 Malwina Brożyna1*†
Malwina Brożyna1*† Zuzanna Stępnicka1,2*†
Zuzanna Stępnicka1,2*† Natalia Tymińska1Bartłomiej Dudek1
Natalia Tymińska1Bartłomiej Dudek1 Katarzyna Kapczyńska3
Katarzyna Kapczyńska3 Adam Matkowski2
Adam Matkowski2 Yanfang Sun4,5
Yanfang Sun4,5 Michał Tomczyk6
Michał Tomczyk6 Adam Junka1
Adam Junka1- 1“PUMA”, Platform for Unique Model Applications, Faculty of Pharmacy, Wroclaw Medical University, Wrocław, Poland
- 2Botanical Garden of Medicinal Plants, Department of Pharmaceutical Biology and Biotechnology, Wroclaw Medical University, Wrocław, Poland
- 3Hirszfeld Institute of Immunology and Experimental Therapy, Polish Academy of Sciences, Wrocław, Poland
- 4Zhejiang International Joint Laboratory of Traditional Medicine and Big Health Products Development, Hangzhou, China
- 5College of Life Sciences and Medicine, Zhejiang Sci-Tech University, Hangzhou, China
- 6Department of Biology and Pharmacognosy, Faculty of Pharmacy with the Division of Laboratory Medicine, Medical University of Białystok, Białystok, Poland
The rising interest in essential oils (EOs) as antimicrobial agents demands evaluation frameworks that provide structured, reproducible assessments. In this study, we examined the strain-dependent response of Pseudomonas aeruginosa to pharmacopoeia-grade Thyme Essential Oil (obtained from Thymus vulgaris L., TEO) or polyhexamethylene biguanide antiseptic (PHMB) using a panel of ten genetically diversified strains in planktonic and biofilm forms, and by complementary in vitro models. Chemical composition of TEO was assessed using Gas Chromatography-Mass Spectrometry (GC–MS), and the main components were thymol, p-cymene, and γ-terpinene. Despite uniform test conditions, we observed striking inter-strain variability: TEO Minimal Inhibitory Concentrations (MICs) differed by up to 1,000-fold, and biofilm susceptibility profiles ranged from full tolerance to near-complete eradication. Notably, strains with low metabolic activity and sparse cell populations—but high matrix biomass—exhibited reduced responsiveness to TEO, while susceptibility to PHMB was more consistent, though not absolute. These findings highlight the critical influence of both microbial phenotype and agent formulation on antimicrobial outcomes. Rather than framing EOs as superior or inferior alternatives, our results advocate for their integration into a stewardship paradigm—one that values standardization, model-based evaluation, and informed formulation. In this context, we position essential oil stewardship not as a constraint but as a necessary evolution for their credible inclusion in antimicrobial strategies.
1 Introduction
Pseudomonas aeruginosa is a genomically complex and phenotypically diverse Gram-negative pathogen, notorious for its intrinsic resistance, environmental resilience, and clinical adaptability. Its remarkable success across ecological niches—from soil and water to medical devices and chronically infected tissues—is rooted in a large and dynamic genome composed of a conserved core and an expansive accessory repertoire (Klockgether et al., 2011; Grace et al., 2022). Horizontal gene transfer, genomic rearrangements, and microevolutionary processes contribute to a non-clonal population structure, in which even closely related isolates may differ markedly in virulence, metabolic capacity, biofilm behavior, and antimicrobial susceptibility (Romling et al., 1997; Chandler et al., 2019; Grace et al., 2022). These features pose formidable challenges to infection control. P. aeruginosa is listed among the World Health Organization’s highest-priority pathogens due to its pervasive multidrug resistance and its ability to thrive in biofilms—structured, matrix-embedded communities that protect the bacteria from antimicrobials and host immune responses (Fernández-Billón et al., 2023; World Health Organization, 2024). While most experimental studies focus on reference strains such as PAO1 or PA14, clinical outcomes are ultimately shaped by the specific traits of the infecting isolate (Grace et al., 2022). Yet, strain-level variability is often overlooked in antimicrobial research, where either a single strain is profiled using an advanced technique, or a broader panel of strains is tested with lesser methodological depth (Sanchez and Martinez, 2019).
This gap arises from both practical and conceptual constraints. In-depth phenotypic analyses—including biofilm imaging, viability assays, and molecular profiling—are time- and resource-intensive, making comprehensive evaluation of multiple strains logistically difficult (Azeredo et al., 2017). As a result, studies frequently default to using a well-characterized reference strain for comparability and genetic tractability. Conversely, larger isolate panels are commonly assessed using only a limited spectrum of readouts, such as Minimal Inhibitory Concentrations or biomass measurements, which fail to capture the functional heterogeneity within species. Compounding this is the prevailing assumption that findings from a model strain can be generalized across the species—an assumption increasingly at odds with accumulating evidence. This methodological bottleneck has impeded progress in understanding intra-species variability, particularly in response to complex, multicomponent agents such as essential oils (Nysten et al., 2024). Much like P. aeruginosa itself—with its multifactorial resistance mechanisms and strain-specific phenotypes—EOs are not uniform chemical entities, but complex, multicomponent mixtures (Dhifi et al., 2016). Even when pharmacopeial standards define their minimum content of major constituents, such as thymol in Thyme EO, these oils typically contain between several and over a dozen bioactive compounds that interact through synergistic, additive, or even antagonistic mechanisms (Stahl-Biskup and Venskutonis, 2012; Santana de Oliveira et al., 2023).
Thymus vulgaris L. (Lamiaceae) – common thyme–the source material for the pharmacopeial material is not only a medicinal, aromatic plant, but is also a commonly used culinary spice. Cultivated worldwide, this plant is genetically diverse, with numerous races, genotypes, and cultivars that, along with environmental and agronomic factors, may significantly modulate the essential oil profile, including minor compound (Hudaib et al., 2002). These may also add some degree of uncertainty in the desired activity despite conforming to quality standards (Basch et al., 2004). Moreover, the European Pharmacopoeia and the EMA HMPC (European Medicines Agency, Herbal Medicinal Products Committee) monographs also accept T. zygis L. or a mixture of these as valid herbal material (European Medicines Agency Committee on Herbal Medicinal Products, 2019). T. zygis (red thyme) is less widespread in cultivation but also versatile in use and variable in composition (Coimbra et al., 2022). Thymi aetheroleum (Thyme Essential Oil), according to the European Pharmacopoeia, shall contain between 37% and 55% thymol. The typically found minor (non-thymol) compounds of TEO include: carvacrol 0.5%–5.5%, carvacrol methyl ether 0.05%–1.5%, p-cymene 14.0%–28.0%, linalool 1.5%–6.5%, β-myrcene 1.0%–3.0%, α-terpinene 0.9%–2.6%, γ-terpinene 4.0%–12.0%, terpinen-4-ol 0.1%–2.5%, α-thujene 0.2%–1.5%, with an admixture of a sesquiterpene β-caryophyllene (European Medicines Agency Committee on Herbal Medicinal Products, 2019).
Moreover, EOs antimicrobial activity is not limited to the liquid phase: volatile fractions can exert effects on planktonic and biofilm-forming bacteria at a distance, further complicating mechanistic interpretation and reproducibility (Mohammed et al., 2024). The dual-phase nature of EOs activity—airborne and contact-dependent—challenges classical testing paradigms developed for water-soluble antibiotics and necessitates more nuanced, multi-layered assessment strategies (Reyes-Jurado et al., 2015).
Further complicating the landscape is the methodological heterogeneity inherent to in vitro biofilm research (Balaban et al., 2019). Unlike planktonic assays, where standardized protocols exist for determining MICs, or zones of growth inhibition, biofilm studies are subject to considerable variability in experimental design (The European Committee on Antimicrobial Susceptibility Testing, 2020; The European Committee on Antimicrobial Susceptibility Testing, 2025a). A culture medium is one of a factor influencing biofilm structure, it should reflect the chemical and nutritional composition of the infection site, as nutrient-rich laboratory broths (e.g., Tryptic Soy Broth) poorly mimic the physiological conditions found in the infection site (Ruhluel et al., 2022; Su et al., 2022; Brożyna et al., 2024). Moreover, the choice of surface is not merely a technical detail, but a major determinant of experimental outcome, particularly when evaluating agents intended for clinical application in wound, mucosal, or implant-associated infections (Dhekane et al., 2022). Porous and biologically relevant matrices are considered to better mimic the extracellular environment encountered during infection (Birkenhauer et al., 2014; Roberts et al., 2023). Biofilms formed on tissue-like substrates often exhibit greater density, deeper stratification, and altered metabolic gradients, which can significantly affect susceptibility profiles.
Additionally, antimicrobial agents may be introduced at different stages—either during biofilm development (preventive models) or after its establishment (therapeutic models) and their activity assessed via a wide range of readouts (Wilson et al., 2017). Therefore, distinct dimensions of biofilm integrity and susceptibility may be captured, making cross-study comparisons challenging. Thus, the complexity of the biological target and the therapeutic agent is further compounded by the methodological plasticity of the models themselves—underscoring the need for integrative, multi-parametric approaches when evaluating biofilm-active compounds (Azeredo et al., 2017).
Taken together, the evaluation of antimicrobial EOs’ strategies against P. aeruginosa biofilms in vitro is confounded by at least three major overlapping layers of complexity: (i) the extensive intra-species variability in biofilm formation and resistance phenotypes; (ii) the multi-component and phase-diverse nature of EOs and (iii) the methodological heterogeneity of in vitro biofilm models (Romling et al., 1997; Birkenhauer et al., 2014; Su et al., 2022; Mohammed et al., 2024). These challenges have led to fragmented literature and hindered the translational potential of plant-based antimicrobials.
Therefore, in this study, we address these limitations by integrating a genetically diverse panel of Pseudomonas strains, chemically specified TEO, and a suite of complementary biofilm assessment techniques, all embedded within a rigorous statistical framework. Collectively, this constitutes a structured and evidence-driven approach, analogous to the stewardship concepts established for antibiotics and antiseptics. By this analogy, we define our methodology as essential oil stewardship.
2 Materials and methods
2.1 Microorganisms
Ten strains of Pseudomonas aeruginosa were selected for the study purposes. Eight strains included clinical isolates from chronic wounds of various etiology (later referred to as P4, P20, P30E, P34, P44E, P68E, P92E, P856) and two reference ones (marked ATCC 9027, ATCC 15442, American Type Culture Collection). The strains were analyzed with MALDI-TOF (Matrix-Assisted Laser Desorption/Ionization Time-of-Flight Mass Spectrometry) ultrafleXtreme spectrometer (Bruker, Billerica, MA, United States) and identified with Biotyper platform. The tested strains were part of the Strain and Line Collection of the Platform for Unique Models Application, Department of the Pharmaceutical Microbiology and Parasitology, Medical University of Wroclaw. The bioethical approval was granted with the following number: Bioethical Committee of Wroclaw Medical University, protocol # 8/2016.
2.2 Tested compounds
The antimicrobial and antibiofilm activity of a commercial Thyme Essential Oil, thymol chemotype (obtained from the leaves of Thymus vulgaris L., Institute of Aromatherapy, Warsaw, Poland, batch number 33667/1/DW/46), was evaluated. Moreover, the antiseptic Prontosan Wound Irrigation Solution (polyhexanide, B. Braun Melsungen AG, Melsungen, Germany), composed of 0.1% polyhexamethylene biguanide, 0.1% undecylenamidopropyl betaine, and purified water, was used as a comparator substance.
2.3 Assessment of TEO chemical composition using gas chromatography- mass spectrometry (GC–MS)
The analysis was conducted to assess the percentage content of the TEO constituents. The protocol was adapted from the European Pharmacopoeia 11 standards. Firstly, the oil was diluted 50 times using hexane, mixed, and submitted for analysis with an Agilent 7890B GC system coupled with 7000GC/TQ, equipped with a PAL RSI85 autosampler (Agilent Technologies, Santa Clara, CA, United States) and an HP-5 MS column (30 m × 0.25 mm × 0.25 μm). Helium was used as a carrier gas with a total flow of 1 mL/min. The ratio of injection splitting was 1:100. The initial temperature of the analysis was 50 °C maintained for 1 min. Next, the temperature was set to reach 170 °C (4 °C/min) and then to 280 °C (10 °C/min), which was kept for 2 min. The MS detector was set as follows: temperature of transfer line, source, and quadrupole – 320 °C, 230 °C, and 150 °C, respectively, and 70 eV voltage of ionization. Detection was performed in total scan mode at 30–400 m/z. The NIST 17.1 library and literature data were applied to compare the acquired mass spectra and the retention index (RI). Indexes of linear retention were evaluated under the conditions applied for the TEO analysis using a mixture of C8–C20 saturated alkanes. The relative abundance of each constituent was presented as a percentage content based on peak area normalization (according to the MassHunter Workstation software version B.09.00). The analysis was carried out in triplicate.
2.4 Determination of strains phylogentic relationship using ERIC-PCR
In the present study, the Enterobacterial Repetitive Intergenic Consensus–Polymerase Chain Reaction (ERIC-PCR) method was employed to amplify repetitive genetic sequences in the genomes of the analyzed microorganisms (Dawson et al., 2002). The reaction was carried out using two primers–ERIC1 (5′ CACTTAGGGGTCCTCGAATGTA 3′) and ERIC2 (5′ AAGTAAGTAGTGGGGTGAGCG 3′) (Genomed, Warsaw, Poland). Genomic DNA isolated from the investigated strains served as the template, while nuclease-free water (A&A Biotechnology, Gdansk, Poland), a commercial PCR Mix Plus Green kit (A&A Biotechnology, Gdansk, Poland), and distilled water were used to prepare the reaction mixture.
For each ERIC-PCR assay, 25 µL of the reaction mixture was prepared, comprising 0.3 µL of primer ERIC1, 0.3 µL of primer ERIC2, 12.5 µL of PCR Mix Plus Green, 1.9 µL of nuclease-free water, and 10 µL of genomic DNA (final volume: 25 µL). Amplification was performed in a thermal cycler (TaKaRa PCR Thermal Cycler, TaKaRa Bio Inc., Kusatsu, Japan) for 30 cycles under the following conditions: initial denaturation at 95 °C for 10 min, denaturation at 90 °C for 30 s, primer annealing at 52 °C for 1 min, and elongation at 65 °C for 8 min, followed by a final elongation step at 65 °C for 16 min.
The resulting PCR products were analyzed by 2% (w/v) agarose gel electrophoresis (Cleaver PowerPro 300, Thistle Scientific Limited, Uddingston, UK). For this purpose, 2.8 g of agarose (Bio-Rad Laboratories, Hercules, CA, United States) was dissolved in a mixture of 112 mL distilled water and 28 mL 5× TBE (Tris Borate EDTA) buffer (BioShop, Burlington, Canada), and the solution was brought to a boil. Subsequently, 6 µL Midori Green (NIPPON Genetics Europe, Düren, Germany) dye was added, and the gel was poured into a tray once the solution had cooled slightly. Prior to loading, 4 µL of 6× loading buffer (A&A Biotechnology, Gdansk, Poland) was added to each 20 µL PCR product. DNA markers (5 µL of DNA Marker 3, A&A Biotechnology, Gdansk, Poland, and 100 bp DNA Ladder H3 RTU, NIPPON Genetics Europe, Düren, Germany) were applied to the first and last wells. Electrophoresis proceeded for approximately 60 min, and the gels were visualized using the Gel Doc XR+ Gel Documentation System (Bio-Rad Laboratories, Hercules, CA, United States). ERIC-PCR patterns were analyzed via visual assessment, and the dendrograms were generated with the UPGMA (Unweighted Pair Group Method with Arithmetic Mean) method using online software http://insilico.ehu.es/.
The strains were divided into three groups (marked group1, group2, group3) according to their genetic relationship. Subsequently, all parameters characterizing biofilm features and P. aeruginosa tolerance to the tested compounds were analyzed across the groups. For this purpose, mean values for each strain were calculated first.
2.5 Culture conditions
The experiments were performed in a Tryptic Soy Broth medium (TSB, Biomaxima, Lublin, Poland). The bacteria were incubated in the medium overnight at 37 °C, then suspended in saline for each test and 1.5 × 108 CFU/mL (Colony-Forming Unit) was adjusted with a densitometer DEN-1B (SIA Biosan Riga, Latvia). Such suspensions were diluted 1,000× in the medium and used in further analysis.
2.6 Evaluation of biofilm biomass and biofilm metabolic activity using crystal violet and tetrazolium staining
The total biofilm mass and metabolic activity were assessed using crystal violet and tetrazolium staining, respectively, to characterize formed pseudomonal biofilms. Three independent experiments in four replicates were performed, and a mean was calculated for each strain. For both tests, biofilms were cultured as follows. The diluted pseudomonal suspensions prepared as described in the section Culture conditions were added to the wells of 96-well polystyrene plates (Wuxi Nest Biotechnology, Wuxi, China) at the volume of 200 µL and incubated under static conditions for 24 h at 37 °C. After the incubation, the medium was gently removed. For the biofilm biomass evaluation, the 20% (v/v) crystal violet (Chempur, Piekary Slaskie, Poland) water solution was poured into the wells at 200 μL, and the setting was incubated at room temperature for 10 min. Subsequently, the solution was pipetted out, the biofilm cells were washed with 200 µL of 0.9% (w/v) sodium chloride (Chempur, Piekary Slaskie, Poland), and the plates were kept at 37 °C for 10 min. Then, 200 µL of 30% (v/v) acetic acid (Chempur, Piekary Slaskie, Poland) water solution was added, and the plates were subjected to shaking at 450 rpm (Mini-shaker PSU-2T, SIA Biosan Riga, Latvia) at room temperature until the crystals dissolved. Finally, the absorbance of the solution was measured at 550 nm using a spectrophotometer (MultiScan Go, Thermo Fischer Scientific, Waltham, MA, United States). For the biofilm metabolic activity assessment, the formed biofilms were stained with 200 µL of 0.1% (w/v) tetrazolium chloride solution (2,3,5-triphenyl-tetrazolium chloride) Sigma-Aldrich, Saint Louis, MO, United States in TSB medium for 2 h. Subsequently, the solution was removed, and the stained biofilm cells were dried for 10 min at 37 °C. To dissolve formazan crystals, a solution of methanol (Chempur, Piekary Slaskie, Poland) and acetic acid (9:1 ratio) was poured into the wells (200 µL), and the plates were shaken for 30 min at room temperature at 450 rpm. Next, the absorbance of the colour solution was measured with the spectrophotometer at 490 nm.
2.7 Assessment of the antimicrobial activity
2.7.1 Modified disk diffusion method
The modified disk diffusion method was performed according to the protocol included in our previous study (Dydak et al., 2021). To apply the tested compounds for the experiment, biocellulose disks (BC) 15 mm in diameter were prepared by culturing a Komagataeibacter xylinus ATCC 53524 strain in the Herstin–Schramm (H-S) medium. The medium was comprised of 2% (w/v) glucose (Chempur, Piekary Slaskie, Poland), 0.05% (w/v) MgSO4·7H2O (Chempur, Piekary Slaskie, Poland), 0.5% (w/v) bacto-peptone (VWR, Radnor, PA, United States), 0.115% (w/v) citric acid monohydric (Chempur, Piekary Slaskie, Poland), 0.5% (w/v) yeast extract (VWR, Radnor, PA, United States), 0.27% (w/v) Na2HPO4 (POCH), and 1% (v/v) ethanol (Chempur, Piekary Slaskie, Poland). Firstly, the H-S medium was added to the wells of a 24-well plate (Wuxi Nest Biotechnology, Wuxi, China) at the volume of 1 mL, and inoculated with K. xylinus. The plates were incubated under static conditions for 7 days at 28 °C. The BC disks were transferred to a bottle and washed with 0.1 M NaOH (Chempur, Piekary Slaskie, Poland) at 80 °C to remove unadhered cells and cell debris, then washed with double-distilled water until neutral pH. Next, the BC disks were sterilized in an autoclave. For research purposes, the weight of BC disks ranged from 0.6 to 0.7 g. The sterile BC disks were placed into fresh 24-well plates and 1 mL of undiluted TEO or PHMB or PBS (Phosphate-Buffered Saline, Merck, KGaA, Darmstadt, Germany, growth control) and kept for 24 h at 4 °C to soak the disks. To evaluate the concentration of absorbed compounds, six fresh BC disks were dried for 24 h at 37 °C and then weighed. The average difference between wet and dry BC disks was approximately 0.67 g. Therefore, the concentration of TEO/PHMB after the soaking was calculated as follows:
X—a volume of TEO/PHMB (mL).
The disk diffusion test was performed using the undiluted 1.5 × 108 CFU/mL pseudomonal suspensions in saline, prepared as described in the section Culture conditions. The bacteria were seeded onto TSA (Tryptic Soy Agar, Biomaxima, Lublin, Poland, 90 mm diameter, 14.2 mm height, 5 mm agar thickness) Petri dishes (Noex, Komorniki, Poland). Next, the soaked BC disks were put onto the agar, and the plates were incubated for 24 h/37 °C. After the incubation, zones of bacterial growth inhibition were measured (in mm) with a ruler. The BC disk diameter (15 mm) was subtracted from the obtained results. When the measured zone was irregular, a shorter diameter was chosen. One independent experiment in three repetitions was performed.
2.7.2 Microdilution method
The tested compounds’ Minimal Inhibitory Concentration was evaluated in 96-well plates according to the methodology presented in our previous study (Brożyna et al., 2024). TEO was applied as an emulsion in the medium and Tween 20 (VWR, Radnor, PA, United States). Firstly, the stock emulsion was prepared by combining TEO with Tween 20 and vigorous vortexing. Next, the medium was added, and the entire mixture was vortexed. TEO constituted 40% (v/v), and Tween 20 constituted 2% (v/v) of the stock emulsion volume. Subsequently, the stock emulsion was geometrically diluted in falcon tubes with TSB, and 100 µL of each concentration was added to a separate well. In the case of PHMB, the geometrical dilutions were prepared in the medium, and 100 µL was also added to each well. Subsequently, the diluted pseudomonal suspensions prepared as described in the section Culture conditions were added at the volume of 100 µL to the wells with the tested compounds. Therefore, the actual concentration applied to the bacterial cells ranged from 20% to 0.01% (v/v) for TEO and from 50% to 0.1% (v/v) for PHMB (concerning 100% as an undiluted PHMB solution). The plates were incubated for 24 h at 37 °C with continuous shaking at 450 rpm. Moreover, the following controls were prepared: controls of pseudomonal growth (bacteria in TSB), medium sterility (TSB only), control of Tween’s 20 antipseudomonal activity (Tween 20 at geometric concentrations ranging from 1% to 0.002% (v/v) in TSB). Once the incubation was finished, 1% (w/v) tetrazolium chloride solution in the medium was added to each well at 20 μL, and all plates were returned to the incubator for 2 h. The MIC value was found in the first well, where no formazan red colour was detected. The experiment was performed once with six replicates. In the case of differentiated MIC between the replicates, the higher value was chosen if it repeated at least three times. The MIC values were presented as percentage values, concerning 100% as undiluted compounds.
2.8 Assessment of antibiofilm activity and number of biofilm-forming cells using quantitative culturing
The antibiofilm activity of the studied compounds was assessed in 48-well plates (Wuxi Nest Biotechnology, Wuxi, China), and the methodology was adapted from our previous study (Brożyna et al., 2024). The compounds were applied at the highest possible concentrations. TEO was applied at the concentration of 5% (v/v) [as an emulsion in TSB with 2% (v/v) Tween 20], and PHMB was used as 50% (v/v) solution in TSB (concerning 100% as undiluted PHMB solution). At higher TEO concentrations, fragile pseudomonal biofilm was destroyed during TEO removal from above the biofilm structure due to the high density of TEO emulsion. In the first step, the diluted pseudomonal suspensions prepared as described in the section Culture conditions were added to the wells at a volume of 500 µL and incubated under static conditions for 24 h at 37 °C. Subsequently, the medium was removed from the wells, and 500 µL of the TEO, PHMB, or TSB was added. Moreover, the influence of 2% (v/v) Tween 20 on the biofilm of the P4 strain was examined. After 24-hour incubation at 37 °C, quantitative culturing was carried out. For this purpose, the medium was removed from the wells and biofilm cells were unattached from the surface by six-times pipetting and shaking at 600 rpm twice for 30 s with 2 × 500 µL of 0.1% (w/v) saponin (VWR, Radnor, PA, United States) solution in water. Finally, the samples were serially diluted in sodium chloride. Next, 10 µL bacterial spots were seeded onto TSA plates, and the plates were incubated for 24 h/37 °C. The bacterial colonies were counted, and each sample’s CFU/mL was calculated. The experiment was performed in one biological repetition with three technical repeats. The results of the growth control (bacteria in TSB) were additionally used to evaluate the number viable cell number (CFU/mL) as a parameter for biofilm characterization. The volume of seeded bacteria was 10 μL; therefore, a minimal CFU/mL number detected according to the methodology was 100. However, a 100 CFU/mL value was assumed if no colony was observed in this dilution. For each sample treated with TEO or PHMB, the percentage reduction of biofilm cells was calculated in relation to the mean of the growth control. The results for the separate strains were previously changed to a log10 scale to present them on a graph.
2.9 Visualization of live and dead biofilm-forming cells using fluorescent dyes and fluorescence microscopy
The antibiofilm effect of the examined compounds against P. aeruginosa was confirmed microscopically using a fluorescent microscope. The method was presented in our previous study (Brożyna et al., 2022). For this research, P857 and ATCC 15442 strains were selected. The biofilm culturing and its treatment with TEO or PHMB, along with the controls, were performed according to the protocol described in the Assessment of antibiofilm activity and number of biofilm-forming cells using the quantitative culturing section. However, the experiment was conducted on 24-well plates, and the volume of applied reagents was 1 mL. After the biofilm’s incubation with the tested compounds, the staining with a fluorescent dye and microscopic visualization were performed as follows. Filmtracer™ LIVE/DEAD™ Biofilm Viability Kit (Thermo Fischer Scientific, Waltham, MA, United States) prepared according to the manufacturer’s instruction was applied as a dye to assess membrane integrity. A total of 10 µL of the reagent was added to each well of a 24-well plate for 15 min (room temperature, darkness). Next, the cells were washed once with 200 µL of double-distilled water and then fixed with 4% (v/v) formaldehyde (Chempur, Piekary Slaskie, Poland) for 2 h. After fixation, formaldehyde was removed, and the biofilms were left to dry at room temperature in darkness. After drying, samples were analyzed using a fluorescence microscope Etaluma 600 LumaScope (object lens with magnification 4×, Etaluma, Carlsbad, CA, United States). The whole surface (100%) of wells was recorded and combined (tilted) to form an image panel showing changes in biofilm after exposure to the tested compounds.
2.10 Statistical analysis
Calculations were performed using R [Version 4.4.3; Rstudio software, Posit PBC, Boston, MA, United States (2025-02-13)]. The distribution and variance homogeneity were calculated using Shapiro-Wilk and Levene test, respectively. The distribution was also inspected visually. Welch-ANOVA followed by post hoc Games-Howell test was performed to compare differences between biofilm mass and biofilm metabolic activity across Pseudomonas strains. The correlation between biofilm mass and metabolic activity was fitted to the linear and exponential function, with the estimated R2 value. Kruskal–Wallis followed by Dunn’s test was performed to compare differences between inhibition zone values, MIC values and number of colony-forming units across Pseudomonas strains, as well as between conditions (TEO and PHMB) within each strain. The same test was performed to compare differences in investigated parameters across groups of genetically distinct strains of Pseudomonas. The results of statistical analyses with a significance level of p < 0.05 were considered significant.
3 Results
3.1 Evaluation of TEO chemical composition
In the first step of the research, the percentage content of the TEO’s components was assessed using GC-MS methodology (Table 1). Thymol prevailed over other components and constituted half of the percentage composition. Except for carvacrol (which exceeded the standard range at 0.15%) and carvacrol methyl ether (which was not present), all components aligned with the European Pharmacopoeia 11 standards.
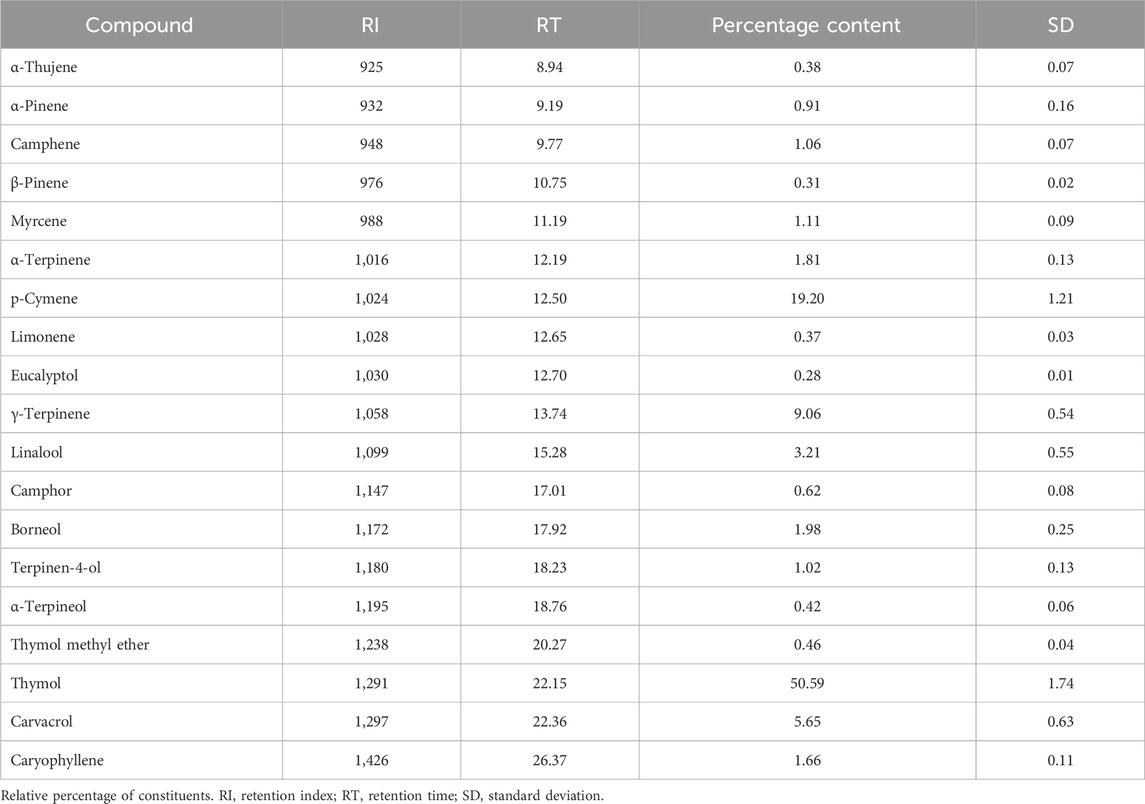
Table 1. Composition of Thyme Essential Oil (TEO) assessed by GC–MS (Gas Chromatography-Mass Spectrometry).
3.2 Strains phylogentic relationship
To explore the genetic diversity among the studied P. aeruginosa strains, ERIC-PCR fingerprinting was performed, and dendrograms were generated using the UPGMA method with Dice similarity coefficients. A ≥90% similarity threshold was used to define clonally related strains, while clusters sharing <80% similarity were considered genetically distinct. Based on banding pattern analysis, three groups (group1, group2, and group3) were delineated (Figure 1). Notably, strains P20 and P68 (group3) exhibited the lowest similarity to all other isolates, suggesting a divergent genetic background.
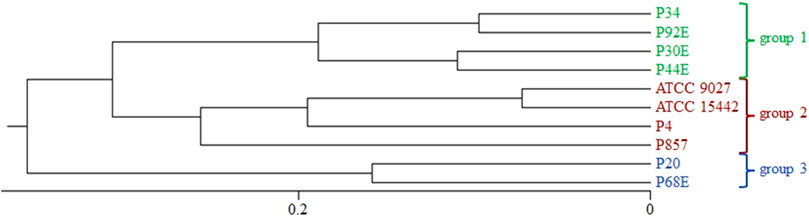
Figure 1. Phylogenetic clustering of P. aeruginosa strains based on ERIC-PCR (Enterobacterial Repetitive Intergenic Consensus–Polymerase Chain Reaction) fingerprinting. The dendrogram was constructed using the UPGMA (Unweighted Pair Group Method with Arithmetic Mean) and Dice similarity index. A ≥90% similarity threshold was applied to define genetically related groups (indicated with green, red and blue colours). Group3 strains (P20, P68) formed a separate cluster with <80% similarity to the remaining isolates.
3.3 Biofilm features
Phenotypic characterization of P. aeruginosa biofilms included quantification of biomass, metabolic activity, and viable cell counts (Figure 2). The distribution of these features across individual strains is detailed in Figure 3, Tables 2 and 3, and Supplementary Tables S1–S4. All strains formed biofilm under the applied conditions, though the extent varied markedly. Metabolic activity exhibited greater inter-strain variability than biomass, with statistically significant differences observed among 70% of strains (Welch’s ANOVA with Games-Howell post hoc, p < 0.05). Notably, strains of group3: P20 and P68, showed the lowest values for both biomass and metabolic activity, differing significantly from group1 and group2 strains. In contrast, strain P34 demonstrated exceptionally high metabolic activity. No correlation was observed between biomass and metabolic activity levels (Supplementary Figure S1). Differences in CFU/mL across strains were less pronounced and not consistently statistically significant, likely due to limited sample size and variability, which reduced the sensitivity of non-parametric tests (Kruskal–Wallis with Dunn’s post hoc, Supplementary Table S4).
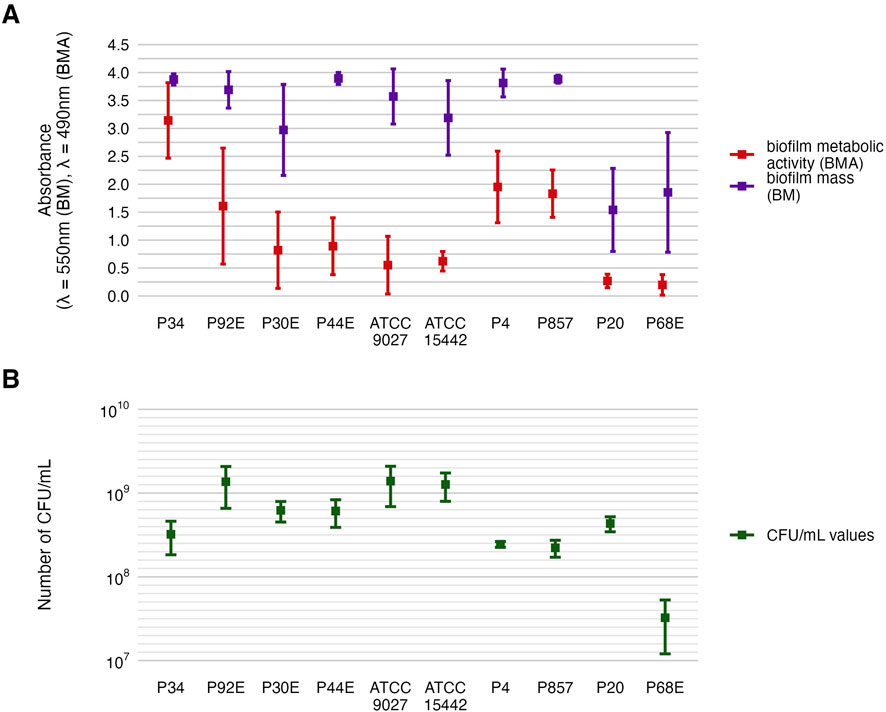
Figure 2. Characterization of P. aeruginosa strains (n = 10) biofilms. The average biofilm biomass and metabolic activity. (A) The mean number of biofilm Colony-Forming Unit/mL (CFU/mL). (B) The error lines represent the standard deviation.

Figure 3. Visual representation of the data distribution for biofilm features of separate P. aeruginosa strains (n = 10). Distribution of biofilm mass. (A,B) Distribution of biofilm metabolic activity. (C,D) The horizontal lines indicate the median value, the points indicate the mean value, the error lines indicate the standard deviation, the boxes indicate the interquartile range.
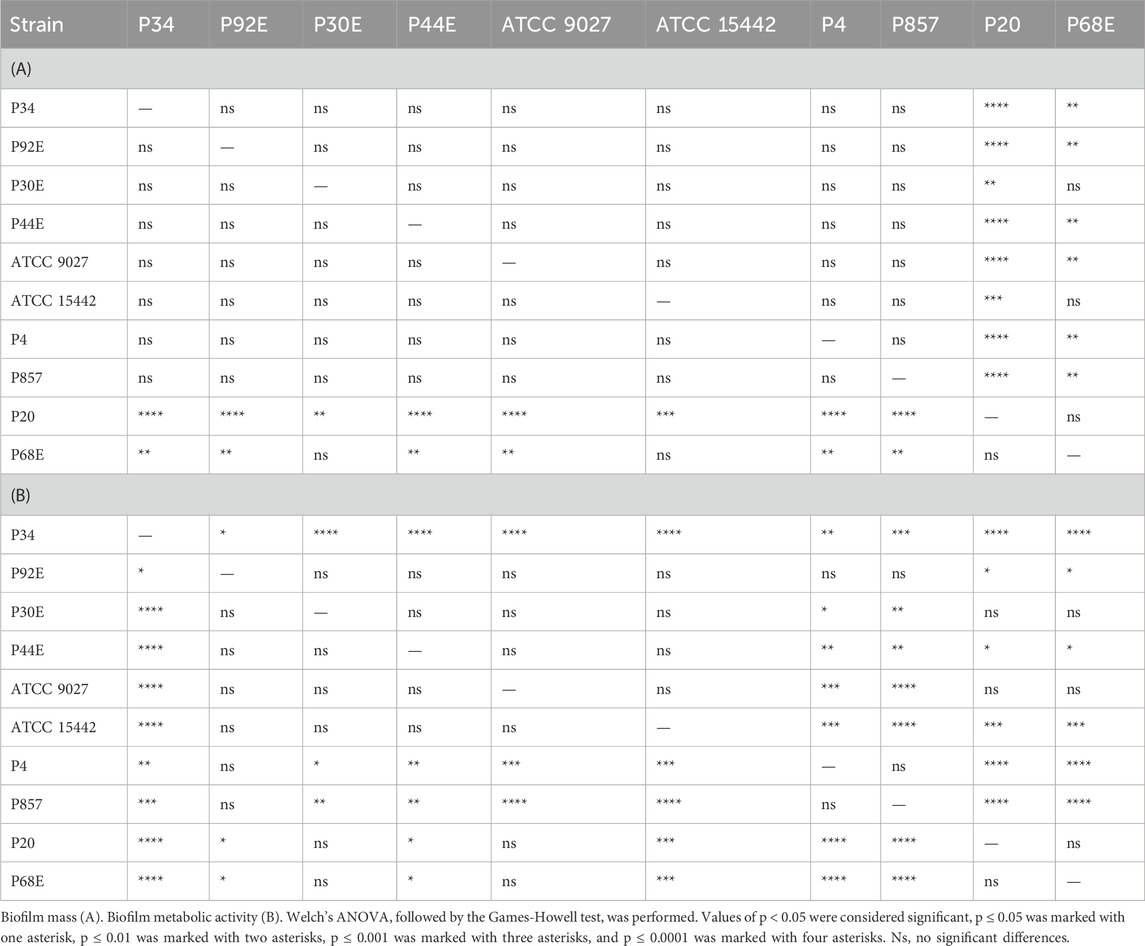
Table 2. Summarized statistical differences in biofilm features between particular P. aeruginosa strains (n = 10).
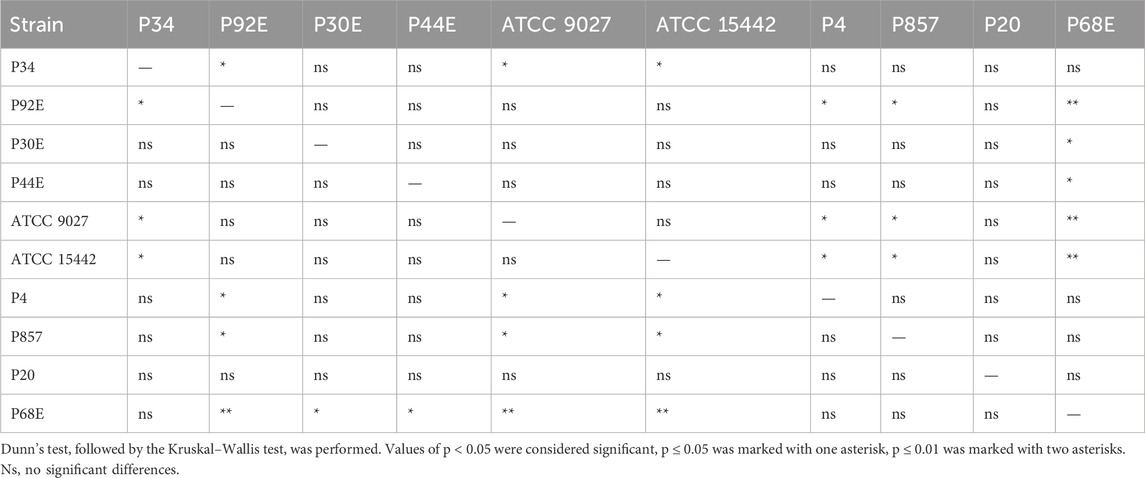
Table 3. Summarized statistical differences in biofilm CFU/mL (Colony-Forming Unit) number between particular P. aeruginosa strains (n = 10).
These findings highlight a pronounced intra-species variability in biofilm-forming capacity among P. aeruginosa strains, with metabolic activity emerging as a particularly discriminative trait. The genetically divergent group3 strains (P20 and P68) consistently exhibited the weakest biofilm-forming phenotypes, aligning with their distinct ERIC-PCR profiles. Conversely, other strains displayed a wide spectrum of biomass and metabolic activity levels independent of viable cell count.
3.4 Antimicrobial and antibiofilm activity
Next, we assessed the susceptibility of P. aeruginosa strains in planktonic forms to TEO and PHMB using two complementary approaches: a modified disk diffusion assay and a microdilution method (Figures 4, 5; Tables 4 and 5; Supplementary Tables S5, S6, S9, and S10). In the disk diffusion setup, bacterial cellulose (BC) disks were soaked with undiluted compounds, delivering an estimated final concentration of 60%. Both methods revealed marked strain-specific variability in response to TEO. The MIC values spanned a 1000-fold range, and inhibition zone diameters ranged from 0 to 7.7 mm, reflecting substantial differences in planktonic tolerance. In general, susceptibility profiles were consistent across methods, with most TEO-tolerant strains in microdilution also displayed reduced inhibition zones, except for strain P857.
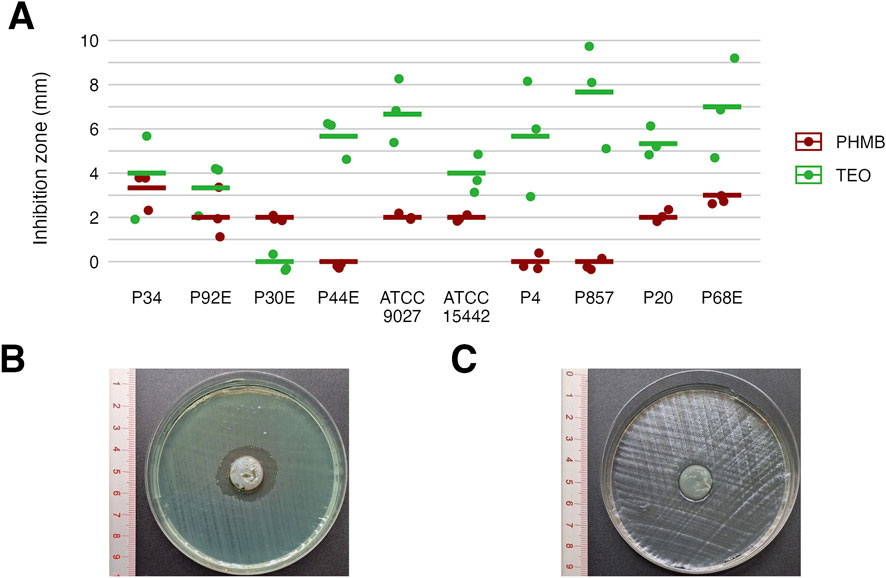
Figure 4. Antimicrobial activity of the tested compounds against P. aeruginosa strains assessed using a modified disk diffusion method. Results for diameters of growth inhibition zones (mm) (n = 10). (A) Representative zones of P857 strain growth inhibition after the treatment with Thyme Essential Oil (TEO). (B) Representative zones of P857 strain growth inhibition after the treatment with polyhexanide (PHMB). (C) The lines represent a mean value.
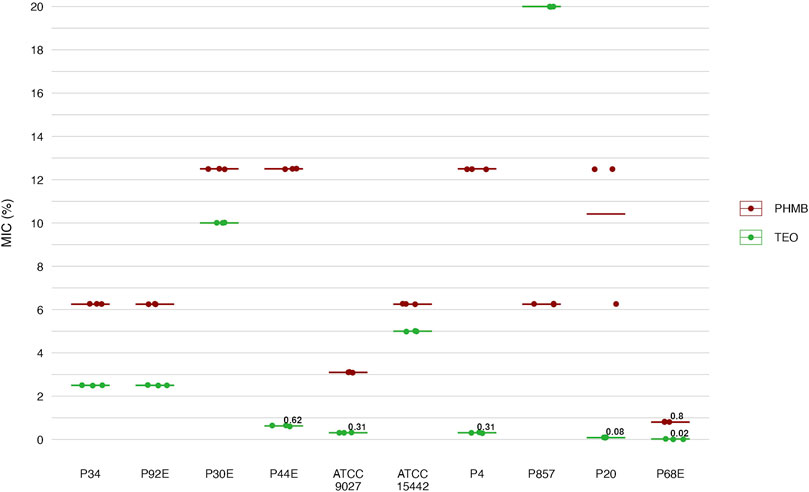
Figure 5. Antimicrobial activity of the tested compounds against P. aeruginosa (n = 10) strains assessed with a microdilution method. MIC, Minimal Inhibitory Concentration (%, v/v). TEO, Thyme Essential Oil; PHMB, polyhexanide. The lines represent a mean value.
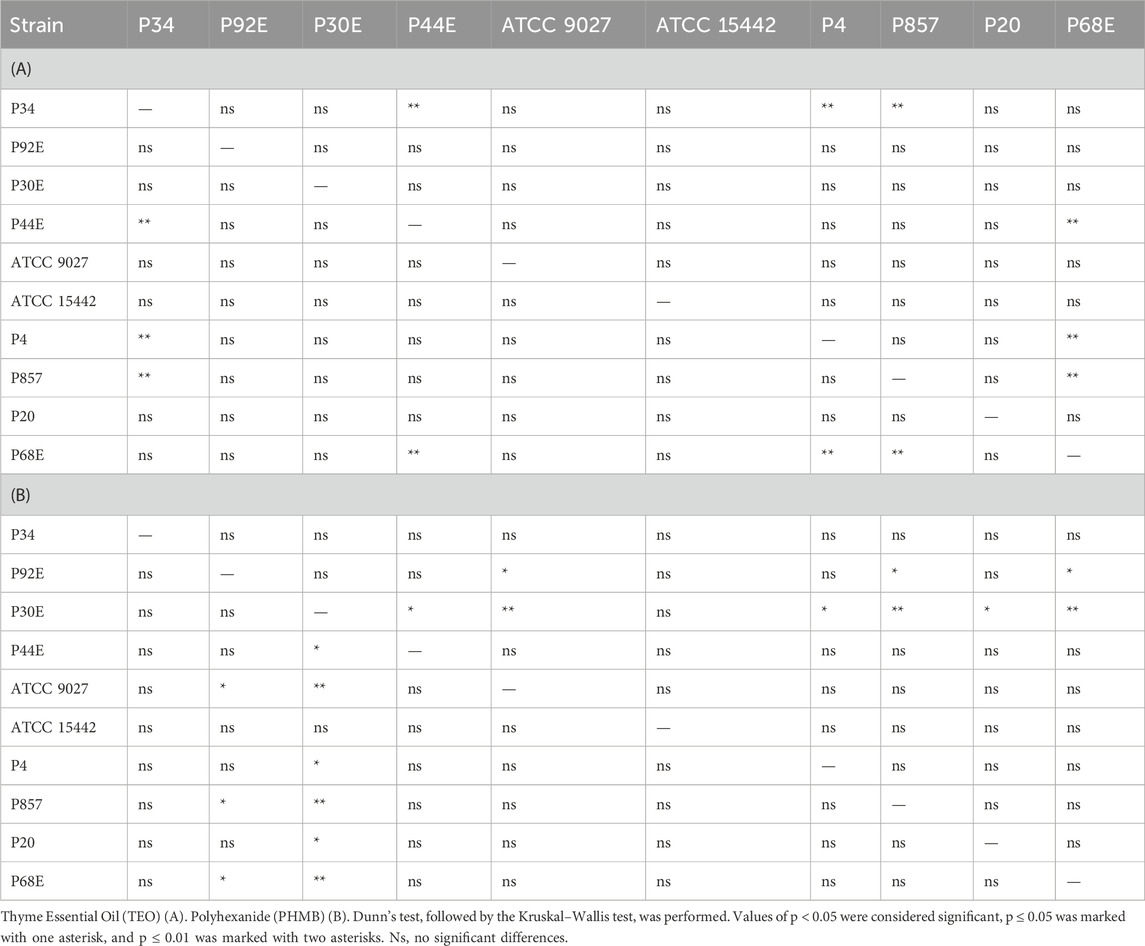
Table 4. Summarized statistical differences in susceptibility to the tested compounds between particular P. aeruginosa strains (n = 10) assessed using a modified disk diffusion method.
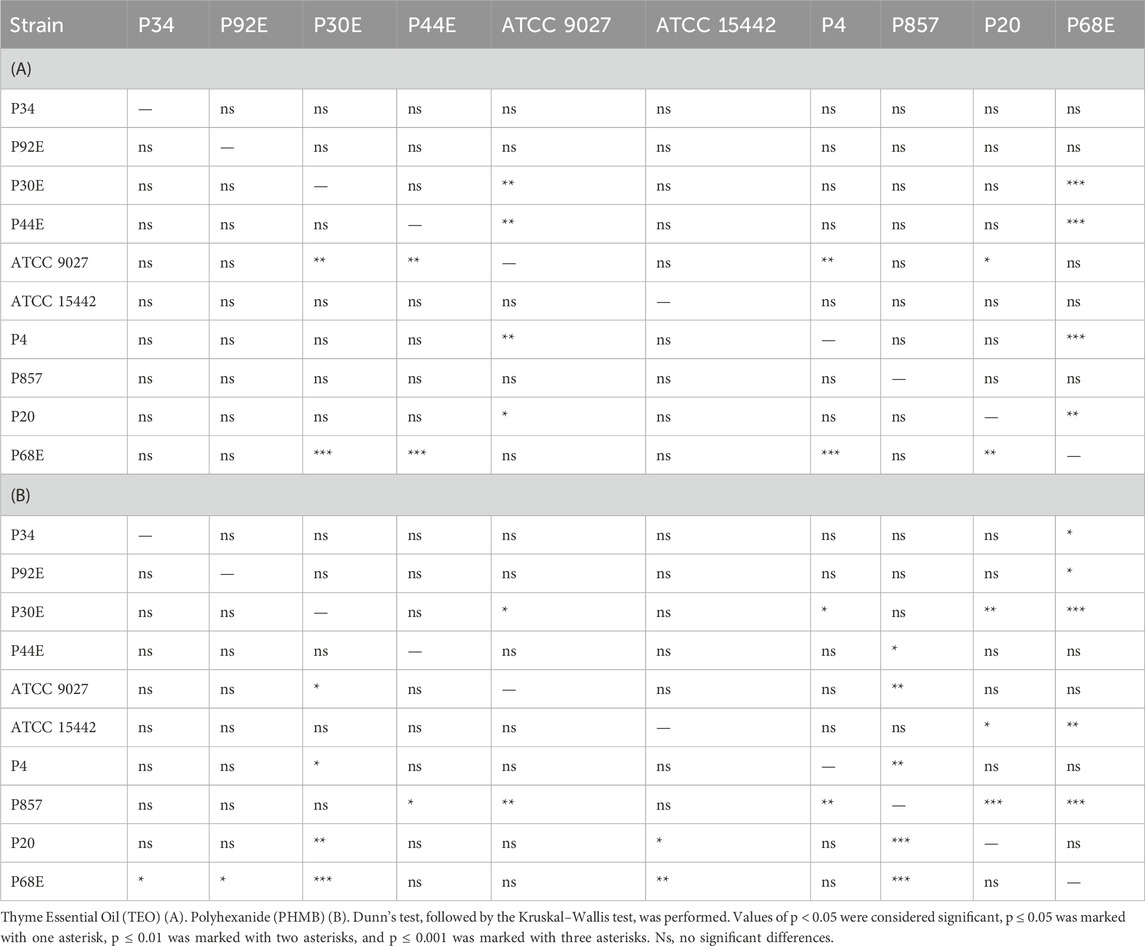
Table 5. Summarized statistical differences in susceptibility to the tested compounds between particular P. aeruginosa strains (n = 10) assessed with a microdilution method.
Notably, strains P20 and P68—previously shown to form the weakest biofilms—were among the most susceptible to TEO. By contrast, PHMB exhibited more uniform activity across the strain panel, with MIC values clustering between 6.3% and 12.5% (v/v) in 80% of isolates and inhibition zones ranging narrowly between 2.0 and 3.3 mm in 70% of cases. Overall, TEO showed superior antipseudomonal activity compared to PHMB in 90% of strains, with statistically significant differences observed in both assays (p < 0.05, Pairwise Dunn’s Test). Interestingly, some strains (e.g., P4, P44) demonstrated high susceptibility to TEO while showing relative resistance to PHMB, highlighting divergent mechanisms of action and potential strain-specific vulnerabilities.
In the subsequent step of the experiment, the antibiofilm activity of the tested compounds was evaluated using quantitative culturing (Figure 6; Table 6; Supplementary Tables S7 and S8). The antibiofilm effect of PHMB was significantly (p < 0.05, Pairwise Dunn’s Test) higher than the one displayed by TEO against six of ten strains (Supplementary Table S11). However, higher variability in the strain’s tolerance to PHMB was observed (Supplementary Table S8). The mean log reduction in biofilm cells after the treatment with PHMB ranged from approx. 2 to 7. In the case of TEO, the log reduction between 2 and 3.3 was determined for all strains except for P68.
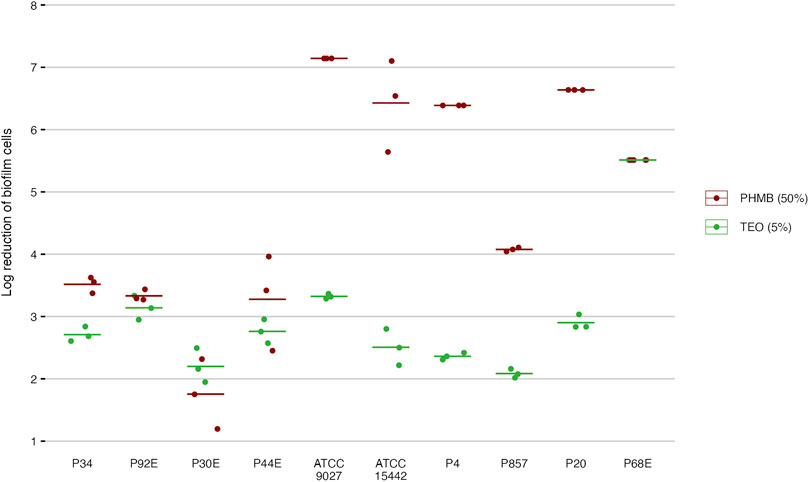
Figure 6. Antibiofilm activity of the tested compounds against P. aeruginosa strains (n = 10) expressed as a reduction (log10) of biofilm cells. TEO, Thyme Essential Oil, PHMB, polyhexanide. The lines represent a mean value.
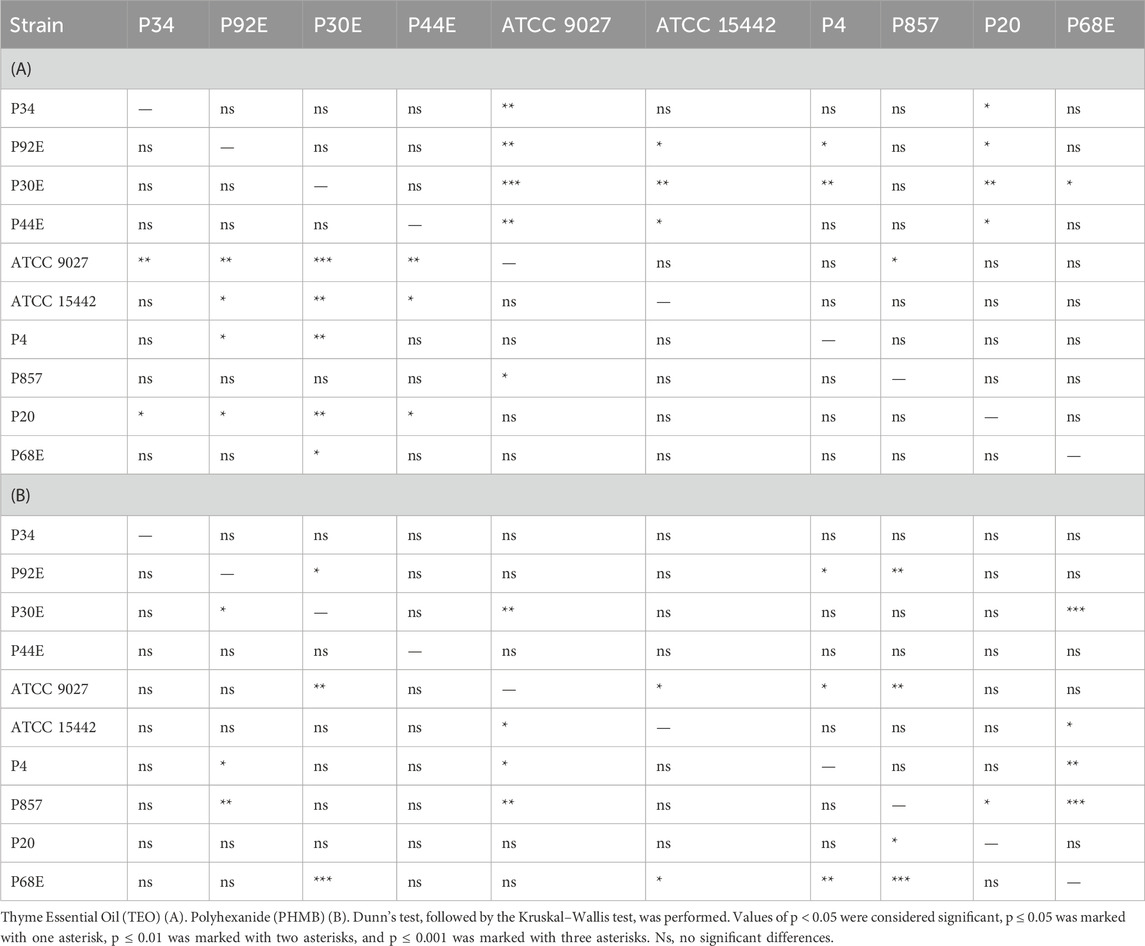
Table 6. Summarized statistical differences in susceptibility to the tested compounds between particular P. aeruginosa strains (n = 10) expressed as biofilm reduction (%).
The activity of TEO and PHMB against P. aeruginosa ATCC 15442 and P857 biofilms was confirmed also using a fluorescence microscope (Figure 7). A higher number of bacterial cells with altered/damaged cell walls (red/orange color) was observed after the biofilms were exposed to TEO than to PHMB.
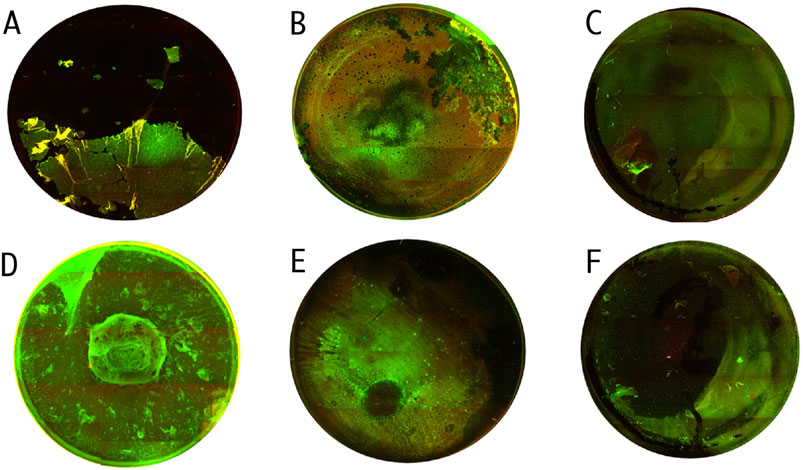
Figure 7. Microscopic visualizations of the P. aeruginosa biofilm (covering the surface of a 24-well plate) stained with a LIVE/DEAD dye. ATCC 15442 strain’s untreated biofilm. (A) ATCC 15442 strain’s biofilm treated with Thyme Essential Oil. (B) ATCC 15442 strain’s biofilm treated with polyhexanide. (C) P857 strain’s untreated biofilm. (D) P857 strain’s biofilm treated with Thyme Essential Oil. (E) P857 strain’s biofilm treated with polyhexanide. (F) The red/orange color shows bacterial cells with altered/damaged cell walls; the green color indicates unaltered cell walls. Fluorescence microscope Etaluma 600 (magnification 4×). The well diameter was 15 mm.
3.5 Analysis across the groups
Subsequently, all parameters characterizing biofilm features and P. aeruginosa tolerance to the tested compounds were analyzed across the groups (Figures 8–11; Supplementary Tables S12–S15). The biofilm mass and metabolic activity of group3 were significantly lower in comparison to group1 and group2 (p < 0.05, Dunn’s test, Supplementary Table S14). No statistically significant differences in biofilm-forming cell numbers were observed between groups. However, the parameter was at the lowest level in group3.
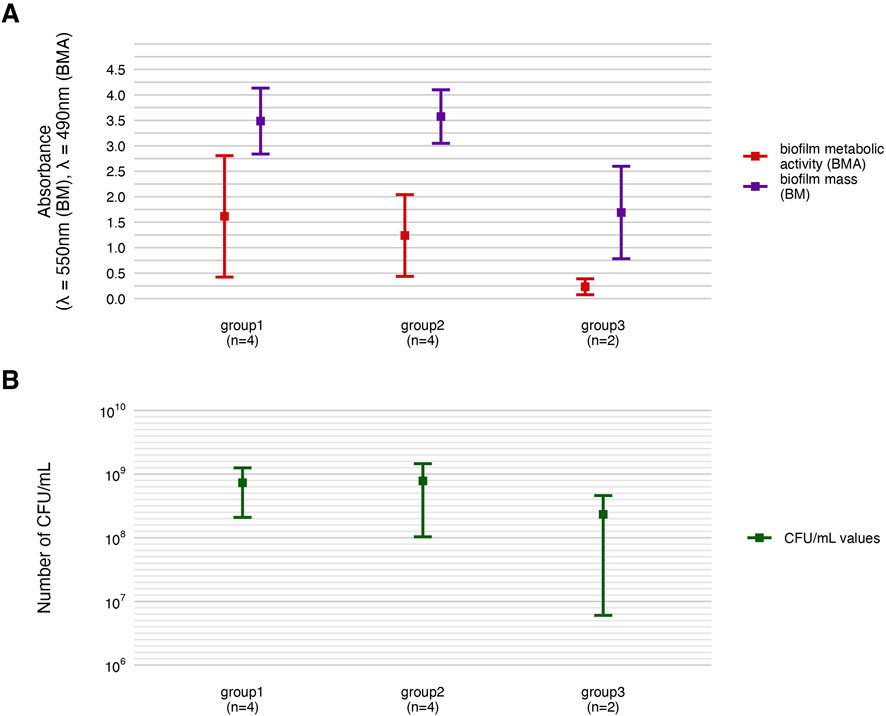
Figure 8. Comparison of biofilm features between genetically distinct groups of P. aeruginosa strains. The mean biofilm biomass and metabolic activity. (A) The mean number of biofilm CFU/mL (Colony-Forming Unit). (B) The error lines represent the standard deviation.
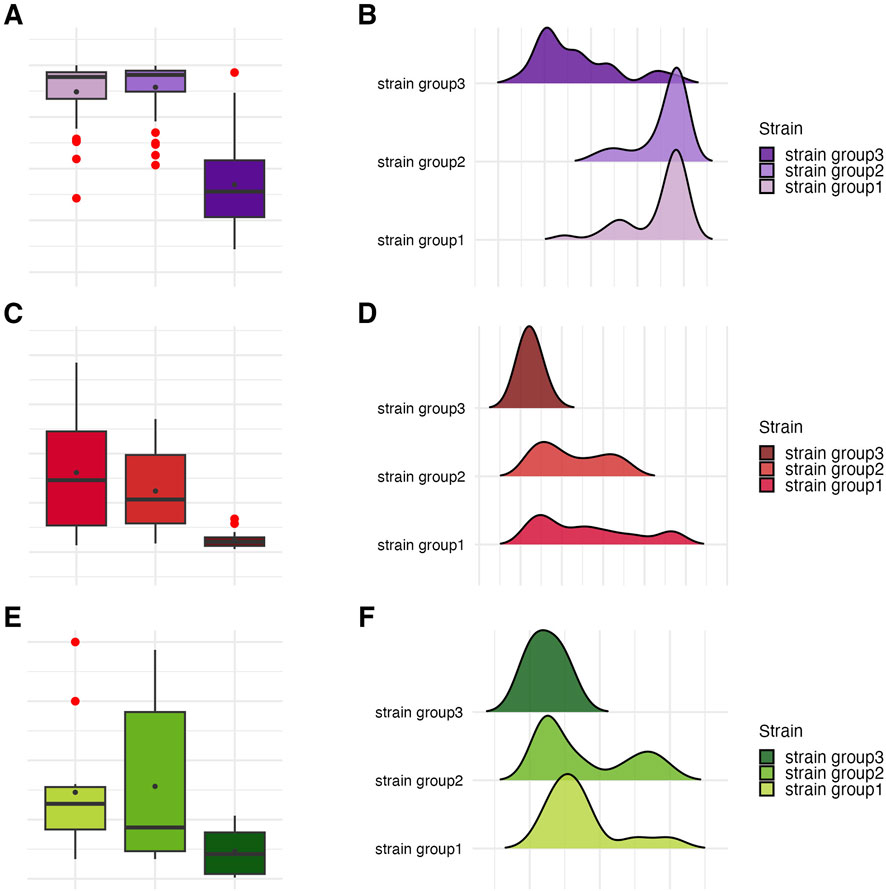
Figure 9. Visual representation of the data distribution for biofilm features across genetically distinct groups of P. aeruginosa strains. Distribution of biofilm mass. (A,B) Distribution of biofilm metabolic activity. (C,D) Distribution of biofilm Colony-Forming Unit number (CFU/mL). (E,F) The horizontal lines indicate the median value, the points indicate the mean value, the error lines indicate the standard deviation, the boxes indicate the interquartile range.
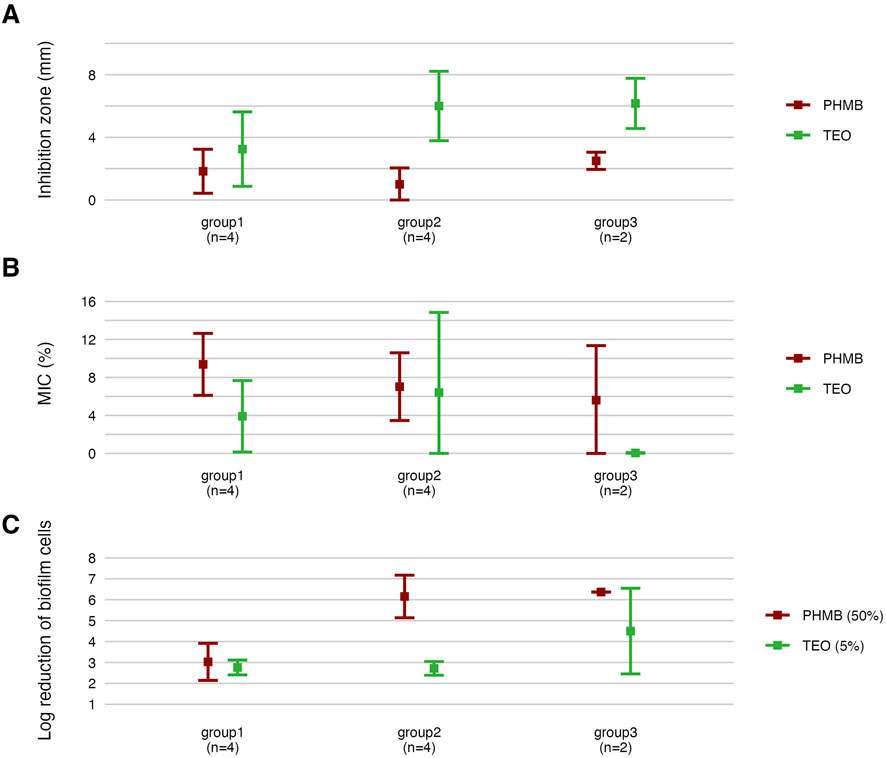
Figure 10. Comparison of antimicrobial and antibiofilm activity of the tested compounds against genetically distinct groups of P. aeruginosa strains. The mean diameters of growth inhibition zones (mm). (A) The mean values of Minimal Inhibitory Concentration (%, v/v) (MIC). (B) The mean reduction (log10) of biofilm cells. (C) TEO, Thyme Essential Oil, PHMB, polyhexanide. The error lines represent a standard deviation.
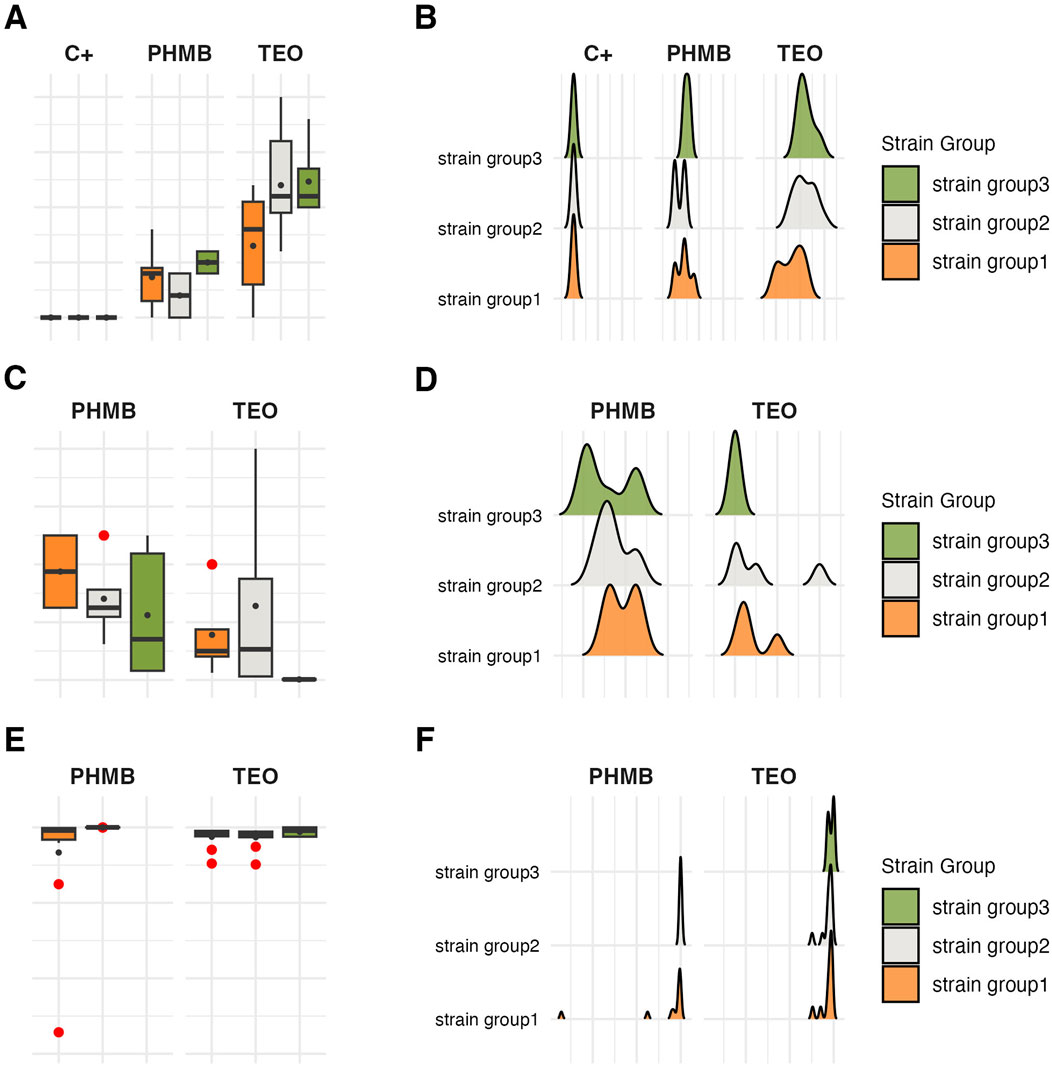
Figure 11. Visual representation of the data distribution for antimicrobial and antibiofilm activity of the tested compounds against genetically distinct groups of P. aeruginosa strains. Distribution of growth inhibition zones values (mm). (A,B) Distribution of Minimal Inhibitory Concentration (MIC) (%, v/v). (C,D) Distribution of biofilm cells reduction (%). (E,F) The horizontal lines indicate a median value, the spots indicate a mean value, the error lines indicate a standard deviation, and the boxes indicate the interquartile range. TEO, Thyme Essential Oil; PHMB, polyhexanide C+, growth control.
Strains in group3 exhibited significantly lower tolerance (except for no significant difference with group2 in the growth inhibition zones) to TEO than group1 and group2 in their planktonic forms and biofilms (p < 0.05, Dunn’s test, Figure 10 and Supplementary Table S15). However, in the case of group3, tolerance to PHMB was not as significantly different from other groups as in TEO.
The summarization of results is presented in a heat-map (Figure 12).
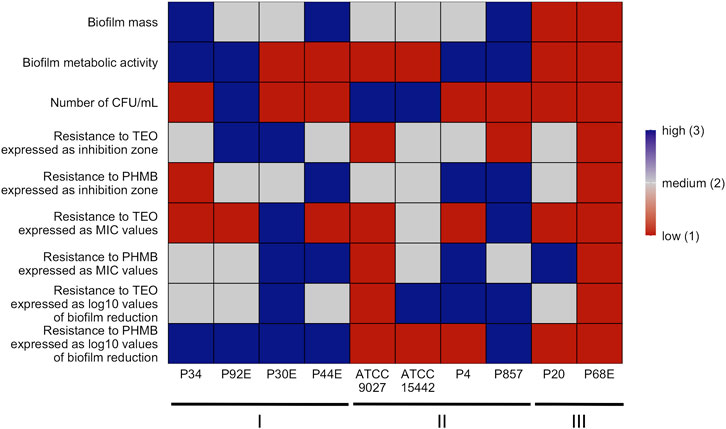
Figure 12. Summary of P. aeruginosa strains’ biofilm features and pseudomonal resistance to Thyme Essential Oil (TEO) or PHMB (polyhexanide) in genetically distinct groups. A heatmap was generated by setting a mean of all strains for each parameter ± 25% as high when the value was above the range (blue), low when the value was below the range (red) and medium when the value was equal to the range (grey). CFU/mL, Colony-Forming Unit/mL. MIC, Minimal Inhibitory Concentration.
The integrative comparison of biofilm phenotypes and antimicrobial susceptibility across genetically defined groups revealed consistent trends supporting the biological relevance of ERIC-PCR-based clustering. Strains belonging to group3 demonstrated overall attenuated biofilm-forming capacity, reflected in significantly lower biomass, metabolic activity, and viable cell counts compared to group1 and group2. In antimicrobial assays, the modified disk diffusion (MDD) method identified 30% of strains as exhibiting low resistance to TEO and 20% as showing low resistance to PHMB. Notably, half of the strains in groups2 and 3 were classified as low TEO-resistant, whereas half of the group1 strains exhibited high resistance to this compound. Consistent with this, the microdilution method revealed that 75% of group1 strains displayed low resistance to TEO. Across the entire panel, 7 out of 10 strains demonstrated low resistance to TEO, while only 2 strains exhibited low resistance to PHMB. When assessing biofilm-associated tolerance, a greater proportion of strains showed reduced susceptibility to PHMB than to TEO. Only 20% of isolates displayed low biofilm-level resistance to TEO. Remarkably, all group1 strains were highly resistant to PHMB in the biofilm model, while 75% of group2 strains showed the inverse pattern—high resistance to TEO and low resistance to PHMB. Group3 strains exhibited uniformly low or intermediate resistance to both compounds. Individual strain-level profiles further emphasized this heterogeneity: P34, P44E, and P857, which formed dense biofilms, were markedly resistant to PHMB in the biofilm state; ATCC 9027 displayed low-to-moderate resistance to both agents across all models; strain 30E was consistently less susceptible to TEO regardless of the method; and P68E combined weak biofilm formation with low resistance to both compounds, making it the most responsive strain in the panel.
4 Discussion
The rationale for this study arose from the growing recognition that antimicrobial efficacy cannot be reliably inferred from single-strain or single-method experiments—particularly when both the pathogen (Pseudomonas aeruginosa) and the antimicrobial agent (Thyme Essential Oil) exhibit inherent biological and chemical heterogeneity, respectively. Our findings confirmed extensive intra-species variability in biofilm-forming capacity and susceptibility to both Thyme Essential Oil and chemically synthesized polyhexanide antiseptic, affecting planktonic and biofilm-associated phenotypes alike. Importantly, this variability was not random but followed, to a certain extent, genetic stratification patterns, suggesting that distinct lineages may harbor intrinsic differences in tolerance mechanisms. Moreover, susceptibility profiles were strongly influenced by the assay system used, underscoring the importance of model selection in evaluating complex antimicrobials, such as essential oils.
These observations resonate with a broader shift in the field toward antimicrobial precision and stewardship. For antibiotics, susceptibility testing is already governed by international standards, with clearly defined breakpoints and reference strains that enable reproducible, clinically meaningful assessments (The European Committee on Antimicrobial Susceptibility Testing, 2025b). In the case of antiseptics, the concept of “antiseptic stewardship” has recently gained traction, driven by reports of existing or emerging tolerance to compounds such as chlorhexidine, octenidine, and PHMB (Kampf, 2024). Yet essential oils—despite their high potential in topical therapy—remain largely exempt from such critical scrutiny. This is particularly concerning given that they are often marketed and applied under the assumption of universal efficacy and low resistance potential (Golus et al., 2016; Orchard and Van Vuuren, 2017).
Our data challenges this assumption. We demonstrate that even a pharmacopoeia-standardized essential oil, dominated by thymol, exhibits highly variable activity depending on the genetic background of the strain and the biological context in which it is tested. In this light, we advocate for a new conceptual framework: essential oil stewardship. This would entail systematic evaluation of chemically defined oils against diverse and clinically relevant microbial panels supported by multi-model testing strategies (Hulankova, 2024). Such an approach is urgently needed to move beyond anecdotal efficacy claims and establish a rational, evidence-based foundation for phytocompound deployment in infection control (Ali et al., 2025).
In this study, we controlled for chemical composition by using a pharmacopoeia-grade TEO batch characterized via GC–MS (Table 1), thereby eliminating compound heterogeneity as a confounder. This allowed us to isolate and examine three primary variables driving the observed differences in antimicrobial outcomes: (i) the genetic identity of the P. aeruginosa strain, (ii) the physiological state of the bacteria-planktonic versus biofilm-associated, and (iii) the specific in vitro model employed. These basic dimensions, yet they can be further developed, shaped both the magnitude and pattern of susceptibility.
As mentioned, P. aeruginosa strains were stratified into three distinct genetic groups (Figure 1). The phenotypic profiling of biofilm-forming capacity (Figures 2, 3) revealed marked inter-strain differences in both biomass and metabolic activity among all of them, despite identical growth conditions. While all ten P. aeruginosa strains formed detectable biofilms, the magnitude and functional characteristics varied significantly. Notably, metabolic activity exhibited a broader dynamic range than biomass, and no consistent correlation was observed between the two parameters, suggesting divergent regulatory pathways underpinning biofilm quantity and viability. Group3 strains (P20, P68) exhibited consistently attenuated biofilm traits, while others (e.g., P34) showed disproportionately high metabolic output. Viable cell counts (Table 3), although less discriminatory, reinforced this heterogeneity. These divergent biofilm phenotypes may have critical implications for susceptibility to TEO, whose antimicrobial activity depends on both direct contact and vapor-phase diffusion (Reyes-Jurado et al., 2020; Brożyna et al., 2022). Strains producing high biofilm biomass—with abundant extracellular matrix—may experience reduced diffusion of TEO components, particularly hydrophobic monoterpenes like thymol, thereby conferring a physical barrier to penetration (Akbarian et al., 2022). However, if metabolic activity within the biofilm remains high, increased uptake of volatile or partially soluble fractions may paradoxically sensitize certain subpopulations (Castaneda et al., 2016). Conversely, low-biomass, low-activity biofilms, such as those formed by group3 strains, likely lack such structural protection and may be more uniformly exposed to the active compounds—potentially explaining their higher susceptibility in subsequent assays (Sadiq et al., 2017). In turn, strains such as P34, which produced metabolically hyperactive but structurally less complex biofilms might remain vulnerable due to elevated uptake or metabolic engagement with the oil components. These nuanced relationships suggest that resistance to EOs is not solely a function of matrix density or cell number, but rather an emergent property of the interplay between biofilm architecture, physiological state, and compound delivery mode (Figures 4–6; Tables 4–6).
In turn, PHMB, obtained via chemical synthesis, is a cationic polymer that exerts its antimicrobial effect through strong electrostatic interactions with the negatively charged bacterial surfaces, leading to membrane disruption, increased permeability, and, ultimately, intracellular interference—particularly with DNA (Renzoni et al., 2017). Notably, PHMB does not require active metabolism for uptake and retains efficacy irrespective of bacterial physiological state. This may explain why strains forming dense or metabolically quiescent biofilms (e.g., P44E, P857) remained susceptible to PHMB despite showing reduced sensitivity to TEO. In our study, PHMB was applied as a commercial wound irrigation solution, also containing the amphiphilic surfactant undecylenamidopropyl betaine. This additive likely enhanced biofilm penetration by reducing the surface tension and improving compound dispersion, which may have contributed to the overall uniformity of PHMB’s antibiofilm activity across strains (Machuca et al., 2019). This observation offers a relevant translational insight: the inclusion of surfactants or other delivery-enhancing excipients may represent a promising strategy for improving the performance of EOs, not only by facilitating their penetration into biofilm structures, but also by reducing the variability of outcomes. The mechanistic divergence between PHMB and TEO, thus underscores not only their differential interaction with biofilm architectures, but also the importance of formulation in optimizing and standardizing antimicrobial efficacy. Our findings emphasize that compound-specific delivery profiles must be carefully considered when interpreting biofilm susceptibility data, especially for complex, multicomponent agents such as EOs.
Our results also highlight critical methodological limitations associated with plate-based biofilm assays. Repeated washing steps, often necessary in classical protocols, can disrupt or incompletely remove loosely attached biomass, leading to an underestimation of actual biofilm architecture (Kragh et al., 2019). This effect was clearly visualized in the performed fluorescence microscopy panel (Figure 7), where the peripheral and central regions of the same well displayed markedly different biofilm structures—even under uniform culture conditions. Such heterogeneity implies that selective imaging of “representative” areas, a common practice in low-throughput settings may introduce substantial observational bias. In contrast, our approach, i.e., mapping entire wells and correlating imaging data with quantitative metrics, revealed spatial variation that would likely be missed by partial sampling. This well-to-well and intra-well structural variability adds yet another layer to the complex landscape of antimicrobial response diversity. In future work, we aim to further dissect this spatial dimension of biofilm behavior to refine the interpretation of biofilm susceptibility results and reduce methodological bias.
The integrative heat map (Figure 12) reflects the cumulative outcome of all prior phenotypic, susceptibility, and structural analyses, serving as a functional convergence point of the study. Patterns observed in biofilm mass, metabolic activity, and viable cell counts (Figures 2, 3; Table 3) translate directly into biofilm-specific resistance profiles. Strains from group3 exhibited a unique signature: high overall biofilm biomass, yet low cell density and metabolic activity, implying a disproportionately high extracellular matrix content. This matrix-dominant architecture likely hindered the diffusion of the hydrophobic components of TEO, resulting in moderate resistance, but paradoxically did not protect as effectively against PHMB, whose cationic, amphiphilic nature may enable adsorption and deeper matrix penetration (Campanac et al., 2002; Li et al., 2015). In contrast, group1 strains, particularly P34 and P30E, demonstrated high resistance to TEO, correlating with elevated metabolic activity and denser, more viable biofilms, which may support active detoxification or efflux (Crabbé et al., 2019). The heat map also exposes strain-specific nuances within genetically related groups, as exemplified by ATCC 9027 or P857 (group2), which displayed an atypically high tolerance to PHMB. The heat map confirms that biofilm-mediated tolerance is not determined by any single parameter but arises from the interplay of genotypic background, matrix-to-cell ratio, and biofilm vitality—further modulated by the molecular and formulation properties of the tested compounds.
In summary, our findings underscore the complexity of assessing antimicrobial efficacy against P. aeruginosa, a species marked by substantial intra-species heterogeneity in both planktonic and biofilm states. We demonstrate that resistance to antiseptic agents—even those of pharmacopeial grade—depends not solely on compound potency, but on the interplay between microbial architecture and compound formulation.
Although the present study is extensive, encompassing a genetically stratified P. aeruginosa panel, complementary biofilm models, and detailed formulation parameters, we consider its limitations as challenges that guide the next stages of our essential oil stewardship programme. First, all experiments were performed under controlled in vitro conditions, which allowed us to isolate and compare genotypic and phenotypic variables, but did not capture host–pathogen interactions or physiological complexity. Building on our growing experience with Galleria mellonella infection models, we plan to extend these evaluations into in vivo settings that incorporate immune response dynamics (Brożyna et al., 2024; Krzyżek et al., 2025). Second, our endpoints focused on phenotypic outcomes (biomass, metabolic activity, CFU counts, microscopy), whereas the underlying molecular mechanisms remain to be elucidated. Future work will integrate metabolomics, proteomics, lipidomics, and genomics to reveal pathways beyond membrane disruption, particularly in the context of the hormetic responses we observed, where certain strains increased metabolic activity at sub-MIC concentrations of TEO. Third, we intentionally limited our scope to a single essential oil of pharmacopoeia grade and one bacterial species to ensure control and interpretability. In subsequent studies, we aim to expand the framework to other essential oils and clinically relevant microorganisms, enabling cross-comparison and the refinement of standardised evaluation protocols. Finally, we did not evaluate the potential cytotoxicity of TEO or PHMB towards eukaryotic cells. We expect that cytotoxicity testing will require the same level of standardisation as antimicrobial testing, and its integration into the stewardship framework will be crucial for delivering a complete and translationally relevant profile of essential oil-based interventions.
Beyond these study-specific considerations, adapting or adopting essential oil stewardship in a global context presents challenges that extend beyond the scope of a single oil–pathogen combination. The first is the intrinsic chemical variability of essential oils, driven by plant genotype, cultivation practices, harvest timing and extraction methods, which complicates standardisation even when the same botanical species is used. Second, there are no internationally agreed reference strain panels, interpretive criteria, or breakpoints for EO activity, making cross-study and cross-laboratory comparisons difficult. Third, regulatory frameworks for EO classification differ widely between regions, ranging from medicinal product to food additive or cosmetic, which directly influences testing requirements and acceptable endpoints. Fourth, different microbial targets and infection environments (e.g., wound biofilms versus respiratory tract biofilms) require distinct model systems, each with its own strengths and limitations. Fifth, the issue of optimal EO delivery - whether in liquid formulations with emulsifiers, nanoemulsions, or as volatile vapours - remains insufficiently addressed, despite clear evidence that formulation profoundly influences antimicrobial outcomes. Finally, advanced analytical and modelling approaches (e.g., GC–MS, omics profiling, in vivo infection models) may be inaccessible to laboratories in resource-limited settings, highlighting the need for simplified yet reproducible alternatives that preserve the principles of stewardship.
Despite these challenges, the proposed stewardship framework offers clear opportunities. The approach can be extended to other essential oils and a broader range of microorganisms, including Gram-positive bacteria, fungi, and viruses, while recognising that these organisms form structurally and functionally distinct biofilms. Therefore, model systems and evaluation criteria will need to be tailored to reflect the specific architecture, physiology, and microenvironment of each biofilm type to ensure that the stewardship framework remains both accurate and applicable. Coordinated, multi-centre studies could generate global databases of EO chemical profiles and corresponding antimicrobial outcomes, providing the basis for evidence-based breakpoints and reference panels. Advances in formulation science, such as nanoemulsions, encapsulation, surfactant-assisted delivery, or controlled-release matrices, could reduce variability in biofilm penetration and enhance reproducibility. Integration of EO stewardship into One Health strategies could align plant-derived antimicrobials with efforts to mitigate antimicrobial resistance across human, animal, and environmental health sectors. Finally, establishing minimal international standards for EO testing - analogous to EUCAST (European Committee on Antimicrobial Susceptibility Testing) or CLSI (Clinical and Laboratory Standards Institute) guidelines for antibiotics -would facilitate regulatory harmonisation and accelerate the responsible translation of EO-based interventions into clinical and biotechnological practice.
The intention of this study was not to present Thymus vulgaris essential oil as an immediately deployable antimicrobial solution, but rather to use it as a model to demonstrate the variability, limitations, and methodological challenges that currently affect essential oil research. This approach is deliberate: it aims to avoid the trend of over-interpreting preliminary activity data and instead to establish a framework - the essential oil stewardship concept - that prioritises standardisation, reproducibility, and systematic development before translation to clinical or practical use. Recognising the constraints and variability of EO activity is, in our view, an essential prerequisite for responsibly identifying, optimising, and ultimately validating formulations with real-world potential.
Data availability statement
All data generated or analyzed during this study are included in the article/Supplementary Material, further inquiries can be directed to the corresponding authors. The data is also openly available at the PPM repository at https://ppm.umw.edu.pl/info/researchdata/UMW78858a5e11404ade9fb2debcc652bd59/.
Author contributions
MB: Conceptualization, Data curation, Formal Analysis, Funding acquisition, Investigation, Methodology, Project administration, Supervision, Visualization, Writing – original draft. ZS: Conceptualization, Data curation, Investigation, Software, Visualization, Writing – original draft. NT: Investigation, Writing – original draft. BD: Investigation, Writing – original draft. KK: Investigation, Writing – original draft. AM: Writing – review and editing. YS: Writing – review and editing. MT: Writing – original draft, Writing – review and editing. AJ: Conceptualization, Formal Analysis, Funding acquisition, Investigation, Project administration, Supervision, Visualization, Writing – original draft, Writing – review and editing.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. This research was funded (regarding biofilm features testing) by the National Science Centre, Poland (Grant No. 2021/41/N/NZ6/03305) and by Wroclaw Medical University, Poland (regarding antimicrobial testing) subsidy funds no SUBK.D230.22.040.
Acknowledgments
The Authors would like to express our gratitude to Weronika Kozłowska from Division of Pharmaceutical Biotechnology, Department of Pharmaceutical Biology and Biotechnology, Wroclaw Medical University, Wroclaw, Poland for the help in the evaluation of essential oils composition. For the purpose of Open Access, the author has applied a CC-BY public copyright license to any Author Accepted Manuscript (AAM) version arising from this submission.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fphar.2025.1659096/full#supplementary-material
References
Akbarian, M., Chen, S. H., Kianpour, M., Farjadian, F., Tayebi, L., and Uversky, V. N. (2022). A review on biofilms and the currently available antibiofilm approaches: matrix-destabilizing hydrolases and anti-bacterial peptides as promising candidates for the food industries. Int. J. Biol. Macromol. 219, 1163–1179. doi:10.1016/j.ijbiomac.2022.08.192
Ali, S. S., Al-Tohamy, R., Al-Zahrani, M., Badr, A., and Sun, J. (2025). Essential oils and plant-derived bioactive compounds: a comprehensive review of their therapeutic potential, mechanisms of action, and advances in extraction technologies. Phytochem. Rev. doi:10.1007/s11101-025-10123-8
Azeredo, J., Azevedo, N. F., Briandet, R., Cerca, N., Coenye, T., Costa, A. R., et al. (2017). Critical review on biofilm methods. Crit. Rev. Microbiol. 43, 313–351. doi:10.1080/1040841X.2016.1208146
Balaban, N. Q., Helaine, S., Lewis, K., Ackermann, M., Aldridge, B., Andersson, D. I., et al. (2019). Definitions and guidelines for research on antibiotic persistence. Nat. Rev. Microbiol. 17, 441–448. doi:10.1038/s41579-019-0196-3
Basch, E., Ulbricht, C., Hammerness, P., Bevins, A., and Sollars, D. (2004). Thyme (Thymus vulgaris L.), thymol. J. Herb. Pharmacother. 4, 49–67. doi:10.1080/j157v04n01_07
Birkenhauer, E., Neethirajan, S., and Weese, J. S. (2014). Collagen and hyaluronan at wound sites influence early polymicrobial biofilm adhesive events. BMC Microbiol. 14, 191. doi:10.1186/1471-2180-14-191
Brożyna, M., Paleczny, J., Kozłowska, W., Ciecholewska-Juśko, D., Parfieńczyk, A., Chodaczek, G., et al. (2022). Chemical composition and antibacterial activity of liquid and volatile phase of essential oils against planktonic and biofilm-forming cells of Pseudomonas aeruginosa. Molecules 27, 4096. doi:10.3390/molecules27134096
Brożyna, M., Dudek, B., Kozłowska, W., Malec, K., Paleczny, J., Detyna, J., et al. (2024). The chronic wound milieu changes essential oils’ antibiofilm activity—an in vitro and larval model study. Sci. Rep. 14, 2218. doi:10.1038/s41598-024-52424-6
Campanac, C., Pineau, L., Payard, A., Baziard-Mouysset, G., and Roques, C. (2002). Interactions between biocide cationic agents and bacterial biofilms. Antimicrob. Agents Chemother. 46, 1469–1474. doi:10.1128/AAC.46.5.1469-1474.2002
Castaneda, P., McLaren, A., Tavaziva, G., and Overstreet, D. (2016). Biofilm antimicrobial susceptibility increases with antimicrobial exposure time. Clin. Orthop. Relat. Res. 474, 1659–1664. doi:10.1007/s11999-016-4700-z
Chandler, C. E., Horspool, A. M., Hill, P. J., Wozniak, D. J., Schertzer, J. W., Rasko, D. A., et al. (2019). Genomic and phenotypic diversity among ten laboratory isolates of Pseudomonas aeruginosa PAO1. J. Bacteriol. 201, e00595-18. doi:10.1128/JB.00595-18
Coimbra, A., Ferreira, S., and Duarte, A. P. (2022). Biological properties of Thymus zygis essential oil with emphasis on antimicrobial activity and food application. Food Chem. 393, 133370. doi:10.1016/j.foodchem.2022.133370
Crabbé, A., Jensen, P. Ø., Bjarnsholt, T., and Coenye, T. (2019). Antimicrobial tolerance and metabolic adaptations in microbial biofilms. Trends Microbiol. 27, 850–863. doi:10.1016/j.tim.2019.05.003
Dawson, S. L., Fry, J. C., and Dancer, B. N. (2002). A comparative evaluation of five typing techniques for determining the diversity of fluorescent pseudomonads. J. Microbiol. Methods 50, 9–22. doi:10.1016/s0167-7012(02)00003-9
Dhekane, R., Mhade, S., and Kaushik, K. S. (2022). Adding a new dimension: multi-level structure and organization of mixed-species Pseudomonas aeruginosa and Staphylococcus aureus biofilms in a 4-D wound microenvironment. Biofilm 4, 100087. doi:10.1016/j.bioflm.2022.100087
Dhifi, W., Bellili, S., Jazi, S., Bahloul, N., and Mnif, W. (2016). Essential oils’ chemical characterization and investigation of some biological activities: a critical review. Medicines 3, 25. doi:10.3390/medicines3040025
Dydak, K., Junka, A., Dydak, A., Brożyna, M., Paleczny, J., Fijalkowski, K., et al. (2021). In vitro efficacy of bacterial cellulose dressings chemisorbed with antiseptics against biofilm formed by pathogens isolated from chronic wounds. Int. J. Mol. Sci. 22, 3996. doi:10.3390/ijms22083996
European Medicines Agency Committee on Herbal Medicinal Products (2019). Assessment report on Thymus vulgaris L., Thymus zygis L., aetheroleum. Available online at: https://www.fitoterapia.net/archivos/201910/draft-assessment-report-thymus-vulgaris-l-thymus-zygis-l-aetheroleum-revision-1_en.pdf (Accessed June 10, 2025).
Fernández-Billón, M., Llambías-Cabot, A. E., Jordana-Lluch, E., Oliver, A., and Macià, M. D. (2023). Mechanisms of antibiotic resistance in Pseudomonas aeruginosa biofilms. Biofilm 5, 100129. doi:10.1016/j.bioflm.2023.100129
Golus, J., Sawicki, R., Widelski, J., and Ginalska, G. (2016). The agar microdilution method – a new method for antimicrobial susceptibility testing for essential oils and plant extracts. J. Appl. Microbiol. 121, 1291–1299. doi:10.1111/jam.13253
Grace, A., Sahu, R., Owen, D. R., and Dennis, V. A. (2022). Pseudomonas aeruginosa reference strains PAO1 and PA14: a genomic, phenotypic, and therapeutic review. Front. Microbiol. 13, 1023523. doi:10.3389/fmicb.2022.1023523
Hudaib, M., Speroni, E., Di Pietra, A. M., and Cavrini, V. (2002). GC/MS evaluation of thyme (Thymus vulgaris L.) oil composition and variations during the vegetative cycle. J. Pharm. Biomed. Anal. 29, 691–700. doi:10.1016/S0731-7085(02)00119-X
Hulankova, R. (2024). Methods for determination of antimicrobial activity of essential oils in vitro—a review. Plants 13, 2784. doi:10.3390/plants13192784
Kampf, G. (2024). Antiseptic stewardship: biocide resistance and clinical implications. 2nd Edn. Cham: Springer.
Klockgether, J., Cramer, N., Wiehlmann, L., Davenport, C. F., and Tümmler, B. (2011). Pseudomonas aeruginosa genomic structure and diversity. Front. Microbiol. 2, 150. doi:10.3389/fmicb.2011.00150
Kragh, K. N., Alhede, M., Kvich, L., and Bjarnsholt, T. (2019). Into the well—a close look at the complex structures of a microtiter biofilm and the crystal violet assay. Biofilm 1, 100006. doi:10.1016/j.bioflm.2019.100006
Krzyżek, P., Dudek, B., Brożyna, M., Krzyżanowska, B., and Junka, A. (2025). Galleria mellonella larvae as a model for Helicobacter pylori biofilm formation under antibiotic stress. Microb. Pathog. 198, 107121. doi:10.1016/j.micpath.2024.107121
Li, X., Yeh, Y. C., Giri, K., Mout, R., Landis, R. F., Prakash, Y. S., et al. (2015). Control of nanoparticle penetration into biofilms through surface design. Chem. Commun. 51, 282–285. doi:10.1039/c4cc07737g
Machuca, J., Lopez-Rojas, R., and Fernandez-Cuenca, F. (2019). Comparative activity of a polyhexanide–betaine solution against biofilms produced by multidrug-resistant bacteria belonging to high-risk clones. J. Hosp. Infect. 103, e92–e96. doi:10.1016/j.jhin.2019.04.008
Mohammed, H. A., Sulaiman, G. M., Khan, R. A., Al-Saffar, A. Z., Mohsin, M. H., Albukhaty, S., et al. (2024). Essential oils pharmacological activity: chemical markers, biogenesis, plant sources, and commercial products. Process Biochem. 144, 112–132. doi:10.1016/j.procbio.2024.05.021
Nysten, J., Sofras, D., and Van Dijck, P. (2024). One species, many faces: the underappreciated importance of strain diversity. PLoS Pathog. 20, e1011931. doi:10.1371/journal.ppat.1011931
Orchard, A., and Van Vuuren, S. (2017). Commercial essential oils as potential antimicrobials to treat skin diseases. Evidence-based Complementary Altern. Med. 2017, 4517971. doi:10.1155/2017/4517971
Renzoni, A., Von Dach, E., Landelle, C., Diene, S. M., Manzano, C., Gonzales, R., et al. (2017). Impact of exposure of methicillin-resistant Staphylococcus aureus to polyhexanide in vitro and in vivo. Antimicrob. Agents Chemother. 61, e00272-17. doi:10.1128/AAC.00272-17
Reyes-Jurado, F., Franco-Vega, A., Ramírez-Corona, N., Palou, E., and López-Malo, A. (2015). Essential oils: antimicrobial activities, extraction methods, and their modeling. Food Eng. Rev. 7, 275–297. doi:10.1007/s12393-014-9099-2
Reyes-Jurado, F., Navarro-Cruz, A. R., Ochoa-Velasco, C. E., Palou, E., López-Malo, A., and Ávila-Sosa, R. (2020). Essential oils in vapor phase as alternative antimicrobials: a review. Crit. Rev. Food Sci. Nutr. 60, 1641–1650. doi:10.1080/10408398.2019.1586641
Roberts, E. L., Abdollahi, S., Oustadi, F., Stephens, E. D., and Badv, M. (2023). Bacterial-nanocellulose-based biointerfaces and biomimetic constructs for blood-contacting medical applications. ACS Mater. Au 3, 418–441. doi:10.1021/acsmaterialsau.3c00021
Romling, U., Schmidt, K. D., and Tummler, B. (1997). Large genome rearrangements discovered by the detailed analysis of 21 Pseudomonas aeruginosa clone C isolates found in environment and disease habitats. J. Mol. Biol. 271, 386–404. doi:10.1006/jmbi.1997.1186
Ruhluel, D., O’Brien, S., Fothergill, J. L., and Neill, D. R. (2022). Development of liquid culture media mimicking the conditions of sinuses and lungs in cystic fibrosis and health. F1000Res 11, 1007. doi:10.12688/f1000research.125074.2
Sadiq, F. A., Flint, S., Li, Y. J., Ou, K., Yuan, L., and He, G. Q. (2017). Phenotypic and genetic heterogeneity within biofilms with particular emphasis on persistence and antimicrobial tolerance. Future Microbiol. 12, 1087–1107. doi:10.2217/fmb-2017-0042
Sanchez, D. A., and Martinez, L. R. (2019). Underscoring interstrain variability and the impact of growth conditions on associated antimicrobial susceptibilities in preclinical testing of novel antimicrobial drugs. Crit. Rev. Microbiol. 45, 51–64. doi:10.1080/1040841X.2018.1538934
Santana de Oliveira, M., Vostinaru, O., Rigano, D., and de Aguiar Andrade, E. H. (2023). Editorial: bioactive compounds present in essential oils: advances and pharmacological applications. Front. Pharmacol. 14, 1130097. doi:10.3389/fphar.2023.1130097
Stahl-Biskup, E., and Venskutonis, R. P. (2012). “Thyme,” in Handbook of herbs and spices. 2nd Edn, Editor K. V. Peter (United Kingdom: Woodhead Publishing), 499–525.
Su, Y., Yrastorza, J. T., Matis, M., Cusick, J., Zhao, S., Wang, G., et al. (2022). Biofilms: formation, research models, potential targets, and methods for prevention and treatment. Adv. Sci. 9, e2203291. doi:10.1002/advs.202203291
The European Committee on Antimicrobial Susceptibility Testing (2020). EUCAST reading guide for broth microdilution. 1–20. Available online at: https://www.eucast.org/eucast_news/news_singleview?tx_ttnews%5Btt_news%5D=362&cHash=f2196d399c8618b7313ccb7f048f7af5 (Accessed June 10, 2025).
The European Committee on Antimicrobial Susceptibility Testing (2025a). EUCAST disk diffusion method for antimicrobial susceptibility testing reading guide. 1–30. Available online at: https://www.eucast.org/ast_of_bacteria/disk_diffusion_methodology (Accessed June 10, 2025).
The European Committee on Antimicrobial Susceptibility Testing (2025b). European committee on antimicrobial susceptibility testing breakpoint tables for interpretation of MICs and zone diameters. 1–118. Available online at: https://www.eucast.org/clinical_breakpoints (Accessed June 10, 2025).
Wilson, C., Lukowicz, R., Merchant, S., Valquier-Flynn, H., Caballero, J., Sandoval, J., et al. (2017). Quantitative and qualitative assessment methods for biofilm growth: a mini-review. Res. Rev. J. Eng. Technol. 6.
World Health Organization (2024). WHO bacterial priority pathogens list. Available online at: https://www.who.int/publications/i/item/9789240093461 (Accessed June 10, 2025).
Keywords: thyme essential oil, biofilm resistance, Pseudomonas aeruginosa, antimicrobial variability, essential oil stewardship
Citation: Brożyna M, Stępnicka Z, Tymińska N, Dudek B, Kapczyńska K, Matkowski A, Sun Y, Tomczyk M and Junka A (2025) Toward essential oil stewardship: strain-resolved evaluation of thyme oil activity against Pseudomonas aeruginosa. Front. Pharmacol. 16:1659096. doi: 10.3389/fphar.2025.1659096
Received: 03 July 2025; Accepted: 29 August 2025;
Published: 26 September 2025.
Edited by:
Cheorl-Ho Kim, Sungkyunkwan University, Republic of KoreaReviewed by:
Cynthia G. Suarez, Finzelberg GmbH & Co. KG, GermanyBanaz Jalil, University College London, United Kingdom
Copyright © 2025 Brożyna, Stępnicka, Tymińska, Dudek, Kapczyńska, Matkowski, Sun, Tomczyk and Junka. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Malwina Brożyna, bWFsd2luYS5icm96eW5hQHVtdy5lZHUucGw=; Zuzanna Stępnicka, enV6YW5uYS5zdGVwbmlja2FAdW13LmVkdS5wbA==
†These authors share first authorship