. (2013). Acute toxicity and the effect of andrographolide on porphyromonas gingivalis-Induced hyperlipidemia in rats. Biomed. Res. Int.2013, 594012. 10.1155/2013/594012
8
AnandhanA.DodsonM.ShakyaA.ChenJ.LiuP.WeiY.et al (2023). NRF2 controls iron homeostasis and ferroptosis through HERC2 and VAMP8. Sci. Adv.9, eade9585. 10.1126/sciadv.ade9585
9
Anti-angiogenic effect of Shikonin (2020). Anti-angiogenic effect of shikonin in rheumatoid arthritis by downregulating PI3K/AKT and MAPKs signaling pathways (2020). J. Ethnopharmacol.260, 113039. 10.1016/j.jep.2020.113039
10
Apigenin inhibits recurrent bladder cancer (2025). Apigenin inhibits recurrent bladder cancer progression by targeting VEGF-β. Cancer Lett.620, 217676. 10.1016/j.canlet.2025.217676
11
ArkanC.Akcora-YildizD. (2025). FDA-Approved disulfiram induces ferroptosis via alteration of redox balance and lipid peroxidation and overcomes carfilzomib resistance in multiple myeloma. Leukemia and Lymphoma66, 250–261. 10.1080/10428194.2024.2422843
12
BaiX.DuanZ.DengJ.ZhangZ.FuR.ZhuC.et al (2024). Ginsenoside Rh4 inhibits colorectal cancer via the modulation of gut microbiota-mediated bile acid metabolism. J. Adv. Res.72, 37–52. 10.1016/j.jare.2024.06.028
13
BanaszkiewiczM.MałyszkoJ.BatkoK.Koc-ŻórawskaE.ŻórawskiM.DumnickaP.et al (2022). The key role of Hepcidin-25 in anemia in multiple myeloma patients with renal impairment. Med. Kaunas.58, 417. 10.3390/medicina58030417
14
BersukerK.HendricksJ. M.LiZ.MagtanongL.FordB.TangP. H.et al (2019). The CoQ oxidoreductase FSP1 acts parallel to GPX4 to inhibit ferroptosis. Nature575, 688–692. 10.1038/s41586-019-1705-2
15
BodineauC.ToméM.MurdochP. del S.DuránR. V. (2022). Glutamine, MTOR and autophagy: a multiconnection relationship. Autophagy18, 2749–2750. 10.1080/15548627.2022.2062875
16
BolatliG.TaşF.AlayuntN. Ö. (2023). Effects of Thymus vulgaris oil on sodium hypochlorite-induced damage in rats. Molecules28, 2164. 10.3390/molecules28052164
17
BordiniJ.GalvanS.PonzoniM.BertilaccioM. T. S.ChesiM.BergsagelP. L.et al (2017). Induction of iron excess restricts malignant plasma cells expansion and potentiates bortezomib effect in models of multiple myeloma. Leukemia31, 967–970. 10.1038/leu.2016.346
18
BordiniJ.MorisiF.CerrutiF.CascioP.CamaschellaC.GhiaP.et al (2020). Iron causes lipid oxidation and inhibits proteasome function in multiple myeloma cells: a proof of concept for novel combination therapies. Cancers (Basel)12, 970. 10.3390/cancers12040970
19
CamioloG.BarbatoA.GiallongoC.VicarioN.RomanoA.ParrinelloN. L.et al (2020). Iron regulates myeloma cell/macrophage interaction and drives resistance to bortezomib. Redox Biol.36, 101611. 10.1016/j.redox.2020.101611
20
CampanellaA.SantambrogioP.FontanaF.FrenquelliM.CenciS.MarcattiM.et al (2013). Iron increases the susceptibility of multiple myeloma cells to bortezomib. Haematologica98, 971–979. 10.3324/haematol.2012.074872
21
CaoX.XiangJ.ZhangQ.LiuJ.ZhouD.XuY.et al (2024). Multidimensional role of adapalene in regulating cell death in multiple myeloma. Front. Pharmacol.15, 1415224. 10.3389/fphar.2024.1415224
22
Cengiz SevalG.BeksacM. (2018). The safety of bortezomib for the treatment of multiple myeloma. Expert Opin. Drug Saf.17, 953–962. 10.1080/14740338.2018.1513487
23
ChenC.HeL.WangX.XiaoR.ChenS.YeZ.et al (2023). Leonurine promotes the maturation of healthy donors and multiple myeloma patients derived-dendritic cells via the regulation on arachidonic acid metabolism. Front. Pharmacol.14, 1104403. 10.3389/fphar.2023.1104403
24
ChenJ.DuanZ.LiuY.FuR.ZhuC. (2022a). Ginsenoside Rh4 suppresses metastasis of esophageal cancer and expression of c-Myc via targeting the Wnt/β-Catenin signaling pathway. Nutrients14, 3042. 10.3390/nu14153042
25
ChenJ.LiX.GeC.MinJ.WangF. (2022b). The multifaceted role of ferroptosis in liver disease. Cell. Death Differ.29, 467–480. 10.1038/s41418-022-00941-0
26
ChenJ.ZaalE. A.BerkersC. R.RuijtenbeekR.GarssenJ.RedegeldF. A. (2021a). Omega-3 fatty acids DHA and EPA reduce bortezomib resistance in multiple myeloma cells by promoting glutathione degradation. Cells10, 2287. 10.3390/cells10092287
27
ChenQ.KangJ.FuC. (2018). The Independence of and associations among apoptosis, autophagy, and necrosis. Signal Transduct. Target Ther.3, 18. 10.1038/s41392-018-0018-5
28
ChenX.KangR.KroemerG.TangD. (2021b). Broadening Horizons: the role of ferroptosis in cancer. Nat. Rev. Clin. Oncol.18, 280–296. 10.1038/s41571-020-00462-0
29
ChenY.LiuS.LiJ.LiZ.QuanJ.LiuX.et al (2020). The latest view on the mechanism of ferroptosis and its research progress in spinal cord injury. Oxid. Med. Cell. Longev.2020, 6375938. 10.1155/2020/6375938
30
ChenY.XuW.JinH.ZhangM.LiuS.LiuY.et al (2024). Nutritional glutamine-modified iron-delivery system with enhanced endocytosis for ferroptosis therapy of pancreatic tumors. ACS Nano18, 31846–31868. 10.1021/acsnano.4c08083
31
ChengY.TangS.ChenA.ZhangY.LiuM.WangX. (2019). Evaluation of the inhibition risk of shikonin on human and rat UDP-Glucuronosyltransferases (UGT) through the cocktail approach. Toxicol. Lett.312, 214–221. 10.1016/j.toxlet.2019.05.017
32
ChiangS.-K.ChenS.-E.ChangL.-C. (2018). A dual role of heme Oxygenase-1 in cancer cells. Int. J. Mol. Sci.20, 39. 10.3390/ijms20010039
33
ChuM.AnX.FuC.YuH.ZhangD.LiQ.et al (2023). Disulfiram/copper induce ferroptosis in triple-negative breast cancer cell line MDA-MB-231. Front. Biosci. Landmark Ed.28, 186. 10.31083/j.fbl2808186
34
CuiS.GhaiA.DengY.LiS.ZhangR.EgbulefuC.et al (2023). Identification of hyperoxidized PRDX3 as a ferroptosis marker reveals ferroptotic damage in chronic liver diseases. Mol. Cell.83, 3931–3939.e5. 10.1016/j.molcel.2023.09.025
35
CuiY.LiY.JiJ.HuN.MinK.YingW.et al (2024). Dynamic single-cell RNA-seq reveals mechanism of selinexor-resistance in chronic myeloid leukemia. Int. Immunopharmacol.134, 112212. 10.1016/j.intimp.2024.112212
36
DengY.-Q.GaoM.LuD.LiuQ.-P.ZhangR.-J.YeJ.et al (2024). Compound-composed Chinese medicine of huachansu triggers apoptosis of gastric cancer cells through increase of reactive oxygen species levels and suppression of proteasome activities. Phytomedicine123, 155169. 10.1016/j.phymed.2023.155169
37
DiergeE.DebockE.GuilbaudC.CorbetC.MignoletE.MignardL.et al (2021). Peroxidation of n-3 and n-6 polyunsaturated fatty acids in the acidic tumor environment leads to ferroptosis-mediated anticancer effects. Cell. Metab.33, 1701–1715.e5. 10.1016/j.cmet.2021.05.016
38
DixonS. J.LembergK. M.LamprechtM. R.SkoutaR.ZaitsevE. M.GleasonC. E.et al (2012). Ferroptosis: an iron-dependent form of nonapoptotic cell death. Cell.149, 1060–1072. 10.1016/j.cell.2012.03.042
39
DollS.FreitasF. P.ShahR.AldrovandiM.da SilvaM. C.IngoldI.et al (2019). FSP1 is a glutathione-independent ferroptosis suppressor. Nature575, 693–698. 10.1038/s41586-019-1707-0
40
DollS.PronethB.TyurinaY. Y.PanziliusE.KobayashiS.IngoldI.et al (2017). ACSL4 dictates ferroptosis sensitivity by shaping cellular lipid composition. Nat. Chem. Biol.13, 91–98. 10.1038/nchembio.2239
41
DolmaS.LessnickS. L.HahnW. C.StockwellB. R. (2003). Identification of genotype-selective antitumor agents using synthetic lethal chemical screening in engineered human tumor cells. Cancer Cell.3, 285–296. 10.1016/S1535-6108(03)00050-3
42
DongF.QuL.DuanZ.HeY.MaX.FanD. (2023). Ginsenoside Rh4 inhibits breast cancer growth through targeting histone deacetylase 2 to regulate immune microenvironment and apoptosis. Bioorg. Chem.135, 106537. 10.1016/j.bioorg.2023.106537
43
DongH.QiangZ.ChaiD.PengJ.XiaY.HuR.et al (2020). Nrf2 inhibits ferroptosis and protects against acute lung injury due to intestinal ischemia reperfusion via regulating SLC7A11 and HO-1. Aging12, 12943–12959. 10.18632/aging.103378
44
DuS.ZengF.DengG. (2023). Tumor neutrophils ferroptosis: a targetable immunosuppressive mechanism for cancer immunotherapy. Signal Transduct. Target Ther.8, 77. 10.1038/s41392-023-01357-z
45
FABP5 (2024). Artesunate-binding FABP5 promotes apoptosis in lung cancer cells via the PPARγ-SCD pathway (2024). Int. Immunopharmacol.143, 113381. 10.1016/j.intimp.2024.113381
46
FanX.YanY.LiY.SongY.LiB. (2024). Anti-tumor mechanism of artesunate. Front. Pharmacol.15, 1483049. 10.3389/fphar.2024.1483049
47
FangX.WangJ.HaoJ.LiX.GuoN. (2015). Simultaneous extraction, identification and quantification of phenolic compounds in Eclipta prostrata using microwave-assisted extraction combined with HPLC–DAD–ESI–MS/MS. Food Chem.188, 527–536. 10.1016/j.foodchem.2015.05.037
48
FengH.SchorppK.JinJ.YozwiakC. E.HoffstromB. G.DeckerA. M.et al (2020). Transferrin receptor is a specific ferroptosis marker. Cell. Rep.30, 3411–3423. 10.1016/j.celrep.2020.02.049
49
FengY.-Q.LiuX.ZuoN.YuM.-B.BianW.-M.HanB.-Q.et al (2024). NAD+ precursors promote the restoration of spermatogenesis in busulfan-treated mice through inhibiting Sirt2-regulated ferroptosis. Theranostics14, 2622–2636. 10.7150/thno.92416
50
ForcinaG. C.DixonS. J. (2019). GPX4 at the crossroads of lipid homeostasis and ferroptosis. PROTEOMICS19, 1800311. 10.1002/pmic.201800311
51
FreitasF. P.AlborziniaH.dos SantosA. F.NepachalovichP.PedreraL.ZilkaO.et al (2024). 7-Dehydrocholesterol is an endogenous suppressor of ferroptosis. Nature626, 401–410. 10.1038/s41586-023-06878-9
52
FuB.ShaoR.WangH.ChenG.BaiS.WangH. (2022). Integrated assessment of the clinical and biological value of ferroptosis-related genes in multiple myeloma. Cancer Cell. Int.22, 326. 10.1186/s12935-022-02742-4
53
FuX.WuH.LiC.DengG.ChenC. (2024). YAP1 inhibits RSL3-induced castration-resistant prostate cancer cell ferroptosis by driving glutamine uptake and metabolism to GSH. Mol. Cell. Biochem.479, 2415–2427. 10.1007/s11010-023-04847-4
54
GalyB.ConradM.MuckenthalerM. (2024). Mechanisms controlling cellular and systemic iron homeostasis. Nat. Rev. Mol. Cell. Biol.25, 133–155. 10.1038/s41580-023-00648-1
55
GaoD.LiuR.LvY.FengY.HongF.XuX.et al (2023). A novel ferroptosis-related gene signature for predicting prognosis in multiple myeloma. Front. Oncol.13, 999688. 10.3389/fonc.2023.999688
56
GaoM.MonianP.QuadriN.RamasamyR.JiangX. (2015). Glutaminolysis and transferrin regulate ferroptosis. Mol. Cell.59, 298–308. 10.1016/j.molcel.2015.06.011
57
GuoC.HuangQ.WangY.YaoY.LiJ.ChenJ.et al (2023a). Therapeutic application of natural products: NAD+ metabolism as potential target. Phytomedicine114, 154768. 10.1016/j.phymed.2023.154768
58
GuoH.SunJ.LiD.HuY.YuX.HuaH.et al (2019). Shikonin attenuates acetaminophen-induced acute liver injury via inhibition of oxidative stress and inflammation. Biomed. and Pharmacother.112, 108704. 10.1016/j.biopha.2019.108704
59
GuoY.ZhouM.MuZ.GuoJ.HouY.XuY.et al (2023b). Recent advances in shikonin for the treatment of immune-related diseases: anti-Inflammatory and immunomodulatory mechanisms. Biomed. and Pharmacother.165, 115138. 10.1016/j.biopha.2023.115138
60
Hasan AliO.BernerF.AckermannC. J.RingS. S.MoulinA.MüllerJ.et al (2020). Fingolimod and tumor-infiltrating lymphocytes in checkpoint-inhibitor treated cancer patients. Cancer Immunol. Immunother.70, 563–568. 10.1007/s00262-020-02693-7
61
HeY.-J.LiuX.-Y.XingL.WanX.ChangX.JiangH.-L. (2020). Fenton reaction-independent ferroptosis therapy via glutathione and iron redox couple sequentially triggered lipid peroxide generator. Biomaterials241, 119911. 10.1016/j.biomaterials.2020.119911
62
HeatherN. (1989). Disulfiram treatment for alcoholism. BMJ299, 471–472. 10.1136/bmj.299.6697.471
63
HenningY.BlindU. S.LarafaS.MatschkeJ.FandreyJ. (2022). Hypoxia aggravates ferroptosis in RPE cells by promoting the fenton reaction. Cell. Death Dis.13, 662. 10.1038/s41419-022-05121-z
64
HillK. S.SchulerE. E.EllingsonS. R.KolesarJ. M. (2023). Artesunate acts through cytochrome c to inhibit growth of pediatric AML cells. Sci. Rep.13, 22383. 10.1038/s41598-023-49928-y
65
HuangQ.RuY.LuoY.LuoX.LiuD.MaY.et al (2024). Identification of a targeted ACSL4 inhibitor to treat ferroptosis-related diseases. Sci. Adv.10, eadk1200. 10.1126/sciadv.adk1200
66
ImranM.Aslam GondalT.AtifM.ShahbazM.Batool QaisaraniT.Hanif MughalM.et al (2020). Apigenin as an anticancer agent. Phytotherapy Res.34, 1812–1828. 10.1002/ptr.6647
67
JiangH.WangL.ZhangQ.WangS.JiaL.ChengH.et al (2024). Bone marrow stromal cells dictate lanosterol biosynthesis and ferroptosis of multiple myeloma. Oncogene43, 1644–1653. 10.1038/s41388-024-03020-5
68
JiangL.KonN.LiT.WangS.-J.SuT.HibshooshH.et al (2015). Ferroptosis as a p53-mediated activity during tumour suppression. Nature520, 57–62. 10.1038/nature14344
69
JiangR.LuoS.ZhangM.WangW.ZhuoS.WuY.et al (2023). Ginsenoside Rh4 inhibits inflammation-related hepatocellular carcinoma progression by targeting HDAC4/IL-6/STAT3 signaling. Mol. Genet. Genomics298, 1479–1492. 10.1007/s00438-023-02070-w
70
JiangX.StockwellB. R.ConradM. (2021a). Ferroptosis: mechanisms, biology, and role in disease. Nat. Rev. Mol. Cell. Biol.22, 266–282. 10.1038/s41580-020-00324-8
71
JiangZ.-B.WangW.-J.XuC.XieY.-J.WangX.-R.ZhangY.-Z.et al (2021b). Luteolin and its derivative apigenin suppress the inducible PD-L1 expression to improve anti-tumor immunity in KRAS-Mutant lung cancer. Cancer Lett.515, 36–48. 10.1016/j.canlet.2021.05.019
72
JiaqiL.SiqingH.qinW.diZ.beiZ.jialinY. (2023). Andrographolide promoted ferroptosis to repress the development of non-small cell lung cancer through activation of the mitochondrial dysfunction. Phytomedicine109, 154601. 10.1016/j.phymed.2022.154601
73
JohnsonJ. L.Gonzalez de MejiaE. (2013). Interactions between dietary flavonoids apigenin or luteolin and chemotherapeutic drugs to potentiate anti-proliferative effect on human pancreatic cancer cells, in vitro. Food Chem. Toxicol.60, 83–91. 10.1016/j.fct.2013.07.036
74
JuJ.LiX.-M.ZhaoX.-M.LiF.-H.WangS.-C.WangK.et al (2023). Circular RNA FEACR inhibits ferroptosis and alleviates myocardial ischemia/reperfusion injury by interacting with NAMPT. J. Biomed. Sci.30, 45. 10.1186/s12929-023-00927-1
75
KaganV. E.MaoG.QuF.AngeliJ. P. F.DollS.CroixC. S.et al (2017). Oxidized arachidonic and adrenic PEs navigate cells to ferroptosis. Nat. Chem. Biol.13, 81–90. 10.1038/nchembio.2238
76
KanaanM. N.PileggiC. A.KaramC. Y.KennedyL. S.Fong-McMasterC.Cuperlovic-CulfM.et al (2024). Cystine/Glutamate antiporter xCT controls skeletal muscle glutathione redox, bioenergetics and differentiation. Redox Biol.73, 103213. 10.1016/j.redox.2024.103213
77
KapposL.RadueE.-W.O’ConnorP.PolmanC.HohlfeldR.CalabresiP.et al (2010). A placebo-controlled trial of oral fingolimod in relapsing multiple sclerosis. N. Engl. J. Med.362, 387–401. 10.1056/NEJMoa0909494
78
KaufmanJ. L.GasparettoC.SchjesvoldF. H.MoreauP.TouzeauC.FaconT.et al (2021). Targeting BCL‐2 with venetoclax and dexamethasone in patients with relapsed/refractory t(11;14) multiple myeloma. Am. J. Hematol.96, 418–427. 10.1002/ajh.26083
79
KimJ.KimY. C.FangC.RussellR. C.KimJ. H.FanW.et al (2013). Differential regulation of distinct Vps34 complexes by AMPK in nutrient stress and autophagy. Cell.152, 290–303. 10.1016/j.cell.2012.12.016
80
KimJ.KunduM.ViolletB.GuanK.-L. (2011). AMPK and mTOR regulate autophagy through direct phosphorylation of Ulk1. Nat. Cell. Biol.13, 132–141. 10.1038/ncb2152
81
KuroeT.WatanabeR.KojimaM.MorisueR.SuganoM.KuwataT.et al (2021). Evaluation of the morphological features and unfavorable prognostic impact of dirty necrosis in renal cell carcinoma. J. Cancer Res. Clin. Oncol.147, 1089–1100. 10.1007/s00432-020-03505-2
82
LeeI.-G.LeeJ.HongS.-H.SeoY.-J. (2023). Apigenin’s therapeutic potential against viral infection. FBL28, 237. 10.31083/j.fbl2810237
83
LeiG.ZhuangL.GanB. (2022). Targeting ferroptosis as a vulnerability in cancer. Nat. Rev. Cancer22, 381–396. 10.1038/s41568-022-00459-0
84
LiF.-J.LongH.-Z.ZhouZ.-W.LuoH.-Y.XuS.-G.GaoL.-C. (2022a). System Xc −/GSH/GPX4 axis: an important antioxidant system for the ferroptosis in drug-resistant solid tumor therapy. Front. Pharmacol.13, 910292. 10.3389/fphar.2022.910292
85
LiH.LiY.YuY.RenX.YangC.JinW.et al (2024a). GSH exhaustion via inhibition of xCT-GSH-GPX4 pathway synergistically enhanced DSF/Cu-induced cuproptosis in myelodysplastic syndromes. Free Radic. Biol. Med.222, 130–148. 10.1016/j.freeradbiomed.2024.06.006
86
LiJ.CaoF.YinH.HuangZ.LinZ.MaoN.et al (2020a). Ferroptosis: past, present and future. Cell. Death Dis.11, 88. 10.1038/s41419-020-2298-2
87
LiJ.LiF.TangF.ZhangJ.LiR.ShengD.et al (2020b). AGS-30, an andrographolide derivative, suppresses tumor angiogenesis and growth in vitro and in vivo. Biochem. Pharmacol.171, 113694. 10.1016/j.bcp.2019.113694
88
LiJ.LiuZ.WuX.LeeS. M.-Y.SetoS.-W.ZhangJ.et al (2024b). Anti-metastatic effects of AGS-30 on breast cancer through the inhibition of M2-like macrophage polarization. Biomed. and Pharmacother.172, 116269. 10.1016/j.biopha.2024.116269
89
LiP.LyuT. (2024). Research progress on ferroptosis in multiple myeloma. Curr. Treat. Options Oncol.25, 1276–1282. 10.1007/s11864-024-01250-z
90
LiS.ZengH.FanJ.WangF.XuC.LiY.et al (2023a). Glutamine metabolism in breast cancer and possible therapeutic targets. Biochem. Pharmacol.210, 115464. 10.1016/j.bcp.2023.115464
91
LiW.FuH.FangL.ChaiH.DingB.QianS. (2023b). Andrographolide induced ferroptosis in multiple myeloma cells by regulating the P38/Nrf2/HO-1 pathway. Archives Biochem. Biophysics742, 109622. 10.1016/j.abb.2023.109622
92
LiW.FuH.FangL.ChaiH.GaoT.ChenZ.et al (2022b). Shikonin induces ferroptosis in multiple myeloma via GOT1-mediated ferritinophagy. Front. Oncol.12, 1025067. 10.3389/fonc.2022.1025067
93
LiW.YinX.FuH.LiuJ.WengZ.MaoQ.et al (2024c). Ethanol extract of Eclipta prostrata induces multiple myeloma ferroptosis via Keap1/Nrf2/HO-1 axis. Phytomedicine128, 155401. 10.1016/j.phymed.2024.155401
94
LiY.ChenF.ChenJ.ChanS.HeY.LiuW.et al (2020c). Disulfiram/copper induces antitumor activity against both nasopharyngeal cancer cells and cancer-associated fibroblasts through ROS/MAPK and ferroptosis pathways. Cancers (Basel)12, 138. 10.3390/cancers12010138
95
LiY.RanQ.DuanQ.JinJ.WangY.YuL.et al (2024d). 7-Dehydrocholesterol dictates ferroptosis sensitivity. Nature626, 411–418. 10.1038/s41586-023-06983-9
96
LiZ.XuZ.ChenW.DuX.OuC.LuoZ.et al (2024e). Tumor-repopulating cells evade ferroptosis via PCK2-dependent phospholipid remodeling. Nat. Chem. Biol.20, 1341–1352. 10.1038/s41589-024-01612-6
97
LiangC.ZhangX.YangM.DongX. (2019). Recent progress in ferroptosis inducers for cancer therapy. Adv. Mater.31, 1904197. 10.1002/adma.201904197
98
LiangL.LiuY.WuX.ChenY. (2023). Artesunate induces ferroptosis by inhibiting the nuclear localization of SREBP2 in myeloma cells. Int. J. Med. Sci.20, 1535–1550. 10.7150/ijms.86409
99
LinL.LiR.CaiM.HuangJ.HuangW.GuoY.et al (2018). Andrographolide ameliorates liver fibrosis in mice: Involvement of TLR4/NF-κB and TGF-β1/Smad2 signaling pathways. Oxid. Med. Cell. Longev.2018, 7808656. 10.1155/2018/7808656
100
LiuD.CaoJ.DingX.XuW.YaoX.DaiM.et al (2024). Disulfiram/Copper complex improves the effectiveness of the WEE1 inhibitor adavosertib in p53 deficient non-small cell lung cancer via ferroptosis. Biochimica Biophysica Acta (BBA) - Mol. Basis Dis.1870, 167455. 10.1016/j.bbadis.2024.167455
101
LiuJ.KangR.TangD. (2022a). Signaling pathways and defense mechanisms of ferroptosis. FEBS J.289, 7038–7050. 10.1111/febs.16059
102
LiuL.ZhangY.WangL.LiuY.ChenH.HuQ.et al (2023). Scutellarein alleviates chronic obstructive pulmonary disease through inhibition of ferroptosis by chelating iron and interacting with arachidonate 15-lipoxygenase. Phytotherapy Res.37, 4587–4606. 10.1002/ptr.7928
103
LiuX.ZhangY.WuX.XuF.MaH.WuM.et al (2022b). Targeting ferroptosis pathway to combat therapy resistance and metastasis of cancer. Front. Pharmacol.13, 909821. 10.3389/fphar.2022.909821
104
LogieE.Van PuyveldeB.CuypersB.SchepersA.BerghmansH.VerdonckJ.et al (2021). Ferroptosis induction in multiple myeloma cells triggers DNA methylation and histone modification changes associated with cellular senescence. Int. J. Mol. Sci.22, 12234. 10.3390/ijms222212234
105
LuQ.LiaoY. (2025). GDF-15 upregulates the SLC7A11/GPX4 signaling axis and promotes mitoxantrone resistance in AML cells. Eur. J. Med. Res.30, 504. 10.1186/s40001-025-02787-x
106
LuQ.LuoS.ShiZ.YuM.GuoW.LiC. (2022). Nitidine chloride, a benzophenanthridine alkaloid from Zanthoxylum nitidum (roxb.) DC., exerts multiple beneficial properties, especially in tumors and inflammation-related diseases. Front. Pharmacol.13, 1046402. 10.3389/fphar.2022.1046402
107
LupescuA.BissingerR.JilaniK.LangF. (2014). In vitro induction of erythrocyte phosphatidylserine translocation by the natural naphthoquinone shikonin. Toxins (Basel)6, 1559–1574. 10.3390/toxins6051559
108
LvY.LiangC.SunQ.ZhuJ.XuH.LiX.et al (2023). Structural insights into FSP1 catalysis and ferroptosis inhibition. Nat. Commun.14, 5933. 10.1038/s41467-023-41626-7
109
MaT.DuJ.ZhangY.WangY.WangB.ZhangT. (2022). GPX4-independent Ferroptosis—A new strategy in disease’s therapy. Cell. Death Discov.8, 434–438. 10.1038/s41420-022-01212-0
110
MaesK.NemethE.RoodmanG. D.HustonA.EsteveF.FreytesC.et al (2010). In anemia of multiple myeloma, hepcidin is induced by increased bone morphogenetic protein 2. Blood116, 3635–3644. 10.1182/blood-2010-03-274571
111
Mahoney-SánchezL.BouchaouiH.AytonS.DevosD.DuceJ. A.DevedjianJ.-C. (2021). Ferroptosis and its potential role in the physiopathology of Parkinson’s disease. Prog. Neurobiol.196, 101890. 10.1016/j.pneurobio.2020.101890
112
MalardF.NeriP.BahlisN. J.TerposE.MoukalledN.HungriaV. T. M.et al (2024). Multiple myeloma. Nat. Rev. Dis. Prim.10, 45–21. 10.1038/s41572-024-00529-7
113
ManzD. H.BlanchetteN. L.PaulB. T.TortiF. M.TortiS. V. (2016). Iron and cancer: recent insights. Ann. N. Y. Acad. Sci.1368, 149–161. 10.1111/nyas.13008
114
MoralesM.XueX. (2021). Targeting iron metabolism in cancer therapy. Theranostics11, 8412–8429. 10.7150/thno.59092
115
MortensenM. S.RuizJ.WattsJ. L. (2023). Polyunsaturated fatty acids drive lipid peroxidation during ferroptosis. Cells12, 804. 10.3390/cells12050804
116
MynottR. L.HabibA.BestO. G.Wallington-GatesC. T. (2023). Ferroptosis in haematological malignancies and associated therapeutic nanotechnologies. Int. J. Mol. Sci.24, 7661. 10.3390/ijms24087661
117
NaponelliV.RocchettiM. T.MangieriD. (2024). Apigenin: molecular mechanisms and therapeutic potential against cancer spreading. Int. J. Mol. Sci.25, 5569. 10.3390/ijms25105569
118
NemethE.GanzT. (2023). Hepcidin and iron in health and disease. Annu. Rev. Med.74, 261–277. 10.1146/annurev-med-043021-032816
119
Nishida Xavier da SilvaT.Friedmann AngeliJ. P.IngoldI. (2022). GPX4: old lessons, new features. Biochem. Soc. Trans.50, 1205–1213. 10.1042/BST20220682
120
NookaA. K.KaufmanJ. L.LonialS. (2024). Glücksklee, the quad, and high-risk myeloma. JCO42, 1–3. 10.1200/JCO.23.02017
121
PanaroniC.FulzeleK.MoriT.SiuK. T.OnyewadumeC.MaebiusA.et al (2022). Multiple myeloma cells induce lipolysis in adipocytes and uptake fatty acids through fatty acid transporter proteins. Blood139, 876–888. 10.1182/blood.2021013832
122
PanaroniC.FulzeleK.SoucyR.SiuK. T.MukaiharaK.HuangC.et al (2018). Arachidonic acid induces ferroptosis-mediated cell-death in multiple myeloma. Blood132, 4498. 10.1182/blood-2018-99-118482
123
ParkJ.-M.LeeD.-H.KimD.-H. (2023). Redefining the role of AMPK in autophagy and the energy stress response. Nat. Commun.14, 2994. 10.1038/s41467-023-38401-z
124
PerisI.Romero-MurilloS.Martínez-BalsalobreE.FarringtonC. C.ArriazuE.MarcoteguiN.et al (2023). Activation of the PP2A-B56α heterocomplex synergizes with venetoclax therapies in AML through BCL2 and MCL1 modulation. Blood141, 1047–1059. 10.1182/blood.2022016466
125
PiérardG. E.Piérard-FranchimontC.PaquetP.QuatresoozP. (2009). Spotlight on adapalene. Expert Opin. Drug Metabolism and Toxicol.5, 1565–1575. 10.1517/17425250903386269
126
QianY.XiaL.WeiL.LiD.JiangW. (2021). Artesunate inhibits Staphylococcus aureus biofilm formation by reducing alpha-toxin synthesis. Arch. Microbiol.203, 707–717. 10.1007/s00203-020-02077-6
127
QinJ.SharmaA.WangY.Tobar-TosseF.DakalT. C.LiuH.et al (2022). Systematic discrimination of the repetitive genome in proximity of ferroptosis genes and a novel prognostic signature correlating with the oncogenic lncRNA CRNDE in multiple myeloma. Front. Oncol.12, 1026153. 10.3389/fonc.2022.1026153
128
QiuC.ZhangX.HuangB.WangS.ZhouW.LiC.et al (2020). Disulfiram, a ferroptosis inducer, triggers lysosomal membrane permeabilization by Up-Regulating ROS in glioblastoma. Onco Targets Ther.13, 10631–10640. 10.2147/OTT.S272312
129
RaafatK. M.El-ZahabyS. A. (2020). Niosomes of active Fumaria officinalis phytochemicals: antidiabetic, antineuropathic, anti-inflammatory, and possible mechanisms of action. Chin. Med.15, 40. 10.1186/s13020-020-00321-1
130
RajkumarS. V.DimopoulosM. A.PalumboA.BladeJ.MerliniG.MateosM.-V.et al (2014). International myeloma working group updated criteria for the diagnosis of multiple myeloma. Lancet Oncol.15, e538–e548. 10.1016/S1470-2045(14)70442-5
131
RajkumarS. V.KumarS. (2020). Multiple myeloma current treatment algorithms. Blood Cancer J.10, 94–10. 10.1038/s41408-020-00359-2
132
RenX.LiY.ZhouY.HuW.YangC.JingQ.et al (2021). Overcoming the compensatory elevation of NRF2 renders hepatocellular carcinoma cells more vulnerable to disulfiram/copper-induced ferroptosis. Redox Biol.46, 102122. 10.1016/j.redox.2021.102122
133
RouzerC. A.MarnettL. J. (2003). Mechanism of free radical oxygenation of polyunsaturated fatty acids by cyclooxygenases. Chem. Rev.103, 2239–2304. 10.1021/cr000068x
134
RuQ.LiY.ChenL.WuY.MinJ.WangF. (2024). Iron homeostasis and ferroptosis in human diseases: mechanisms and therapeutic prospects. Sig Transduct. Target Ther.9, 271–64. 10.1038/s41392-024-01969-z
135
RyanS. K.UgaldeC. L.RollandA.-S.SkidmoreJ.DevosD.HammondT. R. (2023). Therapeutic inhibition of ferroptosis in neurodegenerative disease. Trends Pharmacol. Sci.44, 674–688. 10.1016/j.tips.2023.07.007
136
SainiK. K.ChaturvediP.SinhaA.SinghM. P.KhanM. A.VermaA.et al (2023). Loss of PERK function promotes ferroptosis by downregulating SLC7A11 (system xc−) in colorectal cancer. Redox Biol.65, 102833. 10.1016/j.redox.2023.102833
137
SamovichS. N.Mikulska-RuminskaK.DarH. H.TyurinaY. Y.TyurinV. A.SouryavongA. B.et al (2024). Strikingly high activity of 15-Lipoxygenase towards Di-Polyunsaturated Arachidonoyl/adrenoyl-phosphatidylethanolamines generates peroxidation signals of ferroptotic cell death. Angew. Chem. Int. Ed.63, e202314710. 10.1002/anie.202314710
138
SasakuraM.UrakamiH.TachibanaK.IkedaK.HasuiK.MatsudaY.et al (2024). Topical application of activator protein-1 inhibitor T-5224 suppresses inflammation and improves skin barrier function in a murine atopic dermatitis-like dermatitis. Allergol. Int.73, 323–331. 10.1016/j.alit.2023.12.006
139
ShahS. A. H.RanaS. L.MohanyM.MiloševićM.Al-RejaieS. S.FarooqM. A.et al (2024). Fumaria indica (hausskn.) pugsley hydromethanolic extract: bioactive compounds identification, hypotensive mechanism, and cardioprotective potential exploration. ACS Omega9, 3642–3668. 10.1021/acsomega.3c07655
140
ShangY.LuoM.YaoF.WangS.YuanZ.YangY. (2020). Ceruloplasmin suppresses ferroptosis by regulating iron homeostasis in hepatocellular carcinoma cells. Cell. Signal.72, 109633. 10.1016/j.cellsig.2020.109633
141
SharmaN.DeyS.SinghS.KumarS.KonarM.NaithaniP.et al (2024). Disulfiram-copper potentiates anticancer efficacy of standard chemo-therapy drugs in bladder cancer animal model through ROS-Autophagy-Ferroptosis signalling Cascade. CCDT24, 1145–1157. 10.2174/0115680096325879240815105227
142
ShiJ.ChengY.WangL.XingW.LiY.SunX.et al (2025). SR-B1 deficiency suppresses progression in acute myeloid leukemia via ferroptosis and reverses resistance to venetoclax. Free Radic. Biol. Med.233, 24–38. 10.1016/j.freeradbiomed.2025.03.031
143
ShiJ.JiX.ShanS.ZhaoM.BiC.LiZ. (2023). The interaction between apigenin and PKM2 restrains progression of colorectal cancer. J. Nutr. Biochem.121, 109430. 10.1016/j.jnutbio.2023.109430
144
SongA.WuL.ZhangB.-X.YangQ.-C.LiuY.-T.LiH.et al (2024a). Glutamine inhibition combined with CD47 blockade enhances radiotherapy-induced ferroptosis in head and neck squamous cell carcinoma. Cancer Lett.588, 216727. 10.1016/j.canlet.2024.216727
145
SongG.LiM.FanS.QinM.ShaoB.DaiW.et al (2024b). Boosting synergism of chemo- and immuno-therapies via switching paclitaxel-induced apoptosis to mevalonate metabolism-triggered ferroptosis by bisphosphonate coordination lipid nanogranules. Acta Pharm. Sin. B14, 836–853. 10.1016/j.apsb.2023.08.029
146
SongQ.ZhangY.HuH.YangX.XingX.WuJ.et al (2024c). Augment of ferroptosis with photothermal enhanced fenton reaction and glutathione inhibition for tumor synergistic nano-catalytic therapy. Int. J. Nanomedicine19, 11923–11940. 10.2147/IJN.S480586
147
StockwellB. R.Friedmann AngeliJ. P.BayirH.BushA. I.ConradM.DixonS. J.et al (2017). Ferroptosis: a regulated cell death nexus linking metabolism, redox biology, and disease. Cell.171, 273–285. 10.1016/j.cell.2017.09.021
148
StrikH.EfferthT.KainaB. (2024). Artesunate in glioblastoma therapy: case reports and review of clinical studies. Phytomedicine123, 155274. 10.1016/j.phymed.2023.155274
149
SuQ.LiQ.ZhangW.LiB.ZhuangW. (2022). Integrative analysis of enrichment and prognostic value of ferroptosis-related genes and pathways in multiple myeloma. Carcinogenesis43, 1050–1058. 10.1093/carcin/bgac080
150
SudhakaranM.NavarreteT. G.Mejía-GuerraK.MukundiE.EubankT. D.GrotewoldE.et al (2023). Transcriptome reprogramming through alternative splicing triggered by apigenin drives cell death in triple-negative breast cancer. Cell. Death Dis.14, 824. 10.1038/s41419-023-06342-6
151
SuiX.WangJ.ZhaoZ.LiuB.LiuM.LiuM.et al (2024). Phenolic compounds induce ferroptosis-like death by promoting hydroxyl radical generation in the fenton reaction. Commun. Biol.7, 199. 10.1038/s42003-024-05903-5
152
SunY.LiQ.GuoH.HeQ. (2022). Ferroptosis and iron metabolism after intracerebral hemorrhage. Cells12, 90. 10.3390/cells12010090
153
SuzukiS.VenkateshD.KandaH.NakayamaA.HosokawaH.LeeE.et al (2022). GLS2 is a tumor suppressor and a regulator of ferroptosis in hepatocellular carcinoma. Cancer Res.82, 3209–3222. 10.1158/0008-5472.CAN-21-3914
154
TangD.ChenK.HuangL.LiJ. (2017). Pharmacokinetic properties and drug interactions of apigenin, a natural flavone. Expert Opin. Drug Metabolism and Toxicol.13, 323–330. 10.1080/17425255.2017.1251903
155
TangS.LiuJ.LiF.YanY.LongX.FuY. (2024a). AP-1 inhibitor induces ferroptosis via the PI3K/AKT pathway in multiple myeloma cells. Heliyon10, e34397. 10.1016/j.heliyon.2024.e34397
156
TangS.ZhangF.LiJ.DongH.YangQ.LiuJ.et al (2023). The selective activator protein-1 inhibitor T-5224 regulates the IRF4/MYC axis and exerts cooperative antimyeloma activity with bortezomib. Chemico-Biological Interact.384, 110687. 10.1016/j.cbi.2023.110687
157
TangW.-J.XuD.LiangM.-X.WoG.-Q.ChenW.-Q.TangJ.-H.et al (2024b). Pitavastatin induces autophagy-dependent ferroptosis in MDA-MB-231 cells via the mevalonate pathway. Heliyon10, e27084. 10.1016/j.heliyon.2024.e27084
158
TianS.LiY.-L.WangJ.DongR.-C.WeiJ.MaY.et al (2023). Chinese ecliptae herba (Eclipta prostrata (L.) L.) extract and its component wedelolactone enhances osteoblastogenesis of bone marrow mesenchymal stem cells via targeting METTL3-mediated m6A RNA methylation. J. Ethnopharmacol.312, 116433. 10.1016/j.jep.2023.116433
159
TimalsinaD.DevkotaH. P. (2021). Eclipta prostrata (L.) L. (asteraceae): ethnomedicinal uses, chemical constituents, and biological activities. Biomolecules11, 1738. 10.3390/biom11111738
160
TortiS. V.TortiF. M. (2013). Iron and cancer: more ore to be mined. Nat. Rev. Cancer13, 342–355. 10.1038/nrc3495
161
TuncerC.HaciogluC. (2024). Borax induces ferroptosis of glioblastoma by targeting HSPA5/NRF2/GPx4/GSH pathways. J. Cell. Mol. Med.28, e18206. 10.1111/jcmm.18206
162
VakhrushevaO.ErbH. H. H.BräunigV.MarkowitschS. D.SchuppP.BaerP. C.et al (2022). Artesunate inhibits the growth behavior of docetaxel-resistant prostate cancer cells. Front. Oncol.12, 789284. 10.3389/fonc.2022.789284
163
van de DonkN. W. C. J.PawlynC.YongK. L. (2021). Multiple myeloma. Lancet397, 410–427. 10.1016/S0140-6736(21)00135-5
164
VanHookA. M. (2023). Proinflammatory apoptosis. Sci. Signal.16, eadm8181. 10.1126/scisignal.adm8181
165
WangC.ZhaY.-L.WangH.SunB.QiangW.-G.YuanY.et al (2025). Carfilzomib promotes Iodine-125 seed radiation-induced apoptosis, paraptosis, and ferroptosis in esophageal squamous cell carcinoma by aggravating endoplasmic reticulum stress. Transl. Oncol.57, 102393. 10.1016/j.tranon.2025.102393
166
WangH.ZhangZ.RuanS.YanQ.ChenY.CuiJ.et al (2023a). Regulation of iron metabolism and ferroptosis in cancer stem cells. Front. Oncol.13, 1251561. 10.3389/fonc.2023.1251561
167
WangL.LiuY.DuT.YangH.LeiL.GuoM.et al (2020). ATF3 promotes erastin-induced ferroptosis by suppressing system Xc. Cell. Death Differ.27, 662–675. 10.1038/s41418-019-0380-z
168
WangQ.ZhaoM.ZhangT.ZhangB.ZhengZ.LinZ.et al (2023b). Comprehensive analysis of ferroptosis-related genes in immune infiltration and prognosis in multiple myeloma. Front. Pharmacol.14, 1203125. 10.3389/fphar.2023.1203125
169
WangY.AnX.WangF.JiangY. (2024). Ginsenoside RH4 inhibits ang II-induced myocardial remodeling by interfering with NFIL3. Biomed. and Pharmacother.172, 116253. 10.1016/j.biopha.2024.116253
170
WangY.LvM.ZhaoW. (2023c). Research on ferroptosis as a therapeutic target for the treatment of neurodegenerative diseases. Ageing Res. Rev.91, 102035. 10.1016/j.arr.2023.102035
171
WaughJ.NobleS.ScottL. J. (2004). Adapalene: a review of its use in the treatment of acne vulgaris. Drugs64, 1465–1478. 10.2165/00003495-200464130-00005
172
Weiser DrozdkovaD.Smesny TrtkovaK. (2021). Possible therapeutic potential of disulfiram for multiple myeloma. Curr. Oncol.28, 2087–2096. 10.3390/curroncol28030193
173
WienchB.EichhornT.PaulsenM.EfferthT. (2012). Shikonin directly targets mitochondria and causes mitochondrial dysfunction in cancer cells. Evid. Based Complement. Altern. Med.2012, 726025. 10.1155/2012/726025
174
WuH.LuanY.WangH.ZhangP.LiuS.WangP.et al (2022). Selenium inhibits ferroptosis and ameliorates autistic-like behaviors of BTBR mice by regulating the Nrf2/GPx4 pathway. Brain Res. Bull.183, 38–48. 10.1016/j.brainresbull.2022.02.018
175
WuJ.HanW.ZhangY.LiS.QinT.HuangZ.et al (2024a). Glutamine mitigates oxidative stress-induced matrix degradation, ferroptosis, and pyroptosis in nucleus pulposus cells via deubiquitinating and stabilizing Nrf2. Antioxidants and Redox Signal.41, 278–295. 10.1089/ars.2023.0384
176
WuQ.DengJ.FanD.DuanZ.ZhuC.FuR.et al (2018). Ginsenoside Rh4 induces apoptosis and autophagic cell death through activation of the ROS/JNK/p53 pathway in colorectal cancer cells. Biochem. Pharmacol.148, 64–74. 10.1016/j.bcp.2017.12.004
177
WuQ.GeX.GengZ.WuH.YangJ.CaoS.et al (2024b). HuaChanSu suppresses the growth of hepatocellular carcinoma cells by interfering with pentose phosphate pathway through down-regulation of G6PD enzyme activity and expression. Heliyon10, e25144. 10.1016/j.heliyon.2024.e25144
178
WuX.LiY.ZhangS.ZhouX. (2021). Ferroptosis as a novel therapeutic target for cardiovascular disease. Theranostics11, 3052–3059. 10.7150/thno.54113
179
XianM.WangQ.XiaoL.ZhongL.XiongW.YeL.et al (2024). Leukocyte immunoglobulin-like receptor B1 (LILRB1) protects human multiple myeloma cells from ferroptosis by maintaining cholesterol homeostasis. Nat. Commun.15, 5767. 10.1038/s41467-024-50073-x
180
XieY.KangR.KlionskyD. J.TangD. (2023). GPX4 in cell death, autophagy, and disease. Autophagy19, 2621–2638. 10.1080/15548627.2023.2218764
181
XingK.BianX.ShiD.DongS.ZhouH.XiaoS.et al (2023). miR-612 enhances RSL3-Induced ferroptosis of hepatocellular carcinoma cells via mevalonate pathway. J. Hepatocell. Carcinoma10, 2173–2185. 10.2147/JHC.S433332
182
XingL.LiuX.-Y.ZhouT.-J.WanX.WangY.JiangH.-L. (2021). Photothermal nanozyme-ignited fenton reaction-independent ferroptosis for breast cancer therapy. J. Control. Release339, 14–26. 10.1016/j.jconrel.2021.09.019
183
XuZ.LiuX.ZhuangD. (2022). Artesunate inhibits proliferation, migration, and invasion of thyroid cancer cells by regulating the PI3K/AKT/FKHR pathway. Biochem. Cell. Biol.100, 85–92. 10.1139/bcb-2021-0275
184
XueP.ZhuangH.BaiT.ZengX.DengJ.ShaoS.et al (2024). Iron (II)-Based metal-organic framework nanozyme for boosting tumor ferroptosis through inhibiting DNA damage repair and system Xc. J. Nanobiotechnology22, 228. 10.1186/s12951-024-02508-2
185
YadavS.SharmaA.NayikG. A.CooperR.BhardwajG.SohalH. S.et al (2022). Review of shikonin and derivatives: isolation, chemistry, biosynthesis, pharmacology and toxicology. Front. Pharmacol.13, 905755. 10.3389/fphar.2022.905755
186
YagodaN.von RechenbergM.ZaganjorE.BauerA. J.YangW. S.FridmanD. J.et al (2007). RAS–RAF–MEK-dependent oxidative cell death involving voltage-dependent anion channels. Nature447, 864–868. 10.1038/nature05859
187
YanB.AiY.SunQ.MaY.CaoY.WangJ.et al (2021a). Membrane damage during ferroptosis is caused by oxidation of phospholipids catalyzed by the oxidoreductases POR and CYB5R1. Mol. Cell.81, 355–369.e10. 10.1016/j.molcel.2020.11.024
188
YanC.LiQ.SunQ.YangL.LiuX.ZhaoY.et al (2023). Promising nanomedicines of shikonin for cancer therapy. Int. J. Nanomedicine18, 1195–1218. 10.2147/IJN.S401570
189
YanH.ZouT.TuoQ.XuS.LiH.BelaidiA. A.et al (2021b). Ferroptosis: mechanisms and links with diseases. Sig Transduct. Target Ther.6, 49–16. 10.1038/s41392-020-00428-9
190
YangF.XiaoY.DingJ.-H.JinX.MaD.LiD.-Q.et al (2023a). Ferroptosis heterogeneity in triple-negative breast cancer reveals an innovative immunotherapy combination strategy. Cell. Metab.35, 84–100.e8. 10.1016/j.cmet.2022.09.021
191
YangJ.KimJ. S.KwonY. S.SeongE. S.KimM. J. (2023b). Antioxidant and antiproliferative activities of Eclipta prostrata (L.) L. extract and isolated compounds. Molecules28, 7354. 10.3390/molecules28217354
192
YangJ.WangF.HuZ.LiuX.ZhangW.LiC.et al (2025). Huachansu injection induces ferroptosis in multiple myeloma through NRF2/HO-1 signaling pathway. J. Ethnopharmacol.343, 119454. 10.1016/j.jep.2025.119454
193
YangS.xieZ.PeiT.zengY.XiongQ.WeiH.et al (2022). Salidroside attenuates neuronal ferroptosis by activating the Nrf2/HO1 signaling pathway in Aβ1-42-induced Alzheimer’s disease mice and glutamate-injured HT22 cells. Chin. Med.17, 82. 10.1186/s13020-022-00634-3
194
YangW. S.KimK. J.GaschlerM. M.PatelM.ShchepinovM. S.StockwellB. R. (2016). Peroxidation of polyunsaturated fatty acids by lipoxygenases drives ferroptosis. Proc. Natl. Acad. Sci. U. S. A.113, E4966–E4975. 10.1073/pnas.1603244113
195
YangW. S.SriRamaratnamR.WelschM. E.ShimadaK.SkoutaR.ViswanathanV. S.et al (2014). Regulation of ferroptotic cancer cell death by GPX4. Cell.156, 317–331. 10.1016/j.cell.2013.12.010
196
YangW. S.StockwellB. R. (2008). Synthetic lethal screening identifies compounds activating iron-dependent, nonapoptotic cell death in Oncogenic-RAS-Harboring cancer cells. Chem. and Biol.15, 234–245. 10.1016/j.chembiol.2008.02.010
197
YangY.GaoH.LiuW.LiuX.JiangX.LiX.et al (2021). Arctium lappa L. roots ameliorates cerebral ischemia through inhibiting neuronal apoptosis and suppressing AMPK/mTOR-mediated autophagy. Phytomedicine85, 153526. 10.1016/j.phymed.2021.153526
198
YaoX.LiW.FangD.XiaoC.WuX.LiM.et al (2021). Emerging roles of energy metabolism in ferroptosis regulation of tumor cells. Adv. Sci.8, 2100997. 10.1002/advs.202100997
199
YinZ.LvY.DengL.LiG.OuR.ChenL.et al (2023). Targeting ABCB6 with nitidine chloride inhibits PI3K/AKT signaling pathway to promote ferroptosis in multiple myeloma. Free Radic. Biol. Med.203, 86–101. 10.1016/j.freeradbiomed.2023.04.003
200
YingQ.LouJ.ZhengD. (2023). Ginsenoside Rh4 inhibits the malignant progression of multiple myeloma and induces ferroptosis by regulating SIRT2. Clin. Exp. Pharmacol. Physiology50, 757–765. 10.1111/1440-1681.13805
201
YosriN.AlsharifS. M.XiaoJ.MusharrafS. G.ZhaoC.SaeedA.et al (2023). Arctium lappa (burdock): insights from ethnopharmacology potential, chemical constituents, clinical studies, pharmacological utility and nanomedicine. Biomed. and Pharmacother.158, 114104. 10.1016/j.biopha.2022.114104
202
YuF.TanW.ChenZ.ShenX.MoX.MoX.et al (2022). Nitidine chloride induces caspase 3/GSDME-dependent pyroptosis by inhibting PI3K/Akt pathway in lung cancer. Chin. Med.17, 115. 10.1186/s13020-022-00671-y
203
ZengC.NieD.WangX.ZhongS.ZengX.LiuX.et al (2024). Combined targeting of GPX4 and BCR-ABL tyrosine kinase selectively compromises BCR-ABL+ leukemia stem cells. Mol. Cancer23, 240. 10.1186/s12943-024-02162-0
204
ZhangD.Duque-JimenezJ.FacchinettiF.BrixiG.RheeK.FengW. W.et al (2024a). Transferrin receptor targeting chimeras for membrane protein degradation. Nature638, 787–795. 10.1038/s41586-024-07947-3
205
ZhangG.LvS.ZhongX.LiX.YiY.LuY.et al (2024b). Ferroptosis: a new antidepressant pharmacological mechanism. Front. Pharmacol.14, 1339057. 10.3389/fphar.2023.1339057
206
ZhangJ.LiuY.LiQ.ZuoL.ZhangB.ZhaoF.et al (2023). ACSL4: a double-edged sword target in multiple myeloma, promotes cell proliferation and sensitizes cell to ferroptosis. Carcinogenesis44, 242–251. 10.1093/carcin/bgad015
207
ZhangJ.LiuY.ZuoL.FanF.YanH.ZhaoF.et al (2024c). Class II ferroptosis inducers are a novel therapeutic approach for t(4;14)-positive multiple myeloma. Blood Adv.8, 5022–5038. 10.1182/bloodadvances.2023010335
208
ZhangT.-C.LinY.-C.SunN.-N.LiuS.HuW.-Z.ZhaoY.et al (2024d). Icariin, astragaloside a and puerarin mixture attenuates cognitive impairment in APP/PS1 mice via inhibition of ferroptosis-lipid peroxidation. Neurochem. Int.175, 105705. 10.1016/j.neuint.2024.105705
209
ZhangW.LiuY.LiaoY.ZhuC.ZouZ. (2024e). GPX4, ferroptosis, and diseases. Biomed. and Pharmacother.174, 116512. 10.1016/j.biopha.2024.116512
210
ZhangW.-X.ZhangZ.-M.ZhangZ.-Q.WangY.ZhouW. (2014). Andrographolide induced acute kidney injury: analysis of 26 cases reported in Chinese literature. Nephrology19, 21–26. 10.1111/nep.12172
211
ZhangY.HeF.HuW.SunJ.ZhaoH.ChengY.et al (2024f). Bortezomib elevates intracellular free Fe2+ by enhancing NCOA4-mediated ferritinophagy and synergizes with RSL-3 to inhibit multiple myeloma cells. Ann. Hematol.103, 3627–3637. 10.1007/s00277-024-05762-4
212
ZhangY.WangX.LiX.XiongX.XueR.ZangL.et al (2024g). Novel methyltransferase G9a inhibitor induces ferroptosis in multiple myeloma through Nrf2/HO-1 pathway. Ann. Hematol.103, 2405–2417. 10.1007/s00277-024-05728-6
213
ZhangY.XinL.XiangM.ShangC.WangY.WangY.et al (2022). The molecular mechanisms of ferroptosis and its role in cardiovascular disease. Biomed. and Pharmacother.145, 112423. 10.1016/j.biopha.2021.112423
214
ZhaoC.LiuZ.ChangC.-C.ChenY.-C.ZhangQ.ZhangX.-D.et al (2023). Near-infrared phototheranostic iron pyrite nanocrystals simultaneously induce dual cell death pathways via enhanced fenton reactions in triple-negative breast cancer. ACS Nano17, 4261–4278. 10.1021/acsnano.2c06629
215
ZhaoY.WangQ.ZhuJ.CaiJ.FengX.SongQ.et al (2024). Identification of KW-2449 as a dual inhibitor of ferroptosis and necroptosis reveals that autophagy is a targetable pathway for necroptosis inhibitors to prevent ferroptosis. Cell. Death Dis.15, 764. 10.1038/s41419-024-07157-9
216
ZhongY.TianF.MaH.WangH.YangW.LiuZ.et al (2020). FTY720 induces ferroptosis and autophagy via PP2A/AMPK pathway in multiple myeloma cells. Life Sci.260, 118077. 10.1016/j.lfs.2020.118077
217
ZhouZ.HeW.TianH.ZhanP.LiuJ. (2023). Thyme (Thymus vulgaris L.) polyphenols ameliorate DSS-Induced ulcerative colitis of mice by mitigating intestinal barrier damage, regulating gut microbiota, and suppressing TLR4/NF-κB-NLRP3 inflammasome pathways. Food Funct.14, 1113–1132. 10.1039/D2FO02523J
218
ZhuL.HuS.YanX.ZengQ.ZhangB.JiangL.et al (2023). Ugi reaction-assisted assembly of covalent PROTACs against glutathione peroxidase 4. Bioorg. Chem.134, 106461. 10.1016/j.bioorg.2023.106461
219
ZhuX.ChenQ.XieL.ChenW.JiangY.SongE.et al (2022). Iron ion and sulfasalazine-loaded polydopamine nanoparticles for fenton reaction and glutathione peroxidase 4 inactivation for enhanced cancer ferrotherapy. Acta Biomater.145, 210–221. 10.1016/j.actbio.2022.04.024
Summary
Keywords
multiple myeloma, ferroptosis, pharmacological mechanism, natural products, antitumor effects
Citation
Shi J, Lu Y, Wei W, Ma G, Li C, Li L, Wang Y, Wang Y, Xu R and Cui S (2025) Ferroptosis: a novel pharmacological mechanism against multiple myeloma. Front. Pharmacol. 16:1606804. doi: 10.3389/fphar.2025.1606804
Received
06 April 2025
Accepted
27 June 2025
Published
15 July 2025
Volume
16 - 2025
Edited by
Venkatesh Pooladanda, Harvard Medical School, United States
Reviewed by
Lopmudra Sarode, Nagpur Veterinary College, India
Sreelakshmi Badavenkatappa Gari, Jawaharlal Nehru Technological University, India
Updates
Copyright
© 2025 Shi, Lu, Wei, Ma, Li, Li, Wang, Wang, Xu and Cui.
This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Siyuan Cui, csytcm@126.com
Disclaimer
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.