 Mihai Avram
Mihai Avram Helena Rogg
Helena Rogg Alexandra Korda
Alexandra Korda Christina Andreou
Christina Andreou Felix Müller
Felix Müller Stefan Borgwardt
Stefan Borgwardt- 1Department of Psychiatry and Psychotherapy, Schleswig Holstein University Hospital, University of Lübeck, Lübeck, Germany
- 2Department of Psychiatry (UPK), University of Basel, Basel, Switzerland
Psychiatry has a well-established tradition of comparing drug-induced experiences to psychotic symptoms, based on shared phenomena such as altered perceptions. The present review focuses on experiences induced by classic psychedelics, which are substances capable of eliciting powerful psychoactive effects, characterized by distortions/alterations of several neurocognitive processes (e.g., hallucinations). Herein we refer to such experiences as psychedelic states. Psychosis is a clinical syndrome defined by impaired reality testing, also characterized by impaired neurocognitive processes (e.g., hallucinations and delusions). In this review we refer to acute phases of psychotic disorders as psychotic states. Neuropharmacological investigations have begun to characterize the neurobiological mechanisms underpinning the shared and distinct neurophysiological changes observed in psychedelic and psychotic states. Mounting evidence indicates changes in thalamic filtering, along with disturbances in cortico-striato-pallido-thalamo-cortical (CSPTC)-circuitry, in both altered states. Notably, alterations in thalamocortical functional connectivity were reported by functional magnetic resonance imaging (fMRI) studies. Thalamocortical dysconnectivity and its clinical relevance are well-characterized in psychotic states, particularly in schizophrenia research. Specifically, studies report hyperconnectivity between the thalamus and sensorimotor cortices and hypoconnectivity between the thalamus and prefrontal cortices, associated with patients' psychotic symptoms and cognitive disturbances, respectively. Intriguingly, studies also report hyperconnectivity between the thalamus and sensorimotor cortices in psychedelic states, correlating with altered visual and auditory perceptions. Taken together, the two altered states appear to share clinically and functionally relevant dysconnectivity patterns. In this review we discuss recent findings of thalamocortical dysconnectivity, its putative extension to CSPTC circuitry, along with its clinical implications and future directions.
Introduction
The idea of investigating drug-induced effects that mimic symptoms of psychiatric disorders (i.e., psychosis) was spurred by the discovery of lysergic acid diethylamide (LSD) in 1943, which led to the first substance-induced model of psychosis (1), and later catalyzed the serotonin hypothesis of schizophrenia (2) [for further details see (3, 4)]. Although this hypothesis lacks supporting evidence, more recent models have suggested that drug-induced effects may shed light on the mental state of emerging psychosis and that the idea of using drug-induced effects as a model for psychosis might be still worth exploring (2, 5, 6). With the recent revival of psychedelic research, newly generated theories and supporting data may clarify whether the converging phenomena seen in both drug-induced states and endogenous psychosis share neurophysiological mechanisms.
Classic psychedelics or serotonergic hallucinogens (e.g., psilocybin, dimethyltryptamine (DMT), and LSD) are substances that can induce powerful psychoactive effects, by acting as agonists or partial agonists on serotonin 2A (5-HT2A) receptors (7–9). These psychoactive effects constitute so-called altered states of consciousness (ASC), which reflect temporary changes in an individual's mental state, and are characterized by distortions or alterations in several neurocognitive processes (e.g., perception, thoughts, mood) (10, 11). A variety of experiences can be elicited in this manner, which are influenced by several factors such as dose, environment, but also individual factors (12). We refer to psychedelic-induced ASC in this review as psychedelic states. Similarly, psychosis is a clinical syndrome including several symptoms such as alterations in perception (e.g., hallucinations), abnormal thinking (e.g., delusions), and bizarre behaviors (13), which are characterized by impaired reality testing, reflecting the ability to differentiate the external environment from one's internal world (14). While psychosis can be drug-induced (13), we refer to psychotic states as acute phases of so-called psychotic disorders (unless otherwise specified). In a sense, psychotic states can be understood as ASC (10, 15). Although psychotic states have mainly been associated with alterations in dopamine function, recent research indicates that other neurotransmitter systems may also be involved. For instance, substantial evidence demonstrates that alterations in glutamatergic transmission are relevant for schizophrenia—ranging from postmortem findings to in vivo imaging—and, importantly, that antagonists (e.g., ketamine) to specific glutamate receptors [i.e., N-methyl-D-aspartate (NMDA)] induce psychotic symptoms (16).
Psychedelic and psychotic states are accompanied by a plethora of phenomena, some of which are shared by both while other phenomena are distinct [(4, 17); for details see below]. For instance, a core characteristic of both altered states are perceptual disturbances, mainly hallucinations, however, other perceptual alterations (e.g., of time and space) and experiences with a higher power (e.g., mystical experiences) can also occur (4). Nevertheless, the perceptual disturbances are mainly characterized by distinct features in psychedelic and psychotic states, i.e.,—they are predominantly visual in psychedelic states and auditory in psychotic states (2). Furthermore, reality testing is not impaired in psychedelic states, meaning that subjects are (usually) aware that the experienced phenomena are drug-induced (4, 17); in contrast, patients suffering from psychosis are not able to trace the phenomena—which is considered real—to their medical condition (i.e., reality testing is impaired) (14). It remains to be determined, whether psychedelic and psychotic states reflect distinct or overlapping neural mechanisms. Following the “thalamic filter” model (18), a potential candidate for a shared neural mechanism in psychotic and psychedelic states is a disrupted thalamic filter function. In more detail, this model posits that the thalamic filtering of sensory information to the cortex is modulated by several sources (i.e., cortico-striatal pathways), which are, in turn, modulated by distinct neurotransmitter systems (e.g., dopaminergic, serotonergic). Aberrant modulation may lead to filtering deficits, resulting in an overload of exteroceptive and interoceptive stimuli, thereby bringing about psychedelic or psychotic phenomena (e.g., hallucinations). For a schematic of this model see Figure 1. This model has received recent support from functional magnetic resonance imaging (fMRI) studies pertaining both to psychotic and psychedelic states (22, 27–30). Indeed, recent efforts made to corroborate neuroimaging findings regarding altered perceptions concerning psychotic disorders and psychedelic substances, have highlighted altered thalamocortical connectivity—measured via resting-state fMRI (rsfMRI) functional connectivity—as a common finding (17). This review aims to discuss these recent findings of thalamocortical dysconnectivity, the putative extension of thalamocortical dysconnectivity to CSPTC circuitry, and clinical implications along with future directions.
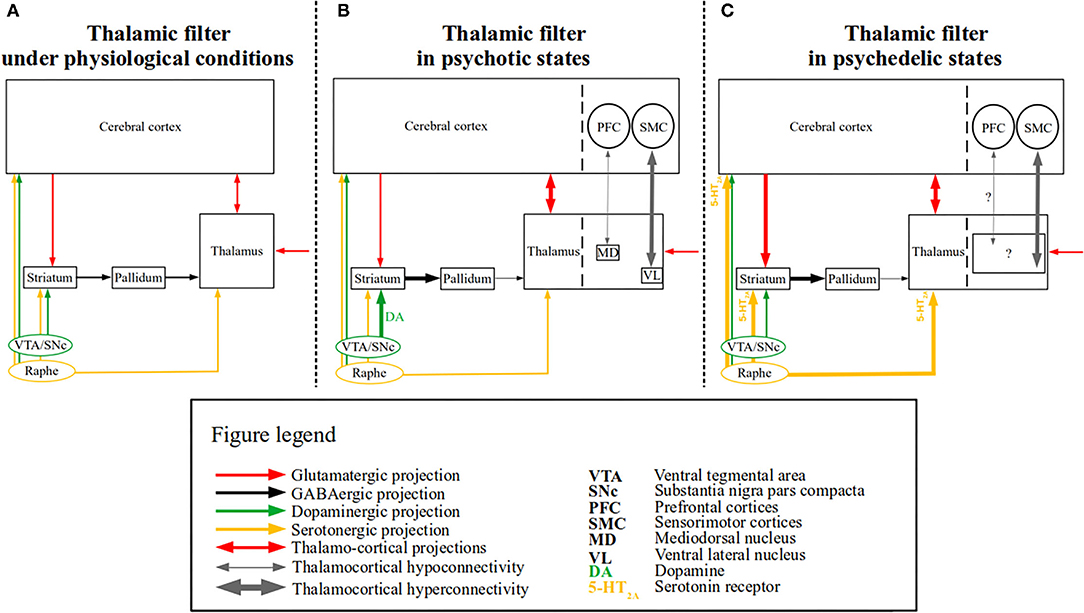
Figure 1. The “thalamic filter” model in psychotic and psychedelic states. A simplified schematic of the “thalamic filter” model is depicted along with the associated thalamocortical dysconnectivity in psychotic and psychedelic states. (A) Depicts thalamic filtering under physiological conditions, in which thalamic activity is modulated by several sources (e.g., cortico-striatal), which are in turn modulated by distinct neurotransmitter systems. Glutamatergic (red arrows), GABAergic (black arrows), dopaminergic (green arrows), and serotonergic projections (yellow arrows) are shown. Cortico-striatal projections are depicted by a unidirectional red arrow from the cerebral cortex to the striatum, whereas the reciprocal projections between the thalamus and the cerebral cortex are depicted by a bidirectional red arrow. Finally, incoming (glutamatergic) sensory input to the thalamus is depicted by a red unidirectional arrow. (B) Depicts a proposed model of altered thalamic filtering in psychotic states, induced via aberrant dopamine function, based on the model provided by Carlsson et al. (19). The left side shows the impaired thalamic filter function, which is induced by excess dopamine (shown as DA) and increased dopaminergic signaling (depicted by the thick green arrow), which leads to an increased inhibition of the pallidum via the striatum (depicted by the thick black arrow) and reduced thalamic inhibition via the pallidum (depicted by the thin black arrow). The enhanced thalamic disinhibition (i.e., “opening” of the thalamic filter) results in sensory flooding (depicted by the thick bidirectional red arrow between the thalamus and the cerebral cortex), presumably bringing about (psychotic) symptoms [for similar models see (20, 21)]. On the right side, separated by the dashed line both in the cerebral cortex and thalamus, rsfMRI findings of thalamocortical dysconnectivity in psychotic disorders are summarized [e.g., (22, 23)]. Thalamocortical hyperconnectivity (depicted by the thick bidirectional gray arrow) between the ventrolateral thalamus (VL) and sensorimotor cortices (SMC) and hypoconnectivity (depicted by the thin bidirectional gray arrow) between mediodorsal thalamus (MD) and prefrontal cortices (PFC), are shown. (C) Depicts a proposed model of altered thalamic filtering in psychedelic states, slightly modified from (9, 18). The left side shows the impaired thalamic filter function induced by psychedelic stimulation of 5-HT2A receptors, which leads to increased extracellular glutamate levels in the prefrontal cortex and inhibitory activity of interneurons in the basal ganglia, thereby increasing the excitatory effect of pyramidal neurons both on the basal ganglia (depicted by the thick unidirectional red arrow between the cerebral cortex and the striatum) and the thalamus (depicted by the thick bidirectional red arrow between the cerebral cortex and the thalamus) (24, 25). Recent evidence also indicates that psychedelics (i.e., LSD) modulate activity in the reticular thalamus leading to the disinhibition of other thalamic nuclei (i.e., mediodorsal nucleus—not shown) (26). In parallel, the increased striatal activity leads to a similar chain of events as seen in psychotic states (i.e., also resulting in enhanced thalamic disinhibition) (18). On the right side, separated by the dashed line both in the cerebral cortex and thalamus, rsfMRI findings of thalamocortical dysconnectivity in psychedelic states are summarized [e.g., (27, 28)]. The driving thalamic regions involved in these phenomena have not yet been identified (depicted by the black square containing “?”), nor is it clear whether thalamocortical hypoconnectivity (thin gray bidirectional arrow with “?”) is present.
The Thalamocortical System
The thalamocortical system refers to reciprocally connected pathways between the cortex and thalamus (31). These pathways are organized topographically, meaning that specific thalamic nuclei project to specific cortical areas and vice versa and that these connections are preferentially related to certain functions (32). Based on the main input (i.e., driver) thalamic nuclei can be classified into two categories, namely first-order and higher-order nuclei (33, 34). First-order nuclei (e.g., medial and lateral geniculate nuclei, ventral lateral nuclei) receive excitatory input from peripheral or subcortical structures and relay this information primarily to layer IV cortical neurons of primary sensory and motor areas. In turn, these cortical regions project back from neurons located in layer VI to the thalamic nuclei they received input from, thereby providing reciprocal feedback. In contrast, higher-order nuclei (e.g., pulvinar, mediodorsal nucleus) receive driving input from layer V cortical neurons and relay this information to other cortical areas, again primarily to layer IV. Based on this cortico-thalamo-cortical pattern of projections, higher-order nuclei have an important function in cortico-cortical communication (33). Additionally, all thalamic nuclei receive modulatory projections from cortical layer VI. This pathway has a 2-fold mode of action: sending excitatory projections to relay cells and inhibiting them via the reticular nucleus, a thin GABAergic sheet surrounding the thalamus (31, 35). In this framework, the thalamus modulates, or gates, both bottom-up sensory- and top-down cortical-information. These thalamocortical interactions, and the information processing they support, are carried out by glutamatergic neurotransmission, which is in turn modulated (e.g., changes in synaptic strength) by other neurotransmitter systems (e.g., dopaminergic) (36).
Notably, by employing MRI techniques, several aspects of the human thalamocortical system have been investigated in vivo. Thalamocortical structural connectivity has been investigated via diffusion tensor imaging (DTI) tractography (37–39), and, of particular interest, functional aspects of the thalamocortical system have been investigated via functional connectivity with rsfMRI, in both patients and healthy subjects (40, 41). Functional connectivity reflects the coherence of infra-slow fluctuations of ongoing brain activity, measured by correlating rsfMRI signal time-courses (42). Abnormal patterns of connectivity (e.g., decreased or increased) are referred to as dysconnectivity and should not be confused with disconnectivity, which reflects rather a decrease in function (e.g., cognitive) (43).
Thalamocortical Connectivity in Psychotic States
Overwhelming evidence indicates that the thalamus is altered in psychotic disorders, particularly in schizophrenia, including lower cell count in some thalamic nuclei (i.e., pulvinar) (44, 45), volume reduction (46), altered activity during cognitive tasks (47, 48), and reduced structural thalamocortical connectivity (49). However, one of the most consistent large-scale in vivo brain imaging findings in psychotic states is altered thalamocortical functional connectivity, measured via rsfMRI. Altered thalamocortical connectivity has been consistently reported in patients with psychotic disorders (50, 51), including in individuals at clinical high risk for psychosis (52, 53), first-episode psychosis (23), and patients with established bipolar disorder (30, 54) and schizophrenia (22, 29, 55). Specifically, thalamocortical connectivity is characterized by two distinct patterns in patients with psychosis: (i) compared to healthy controls, patients show increased functional connectivity (i.e., hyperconnectivity) between the thalamus and sensorimotor regions, including motor, temporal, and occipital areas, and, conversely, (ii) decreased functional connectivity (i.e., hypoconnectivity) between the thalamus and prefrontal and cerebellar regions (22, 23, 29, 30, 52). Remarkably, despite the different approaches used to investigate thalamocortical connectivity and distinct clinical populations, findings consistently indicate that sensorimotor areas are preferentially hyperconnected with ventral lateral/posterior nuclei, whereas prefrontal areas show hypoconnectivity primarily with mediodorsal/ anterior nuclei (22, 29, 45). Intriguingly, findings indicate that these altered thalamocortical connectivity patterns are in line with the topographical organization of thalamocortical projections [for details on topography see (32)]. In support of such topographical organization, prefrontal hypoconnectivity with the higher-order mediodorsal nucleus, which is involved in specific cognitive domains (e.g., memory, executive function) (56), was associated with cognitive disturbances in patients (22, 23). Similarly, evidence indicates that sensorimotor areas are hyperconnected with first-order ventral lateral nuclei (22, 23), which are involved in sensorimotor processing (57). Moreover, this hyperconnectivity pattern has been reported to correlate with psychotic symptoms (22). Although not restricted to the ventral lateral nuclei, several findings indicate a link between thalamocortical hyperconnectivity and psychotic symptoms (52, 58) or transition to psychosis (53). In support, a recent meta-analysis on altered thalamocortical connectivity in psychosis reported a significant, albeit non-specific, relationship between hyperconnectivity and psychotic symptoms across several studies (51). This non-specificity might be related to the rather integrative nature of CSPTC circuitry (see below), corresponding to recently identified integration zones (i.e., areas that combine information from several cortices), as well as individual variations in connectivity patterns (59, 60).
Beyond altered thalamocortical connectivity, altered cortico-striatal and cortico-pallidal connectivity have also been reported in psychotic states (61–64). These findings indicate that thalamocortical connectivity might be embedded in larger, topographically organized feedback circuits between the cortex and both the thalamus and basal ganglia (65). In fact, research suggests that thalamocortical hyper- and hypoconnectivity with intrinsic brain networks extend topographically to the basal ganglia (22). These findings lend support to an older theoretical model that suggested a link between psychotic disorders and CSPTC circuitry alterations (66). Particularly, this model posited that in psychotic disorders, dopaminergic hyperactivity in the striatum would lead, via pallidal projections, to altered cortico-thalamic activity. In line with the model's prediction, a study combining rsfMRI with fluorodopa positron emission tomography (PET) demonstrated that thalamic hypo- and hyperconnectivity with prefrontal and sensorimotor areas are associated with dopamine synthesis capacity and storage in patients with schizophrenia, respectively (55). This suggests that altered dopaminergic transmission modulates CSPTC circuitry, including thalamocortical dysconnectivity, in psychotic states. Specifically, changes in dopaminergic transmission have been shown to modify connectivity strength [see Cole et al. (67)].
In summary, in psychotic states thalamocortical connectivity is altered, extends to the basal ganglia and appears to be modulated by altered dopaminergic transmission, while also being associated preferentially with distinct symptom dimensions (i.e., hyperconnectivity with psychotic symptoms/ hypoconnectivity with cognitive disturbances).
Thalamocortical Connectivity in Psychedelic States
Although findings of altered thalamocortical connectivity in psychedelic states are somewhat less well-established, considerable evidence indicates thalamic involvement. Animal studies have shown that the thalamus, mainly its higher-order nuclei, is modulated by serotonergic afferents from the dorsal and medial raphe and is, therefore, a relevant site for serotonergic transmission (68–70). Furthermore, psychedelic states might be induced via modulatory effects of 5-HT2A receptors located presynaptically on thalamocortical afferents to the prefrontal cortex (9, 24). In humans, following the administration of psychedelics, PET and single-photon emission computed tomography (SPECT) studies have, on the one hand, reported alterations in thalamic glucose metabolism (71, 72), while, on the other hand, reported decreased cerebral blood flow in the thalamus (73–75). Indirect evidence also implicates the thalamus in psychedelic states, as sensorimotor gating is altered, and this can be achieved by disrupting CSPTC circuitry (18, 76–78).
More recent evidence of thalamic involvement in psychedelic states comes from rsfMRI-based neuropharmacological investigations, which typically employ a crossover design. Specifically, participants are scanned on several occasions, in drug-induced states vs. placebo, which are then contrasted to one another (see below). Such studies mainly rely on the investigation of functional connectivity, and despite methodological differences across studies, a pattern of psychedelic-induced altered thalamocortical connectivity has emerged (27, 28, 79–82). In the following, we will focus on studies that investigated thalamocortical connectivity with seed-based correlation analyses, using the thalamus as the seed (see Table 1 for details).
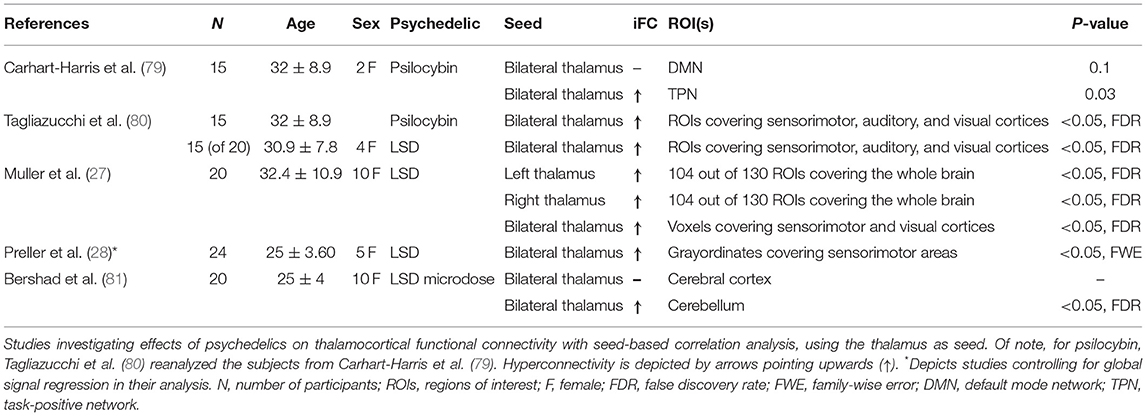
Table 1. Thalamocortical connectivity after administration of psychedelics.
Carhart-Harris et al. (79) investigated thalamic connectivity with the default-mode network (DMN) and a task-positive network (TPN, i.e., regions negatively correlated with a seed of the ventral medial-prefrontal cortex) after psilocybin administration. Specifically, the authors computed seed-based analyses, in which the time-course of the bilateral thalamus was correlated with the time-course of the DMN and TPN, respectively. They found no changes in connectivity between the thalamus and DMN after psilocybin administration, but did observe an increase in connectivity between the thalamus and the TPN—which included areas of the posterior parietal cortex and sensorimotor regions.
In the study by Tagliazucchi et al. (80), thalamocortical connectivity was investigated via seed-based analysis after LSD administration. The time series of the bilateral thalamus was correlated with the time series of 401 regions-of-interest (ROIs), covering cortical and subcortical gray matter. The authors found increased connectivity between the thalamus and primary sensory cortices, including sensorimotor, auditory, and visual cortices. Remarkably, by reanalyzing a previous dataset in which psilocybin was administered instead of LSD [the same subjects as in (79) described above], the authors reported the same pattern of thalamocortical hyperconnectivity with primary sensorimotor areas, which suggests that thalamic hyperconnectivity does not reflect the effect of a specific substance, but rather a broader psychedelic-induced phenomenon.
With a somewhat different approach, Muller et al. (27) investigated thalamocortical connectivity after LSD administration with several methods: ROI-to-ROI, ROI-to-voxel, and global correlation analysis. In the ROI-to-ROI analysis, they investigated thalamocortical connectivity by correlating the time-course of the left and right thalamus with the time-courses of 130 ROIs, covering the whole brain. They found very similar connectivity profiles for the left and right thalamus, and that, compared to placebo, thalamocortical connectivity was increased with the majority of the ROIs, akin to the study by Tagliazucchi et al. (80). This analysis was then followed by the ROI-to-voxel analysis, in which they combined the left and right thalamus and correlated the time-course of this bilateral mask with every other voxel in the brain. Consistent with their previous analyses, the authors found increased thalamic connectivity with many brain regions following LSD administration, particularly with primary sensory regions (e.g., occipital cortex). Finally, Muller and colleagues computed a global correlation analysis by averaging the correlation coefficients of each voxel to each other voxel. The authors found that beyond increased thalamic connectivity, the basal ganglia also had increased connectivity with the rest of the brain, indicating LSD-induced alterations in the CSPTC circuitry. Remarkably, Muller and colleagues also found that LSD-induced thalamic hyperconnectivity with multimodal sensory cortices (i.e., fusiform gyrus and insular cortex) was positively associated with alterations in sensory perceptions [i.e., “visionary restructuralization” and “auditory alterations” sub-scales of the five dimensions ASC scale (5D-ASC)], indicating functional relevance of the altered thalamocortical connectivity patterns.
Finally, using a more ambitious experimental design, Preller et al. (28) investigated thalamocortical connectivity across three different conditions: placebo, LSD, and LSD + ketanserin (i.e., ketanserin is a 5-HT2A receptor antagonist that blocks LSD effects). Specifically, the authors correlated the average time-courses of all grayordinates (i.e., gray matter location represented by a surface vertex or a volume voxel) in the bilateral thalamus with all grayordinates in the brain. Additionally, the authors investigated the effect of global signal regression (GSR) on these analyses. GSR, a controversial preprocessing strategy for rsfMRI, removes global effects elicited by motion and respiration, but possibly also valuable neuronal effects (i.e., distributed neural information) (83). Although GSR had a significant effect on the LSD-induced altered connectivity [the discussion of this effect is beyond the scope of this review, see (28) for details], increased connectivity between the thalamus and sensorimotor areas were found following LSD administration, irrespective of GSR. Additionally, the authors found that ketanserin efficiently obstructed both LSD-induced subjective experiences and changes in brain connectivity, supporting the notion that LSD effects are elicited via 5-HT2A receptors.
In summary, in psychedelic states thalamocortical connectivity with sensorimotor cortices is increased, possibly extending to the basal ganglia via alterations in CSPTC circuitry, which appear to be (at least partially) modulated by 5-HT2A receptors. Furthermore, thalamocortical hyperconnectivity with sensorimotor cortices is associated with subjective visual and auditory perceptual alterations.
Thalamocortical Hyperconnectivity as the Common Denominator in Psychotic and Psychedelic States
Thalamic hyperconnectivity with sensorimotor cortices appears to be a common denominator in psychotic and psychedelic states, possibly reflecting a shared biological mechanism involved in abnormal perception (18). In line with this idea, thalamic hyperconnectivity with sensorimotor cortices has been associated with psychotic symptoms in psychotic states in addition to being associated with altered subjective visual and auditory perceptions in psychedelic states, respectively (27, 51). Furthermore, consistent with thalamocortical findings, the same cortical sensorimotor regions have been reported to be hyperconnected to the basal ganglia in both altered states, indicating connectivity alterations along the CSPTC circuitry (22, 27, 55). In line with the thalamic filter model, such alterations might be induced via endogenous (e.g., elevated dopaminergic transmission as seen in psychosis) and/or exogenous modulation (e.g., elevated serotonergic transmission following psychedelic administration) of cortico-striatal pathways, which in turn modulate thalamocortical connectivity (2, 9, 18) (also see Figure 1). This model has recently received support from a study that investigated resting-state effective connectivity of CSPTC circuitry, following administration of LSD, LSD + ketanserin, and for control, placebo (84). Compared to functional connectivity, effective connectivity, quantified with spectral dynamic causal modeling, can be used to infer the direction or causality of connectivity (e.g., from the thalamus to cortex or vice-versa). The authors found that LSD increased connectivity from the thalamus to the posterior cingulate cortex (PCC)—part of the DMN comprising association cortices rather than sensorimotor ones (85)—and decreased connectivity from the PCC to the thalamus. These effects were dependent on 5-HT2A receptors (i.e., were blocked by ketanserin). Additionally, independent of 5-HT2A receptors, LSD decreased connectivity from the (ventral) striatum to the thalamus, perhaps reflecting striatal induced thalamic disinhibition. Ketanserin blocked the increased connectivity from the thalamus to the PCC, suggesting that thalamocortical hyperconnectivity might be driven by serotonergic effects on 5-HT2A receptors and might therefore reflect secondary effects of increased serotonergic transmission in psychedelic states. Apparently supporting this idea, it has been shown that a single dose of selective serotonin reuptake inhibitors (SSRIs) leads to increased degree centrality in the thalamus (86). One should note, however, that increased degree centrality reflects an increased number of connections between, in this case, the thalamus and the rest of the brain. This differs from thalamic hyperconnectivity with sensorimotor cortices, which rather reflects an increase in connectivity strength—i.e., increased functional coupling. Furthermore, as mentioned above, we posit that thalamocortical hyperconnectivity with sensorimotor cortices has functional relevance regarding altered perceptions, which is supported by the fact that ketanserin blocked not only some of the neural effects but also the subjective ones [also see (28)]. In accordance with this, LSD microdosing (i.e., 10–15 μg vs. 100+ μg LSD), which did not induce psychedelic phenomena (e.g., perceptual distortions) also did not elicit thalamocortical hyperconnectivity (81).
As an interesting sidenote, thalamocortical hyperconnectivity with sensorimotor areas has also been identified during distinct stages of sleep, possibly indicative of dream phenomena (87, 88). It would be interesting for future research to explore whether thalamocortical hyperconnectivity with sensorimotor areas reflects shared biological underpinnings of dream-like and psychedelic-induced phenomena.
Variations of Psychotic and Psychedelic States
Despite sharing common features, there are also marked differences between psychotic and psychedelic states regarding both phenomenology (e.g., perceptual disturbances) and neural correlates. Perhaps one of the most important distinctions is the duration of the experience, with drug effects typically subsiding within a couple of hours. The duration will affect how the experience is incorporated into the subjects' Weltanschauung, with longer-lasting perceptual abnormalities possibly leading to delusional belief systems capable of “explaining” the subjective experience, as seen in psychosis (89). Additionally, although similar perceptual disturbances have been reported in psychotic (both endogenous and drug-induced) and psychedelic states (90), a remarkable distinction is related to the type of perceptual distortion. Specifically, in psychotic states, auditory hallucinations reflect the usual perceptual disturbances, whereas visual hallucinations are more common in psychedelic states (2). Furthermore, psychotic states are often accompanied by negative symptoms and cognitive disturbances, which often precede and outlast the psychotic episodes (16), whereas negative emotions and cognitive disturbances are transient in psychedelic states, and are typically associated with a “bad trip” or higher dosages (17). Finally, and in contrast with most psychotic states, subjects experiencing psychedelic states are aware of the transient nature of their experience and the cause thereof (91, 92).
Notably, psychotic and psychedelic states are accompanied by a plethora of phenomena in addition to altered perception (4). For instance, in psychedelic states, experiential changes appear to be dependent on dosage and include, beyond changes in perception, derealization and depersonalization (i.e., ego dissolution), and spiritual experiences (e.g., insightfulness, blissful state), at higher dosages (12). However, in this review we focused on phenomena that are (i) characteristic and (ii) shared by both altered states, and (iii) functionally relevant for thalamocortical connectivity changes [e.g., psychotic symptoms in psychotic disorders (51) and alterations of subjective visual or auditory perceptions (27)]. Whereas, derealization and depersonalization, in addition to ascribing altered perceptions to a higher power are also present in psychotic states (17), these phenomena appear to be related to DMN alterations rather than to changes in thalamocortical connectivity in psychedelic states (93, 94), and were therefore not discussed.
There are additional differences observed in the neural correlates of psychotic and psychedelic states. Particularly, thalamocortical hyperconnectivity with sensorimotor cortices is only transient in subjects receiving psychedelics (i.e., hyperconnectivity is present after psychedelic administration but not during placebo conditions) but is stable in psychotic states. Interestingly, thalamocortical hyperconnectivity with sensorimotor cortices is already present before the onset of psychosis in subjects at clinical high risk (52), in first-degree relatives of patients with psychosis (54), and in patients with schizophrenia in remission of psychotic symptoms (55). In short, non-transient thalamocortical hyperconnectivity with sensorimotor cortices is present even if the subjects are not currently psychotic (but see below). This suggests that thalamocortical hyperconnectivity also reflects proneness to psychosis, in addition to psychotic phenomena per se. Put differently, while thalamocortical hyperconnectivity might reflect a state marker in psychedelic states, it seems to reflect a trait marker in psychotic states. In support, it has been shown that thalamocortical hyperconnectivity in subjects at clinical high risk predicts later transition to full-blown psychosis (52, 53). Furthermore, thalamocortical hyperconnectivity is usually accompanied by thalamocortical hypoconnectivity with prefrontal cortices, which appears to be missing in psychedelic states. Finally, in contrast to psychedelic states, structural connectivity studies (based on DTI or diffusion weighted imaging—DWI) revealed a similar pattern of thalamocortical dysconnectivity in patients with psychosis: reduced structural connectivity between the thalamus and prefrontal areas (49, 95–99) and increased structural connectivity between the thalamus and sensorimotor areas (49, 95, 98, 99). Crucially, the combined functional and structural dysconnectivity findings indicate substantial disorganization of the thalamocortical system in psychosis [i.e., shared functional and structural alterations—see Brandl et al. (50)]. Both functional (100) and structural connectivity alterations (99) have also been reported for the unaffected siblings of patients, indicating a link between predisposition for psychosis and thalamocortical dysconnectivity. However, while both functional (100) and structural studies (99) reported thalamocortical hypoconnectivity with prefrontal areas in unaffected siblings, thalamocortical hyperconnectivity with sensorimotor areas was not found [but see Lui et al. (54)]. This suggests that thalamocortical hyperconnectivity is specifically associated with disorder-related processes (e.g., perhaps perceptual alterations). We speculate that the stability of the thalamocortical dysconnectivity patterns observed in psychosis (i.e., across the stages of psychosis, persistence despite treatment with antipsychotic medication) is grounded in the substantial disorganization of the thalamocortical system. In contrast, thalamocortical dysconnectivity in psychedelic states is not only transient, but can be blocked by ketanserin (28) and perhaps antipsychotics (26).
In summary, psychotic states differ from psychedelic states in several aspects of phenomenology, including the absence of thalamocortical hypoconnectivity, and the persistence of thalamocortical hyperconnectivity.
Hierarchical Predictive Coding in Psychotic and Psychedelic States
Both psychotic and psychedelic states have been discussed in the context of hierarchical predictive coding (5, 11, 17, 89, 101, 102). Put briefly, hierarchical predictive coding refers to Bayesian approaches that highlight processing of information as an interplay between prior expectations (i.e., top-down predictions about the world) and current inputs (e.g., bottom-up sensory information). Discrepancies between hierarchically organized predictions and outcomes (i.e., so-called prediction errors) lead to an update in the system, resulting in the generation of new predictions. The prediction errors, reflecting uncertainty, are weighted in the system with regard to their precision. It has been proposed that predictions and prediction errors might rely on glutamatergic projections, whereas the weighting of the prediction errors (i.e., their precision) might be driven via neuromodulators such as dopamine, serotonin, and acetylcholine (11). In this context, distinct aspects of the thalamocortical system would organize both feedforward (i.e., bottom-up) and feedback (i.e., top-down) information processing. Notably, however, first-order and higher-order thalamic nuclei play different roles. For instance, cortico-cortical communication could represent the priors, while thalamocortical projections originating in first-order thalamic nuclei reflect feedforward information (i.e., relaying sensory information to the cortex), and cortico-thalamic projections (i.e., form neurons in cortical layer VI) provide reciprocal feedback (101). On the other hand, higher-order nuclei might have a role in modulating feedforward and feedback information-flows by providing precision expectations for the prediction errors (e.g., establishing context) (103). Furthermore, feedforward and feedback information-flows are also modulated by striato-pallidal projections, which are themselves influenced either by cortical projections or, as mentioned above, distinct neuromodulators (65).
It has been argued that (endogenous or exogenous) alterations of the bottom-up/top-down balance might lead to altered perceptions, as seen in psychotic and psychedelic states (11). In this context it is important to consider the mechanisms through which such alterations can be elicited. For instance, psychedelics elicit their effects primarily by acting on 5-HT1A and 5-HT2A receptors (9, 104), which have a much higher expression in limbic (e.g., hippocampal) and cortical areas (e.g., association cortices), and lower receptor density in subcortical areas such as the thalamus (105, 106). By acting on 5-HT1A and 5-HT2A receptors, psychedelics alter the function of layer V pyramidal neurons, which in turn change the activity of other cortical areas. Such changes might increase entropy of cortical activity, leading to an increased relaxation of high-level priors (106). The relaxation of high-level priors results in weaker control over the bottom-up information-flow (89), possibly leading to hallucinations [for discussion of strong high-level priors leading to hallucinations see Sterzer et al. (102)]. Such an altered top-down mechanism is postulated in the so-called REBUS model (i.e., relaxed beliefs under psychedelics) (89). This hypothesis appears to be in contrast with the previously mentioned thalamic filter model (18), which rather suggests a bottom-up alteration (i.e., opening of the thalamic filter leading to sensory flooding). However, layer V pyramidal neurons also project to the thalamus (107), and can therefore alter thalamic activity, despite the lower 5-HT1A/2A receptor density in the thalamus (106). Both models are supported by previous findings (84) and the reports reviewed herein (i.e., thalamocortical connectivity lacks information on directionality).
Future Directions and Conclusions
In contrast to psychotic states, the involvement of specific thalamic nuclei in the hyperconnectivity with sensorimotor cortices (i.e., driving the effect) is unclear in psychedelic states. This limitation is based on the methodology employed so far in psychedelic neuroimaging research (i.e., seed-based functional connectivity from the thalamus). We suggest future studies also investigate the connectivity from distinct cortical areas to the thalamus in a voxel-wise manner. This could allow for the identification of specific thalamic sub-regions [i.e., by matching the findings with fine-grained thalamic subdivisions (108)] and possibly also alterations in functional topography. This could reveal how the mechanisms leading to thalamocortical hyperconnectivity might differ in the two altered states. In line with this idea, psychedelics might also induce thalamocortical hyperconnectivity via effects elicited directly on the thalamus. Inserra et al. (26) found that LSD modulates neurons of the reticular thalamus in mice, which in turn disinhibits the mediodorsal nucleus. Such disinhibition of higher-order nuclei might very well be associated with increased thalamocortical interaction (indeed, an increase in the firing of prefrontal pyramidal neurons was also observed), possibly corresponding to thalamocortical hyperconnectivity. Interestingly, Inserra et al. (26) suggest that the LSD-induced disinhibition of the mediodorsal nucleus was mediated by dopaminergic transmission (i.e., the effect was reversed by antipsychotics). This might be in contrast to the findings in humans, as antipsychotic medication does not appear to normalize thalamocortical dysconnectivity in patients with schizophrenia (55). It is not yet known whether the mediodorsal nucleus is hyperconnected to prefrontal areas in humans, as this has not yet been evaluated directly. However, such hyperconnectivity would be in contrast to psychosis findings, which report hypoconnectivity for this nucleus. The mediodorsal nucleus has been consistently implicated in the pathophysiology of schizophrenia (109), and appears to play a crucial role in the impaired cognitive function of patients (47, 109–111). Furthermore, as previously stated, the hypoconnectivity of this nucleus with prefrontal areas and its link to cognition is well in line with the topographical organization of the thalamocortical system (22, 29, 55). It is possible that such topographical organization is not preserved in psychedelic states. In support, there is evidence that connectivity increases between distinct and normally segregated intrinsic brain networks, following psychedelic administration (80, 112). An altered topographical organization might enable crosstalk between distinct and normally segregated thalamic nuclei (and their cortical connections), possibly resulting into psychedelic phenomena. Future studies investigating the connectivity of distinct thalamic nuclei with the cortex following psychedelic administration might shed some light on this issue.
It is worth mentioning that although dopamine appears to play a central role in the pathophysiology of psychosis and psychedelics mainly act on the serotonergic system, there is substantial evidence indicating that the two systems are highly interconnected (113). For instance, while all licensed antipsychotics have antagonistic effects at dopamine D2 receptors (16), atypical antipsychotics have additional therapeutic benefits (e.g., reduced extrapyramidal side-effects) by also blocking 5-HT2A receptors (114). Interestingly, blocking 5-HT2A receptors also appears to reverse acute effects of NMDA antagonists (see below), indicating complex interactions between the serotonergic, dopaminergic, and glutamatergic systems (24, 114). Such three-way interactions are well in line with the mounting evidence that glutamatergic dysfunction is also implicated at least in some subgroups of patients with psychosis or stages of the disorder (115). Moreover, some 5-HT2A receptor agonists (i.e., psilocybin) appear to modulate dopamine release in the striatum of healthy volunteers (116). Despite these findings, several aspects regarding dopamine-serotonin interactions following psychedelic administration remain unclear. For instance, highly relevant to the topic at hand, it remains to be determined whether antipsychotics block all neural and psychological effects of psychedelics.
In apparent contrast to the findings reported in this review, techniques employing perceptual deprivation such as the multimodal Ganzfeld exposure (117)—which are capable of eliciting visual and auditory perceptual changes—report thalamocortical hypoconnectivity with sensory cortices (i.e., auditory, visual regions). However, the differentially induced subjective experiences differ both qualitatively and quantitatively, and presumably also the associated connectivity changes. Not only are several dimensions assessed with the ASC scale absent from the perceptual deprivation induced effects (e.g., insightfulness, synesthesia etc.) but even those that are present, are markedly reduced in comparison to the psychedelic-induced effects (118). Furthermore, the perceptual changes following perceptual deprivation techniques have not yet been associated with altered thalamocortical connectivity (in contrast to psychedelic-induced effects) and might rather reflect an imbalance between typical top-down signaling and atypical (i.e., unstructured) bottom-up input (117).
A potential limitation of our review concerns the fact that LSD studies dominate the findings of thalamocortical dysconnectivity in psychedelic states (Table 1). There is a paucity of studies investigating the acute effects of other classic psychedelics such as DMT and mescaline, or even entactogens such as 3,4-methylenedioxymethamphetamine (MDMA) on thalamocortical functional connectivity. We cannot therefore exclude that the findings on thalamocortical dysconnectivity in psychedelic states reported herein reflect rather LSD-specific changes than effects of psychedelics in general. Ketamine was not included in our review as it is not a “classic” psychedelic, but rather a hallucinogenic anesthetic, which elicits its effects mainly via NMDA antagonism—in contrast to the classic psychedelics, which are 5-HT2A (partial) agonists (6, 8). However, ketamine is relevant in this context, as it has been associated with both psychotic- and negative-like symptoms in healthy volunteers, and appears, therefore, to reflect the most comprehensive model of schizophrenia up to date (119). Crucially and providing an important incentive for future neuroimaging research investigating the effects of psychedelics, studies investigating the acute effects of ketamine on thalamocortical connectivity followed approaches from psychosis research. For instance, using the methods reported in (29), Höflich et al. (120) demonstrated that acute ketamine effects elicit thalamocortical hyperconnectivity with temporal and sensorimotor cortices in healthy volunteers. In support, a recent multi-site study, following the approach reported in (121), demonstrated hyperconnectivity between the mediodorsal nucleus and auditory cortices and the parahippocampal gyrus (122). Furthermore, both studies demonstrated that transient psychotic-like symptoms accompanied the thalamocortical hyperconnectivity. Taken together, these findings indicate an overlap in altered thalamocortical connectivity with sensorimotor cortices induced both by ketamine and classic psychedelics (i.e., LSD and psilocybin), supporting the idea of thalamocortical dysconnectivity as a biological mechanism presumably underlying altered perceptions. Notably, while several substances (e.g., alcohol, illicit drugs etc.) are capable of eliciting psychotic-like phenomena, possibly via an impaired thalamic filter function [see for example amphetamine and ketamine in (18)], psychedelics appear to also have long-lasting positive effects, in addition to the transient psychotic-like phenomena they elicit. For instance, beyond stable improvements in depression and addiction (106), psychedelics have been shown to increase optimism and trait openness in healthy volunteers (123), and enhance (self-reported) positive behavioral changes that lasted up to 1 year (124).
Another possible limitation of our argument regarding thalamocortical hyperconnectivity with sensorimotor cortices as shared neural correlate of altered perceptions, is the apparent ubiquity of this phenomena in psychiatric disorders (125, 126). This begs the question whether this pattern of altered connectivity is specific for phenomena like altered perceptions, as seen in psychotic and psychedelic states. Large meta-analyses on rsfMRI data have shown that this connectivity pattern is consistent in schizophrenia (50) and bipolar disorder (127), but not other conditions occasionally reporting thalamocortical dysconnectivity (128–130). Additionally, evidence suggests that thalamocortical hyperconnectivity is more substantial in schizophrenia and bipolar disorder than in major depression disorder (131), indicating a continuum, with schizophrenia at the end of the spectrum. We speculate that a similar continuum is also present across distinctly induced psychedelic states, with LSD presumably toward the end of the spectrum. Future studies might elucidate this issue by contrasting between thalamocortical connectivity elicited by distinct substances in the same participants.
A note of caution regarding the findings of thalamocortical dysconnectivity in psychotic states concerns the medication status of the patients. Antipsychotic medication is known to affect functional connectivity in patients with psychotic disorders (132–134) and the majority of studies reviewed herein based their findings on medicated patients [e.g., (22, 23, 55)]. However, there is also evidence that thalamocortical dysconnectivity is present in unmedicated individuals at clinical high risk (52) and in drug-naive adolescents with early-onset schizophrenia (135), indicating that the patterns of dysconnectivity might be independent of antipsychotic medication. Nevertheless, an influence of antipsychotic medication on thalamocortical dysconnectivity in psychotic states cannot be excluded completely.
The relationships between drug-induced alterations in distinct neurocognitive processes (and their meaning to the subject) and underlying biological mechanisms are highly relevant for psychotic research. Therefore, it is necessary to determine not only differences in phenomenology but also in the underlying biological mechanisms. In this review, we identified similar patterns of thalamocortical hyperconnectivity with sensorimotor areas in psychedelic and psychotic states, indicating a shared biological mechanism. In contrast, thalamocortical hypoconnectivity with prefrontal cortices was not observed in psychedelic states (see Table 1). It is unclear whether this pattern of dysconnectivity may not be elicited by psychedelics or whether reports in this regard are simply missing. Consistent with the former, however, thalamocortical hypoconnectivity is not elicited by ketamine either (120, 122). As thalamocortical hypoconnectivity with prefrontal cortices is one of the most robust imaging findings in psychosis research (51), it is important to establish why psychedelics might not elicit this phenomenon. As discussed above, it is possible that thalamocortical hypoconnectivity may not be reflective of psychosis per se but rather of associated phenomena such as cognitive impairment or vulnerability for psychosis, and that thalamocortical hyperconnectivity with sensorimotor cortices might be more evocative of psychotic phenomena, possibly reflecting a biomarker of psychosis (136). It is, therefore, remarkable that only this pattern of dysconnectivity appears to be elicited by classic psychedelics and ketamine, which more accurately model psychotic symptoms than negative or cognitive symptoms (119, 123). We conclude that the findings reviewed in our paper indicate that at least some aspects of psychosis can be modeled by psychedelics, regarding both alterations of neurocognitive processes (i.e., perception) and underlying biological mechanisms (thalamocortical hyperconnectivity). Nevertheless, additional research is needed to better characterize both shared and distinct aspects of psychotic and psychedelic states before valid psychedelic-based pharmacological models can be established for psychotic disorders.
Author Contributions
MA and SB conceived the original idea and outlined the study. MA, HR, AK, CA, FM, and SB contributed to the literature review and edited the manuscript. All authors contributed to the article and approved the submitted version.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher's Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Acknowledgments
The authors thank Alyssa Torske from the Technical University of Munich, School of Medicine for proofreading the manuscript.
References
1. Osmond H. A review of the clinical effects of psychotomimetic agents. Ann N Y Acad Sci. (1957) 66:418–34. doi: 10.1111/j.1749-6632.1957.tb40738.x
2. Geyer MA, Vollenweider FX. Serotonin research: contributions to understanding psychoses. Trends Pharmacol Sci. (2008) 29:445–53. doi: 10.1016/j.tips.2008.06.006
3. Nichols DE, Walter H. The history of psychedelics in psychiatry. Pharmacopsychiatry. (2021) 54:151–66. doi: 10.1055/a-1310-3990
4. Hermle L, Kraehenmann R. Experimental psychosis research and schizophrenia-similarities and dissimilarities in psychopathology. Curr Top Behav Neurosci. (2018) 36:313–32. doi: 10.1007/7854_2016_460
5. Corlett PR, Honey GD, Fletcher PC. From prediction error to psychosis: ketamine as a pharmacological model of delusions. J Psychopharmacol. (2007) 21:238–52. doi: 10.1177/0269881107077716
6. Corlett PR, Honey GD, Krystal JH, Fletcher PC. Glutamatergic model psychoses: prediction error, learning, and inference. Neuropsychopharmacology. (2011) 36:294–315. doi: 10.1038/npp.2010.163
7. Nichols DE. Hallucinogens. Pharmacol Ther. (2004) 101:131–81. doi: 10.1016/j.pharmthera.2003.11.002
9. Vollenweider FX, Preller KH. Psychedelic drugs: neurobiology and potential for treatment of psychiatric disorders. Nat Rev Neurosci. (2020) 21:611–24. doi: 10.1038/s41583-020-0367-2
10. Revonsuo A, Kallio S, Sikka P. What is an altered state of consciousness? Philos Psychol. (2009) 22:187–204. doi: 10.1080/09515080902802850
11. Corlett PR, Frith CD, Fletcher PC. From drugs to deprivation: a Bayesian framework for understanding models of psychosis. Psychopharmacology. (2009) 206:515–30. doi: 10.1007/s00213-009-1561-0
12. Hirschfeld T, Schmidt TT. Dose-response relationships of psilocybin-induced subjective experiences in humans. J Psychopharmacol. (2021) 35:384–97. doi: 10.1177/0269881121992676
13. Gaebel W, Zielasek J. Focus on psychosis. Dialogues Clin Neurosci. (2015) 17:9–18. doi: 10.31887/DCNS.2015.17.1/wgaebel
14. American Psychiatric Association. Diagnostic and Statistical Manual of Mental Disorders. 5th ed. Arlington, VA: American Psychiatric Association (2013).
15. Schmidt TT, Maji T. Empirische Untersuchung veränderter Bewusstseinszustände. In: von Heyden M, Jungaberle H, Majić T, editors. Handbuch Psychoaktive Substanzen. Springer Reference Psychologie. Berlin; Heidelberg: Springer (2016). 1–25. doi: 10.1007/978-3-642-55214-4_65-1.p
16. McCutcheon RA, Krystal JH, Howes OD. Dopamine and glutamate in schizophrenia: biology, symptoms and treatment. World Psychiatry. (2020) 19:15–33. doi: 10.1002/wps.20693
17. Leptourgos P, Fortier-Davy M, Carhart-Harris R, Corlett PR, Dupuis D, Halberstadt AL, et al. Hallucinations under psychedelics and in the schizophrenia spectrum: an interdisciplinary and multiscale comparison. Schizophr Bull. (2020) 46:1396–408. doi: 10.1093/schbul/sbaa117
18. Vollenweider FX, Geyer MA. A systems model of altered consciousness: integrating natural and drug-induced psychoses. Brain Res Bull. (2001) 56:495–507. doi: 10.1016/S0361-9230(01)00646-3
19. Carlsson A, Waters N, Carlsson ML. Neurotransmitter interactions in schizophrenia-therapeutic implications. Eur Arch Psychiatry Clin Neurosci. (1999) 249(Suppl. 4):37–43. doi: 10.1007/PL00014183
20. Dandash O, Pantelis C, Fornito A. Dopamine, fronto-striato-thalamic circuits and risk for psychosis. Schizophr Res. (2017) 180:48–57. doi: 10.1016/j.schres.2016.08.020
21. McCutcheon RA, Abi-Dargham A, Howes OD. Schizophrenia, dopamine and the striatum: from biology to symptoms. Trends Neurosci. (2019) 42:205–20. doi: 10.1016/j.tins.2018.12.004
22. Avram M, Brandl F, Bauml J, Sorg C. Cortico-thalamic hypo- and hyperconnectivity extend consistently to basal ganglia in schizophrenia. Neuropsychopharmacology. (2018) 43:2239–248. doi: 10.1038/s41386-018-0059-z
23. Woodward ND, Heckers S. Mapping thalamocortical functional connectivity in chronic and early stages of psychotic disorders. Biol Psychiatry. (2016) 79:1016–25. doi: 10.1016/j.biopsych.2015.06.026
24. Marek GJ. Interactions of hallucinogens with the glutamatergic system: permissive network effects mediated through cortical layer v pyramidal neurons. Curr Top Behav Neurosci. (2018) 36:107–35. doi: 10.1007/7854_2017_480
25. Llado-Pelfort L, Celada P, Riga MS, Troyano-Rodriguez E, Santana N, Artigas F. Effects of hallucinogens on neuronal activity. Curr Top Behav Neurosci. (2018) 36:75–105. doi: 10.1007/7854_2017_473
26. Inserra A, De Gregorio D, Rezai T, Lopez-Canul MG, Comai S, Gobbi G. Lysergic acid diethylamide differentially modulates the reticular thalamus, mediodorsal thalamus, and infralimbic prefrontal cortex: an in vivo electrophysiology study in male mice. J Psychopharmacol. (2021) 35:469–82. doi: 10.1177/0269881121991569
27. Muller F, Lenz C, Dolder P, Lang U, Schmidt A, Liechti M, et al. Increased thalamic resting-state connectivity as a core driver of LSD-induced hallucinations. Acta Psychiatr Scand. (2017) 136:648–57. doi: 10.1111/acps.12818
28. Preller KH, Burt JB, Ji JL, Schleifer CH, Adkinson BD, Stampfli P, et al. Changes in global and thalamic brain connectivity in LSD-induced altered states of consciousness are attributable to the 5-HT2A receptor. Elife. (2018) 7:e35082. doi: 10.7554/eLife.35082
29. Woodward ND, Karbasforoushan H, Heckers S. Thalamocortical dysconnectivity in schizophrenia. Am J Psychiatry. (2012) 169:1092–9. doi: 10.1176/appi.ajp.2012.12010056
30. Anticevic A, Cole MW, Repovs G, Murray JD, Brumbaugh MS, Winkler AM, et al. Characterizing thalamo-cortical disturbances in schizophrenia and bipolar illness. Cereb Cortex. (2014) 24:3116–30. doi: 10.1093/cercor/bht165
32. Temereanca S, Simons DJ. Functional topography of corticothalamic feedback enhances thalamic spatial response tuning in the somatosensory whisker/barrel system. Neuron. (2004) 41:639–51. doi: 10.1016/S0896-6273(04)00046-7
33. Sherman SM. Thalamus plays a central role in ongoing cortical functioning. Nat Neurosci. (2016) 19:533–41. doi: 10.1038/nn.4269
34. Sherman SM, Guillery RW. Exploring the Thalamus and Its Role in Cortical Function. 2nd ed. Cambridge, MA: MIT Press (2006).
35. Lam YW, Sherman SM. Functional organization of the somatosensory cortical layer 6 feedback to the thalamus. Cereb Cortex. (2010) 20:13–24. doi: 10.1093/cercor/bhp077
36. Sherman SM. The function of metabotropic glutamate receptors in thalamus and cortex. Neuroscientist. (2014) 20:136–49. doi: 10.1177/1073858413478490
37. Behrens TE, Johansen-Berg H, Woolrich MW, Smith SM, Wheeler-Kingshott CA, Boulby PA, et al. Non-invasive mapping of connections between human thalamus and cortex using diffusion imaging. Nat Neurosci. (2003) 6:750–7. doi: 10.1038/nn1075
38. Draganski B, Kherif F, Kloppel S, Cook PA, Alexander DC, Parker GJ, et al. Evidence for segregated and integrative connectivity patterns in the human Basal Ganglia. J Neurosci. (2008) 28:7143–52. doi: 10.1523/JNEUROSCI.1486-08.2008
39. Ball G, Pazderova L, Chew A, Tusor N, Merchant N, Arichi T, et al. Thalamocortical connectivity predicts cognition in children born preterm. Cereb Cortex. (2015) 25:4310–8. doi: 10.1093/cercor/bhu331
40. Zhang D, Snyder AZ, Fox MD, Sansbury MW, Shimony JS, Raichle ME. Intrinsic functional relations between human cerebral cortex and thalamus. J Neurophysiol. (2008) 100:1740–8. doi: 10.1152/jn.90463.2008
41. Yuan R, Di X, Taylor PA, Gohel S, Tsai YH, Biswal BB. Functional topography of the thalamocortical system in human. Brain Struct Funct. (2016) 221:1971–84. doi: 10.1007/s00429-015-1018-7
42. Fox MD, Raichle ME. Spontaneous fluctuations in brain activity observed with functional magnetic resonance imaging. Nat Rev Neurosci. (2007) 8:700–11. doi: 10.1038/nrn2201
43. Stephan KE, Friston KJ, Frith CD. Dysconnection in schizophrenia: from abnormal synaptic plasticity to failures of self-monitoring. Schizophr Bull. (2009) 35:509–27. doi: 10.1093/schbul/sbn176
44. Dorph-Petersen KA, Lewis DA. Postmortem structural studies of the thalamus in schizophrenia. Schizophr Res. (2017) 180:28–35. doi: 10.1016/j.schres.2016.08.007
45. Steullet P. Thalamus-related anomalies as candidate mechanism-based biomarkers for psychosis. Schizophr Res. (2020) 226:147–57. doi: 10.1016/j.schres.2019.05.027
46. van Erp TG, Hibar DP, Rasmussen JM, Glahn DC, Pearlson GD, Andreassen OA, et al. Subcortical brain volume abnormalities in 2028 individuals with schizophrenia and 2540 healthy controls via the ENIGMA consortium. Mol Psychiatry. (2016) 21:585. doi: 10.1038/mp.2015.118
47. Minzenberg MJ, Laird AR, Thelen S, Carter CS, Glahn DC. Meta-analysis of 41 functional neuroimaging studies of executive function in schizophrenia. Arch Gen Psychiatry. (2009) 66:811–22. doi: 10.1001/archgenpsychiatry.2009.91
48. Huang AS, Rogers BP, Woodward ND. Disrupted modulation of thalamus activation and thalamocortical connectivity during dual task performance in schizophrenia. Schizophr Res. (2019) 210:270–7. doi: 10.1016/j.schres.2018.12.022
49. Giraldo-Chica M, Rogers BP, Damon SM, Landman BA, Woodward ND. Prefrontal-thalamic anatomical connectivity and executive cognitive function in schizophrenia. Biol Psychiatry. (2018) 83:509–17. doi: 10.1016/j.biopsych.2017.09.022
50. Brandl F, Avram M, Weise B, Shang J, Simoes B, Bertram T, et al. Specific substantial dysconnectivity in schizophrenia: a transdiagnostic multimodal meta-analysis of resting-state functional and structural magnetic resonance imaging studies. Biol Psychiatry. (2019) 85:573–83. doi: 10.1016/j.biopsych.2018.12.003
51. Ramsay IS. An activation likelihood estimate meta-analysis of thalamocortical dysconnectivity in psychosis. Biol Psychiatry Cogn Neurosci Neuroimaging. (2019) 4:859–69. doi: 10.1016/j.bpsc.2019.04.007
52. Anticevic A, Haut K, Murray JD, Repovs G, Yang GJ, Diehl C, et al. Association of thalamic dysconnectivity and conversion to psychosis in youth and young adults at elevated clinical risk. JAMA Psychiatry. (2015) 72:882–91. doi: 10.1001/jamapsychiatry.2015.0566
53. Cao H, Chen OY, Chung Y, Forsyth JK, McEwen SC, Gee DG, et al. Cerebello-thalamo-cortical hyperconnectivity as a state-independent functional neural signature for psychosis prediction and characterization. Nat Commun. (2018) 9:3836. doi: 10.1038/s41467-018-06350-7
54. Lui S, Yao L, Xiao Y, Keedy SK, Reilly JL, Keefe RS, et al. Resting-state brain function in schizophrenia and psychotic bipolar probands and their first-degree relatives. Psychol Med. (2015) 45:97–108. doi: 10.1017/S003329171400110X
55. Avram M, Brandl F, Knolle F, Cabello J, Leucht C, Scherr M, et al. Aberrant striatal dopamine links topographically with cortico-thalamic dysconnectivity in schizophrenia. Brain. (2020) 143:3495–505. doi: 10.1093/brain/awaa296
56. Pergola G, Danet L, Pitel AL, Carlesimo GA, Segobin S, Pariente J, et al. The regulatory role of the human mediodorsal thalamus. Trends Cogn Sci. (2018) 22:1011–25. doi: 10.1016/j.tics.2018.08.006
57. Hwang K, Bertolero MA, Liu WB, D'Esposito M. The human thalamus is an integrative hub for functional brain networks. J Neurosci. (2017) 37:5594–607. doi: 10.1523/JNEUROSCI.0067-17.2017
58. Ferri J, Ford JM, Roach BJ, Turner JA, van Erp TG, Voyvodic J, et al. Resting-state thalamic dysconnectivity in schizophrenia and relationships with symptoms. Psychol Med. (2018) 48:2492–9. doi: 10.1017/S003329171800003X
59. Seitzman BA, Gratton C, Marek S, Raut RV, Dosenbach NUF, Schlaggar BL, et al. A set of functionally-defined brain regions with improved representation of the subcortex and cerebellum. Neuroimage. (2020) 206:116290. doi: 10.1016/j.neuroimage.2019.116290
60. Greene DJ, Marek S, Gordon EM, Siegel JS, Gratton C, Laumann TO, et al. Integrative and network-specific connectivity of the basal ganglia and thalamus defined in individuals. Neuron. (2020) 105:742–58 e6. doi: 10.1016/j.neuron.2019.11.012
61. Dandash O, Fornito A, Lee J, Keefe RS, Chee MW, Adcock RA, et al. Altered striatal functional connectivity in subjects with an at-risk mental state for psychosis. Schizophr Bull. (2014) 40:904–13. doi: 10.1093/schbul/sbt093
62. Fornito A, Harrison BJ, Goodby E, Dean A, Ooi C, Nathan PJ, et al. Functional dysconnectivity of corticostriatal circuitry as a risk phenotype for psychosis. JAMA Psychiatry. (2013) 70:1143–51. doi: 10.1001/jamapsychiatry.2013.1976
63. Horga G, Cassidy CM, Xu X, Moore H, Slifstein M, Van Snellenberg JX, et al. Dopamine-related disruption of functional topography of striatal connections in unmedicated patients with schizophrenia. JAMA Psychiatry. (2016) 73:862–70. doi: 10.1001/jamapsychiatry.2016.0178
64. Sarpal DK, Robinson DG, Fales C, Lencz T, Argyelan M, Karlsgodt KH, et al. Relationship between duration of untreated psychosis and intrinsic corticostriatal connectivity in patients with early phase schizophrenia. Neuropsychopharmacology. (2017) 42:2214–21. doi: 10.1038/npp.2017.55
65. Alexander GE, DeLong MR, Strick PL. Parallel organization of functionally segregated circuits linking basal ganglia and cortex. Annu Rev Neurosci. (1986) 9:357–81. doi: 10.1146/annurev.ne.09.030186.002041
66. Swerdlow N, Koob G. Dopamine, schizophrenia, mania, and depression: toward a unified hypothesis of cortico-striatopallido-thalamic function. Behav Brain Sci. (1987) 10:197–208. doi: 10.1017/S0140525X00047488
67. Cole DM, Oei NY, Soeter RP, Both S, van Gerven JM, Rombouts SA, et al. Dopamine-dependent architecture of cortico-subcortical network connectivity. Cereb Cortex. (2013) 23:1509–16. doi: 10.1093/cercor/bhs136
68. Moore RY, Halaris AE, Jones BE. Serotonin neurons of the midbrain raphe: ascending projections. J Comp Neurol. (1978) 180:417–38. doi: 10.1002/cne.901800302
69. Vertes RP, Linley SB, Hoover WB. Pattern of distribution of serotonergic fibers to the thalamus of the rat. Brain Struct Funct. (2010) 215:1–28. doi: 10.1007/s00429-010-0249-x
70. Varela C. Thalamic neuromodulation and its implications for executive networks. Front Neural Circuits. (2014) 8:69. doi: 10.3389/fncir.2014.00069
71. Vollenweider FX, Leenders KL, Scharfetter C, Maguire P, Stadelmann O, Angst J. Positron emission tomography and fluorodeoxyglucose studies of metabolic hyperfrontality and psychopathology in the psilocybin model of psychosis. Neuropsychopharmacology. (1997) 16:357–72. doi: 10.1016/S0893-133X(96)00246-1
72. Gouzoulis-Mayfrank E, Schreckenberger M, Sabri O, Arning C, Thelen B, Spitzer M, et al. Neurometabolic effects of psilocybin, 3,4-methylenedioxyethylamphetamine (MDE) and d-methamphetamine in healthy volunteers. A double-blind, placebo-controlled PET study with [18F]FDG. Neuropsychopharmacology. (1999) 20:565–81. doi: 10.1016/S0893-133X(98)00089-X
73. Hermle L, Gouzoulis-Mayfrank E, Spitzer M. Blood flow and cerebral laterality in the mescaline model of psychosis. Pharmacopsychiatry. (1998) 31(Suppl. 2):85–91. doi: 10.1055/s-2007-979352
74. Carhart-Harris RL, Erritzoe D, Williams T, Stone JM, Reed LJ, Colasanti A, et al. Neural correlates of the psychedelic state as determined by fMRI studies with psilocybin. Proc Natl Acad Sci USA. (2012) 109:2138–43. doi: 10.1073/pnas.1119598109
75. Lewis CR, Preller KH, Kraehenmann R, Michels L, Staempfli P, Vollenweider FX. Two dose investigation of the 5-HT-agonist psilocybin on relative and global cerebral blood flow. Neuroimage. (2017) 159:70–8. doi: 10.1016/j.neuroimage.2017.07.020
76. Sipes TE, Geyer MA. DOI disrupts prepulse inhibition of startle in rats via 5-HT2A receptors in the ventral pallidum. Brain Res. (1997) 761:97–104. doi: 10.1016/S0006-8993(97)00316-8
77. Vollenweider FX, Csomor PA, Knappe B, Geyer MA, Quednow BB. The effects of the preferential 5-HT2A agonist psilocybin on prepulse inhibition of startle in healthy human volunteers depend on interstimulus interval. Neuropsychopharmacology. (2007) 32:1876–87. doi: 10.1038/sj.npp.1301324
78. Schmid Y, Enzler F, Gasser P, Grouzmann E, Preller KH, Vollenweider FX, et al. Acute effects of lysergic acid diethylamide in healthy subjects. Biol Psychiatry. (2015) 78:544–53. doi: 10.1016/j.biopsych.2014.11.015
79. Carhart-Harris RL, Leech R, Erritzoe D, Williams TM, Stone JM, Evans J, et al. Functional connectivity measures after psilocybin inform a novel hypothesis of early psychosis. Schizophr Bull. (2013) 39:1343–51. doi: 10.1093/schbul/sbs117
80. Tagliazucchi E, Roseman L, Kaelen M, Orban C, Muthukumaraswamy SD, Murphy K, et al. Increased global functional connectivity correlates with LSD-induced ego dissolution. Curr Biol. (2016) 26:1043–50. doi: 10.1016/j.cub.2016.02.010
81. Bershad AK, Preller KH, Lee R, Keedy S, Wren-Jarvis J, Bremmer MP, et al. Preliminary report on the effects of a low dose of LSD on resting-state amygdala functional connectivity. Biol Psychiatry Cogn Neurosci Neuroimaging. (2020) 5:461–7. doi: 10.1016/j.bpsc.2019.12.007
82. Luppi AI, Carhart-Harris RL, Roseman L, Pappas I, Menon DK, Stamatakis EA. LSD alters dynamic integration and segregation in the human brain. Neuroimage. (2021) 227:117653. doi: 10.1016/j.neuroimage.2020.117653
83. Li J, Kong R, Liegeois R, Orban C, Tan Y, Sun N, et al. Global signal regression strengthens association between resting-state functional connectivity and behavior. Neuroimage. (2019) 196:126–41. doi: 10.1016/j.neuroimage.2019.04.016
84. Preller KH, Razi A, Zeidman P, Stampfli P, Friston KJ, Vollenweider FX. Effective connectivity changes in LSD-induced altered states of consciousness in humans. Proc Natl Acad Sci USA. (2019) 116:2743–8. doi: 10.1073/pnas.1815129116
85. Leech R, Sharp DJ. The role of the posterior cingulate cortex in cognition and disease. Brain. (2014) 137(Pt 1):12–32. doi: 10.1093/brain/awt162
86. Schaefer A, Burmann I, Regenthal R, Arelin K, Barth C, Pampel A, et al. Serotonergic modulation of intrinsic functional connectivity. Curr Biol. (2014) 24:2314–8. doi: 10.1016/j.cub.2014.08.024
87. Chow HM, Horovitz SG, Carr WS, Picchioni D, Coddington N, Fukunaga M, et al. Rhythmic alternating patterns of brain activity distinguish rapid eye movement sleep from other states of consciousness. Proc Natl Acad Sci USA. (2013) 110:10300–5. doi: 10.1073/pnas.1217691110
88. Hale JR, White TP, Mayhew SD, Wilson RS, Rollings DT, Khalsa S, et al. Altered thalamocortical and intra-thalamic functional connectivity during light sleep compared with wake. Neuroimage. (2016) 125:657–67. doi: 10.1016/j.neuroimage.2015.10.041
89. Carhart-Harris RL, Friston KJ. REBUS and the anarchic brain: toward a unified model of the brain action of psychedelics. Pharmacol Rev. (2019) 71:316–44. doi: 10.1124/pr.118.017160
90. Gouzoulis-Mayfrank E, Habermeyer E, Hermle L, Steinmeyer A, Kunert H, Sass H. Hallucinogenic drug induced states resemble acute endogenous psychoses: results of an empirical study. Eur Psychiatry. (1998) 13:399–406. doi: 10.1016/S0924-9338(99)80686-5
91. Shanon B. The Antipodes of the Mind Charting the Phenomenology of the Ayahuasca Experience. Oxford: Oxford University Press (2002).
92. Sanz C, Zamberlan F, Erowid E, Erowid F, Tagliazucchi E. The experience elicited by hallucinogens presents the highest similarity to dreaming within a large database of psychoactive substance reports. Front Neurosci. (2018) 12:229. doi: 10.3389/fnins.2018.00229
93. Carhart-Harris RL, Muthukumaraswamy S, Roseman L, Kaelen M, Droog W, Murphy K, et al. Neural correlates of the LSD experience revealed by multimodal neuroimaging. Proc Natl Acad Sci USA. (2016) 113:4853–8. doi: 10.1073/pnas.1518377113
94. Barrett FS, Griffiths RR. Classic hallucinogens and mystical experiences: phenomenology and neural correlates. Curr Top Behav Neurosci. (2018) 36:393–430. doi: 10.1007/7854_2017_474
95. Cho KI, Shenton ME, Kubicki M, Jung WH, Lee TY, Yun JY, et al. Altered thalamo-cortical white matter connectivity: probabilistic tractography study in clinical-high risk for psychosis and first-episode psychosis. Schizophr Bull. (2016) 42:723–31. doi: 10.1093/schbul/sbv169
96. Csukly G, Szabo A, Polgar P, Farkas K, Gyebnar G, Kozak LR, et al. Fronto-thalamic structural and effective connectivity and delusions in schizophrenia: a combined DTI/DCM study. Psychol Med. (2020) 51:2083–93. doi: 10.1017/S0033291720000859
97. Kubota M, Miyata J, Sasamoto A, Sugihara G, Yoshida H, Kawada R, et al. Thalamocortical disconnection in the orbitofrontal region associated with cortical thinning in schizophrenia. JAMA Psychiatry. (2013) 70:12–21. doi: 10.1001/archgenpsychiatry.2012.1023
98. Sheffield JM, Huang AS, Rogers BP, Giraldo-Chica M, Landman BA, Blackford JU, et al. Thalamocortical anatomical connectivity in schizophrenia and psychotic bipolar disorder. Schizophr Bull. (2020) 46:1062–71. doi: 10.1093/schbul/sbaa022
99. Yao B, Neggers SFW, Kahn RS, Thakkar KN. Altered thalamocortical structural connectivity in persons with schizophrenia and healthy siblings. Neuroimage Clin. (2020) 28:102370. doi: 10.1016/j.nicl.2020.102370
100. Xi C, Liu ZN, Yang J, Zhang W, Deng MJ, Pan YZ, et al. Schizophrenia patients and their healthy siblings share decreased prefronto-thalamic connectivity but not increased sensorimotor-thalamic connectivity. Schizophr Res. (2020) 222:354–61. doi: 10.1016/j.schres.2020.04.033
101. Corlett PR, Taylor JR, Wang XJ, Fletcher PC, Krystal JH. Toward a neurobiology of delusions. Prog Neurobiol. (2010) 92:345–69. doi: 10.1016/j.pneurobio.2010.06.007
102. Sterzer P, Adams RA, Fletcher P, Frith C, Lawrie SM, Muckli L, et al. The predictive coding account of psychosis. Biol Psychiatry. (2018) 84:634–43. doi: 10.1016/j.biopsych.2018.05.015
103. Kanai R, Komura Y, Shipp S, Friston K. Cerebral hierarchies: predictive processing, precision and the pulvinar. Philos Trans R Soc Lond B Biol Sci. (2015) 370:1668. doi: 10.1098/rstb.2014.0169
104. Halberstadt AL, Geyer MA. Multiple receptors contribute to the behavioral effects of indoleamine hallucinogens. Neuropharmacology. (2011) 61:364–81. doi: 10.1016/j.neuropharm.2011.01.017
105. Drago A, Ronchi DD, Serretti A. 5-HT1A gene variants and psychiatric disorders: a review of current literature and selection of SNPs for future studies. Int J Neuropsychopharmacol. (2008) 11:701–21. doi: 10.1017/S1461145707008218
106. Nutt D, Erritzoe D, Carhart-Harris R. Psychedelic psychiatry's brave new world. Cell. (2020) 181:24–8. doi: 10.1016/j.cell.2020.03.020
107. Shepherd GM. Corticostriatal connectivity and its role in disease. Nat Rev Neurosci. (2013) 14:278–91. doi: 10.1038/nrn3469
108. Rolls ET, Huang CC, Lin CP, Feng J, Joliot M. Automated anatomical labelling atlas 3. Neuroimage. (2020) 206:116189. doi: 10.1016/j.neuroimage.2019.116189
109. Pergola G, Selvaggi P, Trizio S, Bertolino A, Blasi G. The role of the thalamus in schizophrenia from a neuroimaging perspective. Neurosci Biobehav Rev. (2015) 54:57–75. doi: 10.1016/j.neubiorev.2015.01.013
110. Parnaudeau S, O'Neill PK, Bolkan SS, Ward RD, Abbas AI, Roth BL, et al. Inhibition of mediodorsal thalamus disrupts thalamofrontal connectivity and cognition. Neuron. (2013) 77:1151–62. doi: 10.1016/j.neuron.2013.01.038
111. Antonucci LA, Taurisano P, Fazio L, Gelao B, Romano R, Quarto T, et al. Association of familial risk for schizophrenia with thalamic and medial prefrontal functional connectivity during attentional control. Schizophr Res. (2016) 173:23–9. doi: 10.1016/j.schres.2016.03.014
112. Muller F, Liechti ME, Lang UE, Borgwardt S. Advances and challenges in neuroimaging studies on the effects of serotonergic hallucinogens: contributions of the resting brain. Prog Brain Res. (2018) 242:159–77. doi: 10.1016/bs.pbr.2018.08.004
113. De Deurwaerdere P, Chagraoui A, Di Giovanni G. Serotonin/dopamine interaction: electrophysiological and neurochemical evidence. Prog Brain Res. (2021) 261:161–264. doi: 10.1016/bs.pbr.2021.02.001
114. Meltzer HY, Huang M. In vivo actions of atypical antipsychotic drug on serotonergic and dopaminergic systems. Prog Brain Res. (2008) 172:177–97. doi: 10.1016/S0079-6123(08)00909-6
115. Egerton A, Grace AA, Stone J, Bossong MG, Sand M, McGuire P. Glutamate in schizophrenia: neurodevelopmental perspectives and drug development. Schizophr Res. (2020) 223:59–70. doi: 10.1016/j.schres.2020.09.013
116. Vollenweider FX, Vontobel P, Hell D, Leenders KL. 5-HT modulation of dopamine release in basal ganglia in psilocybin-induced psychosis in man–a PET study with [11C]raclopride. Neuropsychopharmacology. (1999) 20:424–33. doi: 10.1016/S0893-133X(98)00108-0
117. Schmidt TT, Jagannathan N, Ljubljanac M, Xavier A, Nierhaus T. The multimodal Ganzfeld-induced altered state of consciousness induces decreased thalamo-cortical coupling. Sci Rep. (2020) 10:18686. doi: 10.1038/s41598-020-75019-3
118. Schmidt TT, Prein JC. The Ganzfeld experience-A stably inducible altered state of consciousness: effects of different auditory homogenizations. Psych J. (2019) 8:66–81. doi: 10.1002/pchj.262
119. Beck K, Hindley G, Borgan F, Ginestet C, McCutcheon R, Brugger S, et al. Association of ketamine with psychiatric symptoms and implications for its therapeutic use and for understanding schizophrenia: a systematic review and meta-analysis. JAMA Netw Open. (2020) 3:e204693. doi: 10.1001/jamanetworkopen.2020.4693
120. Hoflich A, Hahn A, Kublbock M, Kranz GS, Vanicek T, Windischberger C, et al. Ketamine-induced modulation of the thalamo-cortical network in healthy volunteers as a model for schizophrenia. Int J Neuropsychopharmacol. (2015) 18:pyv040. doi: 10.1093/ijnp/pyv040
121. Anticevic A, Yang G, Savic A, Murray JD, Cole MW, Repovs G, et al. Mediodorsal and visual thalamic connectivity differ in schizophrenia and bipolar disorder with and without psychosis history. Schizophr Bull. (2014) 40:1227–43. doi: 10.1093/schbul/sbu100
122. Fleming LM, Javitt DC, Carter CS, Kantrowitz JT, Girgis RR, Kegeles LS, et al. A multicenter study of ketamine effects on functional connectivity: large scale network relationships, hubs and symptom mechanisms. Neuroimage Clin. (2019) 22:101739. doi: 10.1016/j.nicl.2019.101739
123. Carhart-Harris RL, Kaelen M, Bolstridge M, Williams TM, Williams LT, Underwood R, et al. The paradoxical psychological effects of lysergic acid diethylamide (LSD). Psychol Med. (2016) 46:1379–90. doi: 10.1017/S0033291715002901
124. Schmid Y, Liechti ME. Long-lasting subjective effects of LSD in normal subjects. Psychopharmacology. (2018) 235:535–45. doi: 10.1007/s00213-017-4733-3
125. Woodward ND, Giraldo-Chica M, Rogers B, Cascio CJ. Thalamocortical dysconnectivity in autism spectrum disorder: an analysis of the Autism Brain Imaging Data Exchange. Biol Psychiatry Cogn Neurosci Neuroimaging. (2017) 2:76–84. doi: 10.1016/j.bpsc.2016.09.002
126. Brown EC, Clark DL, Hassel S, MacQueen G, Ramasubbu R. Thalamocortical connectivity in major depressive disorder. J Affect Disord. (2017) 217:125–31. doi: 10.1016/j.jad.2017.04.004
127. Gong J, Wang J, Chen P, Qi Z, Luo Z, Wang J, et al. Large-scale network abnormality in bipolar disorder: a multimodal meta-analysis of resting-state functional and structural magnetic resonance imaging studies. J Affect Disord. (2021) 292:9–20. doi: 10.1016/j.jad.2021.05.052
128. Kaiser RH, Andrews-Hanna JR, Wager TD, Pizzagalli DA. Large-scale network dysfunction in major depressive disorder: a meta-analysis of resting-state functional connectivity. JAMA Psychiatry. (2015) 72:603–11. doi: 10.1001/jamapsychiatry.2015.0071
129. Lau WKW, Leung MK, Lau BWM. Resting-state abnormalities in Autism Spectrum Disorders: a meta-analysis. Sci Rep. (2019) 9:3892. doi: 10.1038/s41598-019-40427-7
130. Gursel DA, Avram M, Sorg C, Brandl F, Koch K. Frontoparietal areas link impairments of large-scale intrinsic brain networks with aberrant fronto-striatal interactions in OCD: a meta-analysis of resting-state functional connectivity. Neurosci Biobehav Rev. (2018) 87:151–60. doi: 10.1016/j.neubiorev.2018.01.016
131. Tu PC, Bai YM, Li CT, Chen MH, Lin WC, Chang WC, et al. Identification of common thalamocortical dysconnectivity in four major psychiatric disorders. Schizophr Bull. (2019) 45:1143–51. doi: 10.1093/schbul/sby166
132. Sarpal DK, Robinson DG, Lencz T, Argyelan M, Ikuta T, Karlsgodt K, et al. Antipsychotic treatment and functional connectivity of the striatum in first-episode schizophrenia. JAMA Psychiatry. (2015) 72:5–13. doi: 10.1001/jamapsychiatry.2014.1734
133. Wang YC, Tang WJ, Fan XD, Zhang JY, Geng DY, Jiang KD, et al. Resting-state functional connectivity changes within the default mode network and the salience network after antipsychotic treatment in early-phase schizophrenia. Neuropsych Dis Treat. (2017) 13:397–406. doi: 10.2147/NDT.S123598
134. Chopra S, Francey SM, O'Donoghue B, Sabaroedin K, Arnatkeviciute A, Cropley V, et al. Functional connectivity in antipsychotic-treated and antipsychotic-naive patients with first-episode psychosis and low risk of self-harm or aggression: a secondary analysis of a randomized clinical trial. JAMA Psychiatry. (2021) 78:994–1004. doi: 10.1001/jamapsychiatry.2021.1422
135. Zhang M, Palaniyappan L, Deng M, Zhang W, Pan Y, Fan Z, et al. Abnormal thalamocortical circuit in adolescents with early-onset schizophrenia. J Am Acad Child Adolesc Psychiatry. (2021) 60:479–89. doi: 10.1016/j.jaac.2020.07.903
136. Baran B, Karahanoglu FI, Mylonas D, Demanuele C, Vangel M, Stickgold R, et al. Increased thalamocortical connectivity in schizophrenia correlates with sleep spindle deficits: evidence for a common pathophysiology. Biol Psychiatry Cogn Neurosci Neuroimaging. (2019) 4:706–14. doi: 10.1016/j.bpsc.2019.04.012
Keywords: resting-state FC-fMRI, cortico-thalamic connectivity, psychotic states, psychedelic states, serotonergic psychedelics
Citation: Avram M, Rogg H, Korda A, Andreou C, Müller F and Borgwardt S (2021) Bridging the Gap? Altered Thalamocortical Connectivity in Psychotic and Psychedelic States. Front. Psychiatry 12:706017. doi: 10.3389/fpsyt.2021.706017
Received: 06 May 2021; Accepted: 16 September 2021;
Published: 13 October 2021.
Edited by:
Katrin H. Preller, University of Zurich, SwitzerlandReviewed by:
Timo Torsten Schmidt, Free University Berlin, GermanyPierluigi Selvaggi, King's College London, United Kingdom
Tae Young Lee, Pusan National University Yangsan Hospital, South Korea
Copyright © 2021 Avram, Rogg, Korda, Andreou, Müller and Borgwardt. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Mihai Avram, bWloYWkuYXZyYW1AdWtzaC5kZQ==