 Gabriella E. DiCarlo1,2†
Gabriella E. DiCarlo1,2† Samuel J. Mabry3,4†
Samuel J. Mabry3,4† Xixi Cao5
Xixi Cao5 Clara McMillan6
Clara McMillan6 Tiffany G. Woynaroski1,2,7,8
Tiffany G. Woynaroski1,2,7,8 Fiona E. Harrison1,9
Fiona E. Harrison1,9 India A. Reddy10
India A. Reddy10 Heinrich J. G. Matthies3
Heinrich J. G. Matthies3 Charles R. Flynn6
Charles R. Flynn6 Mark T. Wallace1,2,7,8,10
Mark T. Wallace1,2,7,8,10 Hui Wu5‡
Hui Wu5‡ Aurelio Galli3,4*‡
Aurelio Galli3,4*‡- 1Vanderbilt Brain Institute, Vanderbilt University Medical Center, Nashville, TN, United States
- 2Department of Hearing and Speech Sciences, Vanderbilt University Medical Center, Nashville, TN, United States
- 3Department of Surgery, University of Alabama Birmingham, Birmingham, AL, United States
- 4Department of Neurobiology, University of Alabama Birmingham, Birmingham, AL, United States
- 5School of Dentistry, Oregon Health and Science University, Portland, OR, United States
- 6Department of Surgery, Vanderbilt University Medical Center, Nashville, TN, United States
- 7Vanderbilt Kennedy Center, Vanderbilt University Medical Center, Nashville, TN, United States
- 8Frist Center for Autism and Innovation, Vanderbilt University, Nashville, TN, United States
- 9Department of Medicine, Vanderbilt University Medical Center, Nashville, TN, United States
- 10Department of Psychiatry and Behavioral Sciences, Vanderbilt University Medical Center, Nashville, TN, United States
Background: Altered dopamine (DA) signaling has been associated with autism spectrum disorder (ASD), a neurodevelopmental condition estimated to impact 1 in 54 children in the United States. There is growing evidence for alterations in both gastrointestinal function and oral microbiome composition in ASD. Recent work suggests that rare variants of the SLC6A3 gene encoding the DA transporter (DAT) identified in individuals with ASD result in structural and functional changes to the DAT. One such recently identified de novo mutation is a threonine to methionine substitution at position 356 of the DAT (DAT T356M). The DAT T356M variant is associated with ASD-like phenotypes in mice homozygous for the mutation (DAT T356M+/+), including social deficits, hyperactivity, and impaired DA signaling. Here, we determine the impact of this altered DA signaling as it relates to altered oral microbiota, and metabolic and gastrointestinal dysfunction.
Methods: In the DAT T356M+/+ mouse, we determine the oral microbiota composition, metabolic function, and gastrointestinal (GI) function. We examined oral microbiota by 16S RNA sequencing. We measured metabolic function by examining glucose tolerance and we probed gastrointestinal parameters by measuring fecal dimensions and weight.
Results: In the DAT T356M+/+ mouse, we evaluate how altered DA signaling relates to metabolic dysfunction and altered oral microbiota. We demonstrate that male DAT T356M+/+ mice weigh less (Wild type (WT) = 26.48 ± 0.6405 g, DAT T356M+/+ = 24.14 ± 0.4083 g) and have decreased body fat (WT = 14.89 ± 0.6206%, DAT T356M+/+ = 12.72 ± 0.4160%). These mice display improved glucose handling (WT = 32.60 ± 0.3298 kcal/g, DAT T356M+/+ = 36.97 ± 0.4910 kcal/g), and an altered oral microbiota. We found a significant decrease in Fusobacterium abundance. The abundance of Fusobacterium was associated with improved glucose handling and decreased body fat.
Conclusions: Our findings provide new insights into how DAT dysfunction may alter gastrointestinal function, composition of the oral microbiota, and metabolism. Our data suggest that impaired DA signaling in ASD is associated with a number of metabolic and gastrointestinal changes which are common in individuals with ASD.
Introduction
Autism spectrum disorder (ASD) is a neurodevelopmental condition characterized by early-emerging differences in social communication and interaction, and by patterns of restrictive and repetitive interests, behaviors, or activities (1). This condition is estimated to affect 1 in 54 children (2), with males being 4 times more likely to be identified with ASD (2), and represents an economic burden of $11.5 billion—$60.9 billion in the United States (3). There is likely no single cause of ASD. Rather, its diagnosis represents a core set of behavioral symptoms that unifies individuals with a heterogeneous collection of genetic and phenotypic differences.
Within this framework, it is critical to ascertain how the various genetic and environmental risk factors associated with ASD ultimately translate to the core symptoms of this condition and its associated comorbidities. This highlights the importance of using animals to model rare inherited variants and de novo mutations associated with ASD to determine their contribution to the clinical presentation of the disorder. These animal models foster the uncovering of the molecular, neurobiological, and environmental contributors to ASD.
As previously shown, features of ASD might stem from or be exacerbated by abnormal dopamine (DA) signaling (4–8). Therefore, the study of dopaminergic dysfunction as it relates to this disorder is highly relevant. Recent work from our laboratories and other investigators has identified single nucleotide polymorphisms of DAT in individuals with ASD and associated comorbidities (6–11). Among those is an ASD-associated de novo mutation in the SLC6A3 gene resulting in a threonine to methionine substitution at residue 356 (DAT T356M). This mutation impairs central DA signaling and DA-dependent behaviors, promoting repetitive behaviors and hyperlocomotion that reflect behavioral characteristics seen in ASD (6, 8).
In addition to its role in central neurotransmission, DA signaling is involved in regulating functions of the enteric nervous system (ENS), a system of neurons that spans the length of the digestive system and serves to regulate digestive function (12). Dopaminergic neurons are found in the ENS and are important for gut motility, insulin release, and metabolism (13–16). Altered DA signaling promoted by the T356M mutation might not only impact central nervous system (CNS) function, but also ENS function (17). Furthermore, catecholamines, including DA, have been shown to alter the growth of some gram negative microbial species (18), including E. coli and Y. enterocolitica. Thus, changes in DA turnover (as observed in the DAT T356M+/+ mouse) could underlie changes in gut health, microbiota, and metabolism.
Problems with the ENS are reported to co-occur with a number of CNS disorders linked to dysregulation of the DA system. Children with ASD show multiple gastrointestinal (GI) abnormalities (19, 20). Also, they are more likely to experience abdominal pain, constipation, and diarrhea than those without ASD (21). Specifically, 30–70% of individuals with ASD have a functional GI disorder (fGID) (22). The Rome Foundation defines fGIDs as disorders of gut-brain interaction (DGBI). fGIDs are classified by GI symptoms related to any combination of motility disturbances, visceral hypersensitivity, altered gut microbiota, mucosal composition, immune function, and/or central nervous system processing (23). Evidence has suggested that these GI symptoms may exacerbate the behavioral symptoms exhibited by children with ASD by promoting emotional distress (24). Notably, maladaptive behaviors directly correlate with GI issues in individuals with ASD (25). For example, in children with ASD, behavior scores for irritability, social withdrawal, stereotypy, and hyperactivity are significantly higher in children with frequent abdominal pain, gaseousness, diarrhea, and constipation (25).
Beyond alterations in GI function, children with ASD also have an altered oral microbiota. Specifically, they have an increase in Streptococcus levels concomitant with a significant decrease in Fusobacterium (26) relative to neurotypical controls. Fusobacterium nucleatum is an anaerobic filamentous gram-negative bacterial species from the Fusobacterium genus (27, 28). It is a bacterial species implicated in a variety of infections ranging from appendicitis to osteomyelitis, as well as acting as an oncogenic bacterium implicated in colon cancer (28, 29). It is one of the most common oral bacteria found associated with a wide variety of periodontal health conditions, including gingivitis and periodontitis (30). Interestingly, Fusobacterium abundance has been shown to be associated with patients that display obesity and insulin resistance (31) and preterm birth (32), demonstrating a potential role of Fusobacteria in host metabolism.
Evidence is emerging to suggest the DA system is implicated in ASD phenotypes. Furthermore, there is ample data supporting altered gastrointestinal function and both intestinal and oral microbiome composition in ASD. To further probe this, we have generated a mouse harboring a point mutation in the SLC6A3 gene identified from a proband with ASD that displays altered DA neurotransmission (8). In this study, we explore the degree to which oral microbiome composition, metabolism, and glucose tolerance are altered by DA dysfunction in a mouse model of ASD.
Materials and Methods
Generation of DAT T356M+/+ Mouse
Mice were generated by GenOway S.A. These mice were generated and used in a previous study by the lab (8). Briefly, the point mutation was inserted into the exon 8 of the mouse SLC6A3 gene and was expressed under the control of the endogenous SLC6A3 promoter. PCR and southern blot were used to validate the proper integration of the gene. All animals used in the study were derived from matings of DAT T356M+/− parents.
Body Composition
Bruker's minispec Body Composition Analyzer was used to determine the body composition of mice based on Time Domain NMR (TD-NMR). This equipment acquires and analyzes TD-NMR signals from all protons in the entire sample volume and provides a precise method for measurement of lean tissue, fat, and free body fluid in living mice. Body composition was analyzed in mice between 14 and 19 weeks of age. Nineteen male mice and 10 female mice were utilized in these experiments.
Caloric Expenditure
The Promethion from Sable Systems (Las Vegas, NV) was used to assess energy expenditure in 16–19 week-old mice. Ten male mice were used in these experiments. Mice were individually housed in Promethion System cages for 5 days, during which numerous parameters were continuously measured (including O2 consumption, CO2 production, food and water intake, weight, and activity). One week prior to the experiment start date, mice were singly housed for acclimation. The cages were housed in a light and temperature-controlled chamber. The light cycle was set on a 12:12 h cycle (6 am−6 pm). The temperature was maintained at 23°C for the duration of the test.
Oral Glucose Tolerance Test
Oral glucose tolerance testing was performed in mice aged 14–20 weeks of age. 30 mice (20 males and 10 females) were used for these experiments. Animals were fasted for 4 h prior to testing. Fasted blood glucose levels were determined before a solution of 20% dextrose was administered by oral gavage. Mice were given 2 g dextrose/kg body mass. Blood glucose levels were measured at the following time points following oral gavage: 10, 20, 30, 45, 60, 75, 90, and 120 min.
Fecal Measurements
Both male and female mice were used for these experiments. Seven mice were used in these experiments with 21 total fecal pellets measured. Mice were placed in individual transfer buckets for 5 min during the first hour of the light cycle. At the end of the 5-min period, the feces were collected. Each fecal sample was weighed using an analytical scale and measured along the longest axis using a digital caliper.
Oral Microbiome
Sterile swabs were used to collect samples from mice oral cavity at 15–18 weeks of age. Twelve total mice were used for these experiments. Swabs were snap frozen and stored at −80°C until DNA extraction was performed.
DNA samples were extracted from swab samples with ZR Fecal/Soil DNA Miniprep Kit (Zymo Research). PCR was performed with primers specific for the V4 region of the 16S rRNA gene for amplification (33, 34). Sequences were performed on an Illumina Miseq as described previously (35, 36). Sequences were analyzed using the Quantitative Insight into Microbial Ecology (QIIME) suite v1.7 (33) and a QIIME wrapper called QWRAP (35). Operational Taxonomic Units (OTUs) were clustered at 97% sequence similarity. Taxonomic groups were assigned by using the Ribosomal Database Project (RDP) classifier (37) as well as the May 2013 Greengenes 16S rRNA sequence database (38). Samples were tested to determine whether samples clustered differently between the two groups of mice by the permutational multivariate analysis of variance (PERMANOVA) test for significant differences in clustering (p < 0.05). OTUs were grouped by phyla, classes, orders, families, genera and species.
Statistics
All statistical analyses were performed using GraphPad Prism software (version 8.3.0). Data were tested for normality and homogeneity of variance when necessary. Statistical methods are indicated in the figure legends and the results section. All t-tests were run as two-tailed. Data are presented as mean ± SEM. Differences are considered statistically significant at p < 0.05.
Ethics Approval and Consent to Participate
All behavioral and metabolic experiments were performed under a protocol approved by the Vanderbilt University Animal Care and Use Committee.
Results
The DAT T356M+/+ Mouse Has Increased Energy Expenditure
We have previously shown that the DAT T356M+/+ mouse has impaired central DA neurotransmission (8). Since DA is implicated in regulating metabolism, most notably in insulin release (14–16), and ENS function (13), we first measured body weight and body composition of DAT T356M+/+ mice and their wild type (WT) littermates. Body weight was significantly reduced in male DAT T356M+/+ mice compared to male WT mice at 14–19 weeks of age (Figure 1A. WT = 26.48 ± 0.6405 g, DAT T356M+/+ = 24.14 ± 0.4083 g; n = 8 WT, 11 DAT T356M+/+; t = 3.226, df = 17, p = 0.0050 by Student's t-test). However, there was no difference in body weight between female DAT T356M+/+ and WT mice (WT = 22.78 ± 0.8938 g, DAT T356M+/+ = 22.18 ± 0.7658 g; n = 4 WT, 6 DAT T356M+/+; t = 0.5015, df = 8, p = 0.6296 by Student's t-test), suggesting that this effect is sexually dimorphic.
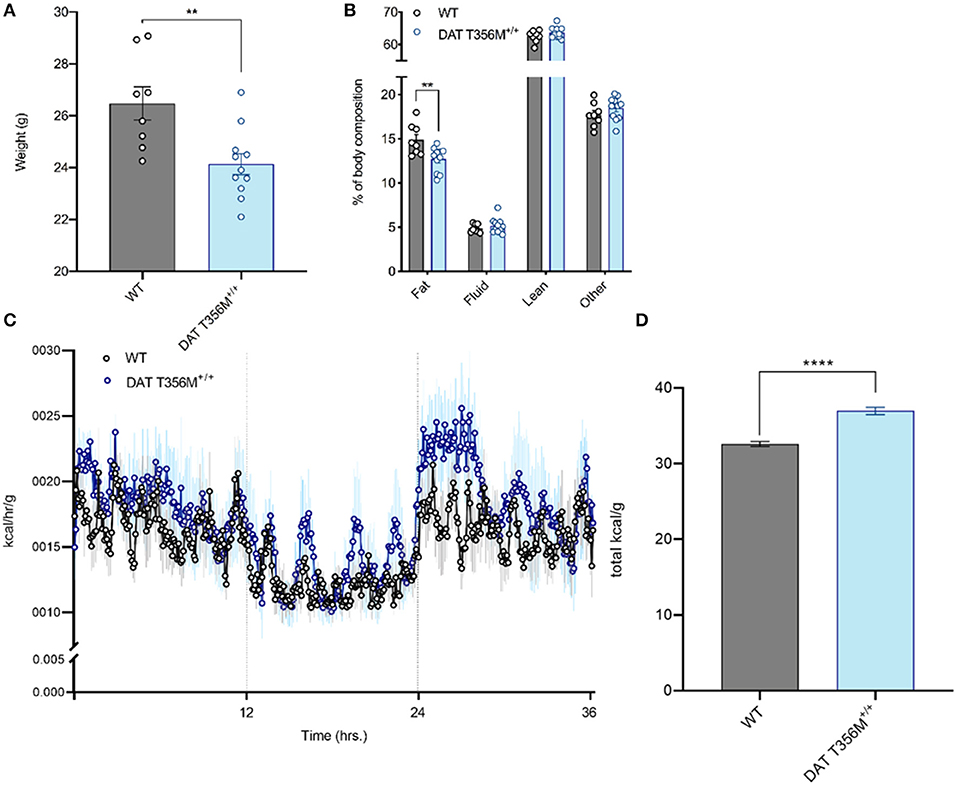
Figure 1. Male DAT T356M+/+ mice have reduced body weight, reduced percent body fat, and increased total caloric expenditure. (A) Total body weight is reduced in male DAT T356M+/+ mice (WT = 26.48 ± 0.6405 g, DAT T356M+/+ = 24.14 ± 0.4083 g; n = 8 WT, 11 DAT T356M+/+; t = 3.226, df = 17, ** = p = 0.0050 by Student's t-test). (B) Percent body fat is reduced in male DAT T356M+/+ mice (WT = 14.89 ± 0.6206%, DAT T356M+/+ = 12.72 ± 0.4160%; n = 8 WT, 11 DAT T356M+/+; F(3, 68) = 5.373, ** = p = 0.0047 by two-way ANOVA followed by Sidak's multiple comparisons test). (C) Caloric expenditure per gram of body weight binned in 5-min intervals over the course of 36 h. Vertical lines at 12 and 24 h represent the start and end of the light cycle, respectively. The blue circles represent the mean value for DAT T356M+/+ mice (n = 5) and the black circles represents the mean value for WT mice (n = 5). The light gray and light blue lines represent the SEM. (D) Total caloric expenditure is increased in male DAT T356M+/+ mice as measured by indirect calorimetry (WT = 32.60 ± 0.3298 kcal/g, DAT T356M+/+ = 36.97 ± 0.4910 kcal/g, n = 5 WT, 5 DAT T356M+/+, t = 7.388, df = 8, **** = p < 0.0001 by Student's t-test).
To better understand how the T356M mutation influences mouse body weight, we measured body composition of male and female DAT T356M+/+ mice and WT mice. In males, we found significantly reduced percent body fat in DAT T356M+/+ mice compared with WT mice (Figure 1B. WT = 14.89 ± 0.6206%, DAT T356M+/+ = 12.72 ± 0.4160%; n = 8 WT, 11 DAT T356M+/+; F(3, 68) = 5.373, p = 0.0047 by two-way ANOVA followed by Sidak's multiple comparisons test). No other components of the body composition (i.e., lean mass, fluid, or other) were different between male DAT T356M+/+ mice and WT mice (Figure 1B). In contrast, no differences in body composition were observed between female DAT T356M+/+ mice and WT mice (Body fat: WT = 13.23 ± 0.5502%, DAT T356M+/+ = 13.17 ± 0.2128%, Fluid: WT = 5.987 ± 0.1256%, DAT T356M+/+ = 5.752 ± 0.3474%, Lean: WT = 64.17 ± 0.776%, DAT T356M+/+ = 63.15 ± 0.3714%; n = 4 WT, 6 DAT T356M+/+, F(1, 24) = 1.631 and p = 0.2137 for genotype, F(2, 24) = 0.7398 and p = 0.4878 for interaction between genotype and tissue type by two-way ANOVA). In male DAT T356M+/+ mice, this decrease in body weight is driven, at least in part, by a decrease in percent body fat and not lean mass (Figure 1B). Exercise can regulate body composition (i.e., decreased body fat and increased lean mass) (39).
In 7 week old heterozygous animals (DAT T356M+/−) we did not observe differences in body weight with respect to WT. To note, is that these animals were younger than the animals used in other experiments in this study. This cohort was made of entirely male mice and there was no significant difference in weight between WT and DAT T356M+/− mice (WT = 21.42 ± 3.57 g, DAT T356M+/− = 20.85 ± 2.75g, n = 13 WT, 11 DAT T356M+/−, t = 0.776, df = 22, p = 0.446 by Students t-test).
We used a metabolic cage (Promethion, Sable Systems, Las Vegas, USA) to measure energy expenditure continuously over 36 h both in male DAT T356M+/+ mice and WT mice (Figure 1C). Total energy expenditure over 36 h was significantly increased in male DAT T356M+/+ mice (Figure 1D. WT = 32.60 ± 0.3298 kcal/g, DAT T356M+/+ = 36.97 ± 0.4910 kcal/g, n = 5 WT, 5 DAT T356M+/+, t = 7.388, df = 8, p < 0.0001 by Student's t-test). As the DAT T356M+/+ mice are hyperactive (8), one would expect lean mass to be increased. However, since we saw no change in lean mass our data indicate that this decrease in body mass is not likely promoted by hyperactivity, but rather by metabolic changes.
DAT T356M+/+ Mice Have Improved Glucose Tolerance
It is well-established that DA also plays an important role in glucose and insulin regulation (16, 40, 41). Insulin-secreting pancreatic β-cells express the enzymes required for DA synthesis and catabolism, as well as all five DA receptors (40, 42). In these cells, DA functions as a negative regulator of glucose-stimulated insulin secretion (GSIS) (41). The D2 receptor (D2R) and D3 receptor (D3R) signaling act in concert to inhibit GSIS (14–16). Consistent with these findings, β-cell-selective D2R knockout mice exhibit marked postprandial hyperinsulinemia (16).
DAT T356M+/+ mice have impaired D2 receptor signaling (8). Thus, we sought to determine whether glucose homeostasis was also affected in these animals. After glucose loading, there was a rapid decrease in blood glucose in DAT T356M+/+ mice, while this rapid decrease in blood glucose is not present in WT mice (Figure 2A). Maximum blood glucose levels were lower in DAT T356M+/+ mice than WTs and returned to baseline faster than WTs, which still had slightly elevated levels even after 120 min. These results highlight that there is significantly improved glucose handling in the DAT T356M+/+ mice (Figure 2B. WT = 129.4 ± 20.80 mg/dL, DAT T356M+/+ = 81.94 ± 11.78 mg/dL; n = 13 WT, 17 DAT T356M+/+; t = 2.100, df = 28, p = 0.0448 by Student's t-test).
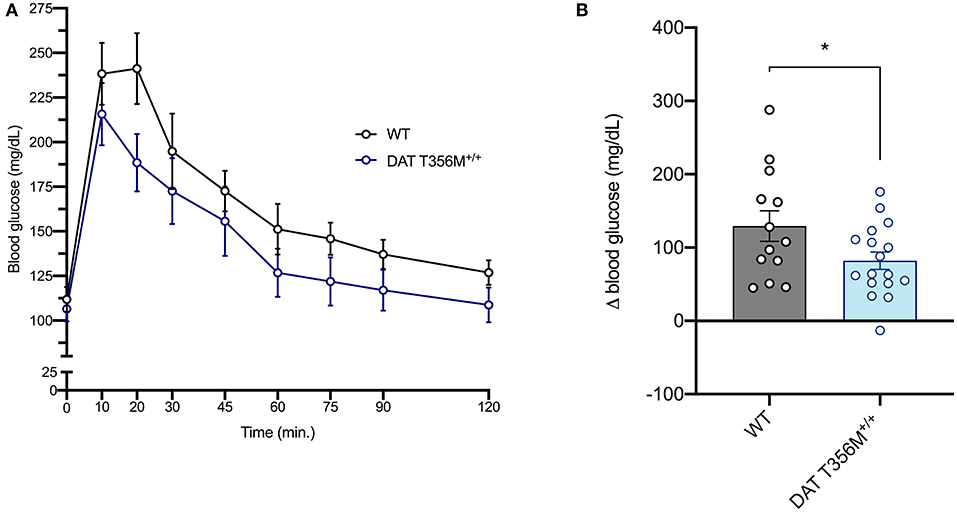
Figure 2. DAT T356M+/+ mice have improved glucose handling after glucose challenge. (A) Blood glucose levels during a glucose tolerance test in WT and DAT T356M+/+ mice. (B) Change in blood glucose from baseline to 20 min (Δ blood glucose) is significantly reduced in DAT T356M+/+ mice compared to WT mice (WT = 129.4 ± 20.80 mg/dL, DAT T356M+/+ = 81.94 ± 11.78 mg/dL; n = 13 WT, 17 DAT T356M+/+; t = 2.100, df = 28, * = p = 0.0448 by Student's t-test).
The DAT T356M+/+ Mouse Displays Reduced Abundance of Oral Fusobacterium
A number of changes in gut and oral microbial populations have been reported in individuals with ASD (26, 43). Strong evidence points to a decrease in the abundance of oral Fusobacterium in persons with ASD (26). Changes in these bacterial populations have also been associated with alterations in various metabolic processes. For example, increased Fusobacterium in the gut correlates with insulin resistance (31). Thus, we sought to determine the composition of the oral microbiota and, specifically, the abundance of Fusobacterium in DAT T356M+/+ mice compared to WT mice. In Figure 3A, we show the top 10 operational taxonomic units (OTUs) isolated from oral swabs from DAT T356M+/+ and WT mice. We found that Fusobacterium was significantly decreased in the DAT T356M+/+ mice (Figure 3B. WT = 0.003716 ± 0.001136 relative units, DAT T356M+/+ = 0.001 ± 0.0003 relative units; n = 5 WT, 7 DAT T356M+/+; t = 2.589, df = 10, p = 0.027 by Student's t-test). However, the decrease in the abundance seen in Fusobacterium is not observed for other bacteria, such as Pseudomonas, the most abundant OTU isolated from our samples (Figures 3A,E. WT = 0.2438 ± 0.05866 relative units, DAT T356M+/+ = 0.329 ± 0.0491 relative units; n = 6 WT, 8 DAT T356M+/+; t = 1.113, df = 12, p = 0.2873 by Student's t-test).
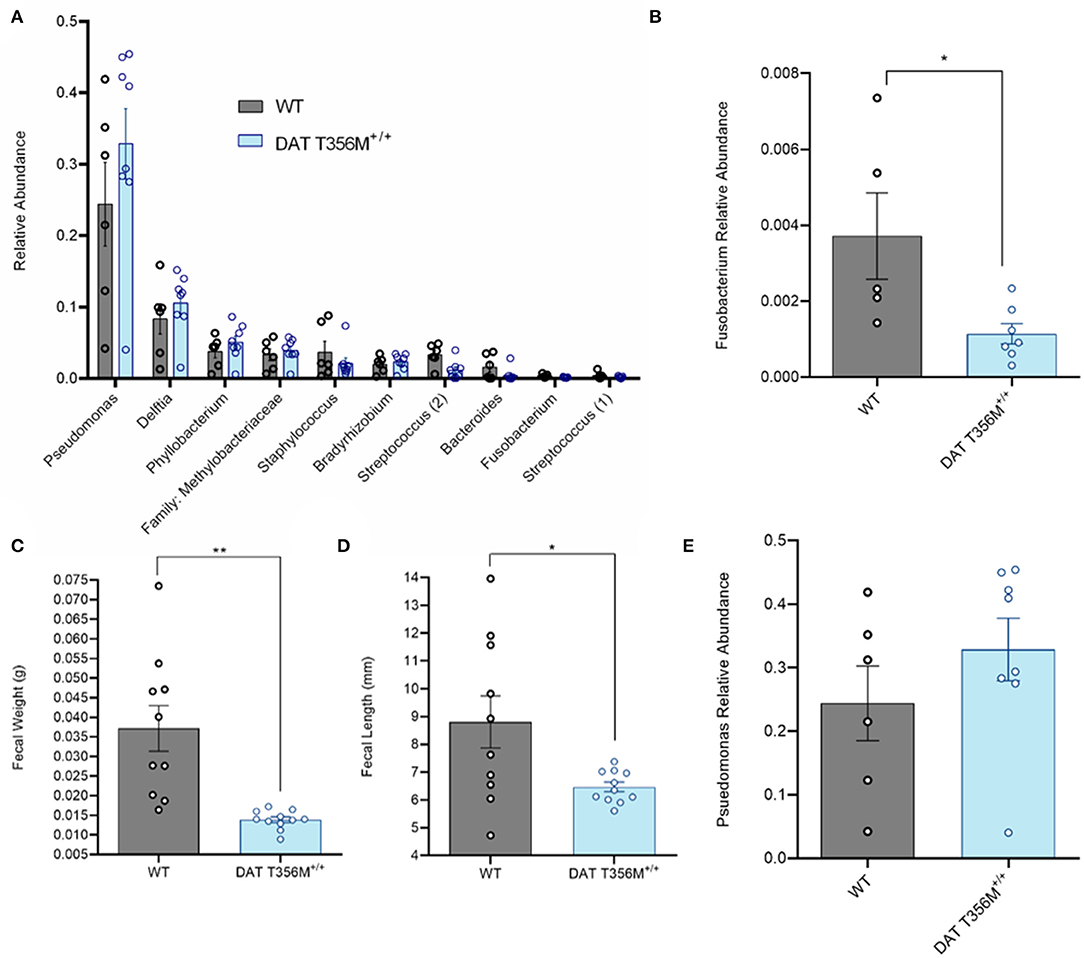
Figure 3. DAT T356M+/+ mice have altered oral microbiota. (A) Relative abundance of top 10 OTUs isolated from oral swabs from WT and DAT T356M+/+ mice. (B) DAT T356M+/+ mice have significantly lower abundance of oral Fusobacterium compared to WT mice (WT = 0.003716 ± 0.001136 relative units, DAT T356M+/+ = 0.001 ± 0.0003 relative units; n = 5 WT, 7 DAT T356M+/+; t = 2.589, df = 10, * = p = 0.027 by Student's t-test). (C) DAT T356M+/+ mice have significantly decreased fecal weight compared to WT mice (WT = 0.0371 ± 0.0058 g, DAT T356M+/+ = 0.0138 ± 0.0139 g; n = 10 fecal pellets from 3 WT mice, 11 fecal pellets from 4 DAT T356M+/+ mice; t = 3.981, df = 9, ** = p = 0.0032 by Welch's t-test). (D) DAT T356M+/+ mice have significantly decreased fecal length compared to WT mice (WT = 8.805 ± 0.9381 mm, DAT T356M+/+ = 6.469 ± 0.1727 mm; n = 10 feces from 3 WT mice, 11 feces from 4 DAT T356M+/+ mice; t = 2.449, df = 9, * = p = 0.0368 by Welch's t-test). (E) There is no difference in the relative abundance of Pseudomonas between WT and DAT T356M+/+ mice (WT = 0.2438 ± 0.05866 relative units, DAT T356M+/+ = 0.3286 ± 0.0491 relative units; n = 6 WT, 8 DAT T356M+/+; t = 1.113, df = 12, p = 0.2873 by Students t-test).
To evaluate the impact of the DAT T356M on GI function, we analyzed fecal weight and length. In a mouse model of constipation, there is a decrease in both fecal weight and length (44), demonstrating that changes in these parameters can be used as a partial readout of GI function. DAT T356M+/+ mice displayed reduced fecal weight (Figure 3C. WT = 0.037 ± 0.0058 g, DAT T356M+/+ = 0.01388 ± 0.01388 g; n = 10 from 3 WT, t = 3.981, df = 9, 11 from 4 DAT T356M+/+; p = 0.0032 by Welch's t-test) and length (Figure 3D. WT = 8.805 ± 0.9381 mm, DAT T356M+/+ = 6.469 ± 0.1727 mm; n = 10 from 3 WT, 11 from 4 DAT T356M+/+; t = 2.449, df = 9, p = 0.0368 by Welch's t-test), suggesting constipation in DAT T356M+/+ mice (44).
Relative Abundance of Oral Fusobacterium Is Positively Correlated With Percent Body Fat and Peak Blood Glucose
In humans, an increased abundance of Fusobacterium correlates with insulin resistance (31). Given the improved glucose handling in the DAT T356M+/+ mice, we sought to determine the relationship between relative abundance of oral Fusobacterium and metabolic parameters in our sample. We observed a positive association between the relative abundance of Fusobacterium in the oral cavity and peak blood glucose during a glucose challenge (Figure 4A, Pearson's r = 0.5923; n = 4 WT, 7 DAT T356M+/+; p = 0.058). Although this correlation did not reach statistical significance, it was notably strong in magnitude. We similarly observed a strong, positive and significant association between the relative abundance of Fusobacterium in the oral cavity and percent body fat (Figure 4B, Pearson's r = 0.6671; n = 4 WT, 7 DAT T356M+/+; p = 0.0249). To determine aforementioned relations were limited to Fusobacterium, we additionally analyzed the associations for relative abundance of Pseudomonas with both percent body fat and peak blood glucose. We found no significant correlation between relative abundance of Pseudomonas and either percent body fat (Pearson's r = 0.1208; n = 6 WT, 8 DAT T356M+/+; p = 0.2233) or peak blood glucose (Pearson's r = 9.7 × 10−6; n = 6 WT, 8 DAT T356M+/+; p = 0.99). These correlations were negligible to small in magnitude.
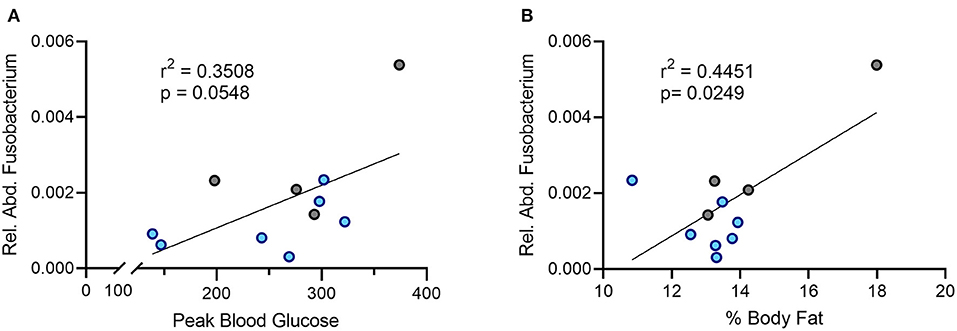
Figure 4. There is a positive correlation between the relative abundance of Fusobacterium and both peak blood glucose and percent body fat. (A) Peak blood glucose is positively correlated with relative abundance of oral Fusobacterium (Pearson's r = 0.5923; n = 4 WT, 7 DAT T356M+/+; p = 0.058). (B) The relative abundance of Fusobacterium in the oral cavity and peak blood glucose are positively and significantly correlated (Pearson's r = 0.6671; n = 4 WT, 7 DAT T356M+/+; p = 0.0249). Blue dots represent T356M+/+ mice and gray dots represent WT mice.
Discussion
The mechanistic understanding of proteins, such as DAT, that regulate DA neurotransmission is vital to pharmacologically target DA dysfunction in autism and other conditions. DAT variants, dysregulation of DA, and altered developmental trajectory of dopaminergic structures have been associated with ASD (6, 9–11, 45, 46). Beyond the importance of proper DA neurotransmission for central function, increasing evidence highlights the role of DA signaling in gut motility, insulin release, and metabolism. In the pancreas, DA is stored in secretory granules in pancreatic beta-islet cells and regulates GSIS (14, 15). Moreover, in the intestines, dopaminergic neurons are found in the ENS and are important for proper intestinal motility (13). As such, it is not surprising that dysfunction in the DA system can affect both metabolism and GI function. Thus, studying DAT variants associated with ASD will facilitate understanding of their role in central and peripheral neurotransmitter homeostasis and will help to determine how transporter dysfunction contributes to GI and metabolic disorders.
Here we explore the impact of an ASD-associated de novo mutation of the DAT (6, 8), which impacts DA neurotransmission, on metabolism, glucose handling, and the oral microbiome. We demonstrate that DAT T356M+/+ mice exhibit altered body composition, energy expenditure, glucose handling, and composition of oral microbiota. This study links altered dopaminergic signaling due to DAT dysfunction caused by an ASD-associated genetic variant with pathophysiological changes that recapitulate aspects of the human disorder in a murine model.
We first explored the impact of the DAT T356M mutation on energy homeostasis. We found that male DAT T356M+/+ mice have reduced body weight and that this reduction in body weight is characterized predominantly by reduced body fat, without a significant change in fluid, lean, or other body tissues. Interestingly, no difference in body composition were observed in female mice. Males have been shown to have sex specific deficits in other animal models of autism (47–49). Thus, a more pronounced phenotype in our male mice is in line with previous data. Since no change in lean mass was observed, it is unlikely that this change in body fat is driven by hyperactivity. Rather, this reduction in body fat is more likely associated with increased total energy expenditure.
As DA signaling regulates insulin release (16), we additionally asked whether the observed reduction in body fat was associated with altered glucose handling. We found that DAT T356M+/+ mice display significant differences in glucose handling (specifically, a significantly lower change in blood glucose 20 min after glucose challenge in male mice). Furthermore, in pancreatic beta-islet cells, the D2 autoreceptor negatively regulates the release of insulin (14, 16). Considering that DAT T356M+/+ mice have reduced D2 receptor signaling (8), we therefore suggest that this improvement in glucose handling may be due to increased insulin release. Furthermore, it is important to note that, when compared to skeletal muscle, adipose tissue is not generally thought as being a major user of enteral (i.e., gastrointestinal) glucose. Therefore, we suggest that this altered glucose handling in the DAT T356M+/+ mice is likely not due to their lowered fat composition, but rather changes in the sensitivity of the D2 receptor. Future studies should directly assess the insulin levels seen in these mice, as well as the sensitivity and expression of D2 receptor in beta-islet cells in the pancreas to determine the underlying physiology of the improved glucose handling seen in our animal model.
In addition to insulin release, DA plays a critical role in GI function (13, 17). Children with ASD display multiple GI abnormalities (19, 20) with 30–70% of individuals with ASD having a fGID (22), a group of disorders classified by motility disturbances, altered mucosal function, and altered gut microbiota (23). As a read out for proper GI function, we measured both the length and weight of feces in DAT T356M+/+ mice and discovered that their feces are lighter and shorter than WT mice. These alterations have been previously observed in mouse models of constipation (44). Notably, children with ASD are more likely to experience abdominal pain and constipation when compared to those without ASD (21).
Many studies have reported differences in the composition of the oral and gut microbiota in patients with ASD (26, 50) and it is possible that these differences in the flora may contribute to both behavioral and GI symptoms associated with autism (21, 24). As communication between the brain and the gut is bidirectional, it is conceivable that dysfunction of key regulators of CNS function associated with ASD may also drive dysfunction of the ENS and thus changes in the composition of the oral and GI microbiota (51, 52). Here, we point to the DAT as one such key regulator. We found that DAT T356M+/+ mice have reduced abundance of Fusobacterium in the oral cavity and no difference in Pseudomonas, the most abundant bacteria in the oral cavity of our animals, as compared to WT mice. It is possible that this change in oral microbiome is due directly to the anomalous DA release seen in this mutant (8), as there is evidence that DA can alter the growth of certain gram-negative bacterial species (18). However, we cannot rule out the possibility that this change is due to dietary differences or differences in metabolism leading to an altered microbiome. Consistent with our data, increased Fusobacterium abundance has been associated with insulin resistance and impaired glucose handling in obese patients (31). Of note, we found a positive association between the relative abundance of Fusobacterium and both peak blood glucose and percent body fat in our mouse model. However, there was no relation between the abundance of Pseudomonas and either peak blood glucose or percent body fat.
Our data demonstrate that there is less Fusobacterium in the oral microbiome in the DAT T356M+/+ model of ASD, a finding that is in agreement with human data from Qiao and colleagues (26). Fusobacterium nucleatum (F. nucleatum) is one of the most common species in periodontal diseases, such as gingivitis (30). These findings, collectively, point toward a potential explanation for the fact that, despite having no difference in quality of teeth brushing (53), children with ASD have been observed to display better oral health and less caries when compared to a control population (53, 54). Given that both humans and our murine model display significantly less oral Fusobacterium, this could potentially be a driving factor in the superior oral hygiene seen in children with ASD, despite no differences in teeth brushing habits (53). Consistent with our hypothesis, a recent study from Schoilew and colleagues demonstrated that patients without a history of caries have significantly less Fusobacterium than patients that previously had caries (55). Alternatively the reduction in F. nucleatum level may mediate some of the phenotypes seen in ASD as F. nucleatum is known to produce butyrate and this is a short chain fatty acid that has been shown to be present at decreased levels in individuals with ASD (56). It is speculated that the alterations in the microbiome and the decrease in butyrate is one of the drivers of the constipation and other GI issues observed in ASD (56). Clearly, this observation requires more research on both human patients and alternative animal models of ASD to corroborate differences presented in other studies and explore underlying mechanisms (57).
Here, we present new evidence of how DAT dysfunction may translate to altered GI function, composition of the microbiota of the oral cavity, and metabolism. This model suggests that impaired DA signaling in ASD drives a number of pathophysiological changes that could explain, in part, at least some of the observed metabolic and GI phenotypes in ASD. This work also suggests that DA signaling, or specific microbial populations in the microbiome, may represent a tractable target for the treatment of GI and metabolic disturbances in persons effected by ASD, and potentially in individuals with other conditions associated with DA dysfunction.
Data Availability Statement
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found in the article/Supplementary Material.
Ethics Statement
The animal study was reviewed and approved by Vanderbilt University Animal Care and Use Committee.
Author Contributions
GD, SM, XC, and CM designed and performed experiment and contributed to the writing of the manuscript. TW, FH, IR, HM, CF, MW, HW, and AG designed the experiments and contributed to writing the manuscript. All authors contributed to the article and approved the submitted version.
Funding
This research is funded by RO1—DA035263 08 (AG and HM) and NIH 5T32NS061788-12 (SM). This research is funded by RO1—DA035263 08 (AG and HM), NIH 5T32NS061788-12 (SM), and 5F30MH115535 (GD).
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
The authors would like to acknowledge Saunders Consulting for the help in editing this manuscript. This work was supported by the Vanderbilt Kennedy Center (P50HD103537; Neul).
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fpsyt.2021.655451/full#supplementary-material
Abbreviations
DA, dopamine; DAT, dopamine transporter; WT, wild type; ASD, autism spectrum disorder; CNS, central nervous system; ENS, enteric nervous system; GI, gastrointestinal; fGID, functional GI disorder; DGBI, disorder of gut brain interaction.
References
1. American Psychiatric Association. Diagnostic and Statistical Manual of Mental Disorders. 5th ed. American Psychiatric Association (2013). doi: 10.1176/appi.books.9780890425596
2. Baio J, Wiggins L, Christensen DL, Maenner MJ, Daniels J, Warren Z, et al. Prevalence of Autism Spectrum Disorder Among Children Aged 8 Years - Autism and Developmental Disabilities Monitoring Network, 11 Sites, United States, 2014. MMWR Surveill Summ. (2018) 67:1–23. doi: 10.15585/mmwr.ss6706a1
3. Lavelle TA, Weinstein MC, Newhouse JP, Munir K, Kuhlthau KA, Prosser LA. Economic burden of childhood autism spectrum disorders. Pediatrics. (2014) 133:e520–e529. doi: 10.1542/peds.2013-0763
4. Rodriguiz RM, Chu R, Caron MG, Wetsel WC. Aberrant responses in social interaction of dopamine transporter knockout mice. Behav Brain Res. (2004) 148:185–98. doi: 10.1016/S0166-4328(03)00187-6
5. Langen M, Durston S, Kas MJ, van Engeland H, Staal WG. The neurobiology of repetitive behavior: …and men. Neurosci Biobehav Rev. (2011) 35:356–65. doi: 10.1016/j.neubiorev.2010.02.005
6. Hamilton PJ, Campbell NG, Sharma S, Erreger K, Herborg Hansen F, Saunders C, et al. De novo mutation in the dopamine transporter gene associates dopamine dysfunction with autism spectrum disorder. Mol Psychiatry. (2013) 18:1315–23. doi: 10.1038/mp.2013.102
7. Campbell NG, Shekar A, Aguilar JI, Peng D, Navratna V, Yang D, et al. Structural, functional, and behavioral insights of dopamine dysfunction revealed by a deletion in SLC6A3. Proc Natl Acad Sci USA. (2019) 116:3853–62. doi: 10.1073/pnas.1816247116
8. DiCarlo GE, Aguilar JI, Matthies HJ, Harrison FE, Bundschuh KE, West A, et al. Autism-linked dopamine transporter mutation alters striatal dopamine neurotransmission and dopamine-dependent behaviors. J Clin Invest. (2019) 130:3407–19. doi: 10.1172/JCI127411
9. Cook EH Jr, Stein MA, Krasowski MD, Cox NJ, Olkon DM, Kieffer JE, et al. Association of attention-deficit disorder and the dopamine transporter gene. Am J Hum Genet. (1995) 56:993–8.
10. Gadow KD, Roohi J, DeVincent CJ, Hatchwell E. Association of ADHD, tics, and anxiety with dopamine transporter (DAT1) genotype in autism spectrum disorder. J Child Psychol Psychiatry. (2008) 49:1331–8. doi: 10.1111/j.1469-7610.2008.01952.x
11. Bowton E, Saunders C, Reddy IA, Campbell NG, Hamilton PJ, Henry LK, et al. SLC6A3 coding variant Ala559Val found in two autism probands alters dopamine transporter function and trafficking. Transl Psychiatry. (2014) 4:e464. doi: 10.1038/tp.2014.90
12. Grundy R, Spiller D. Pathophysiology of the Enteric Nervous System: A Basis for Understanding Functional Diseases. Chichester: John Wiley and Sons (2004).
13. Li ZS, Schmauss C, Cuenca A, Ratcliffe E, Gershon MD. Physiological modulation of intestinal motility by enteric dopaminergic neurons and the D2 receptor: analysis of dopamine receptor expression, location, development, and function in wild-type and knock-out mice. J Neurosci. (2006) 26:2798–807. doi: 10.1523/JNEUROSCI.4720-05.2006
14. Rubí B, Ljubicic S, Pournourmohammadi S, Carobbio S, Armanet M, Bartley C, et al. Dopamine D2-like receptors are expressed in pancreatic beta cells and mediate inhibition of insulin secretion. J Biol Chem. (2005) 280:36824–32. doi: 10.1074/jbc.M505560200
15. Ustione A, Piston DW. Dopamine synthesis and D3 receptor activation in pancreatic β-cells regulates insulin secretion and intracellular [Ca(2+)] oscillations. Mol Endocrinol. (2012) 26:1928–40. doi: 10.1210/me.2012-1226
16. Farino ZJ, Morgenstern TJ, Maffei A, Quick M, De Solis AJ, Wiriyasermkul P, et al. New roles for dopamine D(2) and D(3) receptors in pancreatic beta cell insulin secretion. Mol Psychiatry. (2019) 25:2070–85. doi: 10.1038/s41380-018-0344-6
17. Rao M, Gershon MD. The bowel and beyond: the enteric nervous system in neurological disorders. Nat Rev Gastroenterol Hepatol. (2016) 13:517–28. doi: 10.1038/nrgastro.2016.107
18. Lyte M, Ernst S. Catecholamine induced growth of gram negative bacteria. Life Sci. (1992) 50:203–12. doi: 10.1016/0024-3205(92)90273-R
19. Horvath K, Papadimitriou JC, Rabsztyn A, Drachenberg C, Tildon JT. Gastrointestinal abnormalities in children with autistic disorder. J Pediatr. (1999) 135:559–63. doi: 10.1016/S0022-3476(99)70052-1
20. Lefter R, Ciobica A, Timofte D, Stanciu C, Trifan A. A descriptive review on the prevalence of gastrointestinal disturbances and their multiple associations in autism spectrum disorder. Medicina. (2019) 56:11. doi: 10.3390/medicina56010011
21. McElhanon BO, McCracken C, Karpen S, Sharp WG. Gastrointestinal symptoms in autism spectrum disorder: a meta-analysis. Pediatrics. (2014) 133:872–83. doi: 10.1542/peds.2013-3995
22. Penzol MJG, Salazar de Pablo Llorente C, Moreno C, Hernández P, Dorado ML, Parellada M. Functional gastrointestinal disease in autism spectrum disorder: a retrospective descriptive study in a clinical sample. Front Psychiatry. (2019) 10:179. doi: 10.3389/fpsyt.2019.00179
23. Schmulson MJ, Drossman DA. What is new in Rome IV. J Neurogastroenterol Motil. (2017) 23:151–63. doi: 10.5056/jnm16214
24. Thulasi V, Steer RA, Monteiro IM, Ming X. Overall severities of gastrointestinal symptoms in pediatric outpatients with and without autism spectrum disorder. Autism. (2019) 23:524–30. doi: 10.1177/1362361318757564
25. Chaidez V, Hansen RL, Hertz-Picciotto I. Gastrointestinal problems in children with autism, developmental delays or typical development. J Autism Dev Disord. (2014) 44:1117–27. doi: 10.1007/s10803-013-1973-x
26. Qiao Y, Wu M, Feng Y, Zhou Z, Chen L, Chen F. Alterations of oral microbiota distinguish children with autism spectrum disorders from healthy controls. Sci Rep. (2018) 8:1597. doi: 10.1038/s41598-018-19982-y
27. Bolstad AI, Jensen HB, Bakken V. Taxonomy, biology, and periodontal aspects of Fusobacterium nucleatum. Clin Microbiol Rev. (1996) 9:55–71. doi: 10.1128/CMR.9.1.55
28. Brennan CA, Garrett WS. Fusobacterium nucleatum - symbiont, opportunist and oncobacterium. Nat Rev Microbiol. (2019) 17:156–66. doi: 10.1038/s41579-018-0129-6
29. Bullman S, Pedamallu CS, Sicinska E, Clancy TE, Zhang X, Cai D, et al. Analysis of Fusobacterium persistence and antibiotic response in colorectal cancer. Science. (2017) 358:1443–8. doi: 10.1126/science.aal5240
30. Moore WE, Moore LV. The bacteria of periodontal diseases. Periodontol 2000. (1994) 5:66–77. doi: 10.1111/j.1600-0757.1994.tb00019.x
31. Moreno-Indias I, Sánchez-Alcoholado L, García-Fuentes E, Cardona F, Queipo-Ortuño MI, Tinahones FJ. Insulin resistance is associated with specific gut microbiota in appendix samples from morbidly obese patients. Am J Transl Res. (2016) 8:5672–84.
32. Garcia-So J, Zhang X, Yang X, Rubinstein MR, Mao Y, Kitajewski J, et al. Omega-3 fatty acids suppress Fusobacterium nucleatum-induced placental inflammation originating from maternal endothelial cells. JCI Insight. 4:e125436. doi: 10.1172/jci.insight.125436
33. Caporaso JG, Lauber CL, Walters WA, Berg-Lyons D, Lozupone CA, Turnbaugh PJ, et al. Global patterns of 16S rRNA diversity at a depth of millions of sequences per sample. Proc Natl Acad Sci USA. (2011) 108(Suppl 1):4516–22. doi: 10.1073/pnas.1000080107
34. Kozich JJ, Westcott SL, Baxter NT, Highlander SK, Schloss PD. Development of a dual-index sequencing strategy and curation pipeline for analyzing amplicon sequence data on the MiSeq Illumina sequencing platform. Appl Environ Microbiol. (2013) 79:5112–20. doi: 10.1128/AEM.01043-13
35. Kumar R, Eipers P, Little RB, Crowley M, Crossman DK, Lefkowitz EJ, et al. Getting started with microbiome analysis: sample acquisition to bioinformatics. Curr Protoc Hum Genet. (2014) 82:18.8.1–29. doi: 10.1002/0471142905.hg1808s82
36. Daft JG, Ptacek T, Kumar R, Morrow C, Lorenz RG. Cross-fostering immediately after birth induces a permanent microbiota shift that is shaped by the nursing mother. Microbiome. (2015) 3:17. doi: 10.1186/s40168-015-0080-y
37. Wang Q, Garrity GM, Tiedje JM, Cole JR. Naive Bayesian classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy. Appl Environ Microbiol. (2007) 73:5261–7. doi: 10.1128/AEM.00062-07
38. DeSantis TZ, Hugenholtz P, Larsen N, Rojas M, Brodie EL, Keller K, et al. Greengenes, a chimera-checked 16S rRNA gene database and workbench compatible with ARB. Appl Environ Microbiol. (2006) 72:5069–72. doi: 10.1128/AEM.03006-05
39. Kelly Scott A, Derrick Nehrenberg L, Hua K, Garland T Jr, Pomp D. Exercise, weight loss, and changes in body composition in mice: phenotypic relationships and genetic architecture. Physiol Genomics. (2011) 43:199–212. doi: 10.1152/physiolgenomics.00217.2010
40. Bucolo C, Leggio GM, Drago F, Salomone S. Dopamine outside the brain: The eye, cardiovascular system and endocrine pancreas. Pharmacol Ther. (2019) 203:107392. doi: 10.1016/j.pharmthera.2019.07.003
41. Garcia Barrado MJ, Iglesias Osma MC, Blanco MJ, Hernández MC, Robledo VS, Iniesta LC. Dopamine modulates insulin release and is involved in the survival of rat pancreatic beta cells. PLoS ONE. (2015) 10:e0123197–e97. doi: 10.1371/journal.pone.0123197
42. Chen Y, Hong F, Chen H, Fan RF, Zhang XL, Zhang Y, et al. Distinctive expression and cellular distribution of dopamine receptors in the pancreatic islets of rats. Cell Tissue Res. (2014) 357:597–606. doi: 10.1007/s00441-014-1894-9
43. Mulle JG, Sharp WG, Cubells JF. The gut microbiome: a new frontier in autism research. Curr Psychiatry Rep. (2013) 15:337. doi: 10.1007/s11920-012-0337-0
44. Liang C, Wang KY, Yu Z, Xu B. Development of a novel mouse constipation model. World J Gastroenterol. (2016) 22:2799–810. doi: 10.3748/wjg.v22.i9.2799
45. LaHoste GJ, Swanson JM, Wigal SB, Glabe C, Wigal T, King N, et al. Dopamine D4 receptor gene polymorphism is associated with attention deficit hyperactivity disorder. Mol Psychiatry. (1996) 1:121–4.
46. Langen M, Schnack HG, Nederveen H, Bos D, Lahuis BE, de Jonge MV, et al. Changes in the developmental trajectories of striatum in autism. Biol Psychiatry. (2009) 66:327–33. doi: 10.1016/j.biopsych.2009.03.017
47. Zamarbide M, Mossa A, Muñoz-Llancao P, Wilkinson MK, Pond HL, Oaks AW, et al. Male-Specific cAMP Signaling in the Hippocampus Controls Spatial Memory Deficits in a Mouse Model of Autism and Intellectual Disability. Biol Psychiatry. (2019) 85:760–8. doi: 10.1016/j.biopsych.2018.12.013
48. Roux S, Bailly Y, Bossu JL. Regional and sex-dependent alterations in Purkinje cell density in the valproate mouse model of autism. Neuroreport. (2019) 30:82–8. doi: 10.1097/WNR.0000000000001164
49. Kazlauskas N, Seiffe A, Campolongo M, Zappala C, Depino AM. Sex-specific effects of prenatal valproic acid exposure on sociability and neuroinflammation: Relevance for susceptibility and resilience in autism. Psychoneuroendocrinology. (2019) 110:104441. doi: 10.1016/j.psyneuen.2019.104441
50. Strati F, Cavalieri D, Albanese D, De Felice C, Donati C, Hayek J, et al. New evidences on the altered gut microbiota in autism spectrum disorders. Microbiome. (2017) 5:24. doi: 10.1186/s40168-017-0242-1
51. Collins SM, Surette M, Bercik P. The interplay between the intestinal microbiota and the brain. Nat Rev Microbiol. (2012) 10:735–42. doi: 10.1038/nrmicro2876
52. Sun Y, Zhang M, Chen CC, Gillilland M 3rd, Sun X, El-Zaatari M, Huffnagle GB, et al. Stress-induced corticotropin-releasing hormone-mediated NLRP6 inflammasome inhibition and transmissible enteritis in mice. Gastroenterology. (2013) 144:1478–87, 87.e1–8. doi: 10.1053/j.gastro.2013.02.038
53. Vajawat M, Deepika PC. Comparative evaluation of oral hygiene practices and oral health status in autistic and normal individuals. J Int Soc Prev Community Dent. (2012) 2:58–63. doi: 10.4103/2231-0762.109369
54. Kuter B, Guler N. Caries experience, oral disorders, oral hygiene practices and socio-demographic characteristics of autistic children. Eur J Paediatr Dent. (2019) 20:237–41. doi: 10.5772/intechopen.91809
55. Schoilew K, Ueffing H, Dalpke A, Wolff B, Frese C, Wolff D, et al. Bacterial biofilm composition in healthy subjects with and without caries experience. J Oral Microbiol. (2019) 11:1633194. doi: 10.1080/20002297.2019.1633194
56. Liu S, Li E, Sun Z, Fu D, Duan G, Jiang M, et al. Altered gut microbiota and short chain fatty acids in Chinese children with autism spectrum disorder. Sci Rep. (2019) 9:287. doi: 10.1038/s41598-018-36430-z
Keywords: autism, Fusobacteria, oral microbiome, dopamine transporter, mouse, metabolism
Citation: DiCarlo GE, Mabry SJ, Cao X, McMillan C, Woynaroski TG, Harrison FE, Reddy IA, Matthies HJG, Flynn CR, Wallace MT, Wu H and Galli A (2021) Autism-Associated Variant in the SLC6A3 Gene Alters the Oral Microbiome and Metabolism in a Murine Model. Front. Psychiatry 12:655451. doi: 10.3389/fpsyt.2021.655451
Received: 18 January 2021; Accepted: 16 March 2021;
Published: 15 April 2021.
Edited by:
Zachary Freyberg, University of Pittsburgh, United StatesReviewed by:
Peter Hamilton, Virginia Commonwealth University, United StatesElisa L. Hill-Yardin, RMIT University, Australia
Copyright © 2021 DiCarlo, Mabry, Cao, McMillan, Woynaroski, Harrison, Reddy, Matthies, Flynn, Wallace, Wu and Galli. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Aurelio Galli, YXVyZWxpby5nYWxsaUBnbWFpbC5jb20=
†These authors have contributed equally to this work and share first authorship
‡These authors have contributed equally to this work and share senior authorship